
colombian teachers' expectations of poor and black students ...

ELSEVIER Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
Late Quaternary pollen records from the middle Caqueta river basin incentral Colombian Amazon
Hermann Behling Ł, Juan Carlos Berrio, Henry Hooghiemstra
University of Amsterdam, Hugo de Vries-Laboratory, Department of Palynology and Palaeo=Actuo-ecology, Kruislaan 318,1098 SM Amsterdam (Netherlands Centre for Geo-ecological Research ICG), Netherlands
Received 9 February 1998; revised version received 29 May 1998; accepted 1 June 1998
Abstract
Three pollen records are presented from the Pantano de Monica region (0º420S, 72º040W, 160 m elevation) on thelower terrace of Rio Caqueta of the central Colombian Amazonas. Ten radiocarbon dates from three cores indicate thatthe deposits are of Holocene age, but the pollen data suggest that the record may also contain the Late Glacial. The corePantano de Monica 1 covers the time interval from 11,150 BP (extrapolated) to 4730 BP. During the Late Glacial and earlyHolocene this swamp was smaller in size and waters were more shallow than today, with abundant Mauritia palm trees.This indicates that the lower terrace of the Caqueta River was better drained than today, which might be related to changesin the drainage system and=or drier conditions during that time. Late Glacial and early Holocene vegetational changesin the rain forests surrounding the swamp Pantano de Monica indicate successional stages, probably related to changesin the drainage system and=or climate changes. Presence of Podocarpus pollen grains up to 2.6% of the total sum (andinflux of 78 grains cm�2 yr�1) point to the regional presence of Podocarpus at the beginning of the Holocene. Evidence ofPodocarpus during glacial times in other pollen records from the Amazon basin has been taken as indicative of cooling.Core Pantano de Monica 2 was taken in a small swamp (1000 m distance to Pantano de Monica 1) and starts at 4000 BPwhen this location was well drained. Several changes in the composition of the rain forest are documented, such as increasein disturbance (increase of Cecropia) followed by a quick recovery (increase of Psychotria). Protium and Caryocar treetaxa were frequent, suggesting that the lower terrace area was well drained until 3080 BP, after which small swampsdeveloped. Core Pantano de Monica 3 is from the wet forest in between the two swamps and represents the period since3260 BP. Pollen data show a continuously increasing presence of palms, probably Euterpe, suggesting either a transitionfrom well-drained to less drained conditions in the lower terrace during the late Holocene, and=or a more intensive humanimpact of the rain forest on the lower terraces of Rio Caqueta. The three records from the lower terrace of Rio Caquetashow different forest compositions in the past and indicate that the rain forest environments were not stable during LateGlacial and Holocene times. Comparison with other data from lowland records of northwestern South America suggest thatclimate change is a major factor for environmental change in central Colombian Amazon. 1999 Elsevier Science B.V.All rights reserved.
Keywords: Late Quaternary; pollen analysis; Amazon rain forest; vegetation and climate history; Colombia
Ł Corresponding author. Tel.: C31 20 5257 844; Fax: C31 20 5257 662; E-mail: [email protected]
0031-0182/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 1 - 0 1 8 2 ( 9 8 ) 0 0 1 0 5 - 9

194 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
1. Introduction
Paleoenvironmental conditions of the tropicallowlands and in terms of changes in vegetationalcomposition and climate during last Glacial andHolocene times are poorly known. Data on the pa-leoecology of the Amazonian lowlands, the largesttropical rain forest ecosystem on earth, are lackingfor very large areas which allows for much specula-tion on its natural history. For example, the existenceof Amazonian rain forest refugia is subject to muchdebate in the last decades (e.g. Haffer, 1969; White-more and Prance, 1987; Colinvaux, 1993, 1996; Vander Hammen and Absy, 1994). Palynological studiesin the Amazon basin are rare because of difficult ac-cess and logistical problems. A better understandingof the history of this large area is of great importancefor a wide variety of scientific objectives, such aspalaeodata-model comparisons, sustainable manage-ment of the rain forest ecosystem, and aspects ofevolutionary biology.
The area of the middle Caqueta River is located inthe central part of the Colombian Amazonas. Duringthe last decade various geological, geomorpholog-ical, palynological, and botanical inventory studieshave been made in this area. Miocene palynos-tratigraphical and paleoenvironmental investigationswere done on outcrops along the Caqueta River (e.g.Hoorn, 1994a,b). Late Quaternary vegetation devel-opment, river dynamics, sedimentation, and climaticchange in the Mariname Island area and Quinchearea, both in the Caqueta River valley, have beenstudied by Van der Hammen et al. (1992a,b), Vander Hammen and Cleef (1992), and Urrego (1991,1994/1997).
Several palynological studies exist from the west-ern Amazonian rain forest area in Ecuador. Bush andColinvaux (1988), Bush et al. (1989, 1990), Colin-vaux et al. (1988a,b), Frost (1988), Liu and Colin-vaux (1985, 1988) and Lyons-Weiler (1992) studiedseveral lake sediments and sediment outcrops be-tween 300 and 1100 m elevation in the foothillsof the Andes and at low elevations in the EasternAndes.
Van der Hammen and Oscar Mohr collected in1987 a first core in the area of the Pantano de Mon-ica for a pilot study: the provisional pollen recordincluded two radiocarbon dates of full glacial and
Holocene age and showed an interesting example offorest dynamics in which Ilex played an importantrole in the Late Glacial. This study was not com-pleted but preliminary results are published by Vander Hammen (1998). Our objective is to continue thepalynological study of the sediments of the swampsof Pantano de Monica, to improve the chronologicalframework, to study the modern pollen rain, and tobring more detail on the composition and succes-sional stages of the rain forest during its history.
2. Environmental setting
2.1. Location
The studied swamp area is called ‘Pantano deMonica’ (0º420S, 72º040W, 160 m elevation) and islocated on the lower terrace of the Rio Caqueta,about 5 km south of the research station Pena Rojaof the Tropenbos-Colombia Foundation, and 25 kmsoutheast of the village Araracuara, in the Provinceof Amazonas in the central Colombian Amazon re-gion (Fig. 1). The Caqueta River catchment is themajor drainage system in the Colombian Amazonas.This ‘white water river’ is draining from the Andesinto the Rio Solimoes in Brazil. Different geomor-phological units are found along a transect from theCaqueta River inland toward Pantano de Monica:repeatedly inundated river flood plains, low riverterraces with well and poorly drained areas, high ter-races of the river, and sedimentary plains of Tertiaryage (Duivenvoorden and Lips, 1993, 1995).
For our study three sites were chosen on the lowriver terraces: Site Pantano de Monica 1 (M1) islocated in a large swamp area, about 1 km long and500 m wide. Site Pantano de Monica 2 is from asmall swamp which covers an area of 75 ð 100 m.This second site is about 1000 m distance from thePantano de Monica 1 site. The third site, Pantanode Monica 3, is a forested area, located between thelarge and small swamps at about 500 m distancefrom both Pantano de Monica 1 and 2 sites.
The annual fluctuations of the water level of theRio Caqueta at Araracuara are about 6 to 9 m (datafrom HIMAT, Duivenvoorden and Lips, 1995). Thestudied sites on the low terraces are about 10 to 15m above the low water level of the Rio Caqueta,

H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 195
Fig. 1. Map showing the geographical location of sites Pantano de Monica 1, 2, and 3 in Colombia Amazonas and other mentioned sitesat the Rio Caqueta river basin. The geomorphological situation of the region is based on the map by Duivenvoorden and Lips (1993).
and neither are inundated during high stands ofthe river. The lower terraces in the Rio Caquetavalley are composed of sandy and gravelly sedimentsand includes some organic-rich levels. Radiocarbondates of organic deposits suggest Middle Pleniglacialages (60,000–27,000 BP) (Van der Hammen et al.,1992a).
2.2. Vegetation
The predominant vegetation in the central partof Colombian Amazonia is dense, tall tropical rain
forest, except for some smaller areas on sandstoneplateaus, e.g. locations west of Araracuara, wheresavanna prevails for edaphic reasons. A detailed veg-etation survey and mapping of the middle Caquetaregion, which includes our study area of Pantanode Monica, have been carried out by Duivenvoor-den and Lips (1993, 1995) and Urrego (1994/1997).For the middle Caqueta area Duivenvoorden andLips distinguished 22 different vegetation types, cor-responding to different substrates that range fromwell drained soils of flood plains (varzea, seasonallyinundated areas), to upland systems (low terraces,

