
Male involvement in fertility and factors affecting semen quality ...
6
Key words: breeding soundness evaluation, bull fertility, scrotal/testicu- lar thermoregulation, sperm defects Introduction There is no doubt that there are substantial differences in fertility among individual bulls. In a recent review (Flowers, 2013), normal ranges in re- productive performance from entire herds were compared to those reported for males. For beef cattle, maximum fertility was similar for herds versus males. However, since minimum fertility was less for the latter, it was con- cluded that beef bulls contribute significantly to fertility failures, as bulls with reduced fertility are frequently used. However, for dairy cattle, there were similar minimums for herds and bulls, yet maximum fertility for bulls exceeded that of the herd. Therefore, it was concluded that reproductive failure in dairy cattle was due more to the cow than to the bull. In general, fertility is more important in an individual bull than an indi- vidual cow, as one bull may be used to breed up to 40 females with natural service, or potentially hundreds of thousands via artificial insemination. Although 20 to 40% of bulls may have reduced fertility, few are complete- ly sterile (Coulter and Kastelic, 1999). Subfertile bulls delay conception, prolong the calving season, reduce calf weaning weights, and increase the numbers of females culled, thereby resulting in economic losses and threat- ening sustainability of a livestock operation. Furthermore, infertile bulls can have adverse effects of animal welfare, due to repeated breedings and delayed calving. Whereas some management practices such as multiple- sire breeding groups, low breeding pressure, and extended breeding sea- sons make identification of subfertile bulls difficult, other strategies such as single-sire mating groups, short breeding seasons, and artificial insemina- tion highlight the need to critically evaluate bull fertility. Evaluation of Breeding Soundness The two general methods of evaluating the breeding soundness poten- tial of bulls are either breeding a large number of normal, fertile females and determining pregnancy/calving rates or conducting a breeding sound- ness evaluation. Although a breeding trial is the ultimate test of fertility, it is expensive, particularly if reproductive performance is poor. There- fore, it is strongly recommended to conduct a standard breeding soundness evaluation before the breeding season. These evaluations identify bulls with substantial deficits in fertility, but they do not consistently identify subfertile bulls. In that regard, the use of frozen-thawed semen from bulls in commercial artificial insemination centers that meets minimum quality standards can result in pregnancy rates that differ by 20 to 25% points (Larson and Miller, 2000), as standard semen end points identify grossly abnormal semen but do not consistently identify subfertile bulls with ap- parently normal semen (Gadea et al., 2004). Since bull fertility is influ- enced by a wide range of factors, no single diagnostic test can accurately predict fertility, although an appropriate combination of tests can be more informative (Kastelic and Thundathil, 2008). Based on work in boars, per- haps a means of assessing the post-insemination sperm-oviduct interaction would substantially increase the ability to predict fertility (Flowers, 2013). Seminal plasma proteins apparently have potential as fertility predictors (Flowers, 2013), and more than 400 proteins have been identified in a bull sperm plasma membrane fraction (Byrne et al., 2012). The standards of the Society for Theriogenology (www.therio.org) are intended to assess the likelihood of a bull establishing pregnancy in ≥ 25 healthy, cycling females in a 65- to 70-day breeding season. The classi- fication is based on the bull meeting minimum standards for health and structural soundness, sperm motility ( ≥ 30% progressively motile sperm), sperm morphology ( ≥ 70% normal and ≤ 20% defective heads), and scro- tal circumference. The latter is highly correlated with paired testis weight, which is, in turn, positively correlated with daily sperm production and semen quality (reviewed by Barth, 2007). In that regard, bulls with larger testes produce more and better quality sperm, in general. Furthermore, Male involvement in fertility and factors affecting semen quality in bulls John P. Kastelic Department of Production Animal Health, University of Calgary, Faculty of Veterinary Medicine, Calgary, AB, Canada © Kastelic doi:10.2527/af.2013-0029 Implications • Fertility varies substantially among bulls. In general, methods to predict fertility are better for identifying bulls with low fertility than for ranking bulls with good to excellent fertility. • Compensable sperm abnormalities can be overcome by increas- ing the dose used for artificial insemination; these are attributed to sperm reaching and penetrating the zona pellucida. In contrast, increasing the insemination dose does not improve fertility for uncompensable defects, implying that the sperm are able to cause fertilization and initiate development, but they do not sustain em- bryogenesis. • Bull testes must be 2 to 6°C cooler than core body temperature for fertile sperm; consequently, increased testicular temperature reduces semen quality. • Increased nutrition before 30 weeks of age increased luteiniz- ing hormone pulse frequency, hastened puberty, and increased testicular size at maturity in bulls. However, attempts to correct nutritional deficiencies present during calfhood by supplemental nutrition later in life were unsuccessful. 20 Animal Frontiers Downloaded from https://academic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Male involvement in fertility and factors affecting semen quality ...

Key words: breeding soundness evaluation, bull fertility, scrotal/testicu-lar thermoregulation, sperm defects
Introduction
There is no doubt that there are substantial differences in fertility among individual bulls. In a recent review (Flowers, 2013), normal ranges in re-productive performance from entire herds were compared to those reported for males. For beef cattle, maximum fertility was similar for herds versus males. However, since minimum fertility was less for the latter, it was con-cluded that beef bulls contribute significantly to fertility failures, as bulls with reduced fertility are frequently used. However, for dairy cattle, there were similar minimums for herds and bulls, yet maximum fertility for bulls exceeded that of the herd. Therefore, it was concluded that reproductive failure in dairy cattle was due more to the cow than to the bull.
