
Elevation of cytoskeletal protein breakdown in aged Wistar rat brain
Upload
independentCategory
view
0download
0
Macrophages from lupus-prone MRL mice are characterizedby abnormalities in Rho activity, cytoskeletal organization,and adhesiveness to extracellular matrix proteins
Angelika Longacre, Jason S. Koh, Kevin K-H. Hsiao, Hannah Gilligan, Hanli Fan, Vimal A. Patel,and Jerrold S. Levine1
Section of Nephrology, Department of Medicine, The University of Illinois at Chicago
Abstract: Macrophages (m�) from prediseasedmice of the major murine models of lupus have anidentical defect in cytokine expression that is trig-gered by serum and/or apoptotic cells. It is strikingthat cytokine expression in the absence of serumand apoptotic cells is equivalent to that of nonau-toimmune mice. Here, we show that m� from pre-diseased lupus-prone MRL/MpJ (MRL/�) or MRL/MpJ-Tnfrsf6lpr (MRL/lpr) mice also have reversibleabnormalities in morphology, cytoskeletal organi-zation, and adhesive properties. In the presence ofserum, MRL m� adhered in increased numbers toa variety of extracellular matrix proteins com-pared with m� from two nonautoimmune strains.However, in the absence of serum, adhesion byMRL m� was similar to that of nonautoimmunem�. Increased adhesion by MRL m� was also ob-served in the presence of apoptotic but not ne-crotic cells. The morphology and actin-stainingpattern of adherent MRL m� were consistent withreduced activity of Rho, a cytoskeletal regulator.Indeed, MRL m� cultured in the presence of se-rum had markedly decreased levels of active Rhocompared with nonautoimmune m�. It is remark-able that when cultured in the absence of serum,MRL m� displayed normal Rho activity and cy-toskeletal morphology. Addition of a Rho inhibitorto normal m� reproduced the morphologic andcytoskeletal abnormalities observed in MRL m�.Taken together, our findings support the hypothe-sis that m� from MRL and other systemic lupuserythematosus-prone mice have an apoptotic, cell-dependent, autoimmune phenotype that affects abroad range of m� functions, including cytokinegene expression and Rho-dependent cytoskeletalregulation. J. Leukoc. Biol. 76: 000–000; 2004.
Key Words: rodent � autoimmunity
INTRODUCTION
Adhesive interactions play a critical role in the overall regu-lation of the immune system. Molecules, such as integrins andselectins on the surface of lymphocytes, macrophages (m�),
dendritic cells, and other leukocytes, mediate the interactionsof these cells with one another, extracellular matrix (ECM)proteins, or endothelial and other cells [1–4]. For example, therecruitment of blood monocytes to the peritoneal cavity follow-ing thioglycolate injection is dependent on expression of theintegrin CD11b/CD18 [5, 6]. Differentiation-dependent expres-sion of various selectins and their ligands by T lymphocytesleads to the differential trafficking of naive and effector Tlymphocytes to lymph nodes and inflamed peripheral tissue,respectively [1, 3, 4, 7]. The binding of integrins and otheradhesion molecules such as CD44 to ECM proteins leads to theretention of antigen-stimulated T cells in lymph nodes andperipheral sites of infection or inflammation [1, 3, 4, 7]. Ad-ditionally, adhesive interactions play a critical role in stabili-zation of the immunologic synapse between antigen-presentingcells (APC) and T lymphocytes, thereby modulating intracel-lular signaling events and the overall activation of antigen-specific T cells [8, 9].
Given this broad role for adhesive interactions in immunehomeostasis, it is not surprising that alterations in the functionor expression of various adhesion molecules can affect thestrength and/or target specificity of the immune response inhealth and disease [4, 10, 11]. For example, administration ofa monoclonal antibody (mAb) that blocks adhesion through theintegrin CD11b/CD18, which is critical for the recruitment ofm� [5], significantly decreased the development of overt au-toimmune diabetes from 98% to 3% in a transfer model ofdiabetes using nonobese diabetic (NOD) mice [12]. Similarly,targeted deletion of the integrin intracellular adhesion mole-cule-1 significantly prolonged the lifespan of lupus-proneMRL/MpJ-Tnfrsf6lpr (MRL/lpr) mice and reduced the severityof glomerular disease [13], whereas targeted deletion of selec-tins exacerbated autoimmunity [14].
We have shown that m� from prediseased mice of all majormurine models of systemic lupus erythematosus (SLE) have anidentical defect in cytokine expression, which is triggered byserum [fetal bovine serum (FBS)] lipids and/or apoptotic cells[15–18]. Affected strains include MRL/MpJ (MRL/�), MRL/
1 Correspondence: The University of Illinois at Chicago, Section of Nephrol-ogy, 820 South Wood Street, MC-793, Room 479/CSN, Chicago, IL 60612.E-mail: [email protected]
Received June 17, 2004; revised July 12, 2004; accepted July 19, 2004; doi:10.1189/jlb.0604346
Journal of Leukocyte Biology Volume 76, November 2004 1
Uncorrected Version. Published on August 17, 2004 as DOI:10.1189/jlb.0604346
Copyright 2004 by The Society for Leukocyte Biology.

lpr, NZB, NZW, NZB/W F1, BXSB, and LG/J, all of whichexpress or contribute to the development of autoimmunity. Nosimilar defect was found in 16 nonautoimmune control strains[18, 19]. This defect may be characteristic of autoimmunity ingeneral, as m� from prediseased, diabetes-prone NOD micealso respond abnormally to apoptotic cells, mimicking closelythe apoptotic, cell-dependent abnormality that we have ob-served in SLE-prone strains [19]. Elicitation of this defect byexposing m� from prediseased, autoimmune-prone mice toapoptotic cells leads to the dysregulated expression of multiplecytokines [18, 19]. In probing the basis and consequences ofthis defect, we have used FBS as a surrogate for apoptotic cells,as lipid-containing FBS fully mimics the effect of apoptoticcells in eliciting all aspects of this defect [18, 19].
In the course of these studies, we noticed that m� fromSLE-prone mice also displayed abnormalities in their morphol-ogy and adhesive properties, especially when cultured on bac-teriologic plastic (BP) or surfaces coated with ECM proteinssuch as laminin (LAM), fibronectin (FN), or collagen. In thepresence of FBS, m� from MRL/� and MRL/lpr mice adheredin increased numbers and with a distinct spread morphology ascompared with m� from two nonautoimmune strains. Whencultured in the absence of FBS, the morphology and adhesiveinteractions of MRL m� became normal. Immunofluorescent(IF) staining of adherent MRL m� cultured in the presence butnot the absence of FBS revealed an actin-staining patternconsistent with reduced activity of Rho, a cytoplasmic G pro-tein and cytoskeletal regulator [20, 21]. In confirmation ofthese observations, when MRL m� were cultured in the pres-ence of FBS, the percentage of active Rho was significantlydecreased compared with nonautoimmune m�. In sharp con-trast, culturing MRL m� in the absence of FBS restored theamount of active Rho to normal levels. Addition of a Rhoinhibitor to normal m� reproduced the morphologic and cy-toskeletal abnormalities seen in MRL m�. Taken together,these results support the hypothesis that m� from MRL andother SLE-prone mice have a serum- and/or apoptotic, cell-dependent, autoimmune phenotype that affects a broad rangeof m� functions, including cytokine gene expression and Rho-dependent, cytoskeletal regulation.
MATERIALS AND METHODS
MiceBALB/cByJ (BALB/c), C57BL/6J (C57BL/6), MRL/�, and MRL/lpr mice werepurchased from The Jackson Laboratory (Bar Harbor, ME). Unless otherwiseindicated, these mice were used between 4 and 6 weeks of age. Diseased micewere retired female breeders, greater than 8 months of age (The JacksonLaboratory). All animal protocols were approved by the Institutional AnimalCare and Use Committee.
Materials and reagentsLipopolysaccharide (LPS; Escherichia coli-derived, serotype 0111:B4) wasobtained from List Biological (Campbell, CA). Bovine serum albumin (BSA)and normal goat serum were obtained from Jackson ImmunoResearch Labo-ratories (West Grove, PA). Clostridium botulinum C3 exoenzyme (C3) wasobtained from Cytoskeleton (Denver, CO).
AbThe following mAb were used for flow cytometric analysis: 5C6, rat immuno-globulin G (IgG)2b � anti-mouse CD11b (BioSource International, Camarillo,
CA); MI/70.15.1, rat IgG2b � anti-mouse CD11b (BioSource International);LO-DNP-11, rat IgG2b � control (BioSource International); fluorescein iso-thiocyanate (FITC)-labeled goat F(ab�)2 anti-rat Ig; phycoerythrin (PE)-labeledrat IgG2a � anti-mouse CD18 (PharMingen, San Diego, CA); PE-labeled ratIgG2a � anti-mouse CD49d (PharMingen); and PE-labeled rat IgG2a � anti-mouse CD49e (PharMingen). The following Ab and reagents were used for IF:1624, mouse IgG1 � anti-human vinculin mAb (Chemicon International,Temecula, CA); rhodamine-conjugated sheep polyclonal anti-mouse polyclonalIgG (Cappel Research Reagents, Durham, NC); Oregon green-conjugatedphalloidin (Molecular Probes, Eugene, OR); biotin-conjugated rat IgG2b �anti-mouse CD11b (Caltag Laboratories, Burlingame, CA); and Cy5-conju-gated streptavidin (Caltag Laboratories).
m� culture
Peritoneal exudate cells (PEC) were harvested by lavage 3 days after intra-peritoneal injection of 1.5 ml 4.05% thioglycolate broth [15–19, 22]. Afterbeing washed twice in RPMI-1640, PEC, destined for immediate use in anadhesion assay (see Fig. 1), were resuspended in R.10 culture medium (RPMI1640 plus 10% FBS with 2 mM L-glutamine, 5 mM HEPES, 100 U/mlpenicillin, and 100 �g/ml streptomycin). All other PEC were cultured over-night under nonadherent conditions for use the following day. During over-night, nonadherent culture, PEC were suspended in one of three media: R.10;delipidated R.10 [R.10 with 10% delipidated FBS (dFBS) instead of FBS]; orFBS-free R.0 medium (R.10 minus FBS) supplemented with 100 U/ml humanrecombinant macrophage-colony stimulating factor (rM-CSF; generous gift ofGenetics Institute, Cambridge, MA). For all experiments comparing controlversus SLE-prone MRL strains, we used pooled m� harvested from a minimumof at least five mice per strain.
Preparation of thymocytes
Thymocytes were harvested from the thymuses of BALB/c or MRL/lpr mice andsuspended in R.10 at 5 � 107 cells per ml [18, 19, 22]. Apoptosis was inducedby addition of 5 � 10�6 M hydrocortisone, followed by incubation at 37°C for8 h. Before addition to m� cultures, apoptotic thymocytes were washed threetimes in RPMI 1640 and resuspended in R.0. Necrosis was induced by heatingthymocytes to 70°C for 40 min.
