
Cytoskeletal pattern changes during branch formation in a centrifuged Adiantum protonema
6
J. Plant Res. 111: 53-58, 1998 Journal of Plant Research by The Botanical Society of Japan 1998 C oskeletal Pattern Changes during Branch Formation in a Centrifuged Adiantum Protonema Masamitsu Wada, Kazunari Nozue and Akeo Kadota Biology.Department, Faculty of Science, Tokyo Metropofitan University, Minami Osawa, Hachioji, Tokyo, 192-03 Japan A protonemal branch was induced on a side wall of a fern filamentous protonema by cell centrifugation and subse- quent polarized-red light irradiation as described in a previ- ous paper (Wada 1995, J. Plant Res. 108: 501-509). Changes in microtubule (MT) and microfUament (MF) pat- terns during the branch development were observed under fluorescence microscopy. A ring-like band of cortical MTs (MT-ring) and MFs similar to a preprophase band or a subapical ring structure (Murata et al. 1987) appeared transiently at the future branching site before cell swelling, the first visible step of branch formation. At this stage, the nucleus was located far from the branching site and the MT-ring appeared to be connected to the nucleus by endoplasmic MFs as well as with endoplasmic MTs. The MT-ring disappeared when cell wall swelling occurred. When the cell wall swelling began, a fan-like pattern of cortical MTs emanating from the new growing tip was established and the MTs reached the opposite flank of the protonema. When a new branch started to elongate and the nucleus moved into the branch, a faint subapical ring of MTs appeared at the subapical part of the new branch, Strands of MTs and MFs emanating from the nuclear front end reached a part of the subapical ring. Key words Adiantum m Branching-- Cytoskeleton m Microfilament m Microtubule-- Protonema (fern) Tip growth of filamentous cells, such as root hairs and pollen tubes, is one of the typical growth patterns in plant cells. Overall growth is another type of growth which occurs in most higher plant cells. These two growth pat- terns look very different from each other. Their basic growth mechanisms may however, be shared at various points, such as the involvement of MTs, rossetts (cellulose synthesizing complexes), turgor pressure, cell wall loosening. In a tip-growing cell, the growing region is very much restricted compared to that of overall growth, suggesting that machinery for cell growth is concentrated in a small region, so that we can focus our attention only on the tip region to study the mechanism of cell growth. In this respect, tip growing cells are a good model system for studying plant cell Abbreviations: MT, microtubule; MF, microfilament; DAPI, 4', 6, diamidino-2-phenylindole growth. The cytoskeleton is thought to play an important role in tip growing cells (Wada and Murata 1991) as well as in overall growing cells (Shibaoka 1994), although the mechanisms are not well understood. In tip growing cells, patterns of MTs and/or MFs have been observed in various organisms by immunofluorescence microscopy, for example, in root hairs (Lloyd 1983), pollen tubes (Joos et al. 1994, Terasaka and Niitsu 1994), fern and moss protonemata as well as their rhizoids (Murata et a/. 1987, Murata and Wada 1989b, Kadota and Wada 1989, Doonan 1991), or by electron microscopy in moss caulonemata (Schmiedel and Schnepf 1980), fern protonemata (Stetler and DeMaggio 1972, Wada and O'Brien 1975, Kiss et al. 1995) and Nicotiana pollen (Lancelle et al. 1987). To study the roles of the cytoskeleton, and espe- cially of MTs, however, their distribution should be observed carefully and categories in which they are included, cortical or endoplasmic, be considered because the function of MTs clearly differ depending upon their localization. In fern protonemata, a band of cortical MTs (subapical MT-ring) which forms at the subapical part of growing protonemata (Murata et al. 1987) may play a role in control- ling the orientation of cellulose microfibrils and consequently, the diameter of the protonemata (Murata and Wada 1989b). However such structure have not been found in rhizoids of the same species (Murata et al. 1987) as in the other tip growing cells so far observed. Similar pattern of MTs have only been found in fertilized Arabidopsis eggs (Webb and Gunning 1991). At the growing tip of these cells including fern protonema and rhizoid cells, cortical MTs either show a random pattern or are not localized there, although in the non-growing zone they run roughly parallel to the cell axis. The reason why, among other tip-growing cells, only fern protonemata have subapical MT-rings is unkown. Endoplasmic MTs, on the other hand, are often observed emanating from one or both ends of a spindle-shaped interphase nucleus in tip growing cells (Wada and O'Brien 1975, Mineyuki and Furuya 1986, Kadota and Wada 1995). They may play a role in nuclear migration or nuclear anchor- age (Schmiedel and Schneph 1979, Vogelmann et al. 1981, Mineyuki and Furuya 1986, Llyod et al. 1987, Kadota and Wada 1995) as shown by experiments using cytoskeletal inhibitors, colchicine and cytochalasin B. Doonan et al. (1986) showed that MTs connect the migrating nucleus to the division site during side branch formation.
