
Leukemogenic properties of NUP98-PMX1 are linked to NUP98 and homeodomain sequence functions but not...
12
ORIGINAL ARTICLE Leukemogenic properties of NUP98-PMX1 are linked to NUP98 and homeodomain sequence functions but not to binding properties of PMX1 to serum response factor K Hirose 1 , C Abramovich 1 , B Argiropoulos 1 and RK Humphries 1,2 1 Terry Fox Laboratory, British Columbia Cancer Agency, Vancouver, British Columbia, Canada and 2 Department of Medicine, University of British Columbia, Vancouver, British Columbia, Canada PMX1 is a member of a non-clustered homeobox gene family, not normally expressed in hematopoietic cells, and first identified for its role in enhancing the binding of the serum response factor (SRF) to the serum responsive element (SRE). PMX1 has never been linked to leukemia on its own, raising the possibility of unique mechanisms underlying the oncogenicity of NUP98-PMX1. To elucidate the leukemogenic potential of NUP98-PMX1, we compared the effects of PMX1 and NUP98-PMX1 and, through strategic mutations, the involvement of the SRE in NUP98-PMX1-mediated leukemia. NUP98- PMX1, but not PMX1, had potent ability to impair differentiation, promote proliferation of myeloid progeni- tors, induce lethal myeloproliferative disease and to activate a number of genes previously linked to leukemic stem cells. Similar to NUP98-HOX fusions, the trans- forming potential of NUP98-PMX1 required the NUP98 portion and DNA-binding capability of the PMX1 home- odomain and collaborated with Meis1 to induce more rapid onset myeloproliferative-like myeloid leukemia. The transforming activity of NUP98-PMX1 was independent of its ability to interact with SRF. These findings provide novel evidence of the contributory role of the NUP98 sequence in conferring leukemogenic properties on a partner gene and point to common leukemogenic path- ways for NUP98-PMX1 and NUP98-clustered HOX fusions. Oncogene (2008) 27, 6056–6067; doi:10.1038/onc.2008.210; published online 7 July 2008 Keywords: homeodomain; nucleoporin; hematopoiesis Introduction The nucleoporin gene NUP98 on chromosome 11p15 is involved in chromosomal rearrangements with at least 20 different partners (Slape and Aplan, 2004; Nakamura, 2005; Romana et al., 2006; Jankovic et al., 2008). NUP98 translocations have been described in de novo acute and chronic leukemias, as well as therapy- related myelodysplastic syndrome and therapy-related acute myeloid leukemia (AML). Although these NUP98-associated diseases are relatively rare, they are recurring and are characterized by their aggressiveness and disappointing treatment outcome. NUP98 is a component of the nuclear pore complex that regulates the transport of proteins and RNA– protein complexes between the cytoplasm and nucleus (Griffis et al., 2002, 2003). It belongs to a subgroup of nuclear pore proteins characterized by phenylalanine– glycine repeats (FG repeats), which are interaction sites for karyopherins (importins, exportins) that serve to guide transport through the nuclear pore complex (Radu et al., 1995). The role of nucleoporins in the malignant transformation of hematopoietic cells is only partly understood. The importance of the FG repeats in the etiology of disease is underscored by their preserva- tion in all the fusion proteins (Slape and Aplan, 2004). Moreover, the FG repeats are also conserved in the t(6;9)(p23;q34) chromosome translocation involving NUP214 (or CAN), which is found in patients with AML (von Lindern et al., 1992). In the context of the translocations of NUP98 with HOXA9 and PMX1, the FG repeats of NUP98 have been shown to confer strong transcriptional activity to the chimeric proteins (Kasper et al., 1999; Bai et al., 2006). Moreover, it has been demonstrated that the NUP98 FG repeats of NUP98- PMX1, NUP98-HOXA9 and NUP98-HOXC11 act as a docking site for both a transcriptional co-activator and correpressor (Bai et al., 2006). The most frequently observed NUP98 fusion partners belong to the homeobox gene family, including mostly clustered HOX genes and two non-clustered HOX genes, paired related homeobox 1 and 2 (PMX1 and PMX2) (Slape and Aplan, 2004; Romana et al., 2006). The involvement of these latter two genes in leukemia is intriguing, as in contrast to clustered Hox genes, neither gene has been previously implicated in normal hemato- poiesis or leukemogenesis. PMX1 knockout mice show limb and vertebral skeletal defects, but can survive soon after birth, suggestive of a normal hematopoietic system (Martin et al., 1995; ten Berge et al., 1998). Moreover, PMX1 expression is not detected in hematopoietic cells (Kern et al., 1994). Received 25 July 2006; revised 30 May 2008; accepted 30 May 2008; published online 7 July 2008 Correspondence: Dr RK Humphries, Terry Fox Laboratory, British Columbia Cancer Agency, 675 West 10th Avenue, Vancouver, British Columbia, Canada V5Z 1L3. E-mail: [email protected] Oncogene (2008) 27, 6056–6067 & 2008 Macmillan Publishers Limited All rights reserved 0950-9232/08 $32.00 www.nature.com/onc
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Leukemogenic properties of NUP98-PMX1 are linked to NUP98 and homeodomain sequence functions but not...

ORIGINAL ARTICLE
Leukemogenic properties of NUP98-PMX1 are linked to NUP98
and homeodomain sequence functions but not to binding properties
of PMX1 to serum response factor
K Hirose1, C Abramovich1, B Argiropoulos1 and RK Humphries1,2
1Terry Fox Laboratory, British Columbia Cancer Agency, Vancouver, British Columbia, Canada and 2Department of Medicine,University of British Columbia, Vancouver, British Columbia, Canada
PMX1 is a member of a non-clustered homeobox genefamily, not normally expressed in hematopoietic cells, andfirst identified for its role in enhancing the binding of theserum response factor (SRF) to the serum responsiveelement (SRE). PMX1 has never been linked to leukemiaon its own, raising the possibility of unique mechanismsunderlying the oncogenicity of NUP98-PMX1. Toelucidate the leukemogenic potential of NUP98-PMX1,we compared the effects of PMX1 and NUP98-PMX1and, through strategic mutations, the involvement of theSRE in NUP98-PMX1-mediated leukemia. NUP98-PMX1, but not PMX1, had potent ability to impairdifferentiation, promote proliferation of myeloid progeni-tors, induce lethal myeloproliferative disease and toactivate a number of genes previously linked to leukemicstem cells. Similar to NUP98-HOX fusions, the trans-forming potential of NUP98-PMX1 required the NUP98portion and DNA-binding capability of the PMX1 home-odomain and collaborated with Meis1 to induce morerapid onset myeloproliferative-like myeloid leukemia. Thetransforming activity of NUP98-PMX1 was independentof its ability to interact with SRF. These findings providenovel evidence of the contributory role of the NUP98sequence in conferring leukemogenic properties on apartner gene and point to common leukemogenic path-ways for NUP98-PMX1 and NUP98-clustered HOXfusions.Oncogene (2008) 27, 6056–6067; doi:10.1038/onc.2008.210;published online 7 July 2008
Keywords: homeodomain; nucleoporin; hematopoiesis
Introduction
The nucleoporin gene NUP98 on chromosome 11p15 isinvolved in chromosomal rearrangements with atleast 20 different partners (Slape and Aplan, 2004;Nakamura, 2005; Romana et al., 2006; Jankovic et al.,
2008). NUP98 translocations have been described inde novo acute and chronic leukemias, as well as therapy-related myelodysplastic syndrome and therapy-relatedacute myeloid leukemia (AML). Although theseNUP98-associated diseases are relatively rare, they arerecurring and are characterized by their aggressivenessand disappointing treatment outcome.