196 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
high terraces and terra firme), to poorly drained soils(such as cut off channels or swamps), and to pod-zolised soils. Taxonomic and ecological studies ofthe Arecaceae (Palmae) and Euphorbiaceae family inthe Araracuara region were undertaken by Galeano(1992) and Aldana and Rosselli (1995), respectively.
The study area (Fig. 1) is located on the lowerterraces of the Rio Caqueta, with both well drainedand poorly drained areas. In the central part of thelarge swamp of site Pantano de Monica 1 densestands of 5–8 m tall trees, primarily Clusia sp.and a few other tree species, prevail. Also a fewpalm trees of Mauritia grow in this swamp. Thisapparently water logged swamp has many patches ofopen water and moss posters. A compact root matis formed primarily by Clusia sp. and a few othersmall trees. This mat probably moves up and downduring low and high water stands, respectively. Thesmall swampy area including the site Pantano deMonica 2 is from a less water logged swamp andis covered by ca. 10 m tall trees, mainly from theFabales families. Site Pantano de Monica 3 is locatedin a small shallow basin between both swamps andcovered by 20 to 30 m tall trees. The palm treeEuterpe is frequent at this site.
2.3. Climate
Climatic conditions recorded in Araracuara, about25 km northeast of Pantano de Monica, shows apermanent humid tropical climate without seasonallydry periods (data from HIMAT; after Duivenvoordenand Lips, 1995). The mean annual temperature is25.7ºC and the annual rainfall is 3060 mm. The pe-riod between December and February is slightlydrier. Climate diagrams from central ColombianAmazonia (Juarete, Taracua and Uaupes) indicatesimilar climatic conditions with high annual precip-itation rates and no dry season (Walter and Lieth,1967).
3. Methods and material
3.1. Sampling and pollen analysis
The three cores were hand-drilled with a 25-mmdiameter Dachnowsky sampler. Core Pantano de
Monica 1, from the central part of the large swamparea, represents the interval from 182 to 271 cm be-low the surface. The upper 182 cm contained mainlyroots, very soft and fine detritus mud, and water, andcould not be recovered. Core Pantano de Monica 2,from the small swamp, includes the uppermost 85 cmof the swamp. Core Pantano de Monica 3, from theforested area, starts at the surface and is 90 cm long.
The cores were recovered in 25 cm long sections,wrapped in plastic foil and protected during trans-portation by PVC pipe. Several modern pollen rainsamples, from moss polsters and organic-rich sur-face samples, were collected within approximately10 m distance of the bore holes and stored in plasticbags. In the laboratory all samples were stored underdark and cold (4ºC) conditions. For time control 1cm-thick bulk sediment samples were taken from thecores for Accelerator Mass Spectrometry (AMS) ra-diocarbon dating at Utrecht University (Van der Borget al., 1987). For pollen analysis, samples of 0.5 cm3
were taken at short intervals along the profile, mostlyat 2–5 cm distance.
Samples were prepared for pollen analysis us-ing the standard treatment of sodium pyrophosphate,acetolysis mixture, and heavy liquid separation bybromoform (Faegri and Iversen, 1989). Pollen prepa-ration included addition of tablets with exotic Ly-copodium spores to calculate the pollen concentra-tion (grains=cm�3) and pollen influx (grains cm�2
yr�1). Pollen counts averaged a minimum of 300pollen grains, excluding pollen of aquatics, andspores of ferns, mosses and fungi. Light grey clay de-posits below 234 cm, 66 cm, and 58 cm core depth atsites Pantano de Monica 1, 2 and 3, respectively, didnot contain sufficient pollen grains for a statisticalcount, or were poorly preserved. For pollen identifi-cation we used the reference collection of the Hugode Vries Laboratory as well as pollen morphologicaldescriptions and illustrations published by Behling(1993), Herrera and Urrego (1996), Hooghiemstra(1984), and Roubik and Moreno (1991). The soft-ware TILIA and TILIAGRAPH was used to plot thepollen diagrams and CONISS for cluster analysis ofthe pollen spectra (Grimm, 1987). The pollen per-centage diagrams from the surface samples and thecores represent the most frequent individual pollenand spore taxa. The summary diagrams group thetaxa according to ecological considerations and also

H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 197
give information on pollen counted per sample,pollen concentration and pollen influx, as well asthe cluster analysis dendrogram.
3.2. Stratigraphy
3.2.1. Pantano de Monica 1 (large swamp forest)Not recovered:
0– 20 cm water or mosses20– 60 cm root mat with some plant remains60–¾130 cm water and=or very soft mud
¾130–¾160 cm root and wood deposits¾160– 182 cm very soft organic material
Recovered:182–187 cm brown, decomposed soft organic mate-
rial with plant remains (fragments ofwood, leaves and roots)
187–197 cm brown-grey, decomposed organic ma-terial, somewhat clayey, some plant re-mains
197–215 cm like 182–187 cm215–235 cm grey, compact clay with some plant
remains235–243 cm light grey, compact clay with less plant
remains243–271 cm light grey, very compact sandy clay
with a few roots271– sandy subsurface
3.2.2. Pantano de Monica 2 (small swamp forest)0–22 cm dark brown, decomposed soft organic
material with plant remains (fragmentsof wood, leaves and roots)
22–33 cm brown clayey medium compact or-ganic material, very fine plant remains
33–46 cm light brown clay, compact, somewhatorganic material, more or less no plantremains
46–63 cm grey clay, very compact63–69 cm light brown clay, compact, somewhat
organic69–85 cm light grey clay
3.2.3. Pantano de Monica 3 (forested swamp)0–14 cm dark brown, decomposed soft organic
material with plant remains (fragmentsof wood, leaves and roots)
14–21 cm transition to
21–35 cm brown clayey medium compact organicmaterial, very small plant remains
35–57 cm light brown clay, compact, somewhat or-ganic material, plant remains rare
57–67 cm grey clay very compact67–90 cm light grey clay
4. Results
4.1. Chronology and pollen zones
AMS radiocarbon ages are shown in Table 1. Inevaluating the ages of the organic rich sediments ofPantano de Monica one has to consider that moderncarbon, in form of roots and rootlets, reach deep intothe deposits. Roots and rootlets, probably of Clusiasp., were still observed in the lowermost part ofcore Pantano de Monica 1. The recovered deposits atsites Pantano de Monica 2 and 3 are not very deepand roots from modern vegetation can reach to thebottom of these cores. Carbon originating from thedecomposition of modern rootlets may cause errorsin the radiocarbon ages.
The 4 AMS radiocarbon dates of core Pantanode Monica 1 indicate the deposits are of early andmiddle Holocene age. An age of 8363 š 49 BP at197 cm core depth, and 9720š70 BP at 215 cm coredepth suggests that the age of 7260 š 60 BP at 235cm depth is too young. Excluding the date of 235 cmdepth from the age calculations, the extrapolated ageat 234 cm is 11,150 BP. The top of the core, at 182cm depth, is calculated 4730 BP.
The 3 AMS radiocarbon dates of core Pantano deMonica 2 indicate the deposits are of late Holoceneage. Comparing the age of 4010 š 50 BP at 69cm depth, and 3710 š 50 BP at 33 cm depth, theage of 2900 š 50 BP at 46 cm depth is probablytoo young, or the core is slightly disturbed in thisinterval. Therefore, the date of 2900 š 50 BP hasbeen excluded from the age calculations. The pollenrecord starts at 66 cm depth, with an interpolatedage of 3980 BP. The upper 32 cm of the coreaccumulated during the last 3700 years, but it cannotbe excluded that deposits are missing. The pollenrecord of core Pantano de Monica 3 starts at 58 cmdepth; 3 AMS radiocarbon dates indicate that thesedeposits represent the last 3260 years.