-
Upload
independent -
Category
Documents
-
view
8 -
download
0
Transcript of Cytoskeletal pattern changes during branch formation in a centrifuged Adiantum protonema

J. Plant Res. 111: 53-58, 1998 Journal of Plant Research by The Botanical Society of Japan 1998
C oskeletal Pattern Changes during Branch Formation in a Centrifuged Adiantum Protonema
Masamitsu Wada, Kazunari Nozue and Akeo Kadota
Biology.Department, Faculty of Science, Tokyo Metropofitan University, Minami Osawa, Hachioji, Tokyo, 192-03 Japan
A protonemal branch was induced on a side wall of a fern filamentous protonema by cell centrifugation and subse- quent polarized-red light irradiation as described in a previ- ous paper (Wada 1995, J. Plant Res. 108: 501-509). Changes in microtubule (MT) and microfUament (MF) pat- terns during the branch development were observed under fluorescence microscopy. A ring-like band of cortical MTs (MT-ring) and MFs similar to a preprophase band or a subapical ring structure (Murata e t a l . 1987) appeared transiently at the future branching site before cell swelling, the first visible step of branch formation. At this stage, the nucleus was located far from the branching site and the MT-ring appeared to be connected to the nucleus by endoplasmic MFs as well as with endoplasmic MTs. The MT-ring disappeared when cell wall swelling occurred. When the cell wall swelling began, a fan-like pattern of cortical MTs emanating from the new growing tip was established and the MTs reached the opposite flank of the protonema. When a new branch started to elongate and the nucleus moved into the branch, a faint subapical ring of MTs appeared at the subapical part of the new branch, Strands of MTs and MFs emanating from the nuclear front end reached a part of the subapical ring.
Key words A d i a n t u m m Branching-- Cytoskeleton m Microfilament m Microtubule-- Protonema (fern)
Tip growth of filamentous cells, such as root hairs and pollen tubes, is one of the typical growth patterns in plant cells. Overall growth is another type of growth which occurs in most higher plant cells. These two growth pat- terns look very different from each other. Their basic growth mechanisms may however, be shared at various points, such as the involvement of MTs, rossetts (cellulose synthesizing complexes), turgor pressure, cell wall loosening. In a tip-growing cell, the growing region is very much restricted compared to that of overall growth, suggesting that machinery for cell growth is concentrated in a small region, so that we can focus our attention only on the tip region to study the mechanism of cell growth. In this respect, tip growing cells are a good model system for studying plant cell
Abbreviations: MT, microtubule; MF, microfilament; DAPI, 4', 6, diamidino-2- phenylindole
growth. The cytoskeleton is thought to play an important role in tip
growing cells (Wada and Murata 1991) as well as in overall growing cells (Shibaoka 1994), although the mechanisms are not well understood. In tip growing cells, patterns of MTs and/or MFs have been observed in various organisms by immunofluorescence microscopy, for example, in root hairs (Lloyd 1983), pollen tubes (Joos et al. 1994, Terasaka and Niitsu 1994), fern and moss protonemata as well as their rhizoids (Murata et a/. 1987, Murata and Wada 1989b, Kadota and Wada 1989, Doonan 1991), or by electron microscopy in moss caulonemata (Schmiedel and Schnepf 1980), fern protonemata (Stetler and DeMaggio 1972, Wada and O'Brien 1975, Kiss et al. 1995) and Nicotiana pollen (Lancelle et al. 1987). To study the roles of the cytoskeleton, and espe- cially of MTs, however, their distribution should be observed carefully and categories in which they are included, cortical or endoplasmic, be considered because the function of MTs clearly differ depending upon their localization.