In general, fertility is more important in an individual bull than an indi-vidual cow, as one bull may be used to breed up to 40 females with natural service, or potentially hundreds of thousands via artificial insemination. Although 20 to 40% of bulls may have reduced fertility, few are complete-
ly sterile (Coulter and Kastelic, 1999). Subfertile bulls delay conception, prolong the calving season, reduce calf weaning weights, and increase the numbers of females culled, thereby resulting in economic losses and threat-ening sustainability of a livestock operation. Furthermore, infertile bulls can have adverse effects of animal welfare, due to repeated breedings and delayed calving. Whereas some management practices such as multiple-sire breeding groups, low breeding pressure, and extended breeding sea-sons make identification of subfertile bulls difficult, other strategies such as single-sire mating groups, short breeding seasons, and artificial insemina-tion highlight the need to critically evaluate bull fertility.
Evaluation of Breeding Soundness
The two general methods of evaluating the breeding soundness poten-tial of bulls are either breeding a large number of normal, fertile females and determining pregnancy/calving rates or conducting a breeding sound-ness evaluation. Although a breeding trial is the ultimate test of fertility, it is expensive, particularly if reproductive performance is poor. There-fore, it is strongly recommended to conduct a standard breeding soundness evaluation before the breeding season. These evaluations identify bulls with substantial deficits in fertility, but they do not consistently identify subfertile bulls. In that regard, the use of frozen-thawed semen from bulls in commercial artificial insemination centers that meets minimum quality standards can result in pregnancy rates that differ by 20 to 25% points (Larson and Miller, 2000), as standard semen end points identify grossly abnormal semen but do not consistently identify subfertile bulls with ap-parently normal semen (Gadea et al., 2004). Since bull fertility is influ-enced by a wide range of factors, no single diagnostic test can accurately predict fertility, although an appropriate combination of tests can be more informative (Kastelic and Thundathil, 2008). Based on work in boars, per-haps a means of assessing the post-insemination sperm-oviduct interaction would substantially increase the ability to predict fertility (Flowers, 2013). Seminal plasma proteins apparently have potential as fertility predictors (Flowers, 2013), and more than 400 proteins have been identified in a bull sperm plasma membrane fraction (Byrne et al., 2012).
The standards of the Society for Theriogenology (www.therio.org) are intended to assess the likelihood of a bull establishing pregnancy in ≥ 25 healthy, cycling females in a 65- to 70-day breeding season. The classi-fication is based on the bull meeting minimum standards for health and structural soundness, sperm motility ( ≥ 30% progressively motile sperm), sperm morphology ( ≥ 70% normal and ≤ 20% defective heads), and scro-tal circumference. The latter is highly correlated with paired testis weight, which is, in turn, positively correlated with daily sperm production and semen quality (reviewed by Barth, 2007). In that regard, bulls with larger testes produce more and better quality sperm, in general. Furthermore,
Male involvement in fertility and factors affecting semen quality in bullsJohn P. Kastelic
Department of Production Animal Health, University of Calgary, Faculty of Veterinary Medicine, Calgary, AB, Canada
© Kastelicdoi:10.2527/af.2013-0029
Implications
• Fertility varies substantially among bulls. In general, methods to predict fertility are better for identifying bulls with low fertility than for ranking bulls with good to excellent fertility.
• Compensable sperm abnormalities can be overcome by increas-ing the dose used for artificial insemination; these are attributed to sperm reaching and penetrating the zona pellucida. In contrast, increasing the insemination dose does not improve fertility for uncompensable defects, implying that the sperm are able to cause fertilization and initiate development, but they do not sustain em-bryogenesis.
• Bull testes must be 2 to 6°C cooler than core body temperature for fertile sperm; consequently, increased testicular temperature reduces semen quality.
• Increased nutrition before 30 weeks of age increased luteiniz-ing hormone pulse frequency, hastened puberty, and increased testicular size at maturity in bulls. However, attempts to correct nutritional deficiencies present during calfhood by supplemental nutrition later in life were unsuccessful.
20 Animal Frontiers
Dow
nloaded from https://academ
ic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022

bulls with large testes have female siblings and daughters with earlier pu-berty and better fertility (reviewed by Barth, 2007). Since the heritability of scrotal circumference in young bulls is ~0.5, it responds well to selection (reviewed by Barth, 2007).
Compensable versus Uncompensable Sperm Defects
There are several systems of classifying sperm abnormalities, and di-viding them into compensable and uncompensable defects is a practical approach that is gaining popularity (reviewed in Amann and DeJarnette, 2012). Compensable abnormalities can be overcome by increasing the dose used for artificial insemination. Therefore, these abnormalities are generally believed to be associated with sperm reaching and penetrating the zona pellucida, and as such, are associated with sperm viability mea-sures such as motility, acrosome integrity, and cell membrane integrity. For example, sperm with knobbed acrosomes or bent tails are unlikely to fertil-ize an ovum. In contrast, uncompensable defects are defined as fertility of a male or an insemination dose that does not improve with an increased num-ber of sperm, implying that the sperm are able to cause fertilization and initiate development but are unable to sustain embryogenesis, with most failures occurring before day 8 post breeding. Chromosomal abnormali-ties, protamine status, and perhaps abnormalities of mRNA are believed to be uncompensable defects, as these sperm would be expected to initiate fertilization but not sustain development. Furthermore, laboratory process-ing such as flow-sorting sperm for sex selection can increase the number of uncompensable sperm defects (DeJarnette, 2005).
