
Mapping and diagnostic marker development for Soil-borne cereal mosaic virus resistance in bread...
13
Mapping and diagnostic marker development for Soil-borne cereal mosaic virus resistance in bread wheat Dragan Perovic Jutta Fo ¨rster Pierre Devaux Djabbar Hariri Morgane Guilleroux Kostya Kanyuka Rebecca Lyons Jens Weyen David Feuerhelm Ute Kastirr Pierre Sourdille Marion Ro ¨der Frank Ordon Received: 18 June 2008 / Accepted: 22 January 2009 / Published online: 12 February 2009 Ó Springer Science+Business Media B.V. 2009 Abstract Monogenically-inherited resistance to Soil-borne cereal mosaic virus (SBCMV) in hexa- ploid bread wheat cultivars ‘Tremie’ and ‘Claire’ was mapped on chromosome 5D. The two closest flanking markers identified in the Claire-derived mapping population, Xgwm469-5D and E37M49, are linked to the resistance locus at distances of 1 and 9 cm, respectively. Xgwm469-5D co-segregated with the SBCMV resistance in the Tremie-derived population and with the recently identified Sbm1 locus in the cv. Cadenza. This suggested that Tremie and Claire carry a resistance gene allelic to Sbm1, or one closely linked to it. The diagnostic value of Xgwm469-5D was assessed using a collection of SBCMV resistant and susceptible cultivars. Importantly, all susceptible genotypes carried a null allele of Xgwm469-5D, D. Perovic F. Ordon (&) Julius Kuehn-Institute, Federal Research Centre for Cultivated Plants, Institute for Resistance Research and Stress Tolerance, Erwin-Baur-Str. 27, 06484 Quedlinburg, Germany e-mail: [email protected] D. Perovic e-mail: [email protected] J. Fo ¨rster J. Weyen Saaten-Union Resistenzlabor GmbH, Hovedisser Str. 92, 33818 Leopoldsho ¨he, Germany P. Devaux Florimond Desprez, 3, Rue Florimond Desprez, 59242 Cappelle en Pe ´ve `le, France D. Hariri M. Guilleroux INRA, BIOGER, Route de Saint Cyr, 78026 Versailles Cedex, France K. Kanyuka R. Lyons Department of Plant Pathology and Microbiology, Centre for Sustainable Pest and Disease Management, Rothamsted Research, Harpenden, AL5 2JQ Hertfordshire, UK D. Feuerhelm Elsoms Seeds Ltd, Pinchbeck Road, PE11 1QG Spalding, UK U. Kastirr Institute for Epidemiology and Pathogen Diagnostics, Julius Kuehn-Institute, Federal Research Centre for Cultivated Plants, Erwin-Baur-Str. 27, 06484 Quedlinburg, Germany P. Sourdille UMR 1095 Ame ´lioration et Sante ´ des Plantes, INRA, Domaine de Crouelle 234, Avenue du Bre ´zet, Clermont Ferrand, France M. Ro ¨der Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), Corrensstr. 3, 06466 Gatersleben, Germany 123 Mol Breeding (2009) 23:641–653 DOI 10.1007/s11032-009-9262-2
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Mapping and diagnostic marker development for Soil-borne cereal mosaic virus resistance in bread...

Mapping and diagnostic marker development for Soil-bornecereal mosaic virus resistance in bread wheat
Dragan Perovic Æ Jutta Forster Æ Pierre Devaux Æ Djabbar Hariri Æ Morgane Guilleroux ÆKostya Kanyuka Æ Rebecca Lyons Æ Jens Weyen Æ David Feuerhelm Æ Ute Kastirr ÆPierre Sourdille Æ Marion Roder Æ Frank Ordon
Received: 18 June 2008 / Accepted: 22 January 2009 / Published online: 12 February 2009
� Springer Science+Business Media B.V. 2009
Abstract Monogenically-inherited resistance to
Soil-borne cereal mosaic virus (SBCMV) in hexa-
ploid bread wheat cultivars ‘Tremie’ and ‘Claire’ was
mapped on chromosome 5D. The two closest flanking
markers identified in the Claire-derived mapping
population, Xgwm469-5D and E37M49, are linked to
the resistance locus at distances of 1 and 9 cm,
respectively. Xgwm469-5D co-segregated with the
SBCMV resistance in the Tremie-derived population
and with the recently identified Sbm1 locus in the cv.
Cadenza. This suggested that Tremie and Claire carry
a resistance gene allelic to Sbm1, or one closely
linked to it. The diagnostic value of Xgwm469-5D
was assessed using a collection of SBCMV resistant
and susceptible cultivars. Importantly, all susceptible
genotypes carried a null allele of Xgwm469-5D,
D. Perovic � F. Ordon (&)
Julius Kuehn-Institute, Federal Research Centre for
Cultivated Plants, Institute for Resistance Research and
Stress Tolerance, Erwin-Baur-Str. 27, 06484 Quedlinburg,
Germany
e-mail: [email protected]
D. Perovic
e-mail: [email protected]
J. Forster � J. Weyen
Saaten-Union Resistenzlabor GmbH, Hovedisser Str. 92,
33818 Leopoldshohe, Germany
P. Devaux
Florimond Desprez, 3, Rue Florimond Desprez, 59242
Cappelle en Pevele, France
D. Hariri � M. Guilleroux
INRA, BIOGER, Route de Saint Cyr, 78026 Versailles
Cedex, France
K. Kanyuka � R. Lyons
Department of Plant Pathology and Microbiology, Centre
for Sustainable Pest and Disease Management,
Rothamsted Research, Harpenden, AL5 2JQ
Hertfordshire, UK
D. Feuerhelm
Elsoms Seeds Ltd, Pinchbeck Road, PE11 1QG Spalding,
UK
U. Kastirr
Institute for Epidemiology and Pathogen Diagnostics,
Julius Kuehn-Institute, Federal Research Centre for
Cultivated Plants, Erwin-Baur-Str. 27, 06484
Quedlinburg, Germany
P. Sourdille
UMR 1095 Amelioration et Sante des Plantes, INRA,
Domaine de Crouelle 234, Avenue du Brezet, Clermont
Ferrand, France
M. Roder
Leibniz Institute of Plant Genetics and Crop Plant
Research (IPK), Corrensstr. 3, 06466 Gatersleben,
Germany
123
Mol Breeding (2009) 23:641–653
DOI 10.1007/s11032-009-9262-2

whereas resistant genotypes presumably related to
either Claire and Tremie or Cadenza revealed a 152
or 154 bp allele of Xgwm469-5D, respectively.
Therefore, Xgwm469-5D is well suited for marker
assisted selection for SBCMV resistance.
Keywords Soil-borne cereal mosaic virus
(SBCMV) � Winter wheat (Triticum aestivum) �Resistance � Sbm1 � Marker assisted selection (MAS)
Introduction
Mosaic disease, caused by Soil-borne cereal mosaic
virus (SBCMV), which belongs to the genus Furovi-
rus, is a serious constraint to winter wheat production
in Europe, North America and Asia (Clover et al.
