
Comparison of Two Different Methods for Inoculating VX2 Tumors in Rabbit Livers and Hind Limbs
Upload
independentCategory
view
2download
0
Marine Pollution Bulletin 62 (2011) 1652–1660
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Lysosomal and lipid-associated parameters in the livers of three speciesof arctic seabird chicks: Species differences and relationshipswith contaminant levels
Marit Nøst Hegseth a,b,⇑, Francesco Regoli c, Stefania Gorbi c, Raffaella Bocchetti c, Geir Wing Gabrielsen d,Lionel Camus a
a Akvaplan-NIVA AS, Fram Centre – High North Research Centre for Climate and the Environment, 9296 Tromsø, Norwayb University of Tromsø, Brevika, 9037 Tromsø, Norwayc Universita Polytechnica delle Marche, Facolta di Scienze, Via Brecce Bianche, 60131 Ancona, Italyd Norwegian Polar Institute, Fram Centre – High North Research Centre for Climate and the Environment, 9296 Tromsø, Norway
a b s t r a c t
0025-326X/$ - see front matter � 2011 Elsevier Ltd.doi:10.1016/j.marpolbul.2011.06.011
⇑ Corresponding author at: Akvaplan-NIVA AS,Research Centre for Climate and the Environment,+47 97060492.
E-mail address: [email protected] (M.N. Heg
Lysosomal membrane stability, lipofuscin (LF), malondialdehyde (MDA), neutral lipid (NL) levels, as wellas halogenated organic compounds (HOCs), Cr, Cd, Pb and Fe concentrations were analyzed in liver ofblack-legged kittiwake (BK), herring gull (HG), and northern fulmar (NF) chicks. There were significantspecies differences in the levels of NL, LF and lysosomal membrane stability. These parameters werenot associated with the respective HOC concentrations. LF accumulation was associated with increasingCr, Cd and Pb concentrations. HG presented the lowest lysosomal membrane stability and the highest.
LF and NL levels, which indicated impaired lysosomes in HG compared to NF and BK. Lipid peroxidationwas associated with HOC and Fe2+ levels. Specific HOCs showed positive and significant correlations withMDA levels in HG. The study indicates that contaminant exposure can affect lysosomal and lipid associ-ated parameters in seabird chicks even at low exposure levels. These parameters may be suitable markersof contaminant induced stress in arctic seabirds.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Lysosomal parameters are regularly used as biomarkers of var-ious types of environmental stress in marine animals, such as mol-luscs or fish (Köhler et al., 2002; Moore et al., 2007; Fang et al.,2010). Nutrient restriction, seasonal variations and xenobioticexposure are known to affect lysosomal activities, and early warn-ing signals of cellular pathology can be detected through analysisof these parameters (Moore et al., 2008). During the last three dec-ades, the long range transport of contaminants and the subsequentexposure of Arctic biota to these contaminants have gainedincreasing attention (de Wit et al., 2004; Gabrielsen and Sydnes,2009). In this regard, halogenated organic compounds (HOCs)and heavy metals have been of particular interest, and elevatedlevels of these compounds have been reported in both mammalianand avian top predators (Borga et al., 2001; de Wit et al., 2004;Gabrielsen and Sydnes, 2009). However, to our knowledge, lyso-somal parameters have not previously been applied as tools in
All rights reserved.
Fram Centre – High North9296 Tromsø, Norway. Tel.:
seth).
the context of environmental monitoring in arctic seabirds ormammals.
Lysosomes are intracellular membrane-bound organelles with alow luminal pH that are present in all animal cells. These organ-elles are important parts of the autophagic process, and when nec-essary, cellular components are degraded and recycled in thelysosomes (de Duve, 2005; Luzio et al., 2007). However, any factorthat affects the enzymatic processes or structures in the lysosomemay disturb the integrity of the organelle and thereby preventoptimal lysosomal functioning (Moore et al., 2006; Eskelinen andSaftig, 2009). Foreign substances, such as environmental contami-nants, may affect and harm the lysosomes. Such xenobiotics aretransported through the endocytic pathway to the lysosomeswhere they are stored (Köhler et al., 2002; Moore et al., 2008). Inthis sense, the lysosomes protect the rest of the cell from the po-tential harmful effects of these substances, but at the same time,the presence of harmful substances in the lysosomes poses a threatto the structures and mechanisms occurring within this organelleand may eventually impair autophagic actions in general, whichmay have deleterious effects for the rest of the cell (Moore et al.,2006; Eskelinen and Saftig, 2009). One of the parameters that havebeen found to be sensitive to xenobiotic exposure is the stability ofthe lysosomal membrane (Regoli, 1992; Bocchetti and Regoli,2006; Moore et al., 2008). Through several mechanisms, such as

M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660 1653
the oxidation of membrane lipids, xenobiotics may increase per-meability and destabilise the lysosomal membrane, which caneventually disturb the functioning of this organelle (Hwang et al.,2002; Terman et al., 2006).
Another lysosome-related parameter that has previously beenstudied in the context of contaminant exposure is the presenceand levels of neutral lipids (NLs) (Moore, 1988; Lowe, 1988; Regoli,1992; Köhler et al., 2002; Nesto et al., 2007). In most eukaryoticcells, lipids are stored in specific organelles as NLs, which are oftenreferred as lipid droplets (Murphy, 2001). Hepatic cells containlarge amounts of lipid droplets, which play an important role in en-ergy homeostasis. Dietary changes or other stress factors havebeen shown to mobilise lipids from lipid droplets (Domouhtsidouand Dimitriadis, 2001; Bocchetti and Regoli, 2006; Walther andFarese, 2009), but observations of stress-induced increases of NLlevels have also been reported (Moore, 1988; Köhler et al., 2002).In the latter studies, contaminant-induced lipidosis-like conditionscharacterised by increased levels of lysosome-associated NLs havebeen shown in bivalves and fish sampled along contaminant gradi-ents in different marine habitats in northern Europe. NL levels havethus proven to be a responsive parameter related to contaminantexposure in these organisms under field conditions.
The recycling processes within the lysosome are not perfect anddo not completely degrade the phagocytosed materials (Termanand Brunk, 2004). The incomplete degradation results in the accu-mulation of a non-degradable brown-yellow pigment calledlipofuscin (LF). LF mainly consists of various cell type-specific oxi-dised protein residues and lipid components (Brunk and Terman,2002). The accumulation of LF is suggested to occur through anumber of mechanisms (Terman and Brunk, 2004), and increasedlysosomal activity increases the generation of LF. Any factor thataffects the rate and efficiency of lysosomal activity will thereforeaffect LF generation. Hence, LF levels can be used as a marker ofvarious types of cellular stress, including xenobiotic exposure. Pre-vious studies have found a connection between increased lipofus-cin levels and contaminant exposure, particularly with metals(Regoli, 1992; Moore et al., 2007).
One element involved in lipofuscin generation is the reactionbetween malondialdehyde (MDA) and proteins (Terman andBrunk, 2004). MDA is a by-product of lipid oxidation, which takesplace through the iron-catalysed Fenton reaction and is not specificto lysosomes (Ryan and Aust, 1992). Lipid peroxidation can causedeleterious effects on membranes in the cell (Cheeseman, 1993)including the lysosomal membrane. Lipid peroxidation facilitatedby lipofuscin-associated iron is indeed considered to be one ofthe main mechanisms underlying the impairment of the lysosomalmembrane (Brunk and Terman, 2002). Excess amounts of iron orreactive oxygen species can initiate lipid peroxidation (Halliwelland Gutteridge, 2007), which makes MDA a suitable marker for cel-lular stress caused by exposure to organic contaminants or metals.Lipid peroxidation in relation to contaminants such as brominatedflame retardants (Fernie et al., 2005) and metals (Berglund et al.,2007; Kaminski et al., 2009; Wayland et al., 2010) has previouslybeen examined in birds in which relationships between iron con-centration and MDA levels have been found (Berglund et al.,2007; Kaminski et al., 2009).
