Live-borns from XX but not XY oocytes in the chimeric mouse ovary composed of B6.Y(TIR) and XX cells
9
BIOLOGY OF REPRODUCTION 58, 574-582 (1998) Live-Borns from XX but Not XY Oocytes in the Chimeric Mouse Ovary Composed of B6.Y T' R and XX Cells' Asma Amleh and Teruko Taketo 2 Urology Research Laboratory and Department of Biology, McGill University, Royal Victoria Hospital, Montreal, Quebec, Canada H3A A ABSTRACT When the Y chromosome of some Mus musculus domesticus subspecies is placed onto a C57BL/6J mouse background, the XY (B6.YT'R) progeny develop only ovaries or ovotestes during fetal life. The XY sex-reversed female is infertile mainly because of death of embryos during preimplantation development. In the present study, we constructed female mouse chimera composed of B6.YT r' and XX BALB/c cells to determine whether develop- mental incompetence of XY oocytes can be attributed to defects in the oocytes themselves or in the surrounding XY somatic cells. Distribution of XY cells in chimeric ovaries was examined by in situ hybridization. Of nine XX - XY chimeric females born, eight were composed of B6.YT'R and XX BALB/c cells with a wide range of XY contribution (16-95%), whereas one had 12% XY components of the BALB/c strain. All these females produced progeny exclusively derived from XX oocytes. By comparison, most XX e XX chimeric females produced progeny derived from oocytes of either strain. Two XY <-> XY males also produced prog- eny of both strains. In conclusion, the XY chromosomal com- position in the oocyte appears to be responsible for program- ming its incompetence for postfertilization development. On the other hand, the presence of XY somatic cells in the chimeric ovary allows development of fertile XX oocytes. INTRODUCTION In mammals, a gene on the Y chromosome, named SRY in humans and Sry in mice, appears to be sufficient to initiate a cascade of genetic events leading to testicular organization, whereas its absence results in development of ovaries [1-4]. Once gonadal sex is established, primordial germ cells that have migrated into testicular and ovarian primordia undergo sexual differentiation towards spermatogenesis and oogenesis, respectively, regardless of their own chromosomal sex. While later stages of spermatogenesis require XY sex chromosomal compositions [5-8], oogenesis appears to proceed with variant sex chromosomal compositions in the mouse. For example, XO female mice are invariably fertile although they have a shorter reproductive life than normal XX female mice [9-11]. Some mutant strains of XY females, sex-reversed due to de- letion or perhaps silencing of the Sry gene, also produce lit- ters, although their fertility is impaired to varying extents [12- 15]. An exception is the sterile B6.Y T IR female mouse (pre- viously called B6.YI°M). The B6.YTIR mouse strain has been established by placing the Y chromosome of a Mus musculus domesticus (DOM) mouse (caught in Tirano, Italy) onto the C57BL/6J (B6) in- bred genetic background by repeated backcross [16]. Similar sex reversal was originally described by Eicher et al. [17, 18] using Mus poschiavinus (caught in Poschiavo, Switzerland). Accepted October 6, 1997. Received July 16, 1997. 'Grant support: MRC, Canada (MT-10684). 2Correspondence: Teruko Taketo, Urology Res. Lab., Royal Victoria Hospital, 687 Pine Avenue West, Montreal, PQ, Canada H3A 1Al. FAX: (514) 843-1457; e-mail: [email protected] Despite the presence of the intact Y chromosome and tran- scription of the Sry gene at a normal onset time, the B6.YTIR gonadal primordium differentiates into only an ovary or an ovotestis during fetal life [19]. We have speculated that the Sry gene requires coordination with another gene(s) on an autosome (or the X chromosome) downstream of Sry to ini- tiate normal testicular differentiation. Eicher's group has pos- tulated a dominant Tda (testis-determining autosomal) allele in the DOM strain and a recessive tda allele in the B6 strain, which coordinates with SryB6 but not with SryD ° M [17, 18, 20]. It has been further proposed that the tda trait may involve more than one autosomal locus, probably four or five [21- 24]. Polymorphisms identified in the Sry sequence may be responsible for the coordination with tda alleles [25]; however, no consistent sequence difference has yet been identified to explain the sex reversal involving a variety of DOM subspe- cies [26]. In the B6.YTIR strain, the fetal ovotestis usually develops into a small testis by puberty, and the majority of XY males and some hermaphroditic males become fertile. On the oth- er hand, the XY female with bilateral ovaries fails to pro- duce litters [27]. The eggs ovulated from the XY ovary undergo fertilization efficiently but cannot develop beyond the 2-cell stage either in vivo or in vitro [28]. Isolation of oocytes directly from the juvenile XY ovary and their mat- uration and fertilization in vitro slightly improve the sub- sequent embryonic development, although very few reach the blastocyst stage [29]. Hence, developmental incompe- tence of oocytes appears to be programmed during ovarian differentiation in the B6.YTIR female. We have previously reported that the pituitary of the XY female functions nor- mally in the presence of grafted XX ovaries [27]. In the present study, we constructed chimeric females composed of B6.YTIR and XX cells and asked whether de- velopmental incompetence of XY oocytes can be attributed to their XY chromosomal composition or the surrounding XY ovarian cells. We have previously reported several ab- normal aspects of the XY ovary during development [27, 30]. However, it remained to be clarified whether any of the abnormalities recognized is responsible for the failure of the oocytes in postfertilization development. It was also of interest to find out whether introduction of XX cells corrects some of the abnormal features. It has been well documented that the majority of XX -4 XY chimera develop bilateral testes with normal male phe- notype [31-35], and hence the opportunity to study the con- tribution of XY cells, particularly XY germ cells, to ovarian development has been limited [36-39]. Since half of the B6.YTIR progeny develop bilateral ovaries and female phe- notype, the chimerism with XX cells may favor female de- velopment. Therefore, the mouse chimera composed of B6.YTIR and XX cells gives us an excellent opportunity to examine the interaction between XX and XY cells during ovarian development, even though the B6.YTIR fetal gonad lacks normal testis-determining function. 574
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Live-borns from XX but not XY oocytes in the chimeric mouse ovary composed of B6.Y(TIR) and XX cells

BIOLOGY OF REPRODUCTION 58, 574-582 (1998)
Live-Borns from XX but Not XY Oocytes in the Chimeric Mouse Ovary Composedof B6.Y T'R and XX Cells'
Asma Amleh and Teruko Taketo2
Urology Research Laboratory and Department of Biology, McGill University, Royal Victoria Hospital, Montreal,Quebec, Canada H3A A
ABSTRACT
When the Y chromosome of some Mus musculus domesticussubspecies is placed onto a C57BL/6J mouse background, theXY (B6.YT'R) progeny develop only ovaries or ovotestes duringfetal life. The XY sex-reversed female is infertile mainly becauseof death of embryos during preimplantation development. In thepresent study, we constructed female mouse chimera composedof B6.YTr' and XX BALB/c cells to determine whether develop-mental incompetence of XY oocytes can be attributed to defectsin the oocytes themselves or in the surrounding XY somatic cells.Distribution of XY cells in chimeric ovaries was examined by insitu hybridization. Of nine XX - XY chimeric females born,eight were composed of B6.YT'R and XX BALB/c cells with a widerange of XY contribution (16-95%), whereas one had 12% XYcomponents of the BALB/c strain. All these females producedprogeny exclusively derived from XX oocytes. By comparison,most XX e XX chimeric females produced progeny derived fromoocytes of either strain. Two XY <-> XY males also produced prog-eny of both strains. In conclusion, the XY chromosomal com-position in the oocyte appears to be responsible for program-ming its incompetence for postfertilization development. On theother hand, the presence of XY somatic cells in the chimericovary allows development of fertile XX oocytes.
