
An evolutionary approach to below replacement fertility
24
Evolutionary Approach to Below Replacement Fertility HILLARD KAPLAN, 1 JANE B. LANCASTER, 1 * W. TROY TUCKER, 2 AND K.G. ANDERSON 3 1 Department of Anthropology, University of New Mexico, Albuquerque, New Mexico 2 Department of Anthropology, State University of New York, Stony Brook, Stony Brook, New York 3 Population Studies Center, University of Michigan, Ann Arbor, Michigan ABSTRACT The large human brain, the long period of juvenile dependence, long life span, and male support of reproduction are the co-evolutionary result of the human niche based on skill-in- tensive techniques of resource accrual. The regulation of fertility under traditional conditions is based upon a co-evolved psychology and physiology where adjustments of investment in offspring depend upon the returns to skill and mortality hazards. When all wealth is somatic, the hormonal system controlling ovulation and implantation translates income into genetic descendants. In modern society the existence of extra-somatic wealth is a critical condition to which our evolved proximate physio- logical mechanisms do not respond. However, psychological mechanisms regulating parental in- vestment in offspring quality may lead to greater and greater investment in own and offspring education, a smaller desired family size, a delay in the onset of reproduction, and a reduction in the total numbers of offspring produced. This delay in reproduction can cause many individuals to pro- duce fewer children than desired because fecundity falls during the reproductive part of the life course. As more individuals in a society follow this pattern, more will fail to reach their desired family size. At the same time the effective use of birth control decreases the numbers of families producing more children than desired. Below replacement fertility can result. Predictions from this model were tested using data from the National Survey of Families and Households and the Albuquerque Men study. Am. J. Hum. Biol. 14:233–256, 2002. Ó 2002 Wiley-Liss, Inc Over the past 120 years, there have been dramatic changes in fertility rates throughout the world. In most of Europe, North America, New Zealand, and Aus- tralia, a detectable change in fertility rates (10% reduction in I f ) occurred between 1880 and 1920, with the exception of France where the fertility transition occurred at the beginning of the 19 th Century (Caldwell and Caldwell, 1998; Coale and Treadway, 1986; Knodel and van de Walle, 1979). A second major transition has occurred over the last 30 years. In most countries in Asia, Latin America, and the Middle East, as well as parts of sub-Saharan Africa, marked re- ductions in fertility occurred between 1965 and 1985 (Bongaarts and Watkins, 1996; Caldwell and Caldwell, 1998). During the same period, many countries in developed nations underwent yet another decline in fertility, leading to ÔÔbelow-replacementÕÕ fertility levels (i.e., total fertility rates below 2.2 (Davis et al., 1986; Lesthaeghe and Williems, 1999; Morgan, 1996). This article provides a theoretical frame- work, the embodied capital theory of life history evolution, for explaining the pattern of those fertility transitions, and presents a series of specific hypotheses about the re- duction to below-replacement levels, with a focus on the United States. It is proposed that these fertility transitions result from the interaction of evolved psychological and physiological mechanisms governing hu- man fertility with specific changes in the economy and public health. Our thesis is that natural selection acted on human physiology and psychology to Contract grant sponsor: the University of New Mexico Bio- medical Research Grants; Contract grant number: 1988 and 1989; Contract grant sponsor: the University of New Mexico Research Allocations Committee; Contract grant number: 1988; Contract grant sponsor: the William T. Grant Foundation; Contract grant number: #89130589 and #91130501; Contract grant sponsor: the National Science Foundation 1990–1993; Contract grant number: #BNS-9011723 and #DBS-911552; Contract grant sponsor: National Science Foundation; Contract grant sponsor: National Science Foundation Graduate Research Fellowship (to Kermyt G. Anderson); Contract grant sponsor: a National Institute on Aging grant; Contract grant number: AG15906; Contract grant sponsor: the Andrew W. Mellon Foundation Contract grant sponsor: the National Institute of Child Health and Human Development to the Population Studies Center of the University of Michigan. *Correspondence to: Department of Anthropology, University of New Mexico, Albuquerque, NM 87131. E-mail: [email protected] Received 11 September 2000; Revision received 30 January 2001; Accepted 5 October 2001 AMERICAN JOURNAL OF HUMAN BIOLOGY 14:233–256 (2002); DOI 10.1002/ajhb.10041 ª 2002 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of An evolutionary approach to below replacement fertility

Evolutionary Approach to Below Replacement Fertility
HILLARD KAPLAN,1 JANE B. LANCASTER,1* W. TROY TUCKER,2 AND K.G. ANDERSON3
1Department of Anthropology, University of New Mexico, Albuquerque, New Mexico2Department of Anthropology, State University of New York, Stony Brook, Stony Brook,New York3Population Studies Center, University of Michigan, Ann Arbor, Michigan
ABSTRACT The large human brain, the long period of juvenile dependence, long life span, andmale support of reproduction are the co-evolutionary result of the human niche based on skill-in-tensive techniques of resource accrual. The regulation of fertility under traditional conditions is basedupon a co-evolved psychology and physiology where adjustments of investment in offspring dependupon the returns to skill and mortality hazards. When all wealth is somatic, the hormonal systemcontrolling ovulation and implantation translates income into genetic descendants. In modern societythe existence of extra-somatic wealth is a critical condition to which our evolved proximate physio-logical mechanisms do not respond. However, psychological mechanisms regulating parental in-vestment in offspring quality may lead to greater and greater investment in own and offspringeducation, a smaller desired family size, a delay in the onset of reproduction, and a reduction in thetotal numbers of offspring produced. This delay in reproduction can cause many individuals to pro-duce fewer children than desired because fecundity falls during the reproductive part of the lifecourse. As more individuals in a society follow this pattern, more will fail to reach their desired familysize. At the same time the effective use of birth control decreases the numbers of families producingmore children than desired. Below replacement fertility can result. Predictions from this model weretested using data from the National Survey of Families and Households and the Albuquerque Menstudy. Am. J. Hum. Biol. 14:233–256, 2002. � 2002 Wiley-Liss, Inc
Over the past 120 years, there have beendramatic changes in fertility ratesthroughout the world. In most of Europe,North America, New Zealand, and Aus-tralia, a detectable change in fertility rates(10% reduction in If) occurred between 1880and 1920, with the exception of Francewhere the fertility transition occurred atthe beginning of the 19th Century (Caldwelland Caldwell, 1998; Coale and Treadway,1986; Knodel and van de Walle, 1979). Asecond major transition has occurred overthe last 30 years. In most countries in Asia,Latin America, and the Middle East, as wellas parts of sub-Saharan Africa, marked re-ductions in fertility occurred between 1965and 1985 (Bongaarts and Watkins, 1996;Caldwell and Caldwell, 1998). During thesame period, many countries in developednations underwent yet another decline infertility, leading to ��below-replacement��fertility levels (i.e., total fertility rates below2.2 (Davis et al., 1986; Lesthaeghe andWilliems, 1999; Morgan, 1996).This article provides a theoretical frame-
work, the embodied capital theory of lifehistory evolution, for explaining the patternof those fertility transitions, and presents a
series of specific hypotheses about the re-duction to below-replacement levels, with afocus on the United States. It is proposedthat these fertility transitions result fromthe interaction of evolved psychological andphysiological mechanisms governing hu-man fertility with specific changes in theeconomy and public health.Our thesis is that natural selection acted
on human physiology and psychology to
Contract grant sponsor: the University of New Mexico Bio-medical Research Grants; Contract grant number: 1988 and1989; Contract grant sponsor: the University of New MexicoResearch Allocations Committee; Contract grant number: 1988;Contract grant sponsor: the William T. Grant Foundation;Contract grant number: #89130589 and #91130501; Contractgrant sponsor: the National Science Foundation 1990–1993;Contract grant number: #BNS-9011723 and #DBS-911552;Contract grant sponsor: National Science Foundation; Contractgrant sponsor: National Science Foundation Graduate ResearchFellowship (to Kermyt G. Anderson); Contract grant sponsor: aNational Institute on Aging grant; Contract grant number:AG15906; Contract grant sponsor: the Andrew W. MellonFoundation Contract grant sponsor: the National Institute ofChild Health and Human Development to the PopulationStudies Center of the University of Michigan.*Correspondence to: Department of Anthropology, University
of New Mexico, Albuquerque, NM 87131.E-mail: [email protected] 11 September 2000; Revision received 30 January
2001; Accepted 5 October 2001
AMERICAN JOURNAL OF HUMAN BIOLOGY 14:233–256 (2002); DOI 10.1002/ajhb.10041
ª 2002 Wiley-Liss, Inc.

produce a coordinated and flexible responsesystem that regulates fertility and parentalinvestment. Since all but the last 10,000years of human evolution occurred amongforaging peoples, this system is adapted tothe selection pressures acting on the hunt-ing and gathering life way. Cross-culturalresearch on foraging peoples shows thathumans are adapted to a skill-intensiveforaging niche, requiring years of learningbefore adult competence is reached (Kaplan,1997; Kaplan et al., 2000, 2001). This for-aging niche selected for a co-evolved complexof life-history traits, including prolongeddevelopment and dependency on parents, alarge expensive brain, a very long adult lifespan (during which foraging competenceincreases for 20 or more years), and a three-generational system of resource flows inwhich grandparents as well as parents ac-tively provision children and adolescents.Because of the skill-intensive foragingniche, natural selection acted on humanpsychology in such a way that people aresensitive to environmental variation inlearning opportunities and payoffs. Theyadjust both their own investments inlearning and the amount of time they arewilling to subsidize children in response tothat variation.These behavioral adjustments impact
fertility via two pathways in maternalphysiology. Fecundity (and hence the lengthof inter-birth intervals) is affected by boththe duration of the breastfeeding period andmaternal nutrition, which is, in turn, de-termined by the distribution and productionof food in families. When all wealth is in theform of food energy consumed, there is adirect link between parental investmentand the energetics of reproduction. Thislink mediated adaptive (i.e., fitness maxi-mizing) increases and decreases in fertilityin response to changing payoffs to parentalinvestment.In the context of modern economic, tech-
nological, and social conditions, this evolvedpsychology and reproductive physiologyproduces responses that do not maximizefitness but that are patterned nevertheless.The emergence of extra-somatic wealth (i.e.,land, money, and other forms of physicalcapital) largely broke the link between pa-rental investment and the energetics of re-production. The severing of this linkappears to generate lower fertility andgreater production and consumption of
wealth than would maximize fitness. It ishypothesized that the demographic transi-tion to small families can be understood interms of changing payoffs to human capitalinvestment. These changes are due to theincreased importance of education in thedetermination of wages and to reductions inmortality and to the use of money to pur-chase consumer goods, both of which moti-vate people to delay fertility and to limitfamily size.The subsequent transition to below-re-
placement fertility is due to further changesin technology with multiple effects. First,new technologies changed the distributionof jobs in the economy, increasing in thebenefits of schooling and on-the-job trainingfor both men and women. Second, they in-creased the relative payoffs of market tohome labor for women, increasing the costsof child rearing and decreasing maritalstability. Third, they increased efficiency ofbirth control. These three effects produced agreater tendency to limit fertility to twochildren and further delays in the onset ofreproduction, especially for those individu-als expecting to invest heavily in their ownand in their children�s embodied capital. Itis proposed that below-replacement fertilityis largely a phenomenon of the more edu-cated sector of society, where many indi-viduals involuntarily fall short of their idealfertility due to excessive delay (given thephysiological decline in fecundity with ageand effective birth control). It is hypothe-sized that the delay is due to two factors: 1)increased investments in education and on-the-job training and 2) greater selectivity inobtaining a suitable partner with whom toco-invest in children, due to the criticalnature of both maternal and paternal skills-based investments and to the greater threatof divorce in modern, cash economies. Theprincipal focus of this paper, therefore, is anexamination of the causes of delay in initi-ating reproduction and of reduction in tar-get fertility to two children.The next section presents a brief overview
of the embodied capital theory of life historyevolution. The theory is then applied tohunter-gatherer ecology, with a specificemphasis on the evolved psychology andphysiology of human reproduction. Next,the interaction of this evolutionary heritagewith modern socio-economic conditions isexamined in an attempt to account formodern demographic transitions and below-
234 H. KAPLAN ET AL.