NUP98 is a component of the nuclear pore complexthat regulates the transport of proteins and RNA–protein complexes between the cytoplasm and nucleus(Griffis et al., 2002, 2003). It belongs to a subgroup ofnuclear pore proteins characterized by phenylalanine–glycine repeats (FG repeats), which are interaction sitesfor karyopherins (importins, exportins) that serve toguide transport through the nuclear pore complex(Radu et al., 1995). The role of nucleoporins in themalignant transformation of hematopoietic cells is onlypartly understood. The importance of the FG repeats inthe etiology of disease is underscored by their preserva-tion in all the fusion proteins (Slape and Aplan, 2004).Moreover, the FG repeats are also conserved in thet(6;9)(p23;q34) chromosome translocation involvingNUP214 (or CAN), which is found in patients withAML (von Lindern et al., 1992). In the context of thetranslocations of NUP98 with HOXA9 and PMX1, theFG repeats of NUP98 have been shown to confer strongtranscriptional activity to the chimeric proteins (Kasperet al., 1999; Bai et al., 2006). Moreover, it has beendemonstrated that the NUP98 FG repeats of NUP98-PMX1, NUP98-HOXA9 and NUP98-HOXC11 act as adocking site for both a transcriptional co-activator andcorrepressor (Bai et al., 2006).
The most frequently observed NUP98 fusion partnersbelong to the homeobox gene family, including mostlyclustered HOX genes and two non-clustered HOX genes,paired related homeobox 1 and 2 (PMX1 and PMX2)(Slape and Aplan, 2004; Romana et al., 2006). Theinvolvement of these latter two genes in leukemia isintriguing, as in contrast to clustered Hox genes, neithergene has been previously implicated in normal hemato-poiesis or leukemogenesis. PMX1 knockout mice showlimb and vertebral skeletal defects, but can survive soonafter birth, suggestive of a normal hematopoietic system(Martin et al., 1995; ten Berge et al., 1998). Moreover,PMX1 expression is not detected in hematopoietic cells(Kern et al., 1994).
Received 25 July 2006; revised 30 May 2008; accepted 30 May 2008;published online 7 July 2008
Correspondence: Dr RK Humphries, Terry Fox Laboratory, BritishColumbia Cancer Agency, 675 West 10th Avenue, Vancouver,British Columbia, Canada V5Z 1L3.E-mail: [email protected]
Oncogene (2008) 27, 6056–6067& 2008 Macmillan Publishers Limited All rights reserved 0950-9232/08 $32.00
www.nature.com/onc

The rare t(1;11)(q23;p15) chromosome translocation,juxtaposing PMX1 and NUP98, has been identified intherapy-related AML patients (ten Berge et al., 1998;Nakamura et al., 1999; Zhang et al., 2007). Whether theleukemogenic activity of NUP98-PMX1 is due to theaberrant expression of PMX1 in hematopoieticprecursors driven by the NUP98 ubiquitous promoter,and/or whether there is a contribution of the NUP98moiety to the leukemogenic potential of the fusionprotein remains unknown.
PMX1 has a unique homeodomain (HD), which ishighly similar to the Drosophila Paired domain (Cserjesiet al., 1992; Grueneberg et al., 1992). Over the 60 aminoacids comprising the HD, the PMX1 HD is 70%identical to the Paired domain and 50% to the clusteredHOX HD, raising the possibility of unique mechanismsunderlying its leukemogenic role. This is supported bythe fact that the PMX1 HD, but not the clustered HOXHD, can impart transcriptional activity to the serumresponsive element (SRE) through interaction with theserum responsive factor (SRF), which regulates a varietyof genes including the oncogenes c-fos, fosB and junB(Grueneberg et al., 1992). Moreover, SRF is known tobe activated by oncogenic viral products, such as HCVcore protein, HBV pX protein (Kato et al., 2000) andHTLV1 tax protein (Alexandre and Verrier, 1991). Thisraises the interesting possibility that the leukemogenicprocess induced by NUP98-PMX1 is linked to theability of PMX1 to mediate SRF activity. This notionhas gained further support from the studies indicatingthat NUP98-PMX1 can exert a dominant negative effecton SRF-induced activation of c-fos expression in COS-7and HeLa cell lines (Bai et al., 2006). However, therelevance of these findings, obtained in non-hemato-poietic experimental models, remains unclear.
In this study, we sought to shed light on themechanisms underlying the leukemogenic potential ofNUP98-PMX1. For that purpose, we exploited themurine hematopoietic model and explored and com-pared the effects of PMX1 and NUP98-PMX1 inhematopoiesis; we studied the involvement of the SREin NUP98-PMX1-mediated leukemia, and explored themolecular pathways used by NUP98-PMX1 in theinduction of leukemia.
Results
NUP98-PMX1 but not PMX1 has the ability to impairdifferentiation and promote proliferation of hematopoieticprogenitor cells in vitroTo begin to unravel the role of PMX1 and NUP98-PMX1 in hematopoietic cell growth and differentiation,we exploited a series of in vitro hematopoietic assays.For that purpose, PMX1 or NUP98-PMX1 was over-expressed in primary murine bone marrow (BM) cells(see Figures 1a and b for description of the cDNAs andvectors used). Proviral integrity was verified by Southernblot analysis (data not shown) and the expression ofPMX1 and NUP98-PMX1 was confirmed by westernblot analysis (Figure 1c).
Immediately following retroviral infection and with-out any selection for transduced cells, BM cells wereplated in liquid culture containing interleukin-3, -6 andstem cell factor. The GFPþ population of NUP98-PMX1-transduced BM cells gradually increased fromthe input level of B15% to greater than 70% after 17days of culture (Figure 2a, left panel). The growthadvantage of NUP98-PMX1-transduced BM cells wasmore evident when comparing the in vitro growth ratesof the GFPþ cell populations. NUP98-PMX1-trans-duced BM cells showed a B40-fold increase in total cellnumber over 17 days compared with GFPþ control BMcells (Figure 2a, right panel). In contrast to this, in thePMX1-transduced BM cells, the GFPþ population wasdecreased from 29% of input level to less than 10% after17 days of culture (Figure 2a, left panel) and showeddecreased proliferation compared to GFPþ andNUP98-PMX1-transduced BM cells (Figure 2a, rightpanel), indicating a possible inhibitory/cytotoxic effectof the native protein.
