
Low-cost fermentation medium for alkaline protease production by Bacillus mojavensis A21 using...
7
Low-cost fermentation medium for alkaline protease production by Bacillus mojavensis A21 using hulled grain of wheat and sardinella peptone Anissa Haddar, 1, ⁎ Nahed Fakhfakh-Zouari, 1 Noomen Hmidet, 1 Fakher Frikha, 2 Moncef Nasri, 1 and Alya Sellami Kamoun 1 Laboratoire de Génie Enzymatique et de Microbiologie-Ecole Nationale d'Ingénieurs de Sfax, B.P. 1173-3038 Sfax, Tunisia 1 and Laboratoire de Biochimie et de Génie Enzymatique des Lipases-Ecole Nationale d'Ingénieurs de Sfax, B.P. 1173-3038 Sfax, Tunisia 2 Received 13 January 2010; accepted 31 March 2010 Available online 10 May 2010 Media composition and culture conditions for surfactant stable alkaline protease production by Bacillus mojavensis A21 were optimized using two statistical methods. Plackett–Burman design was applied to find the optimal ingredients and conditions to improve yields. Response surface methodology (RSM), including central composite design, was used to determine the optimal concentrations and conditions. The results indicated that several components, including hulled grain of wheat (HGW), sardinella peptone (SP), NaCl, CaCl 2 , MgSO 4 ,K 2 HPO 4 , KH 2 PO 4 , agitation, culture temperature and initial medium pH, had significant effects on production. The statistical model was constructed via central composite design (CCD) using four selected variables (HGW, NaCl, KH 2 PO 4 and K 2 HPO 4 ). Under the proposed optimized conditions, the protease experimental yield (1860.63 U/mL) closely matched the yield predicted by the statistical model (1838.60 U/mL) with R 2 = 0.98. An overall 14.0-fold increase in protease production was achieved using the optimized medium (HGW 30.0 g/L, SP 1.0 g/L, NaCl 2.0 g/L, KH 2 PO 4 1.0 g/L, K 2 HPO 4 0.3 g/L, CaCl 2 2.0 g/L, MgSO 4 1.0 g/L and pH 9.0, compared with the unoptimized basal medium (starch 10.0 g/L, yeast extract 2.0 g/L, KH 2 PO 4 0.1 g/L, K 2 HPO 4 0.1 g/L, CaCl 2 0.5 g/L and pH 8.0; 137 U/mL). A successful and significant improvement (14-fold) in the production of protease by the A21 strain was accomplished using cheap carbon and nitrogen substrates (HGW and SP), which may result in a significant reduction in the cost of medium constituents. © 2010, The Society for Biotechnology, Japan. All rights reserved. [Key words: Media optimization; Bacillus mojavensis A21; Alkaline detergent proteases; Low-cost media] Microorganisms are vital sources for enzyme production. Among these enzymes, proteases account for nearly 60% of the total industrial enzyme market (1). A large proportion of commercially available proteases are currently derived from Bacillus strains. The use of alkaline proteases has increased remarkably in many industrial processes, including the production of detergents, food processing, in animal feed and in the production of X-ray films (2). It is well known that extracellular protease production by microorganisms is greatly influenced by the media components, especially the carbon and nitrogen sources, but also including additional factors such as the temperature, pH, incubation time, agitation and inoculum density (3). Because approximately 30–40% of the production cost in industrial enzyme production is due to medium growth, optimizing the medium composition is especially vital (4). Optimization of media compounds by the traditional “one-variable- at-a-time” strategy is the most frequently used operation in biotechnology (5). This strategy is extremely time consuming and expensive when a large number of variables need to be considered. Additionally, this method is unable to detect true optimal conditions including the interactions among different production factors. In recent years, the use of statistical approaches involving Plackett–Burman designs and response surface methodology (RSM) has gained momentum in medium optimization. These methods are also useful for understanding interactions among various physico- chemical parameters using a minimal number of experiments. The Plackett–Burman design allows for the screening of the main factors from a large number of variables, and this information can be retained in further optimization. RSM is a collection of statistical techniques that is useful for designing experiments, building models, evaluating the effects of different factors and searching for optimal conditions of studied factors for desirable responses (6). RSM has been successfully applied in many areas of biotechnology, such as α-amylase produc- tion (7) and protease production (8). Bacillus mojavensis A21 has been recently identified as a producer of extracellular bleaching-stable alkaline proteases; three detergent- stable alkaline serine-proteases were purified from the culture supernatant (9,10). In view of the promising applicability of the alkaline protease as a builder for detergents, there is interest in producing this protease in Journal of Bioscience and Bioengineering VOL. 110 No. 3, 288 – 294, 2010 www.elsevier.com/locate/jbiosc ⁎ Corresponding author. Tel.: +216 74 274 088; fax: +216 74 275 595. E-mail address: [email protected] (A. Haddar). 1389-1723/$ - see front matter © 2010, The Society for Biotechnology, Japan. All rights reserved. doi:10.1016/j.jbiosc.2010.03.015
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Low-cost fermentation medium for alkaline protease production by Bacillus mojavensis A21 using...

