
Lymphocyte function in human bone marrow. II. Characterization of an interleukin 2-sensitive T...
12
Journal of Clinical Immunology, Vol. 5, No. 5, 1985 Lymphocyte Function in Human Bone Marrow. Characterization of an Interleukin 2-Sensitive T Precursor-Cell Population II. HANS-MICHAEL DOSCH, 1 CYNTHIA J. LEDGLEY, ~ DANIEL WHITE, 1 PHILIP LAM, 1 and GORDON B. MILLS 1 Accepted: May 13, 1985 In the present study of human bone marrow lymphocytes, we analyze a newly recognized population of T suppressor- cell precursors which are found in marrow only and which have the potential to inhibit immunoglobulin (Ig) production in vitro. Follo~dng exposure to interleukin 2 (IL2), suppres- sor precursors acquire E receptor, T3 determinants, sup- pressor function, and lectin responsiveness. To distinguish this population within the framework of T-ceU ontogeny, it was compared to a previously described population of thymus-dependent helper T-cell precursors which express helper function following exposure to thymus-derived me- diators. The two populations are completely distinct and can be separated on density gradients. Suppressor precur- sors expressed T8 and TAC (IL2-receptor) antigens prior to in vitro induction with IL2. The thymic hormone-dependent cells expressed T4 but not T8 or TAC determinants. In two patients with severe combined immunodeficiency disease (SCID), IL2-responsive precursor cells appeared only late after thymus epithelium transplantation, perhaps best ex- plained by a model in which thymus-dependent differentia- tion pathways precede, induce, or seed pathways of extra- thymic T-cell differentiation. The large pool size of over 1011 suppressor and helper precursor cells present in adult bone marrow suggests that these populations may play an impor- tant role in immune homeostasis. KEY WORDS: T-cell ontogeny; suppressor/helper cells; thymic hormone; interleukin 2 (IL2); severe combined immunodefici- ency disease (SCID). INTRODUCTION Bone marrow is the source of up to two-thirds of circulating immunoglobulin (Ig). Marrow cells can ~Research Institute, Division of Immunology/Rheumatology, The Hospital for Sick Children, 555 University Avenue, Toronto, Ontario, Canada M5G 1X8. secrete large amounts of IgG and IgA in vitro and the frequency of antigen-responsive B cells is high in marrow (1-6). The immunoregulatory rules which govern Ig production in this tissue site are not entirely clear (5-10). In the present report, we examine regulatory T-cell populations present in bone marrow. In an earlier report (11), we de- scribed the detection of a unique suppressor-cell function in bone marrow-cell cultures. We pro- posed a model of a regulatory cascade in which a small marrow T-cell population produces large amounts of interleukin 2 (IL2) in respe.nse to mitogenic activation. This IL2 induces precursors of suppressor effector cells to acquire T-cell surface properties and suppressor function without requir- ing additional activation signals. We now character- ize these IL2-dependent suppressor-cell precursors and their progeny. In an attempt to view these cells within the framework of T-cell ontogeny, IL2-de- pendent marrow cells were compared to T helper- cell precursors previously found to require expo- sure to thymic factors for their functional matura- tion (10). Our data show that the two precursor-cell populations are completely distinct. Only ~[L2-de- pendent suppressor-cell precursors appear as an exclusively marrow resident population. They con- stitutively express T8 and TAC (IL2-receptor) an- tigens (12, 13). They acquire E receptors, T3 deter- minants, suppressor function, and the capacity to proliferate in response to mitogen following as little as 4 hr of incubation with IL2. In two patients with severe combined immunodeficiency disease (SCID), IL2-dependent marrow T precursors appeared only late after receiving thymic epithelial transplants. This could imply an interdepedence of the two precursor-ceU pools within pathways of T-cell dif- 345 0271-9142/85/0900-0345504.50/0 © 1985 Plenum Publishing Corporation
Transcript of Lymphocyte function in human bone marrow. II. Characterization of an interleukin 2-sensitive T...

Journal of Clinical Immunology, Vol. 5, No. 5, 1985
Lymphocyte Function in Human Bone Marrow. Characterization of an Interleukin 2-Sensitive T Precursor-Cell Population
II.
H A N S - M I C H A E L DOSCH, 1 C Y N T H I A J. L E D G L E Y , ~ D A N I E L WH ITE, 1 P H I L I P LAM, 1 and GORDON B. M I L L S 1
Accepted: May 13, 1985
In the present study of human bone marrow lymphocytes, we analyze a newly recognized population of T suppressor- cell precursors which are found in marrow only and which have the potential to inhibit immunoglobulin (Ig) production in vitro. Follo~dng exposure to interleukin 2 (IL2), suppres- sor precursors acquire E receptor, T3 determinants, sup- pressor function, and lectin responsiveness. To distinguish this population within the framework of T-ceU ontogeny, it was compared to a previously described population of thymus-dependent helper T-cell precursors which express helper function following exposure to thymus-derived me- diators. The two populations are completely distinct and can be separated on density gradients. Suppressor precur- sors expressed T8 and TAC (IL2-receptor) antigens prior to in vitro induction with IL2. The thymic hormone-dependent cells expressed T4 but not T8 or TAC determinants. In two patients with severe combined immunodeficiency disease (SCID), IL2-responsive precursor cells appeared only late after thymus epithelium transplantation, perhaps best ex- plained by a model in which thymus-dependent differentia- tion pathways precede, induce, or seed pathways of extra- thymic T-cell differentiation. The large pool size of over 1011 suppressor and helper precursor cells present in adult bone marrow suggests that these populations may play an impor- tant role in immune homeostasis.
KEY WORDS: T-cell ontogeny; suppressor/helper cells; thymic hormone; interleukin 2 (IL2); severe combined immunodefici- ency disease (SCID).
INTRODUCTION
Bone marrow is the source of up to two-thirds of circulating immunoglobulin (Ig). Marrow cells can
~Research Institute, Division of Immunology/Rheumatology, The Hospital for Sick Children, 555 University Avenue, Toronto, Ontario, Canada M5G 1X8.
secrete large amounts of IgG and IgA in vitro and the f requency of antigen-responsive B cells is high in marrow (1-6). The immunoregulatory rules which govern Ig product ion in this tissue site are not entirely clear (5-10). In the present report , we examine regulatory T-cell populations present in bone marrow. In an earlier report (11), we de- scribed the detect ion of a unique suppressor-cell function in bone marrow-cell cultures. We pro- posed a model of a regulatory cascade in which a small marrow T-cell population produces large amounts of interleukin 2 (IL2) in respe.nse to mitogenic activation. This IL2 induces precursors of suppressor effector cells to acquire T-cell surface properties and suppressor function without requir- ing additional activation signals. We now character- ize these IL2-dependent suppressor-cell precursors and their progeny. In an at tempt to view these cells within the f ramework of T-cell ontogeny, IL2-de- pendent marrow cells were compared to T helper- cell precursors previously found to require expo- sure to thymic factors for their functional matura- tion (10). Our data show that the two precursor-cell populations are completely distinct. Only ~[L2-de- pendent suppressor-cell precursors appear as an exclusively marrow resident population. They con- stitutively express T8 and TAC (IL2-receptor) an- tigens (12, 13). They acquire E receptors , T3 deter- minants, suppressor function, and the capacity to proliferate in response to mitogen following as little as 4 hr of incubation with IL2. In two patients with severe combined immunodeficiency disease (SCID), IL2-dependent marrow T precursors appeared only late after receiving thymic epithelial transplants. This could imply an interdepedence of the two precursor-ceU pools within pathways of T-cell dif-
345
0271-9142/85/0900-0345504.50/0 © 1985 Plenum Publishing Corporation

346 DOSCH, LEDGLEY~ WHITE~ LAM, AND MILLS
ferentiation. Considering the population size (4-6% of marrow cells or about 101~ precursors in a normal adult), we feel that the marrow suppressor-cell precursor pool may represent a major source of cells of the suppressor T-cell sublineage.
MATERIALS AND METHODS
Cell Preparation and Culture Conditions
Bone marrow cells were obtained from surgical bone and rib specimens from adult patients. Cells from marrow, tonsil, or blood were enriched for mononuclear cells on Ficotl-Hypaque gradients (6). Cell viabilities were >98% by dye exclusion. Where indicated, cells were further purified by E-rosette depletion on a second Ficoll-Hypaque gradient as described (6).
Complete culture medium consisted of RPMI- 1640 (Ontario Cancer Institute, Toronto, Ontario) supplemented with 10% fetal bovine serum (HiClone Sterile Systems, Inc., Logan, UT), anti- biotics, and glutamine (Difco Laboratories, Detroit, MI). In one series of experiments, unseparated and E rosette-depleted marrow cells or tonsillar lym- phocytes were incubated in the presence of phytohemagglutinin (PHA) (5 txg/ml, Pharmacia, Ltd., Montreal, Quebec) and IL2 (15 U/ml; see below). After 1 or 2 weeks, cultures were har- vested, washed, enriched for viable cells on FicoU-Hypaque gradients, and used for celt surface marker and functional studies. Aliquots were pulsed overnight with [3H]thymidine (I l~Ci/well) and isotope incorporation was determined by liquid scintillation counting.