The relationships between iron and organic contaminant con-centrations, lipid peroxidation, and stress-induced lysosomalparameters have not previously been studied in arctic seabirds.Considering the potential for contaminant exposure that arctic sea-birds experience, these are relevant biological parameters to inves-tigate and characterise and may provide information about earlycellular effects caused by contaminant-related or metabolic stressin these animals.
To investigate the relationships between contaminant exposureand lysosomal parameters in arctic seabirds, chicks from three of
the most abundant seabird species such as the black-legged kitti-wake, Rissa tridactyla (BK), the herring gull, Larus argentatus (HG),and the northern fulmar, Fulmarus glacialis (NF), were sampled.These three species represent two different phyla, in which BKand HG are members of the family Laridae, and NF belongs tothe Procellariidae family. These species have similar food prefer-ences, but because of slightly dissimilar diet compositions, theyare located at different positions in the trophic hierarchy (Mehlumand Gabrielsen, 1993; Dahl et al., 2003; Kubetzki and Garthe,2003). We investigated NL and lipofuscin levels, lysosomal mem-brane stability and MDA content in liver samples from these threespecies, and the results were related to levels of HOCs, Cr, Cd, Pband Fe2+ in the liver.
The aim of this study was to apply methods previously exten-sively employed in other marine animals to study autophagic lyso-somal parameters also in seabird chicks, and to describe potentialspecies differences between BK, NF and HG chicks with regards tothese important hepatic physiological features. In addition werepossible relationships between the lysosomal parameters and con-taminant levels explored.
2. Materials and methods
2.1. Field samples
Tissue samples were collected during August and September in2006 and September in 2007 from three different seabird species:NF aged 65 days, BK aged 35 days and HG aged 75 days. A total of15 chicks from each species were caught and sampled at two dif-ferent sites in Kongsfjorden on Svalbard (78�550N 11�560E) (NFand BK chicks) or near Sommarøy outside Tromsø, Norway(69� 370 55.0700 N, 18� 10 11.1600 E) (HG chicks). Chicks from thesethree seabird species were chosen over adults to minimise theinfluence of unknown factors, such as previous diet and age. Atthe sampling sites, the chicks were sacrificed by decapitation,and their liver was dissected out. Portions of these tissue sampleswere placed in cryotubes and immediately frozen in liquid nitro-gen and then transported to Tromsø, Norway, where the samplesfor histological and biochemical analysis were stored at �80 �C,and the samples for contaminant analysis were stored at �20 �Cuntil analyses were conducted.
2.2. HOC analysis
Details of the methods of HOC analysis of the present study aregiven by Nøst (2009) and Helgason et al. (2010). In brief, approxi-mately 2 g of liver tissue was homogenized and dried in sodiumsulphate. The homogenate mix was transferred to a glass columnwhere it was extracted with cyclohexane/acetone. An additionalcleanup and fractionation step based on a method by Sandangeret al. (2004) was carried out on a Zymark Rapidtrace AutomatedSPE workstation (Caliper Life Sciences, Inc.) using columns packedwith 1 g florisil. MeSO2- and OH-metabolites were additionallycleaned up on acidic silica columns. Three fractions were collected;fraction 1:7.2 mL of dichloromethane/n-hexane (1/4 v/v) as theeluting solvent containing neutral compounds, fraction 2:9 mL ace-tone/n-hexane (1/10 v/v) containing MeSO2-PCBs/MeSO2-DDE, andfinally fraction 3:15 mL metanol/dichloromethane (1/5 v/v) con-taining the OH-PCBs. The third fraction was evaporated to drynessand derivatized using 1 mL of diazomethane in n-hexane. A finalclean-up was performed on the Rapidtrace using a column of0.7 g of 25% sulfuric acid silica with 0.2 g neutral silica on topand extracted using 8 mL of dichloromethane. Evaporation stepswere performed on a Labconco evaporator (Labconco Corp., KansasCity, MO). The amount of extractable organic material (EOM) was

1654 M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660
determined gravimetrically. All plasma extracts were transferredto GC vials with 150 lL inserts, and isooctane was added beforea gentle evaporation with nitrogen gas to the final volume. Finally,10 lL octa chloro naphtalene was added as recovery standard. Theanalysis of the compounds was performed by high-resolution gaschromatography (HRGC) on an Agilent 7890A gas chromatograph(GC) equipped with an Agilent 7683B automatic injector and anAgilent 5975C mass spectrometer (MS) (Agilent, Folsom, CA). TheGC was fitted with a 30 m DB-5 MS column (5% phenyl-methylpolysiloxane; 0.25 mm i.d., 0.25 mm film thickness) fromJ&W Scientific (CA, USA). 1 lL aliquot of the sample extract wassplitless injected and helium used as a carrier gas at a constantflow of 1.5 mL/min. Temperature programme: initial temperature70 �C (2 min), 15 �C/min to 180 �C, 5 �C/min to 280 �C (5 min).The MS was running in the negative ion chemical ionisation mode(NICI) for metabolites. The MS was operated in single ion monitor-ing mode (SIM). All chemical analyses followed internationalrequirements for quality and control (QA/QC), e.g., recommenda-tions of the Arctic Monitoring and Assessment Programme (AMAP)and the requirements in the European quality norm EN 17049. Foreach batch of 10 liver samples one standard reference (NIST Cod oil1588a) and one blank were prepared. These halogenated organiccompounds were detected in one or more species and were sum-marized to RHOCs as a representative measure of the total organiccontaminant burden: PCB 18, 28, 31, 33, 37, 47, 52, 66, 74, 99, 101,105, 114, 118, 123, 128, 138, 141, 149, 153, 156, 157, 167, 170, 180,183, 187, 189, 194, 206, 209, pesticides: o,p0-DDE, p,p0-DDE,p,p0DDD, p,p0DDT, a-hexachlorocyclohexane (HCH), b-HCH, Penta-chlorobenzene (PeCB), hexachlorobenzene (HCB), pentachlorophe-nol (PCP), Tribromoanisole (TBA), polybrominated diphenyl ether(PBDE)-28, 47, 71/49, 99, 100, 119, 154, hydroxylated PCB andPBDEs: 4OH-PCB 107, 146, 187, 3OH-PCB153, 2-OH-BDE 68, meth-ylsulphone-PCB/DDTs: 3/4MeSO2-PCB 49, 52, 91, 87, 101, 110, 132,141, 149, 174, 3-MeSO2-DDE, 5-Methoxy-BDE 47.
2.3. Metal analysis
The method used for the analyses of metal concentrations wasdescribed in detail previously (Fattorini et al., 2010). Briefly, the li-ver samples were dried (60 �C) until achieving constant weight, fol-lowed by digestion under pressure with nitric acid and H2O2 in amicrowave digestor system (Mars CEM, CEM Corporation, Mat-thews, NC, USA). The metal concentrations were assessed by atomicabsorption spectrophotometry with flame and flameless atomisa-tion (SpectrAA 220FS and SpectrAA-240Z Zeeman, Varian, Mul-grave, VIC, Australia). Quality assurance and quality control wereassessed by processing blank samples and reference standardmaterials (lyophilised mussel tissue, Standard Reference MaterialNIST-2977, National Institute of Standards and Technology,Gaithersburg, MD, USA), and the concentrations measured instandard reference materials were always within the 95% confi-dence interval of the certified values.