Journal of Bioscience and BioengineeringVOL. 110 No. 3, 288–294, 2010
www.elsevier.com/locate/jbiosc
Low-cost fermentation medium for alkaline protease production by Bacillusmojavensis A21 using hulled grain of wheat and sardinella peptone
Anissa Haddar,1,⁎ Nahed Fakhfakh-Zouari,1 Noomen Hmidet,1 Fakher Frikha,2
Moncef Nasri,1 and Alya Sellami Kamoun1
⁎ CorrespondE-mail add
1389-1723/$doi:10.1016/
Laboratoire de Génie Enzymatique et de Microbiologie-Ecole Nationale d'Ingénieurs de Sfax, B.P. 1173-3038 Sfax, Tunisia1 and Laboratoire de Biochimie etde Génie Enzymatique des Lipases-Ecole Nationale d'Ingénieurs de Sfax, B.P. 1173-3038 Sfax, Tunisia2
Received 13 January 2010; accepted 31 March 2010Available online 10 May 2010
Media composition and culture conditions for surfactant stable alkaline protease production by Bacillus mojavensis A21were optimized using two statistical methods. Plackett–Burman design was applied to find the optimal ingredients andconditions to improve yields. Response surfacemethodology (RSM), including central composite design, was used to determinethe optimal concentrations and conditions. The results indicated that several components, including hulled grain of wheat(HGW), sardinella peptone (SP), NaCl, CaCl2, MgSO4, K2HPO4, KH2PO4, agitation, culture temperature and initial medium pH,had significant effects on production. The statistical model was constructed via central composite design (CCD) using fourselected variables (HGW, NaCl, KH2PO4 and K2HPO4). Under the proposed optimized conditions, the protease experimental yield(1860.63 U/mL) closely matched the yield predicted by the statistical model (1838.60 U/mL) with R2=0.98. An overall 14.0-foldincrease in protease production was achieved using the optimized medium (HGW 30.0 g/L, SP 1.0 g/L, NaCl 2.0 g/L, KH2PO41.0 g/L, K2HPO4 0.3 g/L, CaCl2 2.0 g/L, MgSO4 1.0 g/L and pH 9.0, comparedwith the unoptimized basal medium (starch 10.0 g/L,yeast extract 2.0 g/L, KH2PO4 0.1 g/L, K2HPO4 0.1 g/L, CaCl2 0.5 g/L and pH 8.0; 137 U/mL). A successful and significantimprovement (14-fold) in the production of protease by the A21 strain was accomplished using cheap carbon and nitrogensubstrates (HGW and SP), which may result in a significant reduction in the cost of medium constituents.
© 2010, The Society for Biotechnology, Japan. All rights reserved.
[Key words: Media optimization; Bacillus mojavensis A21; Alkaline detergent proteases; Low-cost media]
Microorganisms are vital sources for enzyme production. Amongthese enzymes, proteases account for nearly 60% of the total industrialenzyme market (1). A large proportion of commercially availableproteases are currently derived from Bacillus strains.
The use of alkaline proteases has increased remarkably in manyindustrial processes, including the production of detergents, foodprocessing, in animal feed and in the production of X-ray films (2).
It is well known that extracellular protease production bymicroorganisms is greatly influenced by the media components,especially the carbon and nitrogen sources, but also includingadditional factors such as the temperature, pH, incubation time,agitation and inoculum density (3). Because approximately 30–40% ofthe production cost in industrial enzyme production is due tomediumgrowth, optimizing the medium composition is especially vital (4).Optimization of media compounds by the traditional “one-variable-at-a-time” strategy is the most frequently used operation inbiotechnology (5). This strategy is extremely time consuming andexpensive when a large number of variables need to be considered.
ing author. Tel.: +216 74 274 088; fax: +216 74 275 595.ress: [email protected] (A. Haddar).
- see front matter © 2010, The Society for Biotechnology, Japan. Allj.jbiosc.2010.03.015
Additionally, this method is unable to detect true optimal conditionsincluding the interactions among different production factors.
In recent years, the use of statistical approaches involvingPlackett–Burman designs and response surface methodology (RSM)has gained momentum in medium optimization. These methods arealso useful for understanding interactions among various physico-chemical parameters using a minimal number of experiments. ThePlackett–Burman design allows for the screening of the main factorsfrom a large number of variables, and this information can be retainedin further optimization. RSM is a collection of statistical techniquesthat is useful for designing experiments, building models, evaluatingthe effects of different factors and searching for optimal conditions ofstudied factors for desirable responses (6). RSM has been successfullyapplied in many areas of biotechnology, such as α-amylase produc-tion (7) and protease production (8).
Bacillus mojavensis A21 has been recently identified as a producerof extracellular bleaching-stable alkaline proteases; three detergent-stable alkaline serine-proteases were purified from the culturesupernatant (9,10).
In view of the promising applicability of the alkaline protease as abuilder for detergents, there is interest in producing this protease in
rights reserved.

TABLE 1. Experimental variables at different levels used for the production of proteasesby Bacillus mojavensis A21 using Plackett–Burman design.
Variables Units Symbol Levels
289VOL. 110, 2010 ALKALINE PROTEASE PRODUCTION BY BACILLUS MOJAVENSIS A21
high yields in a low-cost fermentation medium. Herein, we report thestatistical optimization of culture conditions for the enhancedproduction of detergent proteases from B. mojavensis A21.
codeLower (−1) Higher (+1)
Hulled grain of wheat g/l X1 5 20Sardinella peptone g/l X2 1 5CaCl2 g/l X3 0.5 2NaCl g/l X4 2 20MgSO4 g/l X5 0.1 1K2HPO4 g/l X6 0.1 1KH2PO4 g/l X7 0.1 1Temperature °C X8 30 37Initial medium pH — X9 7 9Speed of agitation rpm X10 150 250Inoculum size % X11 2 10
TABLE 2. Plackett–Burman experimental design with proteases production levels.
Trials X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X11 Proteolytic ac-tivity (U/ml)
1 +1 −1 +1 −1 −1 −1 +1 +1 +1 −1 +1 672.722 +1 +1 −1 +1 −1 −1 −1 +1 +1 +1 −1 752.523 −1 +1 +1 −1 +1 −1 −1 −1 +1 +1 +1 807.274 +1 −1 +1 +1 −1 +1 −1 −1 −1 +1 +1 1178.185 +1 +1 −1 +1 +1 −1 +1 −1 −1 −1 +1 240.906 +1 +1 +1 −1 +1 +1 −1 +1 −1 −1 −1 1385.457 −1 +1 +1 +1 −1 +1 +1 −1 +1 −1 −1 231.818 −1 −1 +1 +1 +1 −1 +1 +1 −1 +1 −1 350.99 −1 −1 −1 +1 +1 +1 −1 +1 +1 −1 +1 390.010 +1 −1 −1 −1 +1 +1 +1 −1 +1 +1 −1 1405.011 −1 +1 −1 −1 −1 +1 +1 +1 −1 +1 +1 240.912 −1 −1 −1 −1 −1 −1 −1 −1 −1 −1 −1 572.72
MATERIALS AND METHODS
Raw materials All microbiological media components were purchased fromBio-Rad, France. Mirabilis jalaba tuber powder (MJTP), shrimp wastes powder (SWP),feather meal (FM) and sardinella peptone (SP) were prepared in our laboratory. Hulledgrain of wheat (HGW) is a by-product of semolina factories that contains 50–60%starch, 8–12% proteins, 15% cellulose and 5% carbohydrates. To obtain MJTP, the rawmaterial was peeled, grinded and then dried at 80 °C for at least 5 h. The driedMJTPwasminced again to obtain a fine powder and then stored in glass bottles at roomtemperature. The MJTP contained 32.6±2% starch, 17.3±3% proteins, 20±2% ash anda low content of lipids (11). To obtain SWP, the raw material was boiled, pressed toremove water and fat, minced and then dried. The SWP contained high protein content(46–49%), relatively high ash and chitin contents (27–35% and 17–20%, respectively)and low lipid content (5–6.5%) (12). To obtain FM, chicken feathers were washedthoroughly with tap water, dried at 105 °C and then grinded. The FM contained 85.31%protein, 2% lipids and 0.69% ashes. SP was prepared according to Souissi et al. (13) usingBacillus licheniformis NH1 alkaline proteases for protein hydrolysis (degree ofhydrolysis 26%).