replacement fertility. Throughout this sec-tion, the interaction of micro- and macro-level processes and of public and privateinvestments are considered in an attempt tounderstand the historical dynamics.Finally, a series of empirical analyses
regarding changing patterns of fertility,parental investment, and marital dissolu-tion in the U.S. are presented, using both anational probability sample (The NationalSurvey of Families and Households) and asample of men in Albuquerque, New Mexi-co. The results of these analyses provide agood deal of support for hypotheses derivedfrom the embodied capital theory. They donot constitute definitive tests, however, be-cause some of the analyses were conductedprior to the development of the hypothesesand more direct evidence about the causesof delay is required. The paper concludeswith a discussion of further directions fortheory development and empirical research.
EMBODIED CAPITAL THEORY ANDTHE EVOLVED PSYCHOLOGY
AND PHYSIOLOGY OFHUMAN REPRODUCTION
Action of natural selection on fertilityand parental investment
According to the theory of evolution bynatural selection, the evolution of life re-sults from a process in which variant formscompete to harvest energy from the envi-ronment and convert that energy into rep-licates of those forms. Those forms thatcapture more energy and convert that en-ergy more efficiently into offspring becomemore prevalent through time. This rela-tionship between harvesting energy andreproduction generates many complex,time-dependent problems.Fertility contributes most directly to an
organism�s fitness. In fact, all other fitnesscomponents, such as mortality, only affectfitness through their effects on fertility(e.g., mortality rates affect fitness by af-fecting the probability of living to the nextreproductive event). All else being constant,any increase in fertility increases an or-ganism�s fitness. However, there are twotrade-offs that affect natural selection onfertility.The first trade-off balances expenditures
between present and future reproduction.Imagine a sexually reproducing organism
that is newly independent of support fromits parents. It can use its time to harvestenergy from the environment. It can usesome or all of its energy to reproduce.However, another potential use of the en-ergy is to buy more time. By using some ofthe energy for physical maintenance and todefend against diseases and predators, itcan live longer to harvest more energy inthe future. Another use of energy is to growand build tissue. By growing, an organismcan increase its energy capture rates in thefuture and thus increase its future fertility.The allocation of energy to growth also canincrease the length of the life span by low-ering size-dependent mortality and can in-crease success in intra-sexual competitionfor mates, ultimately affecting reproductiverate. For this reason, organisms typicallyhave a juvenile phase in which fertility iszero until they reach a size at which someallocation to reproduction increases fitnessmore than does further growth. Similarly,among organisms that engage in repeatedbouts of reproduction (humans included),some energy during the reproductive phaseis diverted away from reproduction and al-located to maintenance so that it can live toreproduce again. Evolutionary theory ex-pects natural selection on age of first re-production and on adult reproductive rateto maximize total energetic allocations toreproduction. Over the life course, this willdepend on ecological factors, such as mor-tality rates (Charnov, 1993) and the effectsof body size on both energy capture (Char-nov, 1993) and vulnerability to predation(Werner, 1986).The second trade-off is between quantity
and quality of offspring, where quality is afunction of parental investment in an off-spring and reflects its ability to survive andreproduce. In general, natural selection onoffspring number and investment per off-spring is expected to maximize the numberof offspring that survive to reproduce(Smith and Fretwell, 1974). Ecological fac-tors, such as mate pool competition, foodsupply, disease, and predation rates, alsoaffect the optimal fertility rates and optimalexpenditures on offspring.Virtually all complex organisms exhibit
flexibility in both age-at-first-reproductionand fertility rates. Natural selection hasresulted in physiological and psychologicalmechanisms by which individuals adjustfertility onset and fertility rates in relation
BELOW REPLACEMENT FERTILITY 235

to changing environmental conditions. Forexample, plants often exhibit many, thou-sands-fold variation in fertility rates in re-lation to yearly and season variation inrainfall and other environmental factors.Extensive research on many bird specieshas shown that this phenotypic plasticitytracks fitness quite well (Godfray et al.,1991). Birds under variable conditions ad-just clutch sizes in ways that tend to max-imize the number of surviving youngproduced during the life course. The per-vasiveness of adaptive phenotypic plasticityin fertility among non-humans suggeststhat humans are also likely to possess ad-aptations to adjust fertility onset and fer-tility rates in relation to changingenvironmental conditions.
Embodied capital theory
The embodied capital theory generalizesexisting life history theory in biology bytreating the processes of growth, develop-ment, and maintenance as investments instocks of somatic or embodied capital (Ka-plan, 1997; Kaplan et al., 2000). In a phys-ical sense, embodied capital is organizedsomatic tissue—muscles, digestive organs,brains, etc. In a functional sense, embodiedcapital includes strength, immune function,coordination, skill, and knowledge. Becausesuch stocks tend to depreciate with time,allocations to maintenance also representinvestments in embodied capital. Thus, thepresent/future reproduction trade-off can beunderstood in terms of optimal investmentsin own embodied capital vs. reproduction,and the quantity-quality trade-off can beunderstood in terms of investments in theembodied capital of offspring vs. the num-ber of offspring produced.The embodied capital theory allows us to
address problems not yet explored withstandard life history models. First, an ex-clusive focus on physical growth per seprovides an impoverished understanding ofdevelopment. The large human brain, forexample, embodies a stock of capital thatincludes a great deal of skill and knowledgeacquisition during both the juvenile andadult periods. Growth in the form ofknowledge may be as important as growthin body size with respect to providing ben-efits through time. Second, parental in-vestment not only affects survival toadulthood, but also the adult income (pro-
ductivity) of offspring. This is especiallytrue of humans both in modern societiesand among traditional hunter-gatherers.Models of investment in embodied capital
have produced some fundamental results.One set of models shows that investmentsin embodied capital affecting adult incomeor energy capture co-evolve with invest-ments affecting mortality and longevity(Kaplan, 1996; Kaplan et al., 2000; Kaplanand Robson, 2001). The longer the timespent growing and learning prior to repro-ducing, the more natural selection favorsinvestments in staying alive to reap thebenefits (the higher rates of returns fromskills-based productivity) of those invest-ments. Similarly, any investments thatproduce increased energy capture rates inmiddle age select for additional investmentsto reach older ages. The converse is alsotrue. Ecological features or investmentsthat increase the probability of survival toolder ages also produce selection for greaterinvestments in income-related embodiedcapital. These co-evolutionary effects havebeen particularly important in human lifehistory evolution (see below).A second set of models concerns optimal
parental investment in offspring (Kaplan,1996). Those models decompose parentalinvestment into those investments that af-fect offspring survival and those that affectoffspring income in its adulthood. With afixed parental income, increased invest-ment in offspring embodied capital de-creases parental fertility, because parentalfertility is determined by dividing averageinvestment per offspring into total parentalincome. Thus, a quality/quantity trade-off isfundamental to parental decision-makingabout the allocation of parental invest-ments.The models of parental investment also
show that the investments in income/ener-gy production and survival co-evolve. If theaverage level of investment in either off-spring survival or offspring income in-creases, the production of each additionaloffspring is more costly. All things beingequal, this favors further increases in off-spring quality and hence increased invest-ment in both survival-related and income-related embodied capital (see Becker, 1981,and Becker and Lewis, 1973, for the originaltreatment of quantity/quality interactions,and Rogers and Blurton Jones, 1992, for asimilar result). In addition, exogenous
236 H. KAPLAN ET AL.

changes in offspring survival rates affectfertility through two routes. First, increasesin offspring survival during the period ofparental investment increase the expectedcost of raising an offspring, because it willhave a higher likelihood of reaching eachage to receive the investment. With a fixedparental income constraint, this necessarilyreduces offspring quantity. Second, this in-crease in the expected cost of raising anoffspring also favors an increase in optimalinvestments in child quality, in terms ofboth child survival and child income. Theseresults also imply that if payoffs to invest-ments in both offspring income and survivalincrease, large changes in fertility are ex-pected.
Application of embodied capital theoryto traditional humans
Hominids lived as hunter-gatherers forthe vast majority (more than 2 millionyears) of their evolutionary history. Agri-culture originated only 10,000 years agoand has been practiced by the majority ofthe world�s population for just two or threemillennia, a relatively brief period of timefor selection to act. The shift to a fullymercantile, highly urbanized economy withcompetitive wage-labor markets and a vastarray of consumer goods over the last 150years is completely novel. Because naturalselection is an historical process, humansshould be well adapted to the foraging life-style. There is no reason to expect that theywill respond adaptively (in the sense ofmaximizing reproductive fitness) to today�snovel circumstances (see Kaplan et al.,1995, for evidence that modern fertility be-havior does not maximize fitness). There-fore, an evolutionary explanation of humanfertility and parental investment has threerequirements: 1) it must be consistent withour general understanding of evolution bynatural selection, 2) it must explain thebehavior of humans living under traditionalhunting and gathering conditions charac-teristic of our evolutionary history withadaptive models, and 3) it must be able topredict the pattern of modern responses tonovel conditions.Compared with other primates and
mammals, there are at least four distinctivecharacteristics of human life histories: 1) anexceptionally long life span, 2) an extendedperiod of juvenile dependence, 3) support of
reproduction by older post-reproductive in-dividuals, and 4) male support of repro-duction through the provisioning of femalesand their offspring. Kaplan and colleagues(Kaplan, 1997; Kaplan et al., 2000) proposethat a dietary shift towards high-quality,difficult-to-acquire food resources, that arenutrient dense and come in large packages,is responsible for the evolution of those lifehistory characteristics and the large brainsupon which human cultural evolution re-lies.The following logic underlies this pro-
posal. First, high levels of knowledge, skill,coordination, and strength are required toexploit the suite of high-quality, difficult-to-acquire resources that humans consume.The attainment of those abilities requirestime and a significant commitment to de-velopment. This extended learning phaseduring which productivity is low is com-pensated for by higher productivity duringthe adult period with an intergenerationalflow of food from old to young. Becauseproductivity increases with age, the timeinvestment in skill acquisition and knowl-edge leads to selection for lowered mortalityrates and greater longevity. This in turnwill favor a longer juvenile period if thereare important gains in productive abilitywith body size, and growth ceases at sexualmaturity. Second, the characteristics of thefeeding niche, with associated food sharing,provisioning of juveniles, and tool use led tolower mortality during the juvenile andearly adult periods. This too, favors a longerjuvenile period and higher investment infurther mechanisms to increase life span.Thus, it is proposed that the long humanlife span co-evolved with the lengthening ofthe juvenile period, increased brain capaci-ties for information processing and storage,and intergenerational resource flows-all asa result of an important dietary shift. Thesechanges are all consistent with the predic-tions of the models discussed above.In support of this hypothesis, available
data show that: 1) human hunter-gatherersrely almost exclusively on extracted andhunted foods, whereas chimpanzees relymuch more heavily on easily-collected foods;2) the rate at which both hunter-gathererchildren and chimpanzee juveniles acquirefoods depends directly on the difficulty ofacquisition, with extracted and huntedfoods requiring the longest time to master;3) as a result, hunter-gatherer children do
BELOW REPLACEMENT FERTILITY 237