2.4. Histological analyses
For histological analyses (lipofuscin, NLs and lysosomal mem-brane stability), pieces of approximately 100 mg of liver tissuewere mounted on a pre-cooled aluminium cryostat chuck with aSakura Finetek Tissue-Tek optimum cutting temperature (OTC)specimen matrix and kept below �30 �C. From each sample, 8and 10 lM sections were cut for the analysis. 10 individuals ofeach species were analyzed for NL and LF, and three individualsper species were analyzed for lysosomal membrane stability.
For determination of LF content, the cryostat sections werefixed for 15 min in Beker’s fixative (+1.8% NaCl) at 4 �C, followedby rinsing in distilled water. The sections were then mounted in
a glycerol–gelatine mounting medium and analysed microscopi-cally (lipofuscin fluorescent spectra: excitation maxima at 340–390 nm, emission maxima at 430–490 nm, using a DAPI filter).Unsaturated NLs were studied using fixation and staining withoil red O following a method described by Bocchetti and Regoli(2006). Quantification of the staining intensity was performed withKodak 1D Image Software and normalised to the area of liver tis-sue, which was measured with Olympus DP software. Determina-tion of lysosomal membrane stability was performed by thecytochemical method described in ICES, 2004 (Moore et al.,2004). The slides were incubated in 0.1 M citrate buffer (pH 4.5)at 37 �C in a shaking water bath for different time intervals (0, 3,5, 10, 15, 20, 30 and 40 min). For visualization of lysosomal acidphosphatase, sections were incubated for 15 min in 3.5% Polypep,dissolved in 0.1 M citrate buffer, pH 4.5, containing 4 mM naph-thol-AS-BIphosphate dissolved in 2% dimethyl sulfoxide (DMSO).The slides were washed twice in 3% NaCl, and an azo-couplingreaction was performed for 9 min in 0.1% Fast Violet B salt dis-solved in 0.1 M phosphate buffer, pH 7.4. Slides were then rapidlyrinsed under running tap water for 5 min, fixed for 10 min in 10%formaldehyde containing 2% calcium acetate, before beingmounted in glycerine gelatine. The determination of lysosomalmembrane stability was based on the time required for acid labili-zation and therefore producing the maximum staining intensity.This was assessed under a light microscope as the maximal accu-mulation of reaction product associated with lysosomes. Fourdeterminations were made for each animal by dividing each sec-tion in the acid labilization sequence into four approximately equalsegments and assessing the labilization period in each of the corre-sponding segments. A mean value was then derived for each sec-tion, corresponding to each individual organism.
2.5. Lipid peroxidation analysis
Liver tissue was homogenised in 500 ll of ice cold 50 mM PBSpH 7.4 per 100 mg of tissue (1:5) with a Potter–Elvehjem homog-eniser and centrifuged at 14000g for 20 min at 4 �C.
Lipid peroxidation was quantified by spectrophotometricallymeasuring the adduct formed between MDA and thiobarbituricacid using a method based on the description of Aust (1985).MDA tetraethyl acetal was used to generate an MDA standardcurve.
2.6. Statistics
The lysosomal and lipid parameter data were tested for normal-ity using the Shapiro–Wilk test. Differences between the investi-gated species were analysed using the nonparametric Wilcoxonrank sum test due to the small sample size. Multiple regressionswere conducted to evaluate the relationships between the lyso-somal and lipid parameters in relation to the contaminant datafor the field samples. Spearman’s rank correlation coefficient wasused to evaluate the relationships between the antioxidant param-eters and the HOC concentrations. The statistical analyses wereconducted using R software, version 2.9.2.
3. Results
3.1. Contaminant levels
3.1.1. HOCsThe hepatic HOC concentration differed significantly between
the species. BK exhibited the lowest total hepatic HOC concentra-tion (25.56 ng/g w.w. ±7.4), NF showed the highest concentration(193.19 ng/g w.w. ±80.6), and HG had a concentration between

Sum HOC concentration, liver
0
100
200
300
Black-legged kittiwake Herring gull Northern fulmar
ng/g
w.w
.
*
**
Fig. 1. Concentrations of RHOCs (ng/g w.w.) in liver samples from wild living blacklegged kittiwake-, herring gull- and northern fulmar chicks from Norwegian arcticregions.
M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660 1655
those of the other two species (71.80 ng HOCs/g w.w. ±51.7)(Fig. 1).
3.1.2. MetalsSignificant species differences in Fe2+, Cd, Cr and Pb levels were
exhibited. The average BK liver Fe2+ concentration (97.7 lg/g d.w)was significantly lower than those of HG (499.3 lg/g d.w) and NF(700.7 lg/g d.w). For graphical description, see Hegseth et al.(2011). For Cd, Cr and Pb, HG presented the highest concentrations(0.526, 0.154 and 0.703 lg/g d.w respectively) compared to NF(0.301, 0.121 and 0.157 lg/g d.w., respectively) and BK, whichshowed the lowest concentrations of these metals (0.207, 0.047,0.126 lg/g d.w., respectively). The standard deviations and p-val-ues for level of significance for the species differences are givenin Table 1, and Cd, Cr and Pb levels are shown in Fig. 2.
A dose–response-like relationship was observed between spe-cies average values for Cd, Cr and Pb concentrations and speciesaverage values for LF levels, such that BK, the species with the low-
Table 1Average concentrations (lg/g d.w.) of iron, cadmium, chromium and lead in the liver ofNorwegian arctic regions. p-Values describe the level of significance of the species differe
Heavy metals (lg/g d.w.) Black-legged kittiwake North
Average Std. dev. Averag
Fe 98.7 23.7 700.7Cd 0.207 0.092 0.301Cr 0.047 0.009 0.121Pb 0.126 0.069 0.157
Heavy m
0.00
0.20
0.40
0.60
0.80
1.00
Black-legged kittiwake Northe
μg/g
d.w
.
Cd
Cr
Pb
Fig. 2. Concentrations of Cd, Cr and Pb (lg/g d.w.) in liver samples from wild living blackregions.
