
Biochemistry of Alcoholic Fermentation
24
Chapter 1 Biochemistry of Alcoholic Fermentation Fernando Zamora Contents 1.1 Introduction .............................................................. 3 1.2 Yeast Development During Alcoholic Fermentation ............................. 4 1.3 Glycolysis ................................................................ 6 1.4 Fermentation and Respiration ................................................ 8 1.5 Regulation Between Respiration and Fermentation: Pasteur and Crabtree Effects ..... 9 1.6 Alcoholic Fermentation ..................................................... 10 1.7 Glyceropyruvic Fermentation ................................................ 11 1.8 Nitrogen Metabolism ....................................................... 12 1.9 Oxygen and Lipid Biosynthesis .............................................. 14 1.10 Stuck and Sluggish Fermentations: Causes and Solutions ......................... 18 1.11 Other Subproducts of Alcoholic Fermentation .................................. 20 References ............................................................... 22 1.1 Introduction Alcoholic fermentation is the anaerobic transformation of sugars, mainly glucose and fructose, into ethanol and carbon dioxide. This process, which is carried out by yeast and also by some bacteria such as Zymomonas mobilis, can be summarised by this overall reaction. C 6 H 12 O 6 Hexoses → 2 CH 3 CH 2 OH Ethanol + 2CO 2 Carbon dioxide However, alcoholic fermentation is fortunately a much more complex process. At the same time as this overall reaction proceeds, a lot of other biochemical, chemi- cal and physicochemical processes take place, making it possible to turn the grape F. Zamora (B) Departament de Bioqu´ ımica i Biotecnologia, Facultat d’Enologia de Tarragona, Universitat Rovira i Virgili, Campus de Sescelades, C/ Marcel.li Domingo s/n. 43007-Tarragona, Spain e-mail: [email protected] M.V. Moreno-Arribas, M.C. Polo (eds.), Wine Chemistry and Biochemistry, DOI 10.1007/978-0-387-74118-5 1, C Springer Science+Business Media, LLC 2009 3
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of Biochemistry of Alcoholic Fermentation

Chapter 1Biochemistry of Alcoholic Fermentation
Fernando Zamora
Contents
1.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31.2 Yeast Development During Alcoholic Fermentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41.3 Glycolysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61.4 Fermentation and Respiration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81.5 Regulation Between Respiration and Fermentation: Pasteur and Crabtree Effects . . . . . 91.6 Alcoholic Fermentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101.7 Glyceropyruvic Fermentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 111.8 Nitrogen Metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121.9 Oxygen and Lipid Biosynthesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 141.10 Stuck and Sluggish Fermentations: Causes and Solutions . . . . . . . . . . . . . . . . . . . . . . . . . 181.11 Other Subproducts of Alcoholic Fermentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
1.1 Introduction
Alcoholic fermentation is the anaerobic transformation of sugars, mainly glucoseand fructose, into ethanol and carbon dioxide. This process, which is carried out byyeast and also by some bacteria such as Zymomonas mobilis, can be summarised bythis overall reaction.
C6H12O6Hexoses
→ 2 CH3CH2OHEthanol
+ 2CO2Carbon dioxide
However, alcoholic fermentation is fortunately a much more complex process. Atthe same time as this overall reaction proceeds, a lot of other biochemical, chemi-cal and physicochemical processes take place, making it possible to turn the grape
F. Zamora (B)Departament de Bioquımica i Biotecnologia, Facultat d’Enologia de Tarragona,Universitat Rovira i Virgili, Campus de Sescelades, C/ Marcel.li Domingo s/n.43007-Tarragona, Spaine-mail: [email protected]
M.V. Moreno-Arribas, M.C. Polo (eds.), Wine Chemistry and Biochemistry,DOI 10.1007/978-0-387-74118-5 1, C© Springer Science+Business Media, LLC 2009
3

4 F. Zamora
juice into wine. Besides ethanol, several other compounds are produced throughoutalcoholic fermentation such as higher alcohols, esters, glycerol, succinic acid,diacetyl, acetoin and 2,3-butanediol. Simultaneously, some compounds of grapejuice are also transformed by yeast metabolism. Without the production of theseother substances, wine would have little organoleptic interest.
At the start of the winemaking process, several species of yeast may be present inthe grape juice. This biodiversity depends on several factors such as grape variety,the ripening stage at harvest, the antifungal treatments, the climatic conditions of theyear, the development of grey rot or other fungal plagues and the viticultural prac-tices (Sapis-Domerq 1980; Pretorius et al. 1999). However, other factors are alsoimportant. All contact of grapes and must during harvest, transport and, in particularwinery operations significantly influence the final distribution of yeasts at the begin-ning of alcoholic fermentation (Constantı et al. 1997; Mortimer and Polsinelli 1999).
Different yeast species participate in spontaneous alcoholic fermentation evenwhen sulphur dioxide is present (Constantı et al. 1998; Beltran et al. 2002). UsuallyKloeckera, Hanseniaspora and Candida predominate in the early stages of alcoholicfermentation. Later, Pichia and Metschnikowia prevail in the middle stages. Finally,during the latter stages of fermentation, Saccharomyces cerevisiae is the predomi-nant yeast because of its greater resistance to high ethanol concentration (Fleet 1993;Fleet and Heard 1993). Some other yeast, such as Torulaspora, Kluyveromyces,Schizosacchaomyces, Zygosaccharomyces and Brettanomyces may also be presentduring alcoholic fermentation and even in the wine itself, which may cause someorganoleptic defects (Peynaud and Domercq 1959; Ribereau-Gayon et al. 2000a).
Evidently, the succession of these different yeast species throughout alcoholicfermentation influences the final composition of wine in a way that, dependingon which yeasts have grown, may be positive in some cases or negative in oth-ers (Chatonnet et al. 1995; Ribereau-Gayon et al. 2000a). To prevent undesirableyeasts developing, wineries add sulphur dioxide to the grape juice and inoculateselected strains of dry yeasts (Saccharomyces cerevisiae). Sulphur dioxide has adrastic selective effect on yeast development. As Saccharomyces cerevisiae is moreresistant to sulphur dioxide than most other yeasts, using this additive favours itsdevelopment (Lafon-Lafourcade and Peynaud 1974; Romano and Suzzi 1993).
On the other hand, the inoculation of selected dry yeasts greatly increases theinitial population of Saccharomyces cerevisiae. Nowadays, most wineries inoculateselected dry yeast in order to guarantee alcoholic fermentation without any devi-ation. However, other wineries, especially traditional wine cellars, continue to usespontaneous alcoholic fermentation because they believe it gives their wines greatercomplexity.
1.2 Yeast Development During Alcoholic Fermentation
At the beginning of the winemaking process, the yeasts start to metabolize the sugarsand other nutrients present in the grape juice. The yeasts use all these nutrients toobtain energy and increase their population (Boulton et al. 1996; Ribereau-Gayon

1 Biochemistry of Alcoholic Fermentation 5
109
108
107
106
105
104
103
102
10
0
a
b c
d
Time
Yea
st p
opul
atio
n (c
ell/m
l)
Viable cells
Total cells
a: Latency phase
b: Exponential growth phase
c: Quasi-stationary phase
d: Decline phase
Fig. 1.1 Yeasts growth cycle
et al. 2000b). Figure 1.1 shows the classic yeast growth cycle under standard condi-tions (Fleet and Heard 1993; Del Nobile et al. 2003).
During the first hours the yeast population does not increase. During this period,also called the latency phase, it is necessary for the cell to adapt to the new envi-ronmental conditions. The initial population depends on several factors. If no yeastsare inoculated, the population is around 104 cells/ml. However, this population canbe higher if the grapes have been attacked by grey rot or other fungal plagues. Onthe other hand, if selected dry yeasts were inoculated, the initial population wouldalso be higher (around 5 × 106 cells/ml).
Once the yeasts have adapted to the environmental conditions, they begin togrow. This period, named the exponential growth phase, is highly influenced bytemperature (Ough 1964), by the concentration of ammonia, amino acids and othernutrients (Lafon-Lafourcade 1983; Sablayrolles et al. 1996) and by the presence ofoxygen (Sablayrolles and Barre 1986). During the exponential growth phase, theyeasts increase their population up to 107−108 cells/ml. This phase can last from 3to 6 days. After that, yeast stops growing because some nutrients became deficient.During this new phase, called the quasi-stationary phase, the population of yeastremains nearly stable and can last from 2 to 10 days. Later, the decline phase beginsand the population of yeast gradually decreases until it has almost completely dis-appeared. During this period yeasts die because of the lack of nutrients and alsobecause ethanol and other substances produced during alcoholic fermentation aretoxic to them (Lafon-Lafourcade et al. 1984).
The success of an alcoholic fermentation depends on maintaining the popula-tion of viable yeast at sufficient levels until all the fermentable sugars have beenfully consumed (Bisson 1999; Zamora 2004). Otherwise, the winemaker is facedwith the serious problem of stuck and sluggish fermentations. The causes and theways to avoid stuck and sluggish fermentations are discussed later (Bisson andButzke 2000).