INTRODUCTION
In mammals, a gene on the Y chromosome, named SRYin humans and Sry in mice, appears to be sufficient to initiatea cascade of genetic events leading to testicular organization,whereas its absence results in development of ovaries [1-4].Once gonadal sex is established, primordial germ cells thathave migrated into testicular and ovarian primordia undergosexual differentiation towards spermatogenesis and oogenesis,respectively, regardless of their own chromosomal sex. Whilelater stages of spermatogenesis require XY sex chromosomalcompositions [5-8], oogenesis appears to proceed with variantsex chromosomal compositions in the mouse. For example,XO female mice are invariably fertile although they have ashorter reproductive life than normal XX female mice [9-11].Some mutant strains of XY females, sex-reversed due to de-letion or perhaps silencing of the Sry gene, also produce lit-ters, although their fertility is impaired to varying extents [12-15]. An exception is the sterile B6.Y TIR female mouse (pre-viously called B6.YI°M).
The B6.YTIR mouse strain has been established by placingthe Y chromosome of a Mus musculus domesticus (DOM)mouse (caught in Tirano, Italy) onto the C57BL/6J (B6) in-bred genetic background by repeated backcross [16]. Similarsex reversal was originally described by Eicher et al. [17, 18]using Mus poschiavinus (caught in Poschiavo, Switzerland).
Accepted October 6, 1997.Received July 16, 1997.'Grant support: MRC, Canada (MT-10684).2Correspondence: Teruko Taketo, Urology Res. Lab., Royal Victoria
Hospital, 687 Pine Avenue West, Montreal, PQ, Canada H3A 1Al. FAX:(514) 843-1457; e-mail: [email protected]
Despite the presence of the intact Y chromosome and tran-scription of the Sry gene at a normal onset time, the B6.YTIRgonadal primordium differentiates into only an ovary or anovotestis during fetal life [19]. We have speculated that theSry gene requires coordination with another gene(s) on anautosome (or the X chromosome) downstream of Sry to ini-tiate normal testicular differentiation. Eicher's group has pos-tulated a dominant Tda (testis-determining autosomal) allelein the DOM strain and a recessive tda allele in the B6 strain,which coordinates with SryB6 but not with SryD° M [17, 18,20]. It has been further proposed that the tda trait may involvemore than one autosomal locus, probably four or five [21-24]. Polymorphisms identified in the Sry sequence may beresponsible for the coordination with tda alleles [25]; however,no consistent sequence difference has yet been identified toexplain the sex reversal involving a variety of DOM subspe-cies [26].
In the B6.YTIR strain, the fetal ovotestis usually developsinto a small testis by puberty, and the majority of XY malesand some hermaphroditic males become fertile. On the oth-er hand, the XY female with bilateral ovaries fails to pro-duce litters [27]. The eggs ovulated from the XY ovaryundergo fertilization efficiently but cannot develop beyondthe 2-cell stage either in vivo or in vitro [28]. Isolation ofoocytes directly from the juvenile XY ovary and their mat-uration and fertilization in vitro slightly improve the sub-sequent embryonic development, although very few reachthe blastocyst stage [29]. Hence, developmental incompe-tence of oocytes appears to be programmed during ovariandifferentiation in the B6.YTIR female. We have previouslyreported that the pituitary of the XY female functions nor-mally in the presence of grafted XX ovaries [27].
In the present study, we constructed chimeric femalescomposed of B6.YTIR and XX cells and asked whether de-velopmental incompetence of XY oocytes can be attributedto their XY chromosomal composition or the surroundingXY ovarian cells. We have previously reported several ab-normal aspects of the XY ovary during development [27,30]. However, it remained to be clarified whether any ofthe abnormalities recognized is responsible for the failureof the oocytes in postfertilization development. It was alsoof interest to find out whether introduction of XX cellscorrects some of the abnormal features.
It has been well documented that the majority of XX -4XY chimera develop bilateral testes with normal male phe-notype [31-35], and hence the opportunity to study the con-tribution of XY cells, particularly XY germ cells, to ovariandevelopment has been limited [36-39]. Since half of theB6.YTIR progeny develop bilateral ovaries and female phe-notype, the chimerism with XX cells may favor female de-velopment. Therefore, the mouse chimera composed ofB6.YTIR and XX cells gives us an excellent opportunity toexamine the interaction between XX and XY cells duringovarian development, even though the B6.YTIR fetal gonadlacks normal testis-determining function.
574

FERTILITY OF XY OOCYTES IN THE CHIMERIC MOUSE
MATERIALS AND METHODS
Chimera Production
Male mice carrying the yTIR chromosome and the B6genetic background were prepared and maintained in ourmouse colony [16, 27]. B6 females (50-150 days old; TheJackson Laboratory, Bar Harbor, ME) were caged withB6.YTIR male mice (N19-22 backcross generations) to pro-duce embryos that constituted one component of the chi-mera. Mating between BALB/c males and females (CharlesRiver, St-Constance, PQ, Canada) produced embryos to beused as the counterpart of the chimera. Each pair of em-bryos at the 4- to 8-cell stage was placed together on a dentin a plastic Petri dish and allowed to aggregate as previ-ously described [40]. Twenty-eight hours after incubation,95% of aggregates developed to blastocysts, which werethen transferred into uterine horns of pseudopregnant F1(C3H x B6) females (Charles River) at Day 2.5. Each uter-ine horn received 4-8 embryos. In early experiments, upto nine BALB/c embryos with intact zonae pellucidae weretransferred as carriers together with the aggregated embryosinto each female in an attempt to facilitate implantation[41]. We found that this precaution was unnecessary, andcarriers were not included in later experiments. At weaningages, 20-25 days postpartum (dpp), the offspring were ex-amined as to the distribution of coat color (B6, black;BALB/c, albino) and the sex of external genitalia. All ex-periments were conducted in accordance with the Guide tothe Care and Use of Experimental Animals issued by theCanadian Council on Animal Care.
Detection and Quantification of XY Components
Genomic DNA was extracted from a fragment of the tailof each chimeric mouse at weaning and adjusted to a finalconcentration of 1 ig/il. A standard was prepared by mix-ing DNA from B6.YTIR males and BALB/c females eachat 1 ig/jil at varying ratios. DNA samples (2 ig each) werethen blotted onto Hybond-N + membranes (Amersham,Oakville, ON, Canada), denatured, and hybridized with a3 2P-labeled mouse Y-specific probe 145SC5 (gift from Dr.Nishioka, Biology Department, McGill University) as de-scribed previously [27]. The membrane around each DNAdot was cut into a consistent area of 2.25 cm2 , and its 3emissions were determined by a liquid scintillation counter(RackBeta, Turku, Finland). Percentage of XY contributionin each chimeric mouse was deduced from the standardcurve. When the chimeric females were killed, DNA wasextracted from the kidney and quantified as described aboveexcept that the probe was labeled with digoxigenin (Dig),which was detected using the Dig Luminescent DetectionKit (Boehringer Mannheim, Laval, PQ, Canada). Intensityof signals on x-ray films after 12 min of exposure wasanalyzed by ScanJet II cx scanner (Hewlett Packard, Gree-ley, CO) and ImageQuant (Molecular Dynamics, Sunny-vale, CA) programs.