2001). SBCMV is transmitted by the plasmodiophorid
Polymyxa graminis, a eukaryotic soil-borne micro-
organism that has been detected down to a soil depth
of 60 cm and colonises roots of Gramineae plants
(Rao and Brakke 1969). Following transmission by
P. graminis, SBCMV is translocated into the upper
parts of susceptible plants causing stunting and
mosaic symptoms on leaves that are most prominent
in early spring. Winter wheat plants infected in
autumn are particularly sensitive to frost damage
resulting in increased winter killing or reduced vigour
during spring. Chemical control except soil fumiga-
tion, which is unacceptable for economical and
ecological reasons, is ineffective against P. graminis.
Furthermore, as virus-containing resting spores of
P. graminis are distributed by wind, water and
machinery and can survive in the soil for decades
(Brakke and Langenberg 1988), crop rotation is not an
effective option for disease control either. Therefore,
the only possibility of controlling this disease on
infested fields is the growing of resistant cultivars.
Historically, Soil-borne wheat mosaic virus
(SBWMV) was first recognized as a serious winter
wheat disease in Indiana and Illinois in 1919 (Mc-
Kinney 1923) and has subsequently been detected in
other winter wheat growing areas around the globe
(Budge and Henry 2002; Proeseler and Stanarius
1983; Saito et al. 1964). In Europe, however, the most
common causal agent of mosaic disease of wheat
(Triticum aestivum), durum wheat (Triticum durum),
rye (Secale cereale) and triticale is SBCMV, a virus
species related to SBWMV and until recently
considered as a European strain of SBWMV. Com-
plete sequencing of SBWMV and SBCMV genomes
revealed that they share only approximately 70%
nucleotide identity, and therefore should be classified
as separate species (Koenig and Huth 2000; Torrance
and Koenig 2004). In addition, it has been shown that
these two viruses can be distinguished biologically by
infection phenotype on the indicator plant Nicotiana
benthamiana. Whereas SBCMV spreads systemically
and causes leaf chlorosis and subsequent plant
death, SBWMV is restricted to the inoculated leaves
(Kastirr et al. 2006). More recently, it has also been
shown that SBWMV and SBCMV can be distin-
guished by their infection phenotype on some barley
genotypes (Lyons et al. 2008).
SBCMV-resistant wheat cultivars exhibit the
so-called ‘translocation resistance’ (Hariri et al.
1987). In such cultivars, virus accumulates to a
certain degree in the roots, but its movement from the
roots to the stem and leaves is blocked or delayed
such that the viral coat protein antigen is undetect-
able in above ground tissues. Resistant cultivars have
been known in France for many years, but until now
no comprehensive information on the genetic control
of resistance in these cultivars have been available.
Very recently, a locus for resistance to SBCMV,
designated as Sbm1, has been mapped to the long
arm of chromosome 5D in the UK wheat cv. Cadenza
(Bass et al. 2006). However, it appears that Cadenza
shares no common ancestry, regarding donors of
resistance, with SBCMV-resistant cultivars com-
monly grown in France (e.g., Tremie, Claire and
Moulin) (Bayles and Napier 2002). Therefore, the
present study aimed at obtaining detailed information
on the genetics of SBCMV resistance in elite
European wheat cultivars that are unrelated to cv.
Cadenza, and developing molecular markers which
can be employed for efficient marker assisted selec-
tion (MAS) in breeding for SBCMV resistance.
Development of diagnostic markers for rapid identi-
fication of SBCMV resistant genotypes is of special
importance in wheat breeding as field based pheno-
typic selection is expensive, laborious and time
consuming.
642 Mol Breeding (2009) 23:641–653
123

Materials and methods
Plant materials
Doubled haploid (DH) wheat populations segregating
for resistance to SBCMV were derived from crosses
between the resistant cv. Tremie and the susceptible
cultivars Texel, Aztek and Soissons (in total 64 DH
lines), and between the resistant cv. Claire and the
susceptible cv. Savannah (126 DH lines) using inter-
generic hybridisation with maize (Devaux and
Pickering 2005; Table 1). In addition, the population
of 204 DH individuals, derived from a cross between
susceptible cv. Avalon and resistant cv. Cadenza
(Kanyuka et al. 2004) was used for linkage analy-
sis between Sbm1 and the microsatellite marker
Xgwm469. For chromosomal and deletion bin assign-
ment of molecular markers, a set of 21 nulli-tetrasomic
chromosome substitution lines (Sears 1954, 1966) and
one deletion line 5DL5 of the wheat cv. Chinese
Spring (Endo and Gill 1996; Werner et al. 1992) was
used. The diagnostic value of the markers linked to
SBCMV resistance was assessed using a genetically
diverse collection of 99 wheat cultivars that were
tested for resistance to SBCMV (Table 2). Seeds of
these genotypes were obtained primarily from the
National Small Grains Collection, USDA-ARS, Aber-
deen, Idaho, USA.
SBCMV resistance tests
In order to assess resistance of segregating DH
populations, visual scoring and Double Antibody
Sandwich Enzyme-Linked Immunosorbent Assay
(DAS-ELISA) were carried out on plants grown in
highly and uniformly infested soils. DH populations
were evaluated during two growing seasons (2004/
2005 and 2005/2006) at two different locations in
France (Vatan) and Germany (Walternienburg). Each
DH line and parental cultivar was grown as a double-
row (30 plants per row) with two replications at each
location. The method for visual disease scoring was
as follows: all lines were scored on a scale of 0–9 for
appearance of yellow mosaic on leaves. A score of
‘0’ indicated no plants displaying yellow mosaic are
present, and a score of ‘9’ indicated 80–100% of
plants with yellow mosaic symptoms are observed.
DAS-ELISA was performed according to Clark and
Adams (1977). For each DH line ten plants individ-
ually or in two bulks of five plants were ground in a
0.1 M citrate buffer (pH 7.2) containing 0.5 M urea
and centrifuged at 10,000 g for 3 min. The superna-
tants were collected and 100 ll aliquots were added
to each well, which previously had been coated with
SBCMV IgG (1 lg ml-1). After incubation overnight
at 4�C, plates were washed three times and the
alkaline phosphatase-conjugated IgG were added.
After incubation for 4 h at 37�C and a further
washing step, p-nitrophenyl phosphate substrate was
added and extinction was measured at 405 nm.
Samples were considered to be positive when the
E450 value was more than three times that of the
healthy control. In disease scoring three distinct
phenotypic classes were observed as follows: 1)
resistant (with ELISA values B 0.3 and visual symp-
toms scores of ‘0’–‘3’), 2) moderately resistant (with
ELISA values 0.3–0.5 and visual symptoms scores of
‘3’–‘5’), and susceptible (with ELISA values C 0.5
and visual symptoms scores of ‘5’–‘9’).
