
LHCII organization and thylakoid lipids affect the sensitivity of the photosynthetic apparatus to...
7
Research article LHCII organization and thylakoid lipids affect the sensitivity of the photosynthetic apparatus to high-light treatment Kolyo G. Dankov a , Anelia G. Dobrikova a , Bettina Ughy b , Balázs Bogos b , Zoltan Gombos b , Emilia L. Apostolova a, * a Institute of Biophysics and Biomedical Engineering, Bulgarian Academy of Sciences, Sofia 1113, Bulgaria b Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H-6701 Szeged, Hungary article info Article history: Received 16 June 2010 Accepted 18 February 2011 Available online 26 February 2011 Keywords: Thylakoid membrane High-light treatment Lipid composition Photosynthetic apparatus Light-harvesting complex of photosystem II PAM fluorometer Photosynthetic oxygen evolution abstract Pulse-amplitude-modulated (PAM) chlorophyll fluorescence and photosynthetic oxygen evolution were used to investigate the role of the different amount and organization of light-harvesting complexes of photosystem II (LHCII) in four pea species on the susceptibility of the photosynthetic apparatus to high- light treatment. In this work we analyzed the thylakoid membrane lipid composition of the studied pea plants. A relationship between the structural organization of LHCII proteins, the amount of the main lipid classes and the sensitivity of the photosynthetic apparatus to high-light treatment was found. The results reveal that the photosynthetic apparatus, enriched in oligomeric forms of LHCII concomitant with decreased amount of anionic lipids and increased content of the monogalactosyldiacylglycerol (MGDG), is less sensitive to high light. Our data also suggest that the degree of LHCII oligomerization, as well as the lipid composition do not influence the degree of recovery of the PSII photochemistry after excess light exposure. Ó 2011 Elsevier Masson SAS. All rights reserved. 1. Introduction Photosynthesis is a light-driven reaction, but exposure of plants to strong light intensities causes a decrease of photosynthetic efficiency, a phenomenon referred as photoinhibition [1,2]. The susceptibility of plants to photoinhibition depends on plant species and growth light condition. It is well established that acclimation, which is a basic feature of the photosynthetic apparatus of higher plants to adapt to the external environmental conditions, is accompanied with distinct changes in gene expression followed by changes of the organization of the protein complexes, lipids and the function of the thylakoid membranes [3]. The primary site of photodamage is the multiprotein complex of photosystem II (PSII) [1,2]. It is supposed that the extent of the high- light inhibition depends on the balance between the rate of damage and repair of PSII [2,4]. Strong visible light has two effects: a direct effect on photodamage, and an inhibitory effect on repair of PSII via the production of reactive oxygen species [2]. Vass and Cser [5] proposed a model of photoinhibition by visible light in which charge recombination processes are involved not only in damaging, but also in protecting PSII via the modulation of singlet oxygen formation [5]. It is considered that the earlier detectable step of the light-induced photoinhibition is the release of the Mn ion to the thylakoid lumen, which is followed by oxidative damage to the PSII reaction center [6]. Recent investigations developed a new scheme for the molecular mechanisms of photoinhibition, which propose two steps of the photodamage. The first step is the light-dependent destruction in the Mn cluster of the oxygen-evolving complex (OEC), and the second step is the inactivation of the reaction center of PSII [2,7]. It has been shown that phosphorylation of the PSII core protein facilitates the repair of the photodamaged PSII at strong light [8]. Plants have developed several adaptive regulatory mechanisms to optimize the utilization of light energy and to protect themselves against over-excitation-related damage. The dynamic properties of the proteins and pigments of the chlorophyll a/b light-harvesting complex of photosystem II (LHCII) allow excess energy to be dissipated as heat [9]. During the long-term acclimation of plants, the LHCII content in the photosynthetic membranes often changes, which affects the ultrastructure of the grana [10]. The role of light-harvesting antenna size of PSII in susceptibility of the photosynthetic apparatus towards high-light treatment is still * Corresponding author at: Institute of Biophysics and Biomedical Engineering, Bulgarian Academy of Sciences, Acad. G. Bonchev Str., Bl. 21, 1113 Sofia, Bulgaria. Tel.: þ359 2 979 26 21; fax: þ359 2 971 24 93. E-mail address: [email protected] (E.L. Apostolova). Contents lists available at ScienceDirect Plant Physiology and Biochemistry journal homepage: www.elsevier.com/locate/plaphy 0981-9428/$ e see front matter Ó 2011 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.plaphy.2011.02.019 Plant Physiology and Biochemistry 49 (2011) 629e635
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of LHCII organization and thylakoid lipids affect the sensitivity of the photosynthetic apparatus to...

lable at ScienceDirect
Plant Physiology and Biochemistry 49 (2011) 629e635
Contents lists avai
Plant Physiology and Biochemistry
journal homepage: www.elsevier .com/locate/plaphy
Research article
LHCII organization and thylakoid lipids affect the sensitivityof the photosynthetic apparatus to high-light treatment
Kolyo G. Dankov a, Anelia G. Dobrikova a, Bettina Ughy b, Balázs Bogos b,Zoltan Gombos b, Emilia L. Apostolova a,*
a Institute of Biophysics and Biomedical Engineering, Bulgarian Academy of Sciences, Sofia 1113, Bulgariab Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H-6701 Szeged, Hungary
a r t i c l e i n f o
Article history:Received 16 June 2010Accepted 18 February 2011Available online 26 February 2011
Keywords:Thylakoid membraneHigh-light treatmentLipid compositionPhotosynthetic apparatusLight-harvesting complexof photosystem IIPAM fluorometerPhotosynthetic oxygen evolution
* Corresponding author at: Institute of BiophysicsBulgarian Academy of Sciences, Acad. G. Bonchev StrTel.: þ359 2 979 26 21; fax: þ359 2 971 24 93.
E-mail address: [email protected] (E.L. Apo
0981-9428/$ e see front matter � 2011 Elsevier Masdoi:10.1016/j.plaphy.2011.02.019
a b s t r a c t
Pulse-amplitude-modulated (PAM) chlorophyll fluorescence and photosynthetic oxygen evolution wereused to investigate the role of the different amount and organization of light-harvesting complexes ofphotosystem II (LHCII) in four pea species on the susceptibility of the photosynthetic apparatus to high-light treatment. In this work we analyzed the thylakoid membrane lipid composition of the studied peaplants. A relationship between the structural organization of LHCII proteins, the amount of the main lipidclasses and the sensitivity of the photosynthetic apparatus to high-light treatment was found. The resultsreveal that the photosynthetic apparatus, enriched in oligomeric forms of LHCII concomitant withdecreased amount of anionic lipids and increased content of the monogalactosyldiacylglycerol (MGDG),is less sensitive to high light. Our data also suggest that the degree of LHCII oligomerization, as well as thelipid composition do not influence the degree of recovery of the PSII photochemistry after excess lightexposure.
� 2011 Elsevier Masson SAS. All rights reserved.
