
Magnetic field affects meristem activity and cell differentiation in Zea mays roots
8
This article was downloaded by: [Univ Studi Della Calabria] On: 01 December 2011, At: 04:28 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tplb20 Magnetic field affects meristem activity and cell differentiation in Zea mays roots M. B. Bitonti a , S. Mazzuca a , T. Ting b & A. M. Innocenti a a Department of Ecology, University of Calabria b Department of Chemistry, University of Calabria, Rende, Italy Available online: 19 Aug 2006 To cite this article: M. B. Bitonti, S. Mazzuca, T. Ting & A. M. Innocenti (2006): Magnetic field affects meristem activity and cell differentiation in Zea mays roots, Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 140:1, 87-93 To link to this article: http://dx.doi.org/10.1080/11263500500511314 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Magnetic field affects meristem activity and cell differentiation in Zea mays roots

This article was downloaded by: [Univ Studi Della Calabria]On: 01 December 2011, At: 04:28Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Plant Biosystems - An International Journal Dealingwith all Aspects of Plant BiologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tplb20
Magnetic field affects meristem activity and celldifferentiation in Zea mays rootsM. B. Bitonti a , S. Mazzuca a , T. Ting b & A. M. Innocenti aa Department of Ecology, University of Calabriab Department of Chemistry, University of Calabria, Rende, Italy
Available online: 19 Aug 2006
To cite this article: M. B. Bitonti, S. Mazzuca, T. Ting & A. M. Innocenti (2006): Magnetic field affects meristem activity andcell differentiation in Zea mays roots, Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology,140:1, 87-93
To link to this article: http://dx.doi.org/10.1080/11263500500511314
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

ECOLOGY AND ECOPHYSIOLOGY
Magnetic field affects meristem activity and cell differentiationin Zea mays roots
M. B. BITONTI1, S. MAZZUCA1, T. TING2 & A. M. INNOCENTI1
1Department of Ecology, University of Calabria, and 2Department of Chemistry, University of Calabria, Rende, Italy
AbstractExposure of Zea mays seedlings to a continuous electromagnetic field (EMF) for 30 h induced a 30% stimulation in the rateof root elongation compared with the controls. It also resulted in a significant increase of cell expansion, in both the acropetal(metaxylem cell lineage) and basipetal (root cap cells) direction. In addition, in EMF-exposed roots a precocious structuraldisorder was observed both in differentiating metaxylem cells and root cap cells. All these features may be consistent with anadvanced differentiation of root cells that are programmed to die. EMF treatment also resulted in a significant reduction inthe size of the quiescent centre in the root apical meristem. The extent to which these responses are causally linked isdiscussed.
Abbreviations: EMF: electromagnetic field; MES: 2-[N-morpholino]ethanesulfonic acid buffer; QC: quiescentcentre; RAM: root apical meristem
Key words: Magnetic field, metaxylem differentiation, quiescent centre, root cap, root meristem
Introduction
Sessile plants are very sensitive organisms to
evaluate the effects of environment on growth and
development. As a consequence, growth responses to
different exogenous stimuli have been widely inves-
tigated, especially in plant roots (Barlow &
Rathfelder, 1985; Okamoto & Tatara, 1995; Crook
& Ennos, 1996; Goodman & Ennos, 1996). Exposure
of humans, animals and plants to different levels of
electromagnetic field becomes a constant of current
life, due to ever increasing employment of electrical
and technological devices. Notably, even more
evidence has been accumulated showing that electro-
magnetic fields can affect living organisms (Jenrow
et al., 1995; Blank & Goodman, 2002; Piatti et al.,
2002; Hood, 2004).
In plants, the first evidence of the influence of
magnetic field on early growth processes dealt with
the induction of cress root curvature in the seed by a
static magnetic field (Audus, 1960). Later on,
Brulfert et al. (1985) and Inoue et al. (1985) showed
that the growth rate of seedlings could be altered
following exposure to low 60 Hz magnetic field, with
strength being from 70 to 48 mT respectively. Other
evidence demonstrated that changes in root tropism
induced by magnetic field were triggered by intracel-
lular displacement of amyloplasts in the statocytes of
root cap (Kuznetsov & Hasenstein, 1996; Kuznetsov
et al., 1999). More recently, the effects of either
continuous or pulsed magnetic field on plant growth
and development have been investigated in a large
number of plant species (Yano et al., 2001; Es’kov
& Darkov, 2003; Kalinin & Boshkova, 2003).
Generally, the effects of exposure to magnetic field
include the modulation of root growth rates, the
increase in wet and dry weight, the induction of
primary root curvature. By contrast, very little
information exists on the effects induced by magnetic
field at the cellular level (Inoue et al., 1985; Stange
et al., 2002).