An additional collection of 99 wheat genotypes of
European and ‘foreign’ origin with known or putative
resistance to furoviruses was assembled for SBCMV
resistance screening and SSR analyses. Plants were
grown in SBCMV-infested soils collected from an
Table 1 Pedigree and segregation ratios of DH-populations used for mapping SBCMV resistance in wheat
Cross Number of DH
lines in progeny
Number of
resistant DHs
Number of
susceptible DHs
v2(1r:1s) P df
Tremie 9 Aztek 17 – – – – –
Tremie 9 Texel 17 – – – – –
Tremie 9 Soissons 30 – – – – –
Tremie-derived population (combined) 64 27 37 1.56 0.211 1
Claire 9 Savannah 126 69 57 1.14 0.285 1
v2, chi-square test; P, probability; df, degrees of freedom
Mol Breeding (2009) 23:641–653 643
123

Table 2 Winter wheat genotypes screened with SSR WMS469 and their reactions to SBCMV
No. Accession identifiera Acsession name Reaction to SBCMV Xgwm469-5DL allele Xwmc765
1 PI 81793 Akagawa Ako Ichigo R 154 179
2 Alixan R 154 nt
3 PI 340666 Alpha R 152 195
4 Altigo R 154 nt
5 Arsene R 152 nt
6 Autan R 152 nt
7 Avalon S – 199
8 Axima R 152 nt
9 Axona S – 191
10 Aztek S – 193
11 Badger S – 221
12 Biscay S – 193
13 Buster S – 195
14 Cadenza R 154 209
15 Charger R 152 199
16 Chatsworth S – 179
17 Chinese Spring S – 179
18 PI 338915 Ciano F67 S – 191
19 Claire R 152 193
20 Cltr 17882 Line B S – 199
21 Cltr 17886 Line G R 152 199
22 CItr 11673 Comanche R 154 209
23 Consort S – 195
24 Corvus S – 199
25 Deben S – 195
26 Domino S – 191
27 PI 414565 Dong Fang Hong 3 MR – 177
28 PI 361730 Dronning Wilhelmine II S – 189
29 PI 591964 Empraba 16b S – 225
30 Ephoros S – 189
31 PI 262231 Etoile de Choisy MR 154 187
32 PI 476850 F�S. 401 MR 152 167
33 Flame R 152 199
34 Hadm10437-1 S – 193
35 PI 340707 Heines C73 R 154 209
36 Hereward R 152 199
37 CItr 17264 Homestead MR 152 195
38 PI 405702 Hong Liang 5b S – 191
39 PI 495015 IL 77-4259 MR 154 179
40 PI 593688 Jagger R 154 179
41 Kharchia Local S – 167
42 PI 351622 Klein Rendidor MR 152 191
43 PI 566671 KS92WGRC22b S – 179
44 PI 566670 KS92WRGC21b MR – 179
644 Mol Breeding (2009) 23:641–653
123
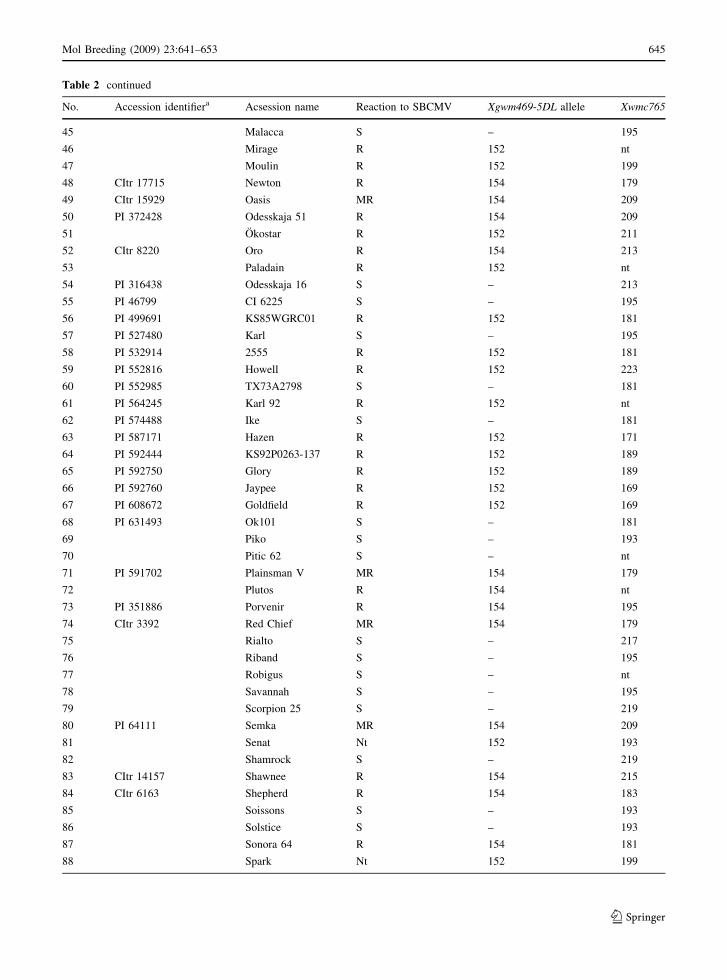
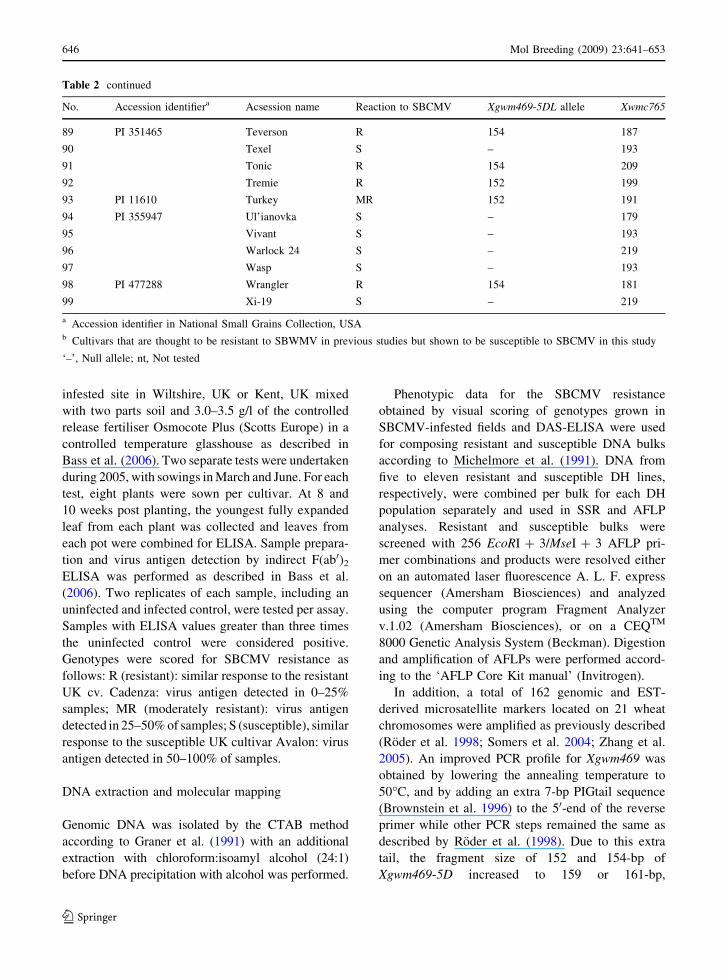
Table 2 continued
No. Accession identifiera Acsession name Reaction to SBCMV Xgwm469-5DL allele Xwmc765
45 Malacca S – 195
46 Mirage R 152 nt
47 Moulin R 152 199
48 CItr 17715 Newton R 154 179
49 CItr 15929 Oasis MR 154 209
50 PI 372428 Odesskaja 51 R 154 209
51 Okostar R 152 211
52 CItr 8220 Oro R 154 213
53 Paladain R 152 nt
54 PI 316438 Odesskaja 16 S – 213
55 PI 46799 CI 6225 S – 195
56 PI 499691 KS85WGRC01 R 152 181
57 PI 527480 Karl S – 195
58 PI 532914 2555 R 152 181
59 PI 552816 Howell R 152 223
60 PI 552985 TX73A2798 S – 181
61 PI 564245 Karl 92 R 152 nt
62 PI 574488 Ike S – 181
63 PI 587171 Hazen R 152 171
64 PI 592444 KS92P0263-137 R 152 189
65 PI 592750 Glory R 152 189
66 PI 592760 Jaypee R 152 169
67 PI 608672 Goldfield R 152 169
68 PI 631493 Ok101 S – 181
69 Piko S – 193
70 Pitic 62 S – nt
71 PI 591702 Plainsman V MR 154 179
72 Plutos R 154 nt
73 PI 351886 Porvenir R 154 195
74 CItr 3392 Red Chief MR 154 179
75 Rialto S – 217
76 Riband S – 195
77 Robigus S – nt
78 Savannah S – 195
79 Scorpion 25 S – 219
80 PI 64111 Semka MR 154 209
81 Senat Nt 152 193
82 Shamrock S – 219
83 CItr 14157 Shawnee R 154 215
84 CItr 6163 Shepherd R 154 183
85 Soissons S – 193
86 Solstice S – 193
87 Sonora 64 R 154 181
88 Spark Nt 152 199
Mol Breeding (2009) 23:641–653 645
123
infested site in Wiltshire, UK or Kent, UK mixed
with two parts soil and 3.0–3.5 g/l of the controlled
release fertiliser Osmocote Plus (Scotts Europe) in a
controlled temperature glasshouse as described in
Bass et al. (2006). Two separate tests were undertaken
during 2005, with sowings in March and June. For each
test, eight plants were sown per cultivar. At 8 and
10 weeks post planting, the youngest fully expanded
leaf from each plant was collected and leaves from
each pot were combined for ELISA. Sample prepara-
tion and virus antigen detection by indirect F(ab0)2
ELISA was performed as described in Bass et al.
(2006). Two replicates of each sample, including an
uninfected and infected control, were tested per assay.
Samples with ELISA values greater than three times
the uninfected control were considered positive.
Genotypes were scored for SBCMV resistance as
follows: R (resistant): similar response to the resistant