Microorganism B. mojavensis A21 producing a bleach-stable alkaline proteasewas isolated from a marine water sample. It was identified on the basis of 16 S rRNAgene sequencing (9).
Protease assay Protease activity was measured according to the method ofKembhavi et al. (4) using casein as a substrate. The enzyme (0.5mL), suitably diluted,was mixed with 0.5 mL of a 100-mM solution of Tris–HCl (pH 10.0) containing 1% (w/v) casein and incubated for 15 min at 60 °C. The reaction was quenched with theaddition of 0.5 mL of trichloroacetic acid (20%; w/v). The mixture was allowed to standat room temperature for 15 min and then centrifuged at 10,000×g for 15 min to removethe precipitate. The soluble fraction was estimated at 280 nm. A standard curve wasgenerated using solutions of 0–50 mg/L of tyrosine. One unit of protease activity wasdefined as the amount of enzyme required to liberate 1 µg of tyrosine permin under theexperimental conditions used.
Selection of the best carbon and nitrogen sources by one-variable-at-a-timeapproach Initial screenings of the most significant carbon and nitrogen sources tomaximize protease production were performed by the one-variable-at-a-timeapproach. Ten simple and complex carbon sources (10 g/L), including sucrose, glucose,maltose, lactose, fructose, potato starch, HGW, MJTP, SWP and FM, and seven differentnitrogen sources (2 g/L), including yeast extract, casein peptone, casein, soy peptone,pastone, SP, NH4Cl and (NH4)2SO4, were tested. Initial M1 medium consisted of thefollowing (g/L): starch, 10.0; yeast extract, 2.0; CaCl2, 0.5; KH2PO4, 0.1; and K2HPO4,0.1. These nitrogen and carbon sources are used in place of yeast extract and starch,respectively. Media were autoclaved at 120 °C for 20 min. Inocula were routinelygrown in Luria-Bertani (LB) broth medium, composed of the following (g/L): peptone,10; yeast extract, 5; and NaCl, 5 (15). Cultivations were conducted in 250-mLErlenmeyer flasks, with a working volume of 25 mL, and incubated at 37 °C on arotatory shaker (200 rpm) for 24 h. The cultures were centrifuged, and the cell-freesupernatants were used for the estimation of proteolytic activity. All experiments werecarried out in duplicate and repeated at least twice.
Identification of the significant variables using Plackett–Burman designs Us-ing Plackett–Burman design is an efficient way to screen the main physico-chemicalparameters among a large number of process variables (16), which is required forelevated protease production. The carbon and nitrogen sources that were previouslyscreened were added to the main culture medium to be optimized. The Plackett–Burmandesign allows for the evaluation of N variables in N+1 experiments; each variable wasexamined at two levels: –1 for a low level and+1 for a high level. The variables chosen forthe present study were HGW, SP, K2HPO4, KH2PO4, CaCl2, NaCl, MgSO4, incubationtemperature, speed of agitation, initial medium pH and inoculum size. Table 1 illustratesthe factors under investigation aswell as the levels of each factor used in the experimentaldesign. Table 2 represents the design matrix. “Design Expert® 7.0” Stat-Ease, Inc.,Minneapolis, USA, was used to analyze the experimental Plackett–Burman design.
Optimization by central composite design The next step in the optimizationof themedium required us to determine the optimal levels of the significant variables inalkaline protease production. To this purpose, response surface methodology (RSM)was adopted for the augmentation of total protease production using a centralcomposite design (CCD). The most significant variables examined were as follows:HGW, NaCl, KH2PO4 and K2HPO4 (Table 3), each of which was assessed at five codedlevels (–1.718, –1, 0, +1 and +1.718), as shown in Table 4.
A total of 30 experiments were conducted and repeated twice. All variables weretaken at a central coded value that was defined as zero. The minimum and maximumranges of the variables were used. The full experimental plans, with regard to the actualand coded values and the corresponding experimental and predicted responses values(Y), are provided in Table 5.
Statistical analysis and modeling The data obtained from the RSM on proteaseproduction were subjected to analysis of variance (ANOVA). Central composite designs
with the four factors (HGW, NaCl, KH2PO4 and K2HPO4) were used. After running theexperiments and measuring the activity levels, the experimental results of RSM werefitted with the response surface regression procedure using the following second-orderpolynomial equation:
Y = α0 + ∑iαiXi + ∑
iiαiiX2
i + ∑ijαijXiXj
In this equation, Y is the predicted proteolytic enzyme production (response), Xi
and Xj are the levels of the independent variables, α0 is the intercept term, αi is thelinear coefficient, αii is the quadratic coefficient and αij is the interaction coefficient.
The statistical significance of the model equation and the model terms wereevaluated via Fisher's test. The quality of fit of the second-order polynomial modelequation was expressed via the coefficient of determination, R2, and the adjusted R2.The fitted polynomial equation was then expressed as three-dimensional surface plotsto illustrate the relationship between the responses and the experimental levels of eachof the variables utilized in this study.
Validation of the experimental model The statistical model was validatedwith respect to all four variables within the design space. A random set of nineexperimental combinations was used to study protease production under theexperimental conditions described above.
Time course of protease production by B. mojavensis A21 To study therelation between protease production and the growth profile of the bacterium, 100 mLof the optimized and unoptimized production media was inoculated in 1-L flasks, andthe growth was measured at regular intervals by viable count (spread plate method)determination. The protease production at different time intervals was determinedusing the standard protease assay.