With a view to providing some new insights into the
influence of electromagnetic fields on root growth, in
the present work we selected Zea mays, a species with
a very well characterized root apical meristem. It is
well known that following specific treatments such as
X-radiation, or prolonged exposure to low tempera-
ture, root growth is guaranteed through the activation
of cell division in the QC (Clowes, 1972; Rost &
Van’t Hof, 1973; Webster & Langenauer, 1973;
Correspondence: M. B. Bitonti, Dipartimento di Ecologia, Universita della Calabria, I-87030 Arcavacata di Rende (CS), Italy. Tel.: þ39 0984 492965.
Fax: þ39 0984 492964. E-mail: [email protected]
Plant Biosystems, Vol. 140, No. 1, March 2006, pp. 87 – 93
ISSN 1126-3504 print/ISSN 1724-5575 online ª 2006 Societa Botanica Italiana
DOI: 10.1080/11263500500511314
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011

Barlow & Pilet, 1983; Muller et al., 1994). Thus, our
attention was mainly drawn to verify the effects
of exposure to a continuous strong magnetic field
on the quiescent centre (QC) of the root apical
meristem. Moreover, laser-ablation studies on roots
of Arabidopsis thaliana, have clearly shown that QC
cells play a key role in root growth by suppressing
differentiation of those cells which lie on its surface
(Van der Berg et al., 1997) and consequently by
controlling vascular bundle repatterning (Sabatini
et al., 1999). Hence we have explored whether the
magnetic field affected the size of QC and the extent
to which such alteration might be linked to changes in
cell expansions, in both acropetal (metaxylem cell
lineage) and basipetal (root cap cells) direction, as an
index of the differentiation process.
Materials and methods
Growth conditions within the magnet cavity
Maize seeds (Zea mays L., Pioneer HI-Bred), pre-
viously sterilized in 3% NaClO, were germinated in
Petri dishes on moistened paper, at 25+ 0.58C in
the dark. Two-day-old seedlings with primary roots
that were about 8 mm long were transferred into vials
and maintained using a perfusion system (Figure 1a),
which was designed to provide continuous nourish-
ment to the seedlings within the cavity of a Bruker
MSL300 NMR spectrometer magnet. The seedlings
were oriented parallel to the 7 Tesla electromagnetic
field (EMF) (see Figure 1b).
Root elongation and mitotic activity
Seedlings were transferred to two vials (n¼ 18 per
vial) containing Mes buffer (10 mM Mes þ 0.2 mM
CaSO4, pH 6). One vial was exposed to EMF for
30 h in darkness. During exposure, roots were
perfused through the peristaltic pump with 2 L of
MES buffer, which was renewed every 10 h. The rate
of perfusion was calibrated in order to maintain a
temperature of 25+ 1.08C in the buffer inside the
vial. The control vial was placed vertically outside the
magnet cavity, in a growth chamber in exactly
the same conditions as described above. Every 10 h,
the length of both EMF-exposed and control roots
was measured. At each sampling time, three roots
were collected, fixed in ethanol:acetic acid 3:1 (v/v)
and the apices were squashed for mitotic index
analysis. Another three roots were fixed in 4%
gluteraldeides in phosphate buffer, embedded in
LR White’s resin and 3mm longitudinal sections of
the root tip (4 mm in length) were stained with
Coomassie Blue for histological analyses. Mitotic
index was estimated by scoring 150 nuclei for each
control and EMF-exposed root. For the histological
analyses, the size (mean area) of root cap cells, the
cell length and the cell number along the metaxylem
cell lineage were measured on median longitudinal
Figure 1. (a) Vial containing seedlings perfused through a peristaltic pump with recycling buffer inside and outside the vial; (b) seedling
orientation in the electromagnetic field. N: North pole of field.
88 M. B. Bitonti et al.
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011
sections of each control and EMF-exposed root by
means of the NIH Image analysis system (National
Institute of Health, Maryland, US). The entire ex-
periment was replicated three times.
Continuous label (CL) with 3H-TdR for QC
visualization
CL was carried out in the EMF and control treatments
by exposing eight seedlings to an oxygenated solu-
tion of tritiated thymidine ([methyl-3H]thymidine,
925 Gbq mmol71, 9.25 104 Bq cm73) in 10 mM MES
for 30 h. Three different replicate experiments were
carried out. At the end of each CL treatment, the
length of both EMF-exposed and control roots
was measured, thereafter the roots were fixed in
ethanol:acetic acid 3:1 (v/v). For the quiescent centre
measurements, the roots (n¼ 4 for each replicate) of
both EMF and controls were wax-embedded and
sectioned longitudinally at 7mm. Sections were
Feulgen stained after hydrolysis in 1M HCl at 608Cfor 7 min and thereafter autoradiographed using Ilford
L4 emulsion diluted 1:1 (w/v) with a 1-month
exposure time. The size of quiescent centre (QC)
was measured from median longitudinal sections as
described by Feldman & Torrey (1975); roots (n¼ 12)
from the different replicates were analyzed and the
number of cells in the QC estimated by scoring the
most median longitudinal sections.