UK cv. Cadenza: virus antigen detected in 0–25%
samples; MR (moderately resistant): virus antigen
detected in 25–50% of samples; S (susceptible), similar
response to the susceptible UK cultivar Avalon: virus
antigen detected in 50–100% of samples.
DNA extraction and molecular mapping
Genomic DNA was isolated by the CTAB method
according to Graner et al. (1991) with an additional
extraction with chloroform:isoamyl alcohol (24:1)
before DNA precipitation with alcohol was performed.
Phenotypic data for the SBCMV resistance
obtained by visual scoring of genotypes grown in
SBCMV-infested fields and DAS-ELISA were used
for composing resistant and susceptible DNA bulks
according to Michelmore et al. (1991). DNA from
five to eleven resistant and susceptible DH lines,
respectively, were combined per bulk for each DH
population separately and used in SSR and AFLP
analyses. Resistant and susceptible bulks were
screened with 256 EcoRI ? 3/MseI ? 3 AFLP pri-
mer combinations and products were resolved either
on an automated laser fluorescence A. L. F. express
sequencer (Amersham Biosciences) and analyzed
using the computer program Fragment Analyzer
v.1.02 (Amersham Biosciences), or on a CEQTM
8000 Genetic Analysis System (Beckman). Digestion
and amplification of AFLPs were performed accord-
ing to the ‘AFLP Core Kit manual’ (Invitrogen).
In addition, a total of 162 genomic and EST-
derived microsatellite markers located on 21 wheat
chromosomes were amplified as previously described
(Roder et al. 1998; Somers et al. 2004; Zhang et al.
2005). An improved PCR profile for Xgwm469 was
obtained by lowering the annealing temperature to
50�C, and by adding an extra 7-bp PIGtail sequence
(Brownstein et al. 1996) to the 50-end of the reverse
primer while other PCR steps remained the same as
described by Roder et al. (1998). Due to this extra
tail, the fragment size of 152 and 154-bp of
Xgwm469-5D increased to 159 or 161-bp,
Table 2 continued
No. Accession identifiera Acsession name Reaction to SBCMV Xgwm469-5DL allele Xwmc765
89 PI 351465 Teverson R 154 187
90 Texel S – 193
91 Tonic R 154 209
92 Tremie R 152 199
93 PI 11610 Turkey MR 152 191
94 PI 355947 Ul’ianovka S – 179
95 Vivant S – 193
96 Warlock 24 S – 219
97 Wasp S – 193
98 PI 477288 Wrangler R 154 181
99 Xi-19 S – 219
a Accession identifier in National Small Grains Collection, USAb Cultivars that are thought to be resistant to SBWMV in previous studies but shown to be susceptible to SBCMV in this study
‘–’, Null allele; nt, Not tested
646 Mol Breeding (2009) 23:641–653
123

respectively. Amplifications were carried out in a
AB9700 Thermal Cycler (Applied Biosystems) or a
DNA Engine PTC-200 Thermal Cycler (MJ
Research) using fluorescently labelled (Cy5 or
DY-681) forward and unlabeled reverse microsatel-
lite primers. Products were resolved either on the
A. L. F. ExpressTM automated laser fluorescence
sequencer (Amersham Biosciences), on the CEQTM
8000 Genetic Analysis System (Beckman), or on the
NEN Model 4300 DNA Analyzer (Li-Cor) as per the
manufacturer’s instructions.
Computational linkage analysis was carried out
using Joinmap v.3.0 (Stam 1993) software.
Results
Disease scoring
Visual evaluation and DAS ELISA-based disease
scoring of 190 DH lines during two field seasons
at two geographical locations (i.e., France and
Germany) yielded a uniform and consistent set of
data for all progeny plants in both mapping popula-
tions. Phenotypic and ELISA data were highly and
significantly correlated, i.e., 0.95*** and 0.89***,
respectively, in the different DH-populations. In
general, two distinct phenotypic classes were
observed, since only a small proportion of DH lines,
five and nine DH lines in Tremie- and Claire-derived
populations, respectively, had an intermediate reac-
tion with visual symptom scores between ‘3’ and ‘5’
and ELISA values 0.3–0.5 Therefore, both resistance
classes (resistant and moderately resistant) were
scored as resistant. Observed segregation of pheno-
types in DH populations were 27R:37S and 69R:57S
for Tremie- and Claire-derived populations, respec-
tively (Table 1). The segregation ratios (v2 = 1.56;
P = 0.211 and v2 = 1.14; P = 0.285; 1df) fit to a
1R:1S segregation indicating that resistance is con-
trolled by one major locus, while occurrence of 14
lines with moderate resistance out of 190 examined
may indicate the presence of another minor gene.