RESULTS
Selection of the most suitable carbon and nitrogen sources bythe one-variable-at-a-time approach A series of experimentswere carried out to study the effects of various simple and complexcarbon and nitrogen sources on protease production by B. mojavensisA21. Cultures were first conducted using medium M1, containingyeast extract (2 g/L) as nitrogen source. In addition, the different

TABLE 3. Identification of significant variables for proteases production by Bacillusmojavensis A21 using Plackett–Burman design.
Values of “ProbN |t|” less than 0.05 indicate model terms are significant.
TABLE 5. Experimental conditions in coded variables of the central composite designand the corresponding experimental responses.
Runorder
Coded values Proteolytic activity (U/ml)
X1 X4 X7 X6 Experimental Predicted
Y1 Y2
1 −1 −1 −1 −1 990.09 975.72 941.192 +1 −1 −1 −1 1720.00 1709.09 1742.273 −1 +1 −1 −1 1013.63 1010.36 978.564 +1 +1 −1 −1 1685.63 1695.09 1723.145 −1 −1 +1 −1 1185.54 1190.54 1210.326 +1 −1 +1 −1 1865.45 1860.63 1838.607 −1 +1 +1 −1 1280.36 1290.09 1251.718 +1 +1 +1 −1 1825.45 1820.09 1823.499 −1 −1 −1 +1 1289.09 1300.54 1288.9110 +1 −1 −1 +1 1784.54 1773.63 1802.8311 −1 +1 −1 +1 1210.09 1247.27 1243.5712 +1 +1 −1 +1 1731.18 1725.72 1700.9913 −1 −1 +1 +1 1429.09 1415.27 1379.8614 +1 −1 +1 +1 1690.36 1695.09 1720.9815 −1 +1 +1 +1 1372.72 1369.63 1338.5416 +1 +1 +1 +1 1596.36 1585.63 1623.1617 −1.718 0 0 0 667.27 656.36 743.2918 +1.718 0 0 0 1730.36 1745.45 1676.3719 0 −1.718 0 0 1728.09 1745.36 1736.7820 0 +1.718 0 0 1660.09 1669.81 1684.8321 0 0 −1.718 0 1474.54 1490.9 1480.8522 0 0 +1.718 0 1625.45 1621.81 1645.2623 0 0 0 −1.718 1517.27 1490.9 1525.9424 0 0 0 +1.718 1676.36 1632.72 1652.6225 0 0 0 0 1649.81 1645.54 1639.0026 0 0 0 0 1640.63 1638.54 1639.0027 0 0 0 0 1643.63 1648.36 1639.0028 0 0 0 0 1645.86 1647.54 1639.0029 0 0 0 0 1638.54 1640.09 1639.0030 0 0 0 0 1631.54 1639.63 1639.00
290 HADDAR ET AL. J. BIOSCI. BIOENG.,
carbon sources were added at a concentration of 10 g/L. B. mojavensisA21 produced a high level of protease in the presence of complexorganic carbon sources (data not shown). The maximum levels ofproteolytic activity were carried out with HGW (410 U/mL), followedby MJTP (291 U/mL), SWP (230 U/mL), starch (137 U/mL) and FM(82 U/mL). Easily assimilated simple carbon sources like fructose,lactose, glucose, sucrose and maltose resulted in weak proteaseproduction, 25, 18, 30, 25 and 34 U/mL, respectively.
In general, both organic and inorganic nitrogen sources were usedefficiently for protease production. In the present study, differentorganic (yeast extract, casein peptone, casein, soy peptone, pastoneand SP) and inorganic ((NH4)2SO4 and NH4Cl) nitrogen sources, at aconcentration of 2 g/L, were tested in M1medium containing HGW at10 g/L as the carbon source. Higher protease production levels wereobtained with SP (475 U/mL) compared to yeast extract, casein, soypeptone, casein peptone and pastone (404, 392, 384, 375 and 342 U/mL, respectively). Inorganic nitrogen sources ((NH4)2SO4 and NH4Cl)showed weak enzyme production levels, 61.4 and 34 U/mL, respec-tively (data not shown).
Among the various carbon and nitrogen sources tested, HGW andSP were the most suitable substrates for the production of proteolyticenzymes. Thus, these substrates were selected for further optimiza-tion steps.
Screening of significant variables using Plackett–Burmandesign Using Plackett–Burman design, a total of eleven variableswere analyzed to determine their effects on protease production(Table 1). The designed matrix was selected to screen the significantvariables for protease production, and the corresponding responsesare shown in Table 2. Statistical analyses of the responses wereperformed and represented in Table 3. Factors having a confidencegreater than 95% (ProbN |t|≤0.05) were considered to have asignificant effect on the protease production.
From the experimental data analysis, all factors tested havesignificant effects on the responses. HGW, CaCl2, MgSO4, K2HPO4,initial alkaline medium pH and speed of agitation exerted a positiveeffect, whereas the other variables, SP, NaCl, KH2PO4, temperature and
TABLE 4. Experimental codes, ranges and levels of the independent variables forresponse surface methodological experiment.
Variables Units Symbolcode
Levels
−1.718 −1 0 +1 +1.718
Hulled grain of wheat g/l X1 2.8 10 20 30 37.2NaCl g/l X4 0.56 2 4 6 7.44KH2PO4 g/l X7 0.048 0.3 0.65 1 1.252K2HPO4 g/l X6 0.048 0.3 0.65 1 1.252
inoculum size, exerted negative effects on protease production. HGW,NaCl, KH2PO4 and K2HPO4 were the most influential factors onenzymes production, as indicated by their t ratio. The optimal levels ofthe four selected variables (HGW, NaCl, KH2PO4 and K2HPO4) andtheir interactions were then examined by a central composite design.
Optimization of significant variables using RSM A total of 30experiments with different combinations of the four selectedvariables were performed (Table 5). The central composite design,employed to determine the optimum levels of the four screenedfactors, and the results of the second-order response surface modelfitting from variance (ANOVA) analysis are presented in Table 6.
The protease production (Y) by B. mojavensis A21 can be expressedin terms of the following regression equation:
Y = 1639:00 F9:61ð Þ + 271:43 F5:21ð Þ X1
–15:11 F5:21ð Þ X4 + 47:82 F5:21ð Þ X7 + 36:85 F5:21ð Þ X6
–14:12 F6:09ð Þ X1X4–43:20 F6:09ð Þ X1X7–71:79 F6:09ð Þ X1X6
+1:01 F6:09ð Þ X4X7–20:68 F6:09ð ÞX4X6–44:55 F6:09ð Þ X7X6
–145:26 F5:83ð Þ X21 + 24:30 F5:83ð Þ X2
4–25:71 F5:83ð Þ X27
–16:83 F5:83ð Þ X26
TABLE 6. Analysis of variance (ANOVA) for the quadratic model.