For the measurement of immunoglobulin produc- tion, 1-2 x 105 marrow cells were cultured for 6-8 days in 200 txl of complete medium. The amounts of IgG, IgA and IgM secreted into culture superna- tants were determined in a standard solid-phase ELISA procedure. Briefly, affinity-purified isotype- specific goat antibodies (Tago Diagnostics, Inc., Burlingame, CA) were bound adhesively to 96-well polystyrene plates. Unbound material was removed and 20 txl of culture supernatant was incubated overnight in the antibody-coated wells. Following a second washing procedure, human Ig bound to the goat antibodies was measured enzymatically using alkaline phosphatase coupled to anti-IgG, -IgA, or -IgM antibodies (Tago Diagnostics, Inc.). Optical densities (OD410nm) were determined in a Dynatech MR600 ELISA reader (Fisher Scientific Co.,
Toronto, Ontario) using Apple IIe software devel- oped by Data Disc Services (Toronto).
Induction o f Precursor-Ceil Differentiation
The following agents were used for the in vitro induction of precursor T-cell differentiation. (i) Thymostimulin 1 (TP: 1) is a calf thymus extract and was a gift from Serono Laboratories (Randolph, MA) (14); a stock solution of 10 ~g/ml in complete culture medium was stored frozen. (ii) Human thy- mic epithelium conditioned medium (HTCM) is supernatant from monolayer cultures of normal human thymic epithelium (HTML) (9, 10). HTCM was stored frozen and used at a 10% (v/v) final concentration. (iii) Theophylline (THEO; Sigma Chemicals, St. Louis, MO) was used at a final concentration of 3 × 10 -3 M. (iv) Tissue culture- derived IL2 was constitutively secreted by the gibbon cell line MLA 144 (11, 15). Culture superna- tants were used without further purification. IL2 derived from the cDNA for human IL2 expressed in Escherichia coIi ("recombinant IL2") (16) was a gift from Biogen (Biogen Research Corp., Cam- bridge, MA). Both sources were standardized in a bioassay using IL2-dependent T lymphoblasts as indicator cells (11). One unit was defined as the amount of IL2 required to stimulate 33% of the maximal [3H]thymidine incorporation in cultures of 104 IL2-dependent lymphoblasts. (v) Supernatant from a subclone of the human histiocyte line U937 (17, 18) served as a control for tissue culture- derived conditioned medium. Secretion of interleu- kin 1 (ILl) was induced in U937 cells by a 12-hr incubation with phorbol myristic acetate (10 -9 m ) , PHA (5 txg/ml), and hydroxyurea (1 pxg/ml) (17). Cells were then thoroughly washed and reincubated in complete medium for another 20 hr, at which time the supernatants were harvested. In some experiments, ILl from activated U937 cells was partially purified using CM-Sephadex (Pharmacia, Ltd.) ion-exchange chromatography, for a more than 100-fold increase in specific activity. ILl ac- tivity was measured by the induction of prolifera- tive responses in mouse thymocytes (18, 19).
To monitor the effects of the various mediators on precursor cells, unseparated or fractionated marrow cells (3-5 x 106/ml) were resuspended in complete culture medium and one of the agents described above was added. Cells were incubated at 37°C for 2-24 hr and then washed. The quantitation of EAET rosette-forming cells served as a screening assay for
Journal of Clinical Immunology, Vol. 5, No. 5, 1985

IL2-SENSITIVE MARROW T CELLS 347
mediator effects and was performed as described (20), except that at least 3000 cells were counted using an eosinophil counting chamber. Where indicated, cyclo- heximide (1 raM; Sigma Chemicals) was added at the beginning of the incubation period.
Cell Separation
Density/pH gradients were prepared from a Tris- buffered solution of 35% bovine serum albumin (BSA; Sigma Chemicals) as described (10). Briefly, the stock solution was diluted in 2% decrements (33, 31%, etc.). Aliquots (1 ml) of each concentra- tion were used to form a discontinuous gradient. Bone marrow cells (1-5 × 107/ml) were suspended in 19% BSA, layered on top of the gradient, and then centrifuged at 3200 rpm for 30 min at 4°C. Cells which sedimented to the different interface regions were harvested and washed. Cell recovery was 70-90% and only in the very high-density fractions (>31% BSA) were cell viabllities below 90%.
Immunofluorescence and Antlbody Studies
Monoclonal anti-T-cell antibodies (T3, T4, T8, T9, T10, T l l ) (12) were obtained from Coulter Electronics, Inc. (Hialeah, FL). The monoclonal anti-TAC antibody directed against the IL2 recep- tor (13) was a gift from Dr. T. A. Waldmann (Na- tional Institutes of Health, Bethesda, MA). The 21w4 monoclonal antibody against nonpolymorphic Ia determinants was provided by Dr. M. Letarte (Toronto). For cytotoxic treatments, appropriate antibody dilutions were added to bone marrow cells (2 × 106/ml RPM 1640) in the presence of rabbit complement (20). After incubations for 1 hr at 10°C and 2 hr at 37°C, cells were washed and used in induction studies. For indirect fluorescent staining, 1 × 106 cells were incubated (0°C, 60 rain) with the appropriate amount of monoclonal antibody, washed, and stained with fluorescinated, affinity-purified F(ab )2 g o a t anti-mouse Ig antibodies (Tago Diagnos- tics, Inc.). Fluorescence studies were done using a cytofluorograph (Epics V, Coulter Electronics, Inc.).
RESULTS
Induction o f T Precursor-Cell Differentiation in Bone Marrow
The appearance of E receptors on marrow cells treated with thymus- or T cell-derived mediators
Table I. Induction of E Rosette-Forming Cells m Bone Marrow a
E-RFC (%) Marrow
donor no. Medium ILl HTCM TP:I IL2
1 1.8 1 6 7.4 7 0 8 9 2 2.1 2~3 8.0 8.4 10.8 3 1.2 1.0 8.5 8.5 9.5 4 3.2 3 4 6.9 7.8 9 7 5 1.4 1.3 7.2 8 8 10 2 6 2 5 2.5 8.0 9 1 11.0 7 0 8 0.9 6 3 6 4 9.2 8 3 9 3 5 8.0 8,2 9.9 9 2.7 2.8 107 100 t0.1
10 3.3 3.5 ND 10 4 9 5 11 3.5 3.1 8.7 9.1 1t.4 t2 4 1 4.2 9.3 10. t 12.0 13 2.9 2.7 ND 8.6 12.21
Mean -+ SD 2.6+-1 2 5 + 1 , 4 8-+-11 8 .6+- i .1 1 0 . 3 - + 1 0
Net E-RFC mduced (%) 0 -0,1 5 4 6.0 7 7
"Bone marrow cells were separated on Ficoll-Hypaque gradients and 3 x 105 cells were incubated (37°C) m the presence of medmm or the medmtors indicated. After 6-8 hr cells were washed and the percentage of E rosette-forming cells (E-RFC) present was determined in the standard assay, The mediators used were supernatant of the human hlstmcyte hne U937 containing mterleukm t ( ILl , 12-15 U/rot), human thymlc eplthehal culture conditioned medium (HTCM; 10%, v/v), thymopoletm 1 (Thymostlmuhn, TP: 1,3 lzg/ml), and mterteukm 2 derived from the MLA 144 cell hne (IL2:12-15 U/mt). ND, not done.
has previously been used to study marrow T precursor-cell differentiation (9-11, 20). It was cho- sen as a screening procedure since the IL2-mediat- ed induction of marrow suppressor-cell activity was paralleled by the acquisition of E receptors in these cells (11 ; see below).
The ability of HTCM, TP: 1, and IL2 to induce the expression of E receptors in 13 marrow samples is shown in Table I. HTCM, TP:I, and IL2 induced the expression of E receptors on 5-8% of the marrow cells. ILl-containing supernatants from U937 cells did not induce E rosette-forming cells (E-RFC). In several experiments, partially purified ILl also did not reduce E-RFC (data not shown). Therefore, within the time period tested (6-8 hr), ILl was unable to induce the appearance of E-RFC in marrow cells.