198 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
Table 1List of AMS radiocarbon dates of samples of the Pantano de Monica cores
Lab. number Depth (cm) 14C yr B.P. a 13C=12C r. Calibrated age (cal BC)
Pantano de Monica 1UtC-4943 187 4855š 38 �30.6 3629–3633UtC-4944 197 8363š 49 �30.6 7484–7451, 7449–7414UtC-4945 215 9720š 70 �29.8 9040–8940UtC-4946 235 7260š 60 �30.0 6165–6132, 6129–6000
Pantano de Monica 2UtC-4940 33 3710š 50 �30.8 2180–2165, 2143–2025, 1998–1984UtC-4941 46 2900š 50 �31.2 1153–1148, 1130–998UtC-4942 69 4010š 50 �30.4 2577–2463
Pantano de Monica 3UtC-4947 21 183š 33 �29.2 cal AD 1667–1686, 1738–1789, 1791–1810, 1929–1952UtC-4948 34 1766š 45 �30.3 cal AD 233–341, 366–371UtC-4949 57 3200 š 70 �31.1 2116–2086, 2040–1941
a 13C-adjusted.
Based on the downcore changes in pollen assem-blages, which are illustrated by the dendrogram ofcluster analysis, there are 3 pollen zones in recordPantano de Monica 1 (zones M1-I to M1-III), 4pollen zones in record Pantano de Monica 2 (zonesM2-I to M2-IV), and 4 pollen zones in record Pan-tano de Monica 3 (zones M3-I to M3-IV). Coredepths, age calculations and number of pollen sam-ples are shown for each core and zone in Table 2.As the three cores represent different age intervals,
Table 2Pollen zones of Pantano de Monica, including depths, age calcu-lations and number of pollen samples for each pollen zone
Zone Depth Age range No. of(cm) (14C yr B.P.) samples
Pantano de Monica 1Zone M1-I 234–215 11150–9720 5Zone M1-II 215–193 9720–6960 7Zone M1-III 193–182 6960–4730 5
Pantano de Monica 2Zone M2-I 66–52.5 3980–3880 5Zone M2-II 52.5–35.5 3880–3740 6Zone M2-III 35.5–27.5 3740–3080 4Zone M2-IV 27.5–0 3080–?–modern 6
Pantano de Monica 3Zone M3-I 58–45 3260–2450 4Zone M3-II 45–33 2450–1640 6Zone M3-III 33–20 1640–180 5Zone M3-IV 20–0 180–modern 4
there is no overlap between the pollen zones exceptfor the uppermost part of core Pantano de Monica 2(Zone M2-IV) with core Pantano de Monica 3 (ZoneM3-IV to M3-I).
4.2. Modern pollen rain data from Pantano deMonica 1, 2 and 3
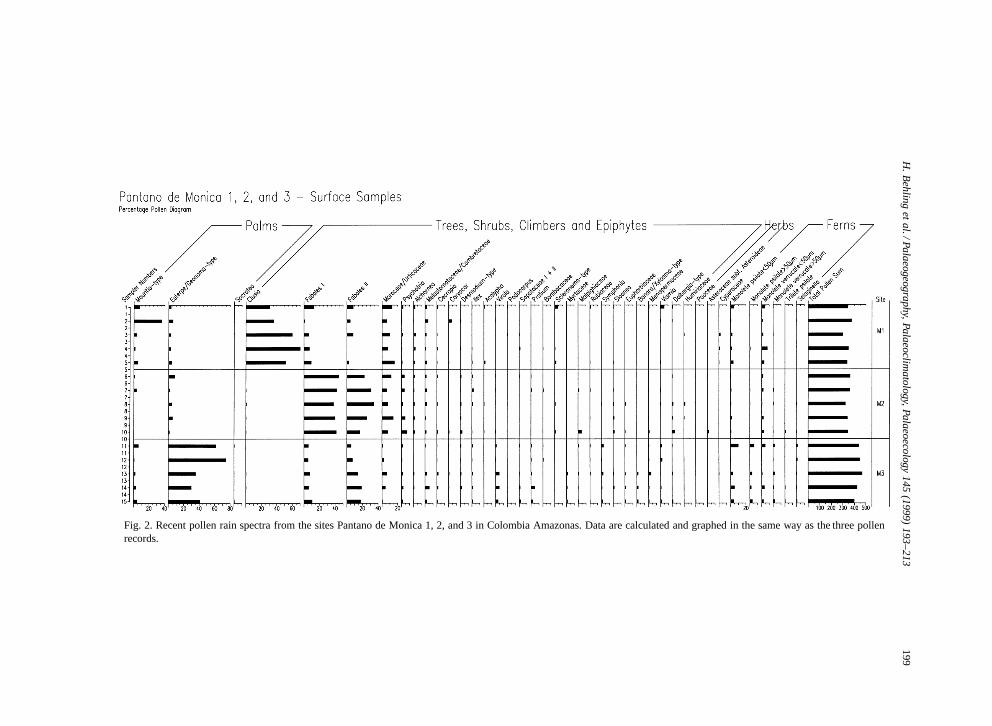
Pollen assemblages in surface samples collected afew meters distance from the three core locations areshown in the percentage diagram (Fig. 2). Samples1–5 are from Pantano de Monica 1 (large swamp),numbers 6–10 are from Pantano de Monica 2 (smallswamp), and numbers 11–15 are from Pantano deMonica 3 (forest).
Samples from site 1 are characterized by highpercentages of Clusia pollen (30–60%), followedby high values of Fabales I and II, Moraceae=Ur-ticaceae, Mauritia-type, Euterpe=Geonoma-type andmany other taxa which occur occasionally with lowpercentages. Pollen grains of herbs and fern sporesare low in numbers.
In site Pantano de Monica 2 modern pollenrain is dominated by Fabales I and II (each 20–40%), followed by Moraceae=Urticaceae, Psycho-tria, Euterpe=Geonoma-type and many other typeswhich occur occasionally with low values.
Modern pollen spectra from site Pantano deMonica 3 show high percentages of Euterpe=Geonoma-type (30–75%), lower percentages of Fa-
H.B
ehlinget
al./Palaeogeography,Palaeoclim
atology,Palaeoecology145
(1999)193–213
199
Fig. 2. Recent pollen rain spectra from the sites Pantano de Monica 1, 2, and 3 in Colombia Amazonas. Data are calculated and graphed in the same way as the three pollenrecords.