Source ofvariation
Sum ofsquares
Degreesof free-dom
Meansquare
F value p-valueProbNF
Significance
Regression 449,6442.625 14 321,174.473 270.320 b0.0001 ⁎
Residuals 53,465.692 45 1,188.126Cor total 4,549,908.317 59R2 0.988
⁎ Significant at the level 95%.

291VOL. 110, 2010 ALKALINE PROTEASE PRODUCTION BY BACILLUS MOJAVENSIS A21
X1, X4, X7 and X6 are HGW, NaCl, KH2PO4 and K2HPO4 concentra-tions, respectively.
The regression coefficients and the analysis of the variance(ANOVA) indicate the high significance of this model. For a goodstatistical model, the R2 value should be close to 1.0 and all of thefactors should be positive and close to each other. The high R2 value(0.988) showed the good agreement between the experimentalresults and the theoretical values predicted by the model (17). Thevalues of the adjusted determination coefficient (adjusted R2=0.985)and predicted determination coefficient (predicted R2=0.977) werealso high, advocating again for a high significance of this model (18).In addition, the model has an adequate precision value of 63.553; thissuggests that the model can be used to navigate the design space. Theprecision value is an index of the signal-to-noise ratio, and a value N4is an essential prerequisite for a model to be a good fit. A lower valueof the coefficient of variation (CV=2.27) showed that the experi-ments conducted were precise and reliable (19).
The three-dimensional (3-D) response surfaces presented in Fig. 1are generally the graphical representation of the regression equation,from which the protease production for different concentration of thevariable could be predicted. The responses are plotted on the Z-axisagainst two variables while other variables were maintained at levelzero. The model predicted that the maximum protease production of1830.60 U/mL appeared at (g/L): HGW, 30; NaCl, 2.0; KH2PO4, 1.0;K2HPO4, 0.3; CaCl2, 2.0; MgSO4, 1.0; SP, 1.0; and pH 9.0. Fig. 1A showsthe prominent effect of HGW on protease activity. Protease activityincreased continuously with an increasing concentration of HGW.Fig. 1B shows that a linear increase in protease secretionwas observedwhen HGW and KH2PO4 concentrations increased, and no concom-itant decline in enzyme production was observed. A similar profilewas observed in Fig. 1C with HGW and K2HPO4 concentrations.
Validation of the model The adequacy of the predicted modelwas verified by nine additional independent experiments. Theconditions of the experiment and predicted responses are representedin Table 7. The experimentally determined production values were inclose agreement with the statistically predicted ones, confirming themodel's authenticity. In fact, the multiple correlation coefficients (R2)and the coefficient of variation (CV) are 0.99 and 1.94, respectively.These denote that the correlation between the experimental and thepredicted values are good and that the experiments performed arehighly reliable. In addition, the average error (difference betweenobserved and predicted value) is close to zero, indicating the absenceof bias in the model's predictions.
Time course of protease production and cell growth during thefermentation The time course of protease activity and the growth ofB. mojavensis A21 for both the optimized (HGW 30.0 g/L, SP 1.0 g/L,NaCl 2.0 g/L, KH2PO4 1.0 g/L, K2HPO4 0.3 g/L, CaCl2 2.0 g/L, MgSO4
1.0 g/L and inoculum size 2.0% (v/v)) and unoptimized (starch 10.0 g/L, yeast extract 2.0 g/L, KH2PO4 0.1 g/L, K2HPO4 0.1 g/L, CaCl2 0.5 g/Land inoculum size 4.0% (v/v)) conditions are shown in Fig. 2. Theseresults show that the secretion of the proteases was associatedwith thecell growth. Protease activity increased during the exponential growthphase and reached activity of 1850 U/mL at 24 h. The production ofproteases increased slightly at the beginning of the stationary phase.However, the protease activity under unoptimized conditions was only135 U/mL after 24 h. By optimizing the medium composition and theculture conditions, the production of alkaline bleaching proteases by B.mojavensis A21 was enhanced from 135 to 1850 U/mL.
DISCUSSION
The A21 strain was found to produce multiple extracellularproteases through the casein-zymography (9). Crude A21 proteasewas highly stable and active at high pH values, showing optimalactivity at pH 8.0–11.0 and 60 °C. This crude enzyme is especially
tolerant and stable in the presence of the various tested laundrydetergents, making it particularly suitable for possible incorporationin liquid and solid laundry detergents.
The improvement ofmicrobial protease production is the purpose ofseveral investigations. In general, no defined medium has been carriedout for the production of alkaline proteases from different microorgan-isms; each strain has its specific required conditions for maximumenzyme production (20). The effect of environmental conditions on theproductionof extracellular proteolytic enzymes could play an importantrole in the induction or repression of the enzymeby specific compounds(21). Protease production is dependent on the availability of both thecarbon and nitrogen sources within the medium and both of theseparameters exert regulatory effects on enzyme synthesis (22). Manyresearchers have attempted to induce protease production by usingglucose or starch, coupledwith expensivenitrogen sources such as yeastextract, peptone or casamino acids. However, far fewer attempts havebeenmade to induce protease production using inexpensive carbon andnitrogen sources (3).
At present, the overall cost of enzyme production is very high dueto high cost of the substrates and mediums used. Therefore,development of novel process to increase the yield of proteaseswith respect to their industrial requirements, coupled with loweringthe production cost, is highly appreciable from the commercial pointof view. The industrial use of proteases, particularly alkalineproteases, is expected to grow tremendously in the coming decade.Therefore, microbial protease producing industries are always insearch of new and cheaper methods to enhance protease productionand decrease the market price of the desired enzyme. To this end,efforts have been directed to explore different means to reduceprotease production costs through improving the yield and usingeither cost-free or low-cost feed stocks or agricultural by-products assubstrate(s) for protease production (24,25).