Results
Root length and mitotic activity
Following the selection of primary roots that were
about 8 mm long, the initial aim was to establish
whether or not EMF affected root elongation and
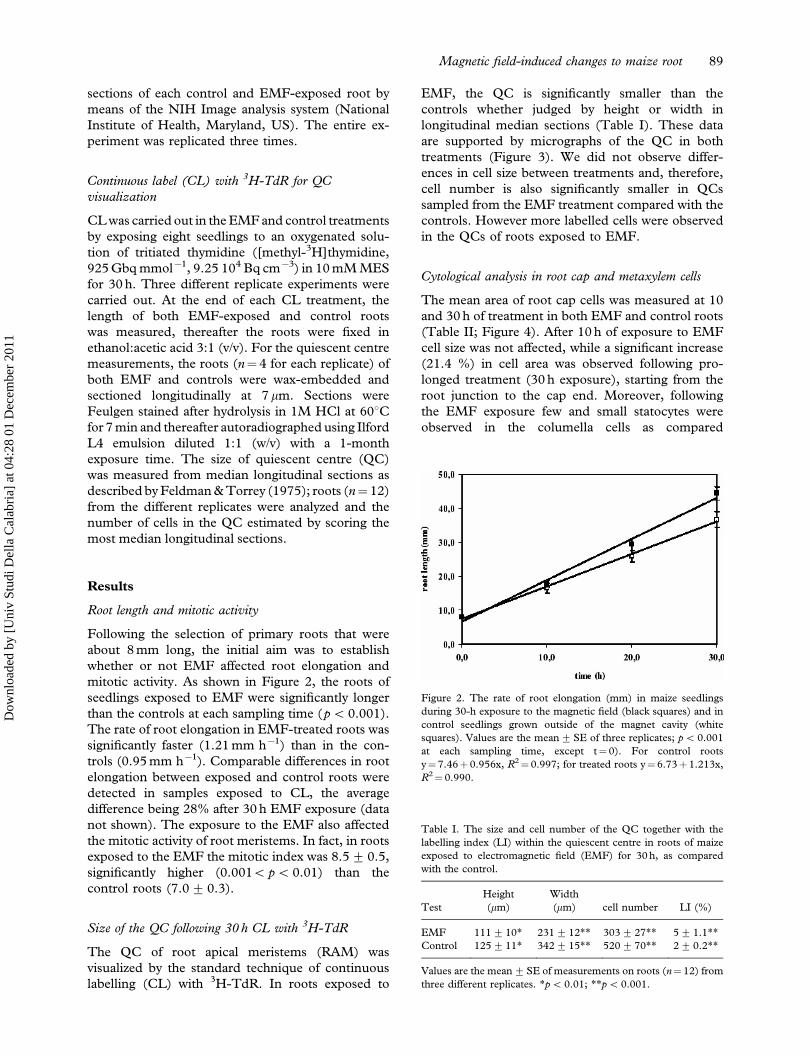
mitotic activity. As shown in Figure 2, the roots of
seedlings exposed to EMF were significantly longer
than the controls at each sampling time (p5 0.001).
The rate of root elongation in EMF-treated roots was
significantly faster (1.21 mm h71) than in the con-
trols (0.95 mm h71). Comparable differences in root
elongation between exposed and control roots were
detected in samples exposed to CL, the average
difference being 28% after 30 h EMF exposure (data
not shown). The exposure to the EMF also affected
the mitotic activity of root meristems. In fact, in roots
exposed to the EMF the mitotic index was 8.5+ 0.5,
significantly higher (0.0015 p5 0.01) than the
control roots (7.0+ 0.3).
Size of the QC following 30 h CL with 3H-TdR
The QC of root apical meristems (RAM) was
visualized by the standard technique of continuous
labelling (CL) with 3H-TdR. In roots exposed to
EMF, the QC is significantly smaller than the
controls whether judged by height or width in
longitudinal median sections (Table I). These data
are supported by micrographs of the QC in both
treatments (Figure 3). We did not observe differ-
ences in cell size between treatments and, therefore,
cell number is also significantly smaller in QCs
sampled from the EMF treatment compared with the
controls. However more labelled cells were observed
in the QCs of roots exposed to EMF.