Thymocytes, marrow cells, and lymphocytes from peripheral tissues were analyzed to determine the tissue distribution of cells responsive either to the thymic extract TP:I or to IL2. To reduce the "background" of spontaneously E-rosetting cells,
Journal o f Chntcal Immunology, Vol 5, No 5, 1985
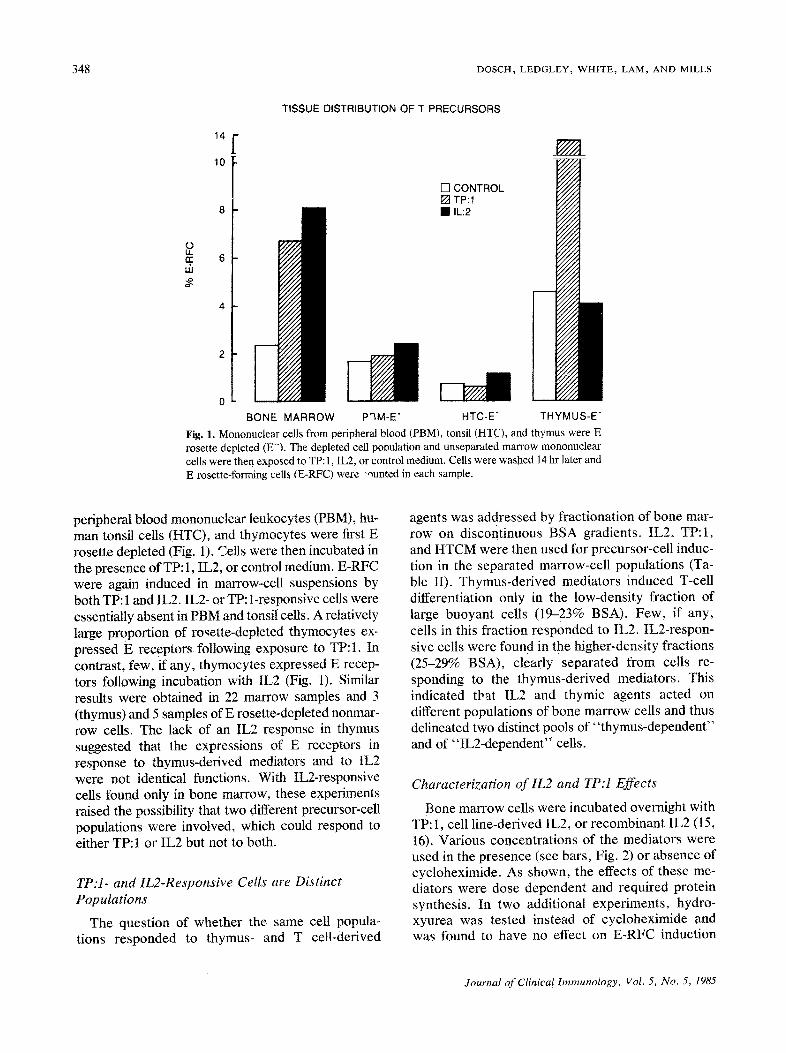
348 DOSCH, LEDGLEY, WHITE~ LAM, AND MILLS
TISSUE DISTRIBUTION OF T PRECURSORS
o t2.
uJ 2
14 r 10
[ ] CONTROL [ ] TP:I • IL:2
I BONE MARROW P'~M-E" HTC-E- THYMUS-E
Fig. 1. Mononuclear cells from peripheral blood (PBM), tonsil (HTC), and thymus were E rosette depleted ( E ) , The depleted cell pooulation and unseparated marrow mononuclear cells were then exposed to TP: 1, IL2, or control medium. Cells were washed 14 hr later and E rosette-forming cells (E-RFC) were -ounted in each sample.
peripheral blood mononuclear leukocytes (PBM), hu- man tonsil cells (HTC), and thymocytes were first E rosette depleted (Fig. 1). Cells were then incubated in the presence of TP: 1, IL2, or control medium. E-RFC were again induced in marrow-cell suspensions by both TP: 1 and IL2. IL2- or TP: 1-responsive cells were essentially absent in PBM and tonsil cells. A relatively large ProPortion of rosette-depleted thymocytes ex- pressed E recePtors foUowing exposure to TP: 1. In contrast, few, if any, thymocytes expressed E recep- tors following incubation with IL2 (Fig. 1). Similar results were obtained in 22 marrow samples and 3 (thymus) and 5 samples of E rosette-depleted nonmar- row cells. The lack of an IL2 response in thymus suggested that the expressions of E receptors in response to thymus-derived mediators and to IL2 were not identical functions. With IL2-responsive cells found only in bone marrow, these experiments raised the possibility that two different precursor-cell populations were involved, which could respond to either TP: 1 or IL2 but not to both.
TP:I- and IL2-Responsive Cells are Distinct Populations
The question of whether the same cell popula- tions responded to thymus- and T cell-derived
agents was addressed by fractionation of bone mar- row on discontinuous BSA gradients. IL2, TP: 1, and HTCM were then used for precursor-cell induc- tion in the separated marrow-cell populations (Ta- ble II). Thymus-derived mediators induced T-cell differentiation only in the low-density fraction of large buoyant cells (1%23% BSA). Few, if any, cells in this fraction responded to IL2. IL2-respon- sive cells were found in the higher-density fractions (25-29% BSA), clearly separated from cells re- sponding to the thymus-derived mediators. This indicated that IL2 and thymic agents acted on different populations of bone marrow cells and thus delineated two distinct pools of "thymus-dependent" and of "IL2-dependent" cells.
Characterization of IL2 and TP:I Effects
Bone marrow cells were incubated overnight with TP: 1, cell line-derived IL2, or recombinant IL2 (15, 16). Various concentrations of the mediators were used in the presence (see bars, Fig. 2) or absence of cycloheximide. As shown, the effects of these me- diators were dose dependent and required protein synthesis. In two additional experiments, hydro- xyurea was tested instead of cycloheximide and was found to have no effect on E-RFC induction
Journal of Clinical Immunology, Vol. 5, No. 5, 1985
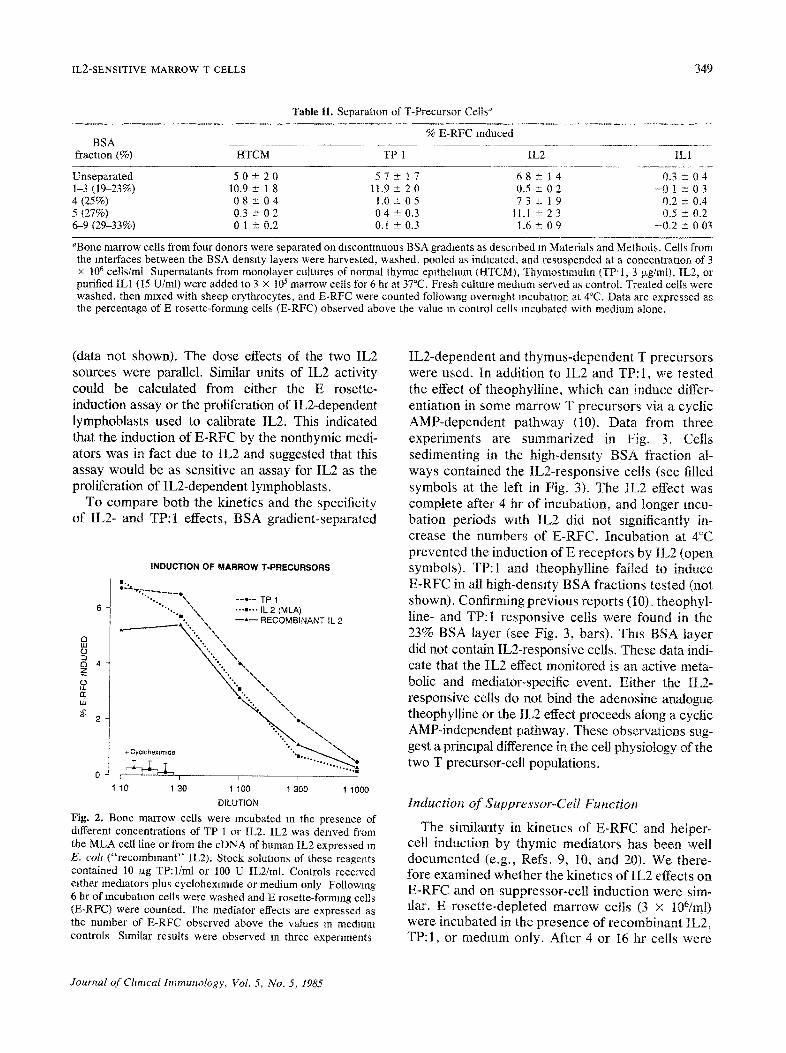
IL2-SENSITIVE MARROW T CELLS 349
Table II. Separatmn of T-Precursor Cells ~
% E-RFC Induced BSA
fraction (%) HTCM TP 1 IL2 ILl
Unseparated 5 0 + - 2 0 57-+ 17 68-+ 14 0 . 3 - + 0 4 1-3 (1%23%) 10.9 ± l 8 11.9 -+ 2 0 0.5 -+ 0 2 - 0 1 ± 0 3 4(25%) 08 +- 04 1.0± 05 73 ± 1 9 0.2 ± 0.4 5 (27%) 0.3 ± 0 2 0 4 ± 0.3 11.1 -+ 2 3 0.5 - 0.2 6-9 (2%33%) 0 1 ± 0.2 0. t -+ 0.3 1.6 ± 0 9 -0 .2 4- 0 03
~Bone marrow cells from four donors were separated on discontinuous BSA gradients as described m Materials and Methods. Cells from the interfaces between the BSA density layers were harvested, washed, pooled as indicated, and resuspended at a concentration of 3 x 106 cells/ml Supernatants from monolayer cultures of normal thymlc epithehum (HTCM), Thymostlmuhn (TP-t, 3 ~g/ml), IL2, or purified ILl (15 U/ml) were added to 3 x 105 marrow cells for 6 hr at 37°C• Fresh culture medium served as control. Treated cells were washed, then mixed with sheep erythrocytes, and E-RFC were counted following overnight incubation at 4°C. Data are expressed as the percentage of E rosette-forming cells (E-RFC) observed above the value in control cells incubated with medium alone.