In this study, complex carbon sources were excellent substratesfor protease production by B. mojavensis A21, and the proteolyticenzyme level was maximized in the presence of HGW. In addition,powders prepared from Mirabilis jalapa tubers and from shrimpwastes were good carbon sources for protease production. Howev-er, protease production was very low when simple carbon sources(i.e., glucose, sucrose, maltose and lactose) were used in the culturemedium. This result confirms earlier reports regarding proteaseproduction, in which it has been observed that these complexcarbon sources constitute better substrates for protease productionthan simple sugars, such as glucose, which can induce catabolicrepression (26). A similar catabolic repression mechanism forextracellular protease production has been described for Bacillus sp.(27). The A21 strain showed a high protease production on SP usedas nitrogen source. These findings are in line with several reportsshowing the enhancement of protease production in the presenceof organic nitrogen sources (2). Fish protein hydrolysate might alsobe used as a nitrogen source for metabolite production (28,29).
Thus, the use of HGW and SP as carbon and nitrogen sources mayresult in a more cost-effective process, and so the production ofdetergent proteases by the A21 strain has been effectively economized.
Recently, the use of statistical models to optimize culture mediumcomponents and conditions has increased due to its ready applicabilityandaccuracy. In thepresent study, the significant variablesnecessary forenhanced protease production were selected using the Plackett–Burman design. A large variation in protease production (232–1405 U/mL) from that mandated by the Plackett–Burman designexperiments suggested a need for further optimization. The RSMapplied to the optimization of protease production in this investigationsuggested the importance of a variety of factors at different levels. TheCCD design plan that we also examined in the present study enabled usto explore culture conditions that resulted in a 14-fold increase inprotease production. A high degree of similarity was observed between

IG. 1. Three-dimensional response surface plot for protease production showing the interactive effects of the HGW and NaCl concentrations (A), HGW and KH2PO4 concentrationsB), and HGW and K2HPO4 concentrations (C).
292 HADDAR ET AL. J. BIOSCI. BIOENG.,
F(
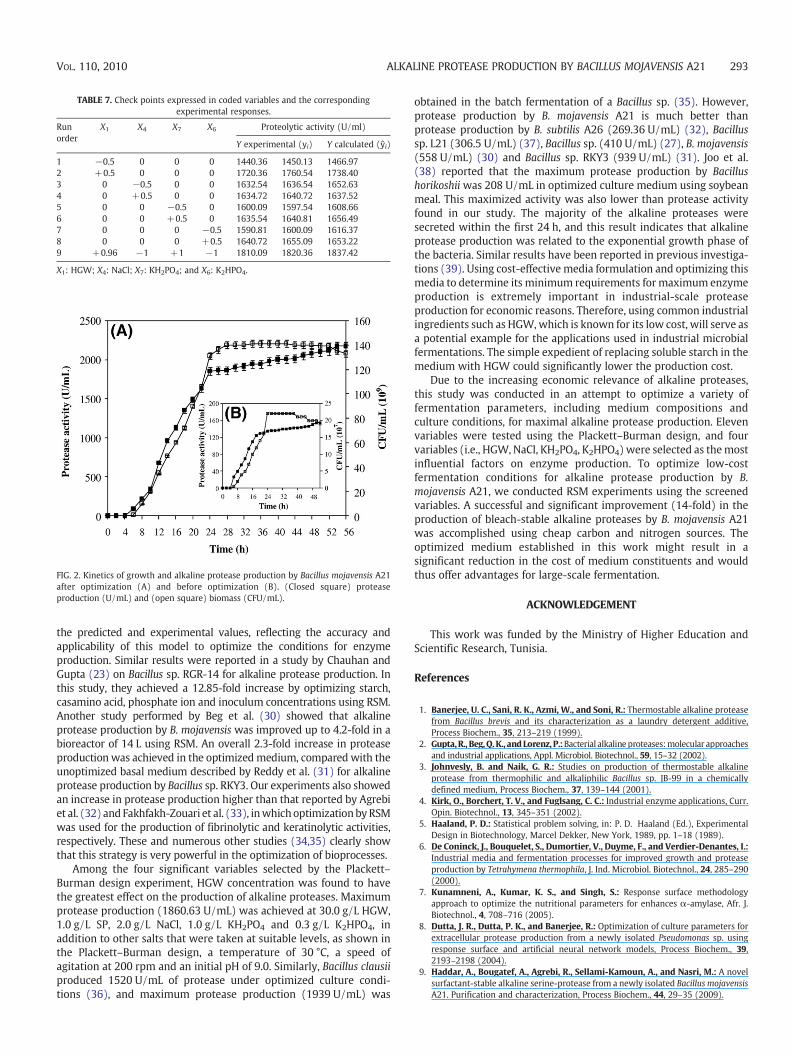
TABLE 7. Check points expressed in coded variables and the correspondingexperimental responses.
Runorder
X1 X4 X7 X6 Proteolytic activity (U/ml)
Y experimental (yi) Y calculated (ŷi)
1 −0.5 0 0 0 1440.36 1450.13 1466.972 +0.5 0 0 0 1720.36 1760.54 1738.403 0 −0.5 0 0 1632.54 1636.54 1652.634 0 +0.5 0 0 1634.72 1640.72 1637.525 0 0 −0.5 0 1600.09 1597.54 1608.666 0 0 +0.5 0 1635.54 1640.81 1656.497 0 0 0 −0.5 1590.81 1600.09 1616.378 0 0 0 +0.5 1640.72 1655.09 1653.229 +0.96 −1 +1 −1 1810.09 1820.36 1837.42
X1: HGW; X4: NaCl; X7: KH2PO4; and X6: K2HPO4.
FIG. 2. Kinetics of growth and alkaline protease production by Bacillus mojavensis A21after optimization (A) and before optimization (B). (Closed square) proteaseproduction (U/mL) and (open square) biomass (CFU/mL).
293VOL. 110, 2010 ALKALINE PROTEASE PRODUCTION BY BACILLUS MOJAVENSIS A21
the predicted and experimental values, reflecting the accuracy andapplicability of this model to optimize the conditions for enzymeproduction. Similar results were reported in a study by Chauhan andGupta (23) on Bacillus sp. RGR-14 for alkaline protease production. Inthis study, they achieved a 12.85-fold increase by optimizing starch,casamino acid, phosphate ion and inoculum concentrations using RSM.Another study performed by Beg et al. (30) showed that alkalineprotease production by B. mojavensis was improved up to 4.2-fold in abioreactor of 14 L using RSM. An overall 2.3-fold increase in proteaseproduction was achieved in the optimizedmedium, compared with theunoptimized basal medium described by Reddy et al. (31) for alkalineprotease production by Bacillus sp. RKY3. Our experiments also showedan increase in protease production higher than that reported by Agrebiet al. (32) and Fakhfakh-Zouari et al. (33), inwhichoptimizationbyRSMwas used for the production of fibrinolytic and keratinolytic activities,respectively. These and numerous other studies (34,35) clearly showthat this strategy is very powerful in the optimization of bioprocesses.