Documentation of thymocyte apoptosis
Viable cells were defined as propidium iodide (PI)-negative cells with faintnuclear Hoechst staining. Apoptotic cells were defined as PI-negative cellswith bright nuclear Hoechst staining and decreased cell size. Post-apoptoticcells (i.e., apoptotic cells that had lost cell-membrane integrity) were definedas PI-positive cells with bright Hoechst staining and decreased cell size. Bythese criteria, �60% of cells in apoptotic cell preparations were apoptotic,�15% were viable, and �25% were post-apoptotic [18, 19, 22]. Necroticcells, as defined by increased cell size in association with uptake of PI andfaint Hoechst staining, comprised 0.1% of apoptotic cell preparations, ascompared with 95% of necrotic cell preparations [18, 19, 22].
Overnight, nonadherent culture
After isolation and washing, PEC were suspended in the appropriate medium(R.10, delipidated R.10, R.0 plus human rM-CSF) and added to 60 � 15 mmtissue-culture dishes at 5–10 � 106 cells per dish. Each dish had beenprecoated the day before with 4.0 ml 1.5% agarose (Gibco, Grand Island, NY)in phosphate-buffered saline (PBS) and then equilibrated over 24 h againstthree changes of the same medium in which PEC were to be resuspended. Afterovernight culture on agarose, PEC were collected, washed in RPMI 1640,resuspended in the same medium as they had been cultured overnight, andthen used for experiments.
FBS delipidation
FBS was delipidated by one of two protocols. FBS was mixed 1:1 (v/v) for 2 hat 22°C with butanol:diethyl ether (1:4, v/v) [23] or chloroform:methanol (2:1,v/v) [24] and then centrifuged to separate organic and aqueous phases. Re-sidual organic solvents were removed from the delipidated aqueous phase byapplying a water-aspirator vacuum for 2 h. The resultant dFBS was sterilelyfiltered for later use.
2 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org

Adhesion assay
PEC suspended in R.10, delipidated R.10, or R.0 plus human rM-CSF (100U/ml) were added at 1–2 � 105 cells per well to 96-well plates in the presenceor absence of LPS (100 ng/ml). Wells were untreated (BP), tissue culture-treated, or coated with FN, LAM, type I collagen (COL I), COL IV, orpoly-D-lysine (P-D-LYS). After 2 or 24 h, wells were washed 3� with RPMI1640, and the number of remaining adherent cells was quantitated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. For allstrains of mice, adherent cells were �98% m�, as confirmed by morphologicanalysis and nonspecific esterase staining. In virtually all cases (except whereadhesion was deliberately inhibited), the percentage of added m� adhering toindividual wells was always �50% for that strain manifesting the greatestdegree of adhesion.
MTT assay
The number of adherent, viable m� was determined using a modification of theMTT assay [25], whose output is proportional to the total number of viable cellspresent. After washing away nonadherent cells, 165 �l MTT dissolved in R.0(1 mg/ml) was added to each well. After incubation at 37°C for 4 h, the MTTformazan was dissolved by adding 165 �l 10% sodium dodecyl sulfate in 0.01N HCl. Aliquots from each well were read using a microELISA plate readerwith a test wavelength of 570 nm and a reference wavelength of 650 nm. Dataare presented as mean � SE of the raw optical density (OD) values, which asseen in Figures 1–3, may be compared, as may those seen in Figures 4 and 5,but because of variations in experimental conditions, OD values betweenfigures from these two groups should not be compared.
IF staining
After overnight, nonadherent culture, 100,00 PEC were allowed to adhere to35 � 15 mm BP dishes or 12 mm round coverslips coated with mouseLAM/P-D-LYS (BD Biosciences, San Diego, CA) in 100 �l medium, R.10, orR.0 plus human rM-CSF (100 U/ml). In some studies, cells were treated with2 �g C3 during adhesion to LAM-coated coverslips. After 6–24 h, nonadherentcells were washed away with PBS, and the remaining adherent m� were fixedwith 4% paraformaldehye in PBS for 10 min at room temperature and thenwashed 3� with PBS. All subsequent steps took place at room temperature.m� were permeabilized with 0.1% Triton X-100 for 10 min, washed 3� withPBS, and then incubated in blocking buffer (PBS containing 5% normal goatserum, 1% BSA, and 0.1% Triton X-100) for 1 h. m� were incubated inblocking buffer containing primary mAb (antivinculin and anti-CD11b) andOregon green-conjugated phalloidin for 1 h, washed 3� with PBS, incubatedin blocking buffer containing Cy5-conjugated streptavidin or rhodamine-con-jugated sheep polyclonal anti-mouse IgG for 1 h, and washed 3� with PBS. Form� adhering to BP, a coverslip was added, whereas for m� adhering toLAM-coated coverslips, the coverslip was inverted onto a glass slide. Cover-slips were then sealed at their edges with clear nail polish and stored overnightat 4°C for viewing the following day.
Microscopy
Cells were viewed under a Zeiss Axiovert microscope equipped with a Achro-stigmat 32�/0.40 Ph1 objective lens and a Micromax digital camera (Univer-sity of Chicago Integrated Microscopy Core, Cancer Center Digital LightFacility, IL). Images from double- or triple-staining were obtained at the sameplane of focus. Image-capture analysis was performed using SlideBook 3.04(Intelligent Imaging Innovations, Denver, CO), run on a Power Macintosh.Images were saved in a tagged image file format and printed on an Epson 1280printer.
Flow cytometry
PEC were stained by one of two protocols, depending on the mAb. PEC stainedwith 5C6 (anti-CD11b) or MI/70 (anti-CD11b) were collected after overnight,nonadherent culture and suspended in R.10 for 30 min at 4°C to block Fcreceptors for IgG. PEC were next washed 3� in RPMI 1640, incubated for 1 hat 4°C in flow cytometry buffer (R.0 containing 0.2% delipidated BSA)containing the relevant mAb, washed 3� in flow cytometry buffer, incubatedfor 1 h at 4°C in flow cytometry buffer containing FITC-labeled secondarymAb, washed 3� in flow cytometry buffer, and finally resuspended in flow
cytometry buffer at 106 cells/ml. PEC stained with C71/16 (anti-CD18), R1-2(anti-CD49d), or 5H10-27 (anti-CD49e) were suspended for 5 min at 4°C inflow cytometry buffer containing Fc Block� (rat anti-mouse CD16/CD32 mAb)at 1 �g per 106 cells. Fc Block� was not washed away. PE-labeled mAb wereadded directly to cell suspensions. PEC were then incubated for 1 h at 4°C,washed 3� in flow cytometry buffer, and finally resuspended in flow cytometrybuffer at 106 cells/ml. Flow cytometric analysis of stained PEC was performedon an Epics ESP flow cytometer (Coulter Electronics, Hialeah, FL). Fluores-cence was achieved with 15 mW 488 nm argon ion laser. Emitted fluorescencewas detected using 530 and 575 nm bandpass optical filters for FITC and PEfluorescence, respectively. Data were analyzed by Epic Elite software (CoulterElectronics).
Rho assay
After overnight, nonadherent culture in R.10 or R.0 plus human rM-CSF, 8 �106 PEC were assayed for active Rho • guanosine 5�-triphosphate (GTP).Active Rho • GTP constitutes only �10% of total Rho in cells. As active Rho• GTP is rapidly hydrolyzed to inactive Rho • guanosine 5�-diphosphate(GDP), a pull-down assay was used to separate active Rho • GTP. We used akit from cytoskeleton containing a fusion protein between glutathione S-transferase and the Rho-binding domain of the rhotekin protein coupled toglutathione agarose beads. Kit directions were followed, and all samples weremaintained on ice throughout. Individual lysates, subjected to the pull-downassay (active Rho•GTP), were then separated by polyacrylamide gel electro-phoresis on a 4–20% tris-glycine gradient gel (Invitrogen, Carlsbad, CA) andtransferred to a nitrocellulose membrane at 100 V for 1 h. After blocking themembrane overnight at room temperature in 5% nonfat milk in 100 mMTris-HCl, pH 7.5, 150 mM NaCl, 0.05% Tween 20 (TBST buffer), the mem-brane was incubated for 1.5 h at room temperature in a 1:500 dilution in TBSTof mouse anti-RhoA IgG1 mAb, supplied with the kit. Blots were washed threetimes with TBST and then incubated with a 1:5000 dilution in TBST of analkaline phosphatase-conjugated goat anti-mouse Ig, supplied as part of adetection kit and used according to the manufacturer’s instructions (chemilu-minescent immunoblot detection system, Applied Biosystems, Bedford, MA).
Statistics
Ten replicate wells per experiment were examined in all adhesion assays, andthe results were averaged. A minimum of three experiments was performed forall data points. Data are expressed as mean � SEM of the averaged valuesobtained from each experiment. Statistical significance was determined by atwo-tailed Student’s t-test.
RESULTS
m� from SLE-prone MRL/� and MRL/lpr miceadhere in increased numbers to a variety ofsurfaces and ECM proteins
Preliminary observations suggested that an increased propor-tion of m� from SLE-prone MRL/� and MRL/lpr mice adheredto a variety of surfaces when m� were plated in the presenceof FBS. To quantify this effect, we used a MTT-based adhesionassay, in which the relative number of m� that remainedadherent after vigorous washing was assessed by mitochondrialconversion of pale yellow MTT to dark blue crystals of MTTformazan. Linearity of the assay was established for cell num-bers ranging from 2000 to 200,000 cells per well by addingknown numbers of PEC followed immediately by MTT withoutintervening washing steps (not shown). Moreover, graphs ofMTT conversion (OD650) versus cell number were roughlysuperimposable with equivalent slopes, irrespective of strain,in the presence and absence of FBS, thereby ensuring thatmitochondrial activity per cell did not vary as a function of
Longacre et al. Abnormalities in Rho and adhesion by MRL macrophages 3

genetic susceptibility to SLE or experimental conditions (notshown).
As shown in Figure 1A, in the presence of FBS, an in-creased proportion of m� from SLE-prone MRL/� and MRL/lpr versus nonautoimmune-prone, control BALB/c andC57BL/6 mice adhered to a variety of surfaces. These surfacesincluded TCP, non-TCP, or BP and plastic coated with COL I,FN, or LAM. The relative increase in the number of adherentMRL m�, compared with BALB/c and C57BL/6 m�, rangedfrom 1.5-fold (TCP, COL I, FN) up to threefold (BP). Adhesionto LAM was marginal for all strains, as it has been shownpreviously that thioglycollate-elicited peritoneal m� requireactivation with phorbol 12-myristate 13-acetate or LPS beforethey will adhere to LAM [24]. Although the results depicted inFigure 1A were obtained after 24 h of adhesion, an identicalpattern of increased adhesion by MRL/� and MRL/lpr m� was
observed after 1 and 4 h of adhesion (not shown). Assay resultswere independent of time, as elicited peritoneal m� are ter-minally differentiated and nonproliferative [22]. No consistent,statistically significant differences in relative adhesion wereseen for MRL/� versus MRL/lpr m� or for BALB/c versusC57BL/6 m�.