Polymerase Chain Reaction (PCR) Analysis of the ZfyPolymorphism
The origin of XY cells (B6.YTIR or BALB/c) was iden-tified by PCR analysis of the polymorphism in a Y-encodedgene, Zfy [42]. The B6.YTR strain carries two copies, Zfy-I and Zfy-2, with an identical molecular size at 618 basepairs (bp). On the other hand, the BALB/c strain has Zfy-1 at 618 bp and Zfy-2 at 600 bp with an 18-bp deletion. Aset of primers (sense primer: 5'-AAGATAAGCTTACAT
AATCACATGGA-3'; antisense primer: 5'-CCTATGAAATCCTTTGCTGCACATGT-3') [42] were synthesized bySheldon Biotechnology Center (McGill University, Montre-al, PQ). PCR amplification conditions were as described byNagamine et al. [42]. The amount of DNA used for eachamplification was 100 ng from most mice except for 120ng from female chimeras #2, #14, and #15, and 200 ngfrom chimeras #18, #22, #30, #31, and #32. Ten microliterseach of the PCR products was size-fractionated by 4%polyacrylamide gel electrophoresis in Tris-borate buffer(pH 8.0) and visualized by ethidium bromide staining.
Fertility of Chimeric Mice
Chimeric females at 35-45 dpp were caged withBALB/c males (100-150 dpp), and the presence of copu-lation plugs was checked for every morning. If no copu-lation plug was found in females within three days, theirmale partners were changed. Females with copulation plugsthat did not become pregnant were subjected to the aboveprocedure again.
Each XY <- XY chimeric male at 49-85 dpp was cagedwith three BALB/c females (42-90 dpp), and the presenceof copulation plugs was checked for every morning up tothree days. Any female with a copulation plug was kept ina separate cage until delivery. The male after a four-daybreak was caged again with females as above. These pro-cedures were repeated until each male copulated with atleast five females. Numbers of live-borns, as well as theireye and skin colors, were recorded at 1-2 days after deliv-ery.
Morphology of Chimeric Ovaries
Ovaries were dissected out from chimeric females at 61-157 dpp, fixed in Bouin's solution overnight, and embeddedin paraffin. Two to three serial sections of 5-,pm thicknesswere mounted on each positively charged slide (Fisher Sci-entific, Ottawa, ON, Canada). Eight sets of slides evenlydistributed between approximately 1/4 and 3/4 depth ofeach ovary were processed for staining with hematoxylinand eosin. The numbers of preantral, antral, and atretic fol-licles that encompassed oocytes, as well as the number ofcorpora lutea were recorded from one representative sectionof each slide. The area of each section was determined bymeasuring the largest and shortest diameters, assimilatingto the closest ellipse shape.
In Situ Hybridization for Detection of the Y Chromosomein Chimeric Ovaries
To analyze the distribution of XY cells in chimeric ova-ries, some representative ovarian sections were processedfor in situ hybridization as described by Musiani et al. [43]with minor changes. Briefly, sections on histology slideswere pretreated with DNase-free RNase to minimize back-ground staining and then with a 30% solution of Pretreat-ment Powder (Oncor, Gaithersburg, MD), and finally di-gested with 125 g/ml of proteinase K (Boehringer Mann-heim) to enhance permeability of the tissue. The sectionson each slide were then covered with 10 l hybridizationsolution (1.6 ng/jl of Dig-labeled 145SC5, 50% deionizedformamide, 10% dextran sulfate, double-strength SSC [sin-gle-strength SSC = 0.15 M sodium chloride, 0.015 M so-dium citrate], and 0.5 jig/jil salmon testis DNA) and de-natured by heating at 87°C for 8 min. Hybridization wasperformed overnight at 37°C, followed by two washes with
575
AMLEH AND TAKETO
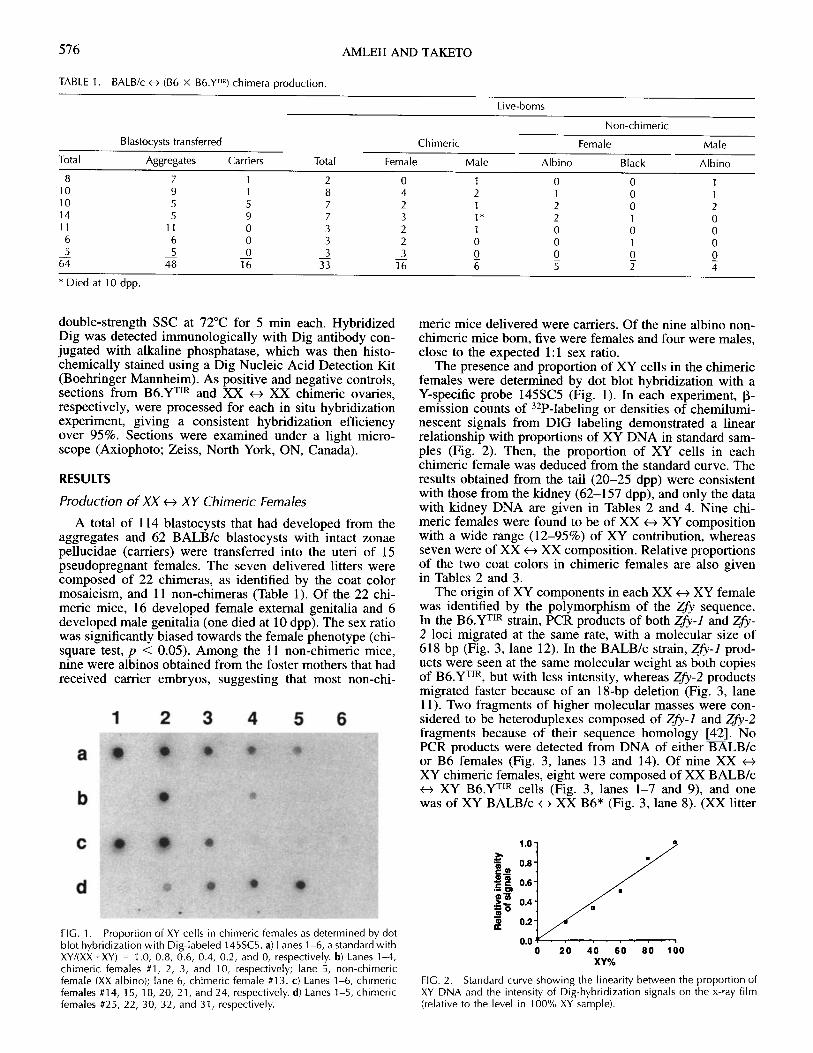
TABLE 1. BALB/c e> (B6 x B6.Y TIR) chimera production.
Live-borns
Non-chimeric
Blastocysts transferred Chimeric Female Male
Total Aggregates Carriers Total Female Male Albino Black Albino
8 7 1 2 0 1 0 0 110 9 1 8 4 2 1 0 110 5 5 7 2 1 2 0 214 5 9 7 3 1* 2 1 011 11 0 3 2 1 0 0 0
6 6 0 3 2 0 0 1 05 5 0 3 3 0 0 0 0
64 48 16 33 16 6 5 2 4
* Died at 10 dpp.
double-strength SSC at 72C for 5 min each. HybridizedDig was detected immunologically with Dig antibody con-jugated with alkaline phosphatase, which was then histo-chemically stained using a Dig Nucleic Acid Detection Kit(Boehringer Mannheim). As positive and negative controls,sections from B6.YTIR and XX <- XX chimeric ovaries,respectively, were processed for each in situ hybridizationexperiment, giving a consistent hybridization efficiencyover 95%. Sections were examined under a light micro-scope (Axiophoto; Zeiss, North York, ON, Canada).