Among the four significant variables selected by the Plackett–Burman design experiment, HGW concentration was found to havethe greatest effect on the production of alkaline proteases. Maximumprotease production (1860.63 U/mL) was achieved at 30.0 g/L HGW,1.0 g/L SP, 2.0 g/L NaCl, 1.0 g/L KH2PO4 and 0.3 g/L K2HPO4, inaddition to other salts that were taken at suitable levels, as shown inthe Plackett–Burman design, a temperature of 30 °C, a speed ofagitation at 200 rpm and an initial pH of 9.0. Similarly, Bacillus clausiiproduced 1520 U/mL of protease under optimized culture condi-tions (36), and maximum protease production (1939 U/mL) was
obtained in the batch fermentation of a Bacillus sp. (35). However,protease production by B. mojavensis A21 is much better thanprotease production by B. subtilis A26 (269.36 U/mL) (32), Bacillussp. L21 (306.5 U/mL) (37), Bacillus sp. (410 U/mL) (27), B. mojavensis(558 U/mL) (30) and Bacillus sp. RKY3 (939 U/mL) (31). Joo et al.(38) reported that the maximum protease production by Bacillushorikoshii was 208 U/mL in optimized culture medium using soybeanmeal. This maximized activity was also lower than protease activityfound in our study. The majority of the alkaline proteases weresecreted within the first 24 h, and this result indicates that alkalineprotease production was related to the exponential growth phase ofthe bacteria. Similar results have been reported in previous investiga-tions (39). Using cost-effective media formulation and optimizing thismedia to determine its minimum requirements for maximum enzymeproduction is extremely important in industrial-scale proteaseproduction for economic reasons. Therefore, using common industrialingredients such as HGW,which is known for its low cost, will serve asa potential example for the applications used in industrial microbialfermentations. The simple expedient of replacing soluble starch in themedium with HGW could significantly lower the production cost.
Due to the increasing economic relevance of alkaline proteases,this study was conducted in an attempt to optimize a variety offermentation parameters, including medium compositions andculture conditions, for maximal alkaline protease production. Elevenvariables were tested using the Plackett–Burman design, and fourvariables (i.e., HGW, NaCl, KH2PO4, K2HPO4) were selected as themostinfluential factors on enzyme production. To optimize low-costfermentation conditions for alkaline protease production by B.mojavensis A21, we conducted RSM experiments using the screenedvariables. A successful and significant improvement (14-fold) in theproduction of bleach-stable alkaline proteases by B. mojavensis A21was accomplished using cheap carbon and nitrogen sources. Theoptimized medium established in this work might result in asignificant reduction in the cost of medium constituents and wouldthus offer advantages for large-scale fermentation.
ACKNOWLEDGEMENT
This work was funded by the Ministry of Higher Education andScientific Research, Tunisia.
References
1. Banerjee, U. C., Sani, R. K., Azmi, W., and Soni, R.: Thermostable alkaline proteasefrom Bacillus brevis and its characterization as a laundry detergent additive,Process Biochem., 35, 213–219 (1999).
2. Gupta,R., Beg,Q.K., andLorenz,P.:Bacterial alkalineproteases:molecular approachesand industrial applications, Appl. Microbiol. Biotechnol., 59, 15–32 (2002).
3. Johnvesly, B. and Naik, G. R.: Studies on production of thermostable alkalineprotease from thermophilic and alkaliphilic Bacillus sp. JB-99 in a chemicallydefined medium, Process Biochem., 37, 139–144 (2001).
4. Kirk, O., Borchert, T. V., and Fuglsang, C. C.: Industrial enzyme applications, Curr.Opin. Biotechnol., 13, 345–351 (2002).
5. Haaland, P. D.: Statistical problem solving, in: P. D. Haaland (Ed.), ExperimentalDesign in Biotechnology, Marcel Dekker, New York, 1989, pp. 1–18 (1989).
6. De Coninck, J., Bouquelet, S., Dumortier, V., Duyme, F., and Verdier-Denantes, I.:Industrial media and fermentation processes for improved growth and proteaseproduction by Tetrahymena thermophila, J. Ind. Microbiol. Biotechnol., 24, 285–290(2000).
7. Kunamneni, A., Kumar, K. S., and Singh, S.: Response surface methodologyapproach to optimize the nutritional parameters for enhances α-amylase, Afr. J.Biotechnol., 4, 708–716 (2005).
8. Dutta, J. R., Dutta, P. K., and Banerjee, R.: Optimization of culture parameters forextracellular protease production from a newly isolated Pseudomonas sp. usingresponse surface and artificial neural network models, Process Biochem., 39,2193–2198 (2004).
9. Haddar, A., Bougatef, A., Agrebi, R., Sellami-Kamoun, A., and Nasri, M.: A novelsurfactant-stable alkaline serine-protease from a newly isolated Bacillus mojavensisA21. Purification and characterization, Process Biochem., 44, 29–35 (2009).

294 HADDAR ET AL. J. BIOSCI. BIOENG.,
10. Haddar, A., Agrebi, R., Bougatef, A., Hmidet, N., Sellami-Kamoun, A., and Nasri,M.: Tow detergent stable alkaline serine-proteases from Bacillus mojavensis A21:purification, characterization and potential application as a laundry detergentadditive, Bioresource Technol., 100, 3366–3373 (2009).
11. Hajji, M., Rebai, A., Gharsallah, N., and Nasri, M.: Optimization of alkaline proteaseproduction byAspergillus clavatus ES1 inMirabilis jalapa tuber powderusing statisticalexperimental design, Appl. Microbiol. Biotechnol., 79, 915–923 (2008).
12. Jellouli, K., Bayoudh, A., Manni, L., Rym, A., and Nasri, M.: Purification,biochemical and molecular characterization of a metalloprotease from Pseudo-monas aeruginosa MN7 grown on shrimp wastes, Appl. Microbiol. Biotechnol., 79,989–999 (2008).