In fern protonemata, a band of cortical MTs (subapical MT-ring) which forms at the subapical part of growing protonemata (Murata et al. 1987) may play a role in control- ling the orientation of cellulose microfibrils and consequently, the diameter of the protonemata (Murata and Wada 1989b). However such structure have not been found in rhizoids of the same species (Murata et al. 1987) as in the other tip growing cells so far observed. Similar pattern of MTs have only been found in fertilized Arabidopsis eggs (Webb and Gunning 1991). At the growing tip of these cells including fern protonema and rhizoid cells, cortical MTs either show a random pattern or are not localized there, although in the non-growing zone they run roughly parallel to the cell axis. The reason why, among other tip-growing cells, only fern protonemata have subapical MT-rings is unkown.
Endoplasmic MTs, on the other hand, are often observed emanating from one or both ends of a spindle-shaped interphase nucleus in tip growing cells (Wada and O'Brien 1975, Mineyuki and Furuya 1986, Kadota and Wada 1995). They may play a role in nuclear migration or nuclear anchor- age (Schmiedel and Schneph 1979, Vogelmann et al. 1981, Mineyuki and Furuya 1986, Llyod et al. 1987, Kadota and Wada 1995) as shown by experiments using cytoskeletal inhibitors, colchicine and cytochalasin B. Doonan et al. (1986) showed that MTs connect the migrating nucleus to the division site during side branch formation.

54 M. Wada et aL
Even if the structure of a growing cell tip is studied very carefully, it may not be possible to understand cell tip and/ or determine the mechanism of tip growth. We recently established a model system for cell branching studies using fern protonemata (Wada 1995) in order to analyze the devel- opment of a new apex, and to understand the characteristics of a growing tip. Studying successive changes in intracel- lular structures such as the cytoskeleton during formation of a new branch on the side wall of a mother protonemal cell may reveal fundamental processes in branch formation, that is, in the development of a cell tip, and also reveal the roles of each cytoskeletal array in development. The site and direction of branch differentiation is easily predicted in Adiantum protonemata when induced as described in our previous paper (Wada 1995). In contrast to the moss Funaria (Schmiedel and Schnepf 1979) and the fern Pteris (Ootaki 1965), nuclear location in Adiantum protonemata does not strictly correlate with the branching site (Wada 1995), indicat- ing that this system is also useful for studying how the nucleus controls the site of branch formation. Here, we report the changes in MT and MF patterns which occur during branch formation in Adiantum.
Matedal and Methods
Plant materials Plant materials, culture and centrifugation methods, and
light conditions were the same as described previously (Wada 1995, Fig. 1). Briefly, protonemata of Adiantum capil- lus-veneris which were grown under continous red light (0.5 Wm 2) for 7 days were irradiated with white light (4 Wm -2) for 7 hr to induce cell division, and then centrifuged basipetally for 20 rain at 2000Xg in the dark. They were cultured for 2 days in the dark and then under continuous polarized red light irradiated vertically with an electric vector vibrating parallel to the cell axis. After various period of time under polarized red light, the cells were fixed, stained and then observed. A branch could also be induced in the protonemata without dark incubation before polarized red light treatment. However, since the cytoskeletal patterns were quite similar after both treatments with or without dark incubation, and moreover, the starting point of branch induc- tion easily regulated in the dark-treated cells by transferring them from darkness to red light, the dark-treated protonemata were used in this work unless otherwise stated.