RESULTS
Production of XX <-> XY Chimeric Females
A total of 114 blastocysts that had developed from theaggregates and 62 BALB/c blastocysts with intact zonaepellucidae (carriers) were transferred into the uteri of 15pseudopregnant females. The seven delivered litters werecomposed of 22 chimeras, as identified by the coat colormosaicism, and 11 non-chimeras (Table 1). Of the 22 chi-meric mice, 16 developed female external genitalia and 6developed male genitalia (one died at 10 dpp). The sex ratiowas significantly biased towards the female phenotype (chi-square test, p < 0.05). Among the 11 non-chimeric mice,nine were albinos obtained from the foster mothers that hadreceived carrier embryos, suggesting that most non-chi-
meric mice delivered were carriers. Of the nine albino non-chimeric mice born, five were females and four were males,close to the expected 1:1 sex ratio.
The presence and proportion of XY cells in the chimericfemales were determined by dot blot hybridization with aY-specific probe 145SC5 (Fig. 1). In each experiment, P-emission counts of 3 2 P-labeling or densities of chemilumi-nescent signals from DIG labeling demonstrated a linearrelationship with proportions of XY DNA in standard sam-ples (Fig. 2). Then, the proportion of XY cells in eachchimeric female was deduced from the standard curve. Theresults obtained from the tail (20-25 dpp) were consistentwith those from the kidney (62-157 dpp), and only the datawith kidney DNA are given in Tables 2 and 4. Nine chi-meric females were found to be of XX <-> XY compositionwith a wide range (12-95%) of XY contribution, whereasseven were of XX <-- XX composition. Relative proportionsof the two coat colors in chimeric females are also givenin Tables 2 and 3.
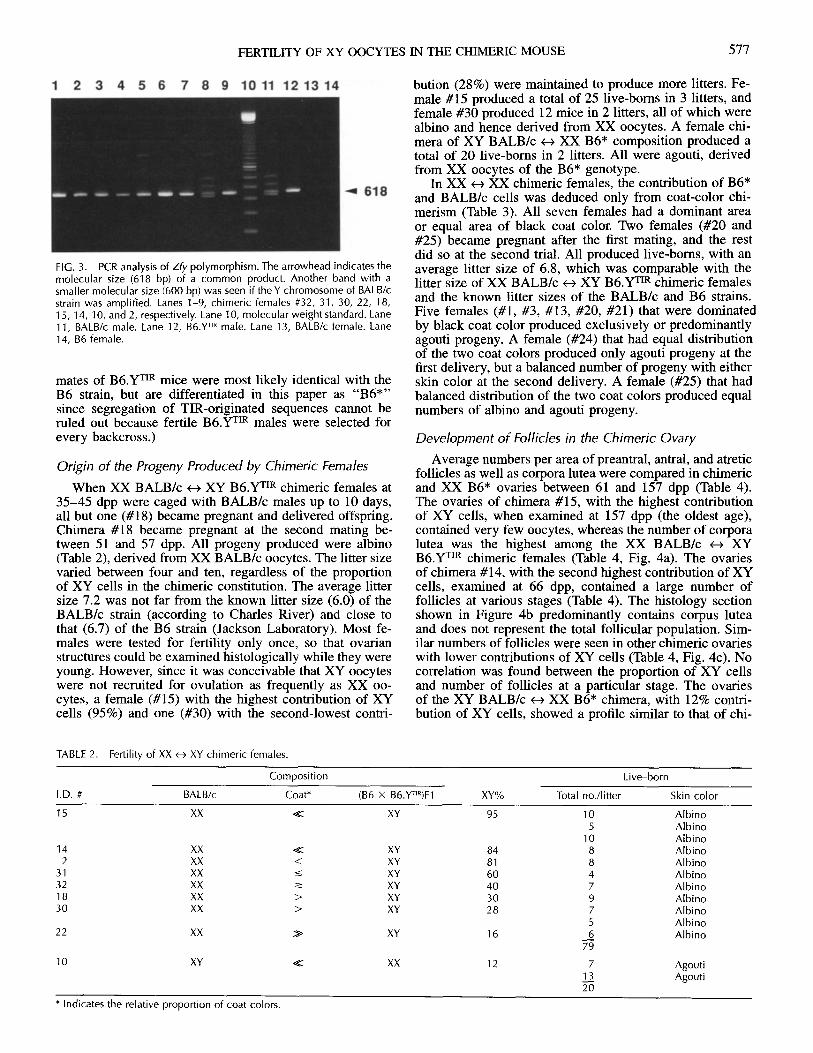
The origin of XY components in each XX <-> XY femalewas identified by the polymorphism of the Zfy sequence.In the B6.YTIR strain, PCR products of both Zfy-l and Zfy-2 loci migrated at the same rate, with a molecular size of618 bp (Fig. 3, lane 12). In the BALB/c strain, Zfy-1 prod-ucts were seen at the same molecular weight as both copiesof B6.YTIR, but with less intensity, whereas Zfy-2 productsmigrated faster because of an 18-bp deletion (Fig. 3, lane11). Two fragments of higher molecular masses were con-sidered to be heteroduplexes composed of Zfy-1 and Zfy-2fragments because of their sequence homology [42]. NoPCR products were detected from DNA of either BALB/cor B6 females (Fig. 3, lanes 13 and 14). Of nine XX <-XY chimeric females, eight were composed of XX BALB/c<- XY B6.YTIR cells (Fig. 3, lanes 1-7 and 9), and onewas of XY BALB/c t< XX B6* (Fig. 3, lane 8). (XX litter
._
-Wr
FIG. 1. Proportion of XY cells in chimeric females as determined by dotblot hybridization with Dig-labeled 145SC5. a) Lanes 1-6, a standard withXY/(XX+XY) = 1.0, 0.8, 0.6, 0.4, 0.2, and 0, respectively. b) Lanes 1-4,chimeric females #1, 2, 3, and 10, respectively; lane 5, non-chimericfemale (XX albino); lane 6, chimeric female #13. c) Lanes 1-6, chimericfemales #14, 15, 18, 20, 21, and 24, respectively. d) Lanes 1-5, chimericfemales #25, 22, 30, 32, and 31, respectively.
0 20 40 60 80 100XY%
FIG. 2. Standard curve showing the linearity between the proportion ofXY DNA and the intensity of Dig-hybridization signals on the x-ray film(relative to the level in 100% XY sample).
576
FERTILITY OF XY OOCYTES IN THE CHIMERIC MOUSE
FIG. 3. PCR analysis of Zfy polymorphism. The arrowhead indicates themolecular size (618 bp) of a common product. Another band with asmaller molecular size (600 bp) was seen if the Y chromosome of BALB/cstrain was amplified. Lanes 1-9, chimeric females #32, 31, 30, 22, 18,15, 14, 10, and 2, respectively. Lane 10, molecular weight standard. Lane11, BALB/c male. Lane 12, B6.YTR male. Lane 13, BALB/c female. Lane14, B6 female.
mates of B6.YTIR mice were most likely identical with theB6 strain, but are differentiated in this paper as "B6*"since segregation of TIR-originated sequences cannot beruled out because fertile B6.YTIR males were selected forevery backcross.)