The “threshold” in sperm numbers is the limit beyond which further increases in sperm numbers per insemination dose fails to further increase fertility, which can be due to reaching the limit of female fertility, or a value below the level of female fertility, due to the presence of uncom-pensable sperm defects (Amann and DeJarnette, 2012). Consequently, the threshold for a particular bull or ejaculate can vary widely and depends on the severity and ratio of compensable and uncompensable sperm defects. Males that have a prolonged linear increase in fertility with increasing numbers of sperm are considered to have primarily compensable sperm defects, whereas those that quickly reach a plateau are considered to have
a large proportion of uncompensable sperm defects. Regardless, it appears that most fertility curves reach their threshold when 70% of sperm have a compensable trait (Flowers, 2013).
Compensable sperm quality attributes are those that affect the ability of a sperm to access and fertilize an ovum. Therefore, measurements of motility, viability, and membrane function are commonly used to estimate compensable sperm quality. However, given that they do not assess un-compensable semen traits, they are most appropriate to determine the num-ber of sperm per insemination dose to reach maximal fertility rather than to predict absolute fertility (Amann and DeJarnette, 2012).
Visual assessment of sperm motility is quick and inexpensive. How-ever, it is highly subjective and often has limited repeatability (DeJarnette, 2005). There are numerous computer-assisted sperm analyzer (CASA) machines that are commercially available. These provide an objective and much more repeatable method of assessing sperm motility, yielding numer-ous end points that characterize sperm motion (Kathiravan et al., 2011).
Acrosomal integrity and sperm viability are commonly measured to predict fertility. The acrosome can be stained using a number of live/dead stains such as eosin/nigrosin or assessed without staining with appropriate optical systems such as differential interference contrast microscopy. Fur-thermore, there are several stains that enable assessments to be made with flow cytometry, allowing rapid and objective evaluation of large numbers of sperm. In addition, sperm from neat semen often is subjected to ad-ditional manipulations that provide more information with regards to its fertilizing potential. Frozen-thawed sperm are often maintained at 37°C for 2 to 4 hours after thawing and then evaluated; this mimics exposure to the female reproductive tract and facilitates detection of latent sperm ab-normalities that may not be apparent immediately post-thaw (DeJarnette, 2005). Furthermore, the hypo-osmotic swelling test (HOST) can be used to determine membrane viability.
Testicular Temperature
It is well established that bull testes must be 2 to 6°C cooler than core body temperature for fertile sperm to be produced. Consequently, increased testicular temperature, regardless of the cause, reduces semen quality (Waites, 1970). Furthermore, increased testicular temperature is a common underlying cause of infertility in bulls (Kastelic, 2013).
Flickr/tausend und eins, fotografik
October 2013, Vol. 3, No. 4 21
Dow
nloaded from https://academ
ic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022
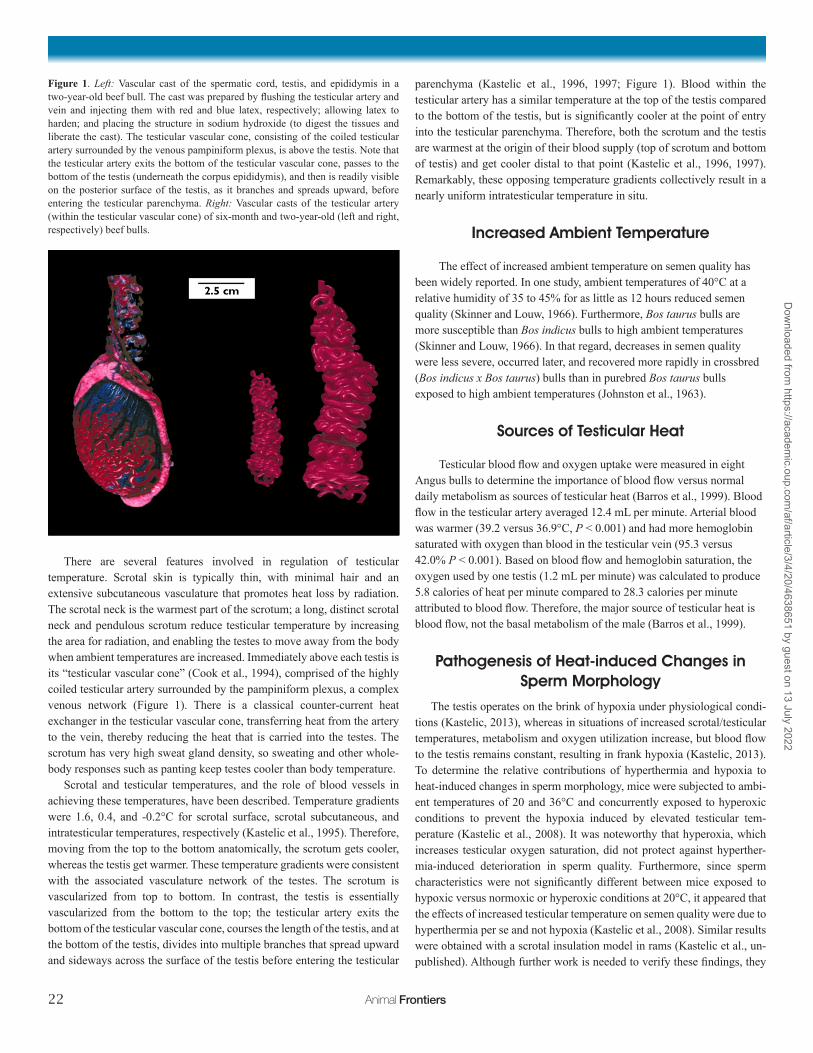
There are several features involved in regulation of testicular temperature. Scrotal skin is typically thin, with minimal hair and an extensive subcutaneous vasculature that promotes heat loss by radiation. The scrotal neck is the warmest part of the scrotum; a long, distinct scrotal neck and pendulous scrotum reduce testicular temperature by increasing the area for radiation, and enabling the testes to move away from the body when ambient temperatures are increased. Immediately above each testis is its “testicular vascular cone” (Cook et al., 1994), comprised of the highly coiled testicular artery surrounded by the pampiniform plexus, a complex venous network (Figure 1). There is a classical counter-current heat exchanger in the testicular vascular cone, transferring heat from the artery to the vein, thereby reducing the heat that is carried into the testes. The scrotum has very high sweat gland density, so sweating and other whole-body responses such as panting keep testes cooler than body temperature.