6 F. Zamora
1.3 Glycolysis
The word glycolysis comes from the Greek terms ������ (glucus = sweet) and����� (lysis = rupture) and the process consists of the intracellular transformationof glucose (and fructose) into pyruvate. This biochemical pathway is the initial pro-cess of carbohydrate catabolism in most organisms and it takes place completelywithin the cytoplasm. This pathway was fully described in 1940 due, in great part,to the contributions of Gustav Embdem and Otto Meyenhorf. For that reason, it isalso called the Embdem-Meyerhoff pathway in their honour although, regrettably,this name excludes other important contributors such as Gerti and Karl Cori, CarlNeuberg, Jacob Parnas, Hans von Euler and Otto Warburg (Kresge et al. 2005).
Yeasts use glycolysis as the main pathway for sugar catabolism (Gancedo 1988).The pentose pathway, which is used by some organisms such as acetic acid bacteriaas the major pathway for sugar catabolism, is only used by yeast as a source ofribose and NADPH (Schaaf-Gersteenschalager and Miosga 1996; Horecker 2002).Ribose is necessary for synthesizing nucleotides and nucleic acids whereas NADPHis required for some metabolic processes such as the lipid synthesis. Thereforeyeasts use the pentose pathway not to obtain energy but rather to provide themselveswith some of the substances indispensable for cell multiplication.
Glycolysis involves a sequence of 11 chemical reactions for breaking down hex-oses and releasing energy in the chemical form of ATP (Barnett 2003). Figure 1.2shows all the reactions in the glycolytic pathway.
Initially, hexoses are transported inside the cell by facilitated diffusion(Lagunas 1993). As the inner sugar concentration is lower than the external sugarconcentration, no energy is necessary for this process.
The first step in glycolysis is the phosphorylation of glucose and fructose by afamily of enzymes called hexokinases to form glucose 6-phosphate and fructose-6-phosphate (Gancedo 1988). This reaction consumes ATP, but it keeps the intracel-lular hexose concentration low and thus favours the continuous transport of sugarsinto the cell through the plasma membrane transporters. After this, phosphoglucoseisomerase converts glucose-6-phosphate into fructose-6-phosphate.
Besides being intermediaries of glycolysis, glucose-6-phosphate and fructose-6-phosphate are also essential substrates for secondary metabolism. In fact, bothhexose-phosphates are needed to synthesize the polysaccharides used to constructthe cell wall (Cabib et al. 1982).
In the following stage, fructose-6-phosphate is phosphorylated again by the actionof phosphofructokinase to form fructose-1,6-diphosphate. This reaction also con-sumes ATP. Later, the enzyme aldolase cleaves to fructose-6-phosphate. As a resultof this reaction two triose phosphates are formed: dihydroxyacetone phosphateand glyceraldehyde-3-phosphate. This reaction produces a much greater proportionof dihydroxyacetone phosphate (96%), which is rapidly transformed into glycer-aldehyde-3-phosphate by triose phosphate isomerase (Heinisch and Rodicio 1996).
Afterwards, the enzyme glyceraldehyde-3-phosphate dehydrogenase transformsglyceraldehyde-3-phosphate into 1,3-diphosphoglycerate. This reaction involves theoxidation of the molecule that is linked to reducing NAD+ to NADH in order toredress the redox balance. Simultaneously, a substrate level phosphorylation takes
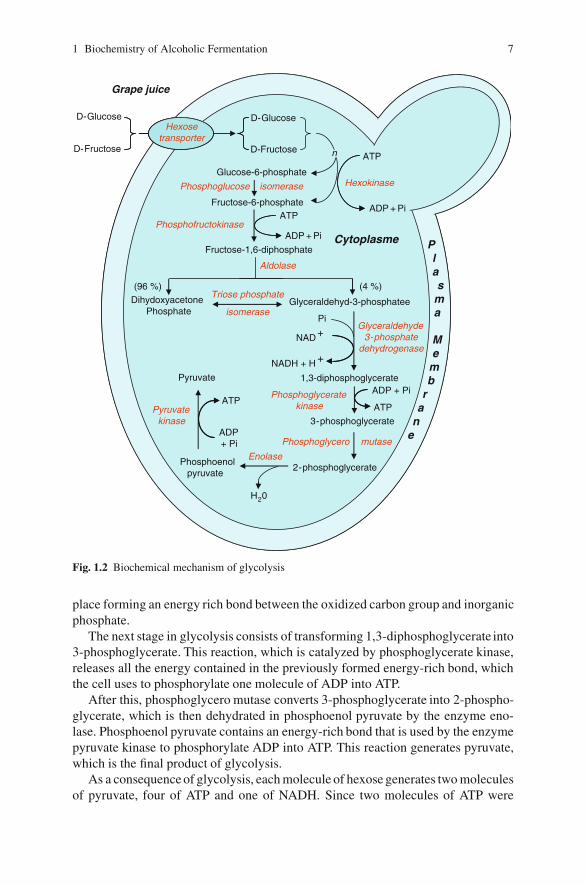
1 Biochemistry of Alcoholic Fermentation 7
D
Hexosetransporter
D-Glucose
D-Fructose
Glucose-6-phosphateHexokinase
Fructose -6-phosphate
ATP
ADP +Pi
Phosphoglucose isomerase
PhosphofructokinaseATP
ADP +PiPl as
ma
Me
mb
ra
ne
CytoplasmeFructose -1,6-diphosphate
DihydoxyacetonePhosphate
Glyceraldehyde -3-phosphate
Aldolase
Triose phosphate
isomerase
Glyceraldehyde3-phosphate
dehydrogenaseNAD +
NADH + H +
ATP
1,3-diphosphoglycerateADP + PiPhosphoglycerate
kinase
3-phosphoglycerate
Phosphoglycero mutase
2-phosphoglyceratePhosphoenol
pyruvate
Pyruvate
ATP
ADP+ Pi
Pyruvate kinase
Enolase
H20
Pi
(4 %)(96 %)
D-Glucose
-Fructose
Hexosetransporter
Hexosetransporter
D-Glucose
D-Fructose
Glucose-6-phosphateHexokinase
Fructose-6-phosphate
ATP
ADP + Pi
ADP + Pi
Phosphoglucose isomerase
Phosphofructokinase
Grape juice
ATP
Pl as
ma
Me
mb
ra
ne
CytoplasmeFructose-1,6-diphosphate
DihydoxyacetonePhosphate
Glyceraldehyd-3-phosphatee
Aldolase
Triose phosphate
isomerase
Glyceraldehyde3-phosphate
dehydrogenaseNAD +
NADH + H +
ATP
1,3-diphosphoglycerateADP + PiPhosphoglycerate
kinase
3-phosphoglycerate
Phosphoglycero mutase
2-phosphoglyceratePhosphoenol
pyruvate
Pyruvate
ATP
ADP+ Pi
Pyruvate kinase
Enolase
H20
Pi
(4 %)(96 %)
n
Fig. 1.2 Biochemical mechanism of glycolysis
place forming an energy rich bond between the oxidized carbon group and inorganicphosphate.
The next stage in glycolysis consists of transforming 1,3-diphosphoglycerate into3-phosphoglycerate. This reaction, which is catalyzed by phosphoglycerate kinase,releases all the energy contained in the previously formed energy-rich bond, whichthe cell uses to phosphorylate one molecule of ADP into ATP.
After this, phosphoglycero mutase converts 3-phosphoglycerate into 2-phospho-glycerate, which is then dehydrated in phosphoenol pyruvate by the enzyme eno-lase. Phosphoenol pyruvate contains an energy-rich bond that is used by the enzymepyruvate kinase to phosphorylate ADP into ATP. This reaction generates pyruvate,which is the final product of glycolysis.
As a consequence of glycolysis, each molecule of hexose generates two moleculesof pyruvate, four of ATP and one of NADH. Since two molecules of ATP were

8 F. Zamora
consumed previously during the phosphorylation of the hexoses, the net energy gainfor the cell is two ATPs per hexose.
Pyruvate produced by glycolysis can be used by yeasts for several metabolic path-ways. However, yeasts must regenerate NAD+ from the NADH to re-establish theoxydoreduction potential of the cell. This can be done by fermentation or respiration.
1.4 Fermentation and Respiration
Yeasts are facultative anaerobic microorganisms because they possess the geneticequipment for metabolizing sugars aerobically or anaerobically (Boulton et al.1996). Therefore, yeasts can consume sugars using two different metabolic path-ways: respiration and fermentation (Racker 1974). Figure 1.3 illustrates these bio-chemical pathways.
Pl as
ma
Me
mb
ra
ne
Dihydoxyacetonephosphate
Glyceraldehyde -3-phosphate
Fermentation
Glycerol
Pyruvate
Hexosetransporter
D-Glucose
D-Fructose
Ethanol
CO2
Ethanal
Pyruvate decarboxylase
Pyruvate dehydrogenase
Acetyl -coA
NAD +
NADH + H +
HScoA
CO2
CO
Oxidizedcoenzymes
Reducedcoenzymes
KrebsCycle
Respiratory chains
O2 H2O
Energy
Energy
Respiration
Pl as
ma
Me
mb
ra
ne
Grape juice
Dihydoxyacetonephosphate Glyceraldehyde-3-phosphate
Fermentation
GlycerolGlycerol
Pyruvate
D-Glucose
D-Fructose
Hexosetransporter
Hexosetransporter
D-Glucose
D-Fructose
D-Glucose
D-Fructose
Ethanol
CO2
EthanalEthanol
Pyruvate decarboxylase
Pyruvate dehydrogenase
Acetyl-coA
NAD +
NADH + H +
HScoA
CO2
CO2CO2
CO2
Oxidizedcoenzymes
Reducedcoenzymes
KrebsCycleKrebsCycle
Respiratory chainsRespiratory chains
O2O2 H2O
Energy
Energy
Respiration
CO2
Fig. 1.3 Fermentation and respiration