Fluorescence microscopy Immunofluorescence microscopy for MT observation was
the same as described by Murata and Wada (1989a, 1989b) except for the method of cell fixation. Protonemal cells were fixed with 3.5% formaldehyde in piperazine-N,N'-bis(2- ethane-sulfonic acid) (PIPES) buffer at room temperature. The protonemal cells were cut near the tip of the apical cell or at the basal part of the basal cell with a small piece of a razor blade to allow antibodies t o enter the cells. After washing with phosphate-buffered saline the cells were treated with anti-chick ~ - and/%tubulin antibodies followed by a fluorescein-linked second antibody. Staining of MFs
with rhodamine phalloidin was performed as described elsewhere (Kadota and Wada 1995). Briefly, cells were prefixed with m-maleimidobenzoic acid N-hydroxysuc- cinimide ester and then post-fixed with 1.2% formaldehyde in PIPES buffer for 30 min each. After washing with PIPES buffer, the cells were stained with rhodamine-phalloidin at 4 C overnight and then observed. Specimens were observed under a conventional epifluorescence microscope (Axioplan; Carl Zeiss Co. Ltd., Germany) or a confocal laser scanning microscope (LSM410, Carl Zeiss; Kadota and Wada 1995).
R ~ u ~
Successive changes in MT patterns were observed during branch development in long apical cells. During interphase, cortical MTs formed a parallel array with the long cell axis in the dark (Fig. 2a) as well as under polarized red light (data not shown). About 10 hr after transfer from darkness to polar- ized red light, when the nucleus may migrate towards the branch site near the septum, a ring-like pattern of cortical MTs (MT-ring) became obvious at the future branching site near septum in the long apical cells (Fig. 3a), although it appeared only transiently (Fig. 7). As was evident in Figs. 4 and 7 in Wada (1995), nuclear migration cannot be detected in living cells because of the difficulty in pinpointing the nucleus within the dense array of chloroplasts and other organelles. The MT-ring was about 5/~m in width on average, and appeared about 30/~m away from the septum (Fig. 8). The MT-ring also appeared near the septum of the basal cell. Double MT-rings were sometimes observed in mirror image on both sides of the septum, although this rarely occurred (data not shown).
The timing of MT-ring formation was studied in detail. The MT-ring could be observed only for a short period around 9 hr after the onset of the polarized red light irradia- tion, obviously before branch development (Fig. 7). The transient appearance of the MT-ring was also observed in the protonemata cultured under continuous polarized red light without dark treatment after cell centrifugation (data not shown).
When a part of a cell wall swelled prior to branching, a fan-like pattern of fine cortical MTs was visible spreading out from the broad branch apex when viewed from the side (Fig. 4a). When this fan-like pattern was observed from the top (Fig. 10), a parallel array of MTs was seen running perpendicular to the cell axis at the top of the banch, but the MTs spread out gradually towards the branch base. When a new branch reached more than 10-15~m in length a subapical ring of cortical MTs (Murata et al. 1987)also appeared (Fig. 5a).