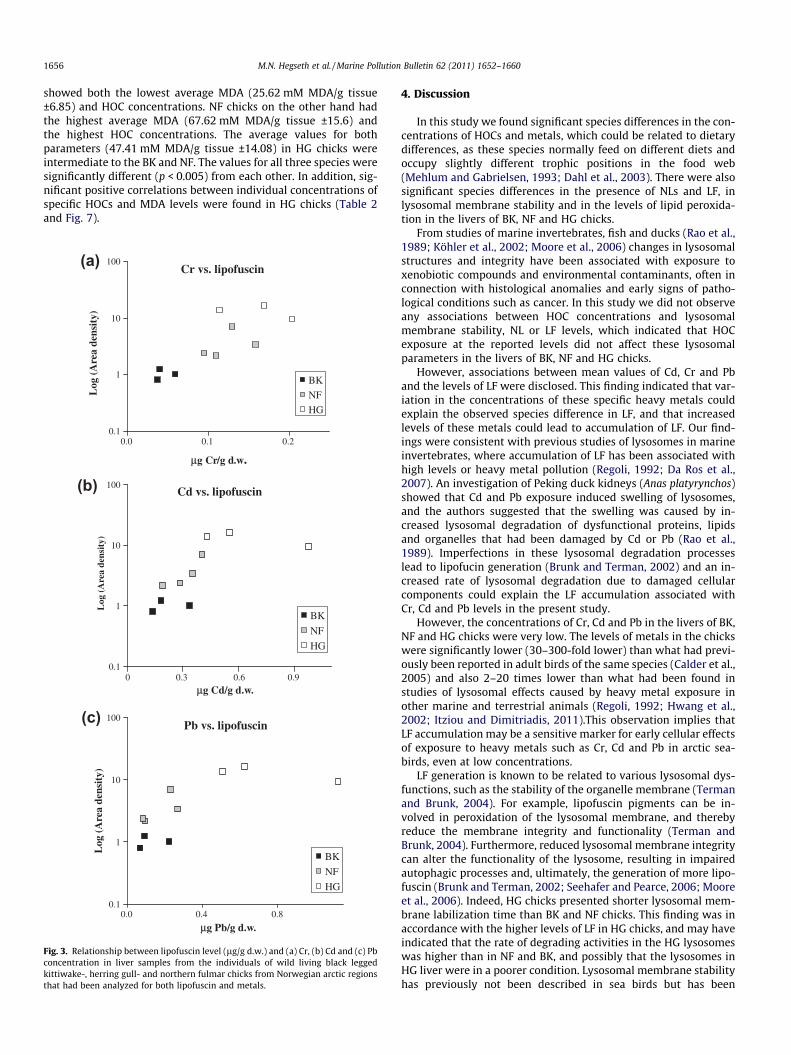
est metal concentrations, also presented the lowest presence of LF,and HG, where the highest mean metal concentrations were found,presented the highest presence of LF. Correlation analyses on anindividual level were unachievable to perform because there wasa mismatch in the selection of samples that were chosen for LFand metal analysis, such that few individuals had been analyzedfor both LF and metals. However, Fig. 3 shows the relationships be-tween LF level and metal concentration in the individuals of thethree species that had been analyzed for both LF and metals, andindicates that there are positive correlations between the metalsand LF levels across the species.
3.2. Neutral lipids, lipofuscin and lysosomal membrane stability
HG exhibited a significantly higher average of both NLs (Fig. 4a–c) and LF (Fig. 4d–f) (NLs: 9.05 area density p < 0.05 ±1.07,p < 0.0001, LF: 19.70 area density ±9.7, p < 0.0001) than NF (NLs:6.48 area density, ±2.03, LF: 5.08 area density ±2.5 p < 0.01), andBK that presented the lowest values for both parameters (NLs:5.59 area density, ±2.06, LF: 1.19 area density ±1.2) The averageNLs area density values of BK and NF were not significantly differ-ent, but the average LF area density values of BK were significantlylower (p < 0.0005) than the average for NF chicks (Fig. 5).
In addition to presenting the highest levels of NLs and LF, HGalso exhibited the shortest average membrane labilization time(8.5 min, range: 7.8–9.3 min) compared to BK (14.17 min, range10–17.5 min) and NF (14.75 min, range 11.8–16.3 min) (Fig. 6).We did not test for statistically significant differences betweenthese values due to the small number of observations (n = 3).
3.3. Lipid peroxidation
We found a concurrence between the average MDA levels andthe total HOC concentrations of the three species. BK chicks
wild living black legged kittiwake-, herring gull- and northern fulmar chicks fromnce.
ern fulmar Herring gull p-Value
e Std. dev. Average Std. dev.
225.5 499.3 71.6 <0.00010.082 0.526 0.269 <0.050.024 0.154 0.037 <0.00010.088 0.703 0.246 <0.001
etals
rn fulmar Herring gull
legged kittiwake-, herring gull- and northern fulmar chicks from Norwegian arctic
1656 M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660
showed both the lowest average MDA (25.62 mM MDA/g tissue±6.85) and HOC concentrations. NF chicks on the other hand hadthe highest average MDA (67.62 mM MDA/g tissue ±15.6) andthe highest HOC concentrations. The average values for bothparameters (47.41 mM MDA/g tissue ±14.08) in HG chicks wereintermediate to the BK and NF. The values for all three species weresignificantly different (p < 0.005) from each other. In addition, sig-nificant positive correlations between individual concentrations ofspecific HOCs and MDA levels were found in HG chicks (Table 2and Fig. 7).
(a) Cr vs. lipofuscin
0.1
1
10
100
0.0 0.1 0.2
μg Cr/g d.w.
Log
(A
rea
dens
ity)
BK
NF
HG
(b) Cd vs. lipofuscin
0.1
1
10
100
0 0.3 0.6 0.9
μg Cd/g d.w.
Log
(A
rea
dens
ity)
BK
NF
HG
(c) Pb vs. lipofuscin
0.1
1
10
100
0.0 0.4 0.8
μg Pb/g d.w.
Log
(A
rea
dens
ity)
BK
NF
HG
Fig. 3. Relationship between lipofuscin level (lg/g d.w.) and (a) Cr, (b) Cd and (c) Pbconcentration in liver samples from the individuals of wild living black leggedkittiwake-, herring gull- and northern fulmar chicks from Norwegian arctic regionsthat had been analyzed for both lipofuscin and metals.
4. Discussion
In this study we found significant species differences in the con-centrations of HOCs and metals, which could be related to dietarydifferences, as these species normally feed on different diets andoccupy slightly different trophic positions in the food web(Mehlum and Gabrielsen, 1993; Dahl et al., 2003). There were alsosignificant species differences in the presence of NLs and LF, inlysosomal membrane stability and in the levels of lipid peroxida-tion in the livers of BK, NF and HG chicks.
From studies of marine invertebrates, fish and ducks (Rao et al.,1989; Köhler et al., 2002; Moore et al., 2006) changes in lysosomalstructures and integrity have been associated with exposure toxenobiotic compounds and environmental contaminants, often inconnection with histological anomalies and early signs of patho-logical conditions such as cancer. In this study we did not observeany associations between HOC concentrations and lysosomalmembrane stability, NL or LF levels, which indicated that HOCexposure at the reported levels did not affect these lysosomalparameters in the livers of BK, NF and HG chicks.
However, associations between mean values of Cd, Cr and Pband the levels of LF were disclosed. This finding indicated that var-iation in the concentrations of these specific heavy metals couldexplain the observed species difference in LF, and that increasedlevels of these metals could lead to accumulation of LF. Our find-ings were consistent with previous studies of lysosomes in marineinvertebrates, where accumulation of LF has been associated withhigh levels or heavy metal pollution (Regoli, 1992; Da Ros et al.,2007). An investigation of Peking duck kidneys (Anas platyrynchos)showed that Cd and Pb exposure induced swelling of lysosomes,and the authors suggested that the swelling was caused by in-creased lysosomal degradation of dysfunctional proteins, lipidsand organelles that had been damaged by Cd or Pb (Rao et al.,1989). Imperfections in these lysosomal degradation processeslead to lipofucin generation (Brunk and Terman, 2002) and an in-creased rate of lysosomal degradation due to damaged cellularcomponents could explain the LF accumulation associated withCr, Cd and Pb levels in the present study.
However, the concentrations of Cr, Cd and Pb in the livers of BK,NF and HG chicks were very low. The levels of metals in the chickswere significantly lower (30–300-fold lower) than what had previ-ously been reported in adult birds of the same species (Calder et al.,2005) and also 2–20 times lower than what had been found instudies of lysosomal effects caused by heavy metal exposure inother marine and terrestrial animals (Regoli, 1992; Hwang et al.,2002; Itziou and Dimitriadis, 2011).This observation implies thatLF accumulation may be a sensitive marker for early cellular effectsof exposure to heavy metals such as Cr, Cd and Pb in arctic sea-birds, even at low concentrations.