These differential growth-promoting effects werefurther evident in colony-forming unit-spleen (CFU-S)assays, which detect primitive hematopoietic progenitorcells capable of forming spleen colonies. BM cellstransduced with GFPþ control, PMX1 or NUP98-PMX1 were isolated by fluorescence-activated cellsorting (FACS) 2 days post-transduction and trans-planted into lethally irradiated recipient mice. Theprogenitor cell frequency was equivalent among thedifferent experimental arms, whereas NUP98-PMX1-transduced BM cells produced much larger spleen
Figure 1 Constructs and retroviral vectors used in this study. (a)Schematic representation of NUP98, full-length PMX1 andNUP98-PMX1. Fusion break points are indicated with arrows.(b) Retroviral vectors used in this study to express the full-lengthFlag-tagged PMX1 and Flag tagged-NUP98-PMX1 fusion inmurine bone marrow cells. (c) Western blot analysis of total celllysates from transfected 293 T cells detected by anti-Flagantibodies.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6057
Oncogene

colonies (data not shown). More strikingly, after 7 daysof liquid culture, NUP98-PMX1-transduced BM cellsincreased the absolute yield of CFU-S 100-fold com-pared with numbers before culture, or 5000-fold whencompared with the number of CFU-S recovered fromGFPþ -transduced control cultures (Figure 2b). Con-versely, PMX1-transduced BM cells had no impact onthe CFU-S numbers after 1 week of liquid culturecompared with GFPþ control (Figure 2b). FACSanalysis of spleen colonies derived from NUP98-PMX1 BM cells indicated a marked increase in theproportion of granulocytic (Gr-1) myeloid cells(74.3±0.7 versus 24.9±3.4% for GFPþ control)and a significant decrease in the proportion ofTER119þ erythroid cells (16.2±1.2 versus 69.0±3.8%in GFPþ control). C-kit expression was also retained ina high proportion of NUP98-PMX1-transduced BMcells in spleen colonies (58.9±3.3 versus 9.4±3.2% in
GFPþ control). These findings suggest that NUP98-PMX1, but not PMX1, impairs BM differentiation.
The effect of NUP98-PMX1 on hematopoietic differ-entiation was further evaluated after 3 weeks in liquidculture by morphology and immunophenotyping.Wright–Geisma stained cytospin preparations andFACS analysis of GFPþ control and PMX1-transducedBM cells showed highly granulated mast cell-likemorphology (data not shown) and high-level expressionof c-Kit and FceR (Figure 2c), consistent with terminaldifferentiation. Under the same condition, the NUP98-PMX1-transduced BM cells showed immature myeloid-like morphology (data not shown) and phenotype(negative expression of FceR and low levels of c-Kit)(Figure 2c).
The ability of NUP98-PMX1, but not PMX1, topromote the proliferation of hematopoietic progenitorswas further confirmed by serial replating of BM
Figure 2 NUP98-PMX1, but not PMX1, promotes proliferation and impairs differentiation of transduced bone marrow (BM) cellsin vitro. (a) Cell expansion of unsorted transduced cells expressed as % GFPþ cells (left panel) or total GFPþ cell number (right panel).Results of a representative experiment from triplicate cultures are shown. (b) Total number of day-12 colony-forming unit-spleenderived from 1� 106 transduced cells after 1 week of liquid culture (mean±s.d., n¼ 5). (c) Fluorescence-activated cell sorting profilesof transduced BM cells cultured for 3 weeks. Representative results are shown. (d) Total numbers of granulo-monocytic progenitorcells in serial replating. Cumulative yield shown for the initial plating of 500 transduced BM cells.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6058
Oncogene

clonogenic progenitor cells in methylcellulose culture.NUP98-PMX1 strongly increased the number of sec-ondary myeloid colony-forming cells in methylcelluloseculture (Figure 2d). Furthermore, most of the colonieswere CFU-granulocyte macrophage (Gr-1, Mac1) withhigh-proliferative potential (made up of several thou-sand cells after 8–10 days culture) and could be seriallyreplated over three passages. In contrast, PMX1-transduced BM cells behaved as the GFPþ control,forming only mast cell colonies that lost their replatingpotential after the secondary plates (Figure 2d).
NUP98-PMX1 but not PMX1 induces a competitivemyeloid growth advantage in vivoTo assess the role of PMX1 and NUP98-PMX1 inhematopoietic development in vivo, PMX1 or NUP98-PMX1-transduced BM cells were transplanted inlethally irradiated recipient mice. Each recipient mousewas transplanted with 5� 105–1� 106 BM cells withoutselection (50–270� 103 GFPþ cells per recipient).Between 4 and 6 months post-transplantation, micetransplanted with NUP98-PMX1-transduced BM cellsexhibited a three- to fourfold expansion of GFPþ
peripheral blood (PB) mononuclear cells compared withthe time of transplantation (Figure 3a). In contrast, thefraction of GFPþ WBC in PMX1 mice was decreased by10-fold (to 3±3.3%) compared with their proportion atthe time of transplant (30±5.3%) (Figure 3a). More-over, the donor cells transduced with NUP98-PMX1preferentially yielded myeloid progeny, as shown by afourfold increase in the percentage of Gr-1/Mac1positive cells and two- to fourfold decrease in thepercentage of the lymphoid markers CD4/CD8 in theGFPþ compartment (Figure 3b). PMX1-transducedcells retained normal multilineage differentiation ability,with proportions of B220, Gr-1/Mac1 and CD4/CD8positive cells similar to GFPþ control mice (Figure 3b).These observations were further supported by anincrease in the absolute numbers of circulating myeloidcells in NUP98-PMX1 mice compared with those inGFPþ control or PMX1 mice (Figure 3c).