1 Biochemistry of Alcoholic Fermentation 9
Both pathways begin with glycolysis (as described above), which generates pyru-vate as a final product. Pyruvate can be transformed into ethanal and carbon dioxideby the enzyme pyruvate decarboxylase and after ethanal can be reduced to ethanol.This process, named alcoholic fermentation, takes place within the cytoplasm. Alco-holic fermentation regenerates the NAD+ consumed during glycolysis and givesyeast an energy gain of only two ATP molecules by metabolized hexose (Barnettand Entian 2005).
Nevertheless, pyruvate can also be transformed into acetyl-coA and carbon diox-ide by pyruvate dehydrogenase. This reaction reduces NAD+ to NADH and mustincorporate the coenzyme A. Acetyl-coA can then be incorporated to the Krebscycle, being transformed into carbon dioxide and producing several molecules ofreduced coenzymes (NADH and FADH2). The reduced coenzymes produced by theKrebs cycle, and also by glycolysis, are later reoxidized in the respiratory chains,reducing molecular oxygen to water (Barnett and Entian 2005). This process, knownas respiration, yields an overall energy gain of 36–38 ATP molecules per metab-olized hexose. Consequently, this process is much more beneficial to yeast thanfermentation, in terms of energy. However, it needs oxygen as a substrate and it isinhibited by high sugar concentration (Crabtree 1929).
The transformation of pyruvate into ethanal or acetyl-coA is therefore a key pointfor regulating yeast metabolism (Ribereau-Gayon et al. 2000c).
1.5 Regulation Between Respiration and Fermentation: Pasteurand Crabtree Effects
Louis Pasteur found that aeration increases biomass production and decreases thekinetics of sugar consumption and ethanol production (Pasteur 1861). He, therefore,concluded that aeration inhibits alcoholic fermentation (Racker 1974).
This phenomenon, which is known as the Pasteur effect, has been attributedto several mechanisms (Barnett and Entian 2005). Respiration needs very highamounts of ADP inside the mitochondria as a subtract for oxidative phosphory-lation. Therefore, when respiration takes place, the cytoplasm lacks ADP and inor-ganic phosphate (Lagunas and Gancedo 1983), which in turn decreases the sugartransport inside the cell (Lagunas et al. 1982). These mechanisms explain how aer-ation inhibits the alcoholic fermentation.
Evidently, once the yeast starts to consume sugars, large quantities of carbondioxide are produced. The release of carbon dioxide displaces the oxygen andcreates semianaerobic conditions that favour fermentation. However, even in thepresence of oxygen, Saccharomyces cerevisiae will not ferment if the sugar concen-tration is higher than 9 g/l. Crabtree first described this phenomenon in 1929 that isknown by different names: the Crabtree effect, catabolic repression by glucose orthe Pasteur contrary effect (Meijer et al. 1998; Ribereau-Gayon et al. 2000c).
When Saccharomyces cerevisiae grow in a high sugar concentration, as is foundin grape juice, their mitochondria degenerate. Simultaneously, the enzymes of the

10 F. Zamora
Krebs cycle and the constituents of respiratory chains are repressed (Gancedo 1992;Polakis et al. 1965; Barnett and Entian 2005). Therefore, under wine fermenta-tion conditions, Saccharomyces cerevisiae can only ferment sugars. Saccharomycescerevisiae can only use respiration when the sugar concentration is really low andwhen oxygen is present in the medium. These conditions are used for the industrialproduction of selected dry yeast.
1.6 Alcoholic Fermentation
As was quoted above, when fermenting grape juice, Saccharomyces cerevisiaemainly directs the pyruvate to produce of ethanol in order to regenerate the NAD+
consumed by glycolysis. This process, called alcoholic fermentation, is shown inFig. 1.4.
Pl as
ma
Me
mb
ra
ne
Dihydoxyacetonephosphate Glyceraldehyde-3-phosphate
AlcoholicFermentation
Hexosetransporter
D-Glucose
D-Fructose
1,3-diphosphoglycerate
Alcoholdehydrogenase
Glyceraldehyde3-phosphate
dehydrogenase
Ethanal
EthanolNAD+
NADH + H+
Pyruvate
CO2
Pyruvate decarboxylase
2 ATP
2 ADP +2 Pi
4 ATP 4 ADP +4 Pi
Pl as
ma
Me
mb
ra
ne
Grape juice
Dihydoxyacetonephosphate
Glyceraldehyde-3-phosphate
AlcoholicFermentation
D-Glucose
D-Fructose
Hexosetransporter
Hexosetransporter
D-Glucose
D-Fructose
D-Glucose
D-Fructose
1,3-diphosphoglycerate
Alcoholdehydrogenase
Glyceraldehyde3-phosphate
dehydrogenase
Ethanal
EthanolNAD+
NADH + H+
Ethanol
Pyruvate
CO2
Pyruvate decarboxylaseCO2
2 ATP
2 ADP +2 Pi
4 ATP 4 ADP +4 Pi
Fig. 1.4 Alcoholic fermentation

1 Biochemistry of Alcoholic Fermentation 11
Pyruvate is initially decarboxylated into ethanal by pyruvate decarboxylase. Thisenzyme needs magnesium and thiamine pyrophosphate as cofactors (Hohmann1996). Thereafter, alcohol dehydrogenase reduces ethanal to ethanol, recyclingthe NADH to NAD+. There are three isoenzymes of alcohol dehydrogenase inSaccharomyces cerevisiae, but isoenzyme I is chiefly responsible for convertingethanal into ethanol (Gancedo 1988). Alcohol dehydrogenase uses zinc as cofactor(Ciriacy 1996).
Both final products of alcoholic fermentation, ethanol and carbon dioxide, aretransported outside the cell by simple diffusion.
1.7 Glyceropyruvic Fermentation
Although the production of ethanol is the most important pathway to regener-ate NAD+, there is an alternative pathway for this purpose. This pathway, calledglyceropyruvic fermentation, generates glycerol as its final product (Prior andHohmann 1996). Figure 1.5 shows the biochemical mechanism of glyceropyruvicfermentation.
The first evidence of this pathway was found by Neuberg (1946). He discoveredthat the fermentation of glucose by yeast in the presence of sulphite produced alot of glycerol. Sulphite combines with ethanal which then prevents NAD+ fromregenerating via alcohol dehydrogenase. Under these conditions, the yeasts need tooxide NADH through an alternative pathway in order to compensate for the NAD+
deficit and the only way to do this is by producing glycerol.Dyhroxyacetone phosphate, the main product of aldolase reaction, can be oxi-
dized to glycerol-3-phosphate by the enzyme glycerol-3-phosphate dehydrogenase.This reaction is coupled to the oxidation of a molecule of NADH to NAD+.Then, glycerol-3-phosphate phosphatase catalyzes the production of glycerol bydephosphorylating glycerol-3-phosphate. The production of glycerol consumes ATPbut it is necessary to compensating for the redox imbalance in the cell (Barreet al. 1998).
Although glyceropyruvic fermentation was first described through the effect ofsulphites, it can also be active in other situations. At the beginning of winemaking,yeasts need a lot of substrates to grow. Cell multiplication implies a very activebiosynthesis of proteins, lipids, nucleotides, etc., and most of these biomoleculesare synthesised using pyruvate as a substrate. Each time a molecule of pyruvate isused anabolically, a NAD+ deficit is produced which must be recovered throughthe glyceropyruvic pathway. For this reason, glycerol is mainly produced during thefirst steps of alcoholic fermentation, when yeasts are growing and they need a largeproportion of pyruvate to increase their biomass (Ribereau-Gayon et al. 2000c).Furthermore, yeasts produce glycerol as a protector against high osmotic pressures(Prior and Hohmann 1996).
For these reasons, glycerol is the third major component of dry wines (after waterand ethanol). Its concentration is usually between 6 and 10 g/l and it improves winequality because it confers sweet and mouthfeel sensations.