It should be stressed that we used only young, prediseasedMRL mice, less than 6 weeks of age, as a source of m�. We didthis to minimize any effects of disease or inflammation, therebyenabling us to distinguish those abnormalities that play afundamental role in the etiology of SLE from those that are aconsequence of established disease. Further evidence againstan effect of disease derives from the absence of a differencebetween MRL/� and MRL/lpr m�, as the presence of the lprmutation accelerates and exacerbates autoimmunity. Moreover,when we compared m� from prediseased (6 weeks) versusovertly diseased (6 months) MRL/lpr mice, the overall pat-tern and magnitude of increased adhesion did not differ (notshown). Nonetheless, the possibility still formally existed thatsubclinical disease or inflammation may have contributed tothe observed differences. We therefore repeated these studiesin the presence of LPS (100 ng/ml), an extremely potentactivator of m�, as a means of achieving a similar degree ofactivation across all m�. As shown in Figure 1B, the presenceof LPS had essentially no effect on the pattern of enhancedadhesion seen with MRL m�. The sole exception was forLAM-coated surfaces, to which activated m� were now able toadhere, as has been previously reported [26]. MRL m� dis-played more than threefold greater adhesion to LAM. As in thecase of unactivated m�, the results in Figure 1B reflect 24 h ofadhesion. An identical pattern of increased adhesion byMRL/� and MRL/lpr m� was also observed after 1 and 4 h ofadhesion (not shown). Taken together, these results suggestthat increased adhesion by MRL m� is independent of diseaseand may therefore represent an inherent abnormality in thesemice.
As opposed to TCP, BP, COL I, FN, and LAM, adhesion toplastic coated with COL IV or P-D-LYS did not differ betweenMRL and control m�, for unactivated and for LPS-activatedm� (Fig. 1). As m� adhere to P-D-LYS nonspecifically, in areceptor-independent manner, the absence of an abnormalityon P-D-LYS-coated surfaces suggests that increased adhesionby MRL m� is likely to be receptor-dependent. Also, asadhesion to TCP, BP, FN, COL I, LAM, and COL IV occurs viaa wide range of different receptors, integrin and nonintegrin innature, our results further suggest that multiple, but not all,adhesion receptors contribute to this abnormality.
Increased adhesion by MRL m� is corrected byremoval or delipidation of FBS
Aberrant cytokine expression by MRL m� is dependent on thepresence of FBS lipids and is fully corrected by culturing MRLm� under FBS-free conditions or in the presence of dFBS [18,19]. We therefore determined whether increased adhesion byMRL m� displayed a similar dependence on FBS. To minimizethe effects of prior in vivo exposure to FBS, m� were culturedovernight under nonadherent conditions in the presence of FBS(R.10), in the presence of dFBS (delipidated R10), or in theabsence of FBS (R.0 plus rM-CSF). After overnight culture,
Fig. 1. MRL m� adhere in increased numbers to a variety of surfaces.Freshly isolated, peritoneal m� from the indicated strains were plated ontovarious surfaces in the presence of 10% FBS (A) without or (B) with LPS (100ng/ml). After 16 h, nonadherent m� were washed away, and the relativenumber of remaining adherent m� was quantitated by MTT assay. All fourpair-wise comparisons between control (BALB/c or C57BL/6) and MRL(MRL/� or MRL/lpr) m� were statistically significant (P0.01) for tissue-culture plastic (TCP), BP, COL I, and FN, irrespective of the presence of LPS.Differences for LAM were statistically significant only in the presence of LPS(P0.01). There were no statistically significant differences in any of the fourpair-wise comparisons between control (BALB/c or C57BL/6) and MRL(MRL/� or MRL/lpr) m� for COL IV and P-D-LYS, irrespective of thepresence of LPS. Data are combined from n � 4 independent experiments.
4 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org
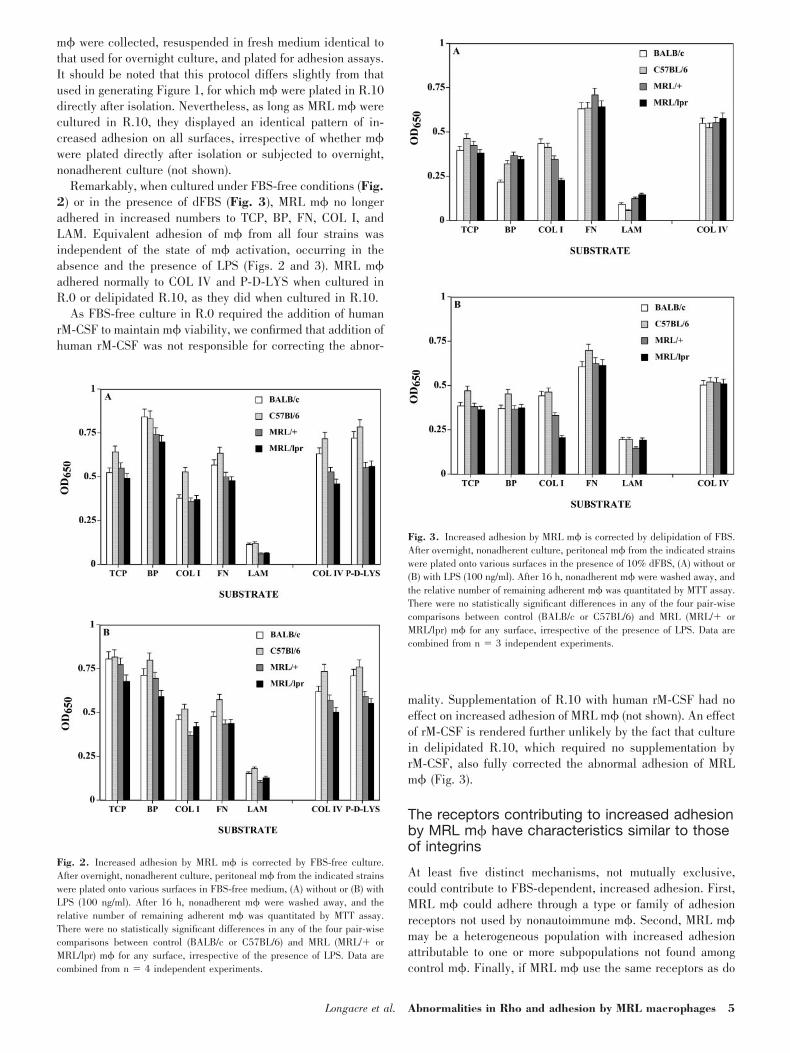
m� were collected, resuspended in fresh medium identical tothat used for overnight culture, and plated for adhesion assays.It should be noted that this protocol differs slightly from thatused in generating Figure 1, for which m� were plated in R.10directly after isolation. Nevertheless, as long as MRL m� werecultured in R.10, they displayed an identical pattern of in-creased adhesion on all surfaces, irrespective of whether m�were plated directly after isolation or subjected to overnight,nonadherent culture (not shown).
Remarkably, when cultured under FBS-free conditions (Fig.2) or in the presence of dFBS (Fig. 3), MRL m� no longeradhered in increased numbers to TCP, BP, FN, COL I, andLAM. Equivalent adhesion of m� from all four strains wasindependent of the state of m� activation, occurring in theabsence and the presence of LPS (Figs. 2 and 3). MRL m�adhered normally to COL IV and P-D-LYS when cultured inR.0 or delipidated R.10, as they did when cultured in R.10.
As FBS-free culture in R.0 required the addition of humanrM-CSF to maintain m� viability, we confirmed that addition ofhuman rM-CSF was not responsible for correcting the abnor-
mality. Supplementation of R.10 with human rM-CSF had noeffect on increased adhesion of MRL m� (not shown). An effectof rM-CSF is rendered further unlikely by the fact that culturein delipidated R.10, which required no supplementation byrM-CSF, also fully corrected the abnormal adhesion of MRLm� (Fig. 3).
The receptors contributing to increased adhesionby MRL m� have characteristics similar to thoseof integrins
At least five distinct mechanisms, not mutually exclusive,could contribute to FBS-dependent, increased adhesion. First,MRL m� could adhere through a type or family of adhesionreceptors not used by nonautoimmune m�. Second, MRL m�may be a heterogeneous population with increased adhesionattributable to one or more subpopulations not found amongcontrol m�. Finally, if MRL m� use the same receptors as do
Fig. 2. Increased adhesion by MRL m� is corrected by FBS-free culture.After overnight, nonadherent culture, peritoneal m� from the indicated strainswere plated onto various surfaces in FBS-free medium, (A) without or (B) withLPS (100 ng/ml). After 16 h, nonadherent m� were washed away, and therelative number of remaining adherent m� was quantitated by MTT assay.There were no statistically significant differences in any of the four pair-wisecomparisons between control (BALB/c or C57BL/6) and MRL (MRL/� orMRL/lpr) m� for any surface, irrespective of the presence of LPS. Data arecombined from n � 4 independent experiments.
Fig. 3. Increased adhesion by MRL m� is corrected by delipidation of FBS.After overnight, nonadherent culture, peritoneal m� from the indicated strainswere plated onto various surfaces in the presence of 10% dFBS, (A) without or(B) with LPS (100 ng/ml). After 16 h, nonadherent m� were washed away, andthe relative number of remaining adherent m� was quantitated by MTT assay.There were no statistically significant differences in any of the four pair-wisecomparisons between control (BALB/c or C57BL/6) and MRL (MRL/� orMRL/lpr) m� for any surface, irrespective of the presence of LPS. Data arecombined from n � 3 independent experiments.
Longacre et al. Abnormalities in Rho and adhesion by MRL macrophages 5
nonautoimmune m�, then MRL m� might possess an in-creased number of one or several adhesion receptors, theaffinity of individual adhesion receptors for their ligands mightbe increased, or cytoskeletal alterations might lead to anoverall increase in the avidity of the interaction between ad-hesion receptors in aggregate and their cognate surface.
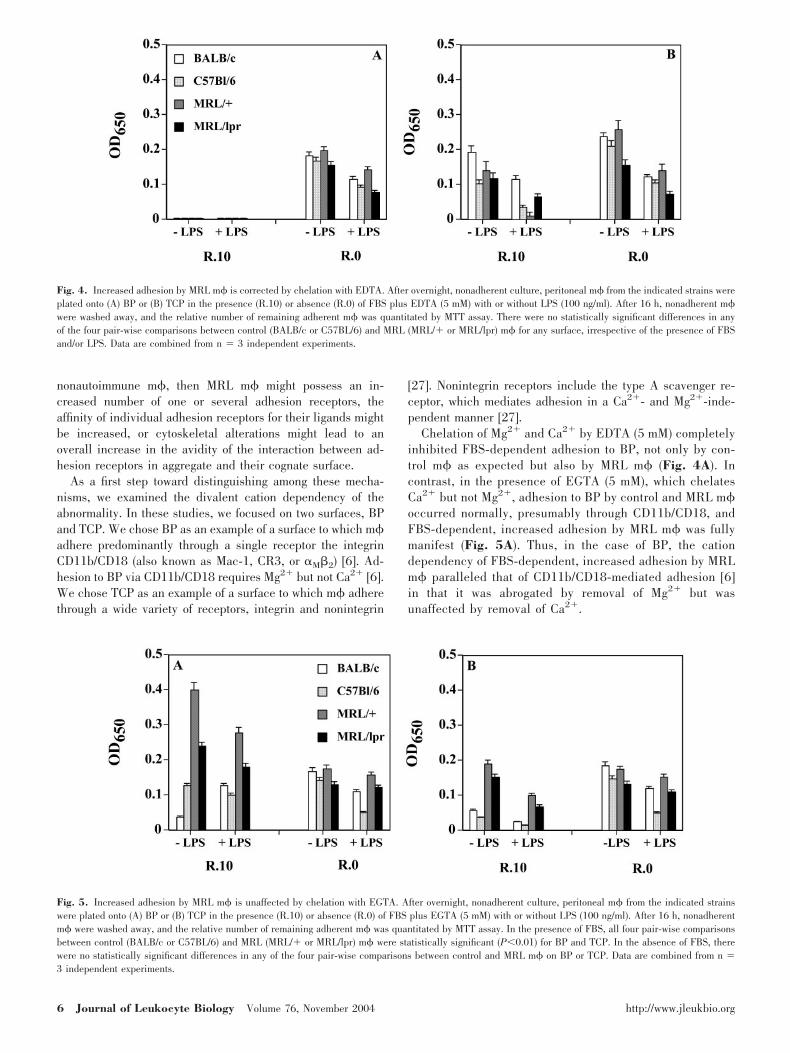
As a first step toward distinguishing among these mecha-nisms, we examined the divalent cation dependency of theabnormality. In these studies, we focused on two surfaces, BPand TCP. We chose BP as an example of a surface to which m�adhere predominantly through a single receptor the integrinCD11b/CD18 (also known as Mac-1, CR3, or M�2) [6]. Ad-hesion to BP via CD11b/CD18 requires Mg2� but not Ca2� [6].We chose TCP as an example of a surface to which m� adherethrough a wide variety of receptors, integrin and nonintegrin
[27]. Nonintegrin receptors include the type A scavenger re-ceptor, which mediates adhesion in a Ca2�- and Mg2�-inde-pendent manner [27].