LF generation is known to be related to various lysosomal dys-functions, such as the stability of the organelle membrane (Termanand Brunk, 2004). For example, lipofuscin pigments can be in-volved in peroxidation of the lysosomal membrane, and therebyreduce the membrane integrity and functionality (Terman andBrunk, 2004). Furthermore, reduced lysosomal membrane integritycan alter the functionality of the lysosome, resulting in impairedautophagic processes and, ultimately, the generation of more lipo-fuscin (Brunk and Terman, 2002; Seehafer and Pearce, 2006; Mooreet al., 2006). Indeed, HG chicks presented shorter lysosomal mem-brane labilization time than BK and NF chicks. This finding was inaccordance with the higher levels of LF in HG chicks, and may haveindicated that the rate of degrading activities in the HG lysosomeswas higher than in NF and BK, and possibly that the lysosomes inHG liver were in a poorer condition. Lysosomal membrane stabilityhas previously not been described in sea birds but has been
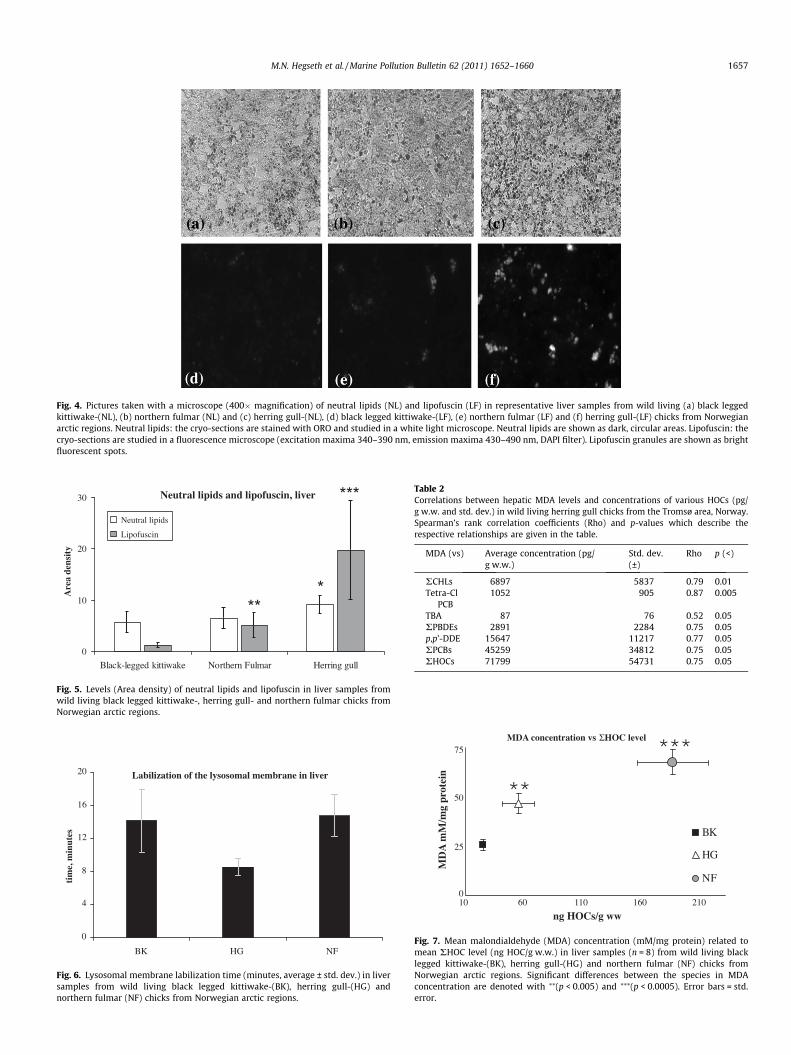
Fig. 4. Pictures taken with a microscope (400� magnification) of neutral lipids (NL) and lipofuscin (LF) in representative liver samples from wild living (a) black leggedkittiwake-(NL), (b) northern fulmar (NL) and (c) herring gull-(NL), (d) black legged kittiwake-(LF), (e) northern fulmar (LF) and (f) herring gull-(LF) chicks from Norwegianarctic regions. Neutral lipids: the cryo-sections are stained with ORO and studied in a white light microscope. Neutral lipids are shown as dark, circular areas. Lipofuscin: thecryo-sections are studied in a fluorescence microscope (excitation maxima 340–390 nm, emission maxima 430–490 nm, DAPI filter). Lipofuscin granules are shown as brightfluorescent spots.
Neutral lipids and lipofuscin, liver
0
10
20
30
Black-legged kittiwake Northern Fulmar Herring gull
Are
a de
nsit
y
Neutral lipids
Lipofuscin
*
***
**
Fig. 5. Levels (Area density) of neutral lipids and lipofuscin in liver samples fromwild living black legged kittiwake-, herring gull- and northern fulmar chicks fromNorwegian arctic regions.
Labilization of the lysosomal membrane in liver
0
4
8
12
16
20
BK HG NF
tim
e, m
inut
es
Fig. 6. Lysosomal membrane labilization time (minutes, average ± std. dev.) in liversamples from wild living black legged kittiwake-(BK), herring gull-(HG) andnorthern fulmar (NF) chicks from Norwegian arctic regions.
Table 2Correlations between hepatic MDA levels and concentrations of various HOCs (pg/g w.w. and std. dev.) in wild living herring gull chicks from the Tromsø area, Norway.Spearman’s rank correlation coefficients (Rho) and p-values which describe therespective relationships are given in the table.
MDA (vs) Average concentration (pg/g w.w.)
Std. dev.(±)
Rho p (<)
RCHLs 6897 5837 0.79 0.01Tetra-Cl
PCB1052 905 0.87 0.005
TBA 87 76 0.52 0.05RPBDEs 2891 2284 0.75 0.05p,p’-DDE 15647 11217 0.77 0.05RPCBs 45259 34812 0.75 0.05RHOCs 71799 54731 0.75 0.05
0
25
50
75
10 60 110 160 210
ng HOCs/g ww
MD
A m
M/m
g pr
otei
n
BK
HG
NF
MDA concentration vs ΣHOC level
Fig. 7. Mean malondialdehyde (MDA) concentration (mM/mg protein) related tomean RHOC level (ng HOC/g w.w.) in liver samples (n = 8) from wild living blacklegged kittiwake-(BK), herring gull-(HG) and northern fulmar (NF) chicks fromNorwegian arctic regions. Significant differences between the species in MDAconcentration are denoted with ⁄⁄(p < 0.005) and ⁄⁄⁄(p < 0.0005). Error bars = std.error.