200 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
bales I and II, Moraceae=Urticaceae, Virola, Mauri-tia-type, and further a number of taxa which occuroccasionally with low percentages. Fern spores areslightly more frequent than in the surface samples ofsites 1 and 2.
4.3. Description of the pollen diagrams
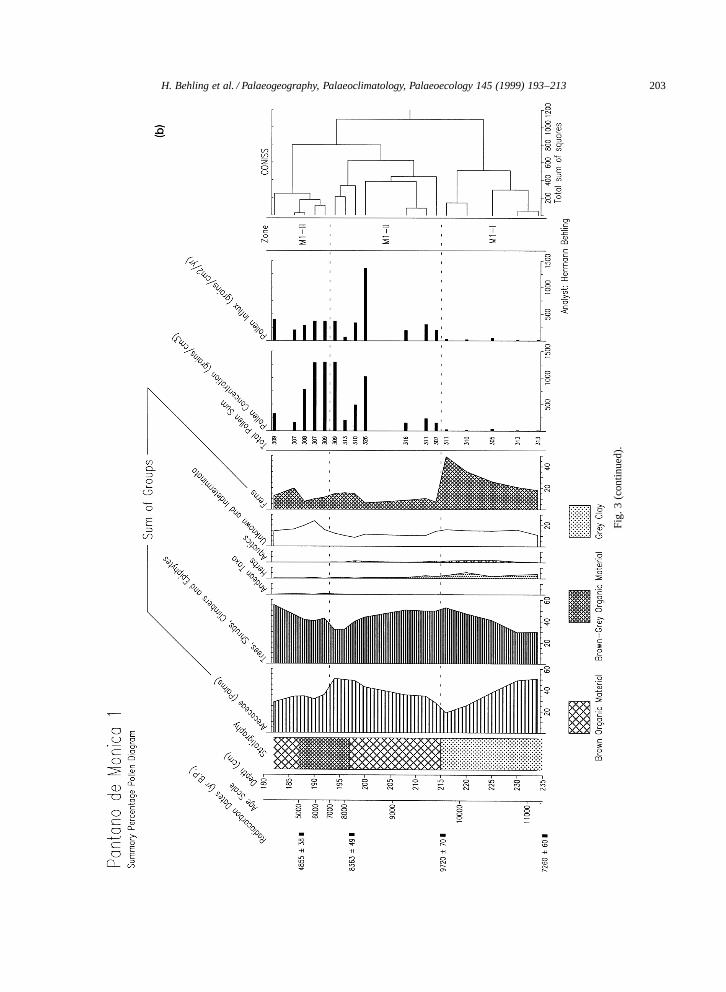
4.3.1. Pollen diagram site Pantano de Monica 1Pollen diagram Pantano de Monica 1 (Fig. 3a,b)
represents the period from 11,150 (extrapolated age)to 4730 BP and shows the 3 pollen zones M1-I,M1-II, and M1-III (Table 2). Fig. 3a illustrates themost important pollen and spore taxa out of about150 different types identified. Pollen concentrationand pollen influx is very low in zone M1-I butmarkedly higher in the upper part of the core.
Characteristic for the diagram is the high rep-resentation of pollen of Mauritia-type (25–50%)throughout the core. Pollen of other palm taxa oc-cur in low numbers, but the Euterpe=Geonoma-typeincreases at the end of zone M1-II, and pollen ofIriartea at the beginning of zone M1-III. Highpercentages of the group including trees, shrubs,climbers and epiphytes (30–55%) also characterizethe three pollen zones. In all zones occur pollenof taxa such as Melastomataceae=Combretaceae, Fa-bales (other), Moraceae=Urticaceae, Sapotaceae, Al-chornea, and many other arboreal taxa with percent-ages below 1% such as Malpighiaceae, Symphonia,Doliocarpus. Certain taxa have their highest repre-sentation in different zones: Posoqueria-type, Rubi-aceae (unknown), and Ilex primarily in the upperpart of zone M1-I; Casearia sylvestris-type, Myr-taceae, Menispermaceae, Podocarpus (maximally2.6%) in the lower part of zone M1-II; Pro-tium, Scleronema, Virola in the upper part of zoneM1-II; Moraceae=Urticaceae, Sapotaceae I–III, Fa-bales (other), Anacardiaceae, and Clusia in zoneM1-III. Some taxa are absent in specific zones:Posoqueria-type and Rubiaceae (unknown) in zonesM1-I and M1-II, and Podocarpus and Caseariasylvestris-type in zone M1-III. Pollen grains ofherbs are represented in trace amounts, except forzone M1-I where Poaceae, Asteraceae and Borre-ria together reach percentages up to 5%. Pollengrains of Andean taxa, such as Myrica and Symplo-cos, are only represented by single grains. Pollen
of aquatic taxa, represented by Cyperaceae, Sagit-taria and Utricularia, are rare. Fern spores occurin moderate percentages, but trilete psilate sporesand spores of Lycopodium curvatum-type are mostfrequent in zone M1-I.
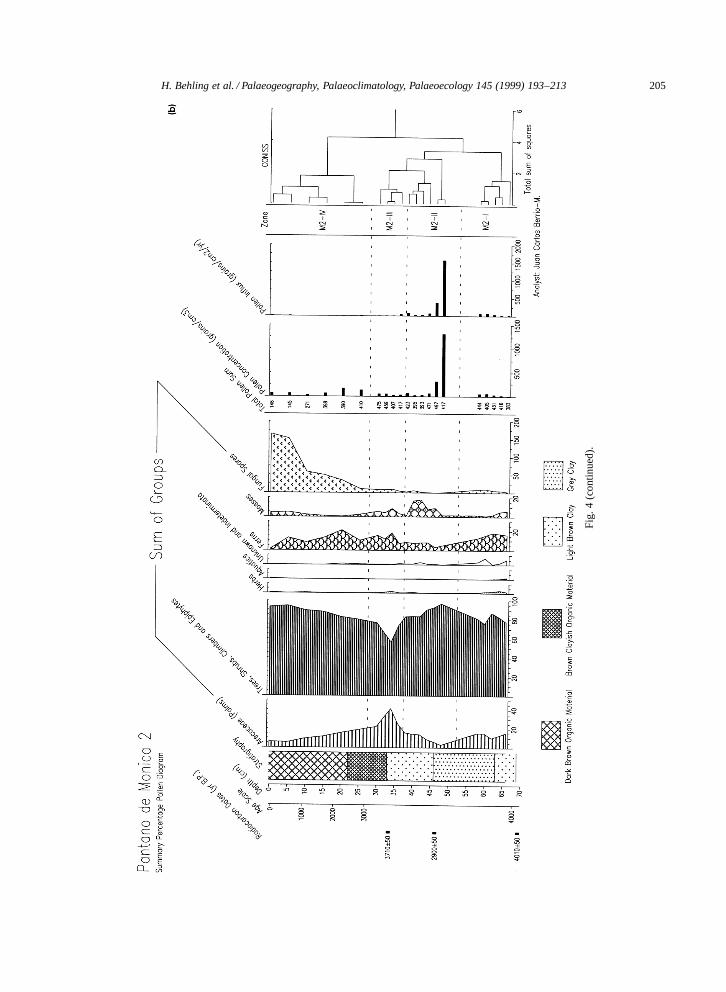
4.3.2. Pollen diagram Pantano de Monica 2Pollen diagram of site Pantano de Monica 2
(Fig. 4a,b) represents the period from 3980 BP topresent-day, but it is possible that there are somegaps in the upper part of the record. The diagramshows four pollen zones, M2-I to M2-IV (Table 2).Fig. 4a shows the most important pollen and sporetaxa. Downcore pollen concentration values are rel-atively constant, except for two samples in zoneM2-II. The pollen influx is very low in the upper partcompared to the lower part of the core.
The pollen record of site Pantano de Mon-ica 2 is marked by high percentages of thegroup of trees, shrubs, climbers and epiphytes(mostly 75–95%), and a lower representationof the group of palms (mostly 5–20%). Pollengrains of Mauritia-type and Euterpe=Geonoma-typeare well represented, whereas Mauritiella-type,Socratea, Astrocarium, and Iriartea occur in lowpercentages only (<1%). One sample in zoneM2-III shows 35% of the Euterpe=Geonoma-type.Pollen grains of other forest taxa are abun-dant including Fabales I and II; or are moder-ately represented including Moraceae=Urticaceae,Melastomataceae=Combretaceae, Psychotria, or rep-resented with low values such as Alchornea, Myr-taceae, Malpighiaceae, Vismia, Clusia, and Virola.Many other taxa only occur as single grains. Inthis second site, forest taxa show strong fluctua-tions: Caryocar and Protium have high percent-ages in zones M2-I and M2-III and low values inzones M2-II and M2-IV. Fabales I and Cecropiashow highest percentages in the lower part of zoneM2-II and Psychotria in the upper part of this zone.Pollen grains of Ilex are more frequent in the lowerpart of zone M2-IV. Moraceae=Urticaceae pollenshow highest frequencies in the upper part of zoneM2-IV. Herb pollen (Poaceae, Asteraceae, Borreria)and pollen grains of aquatic taxa (Cyperaceae) arevery rare in the pollen assemblages. Fern spores arecommon in all zones. In the lower part of the coreSelaginella is less frequent than in the upper part.