This result is in correspondence with previous
findings for inheritance of SBCMV resistance in
wheat (Bass et al. 2006; Kanyuka et al. 2004; Perovic
et al. 2005).
A disease response score of resistance (R), mod-
erate resistance (MR) or susceptibility (S) was
applied to the 99 wheat accessions inoculated with
SBCMV-infested soils from Wiltshire or Kent, UK in
a controlled temperature glasshouse (Table 2). Most
of the accessions were clearly identified as either R or
S confirming the previous published or unpublished
observations by either us or other research groups.
Several very old wheat accessions (e.g., Turkey) that
are thought to be resistant to soil-borne viral mosaic
disease, however, were classified as MR. Also, quite
unexpectedly, some wheat accessions (e.g., Embrapa
16 and KS92WGRC21) that have been previously
described as resistant to soil-borne viral mosaic
disease in North and/or South America (Barbosa
et al. 2001; Cox et al. 1994) were fully susceptible to
SBCMV in our tests.
Genetic mapping and marker saturation of the
SBCMV resistance locus
Due to the monogenic inheritance of SBCMV
resistance and the four populations analysed, eight
distinct DNA bulks for screening with microsatellite
and AFLP markers were constructed. A total of 162
EST-derived and genomic microsatellite markers and
256 EcoRI ? 3/MseI ? 3 AFLP primer combina-
tions were surveyed to identify polymorphisms
between the resistant and the susceptible DNA bulks
and the six parental lines of the mapping populations.
Initially, a set of 130 genomic GWM (Gatersleben
wheat microsatellites) markers distributed over the 21
chromosomes of the wheat genome were surveyed for
polymorphism. Screening revealed 78 monomorphic
markers, while 52 were polymorphic in at least one of
the four parental combinations used. The initial
screening of DNA bulks detected Xgwm469 as
polymorphic on parents as well as on bulks of the
Tremie-derived population, and Xgwm272 as poly-
morphic on parents as well as on bulks of the Claire-
derived population. Genotyping of the three small
Tremie-derived populations did not detect linkage of
the most prominent Xgwm469 fragment of 170-bp
and the SBCMV resistance locus, while careful
scoring of a second dominantly inherited fragment
of 152-bp revealed co-segregation with the resistance
locus. The Xgwm469 fragment of 152-bp represents a
different locus than the 170-bp fragment that maps to
chromosome 6DS in the ITMI population (Roder
et al. 1998) (Figs. 1, 2). Since Xgwm272 mapped to
the long arm of chromosome 5D (Roder et al. 1998),
Mol Breeding (2009) 23:641–653 647
123

screening was conducted with an additional set of 32
EST-derived and genomic microsatellite markers that
mapped to this chromosome arm.
In all DH populations, genetic linkage between the
resistance locus and marker loci from the long arm of
wheat chromosome 5D was detected (Fig. 1). In total,
12 microsatellite loci were integrated into the genetic
map of the Savannah 9 Claire population and 15
microsatellite loci in the consensus map of the
Tremie-derived populations. Five common marker
loci were in collinear order between these two maps
confirming the common position of the SBCMV
resistance locus in cultivars Tremie and Claire.
Deviation of co-linearity in marker order between
two maps could be explained by the fact that the
Tremie-derived map was the consensus of three
populations. To identify additional closely linked
genomic fragments, resistant and susceptible
SBCMV bulks were screened with 256 EcoRI ?
3/MseI ? 3 AFLP primer combinations. Out of 23
AFLP fragments that were polymorphic between
DNA bulks, only one was mapped to 5DL. However,
in the Savannah 9 Claire population the AFLP
E37M49 was the closest proximal marker to the
SBCMV resistance locus. The closest to the resis-
tance locus marker in both populations was Xgwm469
(Fig. 1a, b). This marker co-segregated with the
resistance locus in the Tremie-derived population,
while in the Claire-derived population it mapped
1 cm distal to the resistance locus. To clarify the
position of Xgwm469-5D in relation to a putative
alternative source of SBCMV resistance this marker
was mapped in a population of 204 DH lines derived
from the Avalon 9 Cadenza cross. In this population
Xgwm469-5D co-segregated with Sbm1 (Fig. 1c).
Since the SBCMV resistance locus in Tremie was
located in the same chromosomal position as the
Sbm1 locus, which provides resistance to SBCMV in
the unrelated cv. Cadenza (Bass et al. 2006), it is
most likely that cultivars Tremie and Claire possess
Sbm1 or a gene allelic to Sbm1. However, since this
has not been proven by tests for allelism, the SBCMV
resistance locus in Tremie and Claire is temporarily
designated as Sbm Tremie/Claire.
For the physical assignment to a particular
segment of chromosome 5D, markers flanking the
resistance locus were tested on a set of 21 nulli-
tetrasomic aneuploid lines and the 5DL5 deletion
line. Markers proximal to the resistance locus, i.e.,
Xgpw343 and Xcfd10, as well as the distal marker
Xwmc765 were mapped to the same deletion bin
5DL5 (Fig. 3). As shown in Fig. 3, missing fragments
of flanking markers in lines 3 and 5, i.e., 5D and
5DL5, respectively, indicate the physical position to
Xcfd1830Xgwm8051Xgwm9314
Xgpw34315
Xcfd1035
E37M4939
SbmClaire48Xgwm46949Xwmc76551
Xbbrc14459
Xwmc44363
Xgwm27267
Xcfe30168
Xgpw2323
71
Xgdm630
Xgpw3433
Xgwm93115
Xgwm21222Xgdm133 Xbarc11024Xcfd1026Xcfd18327Xgwm80528
SbmTremie Xgwm46937Xwmc76539
Xwmc44347
Xgwm565 Xgwm269Xgwm65457
Xgdm1530
Xgwm21228
Xgdm11643
Xbarc11066
Xgwm469Sbm181
Xwmc76582Xbarc144 Xwmc44386
A B CFig. 1 Comparison of
genetic linkage maps for
wheat chromosome 5D
constructed using three
mapping populations:
Savannah 9 Claire (a),
Tremie-derived consensus
(b), Avalon 9 Cadenza (c).
SBCMV resistance locus is
indicated in bold.
Corresponding
microsatellite loci are
connected by thin lines
648 Mol Breeding (2009) 23:641–653
123

the sub-telomeric region of chromosome 5D. There-
fore, the SBCMV resistance locus is located on this
chromosome segment of 5DL.
Diagnostic value of SSR Xgwm469-5D
In the mapping study by Bass et al. (2006), genomic
microsatellite markers Xwmc765 and Xbarc110 were
the closest linked markers to Sbm1, whereas in our
study, screening of additional markers from chromo-
some 5D resulted in the identification of Xgwm469-
5D, Xgwm805, and AFLP E37M49, which are more
closely linked to the SBCMV-resistance locus in the
Savannah 9 Claire and Tremie-derived populations.