1. Introduction
Photosynthesis is a light-driven reaction, but exposure of plantsto strong light intensities causes a decrease of photosyntheticefficiency, a phenomenon referred as photoinhibition [1,2]. Thesusceptibility of plants to photoinhibition depends on plant speciesand growth light condition. It is well established that acclimation,which is a basic feature of the photosynthetic apparatus of higherplants to adapt to the external environmental conditions, isaccompanied with distinct changes in gene expression followed bychanges of the organization of the protein complexes, lipids and thefunction of the thylakoid membranes [3].
The primary site of photodamage is the multiprotein complex ofphotosystem II (PSII) [1,2]. It is supposed that the extent of the high-light inhibition depends on the balance between the rate of damageand repair of PSII [2,4]. Strong visible light has two effects: a directeffect on photodamage, and an inhibitory effect on repair of PSII viathe production of reactive oxygen species [2]. Vass and Cser [5]
and Biomedical Engineering,., Bl. 21, 1113 Sofia, Bulgaria.
stolova).
son SAS. All rights reserved.
proposed a model of photoinhibition by visible light in whichcharge recombination processes are involved not only in damaging,but also in protecting PSII via the modulation of singlet oxygenformation [5]. It is considered that the earlier detectable step of thelight-induced photoinhibition is the release of the Mn ion to thethylakoid lumen, which is followed by oxidative damage to the PSIIreaction center [6]. Recent investigations developed a new schemefor the molecular mechanisms of photoinhibition, which proposetwo steps of the photodamage. The first step is the light-dependentdestruction in the Mn cluster of the oxygen-evolving complex(OEC), and the second step is the inactivation of the reaction centerof PSII [2,7]. It has been shown that phosphorylation of the PSII coreprotein facilitates the repair of the photodamaged PSII at stronglight [8].
Plants have developed several adaptive regulatory mechanismsto optimize the utilization of light energy and to protect themselvesagainst over-excitation-related damage. The dynamic properties ofthe proteins and pigments of the chlorophyll a/b light-harvestingcomplex of photosystem II (LHCII) allow excess energy to bedissipated as heat [9]. During the long-term acclimation of plants,the LHCII content in the photosynthetic membranes often changes,which affects the ultrastructure of the grana [10].
The role of light-harvesting antenna size of PSII in susceptibilityof the photosynthetic apparatus towards high-light treatment is still

K.G. Dankov et al. / Plant Physiology and Biochemistry 49 (2011) 629e635630
controversial. Some investigators suggested that photoinhibitiondepends on the size of PSII antenna [11 and refs. therein], whileothers [12] claimed that it is independent of LHCII size. In our recentstudy we have shown that the effect of UV-A radiation on thephotosynthetic apparatus depends not only on the antenna size ofPSII, but also on the degree of LHCII oligomerization [13].
On the other hand, it is well known that the specific interactionsbetween lipids and membrane proteins are involved in the activa-tion of protein function and protein packing of themacrocomplexesin the thylakoid membranes [14e16]. Monogalactosyldiacylglycerol(MGDG) is the major lipid constituent of thylakoid membranes andhas been proposed to be included in various important photosyn-thetic processes [17,18]. Several lines of evidence show that thyla-koid lipid desaturation and composition influence photoinhibition[19e24]. Earlier observations demonstrated that polyunsaturatedfatty acids of membrane lipids are important in the resistance of thephotosynthetic apparatus to low-temperature photoinhibition [15].The role of the digalactosyldiacylglycerol (DGDG) [25], as well as ofthe unsaturated fatty acids of phosphatidylglycerol (PG) [23,26], inrecovery of the PSII protein complex was revealed. In spite ofnumerous studies, the overall role of lipids in the protectionmechanisms of the photosynthetic apparatus against photo-inhibition has not been completely elucidated.
In this study, we used previously described pea plants, whichhave different ratio of oligomeric to monomeric forms of LHCII(LHCIIo/LHCIIm). The ratio increases in the following order: mutantChlorotica XV/1422 (2.45), Auralia wild type (2.85), mutant Costata2/133 (3.34) and Borec wild type (4.87) [27e29]. Earlier studies alsorevealed that the amount of LHCII proteins in Auralia wt and itsmutant Chlorotica XV/1422 is smaller than in Borec wt and themutant Costata 2/133 [28,29]. These differences were accompaniedwith alterations in the pigment composition, as well as in thephotochemical and physicochemical characteristics of thylakoidmembranes [27e29].
In order to get better insight into the role of the degree of LHCIIoligomerization on sensitivity of photosynthetic apparatus to high-light treatment, we studied the effect of excess light on theparameters of the PAM chlorophyll fluorescence and the photo-synthetic oxygen evolution of studied pea plants. In present studywe show that the sensitivity of the photosynthetic apparatus tohigh-light treatment depends on the structural organization ofLHCII and the lipid composition.
2. Materials and methods
2.1. Plant material and thylakoid membrane isolation
The plants from Pisum sativum L. cv. Borec and Auralia and theirmutants (Costata 2/133 and Chlorotica XV/1422) were grown undercontrolled conditions with 16 h light/8 h dark period. The mutantsthat we used in this study are well-defined and stable. In each newseries of studies of these mutants we checked their properties andthey did not reveal any differences in their physicochemical char-acteristics and functions.
The thylakoid membranes were isolated from non-treated andhigh-light irradiated leaves as described in [30] and suspended ina medium containing: 40 mM HEPES (pH 7.6), 10 mM NaCl, 5 mMMgCl2 and 400 mM sucrose.
2.2. High-light treatment
Leaves were cut from plants and the petioles were immersed inwater or in solution of 10 mM lincomycin (Sigma) and incubated indim light for 2 h before high-light treatment. The leaves wereilluminated up to 3 h at room temperature (20e25 �C) with white
high-intensity light (1300 mmolm�2 s�1). For recovery from pho-toinhibitory treatment the leaves were transferred to a dim light atroom temperature.
2.3. Lipid analysis
The analysis of lipids of thylakoid membranes from pea typeswas carried out according to the method of Sato and Murata [31].The total lipid and the various lipid classes, which were fraction-ated on precoated thin-layer chromatography (TLC) plates (Merck5721) developed with CHC13/CH3OH/28% NH4OH (65/35/5, v/v/v),were subjected to methanolysis with 5% HCl in methanol at 85 �Cfor 2 h. The resulting methyl esters were analyzed with a HewlettPackard (Palo Alto, California, USA) HP6890 gas chromatographequipped with a hydrogen flame-ionization detector. Fatty acidmethyl esters were separated on a 30 m� 0.25 mm i.d. SP-2330capillary column (Supelco, Bellefonte, Pennsylvania, USA).Temperatures of the column and the flame-ionization detectorwere 180 and 260 �C, respectively. The relative amounts of fattyacid methyl esters were determined by comparison of areas underthe peaks on the chromatogram to the area of internal fatty acidstandard, pentadecanoic acid (15:0). Fifty mg of pentadecanoic acidwas added to each band of the TLC plate.