Scrotal and testicular temperatures, and the role of blood vessels in achieving these temperatures, have been described. Temperature gradients were 1.6, 0.4, and -0.2°C for scrotal surface, scrotal subcutaneous, and intratesticular temperatures, respectively (Kastelic et al., 1995). Therefore, moving from the top to the bottom anatomically, the scrotum gets cooler, whereas the testis get warmer. These temperature gradients were consistent with the associated vasculature network of the testes. The scrotum is vascularized from top to bottom. In contrast, the testis is essentially vascularized from the bottom to the top; the testicular artery exits the bottom of the testicular vascular cone, courses the length of the testis, and at the bottom of the testis, divides into multiple branches that spread upward and sideways across the surface of the testis before entering the testicular
parenchyma (Kastelic et al., 1996, 1997; Figure 1). Blood within the testicular artery has a similar temperature at the top of the testis compared to the bottom of the testis, but is significantly cooler at the point of entry into the testicular parenchyma. Therefore, both the scrotum and the testis are warmest at the origin of their blood supply (top of scrotum and bottom of testis) and get cooler distal to that point (Kastelic et al., 1996, 1997). Remarkably, these opposing temperature gradients collectively result in a nearly uniform intratesticular temperature in situ.
Increased Ambient Temperature
The effect of increased ambient temperature on semen quality has been widely reported. In one study, ambient temperatures of 40°C at a relative humidity of 35 to 45% for as little as 12 hours reduced semen quality (Skinner and Louw, 1966). Furthermore, Bos taurus bulls are more susceptible than Bos indicus bulls to high ambient temperatures (Skinner and Louw, 1966). In that regard, decreases in semen quality were less severe, occurred later, and recovered more rapidly in crossbred (Bos indicus x Bos taurus) bulls than in purebred Bos taurus bulls exposed to high ambient temperatures (Johnston et al., 1963).
Sources of Testicular Heat
Testicular blood flow and oxygen uptake were measured in eight Angus bulls to determine the importance of blood flow versus normal daily metabolism as sources of testicular heat (Barros et al., 1999). Blood flow in the testicular artery averaged 12.4 mL per minute. Arterial blood was warmer (39.2 versus 36.9°C, P < 0.001) and had more hemoglobin saturated with oxygen than blood in the testicular vein (95.3 versus 42.0% P < 0.001). Based on blood flow and hemoglobin saturation, the oxygen used by one testis (1.2 mL per minute) was calculated to produce 5.8 calories of heat per minute compared to 28.3 calories per minute attributed to blood flow. Therefore, the major source of testicular heat is blood flow, not the basal metabolism of the male (Barros et al., 1999).
Pathogenesis of Heat-induced Changes in Sperm Morphology
The testis operates on the brink of hypoxia under physiological condi-tions (Kastelic, 2013), whereas in situations of increased scrotal/testicular temperatures, metabolism and oxygen utilization increase, but blood flow to the testis remains constant, resulting in frank hypoxia (Kastelic, 2013). To determine the relative contributions of hyperthermia and hypoxia to heat-induced changes in sperm morphology, mice were subjected to ambi-ent temperatures of 20 and 36°C and concurrently exposed to hyperoxic conditions to prevent the hypoxia induced by elevated testicular tem-perature (Kastelic et al., 2008). It was noteworthy that hyperoxia, which increases testicular oxygen saturation, did not protect against hyperther-mia-induced deterioration in sperm quality. Furthermore, since sperm characteristics were not significantly different between mice exposed to hypoxic versus normoxic or hyperoxic conditions at 20°C, it appeared that the effects of increased testicular temperature on semen quality were due to hyperthermia per se and not hypoxia (Kastelic et al., 2008). Similar results were obtained with a scrotal insulation model in rams (Kastelic et al., un-published). Although further work is needed to verify these findings, they
Figure 1. Left: Vascular cast of the spermatic cord, testis, and epididymis in a two-year-old beef bull. The cast was prepared by flushing the testicular artery and vein and injecting them with red and blue latex, respectively; allowing latex to harden; and placing the structure in sodium hydroxide (to digest the tissues and liberate the cast). The testicular vascular cone, consisting of the coiled testicular artery surrounded by the venous pampiniform plexus, is above the testis. Note that the testicular artery exits the bottom of the testicular vascular cone, passes to the bottom of the testis (underneath the corpus epididymis), and then is readily visible on the posterior surface of the testis, as it branches and spreads upward, before entering the testicular parenchyma. Right: Vascular casts of the testicular artery (within the testicular vascular cone) of six-month and two-year-old (left and right, respectively) beef bulls.