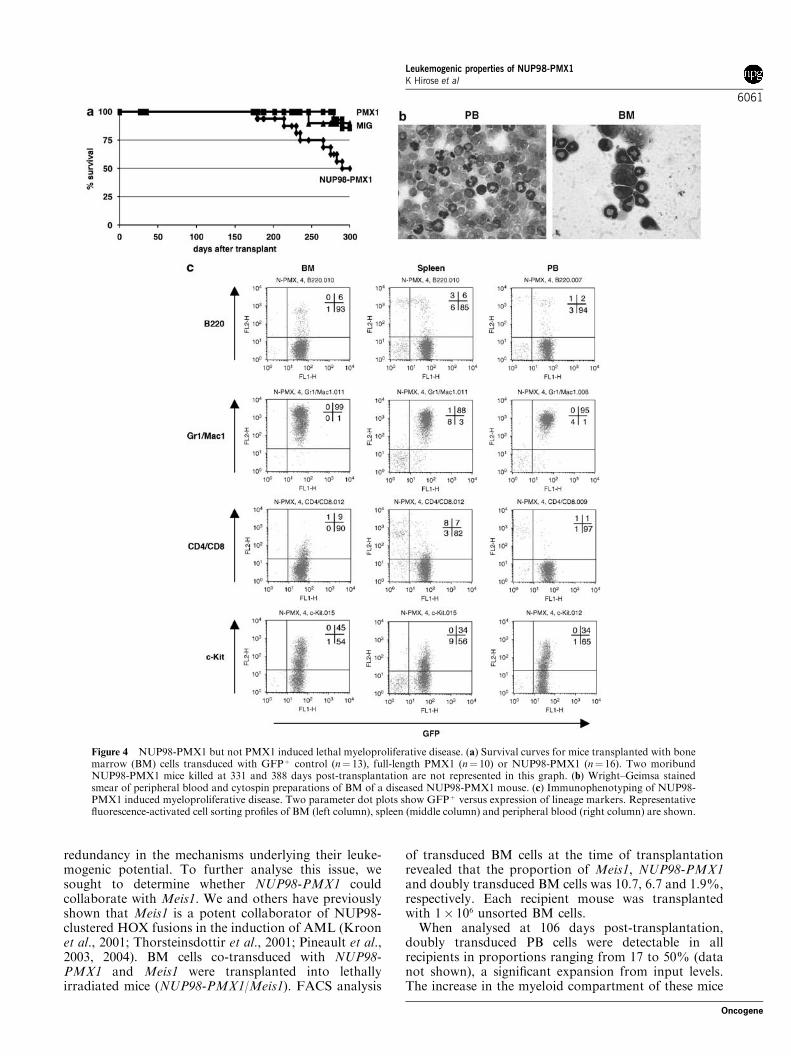
NUP98-PMX1 but not PMX1 causes a lethalmyeloproliferative diseaseConsistent with the ability of NUP98-PMX1 to conferan in vivo growth advantage and to promote myelopoi-esis, a large proportion (8 out of 16) of NUP98-PMX1mice succumbed to a myeloproliferative disorder be-tween 170 and 300 days post-transplantation (meandisease latency of these mice is 246 days) (Figure 4a).Although a small number of GFPþ control and PMX1mice also died in this follow-up period, no hematologi-cal abnormalities were detected in these animals. At thetime of killing, NUP98-PMX1 mice displayed severeanemia, leukocytosis and splenomegaly (SupplementaryTable 1). Morphological analysis of PB and BM smearsshowed predominance of band forms and generally lowpercentage of metamyelocytes (Figure 4b). Cytospinpreparations (data not shown) and FACS analysis(Figure 4c) of splenocytes revealed significant infiltra-
tion by cells with myeloid characteristics. FACS analysisof PB, BM and spleen revealed that the majority of thecells were NUP98-PMX1-transduced cells (GFPþ cells).The myeloid nature of the disease was confirmed by thefinding that most of the GFPþ cells expressed Mac1/Gr-1 but not the lymphoid markers B220, CD4 or CD8(Figure 4c). The accumulation of myeloid cells in BM,PB and spleen confirmed the diagnosis of myeloproli-ferative disease. Proviral integrity and clonality of thedisease was assessed by Southern blot analysis, whichrevealed a mono- or pauci-clonal disease (data notshown). Hematological disease in animals transplantedwith GFPþ control or PMX1 BM cells over 12 months (allthe hematological parameters tested were within normalrange, data not shown) was not detected, further confirm-ing the inability of PMX1 to perturb hematopoiesis.
Taken together, the results presented above demon-strate the ability of NUP98-PMX1 to cause myelopro-liferative disease consistent with perturbeddifferentiation and growth promotion of myeloidprogenitors. Ectopic expression of PMX1 in BM cellsdid not perturb hematopoiesis, in contrast to thereported leukemogenic effects of the native clusteredHOX partners of NUP98.
The transforming potential of NUP98-PMX1 requiresthe NUP98 portion and the DNA binding activityof the PMX1 HD but not the interaction of the PMX1sequence with the SRFOur data indicating that PMX1 lacks leukemogenicactivity suggests a critical role for the NUP98 portion inthe activity of the fusion protein. The NUP98 sequencealone does not confer growth advantage in liquid culture(Gurevich et al., 2004), nor does it promote in vitrotransformation (Kasper et al., 1999), strongly suggestingan essential contribution of the PMX1 portion. Thus, inan attempt to functionally dissect the contributingdomains of the fusion protein to its leukemogenicpotential, a series of NUP98-PMX1 mutants were testedin vitro.
To examine if the transforming ability of NUP98-PMX1 is dependent on the SRF pathway, we generateda NUP98-PMX1 mutant which has two amino-acidsubstitutions in the second helix of the PMX1 HD, thusremoving its ability to interact with the SRF (NUP98-PMX1(ED))(Simon et al., 1997). The NUP98-PMX1(ED) mutant conferred a competitive growthadvantage equivalent to wild-type NUP98-PMX1.Indeed, the total cell numbers of NUP98-PMX1(ED)-transduced BM cells was 17- to 20-fold greater thanGFPþ control cells after 3 weeks of culture, effectscomparable to wild-type NUP98-PMX1 (Figure 5a). InCFU-S assays, NUP98-PMX1(ED) expanded the spleencolony-forming progenitor cells after 7 days in liquidculture to levels similar to the wild-type NUP98-PMX1(data not shown). This was further confirmed inmethylcellulose cultures, in which NUP98-PMX1(ED)behaved similarly to wild-type NUP98-PMX1, forminglarge CFU-GM colonies that could be serially replatedat least three times (Figure 5b).
Leukemogenic properties of NUP98-PMX1K Hirose et al
6059
Oncogene

To confirm that the NUP98 portion plays a criticalrole in leukemogenesis, we made a deletion mutant ofNUP98-PMX1, which lacks the entire portion ofNUP98 (PMX1 exon 2). Furthermore, to assess theimportance of DNA binding of PMX1 in the leukemo-genic potential of NUP98-PMX1, the highly conservedasparagine residue at position 51 in the HD of PMX1was substituted for glutamine (NUP98-PMX1 (N51Q)).This mutation eliminates the DNA-binding activity ofthe HD without affecting its binding to SRF (Gruene-berg et al., 1992; Vershon et al., 1995). In contrast toNUP98-PMX1 and NUP98-PMX1(ED), PMX1 exon 2-and NUP98-PMX1(N51Q)-transduced BM cells had nocompetitive growth advantage in liquid culture andgenerated equal numbers of CFU-S compared withGFPþ control BM cells (Figure 5a and data not shown,respectively). Moreover, in methylcellulose progenitor
assays, PMX1 exon 2- and NUP98-PMX1(N51Q)-transduced BM cells rapidly lost their replating potential(Figure 5b) and produced only mast cell colonies onsecondary replating (data not shown).
Collectively, these results demonstrate that thetransforming activity of NUP98-PMX1 is independentof the SRF pathway, requires the NUP98 portion andmust retain the DNA-binding ability of PMX1, empha-sizing the similarity between NUP98-PMX1 andNUP98-HOX fusions.