Since Xgwm469-5D and Xwmc765 were the two
closest makers to the SBCMV resistance locus in
three populations, the diagnostic value of these
markers was assessed in a collection of SBCMV
resistant and susceptible winter and spring wheat
cultivars originating from different parts of the world
(Table 2). In a set of 99 wheat cultivars Xgwm469-5D
and Xwmc765 detected three and twenty-two different
alleles, respectively. Although, Xwmc765 is located
only 1–3 cm distally from Xgwm469-5D it is of no
180
200
160
140
Cadenza
Claire and Tremie
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 MM
Fig. 2 Molecular analysis of selected wheat genotypes with
the microsatellite marker GWM469 Xgwm469-5DL alleles
linked to SBCMV resistance are indicated by arrows. The
order of genotypes are: 1, Claire; 2, Cadenza; 3, F1 Claire 9
Cadenza; 4, Avalon; 5, Chinese Spring; 6, Ciano F67; 7, Aztek;
8, Tremie; 9, Soisson; 10, Savannah; 11, Axona; 12, Tonic; 13,
Sonora 64; 14, Wasp; 15, Moulin; 16, Flame. M, microSTEP-
20a (700) DNA ladder (Microzone Ltd). Due to extra 7-bp tail,
which was added to the 50 end of the reverse primer the
fragment size of Xgwm469-5D of 152 and 154-bp increased to
159 or 161-bp, respectively
Fig. 3 Assignment of SSR markers Xgpw343, Xcfd10, and
Xwmc765 flanking SbmClaire/Tremie to the chromosome deletion
bin 5DL5. The order of genotypes is: 1, N5A-T5D; 2, N5B-
T5D; 3, N5D-T5A; 4, Chinese Spring; 5, Chinese Spring
deletion line 5DL5, 6, water control. Upper band of Xgpw343maps to chromosome 5D, deletion bin 5DL5, while lower bandmaps to chromosome 5A. Single fragment of Xcfd10 maps to
chromosome 5D, deletion bin 5DL5. Upper fragment of
Xwmc765 maps to chromosome 5B while lowest fragment
maps to chromosome 5D, deletion bin 5DL5. At the left side of
the pictures is DNA ladder pUC19/MspI (Fermentas), while on
the right side of the pictures is the 1 kb DNA ladder
(Fermentas)
c
Mol Breeding (2009) 23:641–653 649
123

diagnostic value for the Sbm1 locus in comparison to
Xgwm469-5D. The Xgwm469-5D allele of 152-bp was
detected in Tremie and Claire as well as in other
resistant wheat cultivars having the resistant cv. Klein
Rendidor in their pedigree. In contrast, facultative
winter wheat cv. Cadenza, spring wheat cv. Tonic and
several related cultivars showed the Xgwm469-5D
allele of 154-bp. In addition, either the 152 or 154-bp
alleles of Xgwm469-5D were detected in a number of
old SBCMV-resistant wheat cultivars of primarily
non-European origin whose relationship to Tonic and
Cadenza or Tremie and Claire is unclear judging from
the available pedigree information. Importantly, in all
susceptible wheat cultivars, a third null allele of this
marker was observed. Therefore, Xgwm469-5D
appears to be an ideal diagnostic marker facilitating
the differentiation between the resistance derived from
cultivars Cadenza/Tonic and Tremie/Claire and the
identification of susceptible cultivars. These results are
in agreement with the pedigree analysis data which
suggest that cv. Cadenza is unrelated to Tremie or
Claire with respect to the donor of resistance. SBCMV
resistance in cultivars Claire and Tremie is most likely
derived from Klein Rendidor, whereas Sbm1 in cv.
Cadenza most likely originates from Sonora 64
(Fig. 4). Therefore, analysis of SBCMV resistance in
cultivars Tremie and Claire resulted in the identifica-
tion of closely linked markers with a high diagnostic
value that will facilitate efficient marker based selec-
tion procedures for SBCMV-resistance in wheat
breeding programs.
Discussion
It was suggested in earlier genetic studies from the
USA (Merkle and Smith 1983; Modawi et al. 1982)
that resistance to SBWMV in many US wheat
Axona----------Spring; null allele
Tonic------------Spring; 154bp
Maris Dove---
Hpg 522/66
H8810-47
Cadenza -------------Spring/Winter; 154bp
Heines Kolben
Sonora 64 Winter; 154bp
CardinalNo data
2772Mgh60-273
Parlo
Heines-Koga 2--
Raeckes White Chaff
Garnet
Heines Kolben
Cb-306-y-70 -------
Yecora 70 ------No data
Maris Hobbit (sib)
Moulin ------------Winter; 152bp
Maris Widgeon
Sib-32 No data
Tremie ----------Winter; 152bp
Siete Cerros 66
Klein RendidorSpring; 152bp
Sonora 64Winter; 154bp
Ciano F67Spring; null allele
Ciano F67Spring; null allele
Taurus
Flame ---Winter; 152bp
Waspnull allele
Moulin ---------Winter; 152bp
Claire -------------Winter; 152bp
Cb-306-y-70 -------
Yecora 70 ------No data
Maris Hobbit (sib)
Maris Widgeon
Ciano F67Spring; null allele
Siete Cerros 66
Klein RendidorSpring; 152bp
Sonora 64Winter; 154bp
Ciano F67Spring; null allele
Fig. 4 Pedigree
relationships of SBCMV
resistant varieties with
assignment of Xgwm469-
5DL donor alleles
650 Mol Breeding (2009) 23:641–653
123

cultivars is derived from the following old cultivars:
(1) the Argentinean cv. Klein Rendidor (this cultivar
is also an ancestor of European SBCMV-resistant
cultivars Tremie and Claire), (2) the US cv. Turkey,
and (3) the Ukrainian cv. Odesskaja 51. Therefore,
the number of detected alleles for Xgwm469-5D is
rather low for such a diverse set of germplasm tested,
but may be explained by the in general low diversity
of the D genome (Balfourier et al. 2007; Wicker et al.
2009). Interestingly, however, Xwmc765 displayed a
much higher allelic diversity (Table 2).
The 152 and 154-bp alleles were linked to
resistance and the null allele was linked to suscep-
tibility in all genotypes tested, suggesting that the
SBCMV-resistance existing in the wheat germplasm
tested may have been derived from two separate
sources. However, further studies (e.g., an allelism
test or the isolation of the SBCMV resistance locus),
will be required to clarify whether Sbm1 in cv.
Cadenza is allelic to the SBCMV resistance locus in
cultivars Tremie and Claire. In addition, several
genotypes with intermediate phenotypes were iden-
tified in the studied collection and very rarely also
within the segregating populations. Occurrence of
intermediate phenotypes after visual scoring suggests
the presence of a major gene as well as other genes
with minor effects contributing to the resistance,
possibly acting as suppressors of resistance. How-
ever, it is also important to mention that the
‘translocation resistance’ is highly influenced by the
environment and the root-shoot translocation block
can be occasionally overcome by the virus (Lyons
et al. 2008; Myers et al. 1993). The exact factors
contributing to the breakdown of translocation resis-
tance are as yet unknown. However, it is conceivable
that the translocation resistance can be overcome
either under high inoculum pressure (i.e., when an
excessively high number of viruliferous P. graminis
zoospores is present close to the plant’s root system)
or when the plant is suffering from other biotic (i.e.,
infection by other phytopathogens) or abiotic stress
(i.e., freezing).
