
Late Pliocene Faunal Turnover in the Turkana Basin, Kenya and Ethiopia
Upload
independentCategory
view
3download
0
ELSEVIER Palaeogeography, Palaeoclimatology, Palaeoecology 134 (1997) 109 125
PALAEO
Late Pliocene climatic change and the global extinction of the discoasters: an independent assessment using oxygen isotope
records Mark R. Chapman a,,, Alex J. Chepstow-Lusty b
a Godwin Laboratory, Department ofiEarth Sciences, University o f Cambridge, Free Sehool Lane, Cambridge, CB2 3RS, UK
b Department o f Plant Sciences, University o f Cambridge, Downing Street, Cambridge, CB2 3EA, UK
Received 15 February 1996; accepted 21 February 1997
Abstract
High-resolution records (2 7 kyr) of Upper Pliocene Discoaster abundances obtained from six ODP/DSDP sites are assessed independently using oxygen isotope stratigraphy. Four Atlantic Ocean sites (DSDP Sites 552 and 607, and ODP Sites 659 and 662) comprise a transect from 56°N to I°S and provide a record of latitudinal variations in Diseoaster biogeography. Low-latitude sites in the Atlantic (ODP Site 662), Pacific (ODP Site 677), and Indian (ODP Site 709) oceans provide additional information about variability in Discoaster abundance patterns within the equatorial region. A common chronology, based on the astronomical time scale developed for ODP Site 677, has been established for all the sites. By integrating oxygen isotope data and Discoaster abundance records at each site we are able to independently evaluate the temporal and spatial distribution of D. brouweri and D. triradiatus in the 500 kyr prior to the extinction of the discoasters near the base of the Olduvai subchron.
Major decreases in abundance are evident during some of the more intense late Pliocene glacial events. In particular, glacial isotope stages 82, 96, 98 and 100 are associated with distinct abundance minima. At these times, large-scale changes in surface hydrographic conditions appear to have suppressed Diseoaster numbers on a global scale. The increase in abundance of D. triradiatus, which precedes the extinction of the discoasters by around 200 kyr, may also be related to the intensification of environmental pressures that accompanied the build-up of Northern Hemisphere ice sheets during the late Pliocene.
In spite of contrasting geographic and oceanographic settings, the various D. brouweri and D. triradiatus records are remarkably similar. This demonstrates that the acme and extinction events are excellent biostratigraphic datums. The simultaneous extinction of D. brouweri and D. triradiatus at 1.95 Ma were synchronous events at both a regional scale within the Atlantic, and on a global scale between the three major oceans. However, the start of the D. triradiatus acme appears to have been diachronous, occurring some 40 kyr earlier in the Atlantic than in the Indo-Pacific, and hence the stratigraphic usefulness of this datum is regional rather than global. © 1997 Elsevier Science B.V.
Keywords: Pliocene; palaeoceanography; Discoaster; isotope stratigraphy; Northern Hemisphere glaciation
* Corresponding author. Fax: 01223 334871; e-mail: [email protected]
0031-0182/97/$17.00 ~5 1997 Elsevier Science B.V. All rights reserved. PII $0031-0182(97 )00035-7

l l0 M.R. Chapman. A.J. (Twpslow-Lusty Pakteo~eo~ral)tly. Pa/aeoc/fllTatoh)JZy, Pakwoeco/o,.,,y 134 (1~97) 109 125
1. Introduction
Discoasters are an extinct group of calcareous nannofossils which are well represented in Tertiary marine sediments for over 55 million years, ranging from the Upper Palaeocene to the Upper Pliocene (Bukry, 1971: Perch-Nielsen, 1985). Their affinities are thought to lie with the extant coccolithophorids which inhabit the photic zone in the modern oceans (Chepstow-Lusty, 1996). Discoasters have been assumed to favour warm surface waters because of their consistently higher abundances in tropical cores and their absence in high-latitude records (Backman and Pestiaux, 1987; Chepstow-Lusty et al., 1991 ). High-resolution studies from a global suite of Upper Pliocene sites, have produced con- vincing evidence that, in addition to favouring warm surface waters, discoasters prefer low- productivity conditions and low-nutrient waters (Chepstow-Lusty et al., 1992).