H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 201
Percentages of moss spores are higher in zone M2-IIthan in other zones. Fungal spores are rare but theirabundance increases markedly in zone M2-IV.
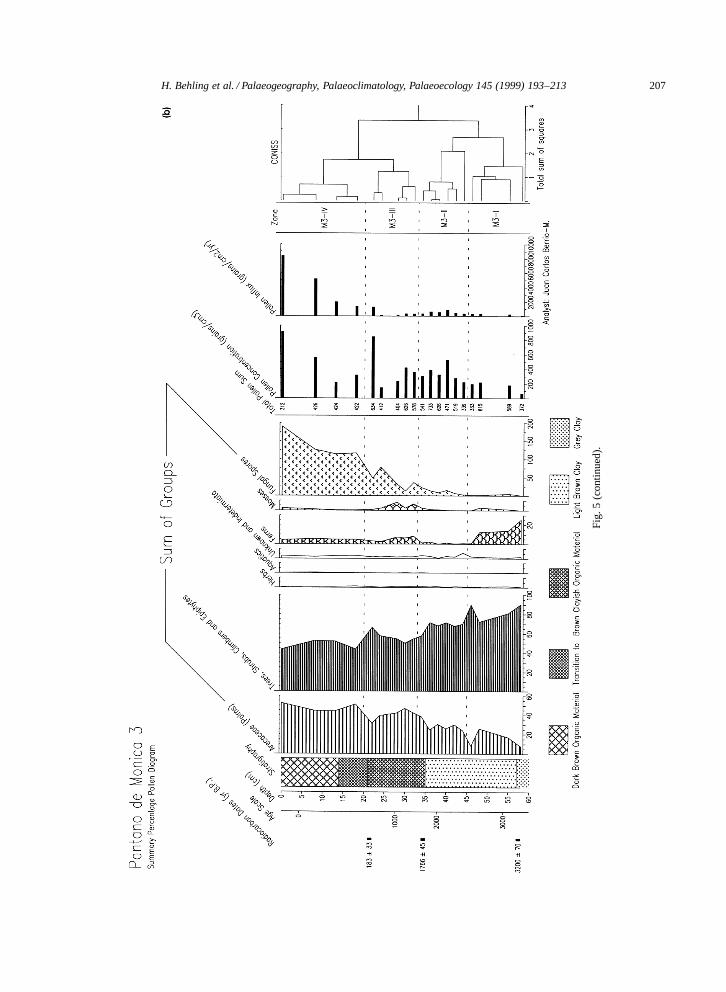
4.3.3. Pollen diagram Pantano de Monica 3The pollen diagram of site Pantano de Monica
3 (Fig. 5a,b) represents the period of the last 3260years and shows four pollen zones M3-I to M3-IV(Table 2). The percentage diagram illustrates themost important pollen and spore taxa. Downcorevalues of pollen concentration and pollen influx arerelatively constant but increase markedly in the upperpart of the core (zone M3-IV).
The pollen record is characterized by agradual increase in palms, mainly pollen ofEuterpe=Geonoma-type which increases from 5% atthe base to 40% at the core-surface. Pollen grains ofMauritia-type and Mauritiella-type are representedin low percentages. Pollen grains of Astrocarium,Socratea and Iriartea are found in trace amounts,except for slightly higher numbers of Astrocar-ium in zones M3-I and M3-II. The contributionof the group of trees, shrubs, climbers, and epi-phytes decreases from 91% at the base to 44% atthe core-surface. Throughout the 4 zones the mostdominant taxa in this group are pollen of Fabales Iand II, followed by Moraceae=Urticaceae, Desmod-ium-type, Protium, Melastomataceae=Combretaceae,Ilex, Menispermaceae, Psychotria, and many othertaxa which occur occasionally with lower per-centages. Some taxa are stronger representedin different zones: Fabales I and II, Malpighi-aceae, Protium, Guarea-type mainly in zone M3-I;Moraceae=Urticaceae, Banara=Xylosma-type, andIlex mainly in zone M3-II; Psychotria, Vismia, Spon-dias-type mainly in zone M3-III; Virola, Apocy-naceae, and Maripa-type mainly in zone M3-IV.Rubiaceae and Cecropia are more frequent in zonesM3-I and M3-II than in the upper two zones. Pollengrains of herbs (Poaceae, Asteraceae) and aquatics(Cyperaceae, Sagittaria) occur only as single grainsin the pollen assemblages. Fern spores are frequentin zone M3-I, very rare in zone M3-II, and morefrequent again in zones M3-III and M3-IV. Mossspores are generally rare but are best represented inzone M3-III. A gradual and significant increase offungal spores is noted from zone M3-II to the top ofthe core.
5. Vegetation development, paleoenvironmentalreconstruction and discussion
The pollen records present tropical rain forest andswamp vegetation on the lower terraces of the RioCaqueta river basin and show vegetational and envi-ronmental change during Late Glacial and Holocenetimes. Opportunities to compare these three sites arelimited because of the different modern vegetationalsettings and the mostly non overlapping time inter-vals (Table 2). The modern pollen rain reflects quiteaccurately the differences in the modern vegetationat the three sites, such as the abundance of Clusia inthe large swamp area of site Pantano de Monica 1,different species of Fabales around the small swampof site Pantano de Monica 2, and the high frequencyof Euterpe in the forest of site Pantano de Monica3.
5.1. Late Glacial to mid-Holocene: site Pantano deMonica 1
Modern pollen rain spectra from site Pantano deMonica 1 (samples 1–5 in Fig. 2) differ markedlyfrom the fossil pollen record during the period of11,500 to 4730 BP (Fig. 3a) which suggests differentvegetational conditions in the swamp. The modernClusia dominated vegetation did not occur duringthat period. This implies that the Clusia dominatedswamp vegetation is locally not older than 4700years. During the recorded period the site was aMauritia-swamp. Many forest taxa are better rep-resented in the pollen record than in the modernpollen rain, which suggests that the rain forest veg-etation was closer to the site than nowadays. Thefloral composition and environmental setting suggestthat the swamp represented in the pollen record wasless water-logged and smaller than today. This indi-cates that the lower terraces in the Rio Caqueta riverbasin were better drained than today, either relatedto a better drainage system or a drier climate. Lowerhigh water levels of the Rio Caqueta as a result ofenhanced seasonality (longer or more intense dry pe-riods), or on the average lower annual rainfall ratesin central Colombian Amazonas, may have occurredduring that time.
Our pollen data do not show any evidence that theearly stage of this swamp was related to a meander

202H
.Behling
etal./Palaeogeography,
Palaeoclimatology,Palaeoecology
145(1999)
193–213Fig. 3. (a) Pollen percentage diagram of core Pantano de Monica 1, Colombia Amazonas. (b) Summary pollen diagram of core Pantano de Monica 1, including pollenconcentration and pollen influx records, and the cluster analysis dendrogram.
H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 203
Fig.
3(c
ontin
ued)
.

204H
.Behling
etal./Palaeogeography,
Palaeoclimatology,Palaeoecology
145(1999)
193–213
Fig. 4. (a) Pollen percentage diagram of core Pantano de Monica 2, Colombia Amazonas. (b) Summary pollen diagram of core Pantano de Monica 2, including pollenconcentration and pollen influx records, and the cluster analysis dendrogram.
H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 205
Fig.
4(c
ontin
ued)
.

206H
.Behling
etal./Palaeogeography,
Palaeoclimatology,Palaeoecology
145(1999)
193–213
Fig. 5. (a) Pollen percentage diagram of core Pantano de Monica 3, Colombia Amazonas. (b) Summary pollen diagram of core Pantano de Monica 3, including pollenconcentration and pollen influx records, and the cluster analysis dendrogram.
H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 207
Fig.
5(c
ontin
ued)
.