Cytological analysis in root cap and metaxylem cells
The mean area of root cap cells was measured at 10
and 30 h of treatment in both EMF and control roots
(Table II; Figure 4). After 10 h of exposure to EMF
cell size was not affected, while a significant increase
(21.4 %) in cell area was observed following pro-
longed treatment (30 h exposure), starting from the
root junction to the cap end. Moreover, following
the EMF exposure few and small statocytes were
observed in the columella cells as compared
Figure 2. The rate of root elongation (mm) in maize seedlings
during 30-h exposure to the magnetic field (black squares) and in
control seedlings grown outside of the magnet cavity (white
squares). Values are the mean+SE of three replicates; p5 0.001
at each sampling time, except t¼0). For control roots
y¼7.46þ 0.956x, R2¼0.997; for treated roots y¼6.73þ 1.213x,
R2¼ 0.990.
Table I. The size and cell number of the QC together with the
labelling index (LI) within the quiescent centre in roots of maize
exposed to electromagnetic field (EMF) for 30 h, as compared
with the control.
Test
Height
(mm)
Width
(mm) cell number LI (%)
EMF 111+10* 231+ 12** 303+ 27** 5+ 1.1**
Control 125+11* 342+ 15** 520+ 70** 2+ 0.2**
Values are the mean+SE of measurements on roots (n¼12) from
three different replicates. *p5 0.01; **p5 0.001.
Magnetic field-induced changes to maize root 89
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011
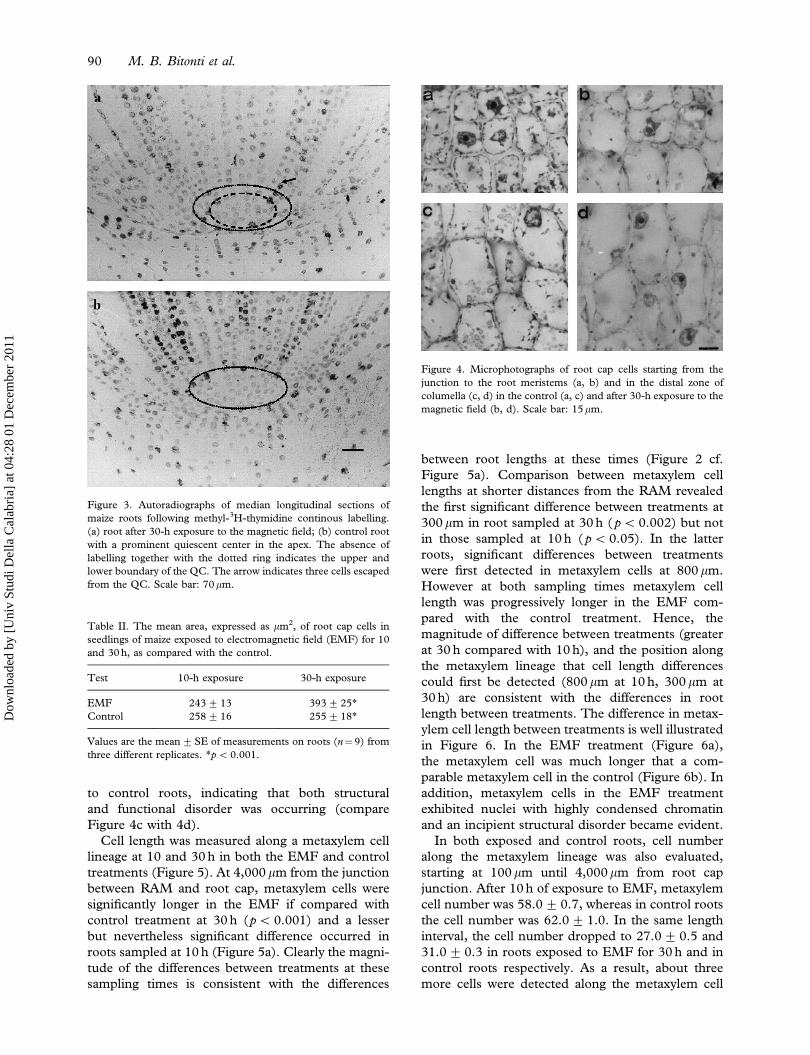
to control roots, indicating that both structural
and functional disorder was occurring (compare
Figure 4c with 4d).