Endoplasmic MTs could also be observed during branch formation. MTs emanating from the nuclear tip seemed to connect the nucleus and the MT-ring (data not shown). When a subapical ring of a new branch appeared, MTs connecting the subapical ring and the nucleus were also detected, although the MTs around the new branch were so dense and complicated it was hard to see the connection clearly (Fig. 5a). These endoplasmic MTs emanating from
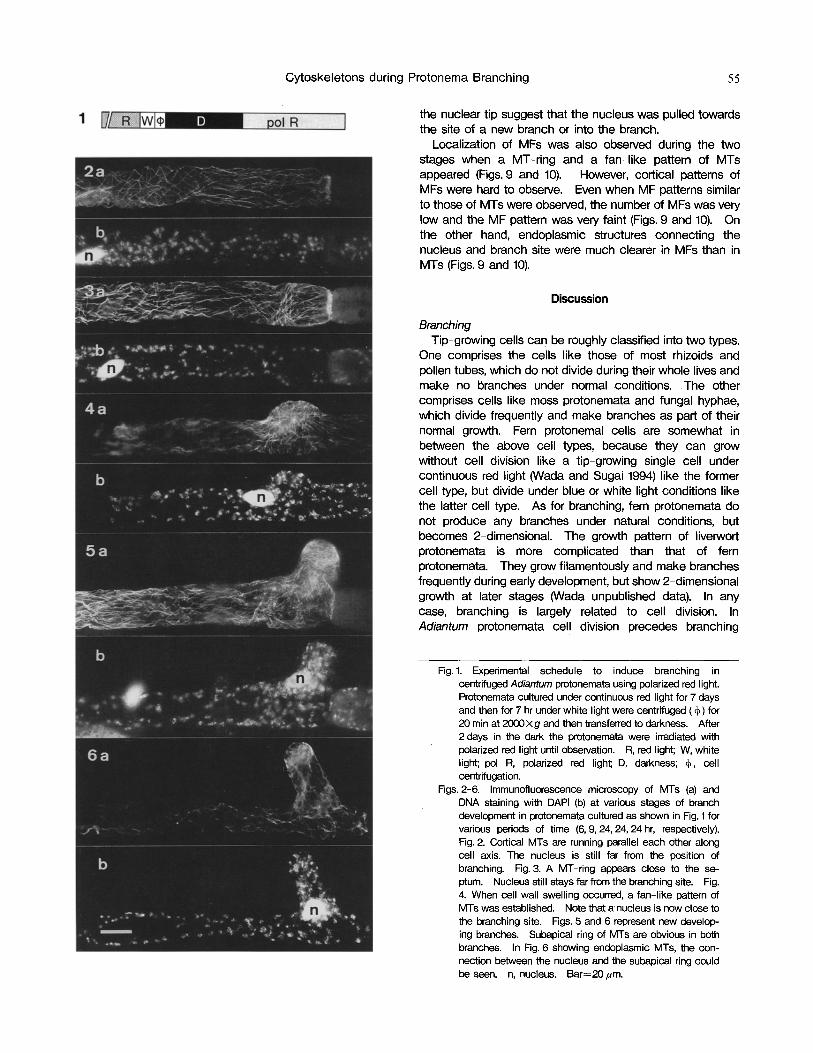
Cytoskeletons during Protonema Branching 55
the nuclear tip suggest that the nucleus was pulled towards the site of a new branch or into the branch.
Localization of MFs was also observed during the two stages when a MT-r ing and a fan- l ike pattern of MTs appeared (Figs. 9 and 10). However, cortical patterns of MFs were hard to observe. Even when MF patterns similar to those of MTs were observed, the number of MFs was very low and the MF pattern was very faint (Figs. 9 and 10). On the other hand, endoplasmic structures connect ing the nucleus and branch site were much clearer in MFs than in MTs (Figs. 9 and 10).
Discussion
Branching Tip growing cells can be roughly classified into two types.
One comprises the cells like those of most rhizoids and pollen tubes, which do not divide during their whole lives and make no branches under normal conditions. The other comprises cells like moss protonemata and fungal hyphae, which divide frequently and make branches as part of their normal growth. Fern protonemal cells are somewhat in between the above cell types, because they can grow without cell division like a t ip-growing single cell under continuous red light (Wada and Sugai 1994) like the former cell type, but divide under blue or white light conditions like the latter cell type. As for branching, fern protonemata do not produce any branches under natural conditions, but becomes 2-dimensional. The growth pattern of liverwort protonemata is more complicated than that of fern protonemata. They grow fi lamentously and make branches frequently during early development, but show 2 dimensional growth at later stages (Wada unpublished data). In any case, branching is largely related to cell division. In Adiantum protonemata cell division precedes branching
Fig. 1. Experimental schedule to induce branching in centrifuged Adiantum protonemata using polarized red light. Protonemata cultured under continuous red light for 7 days and then for 7 hr under white light were centrifuged (~) for 20 min at 2000• and then transferred to darkness. After 2 days in the dark the protonemata were irradiated with polarized red light until observation. R, red light; W, white light; pol R, polarized red light;, D, darkness; ~, cell centrifugation.