208 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
system of Rio Caqueta, or that the site was season-ally inundated. The few recovered pollen grains oftaxa from the mountain forest (Andean forest belt, atpresent occurring from 2300 to 3200 m elevation),such as Weinmannia and Myrica (not shown in thediagrams), were probably transported by wind andnot by the river.
Downcore changes in the pollen assemblages ofcore Pantano de Monica 1 reflect changes in the flo-ral composition of the rain forest. The genus Poso-queria, mid-canopy trees of moist forests (Gentry,1993), and an unknown Rubiaceae-type were onlyfound during the Late Glacial (zone M1-I). Ilex wasmore frequent in the Late Glacial rain forest, butthe palaeoecological significance of this tree taxonis still unclear. The slightly higher presence of herbsin zone M1-I (Borreria is indicative of open forestareas, Croat, 1978; Cleef et al., 1993), may suggestpresence of areas with an open canopy at that timenear this site.
During the period represented in the lower part ofzone M1-II, the pollen type of Casearia sylvestris,possibly reflecting the tree Casearia sylvestris itself,was abundant in the forest. Casearia sylvestris is a2–10 m tall shrub or small tree especially frequent inyoung forests (Croat, 1978). Scleronema (large treesof the Bombacaceae family; Gentry, 1993), Protium(frequent on well drained upland sites; Duivenvoor-den and Lips, 1995) and Virola (especially frequentin older forests and well drained upland sites; Duiv-envoorden and Lips, 1995; Gentry, 1993) were com-mon in the rain forest during the early Holocene(upper part of zone M1-II). These vegetationalchanges at the transition from zone M1-I to M1-II(which might reflect the Late Glacial=Holocenetransition), and also during the period of zoneM1-II (ca. 9700 to 8300 BP), suggest successionalstages of the rain forest composition. These changesmay be caused by climatic change as well as bychanges in the drainage system of this area. Thechanges in floral composition documented at thetransition from zone M1-II to M1-III (increasedpresence of the palms Euterpe=Geonoma-type, Iri-artea, and Moraceae=Urticaceae, Sapotaceae, Anac-ardiaceae and Clusia) are difficult to interpret.
5.1.1. Podocarpus pollen grains in the record ofPantano de Monica 1
The presence of pollen grains of the coniferPodocarpus in low amounts (<1%) in zone M1-Iand up to 2.6% in the lower part of zone M1-IIneeds some discussion. The calculated pollen influxfor 8 grains in the sample at 212 cm depth (D2.6% of the total pollen sum) is 78 grains cm�2
yr�1. Podocarpus pollen can be transported fromtheir main distribution region in the upper mountainforest of the Andes (Torres-Romero, 1988) by windover long distances, but the number of transportedgrains is very low. For instance pollen samples of thesite Laguna Angel in the Llanos Orientales, 600 kmnorth from Pantano de Monica, contain only a fewgrains (not more than 1 grain in each fifth sample)during the Holocene (Behling and Hooghiemstra,1998). The calculated pollen influx in the 5 sampleswhich contained Podocarpus is only between 0.1 and3.7 grains cm�2 yr�1 (average is 2.2). Laguna Angelis about 300 km distant from the Andes and Pantanode Monica about 500 km. During the late Holocenepart of our records (Pantano de Monica 2 and 3) noPodocarpus grains have been found; only one grainwas found in the surface samples (sample no. 3).For comparison, modern pollen rain studies in theAtlantic lowland rain forest in southern Brazil, froma one hectare plot on which four Podocarpus treesoccurred, registered an influx value of 102 grainscm�2 yr�1 (Behling et al., 1997).
All these data and comparisons support that inthe past Podocarpus may have occurred in the studyregion. Podocarpus trees may have rarely grownon the slopes of the higher terraces at that time atsome 100 m distance from the site. The occurrenceof Podocarpus in the Amazonian lowlands duringglacial times has also been interpreted as evidencefor cooling, e.g. in core Morro do Seis Lagos (nearPico da Neblina) and Lagoa da Curuca (near theAmazon mouth) in Brazil (Colinvaux et al., 1996;Behling, 1996).
5.2. Mid-Holocene to recent time: sites Pantano deMonica 2 and 3
The pollen assemblages of the uppermost parts ofcores Pantano de Monica 2 and 3 are quite similarto the assemblages of the modern pollen rain at

H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 209
these two sites. This indicates that modern conditionsprevailed during deposition in the top parts of bothcores. The radiocarbon dates of core Pantano deMonica 2 (Table 1) probably point to sedimentaryhiatuses in the upper part of the core, or the presenceof disturbances in this record.
The pollen record from the small swamp Pan-tano de Monica 2 starts at 4000 BP and reflectschanges in the floral composition of the rain forest.The marked presence of both Protium and Caryocartrees, which grow primarily in well drained uplandsites (Duivenvoorden and Lips, 1995), point to welldrained conditions in the area at the beginning of therecord. The site experienced vegetational disturbanceat the transition from zone M2-I and M2-II, around3000 BP, as indicated by the increased presence ofthe coloning taxon Cecropia. The high presence ofPsychotria, represented by shrubs and small trees ofespecially primary forest (Duivenvoorden and Lips,1995), suggests a quick recovery of the forest (per-haps within 50 years, as far as the radiocarbon datesmay allow this calculation). The rare presence ofthe taxa Protium and Caryocar, may reflect poordrainage of the lower terrace area where the smallswamp developed, probably within the last 3000years (zone M2-IV). A definite age cannot be calcu-lated for the first development because of possiblesedimentary gaps during that time.
The record of site Pantano de Monica 3, from thewet forest area in between both swamps (Fig. 1),probably completely covers the last 3260 years. Thecontinuously increasing presence of palms, probablyEuterpe, in the rain forest at this site is conspic-uous. Euterpe can grow under different conditions,including alluvial flood plains (Galeano, 1992). TheEuterpe record may be interpreted as a change dur-ing the late Holocene from well drained conditionsto poorly drained conditions on the lower terrace.Stronger human impact in this region may alsobe an explanation for the increase of palm trees.Abundance of different palm taxa during the lateHolocene, related to human activity and=or wetterclimatic conditions, is known from other neotropi-cal sites (Balee, 1988; Behling and da Costa, 1997;Behling and Hooghiemstra, 1998).
6. Comparison to other records in northwesternSouth America
6.1. Central Colombian Amazon (middle Caquetariver area)
Two cores from Mariname Island (ca. 0º440S,72º040W), located in the Caqueta River bed (5–6km south of Pantano de Monica) and three coresfrom Quinche area (ca. 0º530S, 71º510W) in theCaqueta River valley (30 km southeast of Pantanode Monica) have been studied by Urrego (1991,1994/1997). These flood plain sites are frequentlyinundated which contrasts the Pantano de Monicasites on lower terraces which are not inundated. Themodern vegetation at these five sites consist of Mau-ritia swamps, which is different from the Pantanode Monica sites. Two cores from site Mariname rep-resent the last ca. 10,000 years (extrapolated), andthe three cores from Quinche contain deposits fromthe early Holocene (ca. 10,000–8000 BP) and fromthe late Holocene (ca. 4000 BP (extrapolated) tothe present). Sedimentation rates and pollen changeshave been interpreted as follows. Between ca. 10,000and 9000 BP open water conditions prevailed andthe Caqueta River valley was permanently inun-dated with rapid sedimentation of clay, suggestingrelatively wet climatic conditions. Around 9000 BPlower water levels with a decreased sedimentationrate are probably related to a drier climate. Cecropiaforest developed on the open flood plains. This pe-riod is followed by seasonally inundated varzea for-est until 6500 on one site and until 4500 BP on asecond site on Mariname Island growing under rela-tively dry conditions. After 4500 BP the sites on theisland became less influenced by direct river flood-ing and a Mauritia swamp developed. The recordsfrom Quinche show a similar succession during thelate Holocene: here Mauritia swamps are only foundduring the last 1000 or 700 years. Urrego concludedthat the documented river dynamics in the study areais at least partly influenced by climatic change.
Several Late Quaternary sediment sequences inthe middle Rio Caqueta valley have been studiedby Van der Hammen et al. (1992a,b). They con-cluded that the depositional and erosional history isrelated to climate change. Gaps in the deposits oforganic material during the Upper Pleniglacial (from