Palaeoclimatic studies suggest that the Earth's climate system underwent a major change between 3.15 and 2.5 Ma (Raymo et al., 1989; Jansen and Sjoholm, 1991; Tiedemann et al., 1994; Shackleton et al., 1995). Benthic 6~80 records indicate both a progressive shift towards more positive values and a gradual increase in the amplitude of fluctuations during this time interval (Raymo et al., 1989; Tiedemann et al., 1994). This long-term trend in 6~sO values reflects changes associated with the large-scale growth of ice sheets in the Northern Hemisphere and a cooling of oceanic deep waters (Tiedemann et al., 1994; Dwyer et al., 1995). The establishment and intensification of Northern Hemisphere glaciation mark an important step in Neogene climatic evolution because it has resulted in a significant increase in the magnitude of global climatic variability over the last 2.5 Myr. In this paper, we examine the response of Discoaster populations to these major changes in palaeocli- matic and palaeoceanographic conditions.
The step-like decline evident in Discoaster diver- sity records (Fig. 1) suggests that major palaeo- ecological changes accompanied the onset of glacial activity in the Northern Hemisphere around 3 Myr ago (Ruddiman et al., 1987; Raymo et al., 1989; Jansen and Sjoholm, 1991 ). The general late Pliocene trend towards lower-species diversity is
reinforced when Discoaster surculus Martini and Bramlette and Discoaster pentaradiatus Tan became extinct following the major expansion of the Northern Hemisphere ice sheets at 2.5 Ma (Backman and Shackleton, 1983; Shackleton et al., 1984; Chepstow-Lusty et al., 1989: Wei, 1993: Tiedemann et al., 1994). The principal remaining taxon, Discoaster brouweri Tan, survives for a further 500 kyr over an interval of time which planktonic and benthic oxygen isotope records suggest was characterised by pronounced glacial interglacial climatic variability (Shackleton et al., 1990). A significant event occurs prior to the extinction of D. brouweri, when abundances of a three-armed form called Discoaster triradiatus Tan increased suddenly to comprise 20% of the total Discoaster population (Backman and Shackleton, 1983). Both of these taxa are illustrated in Fig. 2. The D. triradiatus acme has been repor- ted in numerous marine records (Backman and Shackleton, 1983: Chepstow-Lusty, 1990; Chepstow-Lusty and Chapman, 1995; Flores et al., 1995) and represents the most extreme form of skeletal reduction in the Tertiary evolution of the discoasters.
In this study, we examine high-resolution quanti- tative data obtained from six DSDP/ODP sites (Fig. 3; Table 1) that provide a global record of the distribution of late Pliocene discoasters. Because Discoaster extinction events yield valuable biostratigraphic information and are commonly used in the construction of age models, previous studies have often implicitly assumed these events to be synchronous (Chepstow-Lusty et al., 1989). In this investigation, we avoid this assumption by using oxygen isotope stratigraphy to provide an independent chronology. We then examine the timing of the extinction of D. brouweri and D. triradiatus and the onset and duration of the D. lriradiatus acme at each site, and assess whether these biological events were globally synchronous. Furthermore, the wide geographic coverage and strongly contrasting palaeoceanographic settings of the six sites considered here also enable us to investigate important environmental factors, such as sea surface temperature, surface circulation patterns, and upwelling intensity, which control

M. R, Chapman, A.J. Chepstow-Lusty / Palaeogeography, Palaeoclimatology, Palaeoecology 134 (1997) 109 125 111
n l
s _ G) ==
I m
10 I . _ G) (ll m 0 0
| m
GI
8 "
6
4
2
0
Expansive 91acial ~ icesheets "-
4
icesheets
1 Extinction 2 Extinction
Decline 3 Extinction
Extinction 4 Extinction
Extinction
I I I I I I I I I I I I I I I
1.8 2 2.2 2.4 2.6 2.8 3 3.2
D. variabilis D. tamalis D. asymmetricus D. surculus D. pentaradiatus D. brouweri D. triradiatus
Age (Ma) Fig. 1. Changes in Discoaster assemblage diversity (species number) during the late Pliocene (3.3 to 1.95 Ma).
phytoplankton ecology and may explain the final disappearance of the discoasters.