12 F. Zamora
Pl as
ma
Me
mb
ra
ne
Dihydoxyacetonephosphate
Glyceraldehyde -3-phosphate
Cytoplasme
Triosephosphate
isomerase
Glyceraldehyde3-phosphate
dehydrogenaseNAD +
NADH + H +
Glycerol 3 -phosphatedehydrogenase
1,3-diphosphoglycerateGlycerol -3-phosphate
GlycerolGlycerol Pyruvate
D-Glucose
D-Fructose
Hexosetransporter
D-Glucose
D-Fructose
Alcoholdehydrogenase
Ethanol
CO2
Glycerol 3 -phosphatephosphatase
EthanalOthermetabolites
NAD +
NADH + H +
Pyruvatedecarboxylase
2 ATP
2 ADP +2 Pi
Pl as
ma
Me
mb
ra
ne
Pl as
ma
Me
mb
ra
ne
Grape juice
Dihydoxyacetonephosphate
Glyceraldehyde-3-phosphate
Cytoplasme
Triosephosphate
isomerase
Glyceraldehyde3-phosphate
dehydrogenaseNAD+
NADH + H+
Glycerol 3-phosphatedehydrogenase
1,3-diphosphoglycerateGlycerol-3-phosphate
Glycerol Pyruvate
Hexosetransporter
Hexosetransporter
D-Glucose
D-Fructose
Alcoholdehydrogenase
Ethanol
CO2
Glycerol 3-phosphatephosphatase
EthanalOthermetabolites
NAD +
NADH + H +
Pyruvatedecarboxylase
2 ATP
2 ADP + 2 Pi
Fig. 1.5 Glyceropyruvic fermentation
1.8 Nitrogen Metabolism
When Saccharomyces cerevisiae grows in grape juice it needs significant amountsof assimilable nitrogen to synthesize biomass (Kunkee 1991). Grape juice containsa variety of nitrogen compounds such as ammonia, amino acids, peptides, pro-teins, etc., but only some of them can be assimilated by Saccharomyces cerevisiae(Hensche and Jiranek 1993). When fermenting grape juice fermentation, Saccha-romyces cerevisiae can only use ammonia and amino acids, with the exceptionof proline, as an assimilable source of nitrogen (Barre et al. 1998). Proline canbe assimilated by Saccharomyces cerevisiae but only under aerobic conditions(Boulton et al. 1996). For this reason, the term easily-assimilable nitrogen (EAN)has been proposed to describe collectively all the ammonia and amino acids, except

1 Biochemistry of Alcoholic Fermentation 13
proline. This EAN can simply be determined using the formol index (Taylor 1957;Aerny 1997).
Grape juice is relatively poor in ammonia and amino acids (Kunkee 1991). Con-sequently, the uptake of these compounds by the yeasts may be a critical stagein some cases. At a low concentration of EAN the risk of stuck and sluggish fer-mentations is really high (Bisson 1999; Zamora 2004). For that reason, winemak-ers used to supplement grape juice with ammonium salts (Barre et al. 1998). TheEAN requirement for a complete alcoholic fermentation depends on the yeast strain(Manginot et al. 1998) and the potential alcoholic degree (Bisson and Butzke 2000).Generally, it is considered that an EAN lower than 130 mg/l can seriously affect thecorrect development of alcoholic fermentation.In contrast, excessive nitrogen canlead to the presence of non-assimilated residual nitrogen in the wine, which is a fac-tor in microbiological instability and can even favour the production of ethyl carba-mate and biogenic amines (Ribereau-Gayon et al. 2000b). For that reason, nitrogenmust be supplemented carefully and taking into account the initial EAN concentra-tion of grape juice and its potential alcoholic degree (Bisson and Butzke 2000).
The first step in nitrogen assimilation is its transport inside the cell. Figure 1.6illustrates nitrogen uptake in yeasts.
Ammonium ion is transported inside the cell by facilitated diffusion. How-ever, the intracellular pH causes ammonium ion to release a proton, which mustbe sent outside the cell via H+-ATPase. Once inside the cytoplasm, ammonia
Pl a
sma
Me
mb
ra
ne
NH4+ NH4
+ NH3
ATP
ADP +Pi
H+-ATPase H+
H
Amino acid
H+
Amino acidpool
2-oxoglutarate
Glutamate
Glutamine
NAD +
NADH + H +
Permease
Glutamate
dehydrogenase
Glutamime
synthethase
Pl a
sma
Me
mb
ra
ne
Pl a
sma
Me
mb
ra
ne
NH4+ NH3
ATP
ADP + Pi
H+-ATPaseH+-ATPase H+
Amino acid
H+
HH+
Amino acid
H+
Peptides
Proteins
Polyamides
Nucleic acids
Vitamins
Amino acidpool
Amino acidpool
Grape juice
2-oxoglutarate
Glutamate
Glutamine
NAD+
NADH + H+
PermeasePermease
Glutamate
dehydrogenase
Glutamime
synthethase
Fig. 1.6 Nitrogen metabolism

14 F. Zamora
is incorporated to the amino acid pool via glutamate dehydrogenase or via glu-tamine synthetase, thus producing glutamate or glutamine respectively (Henscheand Jiranek 1993).
On the other hand, amino acids are transported inside the cell by different trans-porters. To date,15 transport systems have been identified for amino acids in Sac-charomyces cerevisiae (Barre et al. 1998) and all of them are symport systemscoupled to the entry of a proton. This proton must also be sent outside the cellin order to maintain the cellular homeostasis. Therefore, the uptake of ammoniumand amino acids must be considered as active transport because it consumes ATPvia H+-ATPase.
All amino acids, except proline, may be used by Saccharomyces cerevisiae ingrape juice fermentation. Amino acids can be directly used to synthesize proteins.However, the amino acid composition of the grape juice is not necessarily similar tothe needs of the cell. For that reason, yeasts must use the remaining amino acidsto synthesize those which it lacks (Hensche and Jiranek 1993; Ribereau-Gayonet al. 2000b). In this case, ammonia is incorporated into other amino acids whereasthe carbon skeleton is metabolized by the cell.
For this reason, the lack of enough EAN can make the yeast use sulphur aminoacids (cysteine and methionine), thus releasing hydrogen sulphite and mercaptans.Thus, supplementing with ammonium salts is recommended not only to avoidstuck and sluggish fermentations but also to prevent reduction off-odours (Jiraneket al. 1995).
Finally, the relationship between the amino acid composition of grapes and thefinal aromatic composition of wine has been recently described (Hernandez-Orteet al. 2002, 2006). Therefore, it is possible that in the near future grape juice willbe complemented with specific mixtures of amino acids in order to improve thearomatic quality of wine.
1.9 Oxygen and Lipid Biosynthesis
As discussed previously,Saccharomyces cerevisiae does not need oxygen to obtainenergy when fermenting grape juice. However, there are some essential biosyntheticpathways that use oxygen as substrate. This is the case for the biosynthesis of sterolsand unsaturated fatty acids (Ratledge and Evans 1989).
During the growth phase, while the cell multiplication is active, yeast needs tobuild new plasma membranes continually. For that reason, yeasts must synthesizegreat amounts of sterols, fatty acids and phospholipids during the first stages ofalcoholic fermentation (Ribereau-Gayon et al. 2000b).
Figure 1.7 illustrates the synthesis of sterols in yeasts. Basically, sterols are syn-thesised by the mevalonate pathway. The key stage in this pathway is, without anydoubt, the reaction catalysed by squalene monooxygenase. This reaction, which usesoxygen as substrate, transforms squalene into squalene 2,3, epoxide. Later, squaleneepoxide lanosterol cyclase catalyses the synthesis of the first sterol of the pathway,

1 Biochemistry of Alcoholic Fermentation 15
Pl asma
Me
mb
ra
ne
Grape juiceCytoplasme
Acetyl-coA
OO
HOHOHOHO
Acetoacetyl-coA
Squalene
Hydroxymethylglutaryl-coA
Mevalonicacid
Lanosterol
Squalenemomooxygenase
NAPD+ NADPH + H+
O2H2O
Squalene2,3-epoxyde
Squaleneepoxidelanosterol
cyclase
Membrane insertion
Ergosterol
Mevalonicacidpathway
Pl asma
Me
mb
ra
ne
Pl asma
Me
mb
ra
ne
Cytoplasme
Acetyl-coA
O
HOHOHOHO
Acetoacetyl-coA
Squalene
Hydroxymethylglutaryl-coA
Mevalonic acid
Lanosterol
Squalenemomooxygenase
NAPD+ NADPH + H+
O2H2O
Squalene2,3-epoxyde
Squalene epoxidelanosterol
cyclase
Membrane insertion
Ergosterol
Mevalonicacidpathway
Fig. 1.7 Synthesis of ergosterol in yeasts
lanosterol. Afterwards, lanosterol is used to obtain ergosterol, which is the mainsterol in Saccharomyces cerevisiae (Nes et al. 1993).
Therefore, in the absence of oxygen, this pathway is completely blocked and itsfinal yeast product, ergosterol, cannot be produced.
Figure 1.8 shows the synthesis of fatty acids. This complex process is catalysedby the multienzymatic complex, fatty acid synthetase. This enzyme uses as sub-strates acetyl-coA and malonyl-coA to produce palmitic acid. Afterwards, palmiticacid, a saturated fatty acid of 16 carbon atoms, can be used to produce other fattyacids (Ratledge and Evans 1989). Fatty acids with more carbon units, such asestearic acid, are obtained by elongation of palmitic acid.