Chelation of Mg2� and Ca2� by EDTA (5 mM) completelyinhibited FBS-dependent adhesion to BP, not only by con-trol m� as expected but also by MRL m� (Fig. 4A). Incontrast, in the presence of EGTA (5 mM), which chelatesCa2� but not Mg2�, adhesion to BP by control and MRL m�occurred normally, presumably through CD11b/CD18, andFBS-dependent, increased adhesion by MRL m� was fullymanifest (Fig. 5A). Thus, in the case of BP, the cationdependency of FBS-dependent, increased adhesion by MRLm� paralleled that of CD11b/CD18-mediated adhesion [6]in that it was abrogated by removal of Mg2� but wasunaffected by removal of Ca2�.
Fig. 4. Increased adhesion by MRL m� is corrected by chelation with EDTA. After overnight, nonadherent culture, peritoneal m� from the indicated strains wereplated onto (A) BP or (B) TCP in the presence (R.10) or absence (R.0) of FBS plus EDTA (5 mM) with or without LPS (100 ng/ml). After 16 h, nonadherent m�were washed away, and the relative number of remaining adherent m� was quantitated by MTT assay. There were no statistically significant differences in anyof the four pair-wise comparisons between control (BALB/c or C57BL/6) and MRL (MRL/� or MRL/lpr) m� for any surface, irrespective of the presence of FBSand/or LPS. Data are combined from n � 3 independent experiments.
Fig. 5. Increased adhesion by MRL m� is unaffected by chelation with EGTA. After overnight, nonadherent culture, peritoneal m� from the indicated strainswere plated onto (A) BP or (B) TCP in the presence (R.10) or absence (R.0) of FBS plus EGTA (5 mM) with or without LPS (100 ng/ml). After 16 h, nonadherentm� were washed away, and the relative number of remaining adherent m� was quantitated by MTT assay. In the presence of FBS, all four pair-wise comparisonsbetween control (BALB/c or C57BL/6) and MRL (MRL/� or MRL/lpr) m� were statistically significant (P0.01) for BP and TCP. In the absence of FBS, therewere no statistically significant differences in any of the four pair-wise comparisons between control and MRL m� on BP or TCP. Data are combined from n �3 independent experiments.
6 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org
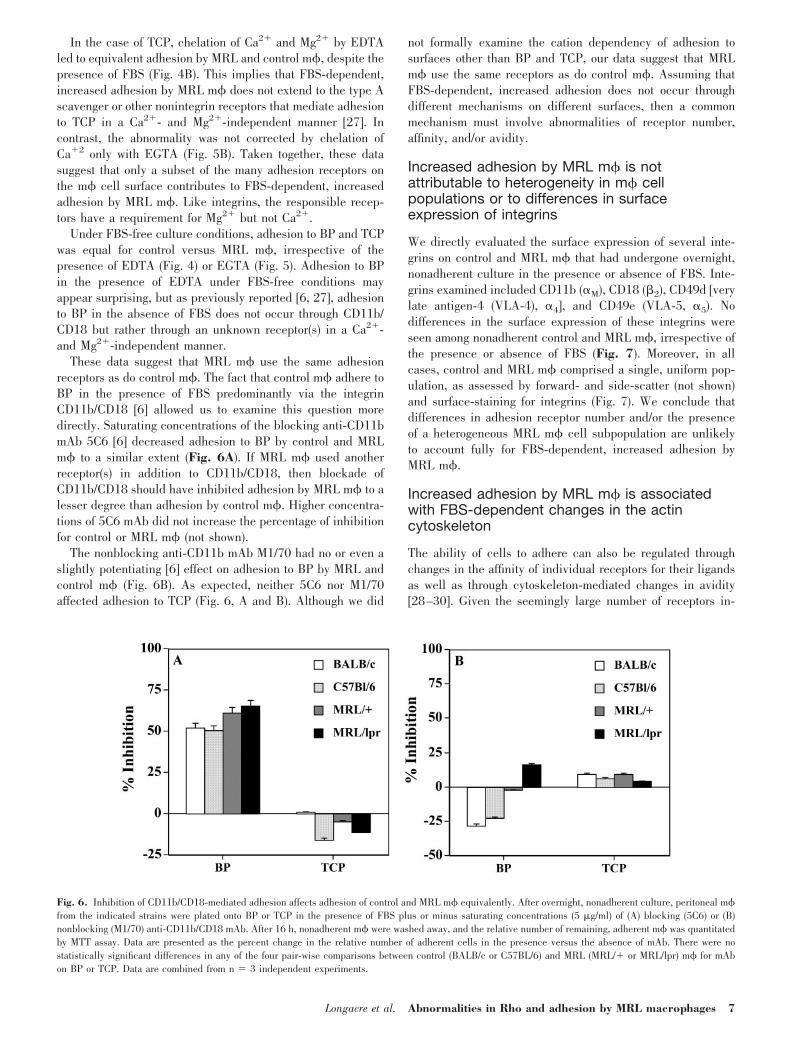
In the case of TCP, chelation of Ca2� and Mg2� by EDTAled to equivalent adhesion by MRL and control m�, despite thepresence of FBS (Fig. 4B). This implies that FBS-dependent,increased adhesion by MRL m� does not extend to the type Ascavenger or other nonintegrin receptors that mediate adhesionto TCP in a Ca2�- and Mg2�-independent manner [27]. Incontrast, the abnormality was not corrected by chelation ofCa�2 only with EGTA (Fig. 5B). Taken together, these datasuggest that only a subset of the many adhesion receptors onthe m� cell surface contributes to FBS-dependent, increasedadhesion by MRL m�. Like integrins, the responsible recep-tors have a requirement for Mg2� but not Ca2�.
Under FBS-free culture conditions, adhesion to BP and TCPwas equal for control versus MRL m�, irrespective of thepresence of EDTA (Fig. 4) or EGTA (Fig. 5). Adhesion to BPin the presence of EDTA under FBS-free conditions mayappear surprising, but as previously reported [6, 27], adhesionto BP in the absence of FBS does not occur through CD11b/CD18 but rather through an unknown receptor(s) in a Ca2�-and Mg2�-independent manner.
These data suggest that MRL m� use the same adhesionreceptors as do control m�. The fact that control m� adhere toBP in the presence of FBS predominantly via the integrinCD11b/CD18 [6] allowed us to examine this question moredirectly. Saturating concentrations of the blocking anti-CD11bmAb 5C6 [6] decreased adhesion to BP by control and MRLm� to a similar extent (Fig. 6A). If MRL m� used anotherreceptor(s) in addition to CD11b/CD18, then blockade ofCD11b/CD18 should have inhibited adhesion by MRL m� to alesser degree than adhesion by control m�. Higher concentra-tions of 5C6 mAb did not increase the percentage of inhibitionfor control or MRL m� (not shown).
The nonblocking anti-CD11b mAb M1/70 had no or even aslightly potentiating [6] effect on adhesion to BP by MRL andcontrol m� (Fig. 6B). As expected, neither 5C6 nor M1/70affected adhesion to TCP (Fig. 6, A and B). Although we did
not formally examine the cation dependency of adhesion tosurfaces other than BP and TCP, our data suggest that MRLm� use the same receptors as do control m�. Assuming thatFBS-dependent, increased adhesion does not occur throughdifferent mechanisms on different surfaces, then a commonmechanism must involve abnormalities of receptor number,affinity, and/or avidity.
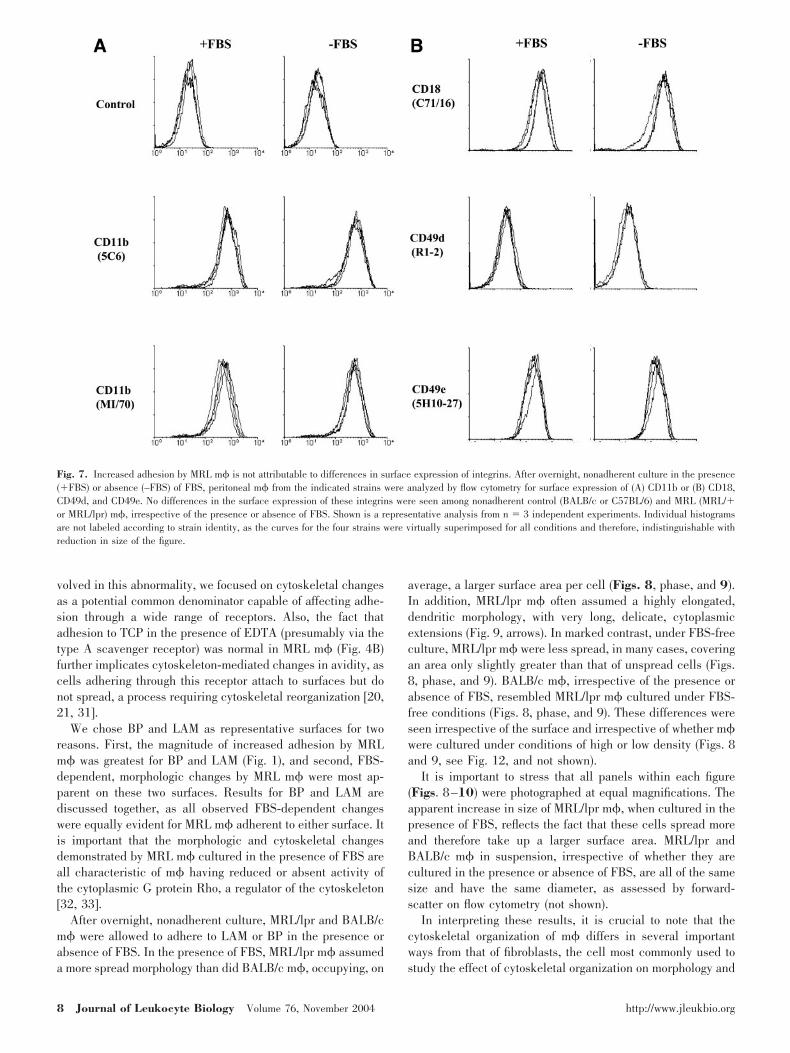
Increased adhesion by MRL m� is notattributable to heterogeneity in m� cellpopulations or to differences in surfaceexpression of integrins
We directly evaluated the surface expression of several inte-grins on control and MRL m� that had undergone overnight,nonadherent culture in the presence or absence of FBS. Inte-grins examined included CD11b ( M), CD18 (�2), CD49d [verylate antigen-4 (VLA-4), 4], and CD49e (VLA-5, 5). Nodifferences in the surface expression of these integrins wereseen among nonadherent control and MRL m�, irrespective ofthe presence or absence of FBS (Fig. 7). Moreover, in allcases, control and MRL m� comprised a single, uniform pop-ulation, as assessed by forward- and side-scatter (not shown)and surface-staining for integrins (Fig. 7). We conclude thatdifferences in adhesion receptor number and/or the presenceof a heterogeneous MRL m� cell subpopulation are unlikelyto account fully for FBS-dependent, increased adhesion byMRL m�.