Both NUP98-PMX1 and clustered NUP98-HOXcollaborate with Meis1 to induce leukemiaThe similarities between the effects of NUP98-PMX1and NUP98-HOX in hematopoietic development andthe essential role of their HDs indicate a strong
Figure 3 Characterization of the transduced peripheral white blood cells from mice transplanted with MIG control, PMX1 orNUP98-PMX1. (a) Peripheral blood (PB) chimerism. Data are plotted as percentage (mean±s.d.) of GFPþ cells in the initialtransplant innoculum (arrow) compared with in PB WBCs 4–6 months post-transplant. Results are shown for two or threeindependent experiments with at least four mice in each group. (b) Immunophenotyping of transduced WBC. Following lysis of the redcells, PB from transplanted mice was collected between 4 and 6 months post-transplantation and stained with antibodies against B220,Gr1/Mac1 and CD4/CD8. Results are shown for mean±s.d. of each cohort of mice from two or three independent experiments. Solidbar, GFPþ cells; open bar, GFP� cells. (c) Absolute numbers of B220, Gr1/Mac1 or CD4/CD8 positive cells in the PB of transducedmice between 4 and 6 months post-transplantation. Results are shown for mean value of mice in two or three independent experiments.Open bar, GFP� cells; solid bar, GFPþ cells.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6060
Oncogene
redundancy in the mechanisms underlying their leuke-mogenic potential. To further analyse this issue, wesought to determine whether NUP98-PMX1 couldcollaborate with Meis1. We and others have previouslyshown that Meis1 is a potent collaborator of NUP98-clustered HOX fusions in the induction of AML (Kroonet al., 2001; Thorsteinsdottir et al., 2001; Pineault et al.,2003, 2004). BM cells co-transduced with NUP98-PMX1 and Meis1 were transplanted into lethallyirradiated mice (NUP98-PMX1/Meis1). FACS analysis
of transduced BM cells at the time of transplantationrevealed that the proportion of Meis1, NUP98-PMX1and doubly transduced BM cells was 10.7, 6.7 and 1.9%,respectively. Each recipient mouse was transplantedwith 1� 106 unsorted BM cells.
When analysed at 106 days post-transplantation,doubly transduced PB cells were detectable in allrecipients in proportions ranging from 17 to 50% (datanot shown), a significant expansion from input levels.The increase in the myeloid compartment of these mice
Figure 4 NUP98-PMX1 but not PMX1 induced lethal myeloproliferative disease. (a) Survival curves for mice transplanted with bonemarrow (BM) cells transduced with GFPþ control (n¼ 13), full-length PMX1 (n¼ 10) or NUP98-PMX1 (n¼ 16). Two moribundNUP98-PMX1 mice killed at 331 and 388 days post-transplantation are not represented in this graph. (b) Wright–Geimsa stainedsmear of peripheral blood and cytospin preparations of BM of a diseased NUP98-PMX1 mouse. (c) Immunophenotyping of NUP98-PMX1 induced myeloproliferative disease. Two parameter dot plots show GFPþ versus expression of lineage markers. Representativefluorescence-activated cell sorting profiles of BM (left column), spleen (middle column) and peripheral blood (right column) are shown.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6061
Oncogene

was donor-derived and was similar to that seen inNUP98-PMX1 mice (data not shown). The leukemiccollaboration between NUP98-PMX1 and Meis1 wassupported by the development of myeloid leukemia inthe NUP98-PMX1/Meis1 mice with a median diseaselatency of 191 days (Figure 6a). These mice presentedwith massive ascites and splenomegaly. The myeloidnature of the disease was confirmed by FACS analysis(Figure 6b). Morphological analysis of cytospin pre-parations revealed an accumulation of myeloblasts inBM (ranging from 11 to 19%, n¼ 4) and PB (Figure 6c),consistent with a diagnosis of myeloproliferative-likemyeloid leukemia (Kogan et al., 2002). Histologicalanalysis of these mice revealed that leukemic cellsinfiltrated the spleen, liver and kidney and destroyedtheir normal architecture (data not shown). The diseasewas transplantable where all secondary recipientstransplanted with doses of 1� 106 and 0.1� 106 BMcells succumbed to disease 40 days post-transplantation(Figure 6a).
Differential transcriptional properties of PMX1and NUP98-PMX1Gene expression profiling on primary AML samples andstudies on mixed lineage leukemia oncogenic fusiongenes have revealed that a common characteristic ofAML is dysregulated expression of select HOX genesand that this may be critical for leukemic stem cell(LSC) function (extensively reviewed by (Eklund, 2006,2007; Argiropoulos and Humphries, 2007)). To gain
insight into the molecular mechanisms underlying theleukemogenicity of NUP98-PMX1, we quantitativelyassessed Hoxa cluster gene expression in mouse stem cellvirus-yellow fluorescent protein (MIY) empty vectorcontrol, PMX1, NUP98-PMX1 and NUP98-PMX1-N51Q-transduced lineage depleted, Sca1þ (Lin-Scaþ )BM cells. Similar to NUP98-HOXD13 (Palmqvist et al.,2007), NUP98-NSD1 (Wang et al., 2007) and NUP98-HHEX (Jankovic et al., 2008), NUP98-PMX1 stronglyactivated multiple Hoxa cluster genes, includingHoxa5 to Hoxa10, compared with MIY control andthe non-leukemogenic PMX1 and NUP98-PMX1(N51Q)(Figure 7a, left panel). Furthermore, onlyNUP98-PMX1 activated expression of Bmi1, Flt3 andCd34 (Figure 7a, right panel), genes associatedwith hematopoietic and LSCs (Wang et al., 2006;Argiropoulos and Humphries, 2007).
To further study the similarities between NUP98-clustered HOX and NUP98-PMX1, we investigatedtheir ability to activate common target genes. We havepreviously shown that overexpression of NUP98-HOXD13 leads to increased levels of Hoxa9 mRNA(Pineault et al., 2003; Palmqvist et al., 2007). Thus, wefirst determined whether this effect is mediated by theability of NUP98-HOXD13 to activate the Hoxa9promoter and whether this can be mimicked byNUP98-PMX1. As shown in Figure 7b (left panel),luciferase assays performed in 293 T cells demonstratedthat both NUP98-PMX1 and NUP98-HOXD13 acti-vated the Hoxa9 promoter linked to a luciferase reportergene. Interestingly, PMX1 was inactive suggesting that
Figure 5 Transforming activity of NUP98-PMX1 is independent of serum response factor binding ability, but dependent on its intacthomeodomain and the NUP98 portion. (a) Fold-expansion of bone marrow (BM) cells transduced with the indicated constructs inliquid culture over 3 weeks are shown (mean±s.d., n¼ 2). (b) Total number of granulo-monocytic colonies in serial replating.Cumulative yield shown for the initial plating of 500 transduced BM cells.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6062
Oncogene

fusion to NUP98 alters its transactivation properties ortarget specificity. In contrast, expression of an SRE-luciferase reporter construct was enhanced by bothPMX1 and NUP98-PMX1, but not by NUP98-HOXD13, providing further evidence of dissociationof the leukemogenic activity of these NUP98-fusions
and the ability to activate expression through the SRFpathway.