not acquire as much food as they consumeuntil late adolescence or early adulthood, incontrast to chimpanzees who do so afterabout age 6; 4) peak production rates forboth human males and females do not occuruntil the mid-thirties for the most difficult-to-acquire resources, but ultimately thesepeaks compensate for the lost energy pro-duction during the learning period; and 5)the provisioning of hunted foods by men isan essential component of human repro-ductive energetics and the support of longjuvenile period (Kaplan et al., 2000). Theanalyses also suggest that the feeding nicheindirectly lowered mortality rates, due tothe effects of provisioning and food sharingon the ability to resist illness and recoverfrom injury, and the use of tools to fend offpredators. It follows that the first majordemographic transition separating ourspecies from apes is a shift to low mortality,an increased life span, a long dependentdevelopment period, a large commitment tolearning and intelligence, and an age-pro-file of production shifted towards older ageswith wealth flows downward between gen-erations (Kaplan et al., 2001).The human adaptation is broad and flex-
ible in one sense and very narrow and spe-cialized in another sense. It is broad in thesense that as hunter-gatherers, humanshave existed successfully in virtually all ofthe world�s major habitats. This has en-tailed eating a very wide variety of foods,both plant and animal, both within andamong environments. It also has entailedlarge flexibility in the contributions of dif-ferent age and sex classes of individuals.The relative contributions of men andwomen to food production vary from groupto group, as do production profiles by age.The human adaptation is narrow and spe-cialized in that it is based on extremely highinvestments in brain tissue and learning. Inevery environment, human foragers con-sume the largest, highest quality and mostdifficult-to-acquire foods, using techniquesthat take years to learn (Kaplan et al.,2000). Therefore, although the specific dietand techniques used to extract resourcesfrom the environment may differ fromgroup to group and global forces have im-pacted all modern groups, the robustness ofthis pattern makes us confident about usingthe niche occupied by modern hunter-gath-erer as a model for the evolutionary past ofour species.
Ecology and the proximate mechanismsgoverning fertility and parental investment
The mortality and production profilesdiscussed above have direct implications forfertility regulation. Because human parentsand grandparents provision children, natu-ral selection probably produced mecha-nisms by which fertility could respond tothe number of children parents could affordto raise in any given socio-ecology. A con-ceptual model of the evolved mechanism offertility regulation in humans and theirresponsiveness to physical, biological, andsocial conditions includes two classes of so-cio-ecological factors affecting survival andembodied capital, respectively, and threedomains of evolved responses; psychologyand culture, behavior, and reproductivephysiology. Two main causal pathwaysregulate fertility, both related to the ener-getic constraints in provisioning offspring;one involves breastfeeding and the otherinvolves the age profiles of work and fooddistribution. Natural selection on the tim-ing and intensity of weaning should reflecta balance between quantity effects (favoringearlier weaning) and quality effects onsurvival and growth of offspring (favoringlater weaning). Socio-ecological conditions,such as the tradeoff between women�s workeffort and child care or conflicts of interestbetween the health of mother and the childwhen one is at the expense of the other(McDade, 2001; Sellen and Smay, 2001)impact the relationship between age atweaning and survival. Similarly, infantcondition is likely to vary individually andbe differentially affected by weaning. It isexpected that natural selection acted onboth infant and maternal psychology tomake them responsive to those variableconditions. For example, sickly infants maybe more dependent on the breast for nutri-tion and larger and healthier babies moreinterested in weaning foods. In the humancase, individual experience and sensitivityto infant�s condition guide maternal per-ceptions regarding breastfeeding in con-junction with the cumulative experience ofthe group, as reflected in cultural conven-tions and ideas about proper breastfeedingpractices. Maternal and infant psychologiesinteract in determining the timing of sup-plementation and the continuation ofbreastfeeding. The evolved reproductivephysiology translates this pattern of
238 H. KAPLAN ET AL.

breastfeeding into differing degrees of fe-cundity.The additional energetic constraints in-
volved in provisioning children form thesecond causal pathway. The age/sex-profileof work and productivity determines thetotal energy available for consumption.Given some age profile of food distributionand of mortality, the requirement that en-ergy consumed does not exceed energy pro-duced constrains fertility rate. The longerchildren are provisioned and the greater theproportion of their food needs subsidized byparents and grandparents, the lower thesupportable reproductive rate with a givenadult income. Natural selection should,therefore, act both on psychological abilitiesto assess the short- and long-term costs andbenefits of different activity profiles and onfertility regulation so that energy flows arebalanced.One fundamental trade-off with respect to
activity profiles is between present and fu-ture production (Bock, 2001; Kaplan, 1996).If children engage in easy-to-perform for-aging activities, such as fruit collection, thisprovides immediate caloric benefits andlowers the parental subsidy required.However, time allocation to those activitiesdetracts from time spent learning moredifficult activities, such as hunting and ex-tractive foraging. During the learningphase, those activities produce very littlefood, but generate high rates of return inthe future. Another fundamental trade-offis between production and mortality risks,since foraging exposes people to predation,accidents, and getting lost. This suggeststhat parental psychology should be respon-sive to age-specific mortality rates and howthey are affected by alternative activityprofiles and on their short- and long-termproductive consequences.A growing body of evidence supports the
view that people in foraging societies aresensitive to these trade-offs and they adjustbehavior accordingly. In a seminal series ofpapers, Blurton Jones and colleagues(Blurton Jones et al., 1994a; 1994b; Hawkeset al., 1995) show that Kung hunter-gath-erer parents consciously assess foragingrisks to children and that differences be-tween !Kung and Hadza children�s produc-tivity result from features of the localecology related to ease of resource extrac-tion and dangers associated with productivetasks.
Recent studies of the productivity ofchildren indicate that the suite of compe-tencies relating to adult capacities variesacross time and place. In general, the pro-ductivity of children in foraging societiesdepends on the availability of foods, such asfruits and shellfish, that can be obtainedefficiently without skill. The productivity ofAche and Hadza children increases dra-matically when fruits are in season (Haw-kes et al., 1989; Kaplan, 1997), and Meriamchildren can be quite productive collectingshellfish and small fish on island reefs (Birdand Bliege Bird, 2001; Bliege Bird and Bird,2001). Bock (1995, 2001), in a study of for-ager/agro-pastoralists in Botswana, showsthat the age-profiles of children�s activitybudgets reflect immediate skill andstrength requirements as well as opportu-nities to learn. Blurton Jones and Marlowe(2001) found that among Hadza childrenthe components of skilled performancessuch as climbing, digging and archery aredependent on individual growth rather thanpractice to mature. However, in all foragingsocieties for which data are available, peo-ple are aware of the many years of knowl-edge acquisition required to become acompetent hunter, and young men are pro-visioned while they learn those skills (Ka-plan et al., 2000). In the future suchvariation in the significance and timing ofgrowth-based and experience-based em-bodied capital will help to define the natureof tradeoffs faced by parents in varying so-cioecological contexts.These time allocation decisions affect
fertility through maternal energetics aswell. In addition to the direct breastfeedingeffects, fecundity responds to women�s en-ergy status, energy balance, and rate ofenergy flow (Ellison, 1990, 1994). Seasonalchanges of food intake (Bailey et al., 1992),net energy expenditure (Hurtado and Hill,1987), and weight (Bailey et al., 1992;Tracer, 1996) all have been shown to predictfecundity as do individual differencesamong women (Hill and Hurtado, 1996).To summarize, in this model the interac-
tion of individual psychology and cumula-tive cultural knowledge, beliefs, andpractices, behavior, and reproductive phys-iology determine fertility. Each of thesedomains co-evolved by natural selection torespond to ecological variability in mortalityregimes and payoffs to parental subsidiza-tion of skill acquisition. The connection
BELOW REPLACEMENT FERTILITY 239

between psychology and fertility is indirectin that psychology adjusts parental invest-ment (through productive behavior, wealthflows, and breastfeeding) and reproductivephysiology translates parental investmentdecisions into fertility. The key to this sys-tem is that maximizing lifetime expectedresource production through the optimalallocation of activities and wealth flowstends also to maximize fitness when allwealth is in the form of food, and extra foodtranslates into higher fertility.
MODERN DEMOGRAPHIC TRANSITIONS
Initial transition from high to low fertility
The next step in the analysis is to deter-mine which aspects of modernization, ininteraction with those evolved psychologicaland physiological mechanisms, lead peopleto lower their fertility. There are two newsocio-ecological factors that precipitated atransition from high to low fertility. Thesecritical new factors are the existence of ex-tra-somatic capital and changes in thepayoffs to embodied capital, especially inthe form of education.
Extra-somatic wealth: breaking the linkbetween wealth and fertility. With few ex-ceptions (e.g., the coastal areas of Californiaand British Columbia and late Pleistocenearcheological sites in the Ukraine), allwealth among hunter-gatherers appears tobe energy stored in people�s bodies. Thiswealth serves to produce offspring and toincrease survival. Under these circum-stances, an evolved psychology and physi-ology that maximizes total wealth producedby the lineage would maximize fitness. Thestorage of wealth in the form of extra-so-matic resources, such as land, cattle, ormoney, probably is a relatively recent phe-nomenon. It is unclear, therefore, whetherselection to have optimized fertility and themanagement of extra-somatic wealth,should be expected, given this pre-existingsuite of evolved proximate mechanisms.Where wealth is held outside the body,there is no mechanism to ensure that it iseventually converted into reproduction. Infact, it may be that the evolved psychologi-cal mechanisms that are sensitive to theimpacts of behavior on future wealth maypromote wealth-maximization at the ex-pense of fertility. In the case of modern
fertility, it appears that people are at-tempting to circumvent the physiologicalmechanisms regulating fertility, by pre-venting pregnancies that would otherwiseoccur.
Changing payoffs to investment in embod-ied capital. Changes in the pay-offs to em-bodied capital occur for two reasons. Duringthe 19th century, there were large changesin both public health practice and scientificunderstanding of disease (Preston and Ha-ines, 1991). This led to a dramatic decline inboth infant, child, and adult mortality ratesthat continued for close to a century. Asscientific advances enabled reductions inmortality rates, there was strong pressureto increase public investments in healthand disease prevention, from the protectionof the water supply at the turn of the cen-tury to the development of vaccines andpublic access to medical care in the 20thcentury. The decline in mortality rates in-creased the return on parental investmentand own investment in embodied capitalaffecting wage rates. Increased survivalrates during the period of parental invest-ment increase the expected costs per childborn, favoring further increases in offspringquality (Kaplan, 1996). Increased survivalduring the adult period increases the ex-pected years of return to educational in-vestments. This further increases theincentive to invest in children�s education[see Meltzer (1992) for supporting data andtheory, but see Preston (1980) who arguesfor a comparatively small effect of changingmortality patterns on rates of return to ed-ucation].At the same time, scientific advances
also led to changes in the economics ofproduction. The economic changes in-creased wage premiums paid for skills andled to very high returns on parental in-vestments in children’s education (Vinov-skis, 1994). Increased private interest ineducation may have inspired more publicinvestment in education. The quality ofpublic education, in turn, increased the rateof return on private investments in educa-tion.These factors, in combination with pa-
rental psychology, result in fertility beingregulated by a consciously determined fer-tility plan realized through birth controltechnology and/or controlled exposure to
240 H. KAPLAN ET AL.