Increased adhesion by MRL m� is associatedwith FBS-dependent changes in the actincytoskeleton
The ability of cells to adhere can also be regulated throughchanges in the affinity of individual receptors for their ligandsas well as through cytoskeleton-mediated changes in avidity[28–30]. Given the seemingly large number of receptors in-
Fig. 6. Inhibition of CD11b/CD18-mediated adhesion affects adhesion of control and MRL m� equivalently. After overnight, nonadherent culture, peritoneal m�from the indicated strains were plated onto BP or TCP in the presence of FBS plus or minus saturating concentrations (5 �g/ml) of (A) blocking (5C6) or (B)nonblocking (M1/70) anti-CD11b/CD18 mAb. After 16 h, nonadherent m� were washed away, and the relative number of remaining, adherent m� was quantitatedby MTT assay. Data are presented as the percent change in the relative number of adherent cells in the presence versus the absence of mAb. There were nostatistically significant differences in any of the four pair-wise comparisons between control (BALB/c or C57BL/6) and MRL (MRL/� or MRL/lpr) m� for mAbon BP or TCP. Data are combined from n � 3 independent experiments.
Longacre et al. Abnormalities in Rho and adhesion by MRL macrophages 7
volved in this abnormality, we focused on cytoskeletal changesas a potential common denominator capable of affecting adhe-sion through a wide range of receptors. Also, the fact thatadhesion to TCP in the presence of EDTA (presumably via thetype A scavenger receptor) was normal in MRL m� (Fig. 4B)further implicates cytoskeleton-mediated changes in avidity, ascells adhering through this receptor attach to surfaces but donot spread, a process requiring cytoskeletal reorganization [20,21, 31].
We chose BP and LAM as representative surfaces for tworeasons. First, the magnitude of increased adhesion by MRLm� was greatest for BP and LAM (Fig. 1), and second, FBS-dependent, morphologic changes by MRL m� were most ap-parent on these two surfaces. Results for BP and LAM arediscussed together, as all observed FBS-dependent changeswere equally evident for MRL m� adherent to either surface. Itis important that the morphologic and cytoskeletal changesdemonstrated by MRL m� cultured in the presence of FBS areall characteristic of m� having reduced or absent activity ofthe cytoplasmic G protein Rho, a regulator of the cytoskeleton[32, 33].
After overnight, nonadherent culture, MRL/lpr and BALB/cm� were allowed to adhere to LAM or BP in the presence orabsence of FBS. In the presence of FBS, MRL/lpr m� assumeda more spread morphology than did BALB/c m�, occupying, on
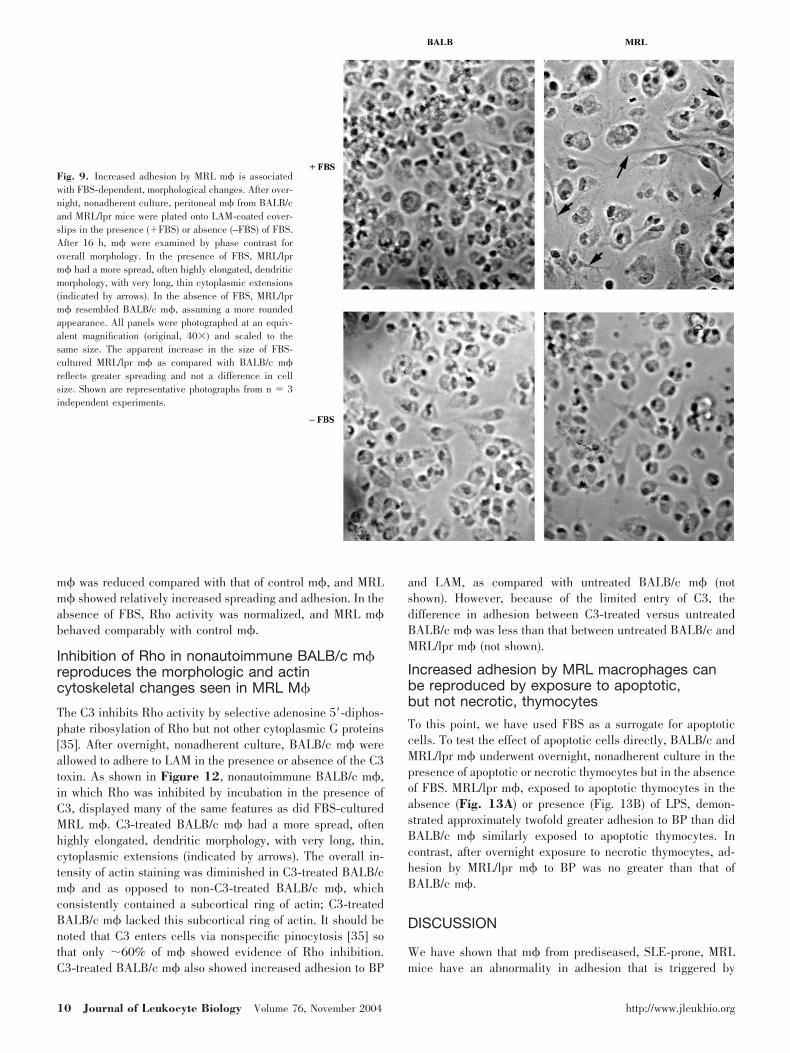
average, a larger surface area per cell (Figs. 8, phase, and 9).In addition, MRL/lpr m� often assumed a highly elongated,dendritic morphology, with very long, delicate, cytoplasmicextensions (Fig. 9, arrows). In marked contrast, under FBS-freeculture, MRL/lpr m� were less spread, in many cases, coveringan area only slightly greater than that of unspread cells (Figs.8, phase, and 9). BALB/c m�, irrespective of the presence orabsence of FBS, resembled MRL/lpr m� cultured under FBS-free conditions (Figs. 8, phase, and 9). These differences wereseen irrespective of the surface and irrespective of whether m�were cultured under conditions of high or low density (Figs. 8and 9, see Fig. 12, and not shown).
It is important to stress that all panels within each figure(Figs. 8–10) were photographed at equal magnifications. Theapparent increase in size of MRL/lpr m�, when cultured in thepresence of FBS, reflects the fact that these cells spread moreand therefore take up a larger surface area. MRL/lpr andBALB/c m� in suspension, irrespective of whether they arecultured in the presence or absence of FBS, are all of the samesize and have the same diameter, as assessed by forward-scatter on flow cytometry (not shown).
In interpreting these results, it is crucial to note that thecytoskeletal organization of m� differs in several importantways from that of fibroblasts, the cell most commonly used tostudy the effect of cytoskeletal organization on morphology and
Fig. 7. Increased adhesion by MRL m� is not attributable to differences in surface expression of integrins. After overnight, nonadherent culture in the presence(�FBS) or absence (–FBS) of FBS, peritoneal m� from the indicated strains were analyzed by flow cytometry for surface expression of (A) CD11b or (B) CD18,CD49d, and CD49e. No differences in the surface expression of these integrins were seen among nonadherent control (BALB/c or C57BL/6) and MRL (MRL/�or MRL/lpr) m�, irrespective of the presence or absence of FBS. Shown is a representative analysis from n � 3 independent experiments. Individual histogramsare not labeled according to strain identity, as the curves for the four strains were virtually superimposed for all conditions and therefore, indistinguishable withreduction in size of the figure.
8 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org

adhesion. As opposed to fibroblasts, cultured m� do not formfocal adhesion complexes [20, 32, 33]. In keeping with theabsence of focal adhesion complexes, Rho-activated m� alsodo not assemble actin into polymerized bundles of stress fibers,which in fibroblasts, converge on focal adhesion complexes.Instead, Rho-activated m� possess a finer contractile networkof actin filaments [20, 32, 33]. Without the anchoring providedby focal adhesion complexes, contraction of this actin filamentnetwork in m� leads to rounding up of m�, as opposed to the
spread-out morphology seen in Rho-activated fibroblasts [20,32, 33].
Staining for the focal adhesion protein vinculin confirmedthe absence of focal adhesion complexes in BALB/c and MRL/lpr m�, irrespective of the presence or absence of FBS (notshown). In accord with the absence of focal adhesion com-plexes, vinculin was distributed uniformly over the m� cellbody, without any differences in the staining pattern betweenBALB/c and MRL/lpr m� (not shown).
It is remarkable that in accord with the increased adhesionshown by MRL/lpr m� (Fig. 1), we observed dramatic FBS-dependent differences in cytoskeletal organization betweenMRL/lpr and BALB/c m�. In the presence of FBS, the overallintensity of actin staining was consistently diminished in MRL/lpr compared with BALB/c m� (Fig. 8, ACTIN-FITC, and Fig.10). In addition, BALB/c m� usually contained a subcorticalactin band, most visible in a higher plane of focus (Fig. 10,arrows). MRL/lpr m�, in the presence of FBS, consistentlylacked this subcortical actin band. Instead of a subcorticalactin band, MRL/lpr m� frequently displayed a concentratedstaining of subcortical actin within dense, plaque-like struc-tures near the cell periphery (Fig. 10). Notably, in the absenceof FBS, the pattern of actin staining did not differ betweenMRL/lpr and BALB/c m� and closely resembled that seen inFBS-cultured BALB/c m� (Figs. 8 and 10).
The micrographs shown in Figures 8–10 were obtained after16 h of adhesion. Identical, although less marked, FBS-depen-dent differences in cytoskeletal organization were observed inMRL/lpr m� after 6 h of adhesion (not shown). The morphologyand staining of C57BL/6 m� paralleled that of BALB/c m�(not shown). Similarly, MRL/� and MRL/lpr m� were indis-tinguishable in morphology and staining (not shown). Finally,to confirm our results for vinculin as well as to rule outdifferences in membrane clustering of integrins, we alsostained BP-adherent MRL/lpr and BALB/c m� for CD11b.Staining for CD11b was distributed uniformly over the m� cellbody and closely resembled that for vinculin, with no differ-ences seen between MRL and control m� (not shown).
MRL m� cultured in the presence of FBS havedecreased, activated Rho
The morphology and cytoskeletal organization of MRL m�,cultured in the presence but not the absence of FBS, wasconsistent with reduced Rho activity compared with that ofBALB/c m� [20, 32, 33] (Figs. 8–10). To determine whetherFBS-dependent dysregulation of Rho activity may underlieFBS-dependent, increased adhesion by MRL m�, we measuredRho activity directly.