2.4. Pulse-amplitude-modulated chlorophyll fluorescence
Modulated chlorophyll fluorescence was measured on leaf discsby a PAM fluorometer (H. Walz, Effeltrich, Germany, model PAM101-103). The leaves were dark adapted for 15 min. The initialfluorescence level (F0) was measured at instrument frequency of1.6 kHz and measuring beam set at 0.120 mmolm�2 s�1 PFD. Forevaluation of maximal fluorescence level (Fm), saturating flashes of3000 mmolm�2 s�1 PFD with duration of 0.8 s were provided bySchott lamp KL 1500 (Schott Glaswerke, Mainz, Germany). Thesaturating flash gives Fm in dark-adapted state and Fm0 in light-adapted state. The interval between two consecutive flashes was60 s. The actinic light illumination (250 mmolm�2 s�1 PFD) wasprovided by second Schott lamp KL 1500 for the induction ofphotosynthesis. The maximum quantum yield of primary photo-chemistry in dark-adapted state was calculated, (Fm� F0)/Fm¼ Fv/Fm [32], which is proportional to de-excitation rate constant ofphotochemical (kp) and non-photochemical processes (kn) of PSIIdeactivation, Fv/Fm¼ kp/(knþ kP) [33]. The non-photochemicalquenching, NPQ¼ (Fm� Fm0)/Fm0, was calculated as in [34].
2.5. Oxygen evolution measurements
Oxygen flash yields and initial oxygen burst of isolated thylakoidmembranes were measured by a home-built polarographic oxygenrate electrode described in [35]. Thylakoid membranes were sus-pended in a medium containing: 40 mM HEPES (pH 7.6), 10 mMNaCl, 5 mM MgCl2 and 400 mM sucrose. The chlorophyll concen-tration was 150 mgml�1. The measurements were performed as in[29]. The induction curves after oxygen burst exhibit biphasicexponential decay. The deconvolution of the oxygen burst decaywas performed as in [13]. Initial S0 and S1 state distribution, missesand double hits were determined by the fitting of the theoreticallycalculated yields according to the model of Kok et al. [36] with theexperimentally obtained oxygen flash yields using the least squaredeviations procedure.
2.6. Statistical analysis
The results aremeanvalues from3 to 5 independent experiments.The statistical differences among the means were determined using
K.G. Dankov et al. / Plant Physiology and Biochemistry 49 (2011) 629e635 631
a two-tailed paired Student’s t-test. Values of P< 0.05 were consid-ered as significant differences between studied plants.
3. Results
3.1. The effect of high-light treatment on the parameters of theroom temperature chlorophyll fluorescence
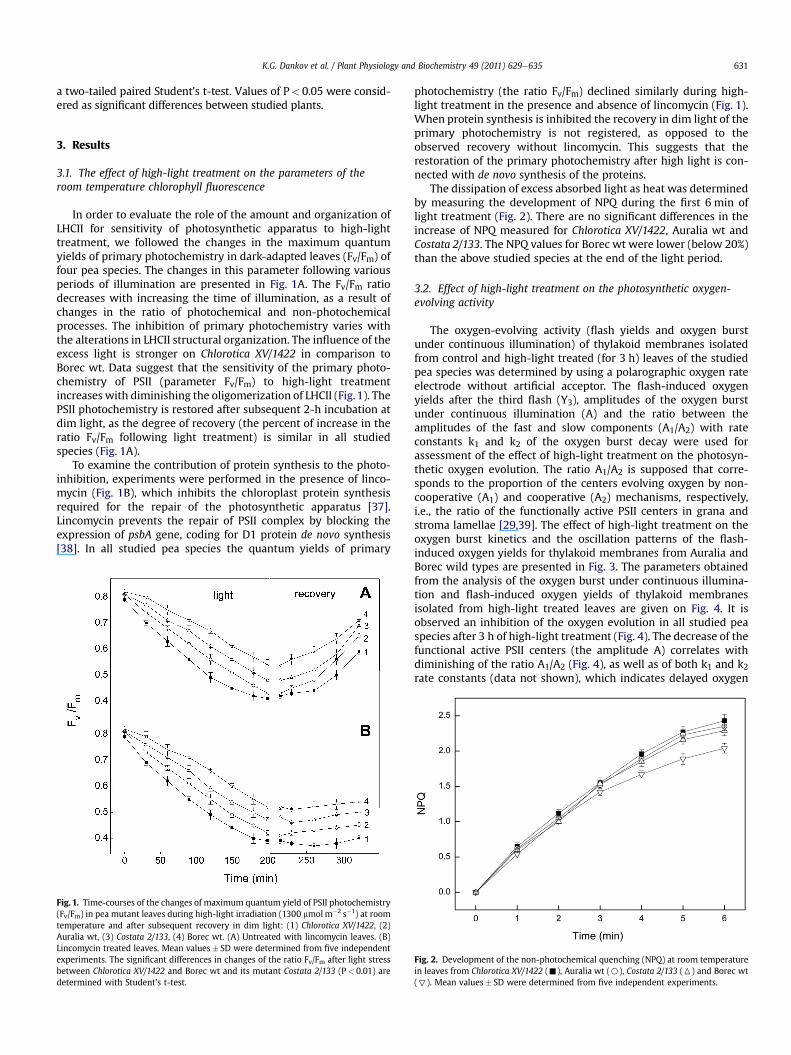
In order to evaluate the role of the amount and organization ofLHCII for sensitivity of photosynthetic apparatus to high-lighttreatment, we followed the changes in the maximum quantumyields of primary photochemistry in dark-adapted leaves (Fv/Fm) offour pea species. The changes in this parameter following variousperiods of illumination are presented in Fig. 1A. The Fv/Fm ratiodecreases with increasing the time of illumination, as a result ofchanges in the ratio of photochemical and non-photochemicalprocesses. The inhibition of primary photochemistry varies withthe alterations in LHCII structural organization. The influence of theexcess light is stronger on Chlorotica XV/1422 in comparison toBorec wt. Data suggest that the sensitivity of the primary photo-chemistry of PSII (parameter Fv/Fm) to high-light treatmentincreases with diminishing the oligomerization of LHCII (Fig.1). ThePSII photochemistry is restored after subsequent 2-h incubation atdim light, as the degree of recovery (the percent of increase in theratio Fv/Fm following light treatment) is similar in all studiedspecies (Fig. 1A).
To examine the contribution of protein synthesis to the photo-inhibition, experiments were performed in the presence of linco-mycin (Fig. 1B), which inhibits the chloroplast protein synthesisrequired for the repair of the photosynthetic apparatus [37].Lincomycin prevents the repair of PSII complex by blocking theexpression of psbA gene, coding for D1 protein de novo synthesis[38]. In all studied pea species the quantum yields of primary
Fig. 1. Time-courses of the changes of maximum quantum yield of PSII photochemistry(Fv/Fm) in pea mutant leaves during high-light irradiation (1300 mmolm�2 s�1) at roomtemperature and after subsequent recovery in dim light: (1) Chlorotica XV/1422, (2)Auralia wt, (3) Costata 2/133, (4) Borec wt. (A) Untreated with lincomycin leaves. (B)Lincomycin treated leaves. Mean values� SD were determined from five independentexperiments. The significant differences in changes of the ratio Fv/Fm after light stressbetween Chlorotica XV/1422 and Borec wt and its mutant Costata 2/133 (P< 0.01) aredetermined with Student’s t-test.
photochemistry (the ratio Fv/Fm) declined similarly during high-light treatment in the presence and absence of lincomycin (Fig. 1).When protein synthesis is inhibited the recovery in dim light of theprimary photochemistry is not registered, as opposed to theobserved recovery without lincomycin. This suggests that therestoration of the primary photochemistry after high light is con-nected with de novo synthesis of the proteins.