Origin of the Progeny Produced by Chimeric Females
When XX BALB/c <-> XY B6.YTIR chimeric females at35-45 dpp were caged with BALB/c males up to 10 days,all but one (#18) became pregnant and delivered offspring.Chimera #18 became pregnant at the second mating be-tween 51 and 57 dpp. All progeny produced were albino(Table 2), derived from XX BALB/c oocytes. The litter sizevaried between four and ten, regardless of the proportionof XY cells in the chimeric constitution. The average littersize 7.2 was not far from the known litter size (6.0) of theBALB/c strain (according to Charles River) and close tothat (6.7) of the B6 strain (Jackson Laboratory). Most fe-males were tested for fertility only once, so that ovarianstructures could be examined histologically while they wereyoung. However, since it was conceivable that XY oocyteswere not recruited for ovulation as frequently as XX oo-cytes, a female (#15) with the highest contribution of XYcells (95%) and one (#30) with the second-lowest contri-
bution (28%) were maintained to produce more litters. Fe-male #15 produced a total of 25 live-borns in 3 litters, andfemale #30 produced 12 mice in 2 litters, all of which werealbino and hence derived from XX oocytes. A female chi-mera of XY BALB/c <- XX B6* composition produced atotal of 20 live-borns in 2 litters. All were agouti, derivedfrom XX oocytes of the B6* genotype.
In XX <- XX chimeric females, the contribution of B6*and BALB/c cells was deduced only from coat-color chi-merism (Table 3). All seven females had a dominant areaor equal area of black coat color. Two females (#20 and#25) became pregnant after the first mating, and the restdid so at the second trial. All produced live-borns, with anaverage litter size of 6.8, which was comparable with thelitter size of XX BALB/c -> XY B6.YTIR chimeric femalesand the known litter sizes of the BALB/c and B6 strains.Five females (#1, #3, #13, #20, #21) that were dominatedby black coat color produced exclusively or predominantlyagouti progeny. A female (#24) that had equal distributionof the two coat colors produced only agouti progeny at thefirst delivery, but a balanced number of progeny with eitherskin color at the second delivery. A female (#25) that hadbalanced distribution of the two coat colors produced equalnumbers of albino and agouti progeny.
Development of Follicles in the Chimeric Ovary
Average numbers per area of preantral, antral, and atreticfollicles as well as corpora lutea were compared in chimericand XX B6* ovaries between 61 and 157 dpp (Table 4).The ovaries of chimera #15, with the highest contributionof XY cells, when examined at 157 dpp (the oldest age),contained very few oocytes, whereas the number of corporalutea was the highest among the XX BALB/c <- XYB6.YTIR chimeric females (Table 4, Fig. 4a). The ovariesof chimera #14, with the second highest contribution of XYcells, examined at 66 dpp, contained a large number offollicles at various stages (Table 4). The histology sectionshown in Figure 4b predominantly contains corpus luteaand does not represent the total follicular population. Sim-ilar numbers of follicles were seen in other chimeric ovarieswith lower contributions of XY cells (Table 4, Fig. 4c). Nocorrelation was found between the proportion of XY cellsand number of follicles at a particular stage. The ovariesof the XY BALB/c <-> XX B6* chimera, with 12% contri-bution of XY cells, showed a profile similar to that of chi-
TABLE 2. Fertility of XX - XY chimeric females.
Composition Live-born
I.D. # BALB/c Coat* (B6 x B6.Y T'R)F1 XY% Total no./litter Skin color
15 XX < XY 95 10 Albino5 Albino
10 Albino14 XX .< XY 84 8 Albino
2 XX < XY 81 8 Albino31 XX XY 60 4 Albino32 XX _ XY 40 7 Albino18 XX > XY 30 9 Albino30 XX > XY 28 7 Albino
5 Albino22 XX > XY 16 6 Albino
79
10 XY << XX 12 7 Agouti13 Agouti20
* Indicates the relative proportion of coat colors.
577
AMLEH AND TAKETO
TABLE 3. Fertility of XX <- XX chimeric females.
Composition Live-born
I.D. # BALB/c Coat* (B6 x B6.YTIR)F1 Total no./litter Albino Agouti
3 XX XX 8 1 713 XX < XX 4 0 4
1 0 120 XX < XX 8 0 821 XX < XX 9 0 9
7 2 51 XX < XX 9 2 7
25 XX < XX 4 2 224 XX = XX 9 0 9
9 5 468 12 56
* Indicates the relative proportion of coat colors.
mera #15, i.e., depletion of follicles and predominant cor- with more XY cells than those away from the basementpora lutea (Table 4, Fig. 4f), except that two advanced an- membrane, which had an XY distribution comparable totral follicles were seen in the XY BALB/c 4 XX B6* that of the interstitium (Fig. 5, b and c). We also observedchimera but not in any section from chimera #15. that within the population of one cell type, such as granu-
The ovaries from three XX BALB/c <-> XX B6* chi- losa cells, certain areas had a higher proportion of XY cellsmeric females (#3, #13, #20) at 87, 157, and 88 dpp, re- than had other areas (Fig. 5c).spectively, were compared (Table 4, Fig. 4, d and e). They Y-Labeling was also found in oocytes of XX BALB/ccontained comparable numbers of follicles at various stages -* XY B6.YTIR chimeric ovaries. In the ovary (#15) withirrespective of age, although the number was slightly lower the highest contribution of XY cells, all three preantral fol-than in the majority of XX BALB/c <- XY B6.YTIR ova- licles identified contained oocytes with hybridization sig-ries. The number of corpora lutea varied with a range sim- nals. In the ovaries (#14 and #2) with a high contributionilar to that in XX BALB/c t+ XY B6.YTIR ovaries. Similar of XY cells, hybridization signals were seen in the majoritynumbers of follicular structures were also observed in the of oocytes encompassed by follicles from primary to antralovaries of an XX B6* female mouse. stages (Fig. 5b). In other chimeric ovaries with a lower
contribution of XY cells, detection of Dig labeling in oo-Distribution of XY Cells in XX -> XY Chimeric Ovaries cytes was less frequent than expected from the contribution
of XY somatic cells. In the ovaries of XY BALB/c <-- XXDNA-to-DNA in situ hybridization indicated that XY B6* chimera, Y labeling was found in various ovarian so-
cells were distributed in all ovarian somatic cell types, i.e., matic cell types in agreement with the proportion (12%) ofsurface epithelium, interstitium, theca cells, and both mural XY cells in nongonadal tissues, except that the surface ep-and cumulus granulosa cells in XX +4 XY chimeric ovaries ithelium appeared to be more populated with XY cells (not(Fig. 5, a-c). Overall contribution of XY cells agreed with shown). No oocyte was found labeled. No hybridizationthe degree of chimerism in the tail and the kidney as well signal was found in negative control, XX BALB/c XXas the coat color. Furthermore, the proportion of XY cells B6* chimeric ovaries (Fig. d).did not differ among follicles at different developmentalstages (Fig. 5a). However, we found considerable differ- Origin of the Progeny Produced by Chimeric Malesences in the frequency of Y-positive cells among ovariancell types. A higher proportion of XY cells was consistently Of the five chimeric males born, two were found to befound in the surface epithelium, granulosa cells, and cor- composed of XY BALB/c e-> XY B6.YTIR cells. Both malespora lutea compared with interstitial cells (Fig. 5, a-c). The- copulated with BALB/c females, producing a mixed pop-ca cells closer to the basement membrane were populated ulation of albino and agouti progeny (Table 5). The mean
TABLE 4. Follicular development in the chimeric and control ovaries.