After overnight, nonadherent culture in the presence orabsence of FBS, lysates prepared from equal numbers of MRL/lpr and BALB/c m� were examined for active Rho, accordingto assay specifications [34]. Consistent with the results ofcytoskeletal staining, active Rho • GTP was reduced in FBS-cultured MRL/lpr m� compared with BALB/c m� (Fig. 11).Under FBS-free conditions, active Rho did not differ betweenMRL/lpr and BALB/c m� and exceeded by several-fold thatfound in FBS-cultured MRL/lpr m� (Fig. 11). Thus, the levelof active Rho correlated with the observed adhesion abnormal-ity of MRL m�. In the presence of FBS, Rho activity of MRL
Fig. 8. Increased adhesion by MRL m� is associated with FBS-dependent,morphologic and actin cytoskeletal changes. After overnight, nonadherentculture, peritoneal m� from BALB/c and MRL/lpr mice were plated ontoLAM-coated coverslips in the presence (�FBS) or absence (–FBS) of FBS.After 16 h, m� were examined by phase contrast for morphology and epiflu-orescence microscopy for staining of actin (ACTIN-FITC). The appearance ofBALB/c m� did not differ with the presence or absence of FBS. In contrast,MRL/lpr m� cultured in the presence of FBS displayed several notablefeatures in comparison with BALB/c m� and FBS-free MRL/lpr m�. In thepresence of FBS, MRL/lpr m� had a more spread, often highly elongated,“dendritic” morphology, with very long, thin cytoplasmic extensions. In addi-tion, the overall intensity of actin staining was consistently diminished inFBS-cultured MRL/lpr m�, which also consistently lacked the subcortical ringof actin seen in BALB/c m� and FBS-free MRL/lpr m�. All panels werephotographed at an equivalent magnification (original, 40�) and scaled to thesame size. Shown are representative photographs from n � 3 independentexperiments.
Longacre et al. Abnormalities in Rho and adhesion by MRL macrophages 9
m� was reduced compared with that of control m�, and MRLm� showed relatively increased spreading and adhesion. In theabsence of FBS, Rho activity was normalized, and MRL m�behaved comparably with control m�.
Inhibition of Rho in nonautoimmune BALB/c m�reproduces the morphologic and actincytoskeletal changes seen in MRL M�
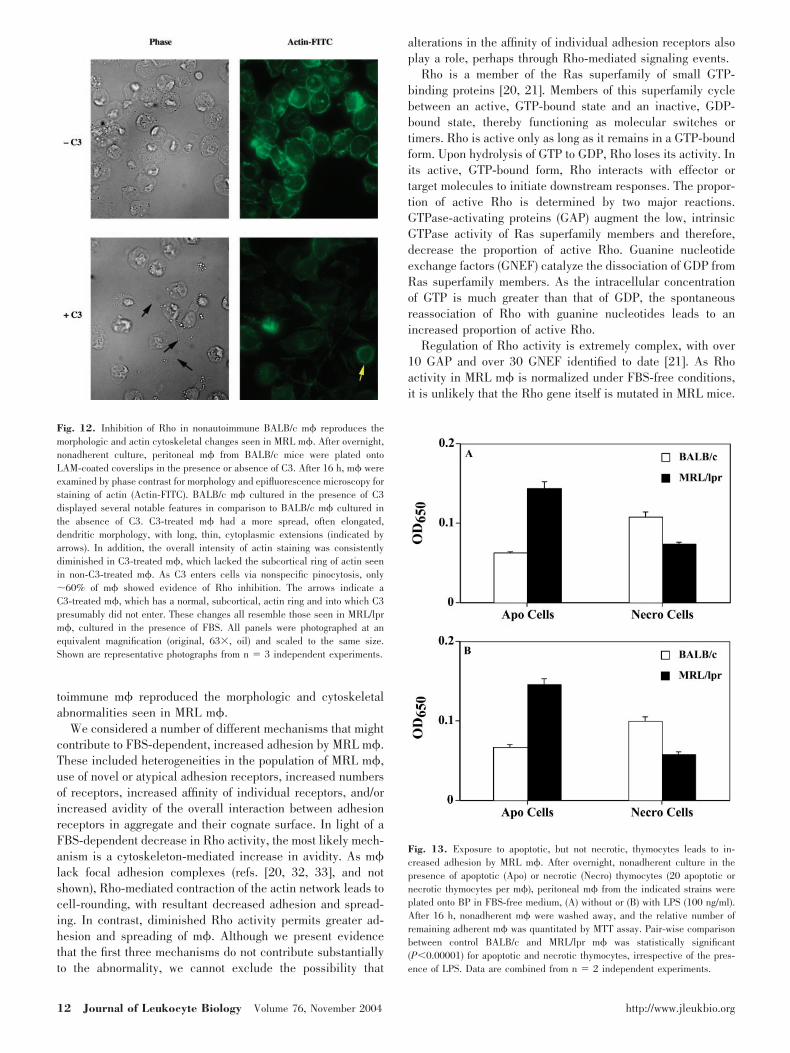
The C3 inhibits Rho activity by selective adenosine 5�-diphos-phate ribosylation of Rho but not other cytoplasmic G proteins[35]. After overnight, nonadherent culture, BALB/c m� wereallowed to adhere to LAM in the presence or absence of the C3toxin. As shown in Figure 12, nonautoimmune BALB/c m�,in which Rho was inhibited by incubation in the presence ofC3, displayed many of the same features as did FBS-culturedMRL m�. C3-treated BALB/c m� had a more spread, oftenhighly elongated, dendritic morphology, with very long, thin,cytoplasmic extensions (indicated by arrows). The overall in-tensity of actin staining was diminished in C3-treated BALB/cm� and as opposed to non-C3-treated BALB/c m�, whichconsistently contained a subcortical ring of actin; C3-treatedBALB/c m� lacked this subcortical ring of actin. It should benoted that C3 enters cells via nonspecific pinocytosis [35] sothat only �60% of m� showed evidence of Rho inhibition.C3-treated BALB/c m� also showed increased adhesion to BP
and LAM, as compared with untreated BALB/c m� (notshown). However, because of the limited entry of C3, thedifference in adhesion between C3-treated versus untreatedBALB/c m� was less than that between untreated BALB/c andMRL/lpr m� (not shown).
Increased adhesion by MRL macrophages canbe reproduced by exposure to apoptotic,but not necrotic, thymocytes
To this point, we have used FBS as a surrogate for apoptoticcells. To test the effect of apoptotic cells directly, BALB/c andMRL/lpr m� underwent overnight, nonadherent culture in thepresence of apoptotic or necrotic thymocytes but in the absenceof FBS. MRL/lpr m�, exposed to apoptotic thymocytes in theabsence (Fig. 13A) or presence (Fig. 13B) of LPS, demon-strated approximately twofold greater adhesion to BP than didBALB/c m� similarly exposed to apoptotic thymocytes. Incontrast, after overnight exposure to necrotic thymocytes, ad-hesion by MRL/lpr m� to BP was no greater than that ofBALB/c m�.
DISCUSSION
We have shown that m� from prediseased, SLE-prone, MRLmice have an abnormality in adhesion that is triggered by
Fig. 9. Increased adhesion by MRL m� is associatedwith FBS-dependent, morphological changes. After over-night, nonadherent culture, peritoneal m� from BALB/cand MRL/lpr mice were plated onto LAM-coated cover-slips in the presence (�FBS) or absence (–FBS) of FBS.After 16 h, m� were examined by phase contrast foroverall morphology. In the presence of FBS, MRL/lprm� had a more spread, often highly elongated, dendriticmorphology, with very long, thin cytoplasmic extensions(indicated by arrows). In the absence of FBS, MRL/lprm� resembled BALB/c m�, assuming a more roundedappearance. All panels were photographed at an equiv-alent magnification (original, 40�) and scaled to thesame size. The apparent increase in the size of FBS-cultured MRL/lpr m� as compared with BALB/c m�reflects greater spreading and not a difference in cellsize. Shown are representative photographs from n � 3independent experiments.
10 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org

serum lipids. In the presence of FBS, m� from MRL/� andMRL/lpr mice adhered in increased numbers to a variety ofsurfaces and ECM proteins, including TCP, BP, COL I, FN,and LAM, as compared with m� from two nonautoimmunestrains. When MRL m� were cultured under FBS-free condi-tions or in the presence of FBS that had been delipidated, theiradhesion was similar to that of m� from nonautoimmune mice.Microscopic examination of adherent MRL m� cultured in thepresence but not the absence of FBS revealed a dendriticspread morphology and an actin-staining pattern consistentwith reduced activity of Rho, a cytoplasmic G protein andcytoskeletal regulator [32, 33]. This was confirmed by thefinding that MRL m�, cultured in the presence of FBS, showeddecreased levels of active Rho when compared with nonauto-immune m�. In sharp contrast, when MRL m� were culturedin the absence of FBS, levels of active Rho were increased tonormal levels. Thus, Rho activity in MRL m� paralleled ex-actly the FBS-dependent abnormalities that we observed inadhesion and cytoskeletal organization: normal in the absenceof FBS and abnormal in the presence of FBS. As in our studieson cytokine gene expression, apoptotic cells and FBS wereequivalent in eliciting the defect. In the presence of apoptoticbut not necrotic cells, MRL m� adhered in increased numbersto BP, similarly to that observed in the presence of FBS.Finally, inhibition of Rho with C3 exoenzyme [35] in nonau-
Fig. 10. Increased adhesion by MRL m� is associatedwith FBS-dependent, actin, cytoskeletal changes. Afterovernight, nonadherent culture, peritoneal m� fromBALB/c and MRL/lpr mice were plated onto LAM-coatedcoverslips in the presence (�FBS) or absence (–FBS) ofFBS. After 16 h, m� were examined by fluorescencemicroscopy for staining of actin. In the presence of FBS,the overall intensity of actin staining was consistentlydiminished in MRL/lpr compared with BALB/c m�,which often contained a subcortical ring of actin (indi-cated by arrows), irrespective of the presence or absenceof FBS. MRL/lpr m�, in the presence of FBS, consis-tently lacked this subcortical ring of actin and instead,frequently displayed a concentrated staining of subcorti-cal actin within dense, plaque-like structures near thecell periphery. The pattern of actin staining in FBS-freeMRL/lpr m� closely resembled that seen in BALB/c m�.All panels were photographed at an equivalent magnifi-cation (original, 40�) and scaled to the same size. Shownare representative photographs from n � 3 independentexperiments.
Fig. 11. MRL m� cultured in the presence of FBS have decreased, activatedRho. After overnight, nonadherent culture in the presence or absence of FBS,lysates prepared from equal numbers of MRL/lpr and BALB/c m� wereexamined for active Rho by a pull-down assay using a fusion protein betweenglutathione S-transferase and the Rho-binding domain of the rhotekin protein,to which only active Rho • GTP binds. The quantity of active Rho • GTP wasreduced in FBS-cultured MRL/lpr m� compared with BALB/c m�, cultured inthe presence or absence of FBS. Under FBS-free conditions, the quantity ofactive Rho in MRL/lpr m� was normalized to levels seen in BALB/c m� andexceeded that found in FBS-cultured MRL/lpr m�. To ensure that lysates usedin the pull-down assay were derived from equivalent numbers of m�, analiquot of the original lysate (before mixing with agarose beads in the pull-down assay) was probed for total Rho (not shown). Shown is a representativeblot from n � 3 independent experiments.