Figs. 2-6, Immunofluorescence microscopy of MTs (a) and DNA staining with DAPI (b) at various stages of branch development in protonemata cultured as shown in Fig. 1 for various periods of time (6, 9, 24, 24, 24 hr, respectively). Fig. 2. Cortical MTs are running parallel each other along cell axis. The nucleus is still far from the position of branching. Fig. 3. A MT-ring appears close to the se- ptum. Nucleus still stays far from the branching site. Fig. 4. When cell wall swelling occurred, a fan-like pattern of MTs was established. Note that a nucleus is now close to the branching site. Figs. 5 and 6 represent new develop- ing branches. Subapical ring of MTs are obvious in both branches. In Fig. 6 showing endoplasmic MTs, the con- nection between the nucleus and the subapical ring could be seen. n, nucleus. Bar=20/~m.

56 M. Wada et al.
60 120
40 A
__=
o 20
0 0 10 20 30 40 50
Time after pol R irradiation (hr)
Fig. 7. Time course of MT-ring appearance at an early stage of branch development. Culture conditions are the same as in Fig. 1. Note that MT-ring (closed circles) appeared only transiently and disappeared before branch formation (open circles). Each point was obtained from 70 to 200 protonemata.
(Wada 1995) Branching of fern protonemata differs significantly from
that of moss protonemata, as discussed above. Branching is an abnormal phenomenon in fern protonemata, despite their ability to branch, but is normal in moss protonemata. Thus, branching is spontaneous developmental step in mosses but is an unexpected event which may occur under unfavorable conditions in fern protonemata. In this respect, branching of fern protonema may be a better model system than moss for studying the mechanism of branch differentia- tion, because fern branching can be induced artificially by changing environmental factors, and can be observed from a very early stage. Branching in fern protonemata is, how- ever, preceeded by several steps which are not related directly to branch formation. These include cessation of growth at the protonemal apex, branching site determination, destruction of the structure of the side wall, etc., and meaning that steps in branch development should be distin- guished carefully from the preceeding phenomena abandon-
100
8 0 E E
60
-~ 40 (
20
J
0 I I I I I
6 7 8 9 10 11 12 Time after pol R irradiation (hr)
Fig. 8. Average width of MT-rings (closed circles) which appeared at different timing after the irradiation with polar- ized red light is shown. The distance between the septum and the center of the MT-ring (open circles) and the distance between the septum and the nucleus (open squares) were also measured in each MT-ring and the averages were plotted. The abscissa is the timing of each MT-ring appearance. Note that the site and width of the MT ring do not change even when the timing of MT-ring appearance is different, although the distance between the septum and the nucleus changes gradually. In other words, in a cell with a nucleus far from the septum, MT-ring appearance delayed. Each data shows the mean _+ standard error.
ing the former apex.
MT-rings Two different kinds of rings of MT bands in fern
protonemata have been reported. One is a preprophase band (Murata and Wada 1989a) observed during (pre-) prophase and the other is a subapical ring (Murata et al. 1987) formed during cell elongation. The MT-r ing identified at the future branching site in the present study might be a new type of MT band having a different function. Although these rings show a similar pattern in protonemal cells (cor- tical MTs running perpendicular to the growing axis), their
Fig. 9. Stereo microphotographs of laser scanning images of MFs in pert of a protonema which was cultured for 27 hr underpolarized red light, after 2 days dark incubation. Optical sections were made every 0.5 ,urn in the Z axis through the cells and stereographs were obtained by Zeiss software. The picture of a long protonemal cell was divided into 2 pieces, including a septum (arrowhead) and a nucleus (arrow). Bar----lO,~m.