The dissipation of excess absorbed light as heat was determinedby measuring the development of NPQ during the first 6 min oflight treatment (Fig. 2). There are no significant differences in theincrease of NPQ measured for Chlorotica XV/1422, Auralia wt andCostata 2/133. The NPQ values for Borec wt were lower (below 20%)than the above studied species at the end of the light period.
3.2. Effect of high-light treatment on the photosynthetic oxygen-evolving activity
The oxygen-evolving activity (flash yields and oxygen burstunder continuous illumination) of thylakoid membranes isolatedfrom control and high-light treated (for 3 h) leaves of the studiedpea species was determined by using a polarographic oxygen rateelectrode without artificial acceptor. The flash-induced oxygenyields after the third flash (Y3), amplitudes of the oxygen burstunder continuous illumination (A) and the ratio between theamplitudes of the fast and slow components (A1/A2) with rateconstants k1 and k2 of the oxygen burst decay were used forassessment of the effect of high-light treatment on the photosyn-thetic oxygen evolution. The ratio A1/A2 is supposed that corre-sponds to the proportion of the centers evolving oxygen by non-cooperative (A1) and cooperative (A2) mechanisms, respectively,i.e., the ratio of the functionally active PSII centers in grana andstroma lamellae [29,39]. The effect of high-light treatment on theoxygen burst kinetics and the oscillation patterns of the flash-induced oxygen yields for thylakoid membranes from Auralia andBorec wild types are presented in Fig. 3. The parameters obtainedfrom the analysis of the oxygen burst under continuous illumina-tion and flash-induced oxygen yields of thylakoid membranesisolated from high-light treated leaves are given on Fig. 4. It isobserved an inhibition of the oxygen evolution in all studied peaspecies after 3 h of high-light treatment (Fig. 4). The decrease of thefunctional active PSII centers (the amplitude A) correlates withdiminishing of the ratio A1/A2 (Fig. 4), as well as of both k1 and k2rate constants (data not shown), which indicates delayed oxygen
Fig. 2. Development of the non-photochemical quenching (NPQ) at room temperaturein leaves from Chlorotica XV/1422 (-), Auralia wt (B), Costata 2/133 (6) and Borec wt(7). Mean values� SD were determined from five independent experiments.

Fig. 3. Time-courses of the initial oxygen burst at continuous irradiation (450 mmol photonm�2 s�1) of pea thylakoid membranes (A and B) isolated from non-irradiated leaves (1)and leaves subjected to 3 h high-light irradiation (1300 mmolm�2 s�1) at room temperature (2). Oscillation patterns of the oxygen flash yields in dark-adapted thylakoid membranes(C and D) from non-irradiated (1) and high-light irradiated leaves (2). (A, C) Auralia wt; (B, D) Borec wt. Thylakoid membranes were suspended in a medium containing: 40 mMHEPES (pH 7.6), 10 mM NaCl, 5 mM MgCl2 and 400 mM sucrose. The chlorophyll concentration was 150 mg ml�1. Time-constant of the electrode is less than 2 ms. Polarographsensitivity is 1.5 V mA�1 for initial oxygen burst measurements and 3 V mA�1 for flash oxygen measurements.
K.G. Dankov et al. / Plant Physiology and Biochemistry 49 (2011) 629e635632
burst kinetics after high-light treatment. Results revealed that theoxygen evolution parameters are stronger influenced in ChloroticaXV/1422 and Auralia wt in comparisonwith Costata 2/133 and Borecwt (Fig. 4). The decrease in the ratio A1/A2 could be attributed to aninhibition of the PSII centers in grana thylakoids, which evolveoxygen through the non-cooperative Kok’s mechanism (the fast A1
Fig. 4. Changes of the kinetic parameters of the oxygen flash yields and the initialoxygen burst of thylakoid membranes, isolated from leaves after 3 h high-lightillumination (1300 mmolm�2 s�1). Values are represented as percentage of the valuesfor untreated control samples, considered as 100%. Y3 e the oxygen flash yield of thethird flash. A e the amplitude of the oxygen burst under continuous illumination. A1
and A2 are the amplitudes of the fast and slow components of the oxygen burst decay,respectively. Average data are from five independent experiments. The statisticaldifferences between values of the parameters are obtained when the mutant ChloroticaXV/1422 is compared with Auralia wt, mutant Costata 2/133 and Borec wt, determinedwith Student’s t-test (*P< 0.05, **P< 0.01).
component) and/or an increased proportion of PSII centers instroma thylakoids (the slow A2 component). On the other hand, thediminution in k1 and k2 rate constants after high-light treatmentcould be consequence from modification of PSII in grana andstroma thylakoid membranes.
The evaluated values of the initial S0eS1 dark distribution ofredox states, misses and double hits (data not shown), using theKok’s model [see Ref. 29], showed that the S0 populations of PSIIcenters increase during high-light treatment for the all investigatedpea species with about 13%, while misses and double hits enhancewith about 8 and 2.5%, respectively.
3.3. Lipid and fatty acid composition of the thylakoid membranes
The lipid composition of the thylakoid membranes from studiedpea plants is shown in Table 1. Comparison of the relative amount ofthe different lipid classes in the studied thylakoid membranesindicates that in the mutant Chlorotica XV/1422 the amount ofmonogalactosyldiacylglycerol (MGDG) is the smallest, whereas thedigalactosyldiacylglycerol (DGDG) content is bigger than that inother investigated pea species (Table 1). These findings could be
Table 1Lipid class composition of thylakoid membranes from different pea lines (molarpercentage of the total lipids). MGDG e monogalactosyldiacylglycerol, DGDG e
digalactosyldiacylglycerol, SQDG e sulfoquinovosyldiacylglycerol, PG e
phosphatidylglycerol.