(data not shown). The dose effects of the two IL2 sources were parallel. Similar units of IL2 activity could be calculated from either the E rosette- induction assay or the proliferation of IL2-dependent lymphoblasts used to calibrate IL2. This indicated that the induction of E-RFC by the nonthymic medi- ators was in fact due to IL2 and suggested that this assay would be as sensitive an assay for IL2 as the proliferation of IL2-dependent lymphoblasts.
To compare both the kinetics and the specificity of IL2- and TP:I effects, BSA gradient-separated
Q IIJ o
4 z O tL
i11 g
2
INDUCTION OF MARROW T-PRECURSORS
ira.,
"'. ",. - - * ' - TP I • .. ,, ...m... IL 2 (MLA)
- - ~ ' ~ RECOMBINANT IL 2
%, %% %,. \%
%. 'o~ %° '%~
%. *-,~
"% "~..
+ Cyclohe×lrngde ' 1 1 ~ ~
0 ~ r @ ~ - F ~ ,
1 10 1 30 1 100 t 300 1 1000
DILUTION
Fig. 2. Bone marrow cells were incubated in the presence of different concentrations of TP 1 or IL2. IL2 was derived from the MLA cell line or from the cDNA of human IL2 expressed in E. colt ("recombinant" IL2). Stock solutions of these reagents contained 10 ~xg TP:I/ml or 100 U IL2/ml. Controls received rather mediators plus cyclohexlmlde or medium only Following 6 hr of incubation cells were washed and E rosette-forming cells (E-RFC) were counted. The mediator effects are expressed as the number o, c E-RFC observed above the values m medium controls S~milar results were observed In three experiments
IL2-dependent and thymus-dependent T precursors were used. In addition to IL2 and TP: 1, we tested the effect of theophylline, which can induce differ- entiatlon in some marrow T precursors via a cyclic AMP-dependent pathway (10). Data from three experiments are summarized in Fig. 3. Cells sedimenting in the high-density BSA fraction al- ways contained the IL2-responsive cells (see filled symbols at the left in Fig. 3). The IL2 effect was complete after 4 hr of incubation, and longer incu- bation periods with IL2 did not significantly in- crease the numbers of E-RFC. Incubation at 4°C prevented the induction of E receptors by IL2 (open symbols). TP:I and theophylline failed to induce E-RFC in all high-density BSA fractions tested (not shown). Confirming previous reports (10L theophyl- line- and TP:I responsive cells were found in the 23% BSA layer (see Fig. 3, bars). This BSA layer did not contain IL2-responsive cells. These data indi- cate that the IL2 effect monitored is an active meta- bolic and mediator-specific event. Either the IL2- responsive cells do not bind the adenosine analogue theophylline or the IL2 effect proceeds along a cyclic AMP-independent pathway. These observations sug- gest a principal difference in the cell physiology of the two T precursor-celt populations,
Induction of Suppressor-Cell Function
The similarity in kinetics of E-RFC and helper- cell induction by thymic mediators has been well documented (e.g., Refs. 9, 10, and 20). We there- fore examined whether the kinetics of IL2 effects on E-RFC and on suppressor-cell induction were sim- ilar. E rosette-depleted marrow cells (3 × 196/ml) were incubated in the presence of recombinant IL2, TP: 1, or medium only. After 4 or I6 hr cells were
Journal o f Chmcal Immunology, Vol. 5, No. 5, 1985
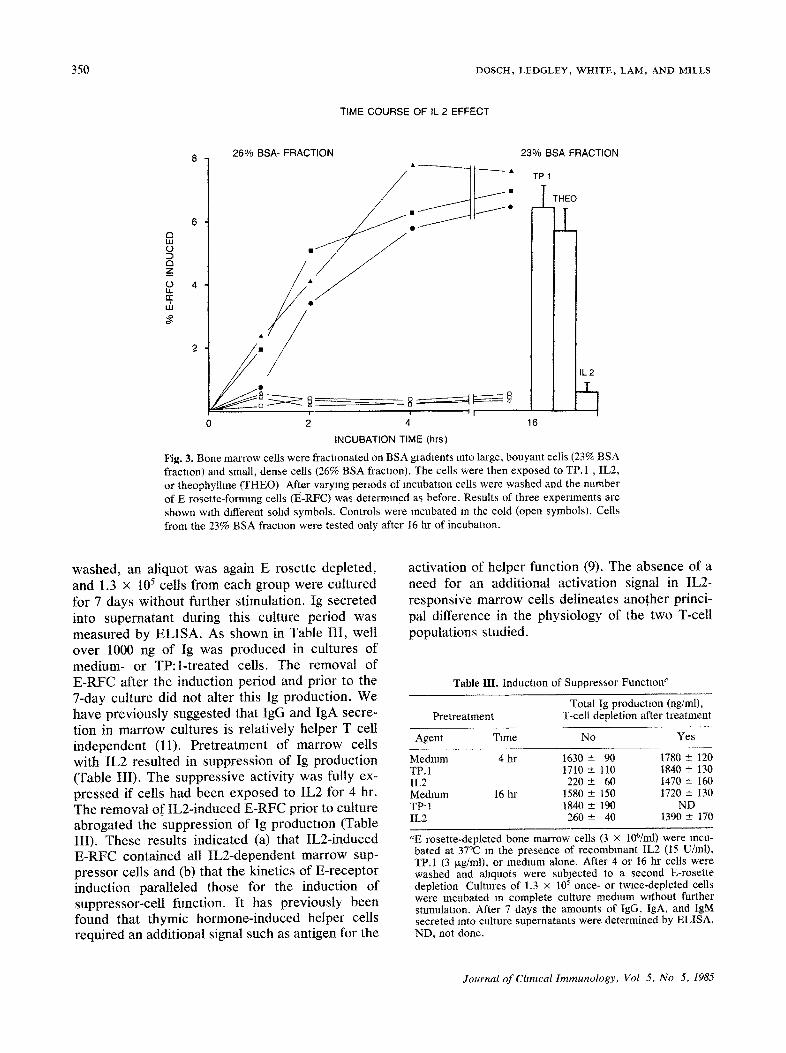
350 D O S C H , L E D G L E Y , W H I T E , L A M , A N D M I L L S
TIME COURSE OF IL 2 EFFECT
8 260/0 BSA- FRACTION 23% BSA FRACTION
THEO o
° l a w o •
_z O 4 • ,, ¢ / u5 * N
2
11_2
~ ~ ~ ~ _L]
J ~ II I 0 2 4 16
INCUBATION TIME (hrs)
Fig. 3. Bone marrow cells were fractlonated on BSA gradients into large, bouyant cells (23% BSA fraction) and small, dense cells (26% BSA fractaon). The cells were then exposed to TP. 1 , IL2, or theophylhne (THEO) After varying periods of mcubataon cells were washed and the number of E rosette-forming cells (t~-RFC) was determined as before. Results of three experiments are shown wath different sohd symbols, Controls were incubated m the cold (open symbols). Cells from the 23% BSA fractaon were tested only after 16 hr of incubation.
washed, an aliquot was again E rosette depleted, and 1.3 x 105 cells from each group were cultured for 7 days without further stimulation. Ig secreted into supernatant during this culture period was measured by ELISA. As shown in Table III, well over 1000 ng of Ig was produced in cultures of medium- or TP:l-treated cells. The removal of E-RFC after the induction period and prior to the 7-day culture did not alter this Ig production. We have previously suggested that IgG and IgA secre- tion in marrow cultures is relatively helper T cell independent (l 1). Pretreatment of marrow cells with IL2 resulted in suppression of Ig production (Table III). The suppressive activity was fully ex- pressed if cells had been exposed to IL2 for 4 hr. The removal of IL2-induced E-RFC prior to culture abrogated the suppression of Ig production (Table III). These results indicated (a) that IL2-induced E-RFC contained all IL2-dependent marrow sup- pressor cells and (b) that the kinetics of E-receptor induction paralleled those for the induction of suppressor-ceU function. It has previously been found that thymic hormone-induced helper cells required an additional signal such as antigen for the
activation of helper function (9). The absence of a need for an additional activation signal in IL2- responsive marrow cells delineates another princi- pal difference in the physiology of the two T-ceU populations studied.
Table III. Induction of Suppressor Function ~
Total Ig production (ng/ml), Pretreatment T-cell depletion after treatment
Agent Time No Yes
Medmm 4 hr 1630 -+ 90 1780 - 120 TP.1 1710 +- 110 1840 +- 130 IL2 220 -+ 60 1470 -+ 160 Medmm 16 hr 1580 -+ 150 1720 -+ 130 TP't 1840 -+ 190 ND IL2 260 +- 40 1390 +- 170
aE rosette-depleted bone marrow cells (3 × 106/ml) were incu- bated at 37°C m the presence of recombinant IL2 (t5 U/ml), TP.1 (3 txg/ml), or medium alone. After 4 or 16 hr cells were washed and ahquots were subjected to a second E-rosette depletion Cultures of 1.3 x 105 once- or twice-depleted cells were incubated in complete culture medmm without further stimulation. After 7 days the amounts of IgG, IgA, and IgM secreted into culture supernatants were determined by ELISA. ND, not done.