To test whether the differences in Hoxa9 transcrip-tional activation reflect differential binding capacities ofPMX1 or NUP98-PMX1 to Hoxa9, chromatin immuno-precipitation experiments were conducted on highly
Figure 6 NUP98-PMX1 cooperates with Meis1 to induce fatal myeloproliferative-like myeloid leukemia. (a) Survival curves formice transplanted with bone marrow (BM) cells transduced with Meis1 alone (n¼ 5), or MEIS1þNUP98-PMX1 (n¼ 10).MEIS1þNUP98-PMX1 mice developed disease significantly earlier when compared to NUP98-PMX1 mice (see Figure 4a), asdetermined by a log-rank test (Po0.0005). (b) Fluorescence-activated cell sorting phenotype for BM and peripheral blood (PB) cells ofa control and diseased NUP98-PMX1/Meis1 mouse. (c) Wright–Geimsa stained smear of a cytospin preparation of BM and PB of aleukemic NUP98-PMX1 mouse showing an accumulation of myeloblasts (arrows).
Leukemogenic properties of NUP98-PMX1K Hirose et al
6063
Oncogene

purified Lin-Scaþ BM cells transduced with PMX1,NUP98-PMX1 and NUP98-PMX1(N51Q). As shown inFigure 7c, PMX1 and NUP98-PMX1 associate with aspecific Hoxa9 regulatory sequence (R1) with similarfold enrichments over their respective controls. TheDNA-binding impaired NUP98-PMX1(N51Q) failed toimmunoprecipitate the R1 fragment, indicating thatPMX1 and NUP98-PMX1 directly bind to the R1region of the Hoxa9 promoter by the HD. These resultsprovide strong evidence that Hoxa9 is a direct target ofboth PMX1 and NUP98-PMX1 and that fusion toNUP98 imparts transactivating functions to PMX1.
Discussion
In this study we demonstrate: (1) that NUP98-PMX1but not native PMX1 has leukemogenic potential; (2)that the NUP98 portion and the PMX1-HD DNAbinding domain but not the SRE binding sequences areimportant for NUP98-PMX1-induced leukemogenesis;(3) that NUP98-PMX1 and NUP98-clustered HOX fusiongenes share leukemogenic pathways; (4) that NUP98-PMX1 but not PMX1 activates a number of genesassociated with an LSC signature and (5) that PMX1and NUP98-PMX1 bind to Hoxa9 regulatory sequencesbut only NUP98-PMX1 activates robust transcription,illustrating the transactivating properties of NUP98.
NUP98 translocations involving Paired mesodermhomeobox genes describe a unique class of homeoboxgene-associated leukemias, linking human leukemia tothe de novo expression of non-clustered HOX genesnormally not involved in hematopoiesis. Indeed, PMX1is normally expressed in a mesodermally restrictedpattern in embryos and most abundantly in cardiac,skeletal and smooth muscle tissues in adults. However,we show here that overexpression of NUP98-PMX1 inBM cells has a potent effect in the differentiation andsurvival of primitive hematopoietic cells, leading to theinduction of a myeloproliferative disease in mice. Theseresults are in striking contrast to the effect of over-expression of PMX1, which resulted in a rapid reductionof the proportion of PMX1-transduced WBCs in vitroand in vivo, suggesting that it decreases the long-termengraftment ability of hematopoietic stem cells and/orrestricts their proliferation.
Many of the in vitro and in vivo effects induced byNUP98-PMX1 are similar to those observed followingoverexpression of NUP98-clustered HOX genes. Com-
mon characteristics include the growth advantage oftransduced BM cells in vitro, as well as the induction of alethal myeloproliferative disease in transplanted miceand the ability to collaborate with Meis1 in theinduction of myeloid leukemia. The possibility thatdifferent NUP98 fusions share common leukemogenicpathways is supported by our findings that NUP98-PMX1 and NUP98-HOXD13 activate Hoxa9 transcri-ption. While the leukemogenic effects of NUP98-PMX1in the murine BM transplantation model appear to beweaker and delayed compared with other reportedNUP98-HOX fusions (Kroon et al., 2001; Thorsteins-dottir et al., 2001; Pineault et al., 2003, 2004), thesediscrepancies could reflect activation of non-overlappinggenetic pathways, susceptibility to additional collabor-ating events and/or differences in experimental metho-dology. The latter includes variations in retroviral genetransfer and the number of BM cells transplanted, bothof which dictate the frequency of the LSC transplantedand thus the potency of the oncogene.
The non-leukemogenic nature of PMX1, which differsfrom the strong leukemogenic potential of the nativeclustered Hox partners of NUP98 (Pineault et al., 2004),clearly underscores the potential of NUP98 to conferoncogenic activity to an otherwise non-leukemogenicpartner. However, the activity of NUP98-PMX1 isclearly dependent on the PMX1 portion. Our structure-function analysis revealed that PMX1-mediated tran-scriptional activation through the SRF pathway isdispensable, but its DNA binding ability was criticalfor transformation and activation of Hoxa9. Theimportance of the PMX1 HD supports previous studiessuggesting that the HD is the most significant contribu-tion of the HOX partner in NUP98 translocationsinvolving clustered Hox genes (Kroon et al., 2001;Pineault et al., 2003). Our results strongly argue that thelinkage of PMX1 to SRF mediated pathways (Bai et al.,2006) may not be relevant to the leukemogenic activityof NUP98-PMX1.
Several non-exclusive mechanisms could explain thecontribution of the NUP98 and the PMX1 sequences toNUP98-PMX1-induced leukemogenesis. First, differentstudies suggest that NUP98-PMX1 could function as anaberrant transcription factor that regulates expressionof downstream target genes critical for transformation.Indeed, Bai et al. (2006) have shown that the NUP98FG repeats of NUP98-PMX1 may act as a docking sitefor both transcriptional co-activators and correpressors,and thus confer aberrant transcription activity to the
Figure 7 NUP98-PMX1, but not PMX1 or NUP98-PMX1(N51S), activates genes previously linked to leukemic stem cells.(a) Multiple Hoxa cluster genes, including Hoxa5 to Hoxa10, are activated in NUP98-PMX1-transduced Lin-Sca1þ BM cellscompared with MIY control, PMX1 or NUP98-PMX1(N51S) transduced cells. Activation of Cd34, Bmi1 and Flt3 by NUP98-PMX1.(b) NUP98-PMX1 and NUP-HOXD13 differ in their ability to activate Hox promoter versus serum response factor (SRE) reporters. Atotal of 293 T cells were transfected with pHoxA9/TAL-Luc (left panel) or SRE-Luc (right panel) reporter plasmids together withPMX1, NUP98-PMX1 or NUP98-HOXD13. Plotted are the relative luciferase activities over the basal activity observed with thereporter plasmid alone. (c) Schematic representation of the murine Hoxa9 locus showing the putative transcription start site (þ 1) andthe relative positions of the amplicons used in the chromatin immunoprecipitation (ChIP) experiments. Lin-Scaþ BM cells weretransduced with the indicated construct, fluorescence-activated cell sorting selected and subjected to ChIP analysis. Immunopreci-pitated DNA was analysed by real-time PCR in triplicate using the indicated primer pairs. The data are expressed as fold enrichment ofDNA associated with the indicated Flag-tagged construct relative to the immunoglobulin G control.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6064
Oncogene

fused partner. In this respect, it has been shown thatNUP98-HOXA9, NUP98-PMX1 and NUP98-NSD1have increased transcriptional activity on specificpromoters when compared with their respective native
proteins (Kasper et al., 1999; Nakamura et al., 1999;Ghannam et al., 2004; Wang et al., 2007). Moreover,the transcriptional repressor HHEX is converted toa transcriptional activator when fused to NUP98
Leukemogenic properties of NUP98-PMX1K Hirose et al
6065
Oncogene

(Jankovic et al., 2008). Such effects are consistent withour observation that NUP98-PMX1 but not PMX1could enhance expression of Hoxa cluster genes. It isalso possible that NUP98 changes the target specificityof the native protein, as previously shown for NUP98-HOXA9 (Ghannam et al., 2004); however, this is notevident in the instance of the target gene Hoxa9, as bothPMX1 and NUP98-PMX1 associate with the samesequences. To date, only a few genes have been reportedto be direct targets of PMX1, such as tenascin-C(McKean et al., 2003), c-FOS (Grueneberg et al., 1992;Martin et al., 1997) and sonic hedgehog (Shh) (ten Bergeet al., 2001); however, it is unlikely that c-FOS or Shhplays a role in NUP98-PMX1-induced leukemia, as theirmRNA levels are not significantly changed in NUP98-PMX1 versus PMX1-transduced Lin-Scaþ BM (datanot shown). We now provide evidence that Hoxa9 is adirect target of both PMX1 and NUP98-PMX1 and thatderegulation of Hoxa9 by NUP98-PMX1 may contri-bute to its leukemogenic activity.