Lipidclasses
Chlorotica XV/1422(mol %)
Auralia wt(mol %)
Costata 2/133(mol %)
Borec wt(mol %)
MGDG 35 45 47 48DGDG 43 35 35 37SQDG 7 6 5 5PG 12 11 11 9Others 3 3 2 1
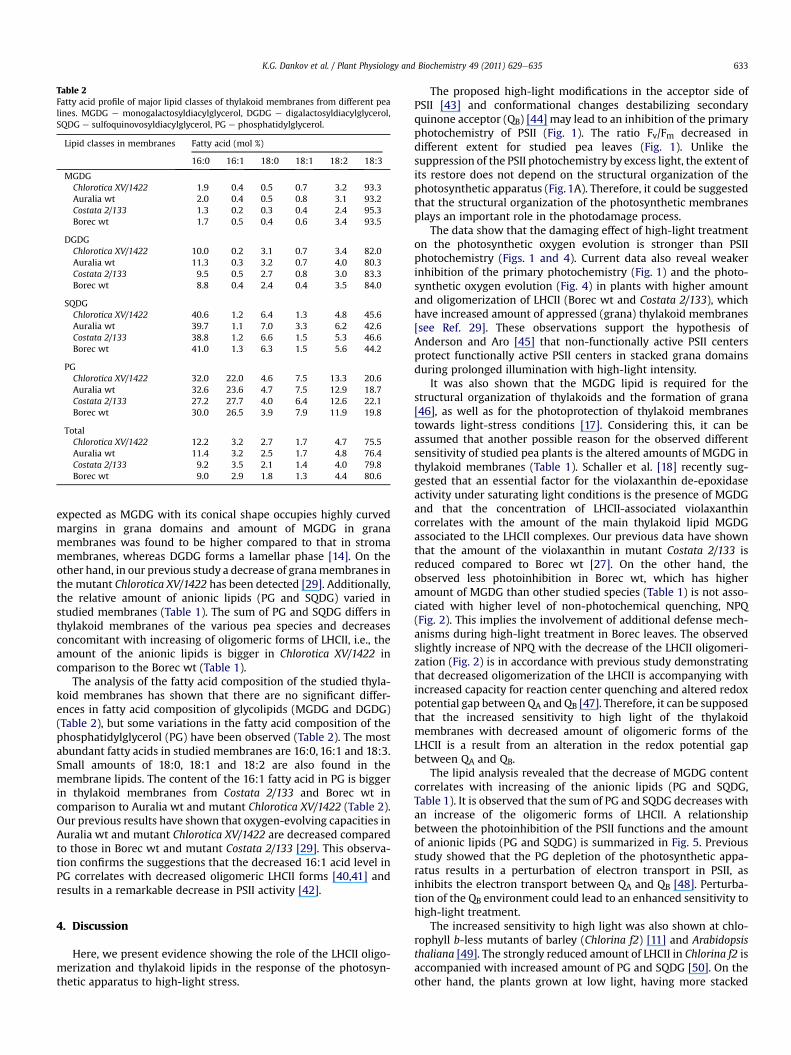
Table 2Fatty acid profile of major lipid classes of thylakoid membranes from different pealines. MGDG e monogalactosyldiacylglycerol, DGDG e digalactosyldiacylglycerol,SQDG e sulfoquinovosyldiacylglycerol, PG e phosphatidylglycerol.
Lipid classes in membranes Fatty acid (mol %)
16:0 16:1 18:0 18:1 18:2 18:3
MGDGChlorotica XV/1422 1.9 0.4 0.5 0.7 3.2 93.3Auralia wt 2.0 0.4 0.5 0.8 3.1 93.2Costata 2/133 1.3 0.2 0.3 0.4 2.4 95.3Borec wt 1.7 0.5 0.4 0.6 3.4 93.5
DGDGChlorotica XV/1422 10.0 0.2 3.1 0.7 3.4 82.0Auralia wt 11.3 0.3 3.2 0.7 4.0 80.3Costata 2/133 9.5 0.5 2.7 0.8 3.0 83.3Borec wt 8.8 0.4 2.4 0.4 3.5 84.0
SQDGChlorotica XV/1422 40.6 1.2 6.4 1.3 4.8 45.6Auralia wt 39.7 1.1 7.0 3.3 6.2 42.6Costata 2/133 38.8 1.2 6.6 1.5 5.3 46.6Borec wt 41.0 1.3 6.3 1.5 5.6 44.2
PGChlorotica XV/1422 32.0 22.0 4.6 7.5 13.3 20.6Auralia wt 32.6 23.6 4.7 7.5 12.9 18.7Costata 2/133 27.2 27.7 4.0 6.4 12.6 22.1Borec wt 30.0 26.5 3.9 7.9 11.9 19.8
TotalChlorotica XV/1422 12.2 3.2 2.7 1.7 4.7 75.5Auralia wt 11.4 3.2 2.5 1.7 4.8 76.4Costata 2/133 9.2 3.5 2.1 1.4 4.0 79.8Borec wt 9.0 2.9 1.8 1.3 4.4 80.6
K.G. Dankov et al. / Plant Physiology and Biochemistry 49 (2011) 629e635 633
expected as MGDG with its conical shape occupies highly curvedmargins in grana domains and amount of MGDG in granamembranes was found to be higher compared to that in stromamembranes, whereas DGDG forms a lamellar phase [14]. On theother hand, in our previous study a decrease of granamembranes inthe mutant Chlorotica XV/1422 has been detected [29]. Additionally,the relative amount of anionic lipids (PG and SQDG) varied instudied membranes (Table 1). The sum of PG and SQDG differs inthylakoid membranes of the various pea species and decreasesconcomitant with increasing of oligomeric forms of LHCII, i.e., theamount of the anionic lipids is bigger in Chlorotica XV/1422 incomparison to the Borec wt (Table 1).
The analysis of the fatty acid composition of the studied thyla-koid membranes has shown that there are no significant differ-ences in fatty acid composition of glycolipids (MGDG and DGDG)(Table 2), but some variations in the fatty acid composition of thephosphatidylglycerol (PG) have been observed (Table 2). The mostabundant fatty acids in studied membranes are 16:0, 16:1 and 18:3.Small amounts of 18:0, 18:1 and 18:2 are also found in themembrane lipids. The content of the 16:1 fatty acid in PG is biggerin thylakoid membranes from Costata 2/133 and Borec wt incomparison to Auralia wt and mutant Chlorotica XV/1422 (Table 2).Our previous results have shown that oxygen-evolving capacities inAuralia wt and mutant Chlorotica XV/1422 are decreased comparedto those in Borec wt and mutant Costata 2/133 [29]. This observa-tion confirms the suggestions that the decreased 16:1 acid level inPG correlates with decreased oligomeric LHCII forms [40,41] andresults in a remarkable decrease in PSII activity [42].
4. Discussion
Here, we present evidence showing the role of the LHCII oligo-merization and thylakoid lipids in the response of the photosyn-thetic apparatus to high-light stress.
The proposed high-light modifications in the acceptor side ofPSII [43] and conformational changes destabilizing secondaryquinone acceptor (QB) [44] may lead to an inhibition of the primaryphotochemistry of PSII (Fig. 1). The ratio Fv/Fm decreased indifferent extent for studied pea leaves (Fig. 1). Unlike thesuppression of the PSII photochemistry by excess light, the extent ofits restore does not depend on the structural organization of thephotosynthetic apparatus (Fig. 1A). Therefore, it could be suggestedthat the structural organization of the photosynthetic membranesplays an important role in the photodamage process.