M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660 1657

1658 M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660
extensively studied in marine bivalves and fish (Köhler et al., 2002;Bocchetti and Regoli, 2006; Cajaraville et al., 2006; Gorbi et al.,2008; Raftopoulou and Dimitriadis, 2010). These studies showedthat exposure to both organic contaminants and metals can desta-bilize the lysosomal membrane at environmentally relevant con-centrations in marine and terrestrial invertebrates (Lowe andPipe, 1994; Reinecke and Reinecke, 2003; Itziou and Dimitriadis,2011), fish (Köhler et al., 2002) and hepatic cell lines (Fotakiset al., 2005). The levels of organic contaminants found in this studywas, however, not related to the changes in lysosomal membranestability in the birds, but increased Cr, Cd and Pb concentrationswere associated with lower lysosomal membrane stability in HGchicks compared to NF and BK. Based on previous studies, exposureto heavy metals, and Cd in particular, is strongly associated withimpairment of lysosomal integrity and function consistently acrossthe phyla, from slime molds to bivalves and fish (Regoli, 1992;Köhler et al., 2002; Reinecke and Reinecke, 2003; Fotakis et al.,2005; Dondero et al., 2006; Moore et al., 2008; Koukouzika andDimitriadis, 2008b; Itziou and Dimitriadis, 2011). The mechanismsbehind the lysosomal impairment by Cd has, however, not beenclearly described, but oxidation of membrane lipids, inactivationof enzymes (Koizumi et al., 1996; Bertin and Averbeck, 2006) or ex-ceeded lysosomal storage capacity due to engulfed Cd-metallothi-onein complexes (Fotakis et al., 2005) have been postulated asunderlying causes. The findings in the present study imply that thismay be the case also in arctic seabirds.
Exposure to contaminants has been associated with lipidosis-like conditions and accumulation of NLs in lysosomes (Moore,1988), but the reported effects of contaminant exposure on NL lev-els have not been consistent, and have varied from no observed ef-fect (Regoli, 1992) to depletion of NLs (Koukouzika and Dimitriadis,2008a). In this study we found associations between the mean con-centrations of Cd, Cr and Pb and the mean levels of NLs in the threespecies, similar to the pattern which was seen for LF. This findingindicated that increased levels of these metals could induce NLaccumulation. However, several stress factors besides contaminantexposure can affect the levels of NL, as these lipids constitute a por-tion of the cellular energy reserves (Moore, 1988; Murphy, 2001).Consequently, the most important influences on NL levels areprobably nutritional factors (Murphy, 2001; Martin and Parton,2006). Lipid droplets are rapidly formed when there are excessamounts of fatty acids, and NL are equally rapidly mobilised fromthese organelles when there is a demand for these compounds,either as membrane components or as an energy source (Martinand Parton, 2005). At the time of sampling, the HG chicks were en-ticed with fish guts as bait in the sampling process and this newlydigested and potentially fat-rich meal could have contributed tothe higher NL levels observed in HG chicks. Additionally, the onlyenergy source for the BK chicks at the time of sampling was thefood fed to them by their parents. The amount of food contributedby the parents may have been insufficient to cover the increasingcalorie needs of these almost fully grown chicks, which would havebeen spending considerable amounts of energy on preparation forfledging at that stage (Jones, 1982; Drent et al., 1992; Gabrielsenet al., 1992). This caloric shortfall may have resulted in a mismatchbetween their energy supply and energy requirements and conse-quently, depleted lipid storages and low levels of NLs which wereseen in BK livers. The difference in NL levels between the investi-gated species was, therefore, probably highly related to dietary fac-tors as well as being associated with contaminant exposure.
As mentioned above, one of the causes of impaired lysosomalfunctions is destabilisation of the lysosomal membrane causedby lipid peroxidation. In this study lipid peroxidation was mea-sured through the levels of the toxic by-product MDA, and wefound that the observed species differences in MDA levels wereconsistent with the species differences in both HOC and Fe2+ levels.
An increased intracellular level of Fe2+ is considered a risk factorfor increased generation of MDA, as the oxidation of lipids by reac-tive oxygen species takes place through Fenton chemistry, and thisreaction is dependent on the presence of Fe2+ (Cheeseman, 1993).Accordingly, the observed differences in Fe2+ concentrations couldexplain the respective MDA levels in the species. Furthermore, wefound inter-species concurrence between MDA concentrations andHOC levels, and there were strong intra-species correlations be-tween MDA levels and several of the specific HOC groups in theHG chicks. More specifically, strong correlations between tetra-chlorinated PCBs and chlordanes and MDA levels were revealed,which were in agreement with data from previous investigationsof molluscs, human cell lines and rats (Fraga et al., 1987; Moore,1988; Saito, 1990; Regoli, 1992). For example, induction of lipidperoxidation was observed in rat liver after exposure to chlordaneand the tetrachlorobiphenyl PCB77 (Ogata and Izushi, 1991; Fadhelet al., 2002). Nevertheless, no statistically significant correlationsbetween MDA and HOCs were seen in the NF or BK chicks, indicat-ing that lipid peroxidation as a direct consequence of HOC expo-sure may have been specific for HG chicks. Oxidised lipids areassumed to cause destabilisation of lysosomal membranes (Boyaand Kroemer, 2008) as well as being an important factor in LF gen-eration (Brunk and Terman, 2002). However, no correlations wereobserved between MDA levels and lysosomal membrane stabilityor LF levels (data not shown). The assay applied in this study mea-sured general cellular MDA generation in the complete S9-fractionof the tissue homogenates and was, therefore, not specific for thelysosomes, which may have concealed possible direct associationsbetween lysosomal parameters and MDA levels.
The BK and NF chicks were sampled on Svalbard in August 2006and the HG chicks were sampled outside Tromsø on mainland Nor-way in September 2007 due to logistical reasons. This 1 year differ-ence in sampling time could potentially have contributed to theobserved species differences. However, intrinsic features such asfeeding behaviour and geographical belonging differ substantiallybetween the species, and do not change from year to year. Theseare important factors with regards to contaminant concentrationand autophagic lysosomal responses; hence, the 1 year differencein sampling time between HG and BK/NF was probably insignifi-cant with regards to the observed species differences in this study.Nevertheless, annual variation in dietary contaminant levels or ac-cess to food at the different sample sites were not accounted for inthis investigation as each species was only sampled once, and cannot completely be ruled out as a contributing factor to the ob-served species differences.
In summary, this study indicated that LF and NL accumulation,and possibly also lysosomal membrane stability were affected byCr, Cd and Pb exposure at low levels, and that these parameters,which are regularly used as biomarkers of various types of environ-mental stress in other marine animals, may be good candidates formarkers of cellular stress also in arctic seabird chicks. Similarly, li-pid peroxidation could act as a marker of oxidative damage in-duced by HOC and Fe2+ exposure, particularly in HG. The findingsin this study are however limited by the small number of observa-tions, and further investigations including larger sample sizes andpreferably larger variation in contaminant levels or controlledexperiments are needed to better assess the effects of contaminantexposure on lysosomal parameters in seabirds.
Acknowledgements
The authors are grateful to the staff of The Norwegian Instituteof Air Research for the HOC analysis and Daniele Fattorini in Anc-ona for Iron analysis. We are also grateful to Hans Wolkers, LindaBakken, Marte Braathen, Christian Wagner, Jonathan Teuchertand Ingeborg Hallanger for helping with the sampling in the field.

M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660 1659
We would also like to thank the helpful staff at Prof. F. Regoli’s labin Ancona for helping with the histological analysis. The projectwas funded by the Norwegian Research Council, and permissionsto sample the birds were given by the governor of Svalbard andthe council governor of Troms County.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.marpolbul.2011.06.011.
References
Aust, S.D., 1985. Lipid peroxidation. In: Greenwald, R.A. (Ed.), Handbook of Methodsfor Oxygen Radical Research. CRC, Boca Raton, Florida, pp. 203–207.
Berglund, M.M., Sturve, J., Förlin, L., Nyholm, N.E.I., 2007. Oxidative stress in piedflycatcher (Ficedula hypoleuca) nestlings from metal contaminatedenvironments in northern Sweden. Environmental Research 105, 330–339.