sex. In response to the increased payoffs toinvestments in education and the expectedcosts of those investments, parents deter-mine the number of children they can affordto raise, given their wealth. The low mor-tality rates also allow parents to plan re-production at the outset, because thenumber of children born accurately predictsthe number of children that will be raised toadulthood.According to this view, the reason why
the transition begins with a negative asso-ciation between wealth and fertility is thatthe payoffs to investment in own and off-spring embodied capital were not the sameacross economic strata. First, infant andchild mortality rates were much loweramong the wealthy than the poor (Prestonand Haines, 1991) in both Europe andAmerica. Second, wealthy parents tended tobe more educated and to have greater ac-cess to educational resources. Our hypoth-esis is that, as a result, wealthier parentscould educate their offspring more effi-ciently than poorer parents could and had agreater return on investments in both ownand offspring education.Public and private sector responses to
these trends then began to change thepayoffs to such investments among thepoorer and less educated sectors of the so-ciety. Increased public investments inhealth and schooling decreased mortalityrates and increased the efficiency of privateinvestments in children�s schooling for allsectors of the society. More effective andless costly forms of birth control technolo-gies allowed for a significant reduction inbirth rates. An increasingly educated pop-ulace increased the expertise in firms withrespect to both management and the tech-nology of production, further increasing thedemand for skilled workers. Ultimately,those interactions produce equal fertilityamong economic strata. The wealthier,more educated people invest more per childthan poorer, less educated, because they arestill more efficient at embodying capital intheir children, but their higher wealth al-lows them to have as many children (al-though more costly) as less wealthyindividuals. Thus, what appears to be agradual decrease in fertility through timeaccording to national-level statistics mayactually be a process in which an increasingproportion of the population exhibits verylow fertility.
Transition to below-replacement fertility
Fertility has not declined steadilythroughout this century. For example, inthe U.S. total fertility rates declined fromapproximately 1880 until the mid 1930s,then rose to a peak in the late 1950s duringthe ��Baby Boom,�� and then declined to be-low-replacement levels by the mid-1970s(Morgan, 1996).Two principal routes by which social,
economic, and technological changes resultin below replacement fertility are proposed.The first route is the changing pay-offs toinvestment in education for the populationas a whole, and the second route dependsspecifically on work force participation bywomen and its impact on family formation.There are also macro-level socio-economicchanges affecting individual decisions.Whereas those macro-level processes arepartially determined by individual decisions(i.e., growth in technology depends, in part,on the educational attainments of theworkforce), individuals must treat thoseconditions as given when making decisionsabout how much to invest in education,when to marry, and how many children tohave. Growth in technology and world tradehas had profound effects on socio-economicconditions in developed countries. First, therelationship between education and incomehas strengthened [see, for example, Boundand Johnson (1992) and Fig. 1 below]. Sec-ond, there has been a changing distributionof jobs in the economy with a shift from theindustrial sector to the service sector, in-creasing the employment opportunities forwomen, and especially increasing theearning power of educated women (Murphyand Welsh, 1989). Third, public invest-ments in education increased substantially,with a great expansion of public institutionsof higher learning. At the same time as op-portunities for college education expanded,the costs of obtaining a college degree in-creased (in real dollars), especially in pri-vate institutions.In responses to those socioeconomic
changes, both men and women increasedtime spent in schooling and presumablytheir expected investments in their chil-dren�s schooling. It is proposed, however,that differences in pre-school experiences,quality of public schools attended, and homeenvironment, all of which are associatedwith parental education, cause there to be
BELOW REPLACEMENT FERTILITY 241

Fig. 1. Continued
242 H. KAPLAN ET AL.
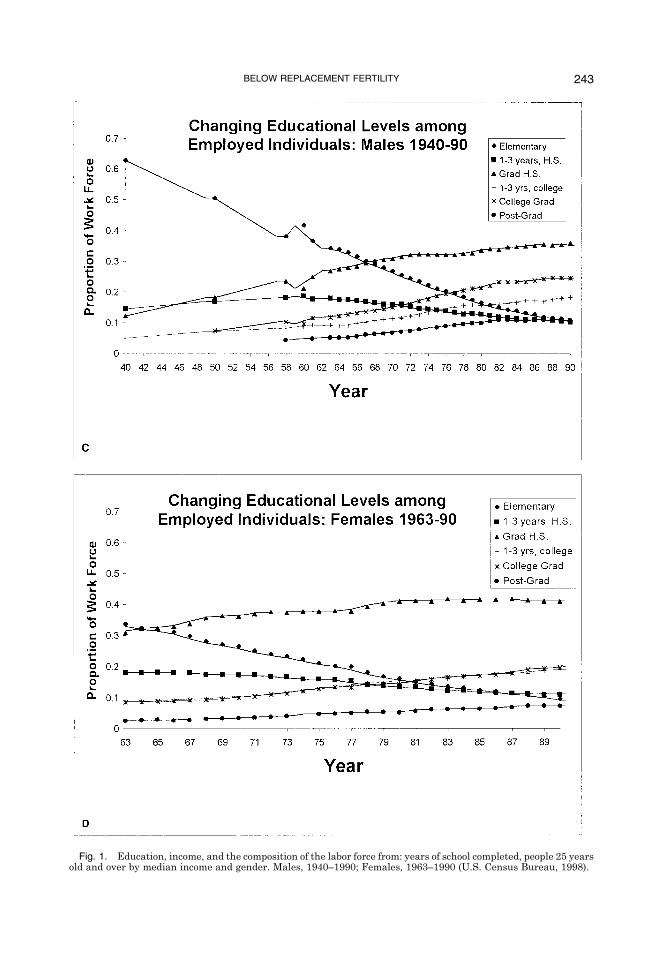
Fig. 1. Education, income, and the composition of the labor force from: years of school completed, people 25 yearsold and over by median income and gender. Males, 1940–1990; Females, 1963–1990 (U.S. Census Bureau, 1998).
BELOW REPLACEMENT FERTILITY 243

large variation in individual rates of returnto education. Those that perform poorly inschool and learn less per time spent inschool will be motivated to terminateschooling investments sooner. It is thisvariation in rates of return and perhaps inthe ability to finance the costs of highereducation (Becker, 1975) that is associatedwith variation in modern life course deci-sions and ultimately fertility.In addition, some macro-level changes
had large impacts specifically on the workforce participation of women. The increasein labor-saving devices in the home de-creased the value of household work. Theincreased efficiency of birth control allowedwomen to delay fertility and exert greatercontrol over the scheduling of reproduction.The increase in expected investments inchildren�s education led to a small targetfamily size and thus less lost time from theworkforce due to the care of young children(with the emergence of the daycare industryaugmenting this effect). All of those changesincreased the relative payoff to wage laborfor women and thus the payoffs to invest-ment in education. It is also hypothesizedthat the shift from the mother-homemaker/father-provider to the dual-earner familystructure decreased marital stability, be-cause women were no longer as financiallydependent on men and men were less de-pendent on homemakers.Finally, the model proposes that those
general changes in rates of return to edu-cation interact with the forces impingingspecifically on women in determining thelife history of fertility. First, increased in-vestment in own embodied capital leads to adelay in family formation for educated menand women. Second, those who invest morein their own embodied capital are also likelyto get higher returns on their investment inchildren�s embodied capital and to investmore as a result. Marital stability is hy-pothesized to be of greater importance topeople if they expect to invest more in theiroffspring, because of both the high costs ofcollege education (requiring two incomes)and the time investments in school-agedchildren. This logic implies that those whoinvest more in their own education will re-spond to the increased risks of marital dis-solution by taking longer to choose asuitable partner (i.e., being more selective)and by increasing the time from marriage toreproduction as a further trial period. Those
delays due to time spent in education andtraining and greater selectivity in matechoice result in some couples never achiev-ing their target fertility, because of thephysiological decrease in female fecunditywith age.Thus, the model proposes that below-re-
placement fertility is primarily a phenome-non of the most educated sector of oursociety and that it is primarily due to delayrather than to a reduction in target fertilityto a below replacement level (Coleman,1996; Tucker et al., 2001). The combinationof delay and increased efficiency of birthcontrol means that more people fail toachieve their target fertility (due to exces-sive delay) than those who exceed it (due tounwanted pregnancies). Fundamental tothis model is the notion that changingpayoffs to investments in education are thedriving force in the movement to below-re-placement fertility augmented by reducedpayoffs to household production and in-creased efficiency and reduced costs of ef-fective birth control.
EMPIRICAL RESULTS
This section of the paper examines howthe fertility behavior of the U.S. populationand the relationship between education andfertility has changed over time. Three birthcohorts (born before 1920, between 1920–1941, and between 1942–1970) are exam-ined, because they correspond roughly tothose reproducing prior to, during, and af-ter the ��Baby Boom��. It is this last cohortthat exhibits below-replacement fertility, atleast among those with higher educationalattainments. Two sets of data are used inthe analysis. The first set is a study of malefertility and parenting conducted by inter-viewing a representative sample of menfrom Albuquerque, New Mexico. The secondis a national probability sample of men andwomen, the National Survey of Familiesand Households.
Albuquerque men and the national survey offamilies and households data sets1
Albuquerque men. The research designfor the Albuquerque Men sample consistedof two complementary interviews, a shortinterview administered to a large repre-sentative sample, and a longer interview
244 H. KAPLAN ET AL.