Journal of Chntcal Immunology, Vol 5, No 5, 1985
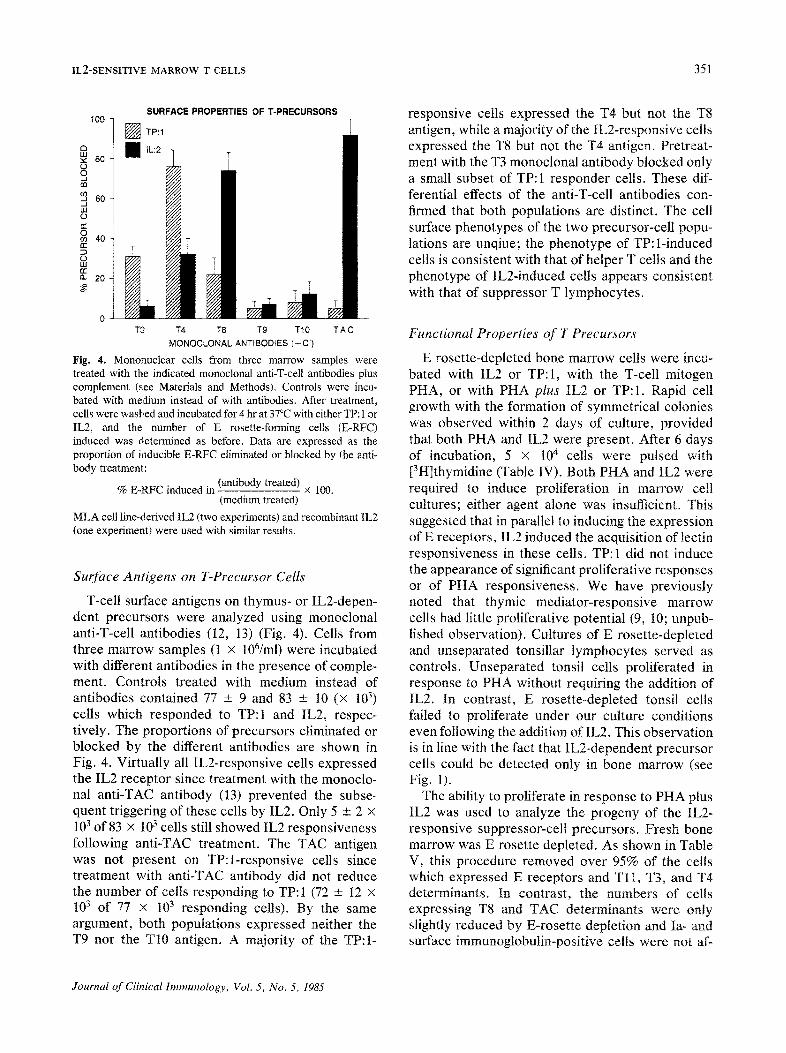
IL2-SENSITIVE MARROW T CELLS 351
SURFACE PROPERTIES OF T-PRECURSORS 100 N TP:I l
_~ 80 l IL:2 X T [ ]
m n .o • •
m m o )
40
20
T3 T4 T8 T9 TIO TAG MONOCLONAL ANTIBODIES (+ C')
Fig. 4. Mononuclear cells from three marrow samples were treated with the indicated monoclonal anti-T-cell antibodies plus complement (see Materials and Methods). Controls were incu- bated with medium instead of with antibodies. After treatment, cells were washed and incubated for 4 hr at 37°C with either TP: 1 or IL2, and the number of E rosette-forming cells (E-RFC) induced was determined as before. Data are expressed as the proportion of inducible E-RFC eliminated or blocked by the anti- body treatment:
% E-RFC induced in (antibody treated) x 100. (medium treated)
MLA cell line-derived IL2 (two experiments) and recombinant IL2 (one experiment) were used with similar results.
Surface Antigens on T-Precursor Cells
T-cell surface antigens on thymus- or IL2-depen- dent precursors were analyzed using monoclonal anti-T-cell antibodies (12, 13) (Fig. 4). Cells from three marrow samples (1 x 106/ml) were incubated with different antibodies in the presence of comple- ment. Controls treated with medium instead of antibodies contained 77 + 9 and 83 -+ 10 (× 103) cells which responded to TP:I and IL2, respec- tively. The proportions of precursors eliminated or blocked by the different antibodies are shown in Fig. 4. Virtually all IL2-responsive cells expressed the IL2 receptor since treatment with the monocto- nal anti-TAC antibody (13) prevented the subse- quent triggering of these cells by IL2. Only 5 -+ 2 x 103 of 83 × 103 cells still showed IL2 responsiveness following anti-TAC treatment. The TAC antigen was not present on TP:l-responsive cells since treatment with anti-TAC antibody did not reduce the number of cells responding to TP: 1 (72 + 12 × 103 of 77 x 103 responding cells). By the same argument, both populations expressed neither the T9 nor the T10 antigen. A majority of the TP:I-
responsive cells expressed the T4 but not the T8 antigen, while a majority of the IL2-responsive cells expressed the T8 but not the T4 antigen. Pretreat- ment with the T3 monoclonal antibody blocked only a small subset of TP: 1 responder cells. These dif- ferential effects of the anti-T-cell antibodies con- firmed that both populations are distinct. The cell surface phenotypes of the two precursor-cell popu- lations are unqiue; the phenotype of TP:l-induced cells is consistent with that of helper T cells and the phenotype of IL2-induced cells appears consistent with that of suppressor T lymphocytes.
Functional Properties of T Precursors
E rosette-depleted bone marrow cells were incu- bated with IL2 or TP:I, with the T-cell mitogen PHA, or with PHA plus IL2 or TP: I. Rapid cell growth with the formation of symmetrical colonies was observed within 2 days of culture, provided that both PHA and IL2 were present. After 6 days of incubation, 5 × l0 4 cells were pulsed with [3H]thymidine (Table IV). Both PHA and IL2 were required to induce proliferation in marrow celt cultures; either agent alone was insufficient. This suggested that in parallel to inducing the expression of E receptors, IL2 induced the acquisition of lectin responsiveness in these cells. TP:I did not induce the appearance of significant proliferative responses or of PHA responsiveness. We have previously noted that thymic mediator-responsive marrow cells had little proliferative potential (9, 10; unpub- lished observation). Cultures of E rosette-depleted and unseparated tonsillar lymphocytes served as controls. Unseparated tonsil cells proliferated in response to PHA without requiring the addition of IL2. In contrast, E rosette-depleted tonsil cells failed to proliferate under our culture conditions even following the addition of IL2. This observation is in line with the fact that IL2-dependent precursor cells could be detected only in bone marrow (see Fig. 1).
The ability to proliferate in response to PHA plus IL2 was used to analyze the progeny of the IL2- responsive suppressor-cell precursors. Fresh bone marrow was E rosette depleted. As shown in Table V, this procedure removed over 95% of the cells which expressed E receptors and TII , T3, and T4 determinants. In contrast, the numbers of cells expressing T8 and TAC determinants were only slightly reduced by E-rosette depletion and Ia- and surface immunoglobulin-positive cells were not af-
Journal o f Clinical Immunology, Vol. 5, No. 5, 1985
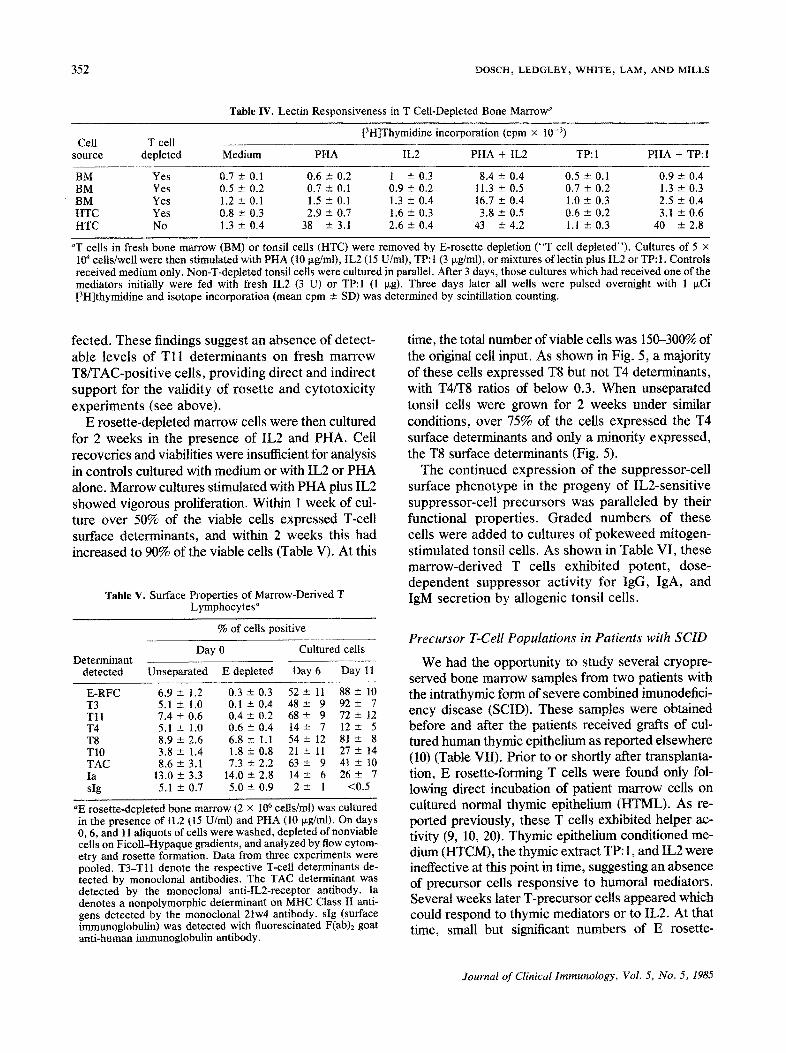
352 DOSCH, LEDGLEY, WI-IITE~ LAM, AND MILLS
Table IV. Lectin Responsiveness in T Cell-Depleted Bone Marrow ~
[3HlThymidine incorporation (cpm × 10 -3) Cell T cell
source depleted Medium PHA IL2 PHA + IL2 TP:I PHA + TP: 1
BM Yes 0.7 ± 0.1 0.6 -+ 0.2 t -+ 0.3 8.4 ± 0.4 0.5 ± 0.1 0.9 ± 0.4 BM Yes 0.5 ± 0.2 0.7 -+ 0.1 0.9 ± 0.2 11.3 ± 0.5 0.7 --- 0.2 1.3 -+ 0.3 BM Yes 1.2 -+ 0.1 1.5 ± 0.1 1.3 ± 0.4 16.7 -+ 0.4 1.0 ± 0.3 2.5 ± 0.4 HTC Yes 0.8 -+ 0.3 2.9 + 0.7 1.6 --- 0.3 3.8 + 0.5 0.6 - 0.2 3.1 ± 0.6 HTC No 1.3 -+ 0.4 38 +- 3.1 2.6 ± 0.4 43 ± 4.2 1.1 --- 0.3 40 --- 2.8
aT cells in fresh bone marrow (BM) or tonsil cells (HTC) were removed by E-rosette depletion ("T cell depleted"). Cultures of 5 × 104 cells/well were then stimulated with PHA (10 txg/ml), tL2 (15 U/ml), TP: 1 (3 ~g/ml), or mixtures of lectin plus IL2 or TP:I. Controls received medium only. Non-T-depleted tonsil cells were cultured in parallel. After 3 days, those cultures which had received one of the mediators initially were fed with fresh IL2 (3 U) or TP:I (1 p.g). Three days later all wells were pulsed overnight with 1 I~Ci [3H]thymidine and isotope incorporation (mean cpm ± SD) was determined by scintillation counting.