16 F. Zamora
Pl as
ma
Me
mb
ra
ne
Cytoplasme
Acetyl-coA
Phospholipids
Malonyl-coA
½ O2 H2O
COOHCOOHCO-ScoA
Acetyl-coA carboxylase
CO2
NAPD+
NADPH + H+
CO2
SaturatedFatty Acid (SFA)
UnsaturatedFatty Acid (UFA)
CO-ScoA
Fatty acid sinthetase
Membrane insertion
Pl as
ma
Me
mb
ra
ne
Cytoplasme
Acetyl-coA
Phospholipids
Malonyl-coA
½ O2 H2O
COOHCOOHCO-ScoACOOHCOOHCO-ScoA
Acetyl-coA carboxylase
CO2
NAPD+
NADPH + H+
CO2
SaturatedFatty Acid (SFA)
UnsaturatedFatty Acid (UFA)
CO-ScoACO-ScoA
Fatty acid sinthetase
Membrane insertion
DesaturaseOLE1
Pl as
ma
Me
mb
ra
ne
Grape juiceCytoplasme
Acetyl-coA
Phospholipids
Malonyl-coA
2 H2O
COOHCOOHCO-ScoA
Acetyl-coA carboxylase
CO2
NAPD+
NADPH + H+
CO2
SaturatedFatty Acid (SFA)
UnsaturatedFatty Acid (UFA)
CO-ScoA
Fatty acid sinthetase
Membrane insertion
Pl as
ma
Me
mb
ra
ne
Cytoplasme
Acetyl-coA
Phospholipids
Malonyl-coA
2 H2O
COOHCOOHCO-ScoACOOHCOOHCO-ScoA
Acetyl-coA carboxylase
CO2
NAPD+
NADPH + H+
CO2
SaturatedFatty Acid (SFA)
UnsaturatedFatty Acid (UFA)
CO-ScoACO-ScoA
Fatty acid sinthetase
Membrane insertion
Desaturase OLE1
SSSSSSSS
Fig. 1.8 Synthesis of fatty acids in yeasts
Nevertheless, unsaturated fatty acids (UFA) need oxygen to be synthesised(Alexandre et al. 1994). In Saccharomyces cerevisiae, UFA production is catalysedby the desaturase OLE1 (Kajiwara et al. 2000). This enzyme is activated by lowtemperatures and the presence of oxygen (Nakagawa et al. 2002). Afterwards, fattyacids are used to synthesize phospholipids, which are inserted into the plasmaticmembranes.
Since oxygen is necessary to synthesize ergosterol and UFA, aeration duringalcoholic fermentation limits the growth of Saccharomyces cerevisiae (Sablayrollesand Barre 1986; Ribereau-Gayon et al. 2000b) and the lack of oxygen may some-times cause stuck and sluggish fermentations (Sablayrolles et al. 1996; Bisson 1999;

1 Biochemistry of Alcoholic Fermentation 17
Zamora 2004). For that reason, aerating the grape juice is recommended during theexponential growth phase of the fermentation in order to encourage yeast to buildtheir membranes and avoid fermentation problems (Barre et al. 1998; Ribereau-Gayon et al. 2000b).
Another aspect to take into account is that all microorganisms need to main-tain adequate fluidity in their membranes (Rodrıguez et al. 2007). Excessive rigid-ity can prevent cellular transport systems from functioning correctly (Los andMurata 2004).In contrast, excessive fluidity can alter the organization and thedynamic properties of phospholipidic bilayer (Laroche et al. 2001).
The fluidity of the plasmatic membrane is considerably affected by temperature(Rodrıguez et al. 2007) and ethanol concentration (Jones and Greenfield 1987).Therefore, during alcoholic fermentationSaccharomyces cerevisiae must adapt thefluidity of the membrane to the changing environmental conditions. It should beemphasized that the temperature of fermentation and aeration depend on the typeof winemaking. Usually, white wines are made at low temperatures (14–18 ◦C) andwithout aeration to conserve aromas whereas red wines are fermented at relativelyhigh temperatures (28–30 ◦C) and are aerated in order to enhance colour extraction.
In white winemaking, Saccharomyces cerevisiae must develop at low temper-ature, which reduces membrane fluidity. To maintain adequate fluidity in theirmembranes, yeasts increase the proportion of UFA in the phospholipids (Thurstonet al. 1981; Torrija et al. 2003). Phospholipids with unsaturated fatty acids havea lower melting point and more flexibility than phospholipids with saturated acylchains (Rodrıguez et al. 2007). Such adaptation involves inducing the fatty aciddesaturase OLE1 which incorporate unsaturated bonds at defined positions in fattyacids (Nakagawa et al. 2002).
However, under normal white fermentation conditions, grape juice is very poorin fatty acids and fermentation is usually carried out under hypoxia conditions(Bertrand and Miele 1984). Under these conditions, yeasts cannot synthesize unsatu-rated fatty acids. Consequently, Saccharomyces cerevisiae need to use another strat-egy to fluidize their membranes and the only possibility is incorporating mediumchain fatty acids (MCFA) within the phospholipids of the membrane (Rozes 1992).The effect of a short chain is similar to that of the double bond of a long chain(Quinn and Chapman 1980) and, therefore, the increased synthesis of MCFA couldalso modulate membrane fluidity.
Nevertheless, some of these MCFA can be released into the medium, reducingyeast viability and leading to stoppages in fermentation (Geneix et al. 1983). Instandard red winemaking, this problem does not exist because fermentations aredone at high temperatures and oxygen is introduced during the racking process.
Moreover, during alcoholic fermentation very important changes take place inthe yeast’s environment. Basically, the ethanol concentration increases progres-sively and yeasts need to adapt their plasmatic membranes to this aggressive newenvironment (Weber and Bont 1996). Apparently, the presence of ethanol in themedium alters drastically the fluidity of the membrane (Jones and Greenfield 1987).Under these conditions, Saccharomyces cerevisiae must increase its proportion ofsterols and unsaturated fatty acids to compensate for this effect and consequently

18 F. Zamora
enhance its tolerance to ethanol (Alexandre et al. 1994). In the particular case ofred winemaking, these changes can be done without problems because oxygen isintroduced during the racking process. Nevertheless, as mentioned previously, whitewines are usually made without aeration, and in that case the lack of oxygen maymake it very difficult for yeast to adapt to ethanol.
1.10 Stuck and Sluggish Fermentations: Causes and Solutions
Sometimes alcoholic fermentation becomes too slow towards the end of the pro-cess. Yeasts drastically reduce their sugar consumption and fermentation may evenstop before all the fermentable sugars have been completely metabolised. When thishappens, oenologists are faced with two problems. First, the wine is not finishedand something has to be done to finish it. Second, the risk of bacterial spoilage isvery high. Heterolactic acid bacteria can metabolize the sugars and produce highamounts of acetic acid (Ribereau-Gayon et al. 2000b). Winemakers are well awareof this problem, and for that reason they are continually concerned about the chancesof having stuck and sluggish fermentation.
The causes of stuck and sluggish fermentations have been the subject of severalstudies (Larue et al. 1982; Ingledew and Kunkee 1985; Alexandre and Charpen-tier 1998; Bisson 1999). Some of them have already been described in previouspoints. The following list summarizes the possible causes and solutions of stuckand sluggish fermentations.
1. Very high sugar concentration: Excessive sugar concentration may be a factorwhich inhibits yeasts. Moreover, during the latter stages of fermentation, anexcessive concentration of ethanol can seriously complicate the full consumptionof the sugars. This is an important problem nowadays, especially with red wines,because winemakers look for complete phenolic maturity in grapes, which isusually attained at very high sugar concentration. In this case, using yeast withhigh ethanol resistance is recommended.
2. Temperature extremes: Yeasts have problems growing when the temperature istoo low. Therefore, too low temperature at the beginning of the process can leadto a deficient yeast population. On the other hand, if the temperature is too high(more than 30 ◦C) the fermentation is at considerable risk of stopping. For that rea-son, thermic control of fermentations is nowadays indispensable. Another aspectto take into account is that sudden changes of temperature may provoke seriousproblems in fermentation. As previously mentioned, yeasts adapt their membranecomposition to maintain the correct fluidity. A drastic temperature decrease canprovoke excessive rigidity in the membranes before yeasts can adapt to it.
3. Complete anaerobiosis: Oxygen is necessary to synthesize ergosterol and MCFA.Without oxygen, yeast will struggle to grow and adapt their membranes to theenvironmental conditions. For that reason, aeration is recommended, at least dur-ing the exponential growth phase.