Follicular structures (No./cm2)Age
Ovary genotype Chimera # XY % (dpp) PA An At CL
XX BALB/c <-- XY B6.YT'R 15 95 157 2.2 0.7 0.2 6.214 84 66 7.2 2.2 0.2 3.62 81 66 7.5 1.5 2.9 3.8
31 60 61 8.0 1.7 1.6 2.432 40 61 9.8 2.0 1.7 4.718 30 75 3.3 1.6 1.4 1.930 28 118 11.5 3.1 1.3 4.722 16 62 4.1 4.1 2.1 4.6
XY BALB/c <-> XX B6* 10 12 141 0.5 0.4 0 7.0XX BALB/c <--> XX B6* 3 0 87 4.8 1.1 2.5 6.1
13 0 157 5.8 1.9 1.2 4.020 0 88 6.0 1.4 1.4 3.2
XX B6* Non-chimera 0 87 5.4 1.2 2.3 4.5
a PA, preantral follicles; An, antral follicles; At, atretic follicles; CL, corpora lutea.
578

FERTILITY OF XY OOCYTES IN THE CHIMERIC MOUSE
FIG. 4. Light micrographs of chimeric ovaries with hematoxylin and eosin staining. Scale bar indicates 1 mm. a) XX BALB/c e> XY B6.YT'R chimericovary (#15) with the highest contribution of XY cells at 157 dpp. Corpora lutea occupy the most area, whereas only a few preantral and atretic folliclesare seen. b) XX BALB/c -> XY B6.YTIR chimeric ovary (#14) with a high contribution of XY cells at 66 dpp. Corpora lutea occupy the most area whereassmall numbers of preantral, antral, and atretic follicles are seen. c) XX Balb/c -> XY B6.YTIR chimeric ovary (#18) with a low contribution of XY cellsat 75 dpp. Note the presence of a large number of follicles. Representative preantral, antral, and atretic follicles as well as a corpus luteum are indicated.d) XX BALB/c -> XX B6* chimeric ovary (#13) at 157 dpp. Progressive developmental stages of follicles are seen among a large area of corpora lutea.Representative preantral, antral, and atretic follicles are indicated. e) XX BALB/c <- XX B6* chimeric ovary (#3) at 87 dpp. Corpora lutea, antral, andrepresentative preantral follicles are indicated. f) XY BALB/c -> XX B6* chimeric ovary (#10) at 141 dpp. Corpora lutea occupy the most area exceptfor two advanced antral follicles. * corpora lutea; PA, preantral follicles; At, atretic follicles; arrowheads, antral follicles.
litter size (7.4) was not different from the known litter size(6.0) of the BALB/c female. The ratio of albino (BALB/c-derived) to agouti (B6.YrTR-derived) progeny varied fromlitter to litter even with litters from the same father. Overall,chimera #5 produced equal numbers, whereas #6 producedmore albino progeny than agouti progeny despite its blackdominant coat color.
DISCUSSION
Predominant Development of Female Chimeras
In the present study, all but one (89%) of the live-bornchimeras, which were composed of B6.YTJR and XXBALB/c cells in varying proportions, developed the femalephenotype. The only exception was a male with bilateraltestes. This strong bias towards the female sex was unex-pected. When chimeric mice are composed of XX and XYcells, the XY cells usually exert a dominating effect onsexual differentiation [32, 33]. Singh et al. [44] have re-
ported that as little as 10% contribution of XY cells is suf-ficient to induce testis formation. Thus, the majority of XX- XY chimeras develop into males and seldom into her-
maphrodites or females [34, 35]. In the B6.YTR strain, thetestis-determining function is partially impaired, resultingin development of adult males, hermaphrodites, and sex-reversed females at a 1:1:2 ratio. Nonetheless, if XX cellsplay only a passive role in gonadal sex determination, thesex ratio in XX BALB/c B6.YTIR chimera would havebeen close to that in the nonchimeric B6.YTIR progeny. In-stead, the presence of XX cells appears to have further re-duced the testis-determining activity of B6.YTrR cells, sinceall but one chimera developed into females and none intohermaphrodites. Overall production of the putative testis-determining gene product may be critical for directing thesexual differentiation in the gonad.
Live-Borns from XX OocytesAfter mating with normal males, all XX - XY chimeric
females produced progeny exclusively derived from XX
579

580
FIG. 5. Distribution of XY cells detectedby in situ hybridization with Dig-labeled145SC5 in chimeric ovaries. a) XX BALB/c<e XY B6.YT'R chimeric ovary (#2) with ahigh contribution of XY cells. Hybridiza-tion signals are seen in all ovarian celltypes: the surface epithelium, interstitium,and follicular components at differentstages (arrowheads). b) XX BALB/c - XYB6.Y T 'R chimeric ovary (#2) at a highermagnification. The surface epitheliumshows a higher frequency of labeled cellsthan the interstitium. A hybridization sig-nal (arrowhead) is seen in an oocyte of apreantral follicle composed of labeled andunlabeled populations of granulosa andtheca cells. Part of corpus luteum is seenwith a high frequency of labeled cells. c)XX BALB/c <-- XY B6.YT'R chimeric ovary(#31) with a medium level of XY contribu-tion. Part of an oocyte but not its nucleusis seen in a preantral follicle. A higherproportion of positive cells are seen intheca layers and the upper area of thegranulosa population than in the lowerarea or the interstitium. d) XX BALB/c -
XX B6* chimeric ovary (#13). No hybrid-ization signals are seen in any of the ovar-ian cell types. The contrast was reduced toshow ovarian structures compared with a-c. Scale bars: a, 300 Im; b-d, 100 ,um.SE, surface epithelium; In, interstitium; Th,theca layers; Gr, granulosa cells; Oo, oo-cyte; CL, corpus luteum.
AMLEH AND TAKETO
TABLE 5. Fertility of XY -> XY chimeric males.
Composition Live-bornAget
I.D. # BALB/c Coat* (B6 x B6.Y T 'R)F1 (dpp) Total no./litter Albino Agouti
5 XY <* XY 52 6 4 254 9 1 869 7 6 169 9 1 871 7 6 1
38 18 20
6 XY < XY 51 3 0 366 8 8 073 9 9 085 6 5 185 9 6 3
35 28 7
t Age at which copulation was noted.* Indicates the relative proportion of coat colors.

FERTILITY OF XY OOCYTES IN THE CHIMERIC MOUSE
oocytes irrespective of the degree of XY contribution inbody formation. Therefore, the fertility of XX oocytes wasnot abolished by the presence of XY somatic cells. We havepreviously reported several abnormal aspects of theB6.YTIR ovary during development, for example, transcrip-tion of the Sry gene in the ovarian primordium [19], post-natal development of abnormal endocrine features [30], andabsence of regular estrous cyclicity at puberty [27]. Sinceall XX <-> XY chimeric females turned out to be fertile, theabnormalities that we have observed in the somatic com-ponents of the B6.YTR female appear to be either not crit-ical for fertility, caused only locally by the XY oocyte, orovercome by the presence of a small number of XX cells.