administered to a subset of the men in theshort interview sample. Between July 1990and July 1993, 7,107 short interviews and1,325 long interviews were conducted. Thetotal short interview sample included 2789Hispanics, 3762 Anglos, and 556 individualsbelonging to other ethnicities. The long in-terview sample consisted of 401 Hispanics,858 Anglos, and 66 others. In New Mexico,the term, ��Hispanic,�� is preferred over��Latino,�� because most people of Hispanicorigin consider themselves native NewMexicans, tracing their family history as farback as living memory to residence in thisarea. In our sample, very few Hispanicswere of recent Mexican origin, and evenfewer were from other Latin countries. An-glo (an ethnic classification peculiar to theSouthwestern U.S.) refers to non-Hispanicmen who classify themselves as white andspeak English rather than Spanish in thehome. Details of the sampling method andthe content of the interviews can be foundin Lancaster and Kaplan (2000).
National survey of families and house-holds. The National Survey of Familiesand Households (NSFH) is a study fundedby the Center for Population Research ofthe National Institute of Child Health andHuman Development with the explicit pur-pose of providing a data set for researchersfrom a broad range of disciplines and theo-retical perspectives. Two rounds of thesurvey were conducted. We analyzed datacollected in the first round (NSFH1) only.The data set consists of a probabilisticsample of 13,007 individuals (men andwomen) from the U.S. Houses were selectedrandomly within stratified sampling blocksand a primary respondent was selectedrandomly from among a sampled house-hold�s permanent occupants. The samplingprocedure, which focused on households, isrobust for the purpose of generalizing to thenational population. Some deficiencies ex-ist, however. In particular, the sample ex-cludes entirely certain populations, such asthe homeless and the institutionalized (i.e.,the sick, insane, incarcerated, and, impor-tantly for this study, the elderly living innursing homes). This sampling techniqueresulted in a nationally representative pri-mary respondent sample of 5,226 men and7,781 women [see Sweet and associates(1988) for details].
Results
Education and income. Figure 1 displaysthe changing relationship between educa-tion and income (in 1997 dollars) between1940 and 1990 for men and 1963 and 1990for women and the changing educationalattainments of U.S. workers, as depicted inthe model (Bureau, 1998). Figure 1, A and Bshow the increase in the wage differentialsassociated with different education and in-come levels over time, especially for men(Fig. 1A). For men without high school de-grees, real wages actually dropped from1958 to 1990. In 1958, men with graduateeducation earned approximately 2.3 timesas much as men with elementary education;by 1990, they earned more than 3.5 times asmuch. Wage differentials among men withsome college education, bachelor�s degrees,and graduate degrees also increased sub-stantially. For women, wage differentialsamong educational attainment levels in-creased substantially in the 1980s.Figure 1, C and D show the changing
educational make-up of the U.S. workforce,presumably in response to the changingpayoffs to education. In 1958, 56% of themale workforce was composed of individualswith less than high school education (38%with only elementary education), whereasonly 16% had college degrees or higher ed-ucation. By 1990, the percentage of themale workforce with less than high schoolhad dropped to 20%, whereas those withcollege degrees increased to 25%. The sametrends are true for women (Fig. 1D). Also,during the same period, the percentage ofthe workforce composed of women increasedsubstantially from 40% in 1963 to 51% in1990.
Fertility. Our fundamental hypothesis isthat excessive delay in the onset of repro-duction, coupled with a target fertility oftwo is largely responsible for below re-placement fertility and that this delay isrelated to investments in own and offspringeducation. Figure 2 shows the changingpatterns of marriage age, onset of repro-duction, and education over the course ofthe twentieth century for the cohorts rep-resented in the NSFH1 sample.Holding education constant, the median
age at marriage and the delay from mar-riage to reproduction was higher for both
BELOW REPLACEMENT FERTILITY 245
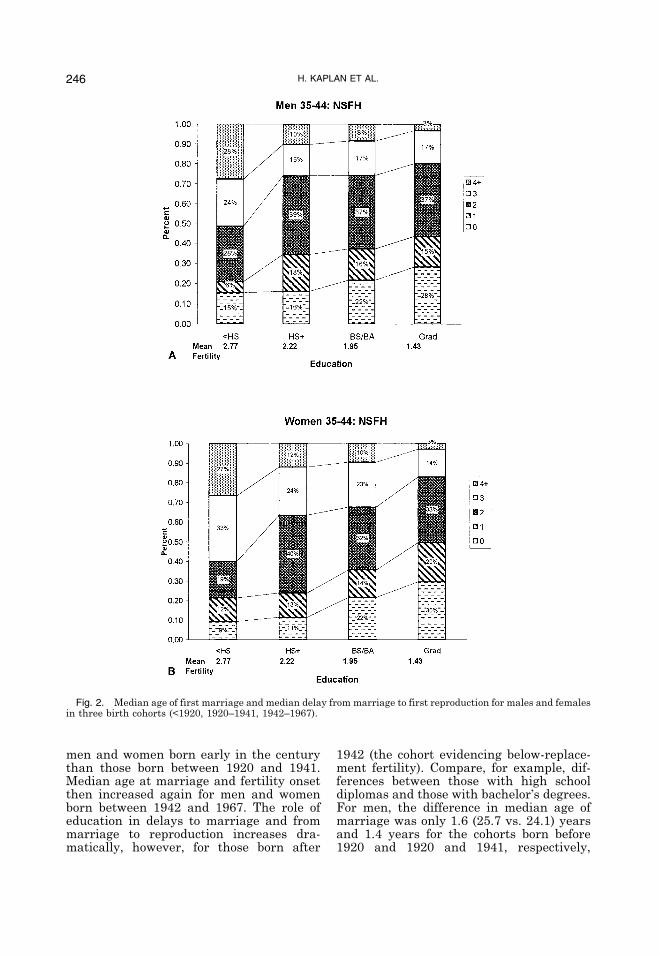
men and women born early in the centurythan those born between 1920 and 1941.Median age at marriage and fertility onsetthen increased again for men and womenborn between 1942 and 1967. The role ofeducation in delays to marriage and frommarriage to reproduction increases dra-matically, however, for those born after
1942 (the cohort evidencing below-replace-ment fertility). Compare, for example, dif-ferences between those with high schooldiplomas and those with bachelor�s degrees.For men, the difference in median age ofmarriage was only 1.6 (25.7 vs. 24.1) yearsand 1.4 years for the cohorts born before1920 and 1920 and 1941, respectively,
Fig. 2. Median age of first marriage and median delay from marriage to first reproduction for males and femalesin three birth cohorts (<1920, 1920–1941, 1942–1967).
246 H. KAPLAN ET AL.