It is interesting to note that although genetically
quite diverse wheat germplasm was tested in this
study, that only three different alleles were detected
at the Xgwm469-5D locus. This number is much
lower than that found in other wheat genotyping
studies with microsatellite markers (Balfourier et al.
2007; Huang et al. 2002; Roder et al. 1998). The low
level of polymorphism at the Xgwm469-5D locus
may be because the wheat D-genome is known to
exhibit a generally low level of polymorphism
(Balfourier et al. 2007; Wicker et al. 2009). It is also
possible that this region on 5DL contains some
(unknown) genes that are essential for crop growth,
development or performance and, therefore, is sub-
jected to a strong purifying selection pressure. A low
level of polymorphism was also detected in barley at
the Bmac29 locus which is closely linked to the
bymovirus resistance locus rym4/rym5 (Graner et al.
1999) and used in marker assisted selection (MAS)
since 1999 (Ordon et al. 2004). Such microsatellite
loci, which are closely linked to the genes of interest
and display low allelic diversity, are best suited for
MAS procedures as they can be reliably used in
breeding programs involving many diverse crosses.
In this study, we identified a previously unmapped
microsatellite locus Xgwm469 as the most closely
linked to the SBCMV resistance gene on 5DL. Here
we also provide evidence that GWM469 is well
suited for use in MAS for reliable selection of
resistant genotypes at early developmental stages and
on a single plant level. This is of special importance
as in many European countries uniformly infested
fields required for reliable resistance screening of
breeding materials are not available. Therefore, the
availability of the GWM469 marker will facilitate
quarantine breeding, i.e., the development of resistant
cultivars in the absence of the virus. This is especially
relevant due to the risk of rapid spreading of these
soil-borne viruses, as seen previously for the soil-
borne viruses of barley (Barley yellow mosaic virus
and Barley mild mosaic virus) in Germany (Huth
1989). However, GWM469 also has some drawbacks.
First, in contrast to many microsatellites, it is a
dominant marker that does not allow the selection of
homozygous resistant genotypes in standard (i.e., not
doubled haploid) segregating heterozygous popula-
tions. In future, this could be of minor importance
when more European wheat breeders will use
doubled haploid populations in which due to homo-
zygosity dominant markers are as informative as co-
dominant ones. Also, null alleles derived from
susceptible lines cannot be distinguished from an
absence of amplification due to a PCR failure. One
way to overcome this would be to use an internal
standard such as an additional monomorphic marker.
In our case, the major fragment of Xgwm469 of
Mol Breeding (2009) 23:641–653 651
123

170-bp is well suited as an internal control of the
PCR reaction. However, the best marker remains the
gene itself since even when the marker is tightly
linked to the gene, the recombination may still occur.
Therefore, the flanking markers developed in this
study may serve as the starting point for the
construction of a high resolution map finally leading
to the map based isolation of Sbm1. When the
resistance gene sequence is revealed it should then be
possible to develop robust allele-specific markers for
reliable and errorless identification of resistant geno-
types in the laboratory.
Acknowledgments The work is supported by a grant in the
European Community’s Sixth Framework Program
WHEATPROTECT (EU contract number COOP-CT-2004-
512703). These results and this publication reflect only the
author’s view and EU is not liable for any use that may be made
of the information contained in this article. The authors like to
thank Prof. B. Gill (Kansas State University, USA) for kindly
providing seeds of NT and 5DL5 Chinese Spring lines, Dr. L.
Sayers (John Innes Centre, UK) for providing Avalon 9
Cadenza DH mapping population, and various European wheat
breeding companies as well as Dr. H. E. Bockelman (National
Small Grains Collection, USDA Aberdeen, USA) and Dr. R.
Singh (CIMMYT, Mexico) for providing wheat genotypes used
in disease resistance tests and/or molecular genetic analyses. We
are also grateful to Dr. R. Bayles (NIAB, Cambridge, UK) and
Mr. M. Oatley (Paxcroft Farms, Trowbridge, UK) for letting us
sample SBCMV-infested soil in Wiltshire site. We would also
like to thank Ms. B. Knupfer and M. Weilepp for excellent
technical assistance. Rothamsted Research receives grant-aided
support from the Biotechnology and Biological Sciences
Research Council of the United Kingdom.
References
Balfourier F, Roussel V, Strelchenko P, Exbrayat F, Sourdille
P, Boutet G, Koenig J, Ravel C, Mitrofanova O, Beckert
M, Charmet G (2007) A worldwide bread wheat core-
collection arrayed in a 384-well plate. Theor Appl Genet
114:1265–1275. doi:10.1007/s00122-007-0517-1
Barbosa MM, Goulart LR, Prestes AM, Juliatti FC (2001)
Genetic control of resistance to soil-borne wheat mosaic
virus in Brazilian cultivars of Triticum aestivum L. Thell.
Euphytica 122:417–422. doi:10.1023/A:1012937116394
Bass C, Hendley R, Adams MJ, Hammond-Kosack KE,
Kanyuka K (2006) The Sbm1 locus conferring resistance
to Soil-borne cereal mosaic virus maps to a gene-rich
region on 5DL in wheat. Genome 49:1140–1148. doi:
10.1139/G06-064
Bayles RA, Napier B (2002) Tolerance of wheat varieties to
Soil-borne wheat mosaic virus (SBWMV). HGCA project
report No. 278. Home-Grown Cereals Authority, London
Brakke MK, Langenberg WG (1988) Experiences with soil-
borne wheat mosaic virus in North America and elsewhere.
In: Cooper JI, Asher MJC (eds) Developments in applied
biology II. Viruses with Fungal Vectors. Association of
Applied Biologists, Wellesbourne, pp 183–202
Brownstein MJ, Carpten JD, Smith JR (1996) Modulation of
non-templated nucleotide addition by Taq DNA poly-
merase: primer modifications that facilitate genotyping.
Biotechniques 20:1004–1010
Budge G, Henry CM (2002) Assessing resistance of UK winter
wheat varieties to Soil-borne wheat mosaic virus and
Wheat spindle streak mosaic virus. HGCA project report,
2002, No. 293. Home-Grown Cereals Authority, London
Clark MF, Adams AN (1977) Characteristics of the microplate
method of enzyme-linked immunosorbent assay for the
detection of plant viruses. J Gen Virol 34:475–483. doi:
10.1099/0022-1317-34-3-475
Clover GRG, Ratti C, Henry CM (2001) Molecular character-
ization and detection of European isolates of Soil-borne
wheat mosaic virus. Plant Pathol 50:761–767. doi:10.1046/
j.1365-3059.2001.00634.x
Cox TS, Sorrells ME, Bergstrom GC, Sears RG, Gill BS,
Walsh EJ, Leath S, Murphy JP (1994) Registration of
KS92WGRC21 and KS92WGRC22 hard red winter wheat
germplasms resistant to wheat streak mosaic virus and
powdery mildew. Crop Sci 34:546
Devaux P, Pickering R (2005) Haploids in the improvement of
Poaceae. In: Palmer D, Keller W, Kasha K (eds) Haploids
in crop improvement II. Springer, Heidelberg, pp 215–242
Endo TR, Gill BS (1996) The deletion stocks of common wheat. J
Hered 87:295–307
Graner A, Jahoor A, Schondelmaier J, Siedler H, Pillen K,
Fischbeck G, Wenzel G, Herrmann RG (1991) Construction
of an RFLP map in barley. Theor Appl Genet 83:250–256.
doi:10.1007/BF00226259
Graner A, Streng S, Kellermann A, Schiemann A, Bauer E,
Waugh R, Pellio B, Ordon F (1999) Molecular mapping and
fine structure of the rym5 locus encoding resistance to dif-
ferent strains of the barley yellow mosaic virus complex.