Our conclusion would be invalid if XY cells were some-how excluded from the formation of ovaries despite thepresence of XY cells in other parts of the body. However,we confirmed the presence of XY somatic cells in chimericovaries by in situ hybridization using a Y-specific DNAprobe. Furthermore, the overall proportion of XY cells inthe chimeric ovary corresponded to the proportion of XYcells determined by the DNA contents in the tail and kid-ney. Therefore, the XY contributions in the kidney, whichshares the embryonic origin with the gonad, should indicatethe varying degrees of chimerism in the ovaries examinedin the present study. We could not determine the exact pro-portion of XY cells in the chimeric ovary that was sub-jected to morphological studies. However, even if it couldbe determined, the percentage of XY cells in the chimericovary after mating and pregnancy may be found to differfrom the proportion at the time of ovulation and, therefore,may not provide any better information than that obtainedfrom the kidney. It is currently impossible to correlate thefate of each oocyte and the composition of the follicle thatencompassed the oocyte.
Absence of Live-Borns from XY Oocytes
Despite the fertility of all chimeric females, no live-borns were produced from XY oocytes. This observationsuggests that the presence of XX cells failed to make theXY oocyte competent for reproduction. Therefore, the de-velopmental failure of the XY oocyte is most likely intrin-sic to the oocyte itself, and not to the surrounding somaticcells in the B6.YTR ovary. Possible competition betweenthe germ cells of two mouse strains in the chimera has beenreported previously [32, 36]. Accordingly, it is conceivablethat XY oocytes were excluded from the chimeric ovariesin competition with XX oocytes. In the present study, how-ever, we confirmed the presence of XY oocytes at differentstages of follicular development in XX BALB/c <-> XYB6.YTIR chimeric ovaries. It remains possible that XX oo-cytes were favorably recruited for ovulation and, therefore,the absence of progeny derived from XY oocytes may beattributable to the paucity of XY-derived eggs ovulatedrather than death of XY-derived embryos. Nonetheless, itmust be emphasized that the XY genotype of the oocyte,and not of somatic cells, appears to be responsible for ex-clusion of XY oocytes from the reproductive process,whether at ovulation or after fertilization.
Unlike XX BALB/c <-4 XY B6.YTIR chimeric ovaries,both germ cell populations in the XY BALB/c <--> XYB6.YTIR chimeric testis contributed to progeny production.Therefore, the absence of progeny derived from the XYoocyte is associated with the infertility of the B6.YTIR fe-male, rather than the results of general competition betweenB6.YTIR and BALB/c germ cells. The BALB/c strain ap-
peared to be favored over the B6.YTIR strain for young-born production if their proportions in the testis were par-allel to those in the coat color. However, it is conceivablethat extensive proliferation of spermatogonia may have dis-torted the proportion. A larger number of chimeric malesis needed to determine whether the BALB/c genotype al-ways has an advantage over the B6.y TIR genotype in sper-matogenesis.
The defect in the XY oocyte remains to be clarified.Hunt and LeMaire [45] have reported a high frequency ofprecocious dissociation between the X and Y chromosomesin oocytes isolated from the B6.YPO female mouse at thediakinesis/metaphase I stage. Accordingly, it is conceivablethat the B6.YTIR female may lose its embryos because ofaneuploidy. However, the XO female mouse is invariablyfertile although it lacks the pairing partner for the single Xchromosome and suffers from a high incidence of aneu-ploidy and embryo death [11, 46]. Furthermore, somestrains of XY sex-reversed female mice demonstrate highfrequencies of aneuploidy and yet produce litters, albeit insmall numbers [12-15]. Therefore, the rate of aneuploidymust be very high to be the sole cause of the completeabsence of progeny in the B6.YTIR female. Actual aneu-ploidy in matured or fertilized eggs from the B6.YTR fe-male remains to be examined.
Reproductive Life Length of XX BALB/c <--> XY B6. yrTR
Chimeric Females
We have previously reported that the ovaries of theB6.YTIR female consistently lose their oocytes by 2 mo ofage [27, 30]. In contrast, the XX BALB/c <- XY B6.YTRchimeric females that were dominated by XY cells con-ceived up to over 4 mo of age in the present study. There-fore, it appears that introduction of normal XX cells pro-longed the reproductive life of the B6.YTIR ovary to someextent. Nonetheless, the chimeric ovary had been deprivedof all but a few oocytes by 5 mo of age, much earlier thanthe normal XX ovary. It remains possible that XY somaticcells exert long-term effects on reproductive functions. Al-ternatively, the number of XX oocytes may limit the repro-duction life length.
In conclusion, the XY chromosomal composition of theoocyte appears to be responsible for the infertility of theB6.YTIR female mouse. On the other hand, the presence ofXY somatic cells allows ovarian differentiation to supportthe XX oocyte for producing live-borns.
ACKNOWLEDGMENTS
We are grateful to Dr. H. Clarke of McGill University and Dr. G.MacGregor of Emory University for their advice on chimera formation,and also to Dr. A. Peterson and I. Tretjakoff of McGill University for theirhelp on carrying out embryo transfers. We thank Dr. Y. Nishioka of McGillUniversity for the gift of DNA probe 145SC5.
REFERENCES
1. Berta P, Hawkins JR, Sinclair AH, Taylor A, Griffiths BL, GoodfellowPN, Fellous M. Genetic evidence equating SRY and the testis-deter-mining factor. Nature 1990; 348:448-450.
2. Gubbay J, Collignon J, Koopman P, Capel B, Economou A, Miin-sterberg A, Vivian N, Goodfellow P, Lovell-Badge R. A gene mappingto the sex-determining region of the mouse Y chromosome is a mem-ber of a novel family of embryonically expressed genes. Nature 1990;346:245-250.
3. Koopman P, Gubbary J, Vivian N, Goodfellow P, Lovell-Badge R.Male development of chromosomally female mice transgenic for Sry.Nature 1991; 351:117-121.
581

AMLEH AND TAKETO
4. Sinclair AH, Berta P, Palmer M, Hawkins JR, Griffiths BL, Smith MJ,Foster JW, Frischauf A-M, Lovell-Badge R, Goodfellow PN. A genefrom the human sex-determining region encodes a protein with ho-mology to a conserved DNA-binding motif. Nature 1990; 346:240-244.
5. Bardoni B, Zuffardi O, Guioli S, Ballabio A, Simi P, Cavalli P, Gri-mold MG, Fraccaro M, Cemerino G. A deletion map of the humanYql region: implications for the evolution of the Y chromosome andtentative mapping of a locus involved in spermatogenesis. Genomics1991; 11:443-451.
6. Burgoyne PS, Mahadevaiah SK, Sutcliffe MJ, Palmer SJ. Fertility inmice requires X-Y pairing and a Y-chromosomal "spermiogenesis"gene mapping to the long arm. Cell 1992; 71:391-398.
7. Kay GE Ashworth A, Penny GD, Dunlop M, Swift S, Brockdorff N,Rastan S. A candidate spermatogenesis gene on the mouse Y chro-mosome is homologous to ubiquitin-activating enzyme El. Nature1991; 354:486-489.
8. Mitchell MJ, Woods DR, Tucker PK, Opp JS, Bishop C. Homologyof a candidate spermatogenic gene from the mouse Y chromosome tothe ubiquitin-activating enzyme El. Nature 1991; 354:483-486.
9. Burgoyne PS, Baker TG. Oocytes depletion in XO mice and their XXsibs from 12 to 200 days post partum. J Reprod Fertil 1981; 61:207-212.
10. Burgoyne PS, Baker TG. Perinatal oocyte loss in XO mice and itsimplications for the aetiology of gonadal dysgenesis in XO women. JReprod Fertil 1985; 75:633-645.