Cytoskeletons during Protonema Branching 57
Fig. 10. Stereo microphotographs of laser scanning image of MTs (left) and MFs (right) during early branch development after 33 hr culture under polarized red light. The branch grows up (i.e. towards us) although a part of the branch crashed down during preparation. The MT pattem spreading from the new branch is obvious. An MF bundle connecting the nucleus and the branching site is also clear. Optical methods are the same as in Fig. 9. n, nucleus. Bar=10.um.
function must be different from others judging by the timing and the intracellular localization of ring appearance, and from their relationship with distinct phenomena. The pre- prophase band may play a role in deciding the fusion site of a new cell wall, although the precise mechanism for this is unknown. The subapical ring is predicted to decide protonemal width by controlling the running direction of cellulose microfibrils (Murata and Wada 1989b) and/or to anchor the nucleus in a specific position by connecting it and the ring with MTs (Kadota and Wada 1995). The func- tion of the MT-ring reported in this work is not yet known but it may differ from those of the two described above. We do not know the molecular structure of these MT rings, but it is possible that each MT band has different MT-associated proteins conferring different functions and stabilities. Actu- ally, the subapical ring is easily destroyed by various factors, such as centrifugation (Murata personal communication) and transfer to darkness (Murata and Wada 1989a) or to blue light (Murata and Wada 1989b), but under the same conditions the MTs of the preprophase band are not depolymerized by centrifugation (Murata and Wada 1992).
The newly formed MT-ring is the second example of transient MT structure predicting the site of a physiological phenomenon. The first case was the preprophase bands found in 1966 by Pickett-Heaps and Northcote (1966a, 1966b). The preprophase band and MT-ring found in this work appeared transiently and predicted the sites of cell division and branch formation, respectively, disappearing before these phenomena occurred. Such MT-bands are thought to leave a message at the site where they de- polymerize. Branching and cell" division are distinct phe- nomena, but in both cases a nucleus moves to the former location of the MT rings. If it is the case, similar mecha- nisms might possibly be involved in additional phenomena such as nuclear anchoring.
This work was supported in part by a Grant-in Aid from the Ministry of Education, Science, Sports and Culture in
Japan (05454023) and by a grant from the Mitsubishi Foun- dation to M.W. We are grateful to Dr. Jane Silverthorne of the University of California, Santa Cruz, for critical reading of the manuscript.
References
Doonan, J.H. 1991. The cytoskeleton and moss mor- phogenesis. In C.W. Lloyd, ed., The Cytoskeletal Bases of Plant Growth and Form. Academic Press, London. pp. 289-301.
Doonan, J.H., Jenkins, G.I., Cove, D.J. and Lloyd, C.W. 1986. Microtubule connect the migrating nucleus to the pro- spective division site during side branch formation in the moss, Physcomitrella patens. Eur. J. Cell Biol. 41: 157- 164.
Joos, U., van Aken, J. and Kristen, U. 1994. Microtubules are involved in maintaining the cellular polarity in pollen tubes of Nicotiana sylvestris. Protoplasma 179: 5-15.
Kadota, A. and Wada, M. 1989. Circular arrangement of cortical F actin around the subapical region of a tip- growing fern protonemal cell. Plant Cell Physiol. 30: 1183-1186.
Kadota, A. and Wada, M. 1995. Cytoskeletal aspects of nuclear migration during tip-growth in the fern Adiantum protonemal cell. Protoplasma 188: 170-179.
Kiss, J.Z., Wang Cahill, F. and Kiss, H.G. 1995. The ultra- structure of the early development of Schizaea pusilla gametophytes. Inter. J. Plant Sci. 156: 131-142.
Lance,e, S.A., CresU, M. and Hepler, P.K. 1987. Ultra- structure of the cytoskeleton in freeze-substituted pollen tubes of Nicotiana alata. Protoplasma 140: 141- 150.
Lloyd, C.W. 1983. Helical microtubular arrays in onion root hairs. Nature 305: 311-313.
Lloyd, C.W., Pearce, K.J., Raulins, D.J., Ridge, R.W. and Shaw, P.J. 1987. Endoplasmic microtubules connect the advancing nucleus to the tip of legume root hairs, but F-actin is involved in basipetal migration. Cell Motil. Cytoskel. 8: 17-36.