fected. These findings suggest an absence of detect- able levels of T11 determinants on fresh marrow T8/TAC-positive cells, providing direct and indirect support for the validity of rosette and cytotoxicity experiments (see above).
E rosette-depleted marrow cells were then cultured for 2 weeks in the presence of IL2 and PHA. Cell recoveries and viabilities were insufficient for analysis in controls cultured with medium or with IL2 or PHA alone. Marrow cultures stimulated with PHA plus IL2 showed vigorous proliferation. Within ! week of cul- ture over 50% of the viable cells expressed T-cell surface determinants, and within 2 weeks this had increased to 90% of the viable cells (Table V). At this
Table V. Surface Properties of Marrow-Derived T Lymphocytes a
% of cells positive
Day 0 Cultured cells Determinant
detected Unseparated E depleted Day 6 Day 11
E-RFC 6.9--- 1.2 0 . 3 - 0 . 3 5 2 ± 11 88-+ 10 T3 5.1+- 1.0 0.1 ---0.4 4 8 ± 9 9 2 ± 7 T l l 7 . 4 ± 0 . 6 0 . 4 ± 0 . 2 6 8 ± 9 7 2 ± 12 T4 5.1 --- 1.0 0.6 ± 0 . 4 14+- 7 12-± 5 T8 8.9 ± 2,6 6.8 - 1.1 54 --- 12 81 - 8 T10 3.8 +-- 1.4 1.8 -+ 0.8 21 +-- 11 27 ± 14 TAC 8 . 6 - 3 . 1 7.3 ± 2 . 2 6 3 ± 9 4 1 ± I0 Ia 13.0± 3.3 1 4 . 0 ± 2 . 8 14± 6 2 6 ± 7 slg 5.1 ± 0.7 5.0 ± 0.9 2 ± 1 <0.5
aE rosette-depleted bone marrow (2 × 106 cells/ml) was cultured in the presence of IL2 (15 U/ml) and PHA (10 ixg/ml). On days 0, 6, and 11 aliquots of cells were washed, depleted of nonviable cells on Ficoll-Hypaque gradients, and analyzed by flow cytom- etry and rosette formation. Data from three experiments were pooled. T3-T11 denote the respective T-cell determinants de- tected by monoclonal antibodies. The TAC determinant was detected by the monoclonal anti-IL2-receptor antibody. Ia denotes a nonpolymorphic determinant on MHC Class II anti- gens detected by the monoclonal 21w4 antibody, slg (surface immunoglobulin) was detected with fluorescinated F(ab)2 goat anti-human immunoglobulin antibody.
time, the total number of viable cells was 150-300% of the original cell input. As shown in Fig. 5, a majority of these cells expressed T8 but not T4 determinants, with T4/T8 ratios of below 0.3. When unseparated tonsil cells were grown for 2 weeks under similar conditions, over 75% of the cells expressed the T4 surface determinants and only a minority expressed, the T8 surface determinants (Fig. 5).
The continued expression of the suppressor-cell surface phenotype in the progeny of IL2-sensitive suppressor-cell precursors was paralleled by their functional properties. Graded numbers of these cells were added to cultures of pokeweed mitogen- stimulated tonsil cells. As shown in Table VI, these marrow-derived T cells exhibited potent, dose- dependent suppressor activity for IgG, IgA, and IgM secretion by allogenic tonsil cells.
Precursor T-Cell Populations in Patients with SCID
We had the opportunity to study several cryopre- served bone marrow samples from two patients with the intrathymic form of severe combined imunodefici- ency disease (SCID). These samples were obtained before and after the patients received grafts of cul- tured human thymic epithelium as reported elsewhere (10) (Table VII). Prior to or shortly after transplanta- tion, E rosette-forming T cells were found only fol- lowing direct incubation of patient marrow cells on cultured normal thymic epithelium (HTML). As re- ported previously, these T cells exhibited helper ac- tivity (9, 10, 20). Thymic epithelium conditioned me- dium (HTCM), the thymic extract TP: 1, and IL2 were ineffective at this point in time, suggesting an absence of precursor cells responsive to humoral mediators. Several weeks later T-precursor cells appeared which could respond to thymic mediators or to IL2. At that time, small but significant numbers of E rosette-
Journal of Clinical Immunology, Vol. 5, No. 5, 1985
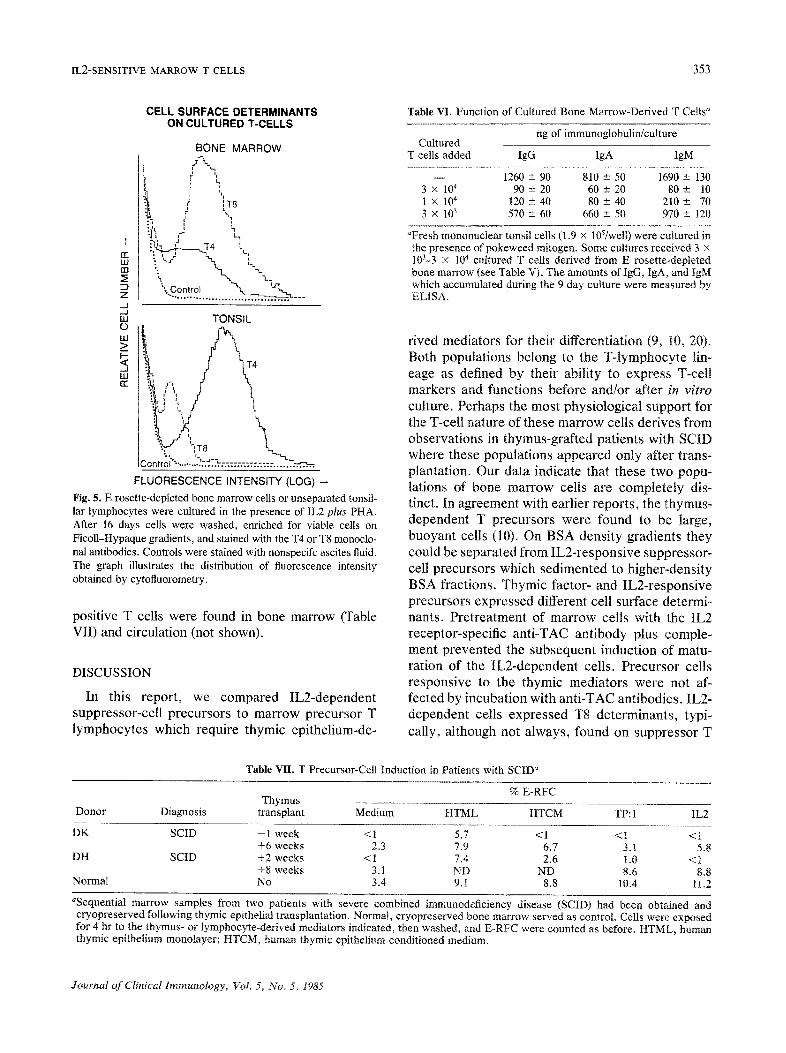
IL2-SENSITIVE MARROW T CELLS 353
CELL SURFACE DETERMINANTS ON CULTURED T.CELLS
BONE MARROW
f \ I '1 r
i
i '1 i d I T8
r
:hi'+ ~ 14 ~_
m ": ' " ~ - n _ t ~ r ? "q" ~' l ) iXl ~ i LLI : "~ -.," L ~ -t..+.