The data show that the damaging effect of high-light treatmenton the photosynthetic oxygen evolution is stronger than PSIIphotochemistry (Figs. 1 and 4). Current data also reveal weakerinhibition of the primary photochemistry (Fig. 1) and the photo-synthetic oxygen evolution (Fig. 4) in plants with higher amountand oligomerization of LHCII (Borec wt and Costata 2/133), whichhave increased amount of appressed (grana) thylakoid membranes[see Ref. 29]. These observations support the hypothesis ofAnderson and Aro [45] that non-functionally active PSII centersprotect functionally active PSII centers in stacked grana domainsduring prolonged illumination with high-light intensity.
It was also shown that the MGDG lipid is required for thestructural organization of thylakoids and the formation of grana[46], as well as for the photoprotection of thylakoid membranestowards light-stress conditions [17]. Considering this, it can beassumed that another possible reason for the observed differentsensitivity of studied pea plants is the altered amounts of MGDG inthylakoid membranes (Table 1). Schaller et al. [18] recently sug-gested that an essential factor for the violaxanthin de-epoxidaseactivity under saturating light conditions is the presence of MGDGand that the concentration of LHCII-associated violaxanthincorrelates with the amount of the main thylakoid lipid MGDGassociated to the LHCII complexes. Our previous data have shownthat the amount of the violaxanthin in mutant Costata 2/133 isreduced compared to Borec wt [27]. On the other hand, theobserved less photoinhibition in Borec wt, which has higheramount of MGDG than other studied species (Table 1) is not asso-ciated with higher level of non-photochemical quenching, NPQ(Fig. 2). This implies the involvement of additional defense mech-anisms during high-light treatment in Borec leaves. The observedslightly increase of NPQ with the decrease of the LHCII oligomeri-zation (Fig. 2) is in accordance with previous study demonstratingthat decreased oligomerization of the LHCII is accompanying withincreased capacity for reaction center quenching and altered redoxpotential gap between QA and QB [47]. Therefore, it can be supposedthat the increased sensitivity to high light of the thylakoidmembranes with decreased amount of oligomeric forms of theLHCII is a result from an alteration in the redox potential gapbetween QA and QB.
The lipid analysis revealed that the decrease of MGDG contentcorrelates with increasing of the anionic lipids (PG and SQDG,Table 1). It is observed that the sum of PG and SQDG decreases withan increase of the oligomeric forms of LHCII. A relationshipbetween the photoinhibition of the PSII functions and the amountof anionic lipids (PG and SQDG) is summarized in Fig. 5. Previousstudy showed that the PG depletion of the photosynthetic appa-ratus results in a perturbation of electron transport in PSII, asinhibits the electron transport between QA and QB [48]. Perturba-tion of the QB environment could lead to an enhanced sensitivity tohigh-light treatment.
The increased sensitivity to high light was also shown at chlo-rophyll b-less mutants of barley (Chlorina f2) [11] and Arabidopsisthaliana [49]. The strongly reduced amount of LHCII in Chlorina f2 isaccompanied with increased amount of PG and SQDG [50]. On theother hand, the plants grown at low light, having more stacked
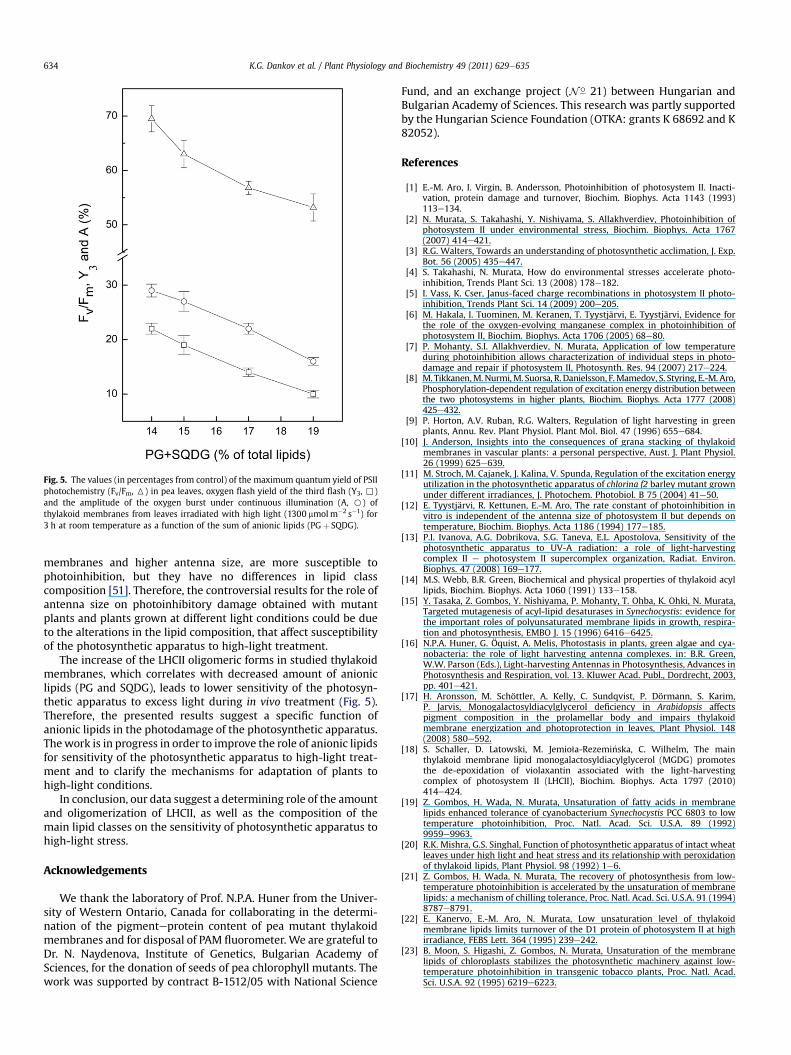
Fig. 5. The values (in percentages from control) of the maximum quantum yield of PSIIphotochemistry (Fv/Fm, 6) in pea leaves, oxygen flash yield of the third flash (Y3, ,)and the amplitude of the oxygen burst under continuous illumination (A, B) ofthylakoid membranes from leaves irradiated with high light (1300 mmolm�2 s�1) for3 h at room temperature as a function of the sum of anionic lipids (PGþ SQDG).
K.G. Dankov et al. / Plant Physiology and Biochemistry 49 (2011) 629e635634
membranes and higher antenna size, are more susceptible tophotoinhibition, but they have no differences in lipid classcomposition [51]. Therefore, the controversial results for the role ofantenna size on photoinhibitory damage obtained with mutantplants and plants grown at different light conditions could be dueto the alterations in the lipid composition, that affect susceptibilityof the photosynthetic apparatus to high-light treatment.
The increase of the LHCII oligomeric forms in studied thylakoidmembranes, which correlates with decreased amount of anioniclipids (PG and SQDG), leads to lower sensitivity of the photosyn-thetic apparatus to excess light during in vivo treatment (Fig. 5).Therefore, the presented results suggest a specific function ofanionic lipids in the photodamage of the photosynthetic apparatus.Thework is in progress in order to improve the role of anionic lipidsfor sensitivity of the photosynthetic apparatus to high-light treat-ment and to clarify the mechanisms for adaptation of plants tohigh-light conditions.