whereas the difference increased to 2.2years for those born after 1942. Similarly,for women, the age difference increasedfrom 2.4 and 2.9 years for the first two co-horts to 3.7 years for the last cohort.Delays from marriage to reproduction
show the same directional effects, but evenmore dramatically. The difference betweenmen with high school diplomas and thosewith bachelor�s degrees was only 0.8 and 0.2years for the first two cohorts and rose to2.3 years for the last cohort. For women, thedifference rose from 0.9 and 0.7 years forthe older two cohorts to 2 years for the lastcohort. The sharpest contrast can be seen incomparison between those with the least tothose with the most education. While delaysto both marriage and reproduction in-creased for all men in the youngest cohortwith high school diplomas or more educa-tion, they actually decreased for high schooldropouts. Among men born after 1942,there is a 4.7-year difference in median ageat marriage and 3.6-year difference in me-dian delay from marriage to reproduction(a total of 8.3 years) between high schooldropouts and those with graduate and pro-fessional degrees. Among women of thesame cohort the corresponding differencesbetween high school dropouts and thosewith graduate and professional degrees are6 years in median age of marriage and 4.5years from marriage to reproduction for atotal of 10.5 years.These delays have powerful effects on
completed fertility for both men and wom-en. For men over 40 years of age, the prob-ability of having no children increases from7% for men who marry or cohabit before theage of 25 to 82% for men who marry afterage 40. The probability of having three ormore children decreases from 54% for menwho marry or cohabit before the age of 25 toclose to zero for men who marry after age40. For women, the probability of having nochildren increases from 5% for women whomarry or cohabit before the age of 25 to 88%for women who marry after age 40. Theprobability of having three or more childrendecreases from 55% for women who marryor cohabit before the age of 25 to close to 8%for women who marry after age 40.The differential impacts of education on
delay across the cohorts are also evidentwhen completed fertility is examined. Forthe older cohorts, the effect is modest. Formen over age 45, the probability of nonre-
production increases from 16% for men withless than high school to 19% for men withgraduate degrees. The probability of havingthree or more children decreases from 48%for men with less than high school to 40%for men with graduate degrees. For women,the probability of nonreproduction increas-es from 11% among those with less thanhigh school to 32% for those with graduatedegrees. The probability of having three ormore children decreases from 55% forwomen with less than high school to 20% forwomen with graduate degrees.The effects of education on fertility com-
pleted by the time of the interview arestronger for the younger cohorts, reproduc-ing in the last 20 years. Figure 3 shows thatmen 35 to 44 years of age with less thanhigh school have a mean of 2.6 children,whereas men with graduate degrees havean average of 1.5 children. Only 21% of highschool dropouts have zero or one child, ris-ing to 34% for those with high school de-grees, 38% for those with bachelor�sdegrees, and 43% for those with graduatedegrees. Conversely, 52% of high schooldropouts have three or more children,dropping to 26% for those with high schooldegrees, 25% for those with bachelor�s de-grees, and 20% for those with graduate de-grees.The effects appear to be somewhat
stronger among women 35 to 44 years ofage. High school dropouts have a mean of2.77 children, dropping to 2.22 for thosewith high school degrees, 1.95 for those withbachelor�s degrees, and 1.43 for those withgraduate degrees. Again, only 21% of highschool dropouts have zero or one child, ris-ing to 24% for those with high school de-grees, 36% for those with bachelor�sdegrees, and 50% for those with graduatedegrees. Finally, 60% of high school drop-outs have three or more children, droppingto 36% for those with high school degrees,33% for those with bachelor�s degrees, and17% for those with graduate degrees. Al-though there may be some catch-up amongthe more educated as this cohort ages, es-pecially for men, all of the evidence suggeststhat the effects of education on male andfemale fertility have increased dramaticallyduring this period of below replacementfertility.The data from the Albuquerque men
study reveal a similar pattern of results.For both Anglos and Hispanics, education
BELOW REPLACEMENT FERTILITY 247
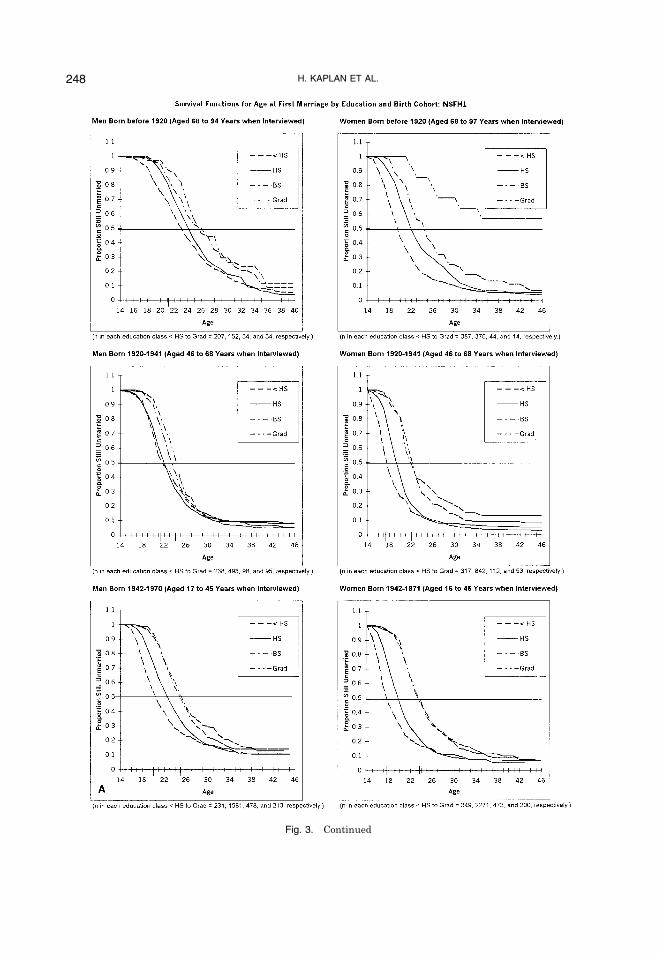
Fig. 3. Continued
248 H. KAPLAN ET AL.
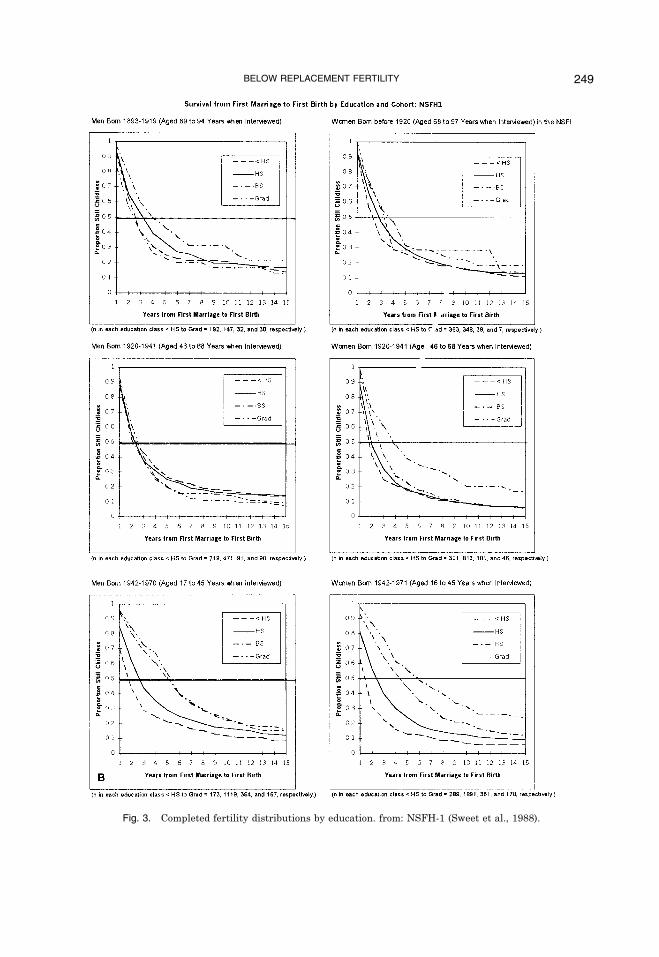
Fig. 3. Completed fertility distributions by education. from: NSFH-1 (Sweet et al., 1988).
BELOW REPLACEMENT FERTILITY 249
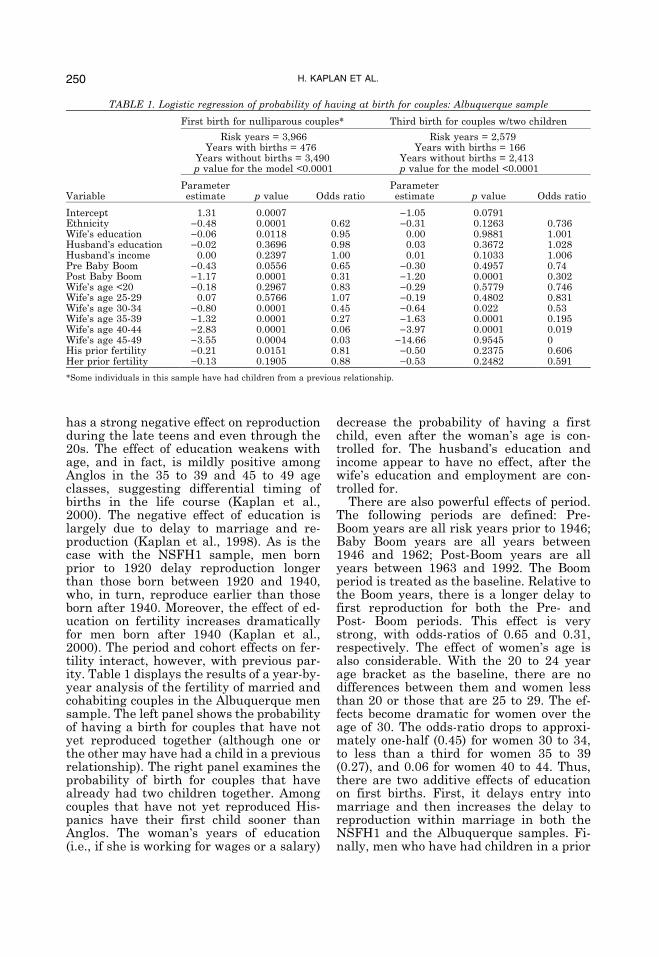
has a strong negative effect on reproductionduring the late teens and even through the20s. The effect of education weakens withage, and in fact, is mildly positive amongAnglos in the 35 to 39 and 45 to 49 ageclasses, suggesting differential timing ofbirths in the life course (Kaplan et al.,2000). The negative effect of education islargely due to delay to marriage and re-production (Kaplan et al., 1998). As is thecase with the NSFH1 sample, men bornprior to 1920 delay reproduction longerthan those born between 1920 and 1940,who, in turn, reproduce earlier than thoseborn after 1940. Moreover, the effect of ed-ucation on fertility increases dramaticallyfor men born after 1940 (Kaplan et al.,2000). The period and cohort effects on fer-tility interact, however, with previous par-ity. Table 1 displays the results of a year-by-year analysis of the fertility of married andcohabiting couples in the Albuquerque mensample. The left panel shows the probabilityof having a birth for couples that have notyet reproduced together (although one orthe other may have had a child in a previousrelationship). The right panel examines theprobability of birth for couples that havealready had two children together. Amongcouples that have not yet reproduced His-panics have their first child sooner thanAnglos. The woman�s years of education(i.e., if she is working for wages or a salary)
decrease the probability of having a firstchild, even after the woman�s age is con-trolled for. The husband�s education andincome appear to have no effect, after thewife�s education and employment are con-trolled for.There are also powerful effects of period.
The following periods are defined: Pre-Boom years are all risk years prior to 1946;Baby Boom years are all years between1946 and 1962; Post-Boom years are allyears between 1963 and 1992. The Boomperiod is treated as the baseline. Relative tothe Boom years, there is a longer delay tofirst reproduction for both the Pre- andPost- Boom periods. This effect is verystrong, with odds-ratios of 0.65 and 0.31,respectively. The effect of women�s age isalso considerable. With the 20 to 24 yearage bracket as the baseline, there are nodifferences between them and women lessthan 20 or those that are 25 to 29. The ef-fects become dramatic for women over theage of 30. The odds-ratio drops to approxi-mately one-half (0.45) for women 30 to 34,to less than a third for women 35 to 39(0.27), and 0.06 for women 40 to 44. Thus,there are two additive effects of educationon first births. First, it delays entry intomarriage and then increases the delay toreproduction within marriage in both theNSFH1 and the Albuquerque samples. Fi-nally, men who have had children in a prior
TABLE 1. Logistic regression of probability of having at birth for couples: Albuquerque sample
First birth for nulliparous couples* Third birth for couples w/two children
Risk years = 3,966 Risk years = 2,579Years with births = 476 Years with births = 166
Years without births = 3,490 Years without births = 2,413p value for the model <0.0001 p value for the model <0.0001
VariableParameterestimate p value Odds ratio
Parameterestimate p value Odds ratio
Intercept 1.31 0.0007 )1.05 0.0791Ethnicity )0.48 0.0001 0.62 )0.31 0.1263 0.736Wife�s education )0.06 0.0118 0.95 0.00 0.9881 1.001Husband�s education )0.02 0.3696 0.98 0.03 0.3672 1.028Husband�s income 0.00 0.2397 1.00 0.01 0.1033 1.006Pre Baby Boom )0.43 0.0556 0.65 )0.30 0.4957 0.74Post Baby Boom )1.17 0.0001 0.31 )1.20 0.0001 0.302Wife�s age <20 )0.18 0.2967 0.83 )0.29 0.5779 0.746Wife�s age 25-29 0.07 0.5766 1.07 )0.19 0.4802 0.831Wife�s age 30-34 )0.80 0.0001 0.45 )0.64 0.022 0.53Wife�s age 35-39 )1.32 0.0001 0.27 )1.63 0.0001 0.195Wife�s age 40-44 )2.83 0.0001 0.06 )3.97 0.0001 0.019Wife�s age 45-49 )3.55 0.0004 0.03 )14.66 0.9545 0His prior fertility )0.21 0.0151 0.81 )0.50 0.2375 0.606Her prior fertility )0.13 0.1905 0.88 )0.53 0.2482 0.591
*Some individuals in this sample have had children from a previous relationship.
250 H. KAPLAN ET AL.

relationship are significantly less likely toreproduce again than men who have not.The result is not significant for women.The right panel shows a similar pattern of
effects in the probability of progressingfrom a parity of two to a parity of three.There are two major differences, however.First, wife�s education has no direct effecton the probability of giving birth, after ageis controlled for. However, there are sharpdeclines in fertility with age of wife. Thus,the effects of education on progression tohigher parities are indirect, operatingthrough the age of the wife. Second, theperiod prior to the Boom is not significantlydifferent from the Baby Boom years, but thePost-Boom years evidence a sharply de-creased probability of progressing from twoto three children, with an odds ratio of 0.02.This reveals an increased tendency to stopat two children in the later cohorts.Our interpretation of these results from
the NSFH1 and Albuquerque men data setsis that the Great Depression of the 1930sand the World Wars delayed fertility for allmen and women who survived to their late1980s or so. During the post-war BabyBoom, low-cost loans for affordable housing,full employment, and the GI bill allowedmen and women to reproduce at earlierages, even though investment in educationwas increasing. Following the Baby Boom,the onset of fertility is again delayed foreverybody, but especially for men andwomen investing more in human capital.These reflect increasing costs of educationas well as increasing importance of educa-tion in the determination of wages (Burck,1976; Herrnstein and Murray, 1994; New-comer, 1955; Vinovskis, 1994). Those effectswere particularly important for women,because greater commitment to career im-plied greater opportunity costs associatedwith reproduction.
INVESTMENT AND OUTCOMES
Our fundamental hypothesis is that de-lays to marriage and reproduction are dueto both investments in own embodied capi-tal and expected investments in offspringembodied capital. We proposed that theexpected investments in offspring wouldinteract with the increased risk of divorce inlater cohorts, with those who expected toinvest more being more reluctant to repro-duce until they were sure of a secure, viable
marriage. We have no direct evidence thatthis reluctance results in a greater selec-tivity in mate choice and a longer latencyperiod from marriage to reproduction.However, there is evidence that the educa-tional status of both men and women affecttime and monetary investments in children,divorce rates when children are young, andultimately, the educational outcomes ofchildren. An analysis of men�s time invest-ments in children during their elementaryschool years showed significant positive ef-fects of a man�s education on both timespent alone with children and their in-volvement in the child�s educational pro-gress (Kaplan et al., 2000).The effects of education on divorce when
children are young are striking. Figure 4[adapted from Lancaster and Kaplan(2000)] shows the probability that a manwill cease to live with a child before the ageof 6 as a function of the child�s birth cohortand its father�s education. The increase indivorce rates is clearly evident in this fig-ure. However, there is a very interestinginteraction effect between a man�s educa-tion and the child�s birth cohort on theprobability of separation between a childand its father. Among children whose fatherhad less than high school education, thepercent of children who ceased to live withhim before age 6 rose from approximately5% for children born before 1960 to 23% forchildren born in the 1980 cohort. Amongchildren whose father had a graduate de-gree, the percent of children who ceased tolive with him before age 6 did not changesignificantly during this period, and re-mained between 5% and 8%.
Fig. 4. Effects of Father�s education on the proba-bility of divorce when children are young.
BELOW REPLACEMENT FERTILITY 251