1 Biochemistry of Alcoholic Fermentation 19
4. Nutrient deficiencies: The lack of some nutrients in the grape juice can causeserious problems during fermentation. Nitrogen, vitamins, minerals, etc. maybe deficient in grape juice. For that reason yeast activators are usually addedin wineries. The standard activators are made up of ammonium salts (phos-phate and/or sulphate), thiamine and their application is certainly very useful.However, as previously mentioned, the nitrogen dose must be selected carefully,taking into account the initial concentration of easily assimilated nitrogen andthe potential alcoholic content of the grape juice. Adding nitrogen will be moreeffective if it is done two or more times and is combined with aeration. Thefirst dose should be added when fermentation starts, the second in the middle ofquasi-stationary phase and finally the third at the end of this stage. During recentyears a new generation of yeast activators has appeared on the market. Thesenew activators are prepared from yeast and provide several other interesting sub-stances such as sterols, UFA, minerals, panthothenic acid, etc. These activatorsare very useful during the latter stages of fermentation.
5. Presence of anti-fungal substances: Sometimes grape juice can contain pesticideresidues which can seriously affect alcoholic fermentation. To avoid this, thevineyard’s treatments must be closely inspected.
6. Presence of medium chain fatty acids: The presence of MCFA can decrease yeastviability and even stop alcoholic fermentation. This problem is more prevalentin white winemaking because fermentation is usually carried out at low temper-atures and without any aeration. Yeast hulls have been very useful for avoidingthis problem. Yeast hulls adsorb MCFA from the media and provide sterols andUFA to the yeasts. Yeast hulls can be used as preventives (20 g/hl) or as curatives(40–50 g/hl) of stuck and sluggish fermentations.
7. Antagonism between microorganisms: The different microorganisms present ingrape juice compete for nutrients. Saccharomyces cerevisiae development isusually encouraged through LSA inoculation and by adding sulphur dioxide.However, sometimes autochthonous yeast or even bacteria can grow and causeorganoleptic deviations and even stuck and sluggish fermentations (Edwardset al. 1999; Gerland 2000; Gao et al. 2002). This problem is greater when thepH of grape juice is high because the antiseptic effect of sulphur dioxide is lesseffective (Beech et al. 1979).
All these causes are possible and can prevent alcoholic fermentation from devel-oping correctly. However, it is usually a synergistic combination of some of thesecauses which brings about stuck and sluggish fermentation. If these causes are elim-inated, then stuck and sluggish fermentations are almost non-existent. Nevertheless,if a fermentation tank presents problems, winemakers must act as soon as possi-ble. Abundant aeration and adding yeast hulls may solve the problem. Finally, iffermentation stops, the yeast must be reinoculated. The choice of the yeast andthe way it is preadapted to ethanol are the key to the success of the inoculum.The use of commercial yeast with a high resistance to ethanol and the use of theclassical protocols for adapting yeast to the champagnization process are highlyrecommended.

20 F. Zamora
1.11 Other Subproducts of Alcoholic Fermentation
As mentioned previously, fermentation produces mainly ethanol and glycerol butalso several other substances that contribute to their complexity (Lambrechts andPretorius 2000). Figure 1.9 shows schematically the metabolic origin of some of themost important subproducts of alcoholic fermentation.
1. Diacetyl, acetoin and 2,3-butanediol: These compounds are produced by con-densing of pyruvate with ethanal. This reaction produces acetolactate whichis later decarboxylated. Diacetyl is produced if the decarboxylation is oxida-tive, whereas acetoin is produced if the decarboxylation is not oxidative. Ace-toin can also be formed by directly reducing diacetyl. Finally, acetoin can bereduced to form 2,3-butanediol. This last reaction is reversible (Ribereau-Gayonet al. 2000c). Acetoin and especially diacetyl give off a buttery smell that may
Hexosetransporter
PyruvateEthanal
Acetolactate
Diacetyle
Acetoine
2,3-Butanediol
Sugars
NAD +
NADH+ H+
Acetyl -coA
Aceticacid
HScoAAmino acid
H+Permease
Keto acids
Fattyacyl-coA
Fattyacids
Higheralcohols
Esters
NAD +NADH+ H+
Ethanol
Esters PartiallyOperative
KrebsCycle
Succinicacid
Grape juice
Hexosetransporter
Hexosetransporter
PyruvateEthanalEthanal
Ethanol
Acetolactate
Diacetyle
Acetoine
2,3-Butanediol
Sugars Sugars
NAD
NADH+ H
NAD+
NADH+ H+
Diacetyle
Acetoine
2,3-Butanediol
Acetyl-coA
Aceticacid
HScoA
Aceticacid
Aminoacid
H+
Amino acid
H+PermeasePermease
Keto acids
Fattyacyl-coA
Fatty acidsFattyacids
Higheralcohols
Esters
NAD+
NADH+ H+
Higheralcohols
Esters
Ethanol
Esters Esters PartiallyOperative
KrebsCycle
PartiallyOperative
KrebsCycle
Succinicacid
Succinicacid
Other acids
Fig. 1.9 Biosynthesis of the other subproducts

1 Biochemistry of Alcoholic Fermentation 21
contribute to a wine’s aroma. Although alcoholic fermentation does not produceenough acetoin and diacetyl to have a real impact on wine aroma, the subsequentdevelopment of lactic acid bacteria can considerably increase its concentration.
2. Ethanal: Ethanal, also called acetaldehyde, is an intermediary of alcoholic fer-mentation obtained by the decarboxylation of pyruvate. Later, ethanal is mainlyreduced to ethanol, but little quantities of it may be released into the wine.Ethanal gives off a characteristic aroma that contributes to the perception that thewine is oxidized, although alcoholic fermentation usually produces only smallamounts of this compound. However, ethanal can also be produced from ethanolby chemical or biological oxidation. Some wines, such as Fino and Manzanillafrom Jerez, or the wine Jaune from Jura, give off a particular aroma which ischaracterized by a high concentration of ethanal. These wines are obtained byaging the wine under a film of Saccharomyces cerevisiae, which is responsiblefor producing ethanal from ethanol.
3. Acetic acid: Acetic acid is the main volatile acid of wine. Its presence at highconcentrations gives off a vinegar odour and a disagreeable sensation in themouth. For that reason, volatile acidity is one of the most important analyti-cal parameters in oenology. Acetic acid may be produced by yeast, lactic acidbacteria and acetic acid bacteria. But normally Saccharomyces cerevisiae onlyproduce small quantities of acetic acid if there are no problems during alcoholicfermentation (0.1–0.3 g/l). However, stuck and sluggish fermentations can gen-erate large amounts of this acid. The high production of acetic acid may be dueto the development of lactic disease or because yeasts produce more acetic acidthan normal by hydrolysis of acetyl-coA.
4. Higher alcohols: Higher alcohols are produced as a deviation of the metabolismof amino acids. Higher alcohols are produced when keto acids correspondingto the carbon skeleton from the different amino acids are decarboxylated andreduced. Higher alcohols are normally below their limit of detection but they arethe precursors of some esters, which have a large sensory impact.
5. Esters: Esters are synthesized from acyl-coA and alcohols by a group of enzymes,the alcohol-acylcoA transferases (Barre et al. 1998). Basically there are two typesof esters in wine: the acetates of higher alcohols and the esters of fatty acids andethanol. The first group are synthesised from acetyl-coA and the different higheralcohols. These esters give off different odours, such as glue (ethyl acetate),banana (isoamyl acetate) or rose (phenylethanol acetate). The other group ofesters are synthesised from the different acyl-coA and ethanol. The differentesters of fatty acids and ethanol give off a fruity aroma. All esters, with theexception of ethyl acetate, give off an agreeable smell and contribute positivelyto the wine aroma. Other esters such as ethyl lactate and diethyl succinate do nothave any sensory impact at normal concentrations.
6. Succinic acid: Succinic acid is quantitatively the third product of alcoholic fer-mentation. Some authors suggest that succinic acid is synthesised via Krebs cyclealthough its functioning is seriously limited (Heerde and Radler 1978). However,other authors think that this cycle is not operative in conditions of grape juicefermentation (Salmon et al. 1987). In any case, succinic acid is present in wine