D o __ %.,... z ._,I ,._I w TONSIL
ILl > : 1 - - -~
< 4 . . J 'ILl ¢c c;
2 L ; , Co -.."::.:: :=.::
FLUORESCENCE INTENSITY (LOG) -
Fig. 5. E rosette-depleted bone marrow cells or unseparated tonsil- lar lymphocytes were cultured in the presence of IL2 plus PHA. After 16 days cells were washed, enriched for viable cells on Ficoll-Hypaque gradients, and stained with the T4 or T8 monoclo- nal antibodies. Controls were stained with nonspecffc ascites fluid. The graph illustrates the distribution of fluorescence intensity obtained by cytofluorometry.
positive T cells were found in bone marrow (Table VII) and circulation (not shown).
DISCUSSION
In this report, we compared IL2-dependent suppressor-cell precursors to marrow precursor T lymphocytes which require thymic epithelium-de-
Table VI, Function of Cultured Bone Marrow-Derived T Cells +
ng of immunoglobulin/culture Cultured
T cells added IgG IgA IgM
- - 1260 ± 90 810 ± 50 I690 ± 130 3 x 104 90 ± 20 60 ± 20 80 ± 10 1 x 104 120 ± 40 80 ± 40 210 ± 70 3 × 103 570 ± 60 660 ± 50 970 ± 120
aFresh mononuclear tonsil cells (1.9 x 105/well) were cultured in the presence of pokeweed mitogen. Some cultures received 3 x 10~-3 x 104 cultured T cells derived from E rosette-depleted bone marrow (see Tabte V). The amounts of IgG, IgA, and IgM which accumulated during the 9 day culture were measured by ELISA.
rived mediators for their differentiation (9, I0, 20). Both populations belong to the T-lymphocyte lin- eage as defined by their ability to express T-cell markers and functions before and/or after in vitro culture. Perhaps the most physiological support for the T-cell nature of these marrow cells derives from observations in thymus-grafted patients with SCID where these populations appeared only after trans- plantation. Our data indicate that these two popu- lations of bone marrow cells are completely dis- tinct. In agreement with earlier reports, the thymus- dependent T precursors were found to be large, buoyant cells (10). On BSA density gradients they could be separated from IL2-responsive suppressor- cell precursors which sedimented to higher-density BSA fractions. Thymic factor- and IL2-responsive precursors expressed different cell surface determi- nants. Pretreatment of marrow cells with the IL2 receptor-specific anti-TAC antibody plus comple- ment prevented the subsequent induction of matu- ration of the IL2-dependent cells. Precursor cells responsive to the thymic mediators were not af- fected by incubation with anti-TAC antibodies. IL2- dependent cells expressed T8 determinants, typi- cally, although not always, found on suppressor T
Table VII. T Precursor-Cell Induction in Patients with SCID +
% E-RFC Thymus
Donor Diagnosis transplant Medium HTML HTCM TP: 1 IL2
DK SCID - 1 week <1 5,7 <1 <1 <1 +6 weeks 2.3 7.9 6.7 3.1 5.8
DH SCID +2 weeks <I 7.4 2.6 t.0 < t +8 weeks 3.1 ND ND 8.6 8.8
Normal No 3.4 9.1 8.8 10.4 11.2
aSequential marrow samples from two patients with severe combined immunodeficiency disease (SCID) had been obtained and cryopreserved following thymic epithelial transplantation. Normal, cryopreserved bone marrow served as control. Cells were exposed for 4 hr to the thymus- or lymphocyte-derived mediators indicated, then washed, and E-RFC were counted as before. HTML, human thymic epithelium monolayer; HTCM, human thymic epithelium conditioned medium.
Journal of Clinical Immunology, Vol. 5, No. 5, 1985

354 DOSCH, LEDGLEY, WHITE, LAM, AND MILLS
cells (12, 22). In contrast, most of the cells respon- sive to thymic factors expressed the T4 determi- nant. In both cases the mediator response was rapid and completed within hours. Only in the IL2-responsive cells could we detect the acquisition of a significant proliferative capacity following me- diator treatment. Because of insufficient cell recov- eries, direct side-to-side experiments with purified precursor-cell populations could not be performed. However, mutually supportive results were ob- tained in functional and patient studies, cell separa- tions, and phenotypic analysis in five experimental systems which monitored rosette formation, bind- ing of monoclonal antibodies, cytotoxicity, cell growth, and suppressor activity.
Thymus-dependent marrow T precursors have been recognized as a source of helper/inducer func- tion, and the T4 molecule itself may play a role in the expression of helper activity (9, 10, 23). In contrast, the IL2-responsive marrow cells de- scribed here exhibit a strong suppressor activity. The expression of the T8-positive suppressor-cell phenotype was stable in the progeny of these mar- row T cells and suppressor activity was still demon- strable following 16 days of culture in the presence of PHA and IL2. Some helper- and suppressor-celt precursors therefore appear functionally committed to their respective effector T-cell sublineage.
Each precursor-cell population represents 4-6% of the marrow cells, suggesting total precursor-cell pool sizes of up to over 1011 cells each in a normal adult (1). The large total size of these cell pools could suggest that bone marrow is a major reposi- tory for helper- and suppressor-cell precursors. If this interpretation is correct, it would then follow that at least later stages in the ontogeny of helper and suppressor cells are different and that, for the suppressor T-cell sublineage, differentiation is not directly thymus dependent. The origin and develop- mental history of the IL2-dependent suppressor precursors are unknown. We previously developed a sequential model of thymus-dependent T-cell dif- ferentiation by monitoring patients with severe combined immunodeficiency disease (SCID) who received and responded to thymic epithelial trans- plantation (10, 20, 28). If our observations of the sequential appearance of T-precursor populations following thymus transplantation in patients with SCID were representative of normal T-cell ontogeny, then the IL2-dependent T precursors found in bone marrow would appear to be a cell population which requires the presence either of
functional thymic epithelium or of thymic epithe- lium processed T lymphocytes (27) for their devel- opment. In analogy to the recent demonstration of both IL2-dependent and IL2-independent T-precur- sor cells in nude mice, we presently favor the latter possibility (reviewed in Ref. 17). These animal studies suggest a two-tiered model of T-cell differ- entiation in which thymus-processed T cells are directly involved in the development of IL2-depen- dent precursor T-cell pools.
Our studies of IL2-dependent marrow T cells raise two questions: (a) Does IL2 induce the differ- entiation of an undifferentiated marrow-cell popula- tion or does it activate a thymus-processed cell which recirculated to the bone marrow? and (b) Does the large precursor-cell pool size reflect a major role in immune homeostasis? The expression of IL2 receptors has generally been considered a postactivation event in cells positive for the T-cell differentiation antigens T3 and T11. T-cell activa- tion is often associated with the expression of Ia determinants, increases in cell volume, and Ca 2+ flux (24). IL2-responsive marrow T precursors were small cells, and Ia, Tt I, and T3 determinants were not detectable. In preliminary experiments, E-RFC induction was not impaired by the removal of external Ca 2+, which prevents transmembrane Ca 2+ flux. In mature T cells, IL2 usually acts as a second signal in concert with another (mitogenic or antigenic) stimulus (25). In the population studied here, however, IL2 directly induced the expression of new surface determinants and new functional properties, without an apparent requirement for additional activation signals. These parameters dis- tinguish the IL2 response in marrow cells from typical IL2 responses in mature T lymphocytes. Perhaps these events are similar to differentiation events observed in nude mice following treatment with IL2 (17, 26, 27). We have used the expression of E-receptor (TI 1) determinants to monitor the IL2 effect. This assay has previously served as a marker of T-cell differentiation, rather than activation events (9, 10). These arguments are indirect, but we presently favor the view that IL2 can induce precursor-cell differentiation in a population of mar- row resident T-lineage cells.
The in vivo role of IL2-dependent T-precursor cells is not known. These cells might be exported from the bone marrow to replenish peripheral ef- lector T-cell pools. Also, IL2-dependent suppressor cells could function within the marrow environment itself and regulate immunoglobulin production in
Journal of Clinical Immunology, Vol. 5, No. 5, 1985

IL2-SENSITIVE MARROW T CELLS 355
the large marrow B-cell compartment (I-5, 7, 11, 29). Bone marrow is a surprisingly good source of IL2, and the IL2 quantities produced by (lectin-) stimulated marrow cells are smilar to the quantities added in the experiments reported here (I I). How- ever, it is unclear whether inducers of IL2 produc- tion (e.g., antigen) normally enter the marrow envi- ronment. A selective impairment of either T helper- or T suppressor-cell sublineages is observed in a growing number of diseases (e.g., Refs. 30-39). It seems attractive to examine whether blocks in the differentiation of thymic factor- or IL2-dependent marrow T-precursor cells would explain the selec- tivity of alterations in the function or number of effector T lymphocytes in some of t.hese conditions.