58 M. Wada et aL
Mineyuki, Y. and Furuya, M. 1986. Involvement of col- chicine-sensitive cytoplasmic elen~ent in premitotic nuclear positioning of Adiantum protonemata. Proto- plasma 130: 83-90.
Murata, T., Kadota, A., Hogetsu, T. and Wada, M. 1987. Circular arrangement of cortical microtubules around the subapical part of a tip-growing fern protonema. Protoplasma 141: 135-138.
Murata, T. and Wada, M, 1989a. Re-organization of mi- crotubules during preprophase band development in Adiantum protonemata. Protoplasma 151: 73-80.
Murata, T. and Wada, M. 1989b. Organization of cortical microtubules and microfibril deposition in reponse to blue-light induced apical swelling in a tip-growing Adiantum protonemal cell. Planta 178: 334-341.
Murata, T. and Wada, M. 1992. Cell cycle specific'disrup- tion of the preprophase band of microtubules in fern protonemata: Effects of displacement of the endoplasm by centrifugation. J. Cell Sci. 101: 93-98.
Ootaki, T. 1965. Branching in fern gametophyte, Pteris vittata, under various light conditions. Cytologia 30: 182-193.
Picket-Heaps, J.D. and Northcote, D.H. 1966a. Organiza- tion of microtubules and endoplasmic reticulum during mitosis and cytokinesis in wheat meristems. J. Cell. Sci. 1:109 120.
Picket-Heaps, J.D. and Northcote, D.H. 1966b. Cell divi- sion in the formation of the stomatal complex of the young leaves of wheat. J. Cell. Sci. 1: 121- 128.
Schmiedel, G. and Schnepf, E. 1979. Side branch forma- tion and orientation in the caulonema of the moss, Funaria hygrometrica: Normal development and fine structure. Protoplasma 100: 357-383.
Schmiedel, G. and Schnepf, E. 1980. Polarity and growth of caulonema tip cells of the moss Funaria hygrometric, a. Planta 147: 405-413.
Shibaoka, H. 1994. Plant hormone-induced changes in the orientation of cortical microtubules: Alterations in the
cross linking between microtubules and the plasma membrane. Annu. Rev. Plant Physiol. Plant Mol. Biol. 45: 527-544.
Stetler, B.A. and DeMaggio, A.E. 1972. An ultrastructural study of fern gametophytes during one- to two-dimen- sional development. Amer. J. Bot. 59: 1011-1017.
Terasaka, O. and Niitsu, T. 1994. Differential roles of microtubule and actin myosin cytoskeleton in the growth of Pinus pollen tubes. Sex Plant Reprod. 7: 264-272.
Vogelmann, T.C., Bassel, A.R. and Miller, J.H. 1981. Effects of micrOtubule inhibitors on nuclear migration and rhizoid differentiation in germinating fern spores (Ono- c/ea sensibilis). Protoplasma 109: 295-316.
Wada, M. 1995. Nuclear behavior during branch formation in a centrifuged Adiantum protonema and the nuclear polarity. J. Plant Reser. 108: 501-509.
Wada, M. and Murata, T. 1991. The cytoskeletons in fern protonemal growth in relation to photomorphogenesis. In C.W. Lloyd, ed., The Cytoskeletal Basis of Plant Growth and Form. Academic Press, London. pp. 277- 288.
Wada, M. and O'Brien, T.P. 1975. Observations on the structure of the protonema of Adiantum capillus-veneris L. undergoing cell division following white-light irradia- tion. Planta 126: 213-237.
Wada, M. and Sugai, M. 1994. Photobiology of ferns. In R.E. Kendrick and G.H.M. Kronenberg, eds., Photomor- phogenesis in Plants. Kluwer Academic Publishers, Dordrecht. pp. 783-802.
Webb, M.C. and Gunning, B.E.S. 1991. The microtubular cytoskeleton during development of the zygote, proem- bryo and free-nuclear endosperm in Arabidopsis thaliana (L.) Heynh. Planta 184: 187-195.
(Received September 24, 1997" Accepted December 12, 1997)




