22 Animal Frontiers
Dow
nloaded from https://academ
ic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022
do call into question the long-standing dogma that the effects of increased testicular temperature on sperm morphology are due to hypoxia.
Effects of Increased Temperature on Testicular Cells
Although heating seems to affect Sertoli and Leydig cell function, germ cells are the most sensitive to heat (Waites and Setchell, 1990). All stages of spermatogenesis are susceptible, with the degree of damage related to the extent and duration of the increased temperature (Waites and Setchell, 1990). Spermatocytes in meiotic prophase are killed by heat, whereas sper-matozoa that are more mature usually have metabolic and structural abnor-malities (Setchell et al., 1971). Heating the testis usually decreases the pro-portion of progressively motile and live sperm and increases the incidence of morphologically abnormal sperm, especially those with defective heads (Barth and Oko, 1989). Although there is considerable variation among bulls in the nature and proportion of defective sperm, the order of appear-ance of specific defects is relatively consistent (Barth and Bowman, 1994). Unless spermatogonia are affected, the interval from cessation of heating to restoration of normal spermatozoa in the ejaculate corresponds to the interval from the beginning of differentiation to ejaculation (Waites and Setchell, 1990). Even though sperm morphology has returned to normal, their utilization may decrease fertilization rates and increase the incidence of embryonic death (Burfening and Ulberg, 1968).
Summary of Increased Testicular Temperature
When scrotal/testicular temperature is increased, sperm morphology is generally unaffected initially (for an interval corresponding to epididymal
transit time) but subsequently declines (Barth and Oko, 1989). However, in some studies, sperm in the epididymis at the time of scrotal heating were morphologically abnormal when collected soon after heating (Wildeus and Entwistle, 1983). In another study, changes in these sperm were manifest only after they were frozen, thawed, and incubated. Sperm morphology usually returns to pre-treatment values within approximately six weeks af-ter the thermal insult (Vogler et al., 1991). However, a prolonged or severe increase in testicular temperature will increase the interval for recovery. In general, the decrease in semen quality following increased testicular temperature is related to the severity and the duration of the thermal insult.
Evaluation of Scrotal Surface Temperature with Infrared Thermography
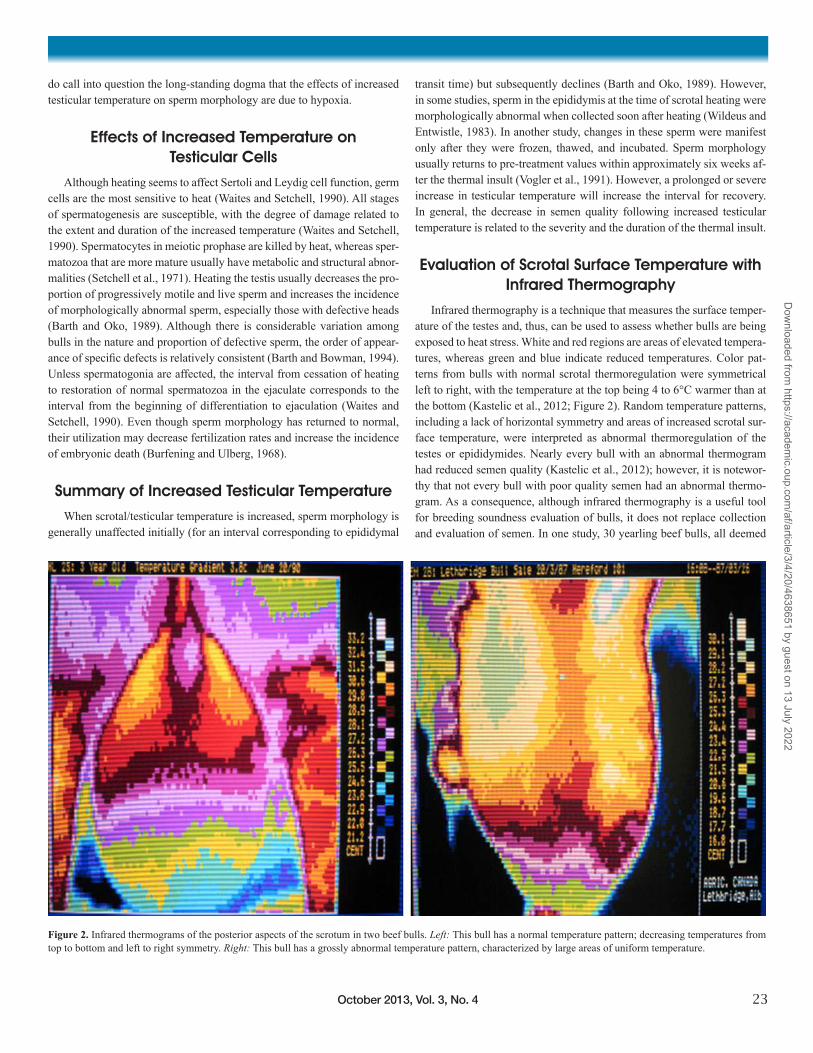
Infrared thermography is a technique that measures the surface temper-ature of the testes and, thus, can be used to assess whether bulls are being exposed to heat stress. White and red regions are areas of elevated tempera-tures, whereas green and blue indicate reduced temperatures. Color pat-terns from bulls with normal scrotal thermoregulation were symmetrical left to right, with the temperature at the top being 4 to 6°C warmer than at the bottom (Kastelic et al., 2012; Figure 2). Random temperature patterns, including a lack of horizontal symmetry and areas of increased scrotal sur-face temperature, were interpreted as abnormal thermoregulation of the testes or epididymides. Nearly every bull with an abnormal thermogram had reduced semen quality (Kastelic et al., 2012); however, it is notewor-thy that not every bull with poor quality semen had an abnormal thermo-gram. As a consequence, although infrared thermography is a useful tool for breeding soundness evaluation of bulls, it does not replace collection and evaluation of semen. In one study, 30 yearling beef bulls, all deemed
Figure 2. Infrared thermograms of the posterior aspects of the scrotum in two beef bulls. Left: This bull has a normal temperature pattern; decreasing temperatures from top to bottom and left to right symmetry. Right: This bull has a grossly abnormal temperature pattern, characterized by large areas of uniform temperature.