22 F. Zamora
in concentrations between 0.6 and 1.2 g/l and it contributes significantly to thewine acidity. Saccharomyces cerevisiae also releases into the wine several otheracids such as lactic acid, isovaleric and isobutyric acids, fatty acids, etc., but onlyin low concentrations.
It can be concluded that alcoholic fermentation is not only the simple transformationof sugars into ethanol. On the contrary, it is a very complex process that allows usto obtain a very pleasant beverage. It represents the transformation of sugars mainlyinto ethanol but also into other subproducts, which can contribute positively or neg-atively to sensory quality. A more complete knowledge about the regulation of allpathways implied that it is necessary to acquire a better understanding of the processand of designing strategies to enhance the quality of the product. Moreover, alco-holic fermentation also implies the transformation of other compounds present inthe grape juice which have a high influence on wine quality. Considerable progresshas been made during recent years and, without doubt, new solutions for new chal-lenges will be developed in the immediate future (Pretorius 2000; Moreno-Arribasand Polo 2005).
Acknowledgments The author gratefully acknowledges CICYT (AGL2007-66338 and AGL2004-02309) for financial support.
References
Aerny, J. (1997) Composes azotes des mouts et des vins. Rev. Suisse Vitic. Hortic., 28, 161–165.Alexandre, H., & Charpentier, C. (1998) Biochemical aspects of stuck and sluggish fermentation
in grape must. J. Ind. Microbiol. Biotechnol., 20, 20–27.Alexandre, H., Rousseaux, I., & Charpentier, C. (1994) Relationship between ethanol tolerance,
lipid composition and plasma membrane fluidity in Saccharomyces cerevisiae and Kloeckeraapiculata. FEMS Microbiol. Lett., 124, 17–22
Barnett, J.A. (2003) A history of research on yeasts 5: the fermentation pathway. Yeast, 20,509–543.
Barnett, J.A., & Entian, K.D. (2005) A history of research on yeasts 9: regulation of sugarmetabolism. Yeast, 22, 835–894.
Barre, P., Blondin, B., Dequin, S., Feuillat, M., Sablayrolles, J.M., & Salmon, J.M. (1998). Lalevure de fermentation alcoolique. In C. Flanzy (Ed.), Oenologie: fondements scientifiques ettechnologiques (pp. 454–497). Paris: Tec Doc Lavoisier.
Beech, F.W., Burroughs, L.F., Timberlake, C.F., & Whiting, G.C. (1979) Progres recents surl’aspect chimique et antimicrobienne de l’ahidride sulfureux. Bull. OIV, 52, 1001–1022.
Beltran, G., Torija, M.J., Novo, M., Ferrer, N., Poblet, M., Guillamon, J.M., Rozes, N., & Mas, A.(2002) Analysis of yeast populations during alcoholic fermentation: a six year follow-up study.Syst. Appl. Microbiol., 25, 287–293.
Bertrand, A., & Miele, A. (1984) Influence de la clarification du mout de raisin sur sa teneur enacides gras. Conn. Vigne Vin, 48, 293–297.
Bisson, L.F. (1999) Stuck and sluggish fermentations. Am. J. Enol. Vitic., 50, 107–119.Bisson, L.F., & Butzke, C.E. (2000) Diagnosis and rectification of stuck and sluggish fermenta-
tions. Am. J. Enol. Vitic., 51, 168–177.Boulton, R.B., Singleton, V.L., Bisson, L.F., & Kunkee, R.E. (1996). Yeast and biochemistry
of ethanol fermentation. In R.B. Boulton (Ed.), Principles and Practices of Winemaking(pp. 139–172). New York: Chapman & Hall.

1 Biochemistry of Alcoholic Fermentation 23
Cabib, E., Roberts, R., & Bowers, B. (1982) Synthesis of the Yeast Cell Wall and its Regulation.Ann. Rev. Biochem., 51, 763–793.
Chatonnet, P., Dubourdieu, D., & Boidron, J.N. (1995) The influence of Brettanomyces/Dekkerasp. yeasts and lactic acid bacteria on the ethylphenol content of red wines. Am. J. Enol. Vitic.,46, 463–468.
Ciriacy, M. (1996) Alcohol dehydrogenases. In F.K. Zimmerman & K.D. Entian (Eds.), YeastSugar Metabolism: Biochemistry, Genetics, Biotechnology, and Applications (pp. 213–224).Boca Raton: CRC Press.
Constantı, M., Poblet, M., Arola, L., Mas, A., & Guillamon, J.M. (1997). Analysis of yeasts pop-ulations during alcoholic fermentation in a newly established winery. Am. J. Enol. Vitic., 48,339–344.
Constantı, M., Reguant, C., Poblet, M., Zamora, F., Mas, A., & Guillamon, J.M. (1998) Molecu-lar analysis of yeast population dynamis: Effect of sulphur dioxide and the inoculum in mustfermentation. Int. J. Food Microbiol., 41, 169–175.
Crabtree, H.D. (1929) Observations on the carbohydrate metabolism of tumours. Biochem. J., 23,536–545.
Del Nobile, M.A., D’Amato, D., Altieri, C., Corbo, M.R., & Sinigaglia, M. (2003) Modeling theyeast Growth-Cycle in a model wine system. J. Food Sci., 68, 2080–2085.
Edwards, C.G., Reynolds, A.G., Rodrıguez, A.V., Semon, M.J., & Mills, J.M. (1999) Implicationof acetic acid in the induction of slow/stuck grape juice fermentation and inhibition of yeast byLactobacillus sp. Am. J. Enol. Vitic., 50, 204–210.
Fleet, G.H. (1993) The microorganisms of winemaking – isolation, enumeration and identifica-tion. In G.H. Fleet (Ed.), Wine Microbiology and Biotechnology (pp. 1–25). Reading: HrawoodAcademic.
Fleet, G.H., & Heard, G.M. (1993) Yeast-growth during fermentation. In G.H. Fleet (Ed.), WineMicrobiology and Biotechnology (pp. 27–54). Reading: Hrawood Academic.
Gancedo, J.M. (1988) La regulation du metabolisme des sucres chez la levure. In P. Bidan &J.R. Bonneviale (Eds.), Application a l’œnologie des progres recents en microbiologie et enfermentation (pp. 133–143). Paris: OIV.
Gancedo, J.M. (1992) Carbon catabolite repression in yeast. Eur. J. Biochem., 206, 297–313.Gao, Y.C., Zhang, G., Krentz, S., Darius, S., Power, J., & Lagarde, G. (2002) Inhibition of spoliage
lactic acid bacteria by lysozyme during wine alcholic fermentation. Aust. J. Grape Wine Res.,8, 76–83.
Geneix, C., Lafon-Lafourcade, S., & Ribereau-Gayon, P. (1983) Effet des acides gras sur la via-bilite des populations de Saccharomyces cerevisiæ. C.R. Acad Sci., 296, 943–947.
Gerland, C. (2000) Gestion de la flore bacterienne lactique: enjeu important pour l’elaboration desvins de qualite. Rev. Oenol., 96, 31–36.
Heerde, E., & Radler, F. (1978) Metabolism of the anaerobic formation of succinic acid by Sac-charomyces cerevisiae. Arch. Microbiol., 117, 269–276.
Heinisch, J.J., & Rodicio, R. (1996) Fructose-1,6 biphospohate aldolase, triose phosphate iso-merase, glyceraldehide-3-phospkate deshidrogenases and phosphoglycerate mutase. In F.K.Zimmerman & K.D. Entian (Eds.), Yeast Sugar Metabolism: Biochemistry, Genetics, Biotech-nology, and Applications (pp. 119–140). Boca Raton: CRC Press.
Hensche, P.A., & Jiranek, V. (1993) Yeast – metabolism of nitrogen compounds. In G.H.Fleet (Ed.), Wine Microbiology and Biotechnology (pp. 77–164). Reading: HrawoodAcademic.
Hernandez-Orte, P., Cacho, J., & Ferreira, V. (2002) Relationship between varietal amino acid pro-file of grapes and wine aromatic composition. Experiments with model solutions and chemo-metric study, J. Agric. Food Chem., 50, 2891–2899.
Hernandez-Orte, P., Ibarz, M.J., Cacho, J., & Ferreira, V. (2006) Addition of amino acids to grapejuice of the Merlot variety: Effect on amino acid uptake and aroma generation during alcoholicfermentation. Food Chem., 98, 300–310.