In conclusion, our data suggest a determining role of the amountand oligomerization of LHCII, as well as the composition of themain lipid classes on the sensitivity of photosynthetic apparatus tohigh-light stress.
Acknowledgements
We thank the laboratory of Prof. N.P.A. Huner from the Univer-sity of Western Ontario, Canada for collaborating in the determi-nation of the pigmenteprotein content of pea mutant thylakoidmembranes and for disposal of PAM fluorometer. We are grateful toDr. N. Naydenova, Institute of Genetics, Bulgarian Academy ofSciences, for the donation of seeds of pea chlorophyll mutants. Thework was supported by contract B-1512/05 with National Science
Fund, and an exchange project (N� 21) between Hungarian andBulgarian Academy of Sciences. This research was partly supportedby the Hungarian Science Foundation (OTKA: grants K 68692 and K82052).
References
[1] E.-M. Aro, I. Virgin, B. Andersson, Photoinhibition of photosystem II. Inacti-vation, protein damage and turnover, Biochim. Biophys. Acta 1143 (1993)113e134.
[2] N. Murata, S. Takahashi, Y. Nishiyama, S. Allakhverdiev, Photoinhibition ofphotosystem II under environmental stress, Biochim. Biophys. Acta 1767(2007) 414e421.
[3] R.G. Walters, Towards an understanding of photosynthetic acclimation, J. Exp.Bot. 56 (2005) 435e447.
[4] S. Takahashi, N. Murata, How do environmental stresses accelerate photo-inhibition, Trends Plant Sci. 13 (2008) 178e182.
[5] I. Vass, K. Cser, Janus-faced charge recombinations in photosystem II photo-inhibition, Trends Plant Sci. 14 (2009) 200e205.
[6] M. Hakala, I. Tuominen, M. Keranen, T. Tyystjärvi, E. Tyystjärvi, Evidence forthe role of the oxygen-evolving manganese complex in photoinhibition ofphotosystem II, Biochim. Biophys. Acta 1706 (2005) 68e80.
[7] P. Mohanty, S.I. Allakhverdiev, N. Murata, Application of low temperatureduring photoinhibition allows characterization of individual steps in photo-damage and repair if photosystem II, Photosynth. Res. 94 (2007) 217e224.
[8] M. Tikkanen,M.Nurmi,M. Suorsa, R.Danielsson, F.Mamedov, S. Styring, E.-M. Aro,Phosphorylation-dependent regulation of excitation energy distribution betweenthe two photosystems in higher plants, Biochim. Biophys. Acta 1777 (2008)425e432.
[9] P. Horton, A.V. Ruban, R.G. Walters, Regulation of light harvesting in greenplants, Annu. Rev. Plant Physiol. Plant Mol. Biol. 47 (1996) 655e684.
[10] J. Anderson, Insights into the consequences of grana stacking of thylakoidmembranes in vascular plants: a personal perspective, Aust. J. Plant Physiol.26 (1999) 625e639.
[11] M. Stroch, M. Cajanek, J. Kalina, V. Spunda, Regulation of the excitation energyutilization in the photosynthetic apparatus of chlorina f2 barley mutant grownunder different irradiances, J. Photochem. Photobiol. B 75 (2004) 41e50.
[12] E. Tyystjärvi, R. Kettunen, E.-M. Aro, The rate constant of photoinhibition invitro is independent of the antenna size of photosystem II but depends ontemperature, Biochim. Biophys. Acta 1186 (1994) 177e185.
[13] P.I. Ivanova, A.G. Dobrikova, S.G. Taneva, E.L. Apostolova, Sensitivity of thephotosynthetic apparatus to UV-A radiation: a role of light-harvestingcomplex II e photosystem II supercomplex organization, Radiat. Environ.Biophys. 47 (2008) 169e177.
[14] M.S. Webb, B.R. Green, Biochemical and physical properties of thylakoid acyllipids, Biochim. Biophys. Acta 1060 (1991) 133e158.
[15] Y. Tasaka, Z. Gombos, Y. Nishiyama, P. Mohanty, T. Ohba, K. Ohki, N. Murata,Targeted mutagenesis of acyl-lipid desaturases in Synechocystis: evidence forthe important roles of polyunsaturated membrane lipids in growth, respira-tion and photosynthesis, EMBO J. 15 (1996) 6416e6425.
[16] N.P.A. Huner, G. Öquist, A. Melis, Photostasis in plants, green algae and cya-nobacteria: the role of light harvesting antenna complexes. in: B.R. Green,W.W. Parson (Eds.), Light-harvesting Antennas in Photosynthesis, Advances inPhotosynthesis and Respiration, vol. 13. Kluwer Acad. Publ., Dordrecht, 2003,pp. 401e421.
[17] H. Aronsson, M. Schöttler, A. Kelly, C. Sundqvist, P. Dörmann, S. Karim,P. Jarvis, Monogalactosyldiacylglycerol deficiency in Arabidopsis affectspigment composition in the prolamellar body and impairs thylakoidmembrane energization and photoprotection in leaves, Plant Physiol. 148(2008) 580e592.
[18] S. Schaller, D. Latowski, M. Jemio1a-Rezemi�nska, C. Wilhelm, The mainthylakoid membrane lipid monogalactosyldiacylglycerol (MGDG) promotesthe de-epoxidation of violaxantin associated with the light-harvestingcomplex of photosystem II (LHCII), Biochim. Biophys. Acta 1797 (2010)414e424.
[19] Z. Gombos, H. Wada, N. Murata, Unsaturation of fatty acids in membranelipids enhanced tolerance of cyanobacterium Synechocystis PCC 6803 to lowtemperature photoinhibition, Proc. Natl. Acad. Sci. U.S.A. 89 (1992)9959e9963.
[20] R.K. Mishra, G.S. Singhal, Function of photosynthetic apparatus of intact wheatleaves under high light and heat stress and its relationship with peroxidationof thylakoid lipids, Plant Physiol. 98 (1992) 1e6.
[21] Z. Gombos, H. Wada, N. Murata, The recovery of photosynthesis from low-temperature photoinhibition is accelerated by the unsaturation of membranelipids: a mechanism of chilling tolerance, Proc. Natl. Acad. Sci. U.S.A. 91 (1994)8787e8791.
[22] E. Kanervo, E.-M. Aro, N. Murata, Low unsaturation level of thylakoidmembrane lipids limits turnover of the D1 protein of photosystem II at highirradiance, FEBS Lett. 364 (1995) 239e242.
[23] B. Moon, S. Higashi, Z. Gombos, N. Murata, Unsaturation of the membranelipids of chloroplasts stabilizes the photosynthetic machinery against low-temperature photoinhibition in transgenic tobacco plants, Proc. Natl. Acad.Sci. U.S.A. 92 (1995) 6219e6223.

K.G. Dankov et al. / Plant Physiology and Biochemistry 49 (2011) 629e635 635
[24] P. Vijyan, J. Browse, Photoinhibition in mutant of Arabidopsis deficient inthylakoid unsaturation, Plant Physiol. 129 (2002) 876e885.