11. Lyon ME Hawker SG. Reproductive lifespan in irradiated and unir-radiated chromosomally XO mice. Genet Res Camb 1973; 21:185-194.
12. Capel B, Rasberry C, Dyson J, Bishop CE, Simpson E, Vivian N,Lovell-Badge R, Rastan S, Cattanach BM. Deletion of Y chromosomesequences located outside the testis determining region can cause XYfemale sex reversal. Nat Genet 1993; 5:301-307.
13. Laval SH, Glenister PH, Rasberry C, Thornton CE, Mahadevaiah SK,Cooke HJ, Burgoyne P, Cattanach BM. Y chromosome short arm-Sxrrecombination in XSxr/Y males causes deletion of Rbm and XY fe-male sex reversal. Proc Natl Acad Sci USA 1995; 92:10403-100407.
14. Lovell-Badge R, Robertson E. XY female mice resulting from a her-itable mutation in the primary testis-determining gene, Tdy. Devel-opment 1990; 109:635-646.
15. Mahadevaiah SK, Lovell-Badge R, Burgoyne PS. Tdy-negative XY,XXY and XYY female mice: breeding data and synaptonemal com-plex analysis. J Reprod Fertil 1993; 97:151-160.
16. Nagamine CM, Taketo T, Koo GC. Studies on the genetics of tda-1XY sex reversal in the mouse. Differentiation 1987; 33:223-231.
17. Eicher EM, Washburn LL. Inherited sex reversal in mice: identifica-tion of a new primary sex-determining gene. J Exp Zool 1983; 228:297-304.
18. Eicher EM, Washburn LL, Whitney III JB, Morrow KE. Mus pos-chiavinus Y chromosome in the C57BL/6J murine genome causes sexreversal. Science 1982; 217:535-537.
19. Lee C-H, Taketo T. Normal onset, but prolonged expression, of Srygene in the B6.YD ° M sex-reversed mouse gonad. Dev Biol 1994; 165:442-452.
20. Eicher EM, Washburn LL. Genetic control of primary sex determi-nation in mice. Annu Rev Genet 1986; 20:327-360.
21. Biddle FG, Eisner JR, Eales BA. The testis-determining autosomaltrait, Tda-1, of C57BL/6J is determined by more than a single auto-somal gene when compared with DBA/2J mice. Genome 1994; 37:296-304.
22. Eales BA, Nahas M, Biddle FG. Directional dominance and a devel-opmental model for the expression of the Tda testis-determining au-tosomal trait of the mouse. Genome 1996; 39:520-527.
23. Eisner JR, Eales BA, Biddle FG. Segregation analysis of the testis-determining autosomal trait, Tda, that differs between the C57BI/6Jand DBA/2J mouse stains suggests a multigenic threshold model. Ge-nome 1996; 39:322-335.
24. Eicher E, Washburn LL, Schork NJ, Lee BK, Shown EP, Xu X,Dredge RD, Pringle MJ, Page DC. Sex-determining genes on mouseautosomes identified by linkage analysis of C57BL/6J-YPOS sex re-versal. Nat Genet 1996; 14:206-209.
25. Coward P, Nagai K, Chen D, Thomas HD, Nagamine CM, Lau YC.Polymorphism of a CAG trinucleotide repeat within Sry correlateswith B6.YDOM sex reversal. Nat Genet 1994; 6:245-250.
26. Carlisle C, Winking H, Weichenhan D, Nagamine CM. Absence ofcorrelation between Sry polymorphisms and XY sex reversal causedby the M. m. domesticus Y chromosome. Genomics 1996; 33:32-45.
27. Taketo-Hosotani T, Nishioka Y, Nagamine C, Villalpando I, Merchant-Larios H. Development and fertility of ovaries in the B6.Y D°M sex-reversed female mouse. Development 1989; 197:95-105.
28. Merchant-Larios H, Clarke HJ, Taketo T Developmental arrest of fer-tilized eggs from the B6.YD°OM sex-reversed female mouse. Dev Genet1994; 15:435-442.
29. Amleh A, Ledee N, Saeed J, Taketo T Competence of oocytes fromthe B6.Y D° M sex-reversed female mouse for maturation, fertilization,and embryonic development in vitro. Dev Biol 1996; 178:263-275.
30. Villalpando I, Nishioka Y, Taketo T Endocrine differentiation of theXY sex-reversed mouse ovary during postnatal development. J SteroidBiochem Mol Biol 1993; 45:265-273.
31. Gearhart J, Oster-Granite ML. Reproduction in a population of chi-meric mice: Relationship of chromosomal sex to functional germ cellsand proportions of chimeric components in several tissues. Biol Re-prod 1981; 24:713-722.
32. McLaren A. Sex chimerism and germ cell distribution in a series ofchimeric mice. J Embryol Exp Morphol 1975; 33:205-216.
33. Mullen RJ, Whitten WK. Relationship of genotype and degree of chi-merism in coat color to sex ratios and gametogenesis in chimeric mice.J Exp Zool 1971; 178:165-176.
34. Mystkowska ET, Tarkowski AK. Observations on CBA-p/CBA-T6T6mouse chimeras. J Embryol Exp Morphol 1968; 20:33-52.
35. Tarkowski AK. Mouse chimeras developed from fused eggs. Nature1961; 190:857-860.
36. Burgoyne PS, Buehr M, McLaren A. XY follicle cells in ovaries ofXX-XY female mouse chimeras. Development 1988; 104:683-688.
37. Evans EP, Ford CE, Lyon ME. Direct evidence of the capacity of theXY germ cell in the mouse to become an oocyte. Nature 1977; 267:430-431.
38. Ford CE, Evans EP, Burtenshaw MD, Clegg HM, Tuffrey M, BarnesRD. A functional "sex-reversed" oocyte in the mouse. Proc R SocLond 1975; B190:187-197.
39. Patek CE, Kerr JB, Gosden RG, Jones KW, Hardy K, Muggleton-Harris AL, Handyside AH, Whittingham DG, Hooper ML. Sex chi-merism, fertility and sex determination in the mouse. Development1991; 113:311-325.
40. Wood SA, Allen ND, Rossant J, Auerbach A, Nagy A. Non-injectionmethods for the production of embryonic stem cell-embryo chimaeras.Nature 1993; 365:87-89.
41. Boland NI, Gosden RG. Clonal analysis of chimeric mouse ovariesusing DNA in situ hybridization. J Reprod Fertil 1994; 100:203-210.
42. Nagamine CM, Chan K, Kozak CA, Lau Y-E Chromosome mappingand expression of a putative testis-determining gene in mouse. Science1989; 243:80-83.
43. Musiani M, Gentilomi G, Zerbini M, Gibellini D, Gallinella G, PileriS, Baglioni P, La Placa M. In situ detection of cytomegalovirus DNAin biopsies of AIDS patients using a hybrido-immunocytochemicalassay. Histochemistry 1990; 94:21-25.
44. Singh L, Matsukuma S, Jones KW. Testis development in a mousewith 10% of XY cells. Dev Biol 1987; 122:287-290.
45. Hunt PA, LeMaire R. Sex-chromosome pairing: evidence that the be-havior of the pseudoautosomal region differs during male and femalemeiosis. Am J Hum Genet 1991; 50:1162-1170.
46. Burgoyne PS, Biggers JD. The consequences of X-dosage deficiencyin the germ line: impaired development in vitro of preimplantationembryos from XO mice. Dev Biol 1976; 51:109-117.
582