2. Materials and methods
Specimens of D. brouweri and D. triradiatus were identified and counted from evenly distrib- uted smear slides using a light microscope, produc- ing abundance values per mm 2 of slide, using the taxonomy and methodology of Backman and Shackleton (1983). The cores were sampled and Discoaster abundance counts generated at 10-cm intervals, except for Sites 607 and 709 which were sampled at 15- and 5-cm intervals, respectively. The mean sampling interval ranges from 2.1 kyr at Site 677 to 7.6 kyr at Site 552 (Table 1). In addition to the two abundance records (Sites 607 and 677) considered by Raffi et al. (1993), four previously published Discoaster abundance records (Chepstow-Lusty et al., 1989, 1992) are re-examined after their age models have been calibrated using oxygen isotope stratigraphy.
The placement of a last occurrence datum is complicated by the effects of sediment mixing and different age estimates may be assigned to the same biostratigraphic event depending on assumptions concerning bioturbation rates (Backman and Shackleton, 1983; Raymo et al., 1989; Raffi et al., 1993; Wei, 1993). The presence or absence of both D. brouweri and D. triradiatus can make the identi- fication of the Discoaster extinction event more reliable, although reworking remains a potential problem. We consider that the continuous presence of both these species is important for defining the true extinction event, and single specimens of either species distributed discontinuously, which also reg- ister as wildly fluctuating percentages in the D. triradiatus acme, are probable indicators of reworking. Hence, an arbitrary but practical cut- off of 5 specimens/mm 2 was applied to counter the effect of low, infrequent occurrences of a few specimens on the stratigraphic placement of the extinction datum. It should be noted that this criterion could not be applied at Site 552 because of the extremely low Discoaster abundances, but

112 M.R. Chapma,, A.J. (Twpston'-Lttsl.v / Palueo~eo~raphy, Palaeoclimatolo~v. PalaeoecoloL~' 134 (1997) 109 125
1 2
I i 1 0 #
Fig. 2. Light microscope photographs of ( 1 ) D. hrouwer i and (2) D. triradiatu.s.
the results from this core are compatible with the other records most probably due to the very low abundances reducing the effects of bioturbation.
The timing of the D. triradiatus acme was also investigated. This event represents a significant increase in the importance of the D. triradiatus population relative to the D. brouweri form: the onset of the acme is defined as the point when D. triradiatus accounts for 20% of the combined pop- ulation (Backman and Shackleton, 1983). Since D. triradiatus occurs only in minor abundances before the acme, reworking is not considered to be an important problem for this event.
The four sites within the Atlantic Ocean com- prise a latitudinal transect. Site 662 is located close to the equator on the upper eastern flank of the Mid-Atlantic Ridge within the equatorial diver- gence zone (Ruddiman et al., 1988) where the surface waters are influenced by the South
Equatorial Current. Because of the effects of equa- torial upwelling and high productivity the sedi- mentation rate at this site is quite high (4 cm/kyr). Site 659 located on the top of the Cape Verde Plateau, off northwest Africa, was selected as a non-upwelling, subtropical open-ocean site affected by the cool Canaries Current and has a sedimentation rate of 3 cm/kyr (Ruddiman et al., 1988). Site 607 is presently located along the northernmost part of the subtropical gyre in the vicinity of the North Atlantic Current and has a lithology comprising interbedded foraminifer nan- nofossil oozes and marls representing the North Atlantic glacial marine cycles (Ruddiman et al., 1987). This site has the second highest sedimenta- tion rate (5 cm/kyr) of the six sites. Site 552 is the northernmost point of our transect and is located on the southwest margin of the Rockall Plateau in the path of the North Atlantic Current (Roberts

M.R. CTmpman, A.J. Chepstow-LusO' / Palaeogeography, Palaeoclh~Tatology, Palaeoecology 134 (1997) 109-125 113
1 2 0 ° 6 0 ° 0 ° 6 0 ° 1 2 0 °
oo
30 °
6O ° I I I I I 1 2 0 ° 6 0 ° 0 ° 6 0 ° 1 2 0 °
Fig. 3. Location of DSDP/ODP sites investigated in this work.