October 2013, Vol. 3, No. 4 23
Dow
nloaded from https://academ
ic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022

sound for breeding via a standard soundness examination, were individu-ally exposed to ~18 heifers for 45 days. Pregnancy rates 80 days after the end of the breeding season were similar (83 versus 85%) for bulls with a normal or questionable scrotal surface temperature pattern, respectively, but were greater (P < 0.01) than pregnancy rates for bulls with an abnormal scrotal surface temperature pattern (68%; Lunstra and Coulter, 1997).
Effects of Nutrition on Male Reproductive Development
In a series of experiments, Angus, Hereford, and Simmental bulls were fed high or medium nutrition from weaning, which occurred at 6 to 7 months of age until they were 12 to 24 months old (reviewed in Coulter and Kastelic, 1999). In general, bulls receiving high nutrition had greater body weight and backfat, but paired testes weight was not affected by diet. Moreover, bulls receiving high nutrition had less daily sperm production and epididymal sperm reserves and greater proportion of sperm abnormali-ties. It was speculated that increased dietary energy may adversely affect sperm production and semen quality due to fat deposition in the scrotum, thereby reducing heat radiated from the scrotal skin and increasing scrotal and testicular temperatures (reviewed in Coulter and Kastelic, 1999). In another study, bulls fed high-nutrition diets had greater scrotal circumfer-ence than those fed medium-nutrition diets, but paired testes weights were the same (Coulter and Kastelic, 1999). Since scrotal weight was greater in bulls fed high nutrition, perhaps fat deposition in the scrotum increased scrotal circumference in these bulls. In addition to the deleterious effects of high-energy diets on reproductive function, these diets may also result in abnormal foot growth due to laminitis, as well as adverse effects on bone and cartilage growth, and may increase the risk of rumen inflammation, liver abscess, and vesicular adenitis (Coulter and Kastelic, 1999).
A series of four experiments were conducted to evaluate the effects of nutrition during calfhood (6 to 8 to 26 to 30 weeks of age) and the subse-quent peripubertal period from approximately 30 to 70 weeks of age, on sexual development and reproductive function in beef bulls (reviewed by
Barth et al., 2008). In these studies, increased nutrition during calfhood resulted in a more sustained increase in luteinizing hormone (LH) pulse frequency early in life (before 25 weeks) and greater testicular develop-ment at maturity. Conversely, low nutrition during calfhood suppressed LH secretion before 25 weeks, delayed puberty, and reduced testicular devel-opment at maturity. For example, for bulls fed low, medium, or high nutri-tion from 10 to 70 weeks of age, age at puberty was 326.9 ± 5.5, 304.7 ± 7.4, and 292.3 ± 4.6 days respectively, and paired testis weights were 523.9 ± 25.8, 552.4 ± 21.1, and 655.2 ± 21.2 g (mean ± SEM). Furthermore, the initial suppression of testicular development and delayed puberty was not corrected in bulls fed reduced nutrition before approximately 26 weeks of age and subsequently placed on the high-nutrition regimen. Therefore, it is not possible to compensate for the effects of low nutrition early in life by subsequently giving supplemental feed.
When low nutrition was accomplished by restricted feed intake, hypo-thalamic and pituitary function were suppressed, with LH secretion most severely affected (Brito, 2006). It appeared that insulin-like growth factor I (IGF-I) was a possible signal to the central “metabolic sensor” involved in translating body nutritional status to the gonadotropin releasing hormone (GnRH) pulse generator (Brito, 2006). Nutrition also affected blood tes-tosterone concentrations, indicating effects on the number and/or function of Leydig cells. Age-related increases in physiological and GnRH-stimu-lated serum testosterone concentrations occurred earlier in bulls receiving high nutrition but were delayed in bulls receiving low nutrition; these ef-fects were probably mediated by both LH and IGF-I (Brito, 2006). Circu-lating leptin and insulin apparently have only permissive roles on GnRH secretion but may enhance testicular development. Growth hormone con-centrations decreased concomitantly with increasing IGF-I concentrations during sexual development in bulls, suggesting the testes could contribute considerable amounts of circulating IGF-I (Brito, 2006).