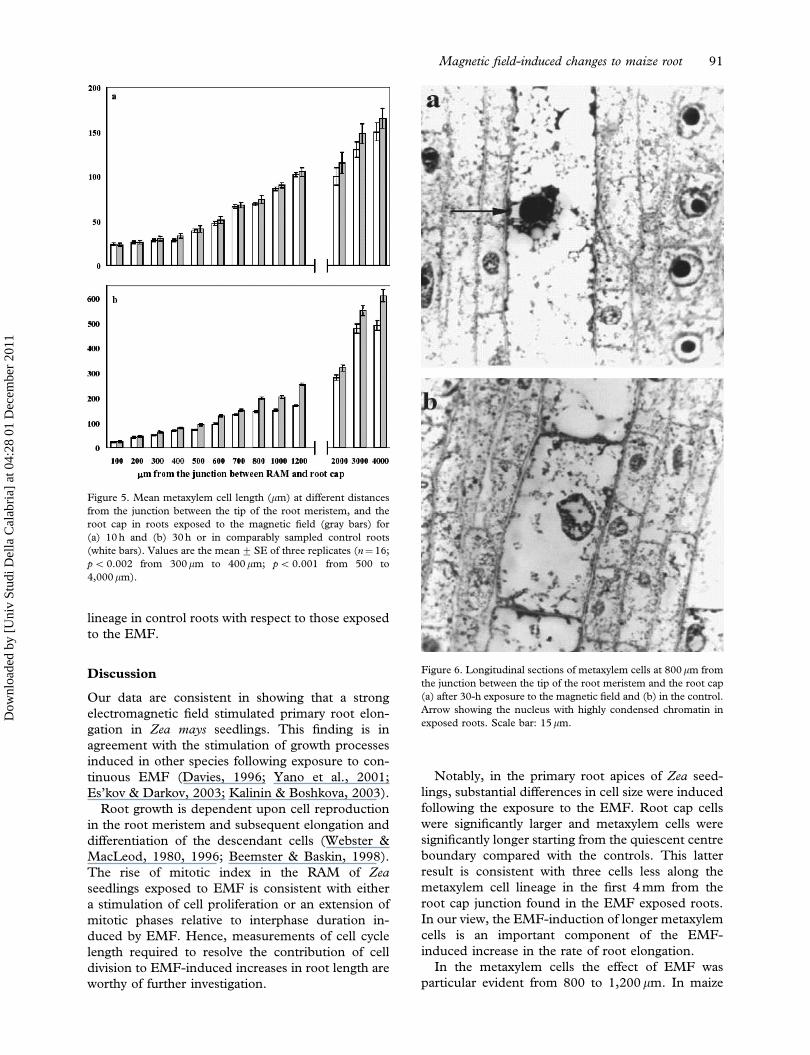
Cell length was measured along a metaxylem cell
lineage at 10 and 30 h in both the EMF and control
treatments (Figure 5). At 4,000 mm from the junction
between RAM and root cap, metaxylem cells were
significantly longer in the EMF if compared with
control treatment at 30 h (p5 0.001) and a lesser
but nevertheless significant difference occurred in
roots sampled at 10 h (Figure 5a). Clearly the magni-
tude of the differences between treatments at these
sampling times is consistent with the differences
between root lengths at these times (Figure 2 cf.
Figure 5a). Comparison between metaxylem cell
lengths at shorter distances from the RAM revealed
the first significant difference between treatments at
300 mm in root sampled at 30 h ( p5 0.002) but not
in those sampled at 10 h ( p5 0.05). In the latter
roots, significant differences between treatments
were first detected in metaxylem cells at 800 mm.
However at both sampling times metaxylem cell
length was progressively longer in the EMF com-
pared with the control treatment. Hence, the
magnitude of difference between treatments (greater
at 30 h compared with 10 h), and the position along
the metaxylem lineage that cell length differences
could first be detected (800 mm at 10 h, 300 mm at
30 h) are consistent with the differences in root
length between treatments. The difference in metax-
ylem cell length between treatments is well illustrated
in Figure 6. In the EMF treatment (Figure 6a),
the metaxylem cell was much longer that a com-
parable metaxylem cell in the control (Figure 6b). In
addition, metaxylem cells in the EMF treatment
exhibited nuclei with highly condensed chromatin
and an incipient structural disorder became evident.
In both exposed and control roots, cell number
along the metaxylem lineage was also evaluated,
starting at 100 mm until 4,000mm from root cap
junction. After 10 h of exposure to EMF, metaxylem
cell number was 58.0+ 0.7, whereas in control roots
the cell number was 62.0+ 1.0. In the same length
interval, the cell number dropped to 27.0+ 0.5 and
31.0+ 0.3 in roots exposed to EMF for 30 h and in
control roots respectively. As a result, about three
more cells were detected along the metaxylem cell
Figure 3. Autoradiographs of median longitudinal sections of
maize roots following methyl-3H-thymidine continous labelling.