In addition to aberrant transcriptional activity, othercontributions of NUP98 to NUP98-PMX1-inducedleukemogenic transformation could not be excluded.For example, FG repeats are known to mediateinteractions with nuclear–cytoplasmic transport recep-tors (Radu et al., 1995) and thus it may be possible thatexpression of the NUP98 fusion affects the nucleartransport machinery leading to transformation. Thispossibility is supported by the selective changes innuclear pore complex stoichiometry and function seenafter disruption of NUP98 expression in mice (Wu et al.,2001). Furthermore, a fusion protein involving the FG-rich nucleoporin NUP214 interacts with hCM1, one ofthe nuclear export factors, causing its aberrant intracel-lular localization and the disorganization of nucleartransport, which might contribute to its oncogenicpotential (Saito et al., 2004). More recently, a deletionmutant of NUP98-HHEX lacking the 5 proximal FGrepeats and the GLEBS domain of NUP98 exhibitednuclear localization differences compared with wild-typeNUP98-HEXX (Jankovic et al., 2008). Moreover, apotential novel mechanism is suggested by the findingthat the GLEBS motif of NUP98 (retained in all the
NUP98 fusions) binds to the mRNA export factor Rae1to regulate chromosome segregation. Indeed, cells frommice that expressed small amounts of both Rae1 andNup98 have severe chromosomal instability, due to thepremature degradation of the mitotic regulator securing(Jeganathan et al., 2005). As the translocation ofNUP98 with PMX1 (or any other HOX partner)produces a nuclear fusion protein that has the potentialto sequester Rae1 in the nucleus and reduce the amountof NUP98 present at the nucleopore, it is tempting tospeculate that the leukemogenic potential of NUP98-HOX fusions could be, in part, mediated by defects inmitosis. The relevance of this potential mechanism inour model needs to be investigated.
In conclusion, we have demonstrated, for the firsttime, the leukemogenic potential of the paired-HOXgene PMX1 when fused to NUP98. Our results supporta critical role for the NUP98 moiety present in thetranslocation, as well as the DNA-binding capacity ofPMX1. Moreover, the discovery of common leukemo-genic pathways for NUP98-PMX1 and NUP98-clustered HOX fusions provides a framework forthe search for common target genes for potentialtherapeutic manipulation.
Materials and methods
Procedural details of cDNA constructs, retroviral vectors,Southern and Western blot analyses, cell cultivation, retroviralinfection, BM transplantation, in vitro hematopoietic assays,spleen colony-forming cell assay, gene expression analysis,transient tranfections, luciferase and chromatin immunopreci-pitation assays are provided in Supplementary methods.
Acknowledgements
This work was supported by the Terry Fox Foundation ofCanada and by the Canadian Institutes of Health forResearch. KH and BA were recipients of fellowships fromthe Leukemia and Lymphoma Society of Canada. We thankDrs Florian Kuchenbauer and Eric Yung for technicalassistance.
References
Alexandre C, Verrier B. (1991). Four regulatory elements in the humanc-fos promoter mediate transactivation by HTLV-1 Tax protein.Oncogene 6: 543–551.
Argiropoulos B, Humphries RK. (2007). Hox genes in hematopoiesisand leukemogenesis. Oncogene 26: 6766–6776.
Bai XT, Gu BW, Yin T, Niu C, Xi XD, Zhang J et al. (2006). Trans-repressive effect of NUP98-PMX1 on PMX1-regulated c-FOS genethrough recruitment of histone deacetylase 1 by FG repeats. Cancer
Res 66: 4584–4590.Cserjesi P, Lilly B, Bryson L, Wang Y, Sassoon DA, Olson EN. (1992).
MHox: a mesodermally restricted homeodomain protein that bindsan essential site in the muscle creatine kinase enhancer. Development
115: 1087–1101.Eklund EA. (2006). The role of HOX genes in myeloid leukemogen-
esis. Curr Opin Hematol 13: 67–73.Eklund EA. (2007). The role of HOX genes in malignant myeloid
disease. Curr Opin Hematol 14: 85–89.
Ghannam G, Takeda A, Camarata T, Moore MA, Viale A, YaseenNR. (2004). The oncogene Nup98-HOXA9 induces gene transcri-ption in myeloid cells. J Biol Chem 279: 866–875.
Griffis ER, Altan N, Lippincott-Schwartz J, Powers MA. (2002).Nup98 is a mobile nucleoporin with transcription-dependentdynamics. Mol Biol Cell 13: 1282–1297.
Griffis ER, Xu S, Powers MA. (2003). Nup98 localizes to both nuclearand cytoplasmic sides of the nuclear pore and binds to two distinctnucleoporin subcomplexes. Mol Biol Cell 14: 600–610.
Grueneberg DA, Natesan S, Alexandre C, Gilman MZ. (1992).Human and Drosophila homeodomain proteins that enhancethe DNA-binding activity of serum response factor. Science 257:1089–1095.
Gurevich RM, Aplan PD, Humphries RK. (2004). NUP98-topo-isomerase I acute myeloid leukemia-associated fusion gene haspotent leukemogenic activities independent of an engineeredcatalytic site mutation. Blood 104: 1127–1136.
Leukemogenic properties of NUP98-PMX1K Hirose et al
6066
Oncogene

Jankovic D, Gorello P, Liu T, Ehret S, La Starza R, Desjobert C et al.(2008). Leukemogenic mechanisms and targets of NUP98/HHEXfusion in acute myeloid leukemia (AML). Blood 111: 5672–5682.