Sexual development and reproductive function were studied from 6 to 16 months of age in 22 Angus x Charolais and 17 Angus bulls (Brito et al., 2012). Associations of average daily gain (ADG) and body weight with ages at puberty and at maturity, scrotal circumference, paired-testes vol-ume and weight, testicular vascular cone diameter and fat thickness, scrotal temperature, sperm production and morphology, and testicular histology were determined. There were no significant correlations between cumula-tive ADG and any of the end points investigated. Body weight at various ages was negatively correlated with ages at puberty and maturity in Angus x Charolais bulls, positively correlated with paired-testes weight in Angus x Charolais and Angus bulls, and positively correlated with seminiferous tubule volume in Angus bulls (P < 0.05). Semen quality improved gradu-ally with age, and the interval between puberty (mean ± SD; 309.4 ± 29.7 and 357 ± 42 days of age) and maturity was approximately 50 days. Age, body weight, scrotal circumference, and paired-testes volume were all good predictors of pubertal and mature status, with moderate to high sensi-tivity and specificity (71.6 to 92.4%). In summary, growth rate between 6 and 16 months of age did not affect sexual development and reproductive function in beef bulls. However, greater body weight at various ages was associated with reduced ages at puberty and maturity and with larger testes at 16 months of age. Therefore, improved nutrition might be beneficial, but only when offered before 6 months of age. Average daily gains of approxi-mately 1.0 to 1.6 kg/day did not result in excessive fat accumulation in the scrotum, increased scrotal temperature, or reduction in sperm production and semen quality and could be considered “safe” targets for growing beef bulls.
An Angus calf. A series of experiments showed that increased nutrition during calf-hood resulted in a more sustained luteinizing hormone pulse frequency increase early in life, as well as improved testicular development and maturity (photo credit: Wikipedia commons).
24 Animal Frontiers
Dow
nloaded from https://academ
ic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022

Conclusions and Future Directions
Bull fertility remains critically important for sustainable cattle produc-tion. Although considerable progress has been made in our understanding of factors affecting bull fertility, and our ability to identify subfertile bulls, there are still considerable gaps in knowledge. For example, there is a need for reliable methods to accurately rank bulls as having good to excel-lent fertility. Increased testicular temperature interferes with production of morphologically normal sperm and decreases fertility; although the patho-genesis has been classically attributed to hypoxia, recent evidence was consistent with a direct effect of increased testicular temperature.
ReferencesAmann, R.P., and J.M. DeJarnette. 2012. Impact of genomic selection of AI dairy
sires on their likely utilization and methods to estimate fertility: A paradigm shift. Theriogenology 77:795–817.
Barros, C.M.Q., E. Oba, L.F.C. Brito, R.B. Cook, G.H. Coulter, G. Groves, M. Olson, and J.P. Kastelic. 1999. [Testicular blood flow and oxygen evaluation in Aberdeen Angus bulls] Avaliação do fluxo sanguineo e do oxigenio testicular em touros Aberdeen Angus. Revista Brasileira de Reprodução Animal 23:218–220.
Barth, A.D. 2007. Evaluation of potential breeding soundness of the bull. In: R.S. Youngquist and W.R. Threlfall, editors, Current Therapy in Large Animal Therio-genology 2. Saunders Elsevier, Philadelphia. p. 228–240.
Barth, A.D., and P.A. Bowman. 1994. The sequential appearance of sperm abnor-malities after scrotal insulation or dexamethasone treatment in bulls. Can. Vet. J. 35:93–102.
Barth, A.D., L.F.C. Brito, and J.P. Kastelic. 2008. The effect of nutrition on sexual development of bulls. Theriogenology 70:485–495.
Barth, A.D., and R.J. Oko. 1989. Abnormal morphology of bovine spermatozoa. Iowa State Univ. Press, Ames.
Brito, L.F.C. 2006. Nutrition, metabolic hormones, and sexual development in bulls. PhD thesis, University of Saskatchewan. http://ecommons.usask.ca/han-dle/10388/etd-04012006-184638.
Brito, L.F.C., A.D. Barth, R.E. Wilde, and J.P. Kastelic. 2012. Effect of growth rate from 6 to 16 months of age on sexual development and reproductive function in beef bulls. Theriogenology 77:1398–1405. http://ecommons.usask.ca/han-dle/10388/etd-04012006-184638. (Accessed 26 August 2013.)
Burfening, P.J., and L.C. Ulberg. 1968. Embryonic survival subsequent to culture of rabbit spermatozoa at 38° and 40°. J. Reprod. Fertil. 15:87–92.
Byrne, K., T. Leahy, R. McCulloch, M.L. Colgrave, and M.K. Holland. 2012. Com-prehensive mapping of the bull sperm surface proteome. Proteomics 12:3559–3579.
Cook, R.B., G.H. Coulter, and J.P. Kastelic. 1994. The testicular vascular cone, scro-tal thermoregulation, and their relationship to sperm production and seminal quality in beef bulls. Theriogenology 41:653–671.
Coulter, G.H., and J.P. Kastelic. 1999. Management programs for developing bulls. In: J.L. Howard and R. Smith, editors, Current Veterinary Therapy–Food Animal Practice 4. W.B. Saunders Co. p. 127–136.