24 F. Zamora
Hohmann, S. (1996) Pyruvate decarboxylases. In F.K. Zimmerman & K.D. Entian (Eds.), YeastSugar Metabolism: Biochemistry, Genetics, Biotechnology, and Applications (pp. 187–212).Boca Raton: CRC Press.
Horecker, B.L. (2002) The Pentose Phosphate Pathway. J. Biol. Chem., 277, 47965–47971.Ingledew, W.M., & Kunkee, R.E. (1985) Factors influencing sluggish fermentations of grape juice.
Am. J. Enol. Vitic., 36, 65–76.Jiranek, V., Langridge, P., & Henschke, P.A. (1995) Regulation of hydrogen sulfite liberation in
wine-producing Saccharomyces cerevisiae strains by assimilable nitrogen. Appl. Environm.Microbiol., 61, 461–467.
Jones, R.P., & Greenfield, P.F., (1987) Ethanol and the fluidity of the yeast plasma membrane.Yeast, 3, 223–232.
Kajiwara, S., Aritomi, T., Suga, K., Ohtaguchi, K., & Kobayashi, O. (2000) Overexpression ofthe OLE1 gene enhances ethanol fermentation by Saccharomyces cerevisiae. Appl. Microbiol.Biotechnol., 53, 568–574.
Kresge, N., Simoni, R.D., & Hill, R.L. (2005) Otto Fritz Meyerhof and the Elucidation of theGlycolytic Pathway. J. Biol. Chem., 280, 124–126.
Kunkee, R.E. (1991). Relationship between nitrogen content of must and sluggish fermentation. InProceedings of the International Symposium of Nitrogen in Grapes and Wine, 18–19 de Junyde 1991, Seattle, Washington (pp. 148–155). Davis CA: American Society of Enology andViticulture.
Lafon-Lafourcade, S. (1983). Wine and brandy. Biotechnology, In H.J. Rehm & G. Reed. (Eds.),Food and Feed Production with Microorganisms, Vol 5, (pp. 81–163). Weinheim: VerlagChemie.
Lafon-Lafourcade, S., & Peynaud, E. (1974) Sur l’action antibacterienne de l’anhidride sulfureuxsous forme libre et sous forme combinee. Conn. Vigne Vin, 8, 187–203.
Lafon-Lafourcade, S., Geneix, C., & Ribereau-Gayon, P. (1984). Inhibition of alcoholic fermen-tation of grape must by fatty acids produced by yeasts and their elimination by yeast ghosts.Appl. Environm. Microbiol., 47, 1246–1249.
Lagunas, R. (1993) Sugar transport in Saccharomyces cerevisiae. FEMS Microbiol. Rev., 16,229–242.
Lagunas, R., & Gancedo, C. (1983) Role of phosphate in the regulation of the Pasteur effect inSaccharomyces cerevisiae. Eur. J. Biochem., 137, 479–483.
Lagunas, R., Dominguez, C., Busturia, A., & Saez, M.J. (1982) Mechanisms of appearance of thePasteur effect in Saccharomyces cerevisiae: inactivation of sugar transport systems. J. Bacte-riol., 152, 19–25
Lambrechts, M.G., & Pretorius, S. (2000) Yeast and its importance to wine aroma – A Review.South Afric. J. Enol. Vitic., 21, 97–128.
Laroche, C., Beney, L., Marechal, P.A., & Gervais, P. (2001) The effect of osmotic pressure on themembrane fluidity of Saccharomyces cerevisiae at different physiological temperatures. Appl.Microbiol. Biotechnol., 56, 249–254.
Larue, F., Lafon-Lafourcade, S., & Ribereau-Gayon, P. (1982). Inhibition de Saccharomyces cere-visiae dans le mout de raisin. C.R. Acad. Sci., 294, 587–590.
Los, D.A., & Murata, N. (2004) Membrane fluidity and its roles in the perception of environmentalsignals. Biochim. Biophy. Acta, 1666, 142–157.
Manginot, C., Roustan, J.L., & Sablayrolles, J.M. (1998) Nitrogen demand of different yeaststrains during alcoholic fermentation. Importance of stationary phase. Enz. Micro. Technol., 23,511–517.
Meijer, M.M.C., Boonstra, J., Verkleij, A.J., & Verrips, C.T. (1998) Glucose repression in Saccha-romyces cerevisiae is related to the glucose concentration rather than the glucose flux. J. Biol.Chem., 273, 24102–24107.
Moreno-Arribas, M.V., & Polo, M.C. (2005) Winemaking biochemistry and microbiology: currentknowledge and future trends. Crit. Rev. Food Sci. Nutr., 45, 265–286.
Mortimer, R., & Polsinelli, M. (1999). On the origin of wine yeast. Res. Microbiol., 150, 199–204.

1 Biochemistry of Alcoholic Fermentation 25
Nakagawa, Y., Sakumoto, N., Kaneko, Y., & Harashima, S. (2002) Mga2p is a putative sensorfor low temperature and oxygen to induce OLE1 transcription in Saccharomyces cerevisiae.Biochem. Biophys. Res. Comm., 291, 707–713.
Nes, D.W., Janssen, G.G., Crumley, F.G., Kalinowska, M., & Akihisa, T. (1993) The structuralrequirements of sterols for membrane function in Saccharomyces cerevisiae. Arch. Biochem.Bioph., 300, 724–733.
Neuberg, C. (1946). The Biochemistry of Yeast. Ann. Rev. Biochem., 15, 435–472.Ough, C.S., (1964). Fermentation rates of juice.I. Effects of temperature and composition on white
juice fermentation rates. Am. J. Enol. Vitic., 15, 167–177.Pasteur, L. (1861) Influence de l’oxygene sur le developpement de la levure et la fermentation
alcoolique. Bulletin de la Societe de Paris (Resume de Seance du 28 juin 1861), 79–80.Peynaud, E., & Domercq, S. (1959) A review of microbiological problems in winemaking in
France. Am. J. Enol. Vitic., 1, 69–77.Polakis, E.S., Bartley, W., & Meek, G.A. (1965) Changes in the activities of respiratory enzymes
during the aerobic growth of yeast on different carbon sources. Biochem. J., 97, 298–302.Pretorius, I.S. (2000) Tailoring wine yeast for the new millennium: novel approcaches to the ancient
art of winemaking. Yeast, 16, 675–729.Pretorius, I.S., Van der Westhuizen, T.J., & Augustyn, O.P.H. (1999) Yeast biodiversity in vineyards
and wineries and its importante to the South African wine industry. S. Afr. J. Enol. Vitic., 20,61–74.
Prior, B.A., & Hohmann, S. (1996) Glycerol production and osmoregulation. In F.K. Zimmerman& K.D. Entian (Eds.), Yeast Sugar Metabolism: Biochemistry, Genetics, Biotechnology, andApplications (pp. 313–338). Boca Raton: CRC Press.
Quinn, A.J., & Chapman, D. (1980) The dynamics of membrane structure. CRC Crit. Rev.Biochem., 8, 1–117.
Racker, E. (1974) History of the Pasteur effect and its pathobiology. Mol. Cell. Biochem., 5, 17–23.Ratledge, C., & Evans, C.T. (1989) Lipids and their metabolism. In A.H. Rose & J.S. Harrison
(Eds.), The Yeasts (2nd ed.), Vol 3 (pp. 367–455). London: Academic Press.Ribereau-Gayon, P., Glories, Y., Maujean, A., & Dubourdieu, D. (2000a) Chemical nature, origins
and consequences of the main organoleptic defects. In P. Ribereau-Gayon (Ed.), Handbook ofEnology, Vol 2, (pp. 209–253). Chichester: John Wiley & sons, Ltd.
Ribereau-Gayon, P., Glories, Y., Maujean, A., & Dubourdieu, D. (2000b) Conditions of yeastdevelopment. In P. Ribereau-Gayon (Ed.), Handbook of Enology, Vol 2, (pp. 75–107). Chich-ester: John Wiley & sons, Ltd.
Ribereau-Gayon, P., Glories, Y., Maujean, A., & Dubourdieu, D. (2000c) Biochemistry of alcoholicfermentation and metabolic pathways of wine yeasts. In P. Ribereau-Gayon (Ed.), Handbookof Enology, Vol 1, (pp. 51–74). Chichester: John Wiley & sons, Ltd.
Rodrıguez, S., Sanchez, A., Martınez, J.M., Prieto, J.A., & Randez, F. (2007) Fluidization of Mem-brane Lipids Enhances the Tolerance of Saccharomyces cerevisiae to Freezing and Salt Stress.Appl. Environm. Microbiol., 73, 110–116.
Romano, P., & Suzzi, G. (1993) Sulfur dioxide and wine microorganisms. In G.H. Fleet (Ed.), WineMicrobiology and Biotechnology (pp. 373–393). Reading: Harwood Academic.
Rozes, N. (1992) Contribution a l’etude du metabolisme lipidique de Saccharomyces cerevisiæ.Application a la differenciation des levures de vin. These de Doctorat de l’Universite de Bor-deaux II, Bordeaux, France.
Sablayrolles, J.M., & Barre, P. (1986) Evaluation des besoins en oxygen de fermentationsalcooliques en conditions oenologiques silees. Sci. Aliments, 6, 373–383.
Sablayrolles, J.M., Dubois, C., Manginot, C., Roustan, J.L., & Barre, P. (1996). Efectiveness ofcombined ammoniacal nitrogen and oxygen additions for completion of sluggish and stuckfermentation. J. Fermen. Bioeng., 82, 377–381
Salmon, J.M., Vezinhet, F., & Barre, P. (1987) Anabolic role of L-malic acid in Saccharomycescerevisiae in anaerobiosis during alcoholic fermentation. FEMS Microbiol. Lett., 42, 213–220.

26 F. Zamora
Sapis-Domerq, S. (1980) Etude de l’influence des produits de traitement de la vigne sur lamicroflore des raisins et des vins. Conn. Vigne Vin, 14, 155–181.
Schaaf-Gersteenschalager, I., & Miosga, T. (1996) The pentose phosphate pathway. In F.K. Zim-merman & K.D. Entian (Eds.), Yeast Sugar Metabolism: Biochemistry, Genetics, Biotechnol-ogy, and Applications (pp. 271–284). Boca Raton: CRC Press.
Taylor, W.H. (1957) Formol Titration: An evaluation of its various modifications. Analyst, 82,488–498.
Thurston, P.A., Taylor, R., & Ahvenainen, J. (1981). Effects of linoleic acid supplements on thesynthesis by yeast of lipids and acetate esters. J. Inst. Brew., 87, 92–95.
Torrija, M.J., Beltran, G., Novo, M.T., Poblet, M., Guillamon, J.M., Mas, A., & Rozes, N. (2003)Effect of fermentation temperature and Saccharomyces species on the cell fatty acid composi-tion and presence of volatile compounds in wine. Int. J. Food Microbiol., 85, 127–136.
Weber, F.J., & Bont, J.A.M., (1996) Adaptation mechanisms of microrganisms to the toxic effectsof organic solvents on membranes. Biochim. Biophys. Acta, 1286, 225–245.
Zamora, F. (2004) Las paradas de fermentacion. Enologos, 29, 28–32.




