210 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
ca. 27,000 to 13,000 BP) are interpreted as relatedto a dry climatic period. Sediment deposition startsat the beginning of the Late Glacial (ca. 13,000 BP),suggesting a wetter climate. Sandy deposits at thebase of the Holocene suggest open water conditions.Later the sediment characteristics suggest seasonalinundations. Finally, most of the early Holocene de-posits may have been removed by river erosion. Themajor part of the deposits are younger than ca. 3500years. Van der Hammen et al. (1992b) concludedthat deposition of more organic rich sediments in theRio Caqueta area during the early Holocene was theresult of a reduced river discharge, suggesting drierconditions during the intervals of ca. 3800–3200 BPand 2700–1900 BP.
In general, lower water levels in the Caqueta Riverduring the early Holocene, fit quite well the paleoen-vironmental conclusions from Pantano de Monica 1.A drier climate, corresponding to well drained con-ditions on the lower terraces from 3800–3200 BP,is supported by the record of Pantano de Monica 2,although the dry period of 2700–1900 BP was notfound.
6.2. Llanos Orientales in Colombia
Evidence of drier climatic conditions during fullglacial times and during the early Holocene andwetter climates during the Late Glacial–Holocenetransition and the late Holocene periods, come fromthe studies in the Colombian savanna region of theLlanos Orientales (Behling and Hooghiemstra, 1998and Behling and Hooghiemstra, in press) about 600km north of Pantano de Monica. Changes to wetterconditions were observed at several sites starting atabout 5000 BP and about 3800 BP. These resultssupport the interpretation of the records from Pan-tano de Monica that changes in the middle Caquetariver area are primarily forced by climate.
6.3. Ecuador
Most of the pollen records from western Ama-zonia are from Ecuadorian lakes along the foothillsof the eastern Andes (500–700 km west to south-west of Pantano de Monica) and represent the lateHolocene. Only the road cut sites Mera (1º290S,77º060W, 1100 m elevation, ca. 31,000–ca. 34,000
BP) and San Juan Bosco (3º030S, 78º270W, 970 melevation, 26,000 – >31,000 BP) represent recordsof Middle Pleniglacial age (Bush et al., 1990). Thepollen records of these two sites show a significantpresence of mountain taxa (e.g. Alnus, Podocarpus),suggesting a cooling of ca. 7.5ºC below present.
The 7000 BP pollen record from Lake Ayauchi(2º50S, 78º10W, at ca. 500 m elevation) reflectsHolocene Amazon rain forest history. The intervalbetween from ca. 4200–3150 BP has been inter-preted as a dry period.
The record of site Lago Kumpaka (3º020S,77º490W, 700 m elevation) represents the environ-mental history of a perturbed rain forest since the last5200 years. The periods of 4300–3800 BP and 1500–800 BP have been interpreted as drier with more sea-sonal precipitation (Liu and Colinvaux, 1988).
The record of lake Anangucocha (0º320S,76º270W, 300 elevation) represents the last 3100years (Frost, 1988). Three distinct intervals couldbe recognized: a Mauritia palm swamp (3100–1300BP), a regional flooding with the destruction of theMauritia swamp (1300–800 BP), and a lake period(800 BP to the present). These changes have beeninterpreted as climatic fluctuations from a dry periodto wetter climatic conditions.
Three lakes at about 300 m elevation have beenstudied by Colinvaux et al. (1988a): Limonococha(0º240S, 76º380W), Lago Agrio (0º070N, 76º550W),and Lago Santa Cecilia (0º040N, 77º010W). Sedimentrecords, which extend back as far as 800 and 1230years, show flood deposits between 1300–800 BPoverlain by lake deposits of the last 800 years.Pollen assemblages indicate disturbance during therecorded periods.
7. Conclusions
In summary, the records from the sites Pantanode Monica 1, 2 and 3 show that the swamp and rainforest environments were not stable during the LateGlacial and Holocene. This is also documented in thefew other Amazonian lowland pollen records. TheLate Glacial occurrence of Podocarpus in the middleRio Caqueta region is evidence for different floralcomposition of the rain forest compared to today.Presence of this conifer suggests a marked cooling in

H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 211
the tropical lowlands, as reported from other pollensites and may also indicate that zone M1-I is stilla part of the Late Glacial. Several studies suggesta cooling between 5 and 7ºC in the neotropicallowlands during glacial times (e.g. Bush et al., 1990;Stute et al., 1995; Colinvaux et al., 1996; Behlingand Lichte, 1997) and cooling is also accepted byadvocates of the forest refugia hypothesis (e.g. Vander Hammen and Absy, 1994).
In general, during the Late Glacial and earlyHolocene the lower terrace is better drained (Pan-tano de Monica 1), than during the interval between4000 and 3080 BP (Pantano de Monica 2), and dur-ing the last 3000 years (Pantano de Monica 2 and3). Observed changes are a result of changes in thedrainage system of the lower terraces in the Caquetariver basin. This might be a consequence of riverdynamics in the valley of the Rio Caqueta, as wellas of climate change (less precipitation and reducedriver discharge). These three sites provide insuffi-cient evidence to evaluate whether climatic changein the central Colombian Amazon was the majorforcing factor for the observed dynamics. Differentsettings of the few sites in the northwestern low-lands of South America, from which we know thepaleoenvironmental and the climatic history, makean evaluation of data difficult. Most of the studiedsites are located near rivers and may be influencedby river dynamics, such as changes in the meanderpattern or strong regional flooding. Such events mayalso cause local environmental change irrespectiveof climatic change.
Comparison with other records which suggest thatthe lower water levels of the Rio Caqueta during earlyHolocene period are the result of drier climatic con-ditions, may support the interpretation that the welldrained lower terraces of Pantano de Monica are alsothe result of drier climates. Also other paleoenviron-mental studies of the Llanos Orientales (Behling andHooghiemstra, 1998, in press) may suggest that cli-matic change is a major factor to explain environmen-tal change in the middle Caqueta River area.
Acknowledgements
The authors thank the people from the village ofPena Roja, Arthur van Dulmen, and Jeroen Groe-
nendyk for assistance during the fieldwork. EllyBeglinger and Annemarie Phillip (Amsterdam) arethanked for preparing the pollen samples. The for-mer director of Tropenbos-Colombia, Juan Saldar-riaga, is thanked for his hospitality and continuouslogistic support during field expeditions. Klaas vander Borg (Utrecht) is acknowledged for the radiocar-bon dates. Joost Duivenvoorden and Antoine Cleef(Amsterdam) provided valuable information to im-prove the interpretation of the data. We thank thereviewers Vera Markgraf and Alan Graham for con-structive and valuable comments on the manuscript.The Netherlands Organization for Scientific Re-search (NWO, branch GOA) is acknowledged forfunding of the current postdoc project (project num-ber 750.195.10 to H. Hooghiemstra).
References
Aldana, J.M., Rosselli, P.F., 1995. Las euforbıaceas de la regionde Araracuara. Euphorbiaceae in the Araracuara Region. Es-tudios en la Amazonia colombiana=Studies of the ColombianAmazonia, Vol. IX. Tropenbos Colombia, Bogota, 191 pp.
Balee, W., 1988. Indigenous adaptation to Amazonian palmforests. Principes 32, 47–54.
Behling, H., 1993. Untersuchungen zur spatpleistozanen undholozanen Vegetations- und Klimageschichte der tropischenKustenwalder und der Araukarienwalder in Santa Cata-rina (Sudbrasilien). Dissertationes Botanicae, 206, J. Cramer,Berlin Stuttgart, 149 pp.
Behling, H., 1996. First report on new evidence for the oc-currence of Podocarpus and possible human presence at themouth of the Amazon during the Late-glacial. Veget. Hist.Archaeobot. 5, 241–246.
Behling, H., da Costa, M.L., 1997. Studies on Holocene tropicalvegetation, mangrove and coast environments in the state ofMaranhao, NE Brazil. Quat. S. Am. Antarct. Peninsula 10,93–118.
Behling, H., Hooghiemstra, H., 1998. Late Quaternary paleoecol-ogy and paleoclimatology from pollen records of the savannasof the Llanos Orientales in Colombia. Palaeogeogr. Palaeocli-matol. Palaeoecol. 139, 251–267.
Behling, H., Hooghiemstra, H., in press. Environmental historyof the Colombian savannas of the Llanos Orientales sincethe Last Glacial Maximum form lake records El Pinal andCarimagua. J. Paleolimnol.
Behling, H., Lichte, M., 1997. Evidence of dry and cold climaticconditions at glacial times in tropical Southeastern Brazil.Quat. Res. 48, 348–358.
Behling, H., Negrelle, R.R.B., Colinvaux, P.A., 1997. Modernpollen rain data from the tropical Atlantic rain forest, ReservaVolta Velha, South Brazil. Rev. Palaeobot. Palynol. 97, 287–299.