60 °
30 °
0 °
30 °
60 °
Table 1 Details of DSDP/ODP sites studied in this work
Site 552 607 659 662 677 709
Water depth (m) 2304 3427 3067 3824 3461 3038 Location 56N, 23'W 41 N, 33W 18N, 21:W 1 S, l l:W 1N, 84'W 4S, 61:E Mean sedimentation rate (cm/kyr) 1.6 4.6 3.2 4.1 4.9 1.1 Mean sampling interval (kyr) 7.6 3.6 3.5 2.5 2.1 5.4
et al., 1984). The l i thology is s imilar to tha t found at Site 607 but the sed imen ta t ion rate is much lower (2 cm/kyr ) .
The two o ther low- la t i tude records are f rom Sites 677 and 709 in the Pacific and Ind ian oceans, respectively. Site 677 is loca ted in the eastern equa to r ia l Pacific between the E c u a d o r and P a n a m a Frac tu re Zones in a m a j o r upwel l ing area (Becker et al., 1988). Site 677 is s i tuated near to the Peru Cur ren t and equa to r i a l d ivergence system and has been shown to have been loca ted in an area o f high biological act ivi ty for at least several hundred t h o u s a n d years ( M o o r e et al., 1973). Pelagic sediments , cons is t ing o f sil iceous and cal- careous nannofoss i l ooze, account for Site 677 having the highest mean sed imen ta t ion rate o f all the sites (5 cm/kyr ) . In contras t , Site 709 has the
lowest sed imen ta t ion rate (1 cm/kyr ) on account of its loca t ion in a region o f the western equa tor ia l Ind ian Ocean affected by stable, warm, low- p roduc t iv i ty surface waters. The Site 709 record was recovered f rom a small basin perched near the summit o f the Mad ing ley Rise, a regional topo- graphic high between the Car l sberg Ridge and the nor the rn Masca rene Pla teau ( B a c k m a n et al., 1988).
3. S t r a t i g r a p h y a n d a g e m o d e l s
Accura t e and rel iable in ter-core s t ra t ig raphic cor re la t ions are required to fully evalua te the geograph ica l extent o f popu l a t i on var ia t ions and the synchrone i ty o f the Discoaster datums. By

114 M.R. (Tlapman, A.J. (7~epstow-Lust.v ,' Pa/aeoL, eo~raphv Palaeoclimalo/o~l', Palaeoecolo£y 134 (1997) 109 125
0
GI 13. 0
0 ¢0
3.0
3.5
4.0
4.5
5.0
2.5
3.0
3.5
4.0
4.5
2.4
3.0
3.6 r
4.2
4.8
-2.3 -
- 1 . 8 -
- 1 . 3
-0.8
-0.3
2.5
3.1
3.7
4.4
5.0
-2.0
-1.5
-1.0
-0.5
0.0 1.8
Site 552
O/M ' I ' I ' I ' I ' I ' I ' I ' I
Site 607 70 72 78 8O 82
M/G I ' I ' I ' I ' I ' I ' I
96 98 100
~ G ' I ' l ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I
Site 659
O/M 6 • ' I ' I ' I ' I " I ' I ' I ' I ' I ' f ' I ' f ' I ' I ' I ' I
Site 662
• I " I " I ' I ' I ' I ' I " I ' I ' I ' I ' I " I " I ' ! ' I
Site 677
' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I ' I = I ' I
Site 709
' I ' I " I " I ' I ' I " I " I ' I " I " I ' I " I ' 1 " I " I
1.9 2 2.1 2.2 2.3 2.4 2 .5 2 .6
Age (Ma)

M.R. Chapman, A.J. Chepstow-LusO" / Palaeogeography, Palaeocl#~atology, Palaeoecology 134 (1997) 109 125 115
integrating the orbitally tuned oxygen isotope records from the sites in this study, which are independent from the Discoaster abundance records, we are able to assess the nature and timing of these biostratigraphic events at a high resolution and on a global scale. The astronomical time scale derived from oxygen isotope data at Site 677 (Shackleton et al., 1990; hereafter referred to as SBP90) provides the template for the development and refinement of the other stratigraphies. The oxygen isotope stratigraphy developed by Tiedemann et al. (1994) for Site 659 is consistent with the SBP90 chronology over our studied time interval. The isotopic data obtained from Site 552 (Shackleton and Hall, 1984), Site 607 (Raymo et al., 1989), Site 662 (Karlin et al., 1989) and Site 709 (Shackleton and Hall, 1990) have been correlated to the Site 677 isotope record. All isotopic records are shown plotted against age in Fig. 4. At three of the sites (552, 607 and 659) magnetostratigraphic data provide an independent means of verifying these correlations.