(a) root after 30-h exposure to the magnetic field; (b) control root
with a prominent quiescent center in the apex. The absence of
labelling together with the dotted ring indicates the upper and
lower boundary of the QC. The arrow indicates three cells escaped
from the QC. Scale bar: 70mm.
Table II. The mean area, expressed as mm2, of root cap cells in
seedlings of maize exposed to electromagnetic field (EMF) for 10
and 30 h, as compared with the control.
Test 10-h exposure 30-h exposure
EMF 243+ 13 393+25*
Control 258+ 16 255+18*
Values are the mean+SE of measurements on roots (n¼9) from
three different replicates. *p50.001.
Figure 4. Microphotographs of root cap cells starting from the
junction to the root meristems (a, b) and in the distal zone of
columella (c, d) in the control (a, c) and after 30-h exposure to the
magnetic field (b, d). Scale bar: 15mm.
90 M. B. Bitonti et al.
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011
lineage in control roots with respect to those exposed
to the EMF.
Discussion
Our data are consistent in showing that a strong
electromagnetic field stimulated primary root elon-
gation in Zea mays seedlings. This finding is in
agreement with the stimulation of growth processes
induced in other species following exposure to con-
tinuous EMF (Davies, 1996; Yano et al., 2001;
Es’kov & Darkov, 2003; Kalinin & Boshkova, 2003).
Root growth is dependent upon cell reproduction
in the root meristem and subsequent elongation and
differentiation of the descendant cells (Webster &
MacLeod, 1980, 1996; Beemster & Baskin, 1998).
The rise of mitotic index in the RAM of Zea
seedlings exposed to EMF is consistent with either
a stimulation of cell proliferation or an extension of
mitotic phases relative to interphase duration in-
duced by EMF. Hence, measurements of cell cycle
length required to resolve the contribution of cell
division to EMF-induced increases in root length are
worthy of further investigation.
Notably, in the primary root apices of Zea seed-
lings, substantial differences in cell size were induced
following the exposure to the EMF. Root cap cells
were significantly larger and metaxylem cells were
significantly longer starting from the quiescent centre
boundary compared with the controls. This latter
result is consistent with three cells less along the
metaxylem cell lineage in the first 4 mm from the
root cap junction found in the EMF exposed roots.
In our view, the EMF-induction of longer metaxylem
cells is an important component of the EMF-
induced increase in the rate of root elongation.
In the metaxylem cells the effect of EMF was
particular evident from 800 to 1,200 mm. In maize
Figure 5. Mean metaxylem cell length (mm) at different distances
from the junction between the tip of the root meristem, and the
root cap in roots exposed to the magnetic field (gray bars) for
(a) 10 h and (b) 30 h or in comparably sampled control roots
(white bars). Values are the mean+SE of three replicates (n¼16;
p50.002 from 300mm to 400 mm; p5 0.001 from 500 to
4,000mm).
Figure 6. Longitudinal sections of metaxylem cells at 800mm from
the junction between the tip of the root meristem and the root cap
(a) after 30-h exposure to the magnetic field and (b) in the control.
Arrow showing the nucleus with highly condensed chromatin in
exposed roots. Scale bar: 15mm.
Magnetic field-induced changes to maize root 91
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011

roots, this zone corresponds to the ‘distal elongation
zone’ characterized by cells that are strongly involved
in the response of roots to a variety of environmental
stimuli (Erickson & Sax, 1956; Ishikawa & Evans,
1995). In this context, intrinsic electric fields have
been found around the root tip (Behrens et al.,
1982, 1985; Ryan et al., 1990; Weisenseel & Meyer,
1997). These minute electric currents seem to be
related to the polar transport of endogenous auxin
in the root tissue (Biro et al., 1982; Lee et al., 1984;
Behrens et al., 1985) and to the cytosolic changes
of Ca2þ. Since both auxin and Ca2þ regulate the
rate of cell expansion (Behrens et al., 1982, 1985;
Lee et al., 1983; Pickard, 1985), our hypothesis is
that EMF may disrupt the pattern of intrinsic ionic
currents by altering the ion movement across the
plasma membrane (Stange et al., 2002), thus altering
the elongation of exposed roots.
The experiment of CL clearly showed that the size
of QC was substantially reduced in the RAM of roots
exposed to EMF with three layers of cells escaping
from QC control. This reduction in QC size, may
have resulted in precocious expansion and differ-
entiation of contacting cells in both basipetal (root
cap) and acropetal (metaxylem lineage) direction. In
fact, in exposed roots the early differences in length
(at 300 mm from 4,000mm) of metaxylem cells were
accompanied by a precocious structural disorder.