Theor Appl Genet 98:285–290. doi:10.1007/s00122005
1070
Hariri D, Courtillot M, Zaoui P, Lapierre H (1987) Multipli-
cation of Soil-borne wheat mosaic virus (SBWMV) in
wheat roots infected by a soil carrying SBWMV and
Wheat yellow mosaic virus (WYMV). Agronomie 7:789–
796. doi:10.1051/agro:19871005
Huang XQ, Borner A, Borner A, Roder MS, Ganal MW (2002)
Assessing genetic diversity of wheat (Triticum aestivum L.)
germplasm using microsatellite markers. Theor Appl Genet
105:699–707. doi:10.1007/s00122-002-0959-4
Huth W (1989) Ein weiterer stamm des barley yellow mosaic
virus in der bundesrepublik Deutschland. Nachrichtenbl
Deut Pflanzenschutzd 40:49–55
Kanyuka K, Lovell D, Mitrofanova OP, Hammond-Kosack K,
Adams MJ (2004) A controlled environment test for resis-
tance to Soil-borne cereal mosaic virus and its use to
determine the mode of inheritance of the resistance in the
UK wheat variety Cadenza, and to screen diverse Triticum
monococcum genotypes for potential sources of improved
disease resistance. Plant Pathol 53:154–160. doi:10.1111/
j.0032-0862.2004.01000.x
Kastirr U, Wortmann H, Ehrig F (2006) Untersuchungen zum
Infektionsverlauf und zur biologischen differenzierung
652 Mol Breeding (2009) 23:641–653
123

von bodenburtigen viren in roggen, triticale und weizen.
Gesund Pflanz 58:231–238. doi:10.1007/s10343-006-01
36-9
Koenig R, Huth W (2000) Soil-borne rye mosaic virus and
European wheat mosaic virus : two names for a furovirus
with variable genome properties which is widely distrib-
uted in several cereal crops in Europe. Arch Virol 145:
689–697. doi:10.1007/s007050050663
Lyons R, Hammnd-Kosack KE, Kanyuka K (2008) Identifi-
cation and characterization of a novel efficient resistance
response to the furoviruses SBWMV and SBCMV in
barley. Mol Plant Microbe Interact 21:1193–1204. doi:
10.1094/MPMI-21-9-1193
McKinney HH (1923) Investigation of the rosette disease of
wheat and its control. J Agric Res 23:771–800
Merkle OG, Smith EL (1983) Inheritance of resistance to soil-
borne mosaic in wheat. Crop Sci 23:1075–1076
Michelmore RW, Paran I, Kesseli RV (1991) Identification of
markers linked to disease resistant genes by bulk segre-
gant analysis: a rapid method to detect markers in specific
genomic regions using segregating populations. Proc Natl
Acad Sci USA 88:9828–9832. doi:10.1073/pnas.88.21.
9828
Modawi RS, Heyne EG, Brunetta D, Willis WG (1982) Genetic
studies of field reaction to wheat soil-borne mosaic virus.
Plant Dis 66:1183–1184
Myers LD, Sherwood JL, Siegerist WC, Hunger RM (1993)
Temperature-influenced virus movement in expression of
resistance to Soilborne wheat mosaic virus in hard red
winter wheat (Triticum aestivum). Phytopathology 83:
548–551. doi:10.1094/Phyto-83-548
Ordon F, Friedt W, Scheurer K, Pellio B, Werner K, Neuhaus G,
Huth W, Habekuss A, Graner A (2004) Molecular markers
in breeding for virus resistance in barley. J Appl Genet 45:
145–159
Perovic D, Weyen J, Schondelmaier J, Forster J, Devaux P,
Hariri D, Feuerhelm D, Stein N, Graner A, Ordon F
(2005) Linkage mapping and transcription profiling of
resistance to soil-borne viruses in hexaploid wheat (Trit-icum vulgare sp. aestivum). Parasitica 61:79–84
Proeseler G, Stanarius A (1983) Nachweis des weizenspin-
delstrichelmosaik-virus (wheat spindle streak mosaic
virus) in der DDR. Arch Phytopathol Pflanzenschutz 19:
345–349
Rao AS, Brakke MK (1969) Relation of soil-borne wheat
mosaic virus and its fungal vector, Polymyxa graminis.
Phytopathology 59:581–587
Roder MS, Korzun V, Wendehake K, Plaschke J, Tixier MH,
Leroy P, Ganal MW (1998) A microsatellite map of
wheat. Genetics 149:2007–2023
Saito Y, Takanashi K, Iwata Y, Okamoto H (1964) Studies on
soil-borne virus disease of wheat and barley. I. Several
properties of viruses. Bull Natl Inst Agr Sci (Japan) Ser C
17:1–19
Sears ER (1954) The aneuploids of common wheat. Univ Mo
Agric Exp Stn Res Bull 572:1–58
Sears ER (1966) Nullisomic-tetrasomic combinations in hexa-
ploid wheat. In: Riley R, Lewis KR (eds) Chromosome
manipulation and plant genetics. Oliver & Boyd, Edinburgh,
pp 29–45
Somers DJ, Peter I, Edwards K (2004) A high-density microsat-
ellite consensus map for bread wheat (Triticum aestivum L.).
Theor Appl Genet 109:1105–1114. doi:10.1007/s00122-
004-1740-7
Stam P (1993) Construction of integrated genetic linkage maps
by means of a computer package: JOINMAP. Plant J 3:
739–744
Torrance L, Koenig R (2004) Furovirus. In: Fauquet CM,
Mayo MA, Maniloff J, Desselberger U, Ball LA (eds)
Virus taxonomy, classification and nomenclature of viru-
ses. Elsevier, Amsterdam, pp 1027–1032
Werner JE, Endo TR, Gill BS (1992) Towards a cytogeneti-
cally based physical map of the wheat genome. Proc Natl
Acad Sci USA 89:11307–11311. doi:10.1073/pnas.89.23.
11307
Wicker T, Krattinger S, Lagudah ES, Komatsuda T, Pourkhei-
randish M, Matsumoto T, Cloutier S, Kanamori H, Sato K,
Perovic D, Stein N, Keller B (2009) Analysis of intraspecies
diversity in wheat and barley genomes identifies break-
points of ancient haplotypes and provides insight in the
structure of diploid and hexaploid Triticeae gene pools
wheat. Plant Physiol 149:258–270. doi:10.1104/pp.108.
129734
Zhang LY, Bernard M, Leroy P, Feuillet C, Sourdille P (2005)
High transferability of bread wheat EST-derived SSRs to
other cereals. Theor Appl Genet 111:677–687. doi:10.1007/
s00122-005-2041-5
Mol Breeding (2009) 23:641–653 653
123




