Additional control points were needed to con- strain the age models at Sites 709 and 662. Oxygen isotope stages 96 to 100 are well-defined events in the benthic isotope records from Sites 552, 607, 659 and 677, but they are not clear in the plank- tonic isotope record at Site 709. A benthic isotope record was generated to confirm the identification of these isotope stages (M. Chapman, unpublished data). Furthermore, the age model at Site 662 is only partly based on isotope stratigraphy because the isotopic record at Site 662 does not extend back to 2.6 Ma. Hence, age estimates are less rigorously constrained over the interval 2.6 2.25 Ma because they are calculated by linear interpolation between the end point of the isotope record and the extinction of Dentoglobigerina altispira at 3.01 Ma (Chapman et al., 1996).
4. Latitudinal patterns within the North Atlantic
Examination of the North Atlantic D. brouweri abundance patterns (Fig. 5) reveals strong evi-
dence of cyclicity over the studied time interval (Chepstow-Lusty et al., 1989). The Discoaster abundances at equatorial Site 662 are relatively high in an Atlantic context, although the D. brouw- eri record is characterised by a marked drop in abundance around 2.5 Ma and a trend towards lower abundances after 2.3 Ma. The rapid increase in D. triradiatus which marks the base of the D. triradiatus acme (Figs. 6 and 7) occurs during isotope stage 8l (2.14 Ma). The extinction of D. brouweri and D. triradiatus occurs within inter- glacial stage 71 (1.94 Ma). The broad patterns observed in the subtropical record from Site 659 are similar to those identified at Site 662. Discoaster production was suppressed around 2.5 Ma and the D. triradiatus acme begins in stage 81 (2.15 Ma). However, the extinction of D. brouweri and D. triradiatus occurs slightly earlier at the end of glacial stage 72 ( 1.95 Ma). Although Discoaster abundances are greatly reduced in the interval after 2.3 Ma, a distinctive peak in abun- dance around 2.0 Ma is evident at Site 659.
Discoaster abundances are significantly reduced at Site 607 and extremely low at Site 552 but the marked abundance fluctuations around 2.5 Ma closely resemble those found in the low-latitude records (Fig. 5). At Site 607 the base of the D. triradiatus acme occurs in the middle of stage 82 (2.17 Ma) but it appears that the D. triradiatus acme, which began in stage 81 (2.14 Ma) at Site 552, is stratigraphically restricted because of the very low and discontinuous distribution of dis- coasters this far north (5CN) . In spite of the lower abundances in these mid-latitude records, the abundance peak around 2.0 Ma observed at Site 659 is clearly present in the records from Sites 607 and 552. The extinction of D. brouweri and D. triradiatus can also be accurately distinguished in stage 72 (1.95 Ma). Magnetostratigraphic data from Site 607 indicate that this event occurred approximately at the base of the Olduvai subchron (Chepstow-Lusty et al., 1989; Raffi et al., 1993).