212 H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213
Bush, M.B., Colinvaux, P.A., 1988. A 7000 year vegetationalhistory form lowland Amazon, Ecuador. Vegetatio 76, 141–154.
Bush, M.B., Piperno, D.R., Colinvaux, P.A., 1989. A 6000 yearhistory of Amazonian maize cultivation. Nature 340, 302–303.
Bush, M.B., Colinvaux, P.A., Wiemann, M.C., Piperno, D.R.,Liu, K.-b., 1990. Late Pleistocene temperature depression andvegetation change in Ecuadorian Amazonia. Quat. Res. 34,330–345.
Cleef, A.M., Van der Hammen, T., Hooghiemstra, H., 1993. Thesavanna relationship in the Andean paramo flora. Op. Bot.121, 285–290.
Colinvaux, P.A., 1993. Pleistocene biogeography and diversityin tropical forests of South America. In: Goldblatt, P. (Ed.),Biological Relationships Between Africa and South America.Yale University Press, New Haven, pp. 473–499.
Colinvaux, P.A., 1996. Quaternary environmental history and for-est diversity in the neotropics. In: Jackson, J.B.C., Budd, A.F.,Coates, A.G. (Eds.), Evolution and Environment in TropicalAmerica. The University of Chicago Press, Chicago, pp. 359–405.
Colinvaux, P.A., Frost, M., Frost, L., Liu, K.-b., Steinnitz-Kan-nan, M., 1988a. Three pollen diagrams of forest disturbance inthe western Amazon. Rev. Palaeobot. Palynol. 55, 73–82.
Colinvaux, P.A., Olson, K., Liu, K.-b., 1988b. Late-glacial andHolocene pollen diagrams from two endorheic lakes of theinter-Andean plateau of Ecuador. Rev. Palaeobot. Palynol. 55,73–81.
Colinvaux, P.A., De Oliveira, P.E., Moreno, J.E., Miller, M.C.,Bush, M.B., 1996. A long pollen record from lowland Ama-zonia: forest and cooling in glacial times. Science 274, 85–87.
Croat, T.C., 1978. Flora of Barro Colorado Island. StandfordUniversity Press, Palo Alto, CA, 943 pp.
Duivenvoorden, J.F., Lips, J.M., 1993. Mapa de la ecologıa delpaisaje del medio Caqueta. Memoria explicativa. Landscapeecological map of the middle Caqueta Basin. ExplanatoryNotes. Estudios en la Amazonia colombiana=Studies on theColombian Amazonia, Vol. III. Tropenbos Colombia, Bogota,301 pp.
Duivenvoorden, J.F., Lips, J.M., 1995. A Land-ecological Studyof Soils, Vegetation, and Plant Diversity in Colombian Ama-zonia. Tropenbos Series 12, Wageningen, 438 pp.
Faegri, K., Iversen, J., 1989. Textbook of Pollen Analysis, 4thed. Wiley, Chichester, 328 pp.
Frost, I., 1988. A Holocene sedimentary record from Anangu-cocha in the Ecuadorian Amazon. Ecology 69, 66–73.
Galeano, G., 1992. Las palmas de la region de Araracuara.The Palms in the Araracuara Region. Estudios en la Amazo-nia colombiana=Studies on the Colombian Amazonia, Vol. I.Tropenbos Colombia, Bogota, 180 pp.
Gentry, A.H., 1993. Woody Plants of Northwest South America.Conservation International, Washington, DC, 895 pp.
Grimm, E.C., 1987. CONISS: a Fortran 77 program for strati-graphically constrained cluster analysis by the method of theincremental sum of squares. Computers Geosci. 13, 13–35.
Haffer, J., 1969. Speciation in Amazonian forest birds. Science165, 131–137.
Herrera, L.F., Urrego, L.E., 1996. Atlas de polen de plantasutiles y cultivadas de la Amazonia colombiana. Pollen atlasof useful and cultivated plants in the Colombian Amazonregion. Estudios en la Amazonia colombiana=Studies on theColombian Amazonia, Vol. XI. Tropenbos Colombia, Bogota,462 pp.
Hooghiemstra, H., 1984. Vegetational and climatic history of thehigh plain of Bogota, Colombia: a continuous record of thelast 3.5 million years. Dissertationes Botanicae 79. J. Cramer,Vaduz, 368 pp.
Hoorn, C., 1994a. Fluvian palaeoenvironments in the intracra-tonic Amazonas Basin (early Miocene–early Middle MioceneColombia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 109, 1–54.
Hoorn, C., 1994b. Miocene palynostratigraphy and paleoenvi-ronments of northwestern Amazonia: evidence for marine in-cursions and the influence of Andean tectonics. Ph.D. thesis,University of Amsterdam, 156 pp.
Liu, K.-b., Colinvaux, P.A., 1985. Forest changes in the Amazonbasin during the last glacial maximum. Nature 318, 556–557.
Liu, K.-b., Colinvaux, P.A., 1988. A 5200-year history of Ama-zon rain forest. J. Biogeogr. 15, 231–248.
Lyons-Weiler, J.F., 1992. Palynological evidence for regional,synchronous, Holocene climate changes in Amazonia and theirinfluence on evolution in the tropics. Thesis at Ohio StateUniversity, Columbus, OH, USA. Rev. Palaeobot. Palynol., inpress.
Roubik, D.W., Moreno, J.E., 1991. Pollen and Spores of BarroColorado Island. Missouri Botanical Garden 36, 270 pp.
Stute, M., Forster, M., Frischkorn, H., Serejo, A., Clark, J.F.,Schlosser, P., Broecker, W.S., Bonani, G., 1995. Cooling oftropical Brazil (5ºC) during the last glacial maximum. Science269, 379–383.
Torres-Romero, J.H., 1988. In: Pinto, P., Lozano, G. (Eds.), Florade Colombia, 5. Podocarpaceae. Editoriales de la ImprentaNational, Bogota, Colombia, pp. 1–73.
Urrego, L.E., 1991. Sucesion Holocenica de un bosque de Mau-ritia flexuosa L.f. en el valle del Rıo Caqueta (AmazoniaColombiana). Colombia Amazonica 5 (2), 99–118.
Urrego, L.E., 1994=1997. Los bosques inundables del medioCaqueta: caracterizacion y sucesion. Ph.D. thesis, Univer-sity of Amsterdam. Published as: Estudios en la Amazo-nia Colombiana=Studies on the Colombian Amazonia, XIV.Tropenbos Colombia, Bogota, 335 pp.
Van der Borg, K., Alderliestern, A., Harnton, C.M., De Jong,A.F., Van Zwol, N.A., 1987. Accelerator mass spectrometrywith 14C and 10B in Utrecht. Nucl. Instr. Methods B 29, 143–145.
Van der Hammen, T., 1998. Paleoecology of Amazonia. Proceed-ings Symposium, Belem, in press.
Van der Hammen, T., Cleef, A.M., 1992. Holocene changes ofrainfall and river discharge in Northern South America and theEl Nino phenomenon. Erdkunde 46, 252–256.
Van der Hammen, T., Absy, M.L., 1994. Amazonia during

H. Behling et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 145 (1999) 193–213 213
the last glacial. Palaeogeogr. Palaeoclimatol. Palaeoecol. 109,247–261.
Van der Hammen, T., Duivenvoorden, J.F., Lips, J.M., Urrego,L.E., Espejo, N., 1992a. The Late Quaternary of the MiddleCaqueta River area (Colombian Amazonia). J. Quat. Sci. 7,45–55.
Van der Hammen, T., Urrego, L.E., Espejo, N., Duivenvoorden,J.F., Lips, J.M., 1992b. Late-glacial and Holocene sedimenta-
tion and fluctuations of river water level in the Caqueta area(Colombian Amazonia). J. Quat. Sci. 7, 57–67.
Walter, H., Lieth, H., 1967. Klimadiagramm-Weltatlas. GustavFischer, Jena.
Whitemore, T.C., Prance, G.T., 1987. Biogeography and Qua-ternary History in Tropical America. Oxford University Press,Oxford, 214 pp.
Copyright © 2022 FDOKUMEN




