Table 2 shows the results of three analysesexamining investments in older childrenand their educational outcomes from theAlbuquerque men sample. Column A showsthe probability of a child over age 18 at-tending college. Anglos are more likely toattend college. The father�s income and ed-ucation are also positively associated withcollege attendance, as is the mother�s edu-cation. Number of stepsiblings, but not bio-logical siblings, is negatively associated withcollege attendance. Parental divorce andseparation have a very strong negative ef-fect on college attendance. Column B showsthe probability of the child receiving finan-cial support for her college education fromher father, given that she attended college.Again, we find that Anglos are more likely toprovide financial support and that father�seducation and income (but not mother�s ed-ucation) are positively associated with sup-port for college. Number of both genetic andstep siblings is negatively associated withsupport. Parents who remain together arealso more likely to provide support.Thus, both marital stability and educa-
tion have strong effects on investments inoffspring and on their educational achieve-ments. Age at marriage and the educationallevels of both men and women are bothstrong predictors of marital stability andcompleted fertility (Wineberg, 1990).
DISCUSSION
This paper has examined the very lowfertility exhibited in recent years in light ofhuman evolutionary history. It is clear thatcurrent fertility behavior does not maximizefitness (Kaplan et al., 1995), but an under-standing of evolutionary history can helpilluminate and predict those patterns ofdeviation from fitness maximization. It wasargued that as a result of human evolu-tionary history, human parents investheavily in the embodied or human capital ofoffspring. The large human brain, the longperiod of juvenile dependence, long lifespan, and parental and post-reproductivesupport of reproduction are the co-evolutionary result of a niche based on skill-intensive techniques of resource accrual.The high level of investment in embodied
capital affects virtually all the life-historycharacters typical of our species. Growth isboth slow and prolonged. Reproduction isdelayed as a result. Increases in productiv-ity continue into mid-adulthood, well afterreproduction has begun, and people remainquite productive well into old age after re-production has ceased. Adult mortalityrates are extremely low, and aging occursvery slowly. Children are provisioned formany years after weaning, and parentssupport multiple, dependent young. The
TABLE 2. Determinants of college attendance and parental support for higher education
A. Probability ofchild attending
college
B. Probability ofchild receivingmoney for college
C. Log of amountof money child
received�
Full sample College sample Funded sample
Logistic regressionN = 1947, v2 = 439.0,df = 7, p < 0.00001
Logistic regressionN = 1132, v2 = 152.7,df = 8, p < 0.00001
OLS regressionN = 637, R2 = 0.201,
F6,630 = 26.4, p < 0.00001
VariableParameterestimate
Partialp
Parameterestimate
Partialp
Parameterestimate
Partialp
Intercept )3.66 0.000 )0.80 0.090 )5.93 0.000Child�s age (censored at 30) 0.04 0.041 )0.44 0.074 0.10 0.000Respondent is Anglo 0.44 0.000 0.54 0.001 — nsRespondent�s income*,� 0.08 0.000 0.11 0.000 0.08 0.000Respondent�s education 0.11 0.000 0.11 0.000 — nsChild�s mother�s education 0.14 0.000 — ns 0.07 0.000No. of genetic kids respondenthas by current mate*
–– ns )0.21 0.000 )0.11 0.000
No. of step kids respondenthas by current mate*
)0.31 0.005 )0.67 0.000 — ns
Respondent and child’s motherare together
0.66 0.000 1.38 0.000 0.82 0.000
*Evaluated when offspring was age 18.�In tens of thousands of 1990 dollars.
252 H. KAPLAN ET AL.

high productivity of males in the provi-sioning of meat and of postreproductivepeople results in a three-generational sys-tem of resources flows, in which bothgrandparents and men assist women in theproduction and support of offspring.The regulation of fertility under tradi-
tional conditions is based upon a co-evolvedpsychology and physiology. The fundamen-tal features of the evolved psychology af-fecting fertility are the adjustments ofinvestment in offspring depending upon thereturns to skill and mortality hazards.When all wealth is somatic, the hormonalsystem controlling ovulation and implanta-tion translates income into genetic de-scendants. When some wealth is extra-somatic, there can be a conflict betweenconsciously desired fertility and the fertilityoutput potential of the hormonal system,leading to the practice of birth control.This commitment to intensive investment
in embodied capital (both for self and off-spring) also appears to be related to re-sponses to modernization. Changingtechnologies of production and improvedpublic health may have interacted to in-crease the payoffs to investments in skilland education, investments in health andlongevity, and investments in child quality.Although such increases in investmentsmay be a predictable consequence of ourevolutionary heritage, they may not beadaptive in the strict sense of being fitnessmaximizing. Thus, the demographic tran-sition may be the result of the interaction ofthe evolved psychology of parental invest-ment and investment in own embodiedcapital with modern conditions in whichmost wealth is held extra-somatically.In the last three decades, education has
become an increasingly important determi-nant of wage rates, fertility behavior, andparental investment. As a result of invest-ment in own education, fertility can be de-layed enough that target fertility is notreached, because of age-specific physiologi-cal reductions in fertility. In that sense,there may be a mismatch between socialmaturity and physiological maturity andreproductive senescence. Most people desireonly two children and exhibit a strong ten-dency to stop reproducing after the birth oftheir second child (Coleman, 1996). In thecontext of effective birth control, below re-placement fertility may be the result of anincrease in individuals involuntarily falling
short of their ideal due to excessive delayand a decrease in those involuntarily ex-ceeding their ideal due to failed birth con-trol.Although the results presented here do
not constitute tests of this hypothesis, theyshow the dramatic effects of investment inown and offspring embodied capital on fer-tility. Investments in own education delaymarriage and delay the onset of fertilityfollowing marriage. These effects haveincreased dramatically in the most recentcohorts, who are displaying below-replace-ment fertility. Individuals pursuing differ-ent levels of education are increasinglydivergent in the timing of family formationand as a result, in completed fertility. Theyare also increasingly divergent in parentalinvestment and marital stability. Thesedifferences directly impact the achievededucation of their children.A particularly striking finding of the
present study is that education is not onlyincreasingly associated with fertility anddelays to marriage and reproduction amongwomen but among men as well. This is im-portant because it suggests that we mustunderstand why men choose to delay as welland why educational homogamy is on therise (Mare, 1991; Oppenheimer, 1988). Menand women appear to be matching in termsof investments in own and children�s capitaland in the delays to marriage and repro-duction. It does not appear that femaleempowerment (Handwerker, 1993) and theopportunity costs of women�s time (Desaiand Waite, 1991) are the only, or evenprincipal, determinants of low fertility.If the hypothesis is correct that below-
replacement fertility is largely the result ofpeople failing to achieve target fertility be-cause of delay in reproduction interactingwith physiological declines in fertility withage, what can we expect for the future? Willadvances in infertility treatment reduce thenumber of people exhibiting below replace-ment fertility and how large will be the ef-fect? Will observations of older peopleexperiencing fertility problems lead young-er people to marry and reproduce earlier? Ifthe model is correct, we should expect to seesome reversals of the effects of education onbelow-replacement fertility. To detect thoseeffects, it will be necessary to distinguishthe effects of education on delay from theself-selection effects of delay on education(i.e., those failing to marry and reproduce
BELOW REPLACEMENT FERTILITY 253

continuing longer with education as a re-sult).Even if this model is correct, we are still
left with the question of why those socialand economic changes and differential in-vestments in capital have such large effectson fertility. We still need a better under-standing of the psychological and culturalprocesses underlying decisions about repro-duction and the expenditures of resourceson consumption goods. Our hypothesis isthat extra-somatic wealth is the principalnovelty to which our evolved psychologyresponds by desiring less than fitness-maximizing fertility. But, why are the ef-fects so strong? One possible hypothesis isthat social dynamics of small groups inhunting and gathering economies resultedin greater fitness for those of higher socialstanding and selected for a psychology inwhich relative social position of self andoffspring is valued highly. There is evidencethat, despite the fact that meat is shared inmost hunter/gatherer communities, thechildren of good hunters fare better thanothers, perhaps due to preferential treat-ment by group members (Hill and Hurtado,1996; Kaplan and Hill, 1985). Such a psy-chology would also emerge if people judgedthe well being of themselves and theirchildren based upon the wealth and con-sumption of others. This psychology mayhave been fitness maximizing under tradi-tional conditions. If relative, as opposed toabsolute, wealth and social standing guidehuman decisions regarding wealth flows,parental investment, and fertility, it ispossible that ��run-away�� consumption andinvestment in children�s education resultfrom the interaction of this psychology andmodern education-based labor markets andconsumption possibilities (Frank, 1985).In modern society, people face a tremen-
dous array of consumption goods, includinghousing, clothing, electronic equipment,vehicles, etc. The medium of exchange forobtaining those goods is money. Money ishighly fungible in that it can be translatedinto any one of those goods. As diminishingreturns to consumption of one category isreached, money can then be allocated toanother. With all the goods available, thereis always some good of which little has beenconsumed and returns to purchasing someamount of it are still high. The same can besaid for investment in offspring�s embodiedcapital, which appears to include not only
investments in schooling but in items asso-ciated with social training and social status,such as hobbies, sports, clothing, and toys.Much of these investments function ascommitments to favorable eventual place-ment of children in the mating market. Ourbest guess at this time is that the low fer-tility exhibited in modern societies is due tothe combined effects of lowered mortality,higher payoffs to investments in offspringincome, a perceived lack of diminishing re-turns to other forms of consumption, andincreasingly effective birth-control technol-ogies.If the existence of extra-somatic wealth is
the critical condition to which our evolvedproximate mechanisms do not respond so asto maximize fitness, it may be that devia-tions from fitness maximization are likely tobe observed in other contexts as well. Whenthere is heritable wealth, such as cattle orland, the breast-feeding/energy-balancesystem may generate higher fertility thanparents� desire. Adjustments to this situa-tion may involve primarily differential in-heritance, such as primogeniture andillegitimacy (Mace, 1998) but may also in-clude late age of marriage (Coale andTreadway, 1986) or even celibacy (Boone,1995) and lowered rates of polygyny bywealthy men (Luttbeg et al., 2000). Thus,perhaps we should not be surprised to finddeviations from fitness maximization soonafter there are forms of extra-somaticwealth and social stratification. We are onlybeginning to investigate the evolved psy-chology and physiology of human repro-duction and parental investment and theirinteraction with changing economic, social,and cultural conditions.
LITERATURE CITED
Bailey RC, Jenike MR, Ellison PT, Bentley GR, Harri-gan AM, Peacock NR. 1992. The ecology of birthseasonality among agriculturalists in Central Africa.Biosoc Sci 24:393–412.
Becker GS. 1975. Human capital. New York: ColumbiaUniversity Press.
Becker GS. 1981. A treatise on the family. Cambridge,MA: Harvard University Press.
Becker GS, Lewis HG. 1973. Interaction betweenquantity and quality of children. In: Schult TW, edi-tor. Economics of the family: marriage, children, andhuman capital. Chicago: University of Chicago Pressfor National Bureau of Economic Research. p 81–90.
Bird DW, Bliege Bird RB. 2001. Children on the reef:slow learning or strategic foraging. Hum Nat 12:
Bliege Bird RB, Bird DW. 2001. Constraints of knowingor constraints of growing? Fishing and collectingamong the children of Mer. Hum Nat 12:
254 H. KAPLAN ET AL.