Jeganathan KB, Malureanu L, van Deursen JM. (2005). The Rae1-Nup98 complex prevents aneuploidy by inhibiting securin degrada-tion. Nature 438: 1036–1039.
Kasper LH, Brindle PK, Schnabel CA, Pritchard CE, Cleary ML,van Deursen JM. (1999). CREB binding protein interacts withnucleoporin-specific FG repeats that activate transcription andmediate NUP98-HOXA9 oncogenicity. Mol Cell Biol 19: 764–776.
Kato N, Yoshida H, Ono-Nita SK, Kato J, Goto T, Otsuka M et al.(2000). Activation of intracellular signaling by hepatitis B and Cviruses: C-viral core is the most potent signal inducer. Hepatology
32: 405–412.Kern MJ, Argao EA, Birkenmeier EH, Rowe LB, Potter SS. (1994).Genomic organization and chromosome localization of the murinehomeobox gene Pmx. Genomics 19: 334–340.
Kogan SC, Ward JM, Anver MR, Berman JJ, Brayton C, Cardiff RDet al. (2002). Bethesda proposals for classification of nonlymphoidhematopoietic neoplasms in mice. Blood 100: 238–245.
Kroon E, Thorsteinsdottir U, Mayotte N, Nakamura T, Sauvageau G.(2001). NUP98-HOXA9 expression in hemopoietic stem cellsinduces chronic and acute myeloid leukemias in mice. EMBO J
20: 350–361.Martin JF, Bradley A, Olson EN. (1995). The paired-like homeo boxgene MHox is required for early events of skeletogenesis in multiplelineages. Genes Dev 9: 1237–1249.
Martin KA, Gualberto A, Kolman MF, Lowry J, Walsh K. (1997).A competitive mechanism of CArG element regulation by YY1 andSRF: implications for assessment of Phox1/MHox transcriptionfactor interactions at CArG elements. DNA Cell Biol 16: 653–661.
McKean DM, Sisbarro L, Ilic D, Kaplan-Alburquerque N,Nemenoff R, Weiser-Evans M et al. (2003). FAK induces expressionof Prx1 to promote tenascin-C-dependent fibroblast migration. J
Cell Biol 161: 393–402.Nakamura T. (2005). NUP98 fusion in human leukemia: dysregulationof the nuclear pore and homeodomain proteins. Int J Hematol 82:21–27.
Nakamura T, Yamazaki Y, Hatano Y, Miura I. (1999). NUP98 isfused to PMX1 homeobox gene in human acute myelogenousleukemia with chromosome translocation t(1;11)(q23;p15). Blood 94:741–747.
Palmqvist L, Pineault N, Wasslavik C, Humphries RK. (2007).Candidate genes for expansion and transformation of hematopoieticstem cells by NUP98-HOX fusion genes. PLoS ONE 2: e768.
Pineault N, Abramovich C, Ohta H, Humphries RK. (2004).Differential and common leukemogenic potentials of multipleNUP98-Hox fusion proteins alone or with Meis1. Mol Cell Biol
24: 1907–1917.Pineault N, Buske C, Feuring-Buske M, Abramovich C, Rosten P,Hogge DE et al. (2003). Induction of acute myeloid leukemia in mice
by the human leukemia-specific fusion gene NUP98-HOXD13 inconcert with Meis1. Blood 101: 4529–4538.
Radu A, Moore MS, Blobel G. (1995). The peptide repeat domain ofnucleoporin Nup98 functions as a docking site in transport acrossthe nuclear pore complex. Cell 81: 215–222.
Romana SP, Radford-Weiss I, Ben Abdelali R, Schluth C, Petit A,Dastugue N et al. (2006). NUP98 rearrangements in hematopoieticmalignancies: a study of the Groupe Francophone de CytogenetiqueHematologique. Leukemia 20: 696–706.
Saito S, Miyaji-Yamaguchi M, Nagata K. (2004). Aberrant intra-cellular localization of SET-CAN fusion protein, associated witha leukemia, disorganizes nuclear export. Int J Cancer 111: 501–507.
Simon KJ, Grueneberg DA, Gilman M. (1997). Protein and DNAcontact surfaces that mediate the selective action of the Phox1homeodomain at the c-fos serum response element. Mol Cell Biol 17:6653–6662.
Slape C, Aplan PD. (2004). The role of NUP98 gene fusions inhematologic malignancy. Leuk Lymphoma 45: 1341–1350.
Ten Berge D, Brouwer A, Korving J, Martin JF, Meijlink F. (1998).Prx1 and Prx2 in skeletogenesis: roles in the craniofacial region,inner ear and limbs. Development 125: 3831–3842.
Ten Berge D, Brouwer A, Korving J, Reijnen MJ, van Raaij EJ,Verbeek F et al. (2001). Prx1 and Prx2 are upstream regulators ofsonic hedgehog and control cell proliferation during mandibulararch morphogenesis. Development 128: 2929–2938.
Thorsteinsdottir U, Kroon E, Jerome L, Blasi F, Sauvageau G. (2001).Defining roles for HOX and MEIS1 genes in induction of acutemyeloid leukemia. Mol Cell Biol 21: 224–234.
Vershon AK, Jin Y, Johnson AD. (1995). A homeo domain proteinlacking specific side chains of helix 3 can still bind DNA and directtranscriptional repression. Genes Dev 9: 182–192.
Von Lindern M, Fornerod M, Soekarman N, van Baal S, Jaegle M,Hagemeijer A et al. (1992). Translocation t(6;9) in acutenon-lymphocytic leukaemia results in the formation of a DEK-CAN fusion gene. Baillieres Clin Haematol 5: 857–879.
Wang GG, Cai L, Pasillas MP, Kamps MP. (2007). NUP98-NSD1links H3K36 methylation to Hox-A gene activation and leukaemo-genesis. Nat Cell Biol 9: 804–812.
Wang GG, Pasillas MP, Kamps MP. (2006). Persistent transactivationby meis1 replaces hox function in myeloid leukemogenesis models:evidence for co-occupancy of meis1-pbx and hox-pbx complexeson promoters of leukemia-associated genes. Mol Cell Biol 26:3902–3916.
Wu X, Kasper LH, Mantcheva RT, Mantchev GT, Springett MJ, vanDeursen JM. (2001). Disruption of the FG nucleoporin NUP98causes selective changes in nuclear pore complex stoichiometry andfunction. Proc Natl Acad Sci USA 98: 3191–3196.
Zhang L, Alsabeh R, Mecucci C, La Starza R, Gorello P, Lee S et al.(2007). Rare t(1;11)(q23;p15) in therapy-related myelodysplasticsyndrome evolving into acute myelomonocytic leukemia: a case reportand review of the literature. Cancer Genet Cytogenet 178: 42–48.
Supplementary Information accompanies the paper on the Oncogene website (http://www.nature.com/onc)
Leukemogenic properties of NUP98-PMX1K Hirose et al
6067
Oncogene




