Longacre et al. Abnormalities in Rho and adhesion by MRL macrophages 11
toimmune m� reproduced the morphologic and cytoskeletalabnormalities seen in MRL m�.
We considered a number of different mechanisms that mightcontribute to FBS-dependent, increased adhesion by MRL m�.These included heterogeneities in the population of MRL m�,use of novel or atypical adhesion receptors, increased numbersof receptors, increased affinity of individual receptors, and/orincreased avidity of the overall interaction between adhesionreceptors in aggregate and their cognate surface. In light of aFBS-dependent decrease in Rho activity, the most likely mech-anism is a cytoskeleton-mediated increase in avidity. As m�lack focal adhesion complexes (refs. [20, 32, 33], and notshown), Rho-mediated contraction of the actin network leads tocell-rounding, with resultant decreased adhesion and spread-ing. In contrast, diminished Rho activity permits greater ad-hesion and spreading of m�. Although we present evidencethat the first three mechanisms do not contribute substantiallyto the abnormality, we cannot exclude the possibility that
alterations in the affinity of individual adhesion receptors alsoplay a role, perhaps through Rho-mediated signaling events.
Rho is a member of the Ras superfamily of small GTP-binding proteins [20, 21]. Members of this superfamily cyclebetween an active, GTP-bound state and an inactive, GDP-bound state, thereby functioning as molecular switches ortimers. Rho is active only as long as it remains in a GTP-boundform. Upon hydrolysis of GTP to GDP, Rho loses its activity. Inits active, GTP-bound form, Rho interacts with effector ortarget molecules to initiate downstream responses. The propor-tion of active Rho is determined by two major reactions.GTPase-activating proteins (GAP) augment the low, intrinsicGTPase activity of Ras superfamily members and therefore,decrease the proportion of active Rho. Guanine nucleotideexchange factors (GNEF) catalyze the dissociation of GDP fromRas superfamily members. As the intracellular concentrationof GTP is much greater than that of GDP, the spontaneousreassociation of Rho with guanine nucleotides leads to anincreased proportion of active Rho.
Regulation of Rho activity is extremely complex, with over10 GAP and over 30 GNEF identified to date [21]. As Rhoactivity in MRL m� is normalized under FBS-free conditions,it is unlikely that the Rho gene itself is mutated in MRL mice.
Fig. 13. Exposure to apoptotic, but not necrotic, thymocytes leads to in-creased adhesion by MRL m�. After overnight, nonadherent culture in thepresence of apoptotic (Apo) or necrotic (Necro) thymocytes (20 apoptotic ornecrotic thymocytes per m�), peritoneal m� from the indicated strains wereplated onto BP in FBS-free medium, (A) without or (B) with LPS (100 ng/ml).After 16 h, nonadherent m� were washed away, and the relative number ofremaining adherent m� was quantitated by MTT assay. Pair-wise comparisonbetween control BALB/c and MRL/lpr m� was statistically significant(P0.00001) for apoptotic and necrotic thymocytes, irrespective of the pres-ence of LPS. Data are combined from n � 2 independent experiments.
Fig. 12. Inhibition of Rho in nonautoimmune BALB/c m� reproduces themorphologic and actin cytoskeletal changes seen in MRL m�. After overnight,nonadherent culture, peritoneal m� from BALB/c mice were plated ontoLAM-coated coverslips in the presence or absence of C3. After 16 h, m� wereexamined by phase contrast for morphology and epifluorescence microscopy forstaining of actin (Actin-FITC). BALB/c m� cultured in the presence of C3displayed several notable features in comparison to BALB/c m� cultured inthe absence of C3. C3-treated m� had a more spread, often elongated,dendritic morphology, with long, thin, cytoplasmic extensions (indicated byarrows). In addition, the overall intensity of actin staining was consistentlydiminished in C3-treated m�, which lacked the subcortical ring of actin seenin non-C3-treated m�. As C3 enters cells via nonspecific pinocytosis, only�60% of m� showed evidence of Rho inhibition. The arrows indicate aC3-treated m�, which has a normal, subcortical, actin ring and into which C3presumably did not enter. These changes all resemble those seen in MRL/lprm�, cultured in the presence of FBS. All panels were photographed at anequivalent magnification (original, 63�, oil) and scaled to the same size.Shown are representative photographs from n � 3 independent experiments.
12 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org

Rather, the responsible abnormality most likely involves one ormore of the GAP and GNEF, which interact directly with Rhoor other pathways further upstream that regulate these GAPand GNEF.
We have also examined the role of apoptotic cells in trig-gering the observed abnormalities in adhesion and demon-strated that increased numbers of MRL m� adhered in thepresence of apoptotic but not necrotic cells. We have previ-ously shown that apoptotic cells fully mimic the effect of FBSin eliciting dysregulated cytokine expression by m� from MRLand other SLE-prone strains [18, 19]. The simplest hypothesisto explain our data is that MRL mice have a mutation or allelicdifference affecting signal transduction within a specific Rho-regulatory pathway, which is triggered by the recognitionand/or uptake of apoptotic cells. Although the identity of theresponsible lipids in FBS that mimic the effects of apoptoticcells remains undefined, the most likely candidates are oxida-tively modified lipids, lipoproteins, or lipid-protein adducts,whose clearance by m� depends on many of the same receptorsused to clear apoptotic cells [36–38].
The consequences of dysregulated Rho activity may not belimited to the adhesive properties of MRL m�. The number ofdownstream targets and effector functions known to be influ-enced by Rho is continually increasing [21]. Many of these aredependent on the actin cytoskeleton and include cell migration[39], complement-dependent phagocytosis [40], and cytokine-sis [41]. It is important that activated Rho also regulates anumber of other signal transduction pathways that are inde-pendent of the cytoskeleton. These include activation of thetranscription factors serum response factor [42] and nuclearfactor-�B [43], cell-cycle progression [44], and cadherin-me-diated, cell-cell contact [45]. Hence, an abnormality in theregulation of Rho could have wide-reaching effects on immunehomeostasis, affecting such m� functions as migration, traf-ficking, antigen uptake, and interaction with other immunecells. All of these functions are crucial to the balance betweentolerance and immunity. Indeed, alterations in the function orexpression of various adhesion molecules have been shown tomodulate the course of disease in several murine models ofautoimmunity [4, 10–14].
Moreover, recent studies have shown that cytoskeletalevents within the APC play a critical role, not only in formationof the immunologic synapse but also in T cell activation [46,47], so that the Rho-dependent cytoskeletal abnormalities wehave described in MRL m� may affect the manner in which Tcells are activated. Given that dysregulated Rho activity isdependent on the presence of FBS (a surrogate for apoptoticcells), abnormalities in T cell activation may be limited tothose antigens that are presented by APC in the context ofapoptotic cells. The potential dependence of dysregulated Rhoactivity on exposure to apoptotic cells is especially noteworthyin that we and others have shown that apoptotic cells and theirproducts are the antigenic targets of multiple autoantibodiesacross a broad spectrum of autoimmune diseases, includingSLE, antiphospholipid syndrome, scleroderma, and antineutro-phil, cytoplasmic, Ab-positive vasculitis [48–55].
In summary, we have shown that m� from prediseased,SLE-prone MRL mice have a FBS-dependent abnormality inadhesion, which appears to be the result of FBS-dependent
dysregulation in the activity of the cytoplasmic G protein Rho.In an earlier study, we showed that m� from MRL and multipleother SLE-prone strains, as well as the autoimmune, diabetes-prone NOD strain, also have a FBS- and apoptotic cell-depen-dent abnormality in the expression of multiple cytokines [18,19]. FBS-dependent dysregulation in Rho activity extends toall of the same SLE-prone strains (MRL/�, MRL/lpr, NZB,NZW, NZB/W F1, BXSB, and LG/J), plus the NOD strain,whereas no similar defect in Rho activity was found in sevennonautoimmune, control strains (H. Fan, A. Longacre, V. A.Patel, and J. S. Levine, manuscript in preparation). Takentogether, the results of these studies suggest that m� from MRLand other autoimmune-prone strains may have a shared auto-immune phenotype that is triggered by the uptake of apoptoticcells and manifests as abnormalities within a broad range ofm� functions. In the absence of apoptotic cells (or theirsurrogate, FBS lipids), m� from SLE-prone mice behave moreor less normally. We hypothesize that MRL and other autoim-mune-prone strains may be linked by a mutation(s) affecting acommon signaling element (or separate elements within acommon signaling pathway), which is activated in response toapoptotic cells. The consequence of such aberrant signalingwould be the “misreading” of apoptotic cells by m� andperhaps other APC in autoimmune-prone mice. Given theimportant role that signaling events induced by apoptotic cellslikely play in the balance between tolerance and immunity, themisreading of apoptotic cells has broad potential to induceautoimmunity.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health (NIH)Grants DK59793 (J. S. L.) and HL69722 (J. S. L.) and NIHTraining Grant T32DK07510 (A. L.). The authors thankCharles Whittaker for assistance in surface staining of macro-phages for flow cytometry; the University of Chicago IntegratedMicroscopy Core, Cancer Center Digital Light Facility, espe-cially Shirley Bond, for assistance in microscopic analysis ofthe cytoskeleton; David Lourim and Srikala Rhagavan forassistance in the design and interpretation of our cytoskeletalstudies; and Joyce Rauch and Martin Alexander Schwartz forcritical review of the manuscript. A. L. and J. S. K. contributedequally to this work.
REFERENCES
1. Miyamoto, Y. J., Andruss, B. F., Mitchell, J. S., Billard, M. J., McIntyre,B. W. (2003) Diverse roles of integrins in human T lymphocyte biology.Immunol. Res. 27, 71–84.
2. Hynes, R. O. (2002) Integrins: bidirectional, allosteric signaling ma-chines. Cell 110, 673–687.
3. Patel, K. D., Cuvelier, S. L., Wiehler, S. (2002) Selectins: critical medi-ators of leukocyte recruitment. Semin. Immunol. 14, 73–81.
4. Ley, K. (2003) The role of selectins in inflammation and disease. TrendsMol. Med. 9, 263–268.
5. Potter, P. K., Cortes-Hernandez, J., Quartier, P., Botto, M., Walport, M. J.(2003) Lupus-prone mice have an abnormal response to thioglycolate andan impaired clearance of apoptotic cells. J. Immunol. 170, 3223–3232.
Longacre et al. Abnormalities in Rho and adhesion by MRL macrophages 13

6. Rosen, H., Gordon, S. (1987) Monoclonal antibody to the murine type 3complement receptor inhibits adhesion of myelomonocytic cells in vitroand inflammatory cell recruitment in vivo. J. Exp. Med. 166, 1685–1701.
7. Von Andrian, U. H., Mackay, C. R. (2000) T cell function and migration:two sides of the same coin. N. Engl. J. Med. 343, 1020–1034.