Blurton Jones N, Marlowe FW. 2001. Selection for de-layed maturity: does it take 20 years to learn to huntand gather? Hum Nat 12:
Blurton Jones NG, Hawkes K, Draper P. 1994a. Dif-ferences between Hadza and !Kung children�s work:Original affluence or practical reason? In: Burch ES,Ellana L, editor. Key issues in hunter gatherer re-search. Oxford: Berg. p 189–215.
Blurton Jones NG, Hawkes K, Draper P. 1994b. Forag-ing returns of !Kung adults and children: why didn�t!Kung children forage? Anthropol Res 50:217–248.
Bock J. 2001. Learning, life history, and productivity:children�s lives in the Okavango Delta, Botswana.Hum Nat 12:
Bock JA. 1995. The determinants of variation in chil-dren�s activities in a southern African community.Doctrate Dissertation, University of New Mexico,Albuquerque.
Bongaarts J, Watkins SC. 1996. Social interactions andcontemporary fertility transitions. Pop Dev Rev22:639–682.
Boone J. 1995. Parental investment and elite familystructure in preindustrial states: a case study of latemedieval-early modern Portuguese genealogies. AmAnthropol 88:259–878.
Bound J, Johnson G. 1992. Changes in the structure ofwages in the 1980�s: an evaluation of alternative ex-planations. Am Econ Rev 82:371–392.
Burck CG. 1976. A group profile of the Fortune 500child executives. Fortune :173–177.
Bureau USC. 1998. Years of School Completed – People25 years old and over by median income and gender:1958-1998. Washington, DC: Department of Com-merce.
Caldwell J, Caldwell P. 1998. Regional paths to fertilitytransition. Conference on Global Fertility Transition.
Charnov EL. 1993. Life history invariants: some ex-planations of symmetry in evolutionary ecology. Ox-ford: Oxford University Press.
Coale AJ, Treadway R. 1986. A summary of thechanging distribution of overall fertility, marital fer-tility and the proportion married in the provinces ofEurope. In: Coale AJ, Watkins SC, editors. The de-cline of fertility in Europe. Princeton, NJ: PrincetonUniversity Press. p 31–181.
Coleman D. 1996. New patterns and trends in Euro-pean fertility: international and sub-national com-parisons. In: Coleman D, editor. Europe�s populationin the 1990s. New York: Oxford University Press. p 1–61.
Davis K, Bernstam MS, Ricardo-Campbell R, editors.1986. Below-replacement fertility in industrial socie-ties: causes, consequences, policies. New York: Pop-ulation and Development Review Supplement.
Desai S, Waite LJ. 1991. Women�s employment duringpregnancy and after the first birth: occupationalcharacteristics and work commitment. Am Sociol Rev56:551–566.
Ellison PT. 1990. Human ovarian function and repro-ductive ecology: new hypotheses. Am Anthropol92:933–952.
Ellison PT. 1994. Advances in human reproductiveecology. Ann Rev Anthropol 23:255–275.
Frank RH. 1985. Choosing the right pond: human be-havior and the quest for status. New York: OxfordUniversity Press.
Godfray HCJ, Partridge L, Harvey PH. 1991. Clutchsize. Ann Rev Ecol Systemat 22:409–429.
Handwerker WP. 1993. Empowerment and fertilitytransition on Antigua, WI: education, employment,and the moral economy of childbearing. Hum Organ52:41–52.
Hawkes K, O�Connell JF, Blurton Jones N. 1989.Hardworking Hadza grandmothers. In: Standen V,Foley RA, editors. Comparative socioecology of hu-mans and other mammals. London: Basil Blackwell.p 341–366.
Hawkes K, O�Connell JF, Blurton Jones N. 1995. Hadzachildren�s foraging: juvenile dependency, social ar-rangements and mobility among hunter-gatherers.Curr Anthropol 36:688–700.
Herrnstein RJ, Murray C. 1994. The bell curve: Intel-ligence and class structure in American life. NewYork: Free Press.
Hill K, Hurtado AM. 1996. Ache life history: The ecol-ogy and demography of a foraging people. Hawthorne,NY: Aldine.
Hurtado AM, Hill KR. 1987. Early dry season subsist-ence ecology of the Cuiva (Hiwi) foragers of Vene-zuela. Hum Ecol 15:163–187.
Kaplan H, Hill K. 1985. Hunting ability and reproduc-tive success among male Ache foragers. Curr An-thropol 26:131–133.
Kaplan H, Lancaster J, Bock J, Johnson S. 1995. Doesobserved fertility maximize fitness among New Mex-ican Men? A test of an optimality model and a newtheory of parental investment in the embodied capitalof offspring. Hum Nat 6:325–360.
Kaplan H, Lancaster JB, Anderson KG. 1998. Humanparental investment and fertility: The life histories ofmen in Albuquerque. In: Booth A, Crouter A, editors.Men in families. Mahwah, NJ: Erlbaum. p 55–109.
Kaplan HS. 1996. A theory of fertility and parentalinvestment in traditional and modern human socie-ties. Yrbk Phys Anthropol 39:91–135.
Kaplan HS. 1997. The evolution of the humanlife course. In: Watcher K, Finch C, editors. BetweenZeus and salmon: the biodemography of aging.Washington, DC: National Academy of Sciences. p175–211.
Kaplan HS, Hill K, Hurtado AM, Lancaster JB.2001. The embodied capital theory of human evolu-tion. In: Ellison PT, editors. Reproductive ecologyand human evolution. Hawthorne, NY: Aldine deGruyter. pp
Kaplan HS, Hill K, Lancaster JB, Hurtado AM. 2000. Atheory of human life history evolution: diet, intelli-gence, and longevity. Evol Anthropol 9:1–30.
Kaplan HS, Robson A. 2001. The embodied capitaltheory of brain-lifespan co-evolution. Albuquerque,NM: Department of Anthropology, University of NewMexico.
Knodel J, van de Walle E. 1979. Lessons from the past:policy implications of historical fertility studies. PopDev Rev 5:217–245.
Lancaster JB, Kaplan HS. 2000. Parenting other men�schildren: costs, benefits and consequences. In: Cha-gnon N, Irons W, editors. Adaptation and humanbehavior: an anthropological perspective. Hawthorne,NY: Aldine de Gruyter. p 173–195.
Lesthaeghe R, Williems P. 1999. Is low fertility a tem-porary phenomenon in the European Union? Pop DevRev 25:211–228.
Luttbeg B, Borgerhoff Mulder M, Mangel MS. 2000. Tomarry or not to marry? A dynamic model of marriagebehavior and demographic transition. In: Cronk L,Chagnon NA, Irons W, editors. Adaptation and hu-man behavior: an anthropological perspective. Haw-thorne, NY: Aldine de Gruyter.
Mace R. 1998. The coevolution of human fertility andwealth inheritance strategies. Phil Trans Roy SocLondon B 353:389–397.
Mare RD. 1991. Five decades of educational assortativemating. Am Sociol Rev 56:15–32.
BELOW REPLACEMENT FERTILITY 255

McDade TW. 2001 Parent-offspring conflict and thecultural ecology of breast feeding. Hum Nat 12:
Meltzer DO. 1992. Mortality decline, the demographictransition and economic growth. Chicago: Universityof Chicago.
Morgan P. 1996. Characteristic features of modernAmerican fertility. In: Casterline JB, Lee RD, FooteKA, editors. Fertility in the United States: new pat-terns, new theories. New York: Population and De-velopment Review Supplement. p 19–66.
Murphy K, Welsh F. 1989. Wage premiums for collegegraduates: recent growth and possible explanations.Ed Res 18:17–27.
Newcomer M. 1955. The big business executive: thefactors that made him. 1890–1950. New York: Co-lumbia University Press.
Oppenheimer VK. 1988. A theory of marriage timing.Am J Sociol 94:563–591.
Preston SH. 1980. Mortality declines in less developedcountries: causes and consequences. In: Easterlin RA,editor. Population and economic change in develop-ing countries. Chicago, IL: University of ChicagoPress.
Preston S, Haines M. 1991. Fatal years: child mortalityin the late nineteenth century America. Princeton,NJ: Princeton University Press.
Sellen DW, Smay DB. 2001. Relationship betweensubsistence and age at weaning in ��pre-industrial��societies. Hum Nat 12:
Smith CC, Fretwell SD. 1974. The optimal balance be-tween size and number of offspring. Am Natural108:499–506.
Sweet J, Bumpass L, Call V. 1988. The design andcontent of the National Survey of Families andHouseholds. Working Paper #1, University of Wis-consin-Madison.
Tracer DO. 1996. Lactation, nutrition, and postpartumamenorrhea in Lowland Papua-New Guinea. HumBiol 68:277–292.
Tucker WT, Kaplan H, Lancaster JB. 2001. A life his-tory theory of male childlessness. Hum Nat 12:
Vinovskis MA. 1994. Education and the economictransformation of Nineteenth Century America. In:Riley MW, Kahn RL, Foner A, editors. Age andstructural lag: Society�s failure to provide meaningfulopportunities in work, family, and leisure. New York:Wiley and Sons. p 171–196.
Werner EE. 1986. Amphibian metamorphosis: growthrate, predation risk and the optimal time to trans-form. Am Natural 128:319–341.
Wineberg H. 1990. Delayed childbearing, childlessnessand marital disruption. J Comp Fam Stud 21:99–110.
256 H. KAPLAN ET AL.




