[25] N. Mizusawa, I. Sakurai, N. Sato, H. Wada, Lack of digalactosyldiacylglycerolincreases the sensitivity of Synechocystis sp. PCC 6803 to high light stress,FEBS Lett. 583 (2009) 718e722.
[26] I. Domonkos, H. Laczkó-Dobos, Z. Gombos, Lipid-assisted proteineproteininteractions that support photosynthetic and other cellular activities, Prog.Lipid Res. 47 (2008) 422e435.
[27] A.G. Dobrikova, R.M. Morgan, A.G. Ivanov, E.L. Apostolova, I.B. Petkanchin,N.P.A. Huner, S.G. Taneva, Electric properties of thylakoid membranes frompea mutants with modified carotenoid and chlorophylleprotein complexescomposition, Photosynth. Res. 65 (2000) 165e174.
[28] A.G. Dobrikova, A.G. Ivanov, E.L. Apostolova, N. Naydenova, I.B. Petkanchin,S.G. Taneva, Contribution of LHCII complex to the electric properties ofthylakoid membranes. in: N. Gözükirmizi (Ed.), Proceedings of the SecondBalkan Botanical Congress. Marmara University, Istanbul, 2001, pp. 75e80.
[29] E.L. Apostolova, A.G. Dobrikova, P.I. Ivanova, I.B. Petkanchin, S.G. Taneva,Relationship between the organization of the PSII supercomplex and thefunctions of the photosynthetic apparatus, J. Photochem. Photobiol. B 83(2006) 114e122.
[30] M.A. Harrison, A. Melis, Organization and stability of polypeptides associatedwith the chlorophyll aeb light-harvesting complex of photosystem-II, PlantCell Physiol. 33 (1992) 627e637.
[31] N. Sato, N. Murata, Membrane lipids, Methods Enzymol. 167 (1988) 251e256.[32] M. Kitajima, W. Butler, Quenching of chlorophyll fluorescence and primary
photochemistry in chloroplasts by dibromothymoquinone, Biochim. Biophys.Acta 376 (1975) 105e115.
[33] M. Havaux, R. Strasser, H. Greppin, A theoretical and experimental analysis ofthe qP and qN coefficients of chlorophyll fluorescence quenching and theirrelation to photochemical and nonphotochemical events, Photosynth. Res. 27(1991) 41e55.
[34] W. Bilger, O. Björkman, Role of the xanthophyll cycle in photoprotectionelucidated by measurements of light-induced absorbance changes, fluores-cence and photosynthesis in leaves of Hedera canariensis, Photosynth. Res. 25(1990) 173e185.
[35] Y. Zeinalov, An equipment for investigations of photosynthetic oxygenproduction reactions, Bulg. J. Plant Physiol. 28 (2002) 57e67.
[36] B. Kok, B. Forbush, M. McGloin, Co-operation of charges in photosynthetic O2evolution. I. A linear four step mechanism, Photochem. Photobiol. 11 (1970)457e475.
[37] R.J. Ellis, Inhibitionof chloroplastproteinsynthesisby lincomycinand2-(4-methyl-2,6-dinitroanilino)-N-methylpropionamide, Phytochemistry 14 (1975) 89e93.
[38] P. Mulo, S. Pursiheimo, C.-X. Hou, T. Tyystjärvi, E.-M. Aro, Multiple effects ofantibiotics on chloroplast and nuclear gene expression, Funct. Plant Biol. 30(2003) 1097e1103.
[39] Y. Zeinalov, L. Maslenkova, Mechanisms of photosynthetic oxygen evolution.in: M. Pessarakli (Ed.), Handbook of Photosynthesis. Marcel Dekker, New York,1996, pp. 129e150.
[40] N.P.A. Huner, M. Krol, J. Williams, E. Maissan, P. Low, D. Roberts, J. Thompson,Low temperature development induces a specific decrease in trans-3-hex-adecenoic acid content which influences LHCII organization, Plant Physiol. 84(1987) 12e18.
[41] G. Dubertret, C. Gerard-Hirne, A. Tremolieres, Importance of trans-D3-hexadecenoic acid containing phosphatidylglycerol in the formation of thetrimeric light-harvesting complex in Chlamydomonas, Plant Physiol. Bio-chem. 40 (2002) 829e836.
[42] A. Maani, G. Dubertret, M. Delrieu, O. Roche, A. Tremolieres, Mutants ofChlamydomonas reinhardtii affected in phosphatidylglycerol metabolism andthylakoid biogenesis, Eur. J. Biochem. 271 (1998) 609e619.
[43] I. Vass, K. Cser, O. Cheregi, Molecular mechanisms of light stress of photo-synthesis, Ann. N.Y. Acad. Sci. 1113 (2007) 114e122.
[44] I. Ohad, N. Adir, H. Koike, D.J. Kyle, Y. Inoue, Mechanism of photoinhibition invivo. A reversible light-induced conformational change of reaction center II isrelated to an irreversible modification of the D1 protein, J. Biol. Chem. 265(1990) 1972e1979.
[45] J. Anderson, E.-M. Aro, Grana stacking and protection of photosystem II inthylakoid membranes of higher plant leaves under sustained high irradiance:an hypothesis, Photosynth. Res. 41 (1994) 315e326.
[46] P. Jarvis, P. Dormann, C.A. Peto, J. Lutes, C. Benning, J. Chory, Galactolipiddeficiency and abnormal chloroplast development in the ArabidopsisMGD synthase 1 mutant, Proc. Natl. Acad. Sci. U.S.A. 97 (2000)8175e8179.
[47] A.G. Ivanov, M. Krol, E.L. Apostolova, R.M. Morgan-Kiss, N. Naydenova,N.P.A. Huner, P.V. Sane, Oligomerization state of LHCII modulates the redoxproperties of the acceptor side of photosystem II in Costata-2/133 mutant ofpea, Physiol. Mol. Biol. Plants 11 (2005) 199e207.
[48] Z. Gombos, Z. Várkonyi, M. Hagio, M. Iwaki, L. Kovács, K. Masamoto, S. Itoh,H. Wada, Phosphatidylglycerol requirement for the function of electronacceptor plastoquinone Q(B) in the photosystem II reaction center,Biochemistry 41 (2002) 3796e3802.
[49] E.H. Kim, X.P. Li, R. Razeghifard, J.M. Anderson, K.K. Niyogi, B.J. Pogson,W.S. Chow, The multiple roles of light-harvesting chlorophyll a/beproteincomplexes define structure and optimize function of Arabidopsis chloroplasts:a study using two chlorophyll b-less mutants, Biochim. Biophys. Acta 1787(2009) 973e984.
[50] P. Bolton, J. Wharte, J.L. Harwood, The lipid composition of a barley mutantlacking chlorophyll b, Biochem. J. 174 (1978) 67e72.
[51] D. Chapman, J. De Felice, J. Barber, Polar lipid composition of chloroplastthylakoids isolated from leaves grown under different lighting conditions,Photosynth. Res. 8 (1986) 257e265.




