Analogously, symptoms of a precocious structural
and functional disorder were observed in the
sloughing cells of the root cap, which exhibited
small statocytes, likely as a consequence of starch
degradation induced by stress imposed by strong
EMF-exposure. Notably, rapid degradation of
amyloplasts in columella cells has been detected
also in water-stressed roots showing a reduced
responsiveness to gravity (Takahashi et al., 2003).
Thus, features described for both metaxylem and
root cap cells may be consistent with an adv-
anced differentiation of cells that are programmed
to die.
Given that QC cells repress differentiation in
contacting apical initial cells (Van der Berg et al.,
1997; Sabatini et al., 1999) we suggest that the onset
of cell differentiation was permitted to occur closer
to the root tip in the EMF, because of cells escaping
from QC control. In conclusion, we have demon-
strated that EMF results in an increase in primary
root elongation, and the onset of metaxylem cell
elongation occurs at a closer distance to the root tip
than in control. The quiescent centre in the RAM
was significantly smaller in the MF compared with
the control treatments. The precocious elongation of
the metaxylem cells and the reduction in QC size
may be casually linked but verification will depend
on experiments that focus on the onset of differ-
entiation in response to MF.
Acknowledgment
This work was funded by the University of Calabria
(ex 60% grant).
References
Audus LJ. 1960. Magnetotropism: a new plant growth response.
Nature 185:132 – 134.
Barlow PW, Pilet PE. 1983. Mitotic activity in the maize root apex
after freeze-decapping. Planta 157:286 – 288.
Barlow PW, Rathfelder EL. 1985. Cell division and regeneration
in primary root meristems of Zea mays recovering from cold
treatment. Environ Experiment Bot 27:303 – 314.
Beemster GTS, Baskin TI. 1998. Analysis of cell division and
elongation underlying the developmental acceleration of root
growth in Arabidopsis thaliana. Plant Physiol 116:515 – 526.
Beherens HM, Wiesenseel MH, Sievers A. 1982. Rapid changes
in the pattern of electric current around the root tip of
Lepidum sativum L. following gravistimulation. Plant Physiol
70:1079 – 1083.
Behrens HM, Gradmann D, Sievers A. 1985. Membrane-
potential responses following gravistimulation in root of
Lepidum sativum L. Planta 163:463 – 472.
Biro RL, Hale CC, Wiegend OF, Roux SJ. 1982. Effects of
chloropromazine on gravitropism in avena coleoptiles. Ann Bot
50:737 – 745.
Blank M, Goodman R. 2002. Insights into electromagnetic
interaction mechanisms. J Cell Physiol 192:16 – 22.
Brulfert A, Miller MW, Robertson D, Dooley DA, Economou P.
1985. A cytohistological analysis of roots whose growth is affected
by a 60-Hz electric field. Bioelectromagnetics 6:283 – 291.
Clowes FAL. 1972. Cell cycle in a complex meristem after
X-irradiation. New Phytol 71:891 – 897.
Crook MJ, Ennos AR. 1996. Mechanical differences between free-
standing and supported wheat plants Triticum aestivum L. Ann
Bot 50:109 – 119.
Davies MS. 1996. Effects of 60 Hz electromagnetic fields on early
growth in three plant species and a replication of previous
results. Bioelectromagnetics 17:154 – 161.
Erickson RO, Sax KB. 1956. Rates of cell division and cell
elongation in the growth of the primary root of Zea mays. Proc
Am Phil Soc 100:499 – 514.
Es’kov EK, Darkov AV. 2003. Effect of high intensity magnetic
field on the processes of early growth in plant seeds and
development of honeybees. Izv Akad Nauk Ser Biol 2003:
617 – 622.
Feldman LJ, Torrey JG. 1975. The quiescent centre and primary
vascular tissue pattern formation in cultured roots of Zea mays.
Can J Bot 53:2796 – 2803.
Goodman AM, Ennos AR. 1996. A comparative study of the
response of the roots and shoots of sunflower and maize to
mechanical stimulation. J Experiment Bot 47:1499 – 1507.
Hood E. 2004. EMFs and DNA effects: potential mechanism
elucidated. Environ Health Perspect 112:368.
Inoue M, Miller MW, Cox C, Carstesen EL. 1985. Growth rate
and mitotic index analysis of Vicia faba L. roots to 60-Hz
electric field. Bioelectromagnetics 6:293 – 303.
Ishikawa H, Evans ML. 1995. Specialized zones of development
roots. Plant Physiol 109:725 – 727.
Jenrow KA, Smith CH, Liboff AR. 1995. Weak extremely low-
frequency magnetic field and regeneration in the planaria
Dugesia tigrina. Bioelectromagnetics 16:106 – 112.