DeJarnette, J.M. 2005. The effect of semen quality on reproductive efficiency. Vet. Clin. North Am. Food Anim. Pract. 21:409–418.
Flowers, W.L. 2013. Sperm characteristics that limit success of fertilization. J. Anim. Sci. 91:3022–3029.
Gadea, J., E. Selles, and M.A. Marco. 2004. The predictive value of porcine seminal parameters on fertility outcome under commercial conditions. Reprod. Domest. Anim. 39:303–308.
Johnston, J.E., H. Naelapaa, and J.B. Frye. 1963. Physiological responses of Hol-stein, Brown Swiss and Red Sindhi crossbred bulls exposed to high temperatures and humidities. J. Anim. Sci. 22:432–436.
Kastelic, J.P. 2013. Thermoregulation of the testes. In: R.M. Hopper, editor, Bovine Reproduction. Wiley-Blackwell, Hoboken. In press.
Kastelic, J.P., R.B. Cook, and G.H. Coulter. 1997. Contribution of the scrotum, testes, and testicular artery to scrotal/testicular thermoregulation in bulls at two ambient temperatures. Anim. Reprod. Sci. 45:255–261.
Kastelic, J.P., G.H. Coulter, and R.B. Cook. 1995. Scrotal surface, subcutaneous, in-tratesticular, and intraepididymal temperatures in bulls. Theriogenology 44:147–152.
Kastelic, J.P., G.H. Coulter, and R.B. Cook. 1996. Contribution of the scrotum and testes to scrotal and testicular thermoregulation in bulls and rams. J. Reprod. Fer-til. 108:81–85.
Kastelic, J.P., and J. Thundathil. 2008. Breeding soundness evaluation and semen analysis for predicting bull fertility. Reprod. Domest. Anim. 43(Suppl. 2):368–373.
Kastelic, J.P., J.C. Thundathil, and L.F.C. Brito. 2012. Bull BSE and semen analysis for predicting bull fertility. Clinical. Theriogenology 4:277–287.
Kastelic, J.P., R. Wilde, A. Bielli, P. Genovese, S. Bilodeau-Goeseels, and J. Thunda-thil. 2008. Hyperthermia is more important than hypoxia as a cause of disrupted spermatogenesis. Reprod. Domest. Anim. 43(Suppl. 3):166.
Kathiravan, P., J. Kalatharan, G. Karthikeya, K. Rengarajan, and G. Kadirvel. 2011. Objective sperm motion analysis to assess dairy bull fertility using computer-aided system–a review. Reprod. Domest. Anim. 46:165–172.
Larson, J.L., and D.J. Miller. 2000. Can relative spermatozoal galactosyltransferase activity be predictive of dairy bull fertility? J. Dairy Sci. 83:2473–2479.
Lunstra, D.D., and G.H. Coulter. 1997. Relationship between scrotal infrared temper-ature patterns and natural-mating fertility in beef bulls. J. Anim. Sci. 75:767–774.
Setchell, B.P., J.K. Voglmayr, and N.T. Hinks. 1971. The effect of local heating on the flow and composition of rete testis fluid in the conscious ram. J. Reprod. Fertil. 24:81–89.
Skinner, J.D., and G.N. Louw. 1966. Heat stress and spermatogenesis in Bos indicus and Bos taurus cattle. J. Appl. Physiol. 21:1784–1790.
Vogler, C.J., R.G. Saacke, J.H. Bame, J.M. Dejarnette, and M.L. McGilliard. 1991. Effects of scrotal insulation on viability characteristics of cryopreserved bovine semen. J. Dairy Sci. 74:3827–3835.
Waites, G.M.H. 1970. Temperature regulation and the testis. In: A.D. Johnson, W.R. Gomes, and N.L. Vandemark, editors, The testis. Academic Press, New York. p. 241–279.
Wildeus, S., and K.W. Entwistle. 1983. Spermiogram and sperm reserves in hybrid Bos indicus x Bos taurus bulls after scrotal insulation. J. Reprod. Fertil. 69:711–716.
Waites, G.M.H., and B.P. Setchell. 1990. Physiology of the mammalian testis. In: G.E. Lamming, editor, Marshall’s Physiology of Reproduction, Fourth Edition, Volume 2, Reproduction in the Male. Churchill Livingstone, Edinburgh. p. 1–105.
About the AuthorDr. John Kastelic holds a DVM with dis-tinction from the University of Saskatch-ewan (1982); a Ph.D. from the University of Wisconsin–Madison (1990), and a Pro-fessor of Honorary Cause from Szent Ist-ván University (2007). He is a diplomate of the American College of Theriogenolo-gists and a former Theriogenologist of the Year (2009). He spent two years in private veterinary practice, 22 years as a research scientist, and recently joined the Univer-sity of Calgary where he is a Professor of Cattle Reproductive Health-Theriogenol-
ogy and Head of the Department of Production Animal Health. For 10 years, Dr. Kastelic was the Co-Editor-in-Chief of the journal Theriogenology. He has published extensively on various aspects of reproduction, primarily in bulls and cows, and has given numerous presentations on animal reproduc-tion and scientific writing around the world. Correspondence: [email protected]
October 2013, Vol. 3, No. 4 25
Dow
nloaded from https://academ
ic.oup.com/af/article/3/4/20/4638651 by guest on 13 July 2022




