13. Souissi, N., Bougatef, A., Triki-Ellouz, Y., and Nasri, M.: Biochemical andfunctional properties of sardinella (Sardinella aurita) by-product hydrolysates,Food Technol. Biotechnol., 45, 187–194 (2007).
14. Kembhavi, A. A., Kulkarni, A., and Pant, A. A.: Salt-tolerant and thermostablealkaline protease from Bacillus subtilis NCIM No 64, Appl. Biochem. Biotechnol., 38,83–92 (1993).
15. Miller, J. H.: Experiments in Molecular Genetics, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY, 1972, pp. 431–435 (1972).
16. Plackett, R. L. and Burman, J. P.: The design of optimum multifactorialexperiments, Biometrika, 33, 305–325 (1946).
17. Weisberg, S.: Applied Linear Regression, John Wiley and Sons Inc, New York, 1985217 pp.
18. Khuri, A. I. and Cornell, J. A.: Response surfaces: design and analysis, MarcelDecker Inc., New York (1987).
19. Box, G. E. P., Hunter, W. G., and Hunter, J. S.: Statistics for experiments: anintroduction to design data analysis and model Building, John Wiley, New York,pp. 291–334 (1978).
20. Gupta, R., Beg, Q. K., Khan, S., and Chauhan, B.: An overview on fermentation,downstream processing and properties of microbial alkaline proteases, Appl.Microbiol. Biotechnol., 60, 381–395 (2002).
21. Wang, Q., Hou, Y., Xu, Z., Miao, J., and Li, G.: Optimization of cold-active proteaseproduction by the psychrophilic bacterium Colwellia sp. NJ341 with responsesurface methodology, Bioresource Technol., 99, 1926–1931 (2007).
22. Chu, I.M., Lee, C., and Li, T. S.: Production anddegradation of alkaline protease in batchcultures of Bacillus subtilis ATCC 14416, Enzyme Microb. Technol., 4, 55–61 (1992).
23. Chauhan, B. and Gupta, R.: Application of statistical experimental design foroptimization of alkaline protease production from Bacillus sp. RGR-14, ProcessBiochem., 39, 2115–2122 (2004).
24. Sandhya, C., Sumantha, A., Szakacs, G., and Pandey, A.: Comparative evaluation ofneutral protease production by Aspergillus oryzae in submerged and solid-statefermentation, Process Biochem., 40, 2689–2694 (2005).
25. Prakasham, R. S., Rao, Ch. S., and Sarma, P. N.: Green gram husk—an inexpensivesubstrate for alkaline protease production by Bacillus sp. in solid-state fermenta-tion, Bioresource Technol., 97, 1449–1454 (2006).
26. Hanlon, G. W., Hodges, N. A., and Russel, A. D.: The influence of glucose,ammonium and magnesium availability on the production of protease andbacitracin by Bacillus licheniformis, J. Gen. Microbiol., 128, 845–851 (1982).
27. Patel, R., Dodia, M., and Singh, S. P.: Extracellular alkaline protease from a newlyisolated haloalkaliphilic Bacillus sp.: production and optimization, ProcessBiochem., 40, 3569–3575 (2005).
28. Triki-Ellouz, Y., Ghorbel, B., Souissi, N., Kammoun, S., and Nasri, M.: Biosynthesisof protease by Pseudomonas aeruginosa MN7 grown on fish substrate, World J.Microbiol. Biotechnol., 19, 41–45 (2003).
29. Ghorbel, S., Souissi, N., Triki-Ellouz, Y., Dufosse, L., Guérard, F., and Nasri, M.:Preparation and testing of sardinella protein hydrolysates as nitrogen source forextracellular lipase production by Rhizopus oryzae, World J. Microb. Biotechnol., 21,33–38 (2005).
30. Beg, Q. K., Sahai, V., and Gupta, R.: Statistical media optimization and alkalineprotease production from Bacillus mojavensis in a bioreactor, Process Biochem., 39,203–209 (2003).
31. Reddy, L. V. A., Wee, Y. J., Yun, J. S., and Ryu, H. W.: Optimization of alkalineprotease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman and response surface methodological approaches, Bioresource Technol.,99, 2242–2249 (2008).
32. Agrebi, R., Haddar, A., Hajji, M., Frikha, F., Manni, L., Jellouli, K., and Nasri, M.:Fibrinolytic enzymes from a newly isolated marine bacterium Bacillus subtilis A26:characterization and statistical media optimization, Can. J. Microbiol., 55,1049–1061 (2009).
33. Fakhfakh-Zouari, N., Haddar, A., Hmidet, N., Frikha, F., and Nasri, M.:Application of statistical experimental design for optimization of keratinasesproduction by Bacillus pumilus A1 grown on chicken feather and some biochemicalproperties, Process Biochem., 45, 617–626 (2010).
34. Adinarayana, K. and Ellaiah, P.: Response surface optimization of the criticalmedium components for the production of alkaline protease by a newly isolatedBacillus sp, J. Pharm. Pharmaceut. Sci., 5, 272–278 (2002).
35. Puri, S., Khali, O., and Gupta, R.: Optimization of alkaline protease production fromBacillus sp. By response surface methodology, Curr. Microbiol., 44, 286–290 (2002).
36. Oskouie, S. F. G., Tabandeh, F., Yakhchali, B., and Eftekhar, F.: Response surfaceoptimization of medium composition for alkaline protease production by Bacillusclausii, Biochem. Eng. J., 39, 37–42 (2008).
37. Tari, C., Genckal, H., and Tokatli, F.: Optimization of a growth medium using astatistical approach for the production of an alkaline protease from a newlyisolated Bacillus sp. L21, Process Biochem., 41, 659–665 (2006).
38. Joo, H. S., Kumar, C. G., Park, G. C., Kim, K. T., Paik, S. R., and Chang, C. S.:Optimization of the production of an extracellular alkaline protease from Bacillushorikoshii, Process Biochem., 38, 155–159 (2002).
39. Uyar, F. and Baysal, Z.: Production and optimization of process parameters foralkaline protease production by a newly isolated Bacillus sp. under solid statefermentation, Process Biochem., 39, 1893–1898 (2004).




