Kalinin LG, Boshkova IL. 2003. Physical model of the plant tissue
response to exposure to the microwave electromagnetic field.
Biofizika 48:122 – 124.
Kuznetsov OA, Hasenstein KI. 1996. Magnetophoretic induction
of root curvature. Planta 198:87 – 94.
92 M. B. Bitonti et al.
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011

Kuznetsov OA, Schwuchow J, Sack FD, Hasenstein KI. 1999.
Curvature induced by amyloplast magnetophoresis in protone-
mata of the moss Ceratodon purpureus. Plant Physiol 19:645 – 650.
Lee JS, Mulkey TJ, Evans ML. 1983. Reversible loss of gravitropic
sensitivity in maize root after tip application of calcium
chelators. Science 220:1375 – 1376.
Lee JS, Mulkey TJ, Evans ML. 1984. Inhibition of polar calcium
movement and gravitropism in roots treated with auxin-
transport inhibitor. Planta 160:536 – 543.
Muller ML, Barlow PW, Pilet PE. 1994. Effect of abscisic acid on
the cell cycle in the growing maize root. Planta 195:10 – 16.
Okamoto H, Tatara A. 1995. Effects of low-dose g-radiation on
the cell cycle duration of barley roots. Environ Experiment Bot
35:379 – 388.
Piatti E, Albertini MC, Baffone W, Fraternale D, Citterio B,
Piacentini MP, et al. 2002. Antibacterial effect of a magnetic
field on Serratia marcescens and related virulence to Hordeum
vulgare and Rubus fruticosus callus cells. Comp Biochem Physiol
B Biochem Mol Biol 132:359 – 365.
Pickard, BG. 1985. Roles of hormones, protons and calcium
in geotropism. In: Pharis RP, Reid DM, editors. Encyclopedia
of Plant Physiology: Hormonal regulation of development III
role of environment factors. Berlin: Springer-Verlag.
pp 193 – 281.
Rost TL, Van’t Hof J. 1973. Radiosensitivity, RNA and protein
metabolism of ‘leaky’ and arrested cells in sunflowers root
meristems. Am J Bot 60:172 – 181.
Ryan PR, Newman IA, Shields B. 1990. Ion fluxes in corn roots
measured by microelectrodes with ionic-specific liquid mem-
branes. J Membrane Sci 53:59 – 69.
Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoy LE,
Malamy J, et al. 1999. An auxin-dependent distal organizer
of pattern and polarity in the Arabidopsis root. Cell 99:
463 – 472.
Stange BC, Rowland RE, Rapley BI, Pod JV. 2002. ELF magnetic
fields increase amino acid uptake into Vicia faba roots and alter
ion movement across the plasma membrane. Bioelectromag-
netics 23:347 – 354.
Takahashi N, Yamazaki Y, Kobayashi A, Higashitani, Takahashi
H. 2003. Hydrotropism interacts with gravitropism by degrad-
ing amyloplasts in seedling roots of Arabidopsis and radish.
Plant Physiol 132:805 – 881.
Van der Berg C, Willemsen VV, Hage W, Weisbeek P, Scheres B.
1997. Short-range control of cell differentiation in the
Arabidopsis root meristem. Nature 390:287 – 289.
Webster PL, Langenauer H. 1973. Experimental control of the
activity of the quiescent center in excised root tips of Zea mays.
Planta 112:91 – 100.
Webster PL, MacLeod RD. 1980. Characteristics of root apical
meristem cell population kinetics: A review of analyses and
concepts. Environ Experiment Bot 20:335 – 358.
Webster PL, MacLeod RD. 1996. The root apical meristems and
its margins. In: Waisel Y, Eshel A, Kafkafi U, editors. Plant
roots. The hidden half. New York: Marcel Dekker, Inc.
pp 51 – 76.
Weisenseel MH, Meyer AJ. 1997. Bioelectricity, gravity and
plants. Planta 203:98 – 106.
Yano A, Hidaka E, Fujiwara K, Imoto M. 2001. Induction of
primary root curvature in radish seedlings in a static magnetic
field. Bioelectromagnetics 22:194 – 199.
Magnetic field-induced changes to maize root 93
Dow
nloa
ded
by [
Uni
v St
udi D
ella
Cal
abri
a] a
t 04:
28 0
1 D
ecem
ber
2011




![Tolerance of Maize (Zea mays L.) and Soybean [Glycine max (L.) Merr.] to Late Applications of Postemergence Herbicides](https://static.fdokumen.com/doc/165x107/63438e5b1a2cfc44fc019bae/tolerance-of-maize-zea-mays-l-and-soybean-glycine-max-l-merr-to-late-applications.jpg)