A C K N O W L E D G M E N T S
We wish to thank Drs. T. A. Waldman (NIH) and M. Letarte (Toronto), the Serono Laboratories (Randolph, MA) and the Biogen Corporation (Cam- bridge, MA), for their generous gifts of reagents and Dr. E. W. Gelfand for reading the manuscript. The Apple IIe ELISA software was a gift from Mr. J. Kurpias, Data Disc Services, Toronto. This work was supported by grants from the Medical Research Council, the National Cancer Institute of Canada, and Physicians Services Inc., of Ontario. GBM is an Alberta Heritage Trust Foundation Fellow.
R E F E R E N C E S
1. McMfllan R, Longmlre RL, Long R. Heath JE, Craddock CG: Immunoglobuhn synthesis by human lymphoid tissues: Normal bone marrow as a major site of IgG production J Immunol t09 1386-1394, 1972
2. Benner R, H1jmans W, Haaljman JJ: The bone marrow as the major source of serum lmmunoglobuhn, but stall a neglected site of antibody formation Chn Exp Immunol 46 1-8, t981
3. Ryser JE, Dutton RW: The humoral immune response of mouse bone marrow lymphocytes In vitro. Immunology 32:811-8t7, 1977
4. Askonas BA. White RG. Formation of specific antibodies and 7-globuhn. A study of the synthetic abdlty of various t~ssues from rabbits lmmumzed by different methods Blochem J 68 252-261. 1958
5. MacDermott RP. Beale MG, Alley CD, Nash GS, Bertovich MJ, Bragdon MJ: Synthesis and secretion of IgA, IgM, and IgG by peripheral blood mononuclear cells in human disease states, by ~solated human Intestinal mononuclear cells, and by human bone marrow mononuclear cells from ribs. NY Acad Scl 409:498-509, 1983
6 Dosch HM, Gelfand EW Generation of human plaque forming cells in culture Tissue distribution, antlgemc and cellular requirements J Immunol 118.302-308, 1977
7. deGast G, Platts-MlllS TAE Functional studies on lympho- cytes in adult human bone marrow, i. Ig production in vitro after fractlonation on a sucrose gradient and T/~aon-T ceil separation J Immunol 122.280-284, 1979
8 Siegel RL, Issekutz T, Schwaber J, Rosen FS. Geha RS: Deficiency of T helper cells in transient hypogammaglobuh- nemla of infancy N Engl J Med 305:1307-t3t3, 1981
9. Gelfand EW, Dosch HM: In vitro functional heterogeneity of humoral and cellular immune-deficiency state~,. In Anti- body Formation m Man, AS Faucl. R Balheux (eds). New York, Academic Press, 1979, pp 30%324
10. Gelfand EW, Dosch HM. Shore A. Llmatlbul S, Lee JWW The role of thymus in human T cell differentlatxon. In The Biological Basis of Immunodeficlency, EW Gelfand, HM Dosch (eds). New York. Raven Press, 1980, pp 39-56
11 Mdls GB, Ledgley C J, Hlbl T, White D, Lam P, Dosch HM Lymphocyte function m human bone marrow. I. Character- ization of two T cell populations regulating lmmuaoglobuhn secretion. J Immuno! 134"3036-3041. 1985
12. Bach MA. Bach JF' The use of monoclonal antlbodles to study T cell imbalances in human disease Clin Exp Immunol 45'449-456, 1981
13. Depper JM. Leonard WJ, Robb RJ. Waldman TA, Greene WC: Blockade of the lnterleukm 2 receptor by anti-TAC antibody: Inhibition of human lymphocyte activatxon J Immunol 131 690-696, 1983
14. Falchettl R, Bergesl G, Eshkol A, Cafiero C, Adonm L, Caprmo L. Pharmacological and biological properties of a calf thymus extract (TP-1). Drugs Exp Chn Res 3.39-42, 1977
15. Henderson LE, Hewetson JF. Hopkins RF III, Sowder RC. Newborne RH. Rabm H: A rapid, large scale purification for Gibbon Interleukm 2 J Immunol 131.810-815. 1983
16. Devos RG, Plaetnik H, Cherontre G, Simmons J, Tavernier E, Remont M, Flers W. Molecular cloning of human inter- leukin 2 cDNA and its expression In E coh. Nucleic Acid Res 11:4307-4319, 1983
17. Dosch HM, White D, Grant C: Reconstltution of nude mouse T cell function m vwo. IL2 independent effect of human T cells. J tmmunot 134.336-342, t984
18 Amento EP, Kurnlk JT, Epstein A, Krane SM. Modulation of synovlal cell products by a factor from a human cefl line T lymphocyte reduction of a mononuclear cell factor Proc Natl Acad Sci USA 79:5307-5311, 1982
19. Gillis A" Interteukm biochemistry and biology Fed Proc 42:2635, 1983
20. Dosch HM. Lee JWW, Gelfand EW, Falk A. Severe com- bined lmmunodeficiency disease: A model of T-cell dysfunc- tion Chn Exp Immunot 34"260-261, 1978
21. Ledgley CJ, Dosch HM. Bone marrow is a highly regulated, major site of lmmunoglobutin production in man Paedlat Res 17 255. 1983
22 Slegal FP, Rambatti P, S~egal M, Lopez C, Smith M, Davies TF, Osband M. Estren S: Helper cell functions of leukemic Leu-2a +, histamine receptor + T~ lymphocytes J Immunol 129:1775-1781, 1982
23. Rogozmskl L, Bass A, Ghckman E, Talle MA, Goldstem G. Wang J. Chen L, Thomas Y The T4 surface antigen is revolved in the induction of helper function J Immunol 132"735-739, 1984
24 Cheung RK, Grmstein S, Gelfand EW" Permissive role of
Journal o f Chmcal Immunology, Vol. 5, No 5, 1985

356 DOSCH, LEDGLEY, WHITE, LAM, AND MILLS
calcmm in the inh~bitlon of T celt mltogenesis by calmoduhn antagomsts. J Immunol 131.229t-2295, 1983
25 Paetkau V, Bleackley RC, Rlendeau D, Harmsh DG. HolowachukEW: Toward the molecular bmlogyofIL2 Mol Immunol (in press). 1985
26. Humg T: T cell function and specificity m athymlc mice. Immunol Today 4:84-87, 1984
27 Lipsick JS, Seruman L, Sato VL, Kaplan NO. Dlfferentm- tion and actlvatlon of nu/nu splemc T ceil precursors by mature peripheral T cells m the absence of thymus J Immunol 129:40--45, 1982
28. Gelfand EW, Dosch HM Differentiatmn of precursor T lymphocytes m man and dehneat~on of the selective abnor- mahtles m severe combined lmmunodefic~ency disease. Chn Immunol Immunopathol 25'303-315, 1982
29. Benner R, van Oudenanen A, Bjorklund M, Ivars F, Hohnberg D: Background lmmunoglobuhn' Measurement, bmlogical stgmficance and regulation Immunol Today 3:243-249, 1982
30. Dosch HM, Shore A: Hypothesis. The role oflnterleukms m lymphopoiesls--lmportant m autolmmune d~sease9 J Rheu- matol 9:353-358, 1982
31. Alcocer-Verela J, Alarcon-Segovla D' Decreased production of and response to lnterleukm 2 by cultured lymphocytes from pat,ents with systemic lupus erythematosus. J Chn Invest 69:1388-1395. 1982
32. Talal N, Dauphmee M. Christadoss P, Ferandes G, Russell
I J, Mlyasaka T- Interleukin-2 and autolmmune disease Adv Nephrol 12:239-246, 1983
33 Honta M, Suzuki H, Onodera T, Gmsberg-Fellner F, Faucl AS, Notkms A Abnormalities of lmmunoregulatory T cell subsets in patients with insulin dependent &abetes melhtus. J Immunol 129:1426-1429. 1982
34 Hoffman SL, Piessens WF, Rotiwanto S, Hussem PR, Kurmavan L' Reduction of suppressor T lymphocytes m the troptcal splenomegaly syndrome N Engl J Med 310 337- 341, 1984
35. Shearer G, Talal N A chmclan and a scientist look at the acqmred lmmunodeficlency syndrome (AIDS). Immunol To- day 4"180-185. 1983
36 Amman AJ, Abrams D, Conant M, Chudwm D, Cowan M, Volberdmg P" Acqmred immune dysfunction m homosexual men. Immunologic profles Chn Immunol Immunopathol 27 315-321. 1983
37. Tsang KY, Fudenberg HH, Galbralth GMP' In vitro aug- mentatlon of mterleukm 2 production and lymphocytes with the TAC antigen marker m patients with AIDS N Engl J Med 310"987, 1984
38 Dardenne M, Savino W, Gastmel LN, Nabarra B, Bach JF: Thym~c dysfunction m the mutant diabetic (db/db) mouse J Immunol 130:1195-1199, 1983
39. Dardenne M, Bach JF, Safal B: Low serum thymlc hormone levels m patients with acquired lmmunodeficlency syn- drome. N Engl J Med 309 48-49, 1983
Journal o f Chnical Immunology, Vol 5, No. 5. 1985




