Bertin, G., Averbeck, D., 2006. Cadmium: cellular effects, modifications of ábiomolecules, modulation of á DNA repair and á genotoxic consequences (aáreview). Biochimie 88, 1549–1559.
Bocchetti, R., Regoli, F., 2006. Seasonal variability of oxidative biomarkers,lysosomal parameters, metallothioneins and peroxisomal enzymes in theMediterranean mussel Mytilus galloprovincialis from Adriatic Sea.Chemosphere 65, 913–921.
Borga, K., Gabrielsen, G.W., Skaare, J.U., 2001. Biomagnification of organochlorinesalong a Barents Sea food chain. Environmental Pollution 113, 187–198.
Boya, P., Kroemer, G., 2008. Lysosomal membrane permeabilization in cell death.Oncogene 27, 6434–6451.
Brunk, U.T., Terman, A., 2002. Lipofuscin: mechanisms of age-related accumulationand influence on cell function. Free Radical Biology and Medicine 33, 611–619.
Cajaraville, M.P., Garmendia, L., Orbea, A., Werding, R., Gmez-Mendikute, A.,Izagirre, U., Soto, M., Marigmez, I., 2006. Signs of recovery of mussels healthtwo years after the Prestige oil spill. Marine Environmental Research 62, S337–S341.
Calder, John, AMAP Working group., 2005. AMAP Assessment 2002: Heavy Metals inthe Arctic. Arctic Monitoring and Assessment Programme (AMAP). xvi-265,Oslo, Norway.
Cheeseman, K.H., 1993. Mechanisms and effects of lipid peroxidation. MolecularAspects of Medicine 14, 191–197.
Da Ros, L., Moschino, V., Guerzoni, S., Halldrsson, H.P., 2007. Lysosomal responsesand metallothionein induction in the blue mussel Mytilus edulis from the south-west coast of Iceland. Environment International 33, 362–369.
Dahl, T.M., Falk-Petersen, S., Gabrielsen, G.W., Sargent, J.R., Hop, H., Millar, R.M.,2003. Lipids and stable isotopes in common eider, black-legged kittiwake andnorthern fulmar: a trophic study from an Arctic fjord. Marine Ecology ProgressSeries 256, 257–269.
de Duve, C., 2005. The lysosome turns fifty. Nature Cell Biology 7, 847–849.de Wit CA, Fisk A, Hobbs K, Muir D, Gabrielsen G, Kallenborn R, Krahn M, Norstrom
R, Skaare J. 2004. AMAP 2002: persistent organic pollutants in the Arctic. Oslo,Norway.
Domouhtsidou, G.P., Dimitriadis, V.K., 2001. Lysosomal and lipid alterations in thedigestive gland of mussels, Mytilus galloprovincialis (L.) as biomarkers ofenvironmental stress. Environmental Pollution 115, 123–137.
Dondero, F., Jonsson, H., Rebelo, M., Pesce, G., Berti, E., Pons, G., Viarengo, A., 2006.Cellular responses to environmental contaminants in amoebic cells of the slimemould Dictyostelium discoideum. Comparative Biochemistry and Physiology PartC: Toxicology & Pharmacology 143, 150–157.
Drent, R.H., Klaassen, M., Zwaan, 1992. Predictive growth budgets in gulls and terns.ARDEA 80, 5–17.
Eskelinen, E.L., Saftig, P., 2009. Autophagy: a lysosomal degradation pathway with acentral role in health and disease. Biochimica et Biophysica Acta (BBA) -Molecular Cell Research 1793, 664–673.
Fadhel, Z., Lu, Z., Robertson, L.W., Glauert, H.P., 2002. Effect of 3,30 ,4,40-tetrachlorobiphenyl and 2,20 , 4,40 ,5,50-hexachlorobiphenyl on the induction ofhepatic lipid peroxidation and cytochrome P-450 associated enzyme activitiesin rats. Toxicology 175, 15–25.
Fang, J.K.H., Wu, R.S.S., Zheng, G.J., Lam, P.K.S., Shin, P.K.S., 2010. Seasonality ofbioaccumulation of trace organics and lysosomal integrity in green-lippedmussel Perna viridis. Science of the Total Environment 408, 1458–1465.
Fattorini, D., Notti, A., Nigro, M., Regoli, F., 2010. Hyperaccumulation of vanadium inthe Antarctic polychaete Perkinsiana littoralis; as a natural chemical defenseagainst predation. Environmental Science and Pollution Research 17, 220–228.
Fernie, K.J., Shutt, J.L., Mayne, G., Hoffman, D., Letcher, R.J., Drouillard, K.G., Ritchie,I.J., 2005. Exposure to polybrominated diphenyl ethers (PBDEs): changes inthyroid, vitamin A, glutathione homeostasis, and oxidative stress in Americankestrels (Falco sparverius). Toxicological Sciences 88, 375–383.
Fotakis, G., Cemeli, E., Anderson, D., Timbrell, J.A., 2005. Cadmium chloride-inducedDNA and lysosomal damage in a hepatoma cell line. Toxicology in Vitro 19,481–489.
Fraga, C.G., Leibovitz, B.E., Tappel, A.L., 1987. Halogenated compounds as inducers oflipid peroxidation in tissue slices. Free Radical Biology and Medicine 3, 119–123.
Gabrielsen, G.W., Klaassen, M., Mehlum, F., 1992. Energetics of Black-leggedkittiwake Rissa Tridactyla chicks. ARDEA 80, 29–40.
Gabrielsen, G.W., Sydnes, L.K., 2009. Pollution in the Barents Sea. In: Sakshaug, E.,Johnsen, G., Kovacs, K. (Eds.), Ecosystem Barents Sea. Tapir Academic Press, pp.497–544.
Gorbi, S., Virno Lamberti, C., Notti, A., Benedetti, M., Fattorini, D., Moltedo, G., Regoli,F., 2008. An ecotoxicological protocol with caged mussels, Mytilusgalloprovincialis, for monitoring the impact of an offshore platform in theAdriatic Sea. Marine Environmental Research 65, 34–49.
Halliwell, B., Gutteridge, J.M.C., 2007. Free radicals in Biology and Medicine. OxfordUniversity Press, New York.
Hegseth, M.Ns., Camus, L., Helgason, L.B., Bocchetti, R., Gabrielsen, G.W., Regoli, F.,2011. Hepatic antioxidant responses related to levels of PCBs and metals inchicks of three Arctic seabird species. Comparative Biochemistry and PhysiologyPart C: Toxicology & Pharmacology 154, 28–35.
Helgason, L.B., Arukwe, A., Gabrielsen, G.W., Harju, M., Hegseth, M.N., Heimstad,Er.S., Jrgensen, E.H., Mortensen, A.S., Wolkers, J., 2010. Biotransformation ofPCBs in Arctic seabirds: characterization of phase I and II pathways attranscriptional, translational and activity levels. Comparative Biochemistryand Physiology Part C: Toxicology & Pharmacology 152, 34–41.
Hwang, H.M., Wade, T.L., Sericano, J.L., 2002. Relationship between lysosomalmembrane destabilization and chemical body burden in eastern oysters(Crassostrea virginica) from Galveston Bay, TX, USA. Environmental Toxicologyand Chemistry 21, 1268–1271.
Itziou, A., Dimitriadis, V.K., 2011. Introduction of the land snail Eobania vermiculataas a bioindicator organism of terrestrial pollution using a battery of biomarkers.Science of the Total Environment 409, 1181–1192.