8. Acuto, O., Cantrell, D. (2000) T cell activation and the cytoskeleton. Annu.Rev. Immunol. 18, 165–184.
9. Dustin, M. L., Cooper, J. A. (2000) The immunologic synapse and the actincytoskeleton: molecular hardware for T cell signaling. Nat. Immunol. 1, 23–29.
10. Wehrle-Haller, B., Imhof, B. A. (2003) Integrin-dependent pathologies.J. Pathol. 200, 481–487.
11. Hamm, C. W. (2003) Anti-integrin therapy. Annu. Rev. Med. 54, 425–435.12. Hutchings, P., Rosen, H., O’Reilly, L., Simpson, E., Gordon, S., Cooke, A.
(1990) Transfer of diabetes in mice prevented by blockade of adhesion-promoting receptor on macrophages. Nature 348, 639–642.
13. Bullard, D. C., King, P. D., Hicks, M. J., Dupont, B., Beaudet, A. L., Elkon,K. B. (1997) Intercellular adhesion molecule-1 deficiency protects MRL/MpJ-Fas(lpr) mice from early lethality. J. Immunol. 159, 2058–2067.
14. Bullard, D. C. (2002) Adhesion molecules in inflammatory diseases:insights from knockout mice. Immunol. Res. 26, 27–33.
15. Donnelly, R. P., Levine, J., Hartwell, D. W., Frendl, G., Fenton, M. J.,Beller, D. I. (1990) Aberrant regulation of interleukin 1 expression inmacrophages from young autoimmune-prone mice. J. Immunol. 145,3231–3239.
16. Levine, J. S., Pugh, B. J., Hartwell, D., Fitzpatrick, J. M., Marshak-Rothstein, A., Beller, D. I. (1993) IL-1 dysregulation is an intrinsic defectin macrophages from MRL autoimmune-prone mice. Eur. J. Immunol. 23,2951–2958.
17. Levine, J. S., Koh, J. S., Hartwell, D., Beller, D. I. (1998) Adhesion elicitsan intrinsic macrophage defect in interleukin-1 expression by autoim-mune-prone MRL mice. J. Autoimmun. 11, 141–150.
18. Koh, J. S., Wang, Z., Levine, J. S. (2000) Cytokine dysregulation inducedby apoptotic cells is a shared characteristic of murine lupus. J. Immunol.165, 4190–4201.
19. Fan, H., Longacre, A., Meng, F., Patel, V., Hsiao, K., Koh, J. S., Levine,J. S. (2004) Cytokine dysregulation induced by apoptotic cells is a sharedcharacteristic of macrophages from NOD and SLE-prone mice. J. Immu-nol. 172, 4834–4843.
20. Hall, A. (1998) Rho GTPases and the actin cytoskeleton. Science 279,509–514.
21. Bishop, A. L., Hall, A. (2000) Rho GTPases and their effector proteins.Biochem. J. 348, 241–255.
22. Reddy, S. M., Hsiao, K. H., Abernethy, V. E., Fan, H., Longacre, A.,Lieberthal, W., Rauch, J., Koh, J. S., Levine, J. S. (2002) Phagocytosis ofapoptotic cells by macrophages induces novel signaling events leading tocytokine-independent survival and inhibition of proliferation: activation ofAkt and inhibition of extracellular signal-regulated kinases 1 and 2.J. Immunol. 169, 702–713.
23. Hardy, R. W. (1986) Purification and characterization of monoclonalantibodies. In Handbook of Experimental Immunology (D. M. Weir, L. A.Herzenberg, C. Blackwell, L. A. Herzenberg, eds.), Oxford, UK, BlackwellScientific, 13.1–13.13.
24. Goldstein, J. L., Basu, S. K., Brown, M. S. (1983) Receptor-mediatedendocytosis of low-density lipoprotein in cultured cells. Methods Enzymol.98, 241–260.
25. Mossmann, T. (1983) Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity assays. J. Immunol.Methods 65, 55–63.
26. Shaw, L., Messier, J. M., Mercurio, A. M. (1990) The activation-dependentadhesion of macrophages to laminin involves cytoskeletal anchoring andphosphorylation of the 6�1 integrin. J. Cell Biol. 110, 2167–2174.
27. Fraser, I., Hughes, D., Gordon, S. (1993) Divalent cation-independentmacrophage adhesion inhibited by monoclonal antibody to murine scav-enger receptor. Nature 364, 343–346.
28. Stewart, M., Hogg, N. (1996) Regulation of leukocyte integrin function:affinity vs. avidity. J. Cell. Biochem. 61, 554–561.
29. Kolanus, W., Seed, B. (1997) Integrins and inside-out signal transduction:converging signals from PKC and PIP3. Curr. Opin. Cell Biol. 9, 725–731.
30. Hughes, P. E., Pfaff, M. (1998) Integrin affinity modulation. Trends CellBiol. 8, 359–364.
31. Hall, A. (1994) Small GTP-binding proteins and the regulation of the actincytoskeleton. Annu. Rev. Cell Biol. 10, 31–54.
32. Aepfelbacher, M., Essler, M., Huber, E., Czech, A., Weber, P. C. (1996)Rho is a negative regulator of human monocyte spreading. J. Immunol.157, 5070–5075.
33. Allen, W. E., Jones, G. E., Pollard, J. W., Ridley, A. J. (1997) Rho, Rac,and Cdc42 regulate actin organization and cell adhesion in macrophages.J. Cell Sci. 110, 707–720.
34. Ren, X-D., Schwartz, M. A. (2000) Determination of GTP loading on Rho.Methods Enzymol. 325, 264–272.
35. Genth, H., Gerhard, R., Maeda, A., Amano, M., Kaibuchi, K., Aktories,K., Just, I. (2003) Entrapment of Rho ADP-ribosylated by Clostridiumbotulinum C3 exoenzyme in the Rho-guanine nucleotide dissociationinhibitor-1 complex. J. Biol. Chem. 278, 28523–28527.
36. Terpstra, V., Kondratenko, N., Steinberg, D. (1997) Macrophages lackingscavenger receptor A show a decrease in binding and uptake of acetylatedlow-density lipoprotein and of apoptotic thymocytes, but not of oxidativelydamaged red blood cells. Proc. Natl. Acad. Sci. USA 94, 8127–8131.
37. Fukasawa, M., Adachi, H., Hirota, K., Tsujimoto, M., Arai, H., Inoue, K.(1996) SRB1, a class B scavenger receptor, recognizes both negativelycharged liposomes and apoptotic cells. Exp. Cell Res. 222, 246–250.
38. Chang, M-K., Bergmark, C., Laurila, A., Horkko, S., Han, K-H., Fried-man, P., Dennis, E. A., Witztum, J. L. (1999) Monoclonal antibodiesagainst oxidized low-density lipoprotein bind to apoptotic cells and inhibittheir phagocytosis by elicited macrophages: evidence that oxidation-spe-cific epitopes mediate macrophage recognition. Proc. Natl. Acad. Sci. USA96, 6353–6358.
39. Hall, A., Nobes, C. D. (2000) Rho GTPases: molecular switches thatcontrol the organization and dynamics of the actin cytoskeleton. Philos.Trans. R. Soc. Lond. B Biol. Sci. 355, 965–970.
40. Caron, E., Hall, A. (1998) Identification of two distinct mechanisms ofphagocytosis controlled by different Rho GTPases. Science 282, 1717–1721.
41. Prokopenko, S. N., Saint, R., Bellen, H. J. (2000) Untying the Gordianknot of cytokinesis: role of small G proteins and their regulators. J. CellBiol. 148, 843–848.
42. Hill, C. S., Wayne, J., Treisman, R. (1995) The Rho family GTPasesRhoA, Rac1, and CDC42Hs regulate transcriptional activity by SRF. Cell81, 1159–1170.
43. Perona, R., Montaner, S., Saniger, L., Sanchez-Perez, I., Bravo, R., Lacal,J. C. (1997) Activation of the nuclear factor-�B by Rho, CDC42, andRac-1 proteins. Genes Dev. 11, 463–475.
44. Olson, M. F., Ashworth, A., Hall, A. (1995) An essential role for Rho, Rac,and Cdc42 GTPases in cell cycle progression through G1. Science 269,1270–1272.
45. Kaibuchi, K., Kuroda, S., Fukata, M., Nakagawa, M. (1999) Regulation ofcadherin-mediated cell-cell adhesion by the Rho family GTPases. Curr.Opin. Cell Biol. 11, 591–596.
46. Al-Alwan, M. M., Rowden, G., Lee, T. D. G., West, K. A. (2001) Thedendritic cell cytoskeleton is critical for the formation of the immunolog-ical synapse. J. Immunol. 166, 1452–1456.
47. Al-Alwan, M. M., Liwski, R. S., Haeryfar, S. M. M., Baldridge, W. H.,Hoskin, D. W., Rowden, G., West, K. A. (2003) Dendritic cell actincytoskeletal polarization during immunological synapse formation ishighly antigen-dependent. J. Immunol. 171, 4479–4483.
48. Casciola-Rosen, L. A., Anhalt, G., Rosen, A. (1994) Autoantigens targetedin systemic lupus erythematosus are clustered in two populations ofsurface structures on apoptotic keratinocytes. J. Exp. Med. 179, 1317–1330.
49. Levine, J. S., Koh, J. S. (1999) The role of apoptosis in autoimmunity:immunogen, antigen, and accelerant. Semin. Nephrol. 19, 34–47.
50. Price, B. E., Rauch, J., Shia, M. A., Walsh, M. T., Lieberthal, W., Gilligan,H. M., O’Laughlin, T., Koh, J. S., Levine, J. S. (1996) Antiphospholipidautoantibodies bind to apoptotic, but not nonapoptotic, thymocytes in a�2-glycoprotein I-dependent manner. J. Immunol. 157, 2201–2208.
51. D’Agnillo, P., Levine, J. S., Subang, R., Rauch, J. (2003) Prothrombinbinds to the surface of apoptotic, but not viable, cells and serves as a targetof lupus anticoagulant autoantibodies. J. Immunol. 170, 3408–3422.
52. Ohlsson, M., Jonsson, R., Brokstad, K. A. (2002) Subcellular redistribu-tion and surface exposure of the Ro52, Ro60 and La48 autoantigensduring apoptosis in human ductal epithelial cells: a possible mechanismin the pathogenesis of Sjogren’s syndrome. Scand. J. Immunol. 56,456–469.
53. Gilligan, H. M., Bredy, B., Brady, H. M., Hebert, M-J., Slayter, H. S., Xu,Y., Rauch, J., Shia, M. A., Koh, J. S., Levine, J. S. (1996) Antineutrophilcytoplasmic autoantibodies interact with primary granule constituents onthe surface of apoptotic neutrophils in the absence of neutrophil priming.J. Exp. Med. 184, 2231–2241.
54. Levine, J. S., Subang, R., Koh, J. S., Rauch, J. (1998) Induction ofanti-phospholipid antibodies by �2-glycoprotein I bound to apoptoticthymocytes. J. Autoimmun. 11, 413–424.
55. Mevorach, D., Zhou, J. L., Song, X., Elkon, K. B. (1998) Systemicexposure to irradiated apoptotic cells induces autoantibody production. J.Exp. Med. 188, 387–392.
14 Journal of Leukocyte Biology Volume 76, November 2004 http://www.jleukbio.org
Copyright © 2022 FDOKUMEN




