Jones, M.M., 1982. Growth of the pectoralis muscle of the house sparrow (Passerdomesticus). Journal of anatomy 135, 719–731.
Köhler, A., Wahl, E.S., Âffker, K., 2002. Functional and morphological changes oflysosomes as prognostic biomarkers of toxic liver injury in a marine flatfish(Platichthys flesus (L.)). Environmental Toxicology and Chemistry 21, 2434–2444.
Kaminski, P., Kurhalyuk, N., Jerzak, L., Kasprzak, M., Tkachenko, H., Klawe, J.J., Szady-Grad, M., Koim, B., Wisniewska, E., 2009. Ecophysiological determinations ofantioxidant enzymes and lipoperoxidation in the blood of White Stork Ciconiaciconia from Poland. Environmental Research 109, 29–39.
Koizumi, T., Shirakura, H., Kumagai, H., Tatsumoto, H., Suzuki, K.T., 1996.Mechanism of cadmium-induced cytotoxicity in rat hepatocytes: cadmium-induced active oxygen-related permeability changes of the plasma membrane.Toxicology 114, 125–134.
Koukouzika, N., Dimitriadis, V.K., 2008a. Aspects of the usefulness of five marinepollution biomarkers, with emphasis on MN and lipid content. Marine PollutionBulletin 56, 941–949.
Kubetzki, U., Garthe, S., 2003. Distribution, diet and habitat selection by foursympatrically breeding gull species in the south-eastern North Sea. MarineBiology 143, 199–207.
Lowe, D.M., Pipe, R.K., 1994. Contaminant induced lysosomal membrane damage inmarine mussel digestive cells: an in vitro study. Aquatic Toxicology 30, 357–365.
Lowe, D., 1988. Alterations in cellular structure of Mytilus edulis resulting fromexposure to environmental contaminants under field and experimentalconditions. Marine Ecology Progress Series 46, 91–100.
Luzio, J.P., Pryor, P.R., Bright, N.A., 2007. Lysosomes: fusion and function. NatureReviews Molecular Cell Biology 8, 622–632.
Martin, S., Parton, R.G., 2005. Caveolin, cholesterol, and lipid bodies. Seminars inCell & Developmental Biology 16, 163–174.
Martin, S., Parton, R.G., 2006. Lipid droplets: a unified view of a dynamic organelle.Nature Reviews Molecular Cell Biology 7, 373–378.
Mehlum, F., Gabrielsen, G.W., 1993. The diet of high-arctic seabirds in coastal andice-covered, pelagic areas near the Svalbard archipelago. Polar Research 12, 1–20.
Moore, M.N., Lowe, D.M., Köhler, A., 2004. Biological effects of contaminants:measurement of lysosomal membrane stability. ICES Techniques in MarineEnvironmental Sciences 36, 31.
Moore, M.N., 1988. Cytochemical responses of the lysosomal system and NADPH-ferrihemoprotein reductase in molluscan digestive cells to environmental andexperimental exposure to xenobiotics. Marine Ecology Progress Series 46, 81–89.
Moore, M.N., Allen, J.I., McVeigh, A., Shaw, J., 2006. Lysosomal and Autophagicreactions as predictive indicators of environmental impact in aquatic animals.Autophagy 2, 217–220.
Moore, M.N., Koehler, A., Lowe, D., Viarengo, A., 2008. Three lysosomes andautophagy in aquatic animals. In: Daniel, J.K. (Ed.), Methods in EnzymologyAutophagy: Lower Eukaryotes and Non-Mammalian Systems Part A. AcademicPress, New York, pp. 581–620, Chapter Thirty[hyphen (true graphic)].
Moore, M.N., Viarengo, A., Donkin, P., Hawkins, A.J.S., 2007. Autophagic andlysosomal reactions to stress in the hepatopancreas of blue mussels. AquaticToxicology 84, 80–91.
Murphy, D.J., 2001. The biogenesis and functions of lipid bodies in animals, plantsand microorganisms. Progress in Lipid Research 40, 325–438.
Nesto, N., Romano, S., Moschino, V., Mauri, M., Da Ros, L., 2007. Bioaccumulationand biomarker responses of trace metals and micro-organic pollutants in

1660 M.N. Hegseth et al. / Marine Pollution Bulletin 62 (2011) 1652–1660
mussels and fish from the Lagoon of Venice, Italy. Marine Pollution Bulletin 55,469–484.
Nøst, TH., Thyroid hormones and HOCs in arctic seabird chicks. 2009. Universitylibrary, Trondheim, Norway, Norwegian University of Technology and Science.
Ogata, M., Izushi, F., 1991. Effects of chlordane on parameters of liver and muscletoxicity in man and experimental animals. Toxicology Letters 56, 327–337.
Raftopoulou, E.K., Dimitriadis, V.K., 2010. Assessment of the health status ofmussels Mytilus galloprovincialis along Thermaikos Gulf (Northern Greece): anintegrative biomarker approach using ecosystem health indices. Ecotoxicologyand Environmental Safety 73, 1580–1587.
Rao, P.V., Jordan, S.A., Bhatnagar, M.K., 1989. Ultrastructure of kidney of ducksexposed to methylmercury, lead and cadmium in combination. Journal ofEnvironmental Pathology, Toxicology and Oncology 9, 19–44.
Regoli, F., 1992. Lysosomal responses as a sensitive stress index in biomonitoringheavy metal pollution. Marine Ecology Progress Series 84, 63–69.
Reinecke, A.J., Reinecke, S.A., 2003. The influence of exposure history to lead on thelysosomal response in Eisenia fetida (Oligochaeta). Ecotoxicology andEnvironmental Safety 55, 30–37.
Ryan, T.P., Aust, S.D., 1992. The role of iron in oxygen-mediated toxicities. CriticalReviews in Toxicology 22, 119–141.
Saito, M., 1990. Polychlorinated biphenyls-induced lipid peroxidation as measuredby thiobarbituric acid-reactive substances in liver subcellular fractions of rats.Biochimica et Biophysica Acta (BBA) - Lipids Lipid Metabolism 1046,301–308.
Sandanger, T., Dumas, P., Berger, U., Burkow, I.C., 2004. Analysis of HO-PCBs and PCPin blood plasma from individuals with high PCB exposure living on theChukotka Peninsula in the Russian Arctic. Journal of Environmental Monitoring6, 758–765.
Seehafer, S.S., Pearce, D.A., 2006. You say lipofuscin, we say ceroid: definingautofluorescent storage material. Neurobiology of Aging 27, 576–588.
Terman, A., Kurz, T., Gustafsson, B., Brunk, U.T., 2006. Lysosomal labilization. IUBMBLife 58, 531–539.
Terman, A., Brunk, U.T., 2004. Lipofuscin. The International Journal of Biochemistry& Cell Biology 36, 1400–1404.
Walther, T.C., Farese, J., 2009. The life of lipid droplets. Biochimica et BiophysicaActa (BBA) - Molecular Cell Biology of Lipids 1791, 459–466.
Wayland, M., Hoffman, D.J., Mallory, M.L., Alisauskas, R.T., Stebbins, K.R., 2010.Evidence of Weak Contaminant-Related Oxidative Stress in Glaucous Gulls(Larus hyperboreus) from the Canadian Arctic. Journal of Toxicology andEnvironmental Health, Part A: Current Issues 73, 1058–1073.
Copyright © 2022 FDOKUMEN




