Comparison of the distribution of discoasters
Fig. 4. Orbitally-tuned ~5180 isotope records for the Atlantic (Sites 552, 607, 659, 662), Pacific (Site 677), and Indian (Site 709) oceans. Key glacial isotope stages are numbered. Position of magnetic reversals are indicated: Olduvai/Matuyama (O/M), Matuyama/Gauss (M/G).

116 M.R. CTutpman, A.J. Ctlepstow-Lu,sly / P(t/ae(~eoq, rap/ y, P(tlaeoclimam/o~ '. Pa/aeoecolo~zy 134 (1997) 109 125
4
3
2
1
0
80
60
A 40
i • 2O
g o 500
~,~ 375
" o 250
125
0
400
300
200
1 O0
o 600
, ~ 450
300
150
1800
1350
900
450
1.8
Site 552 72 82
iiiii • ] " I
96 98 100
' I ' I " ' I ' I ~ I - '
Site 607 72 82 96 98 100
Site 659 t . 72 82 . 96H 98 lOO
iiiii i i i ' I ' I ' I ~ I ' I ' ' I ' I ' ' I ' I ' '
Site 662 ]1
' " I ' " I " I ' I " I " ' I " I "
Site
. . . . . i iiii • I 1 l ~ l ~ I = I " " I " I ' " I " I '
Site 7 0 ~ 7 2 8 2 9 6 9 8 1 0 0
ii~i . . . . '~ . . . . . iiii , iiS ,iiii , I ' I I I I I I I I
1.9 2 2.1 2.2 2.3 2.4 2.5 2.6
Age (Ma) Fig. 5. Abundance variations of Di,~('oaxter hromreri between 2.6 and 1.95 Ma. Shaded areax indicate the position of prominent late Pliocene glacial events discussed in text.

M.R. Chapman, A.J. Chepstow-Lusty / Palaeogeography, Palaeoclimatologv Palaeoecology 134 (1997) 109 125 l 17
0 . 8 -
0 . 6 -
0.4 -
0.2 -
0.0
4 0 -
30~
10-
0
100 -
502 JO 25-
0
2 0 ,
15-
10-
5
~ o 2O
o 15
120
.90
60
30
0
1.8
Site 552
I
Site 607
I
Site 659
I
Site 662
I
Site 677
I
Site 709
I
1.85 1.9
8 i __ ^ a _ i _
r ' I " [ 1 I - r I ' AA ^ .
I " I " I " I
72 82
I I - - ' I I - r - I " ' - - I - ' I ~ - 1 ~ - - I
72
i - - • - 1 = -1 I r T - ' I I 1 - [ - 1 I
I ' . . . . . . .
1,95 2 2.05 2.1 2.15 2.2 2.25 2.3 2.35 2.4
Age (Ma) Fig. 6. Abundance variations of Discoaster triradiatus between 2.4 and 1,95 Ma.

118 M.R. (Tta/~m(m, A.J. (7~elZ~'to~r-Llt.~ O' ,, Pa/aeo~,*eo~rat# U" Pa/aeoc/imato/o~r Palaeoecolo~)" 134 (1997~ 109 125
0 ¢.. ¢0
'10 ¢=
,¢, ca
?,
100 -
7 5 -
5 0 -
25 -
0
1 O0 -
7 5 -
5 0 -
2 5 -
0
1 O0 -
7 5 -
5 0 -
25-
0
100 -
75-
50
25
0
1 O0 -
7 5 -
50-
25
0
100
75
50
25
0
1.8
Site 552
I ' | i ' - - 1 1
Base < D. triradiatus | |
acme M
. . . . . . . . . A _ _~ 1 - r - - I ' f ~ I ~ - - I - - r I
Site 607 Base
D. triradiatus acme
Site 659 Base D. triradiatus
acme
Site 662 ~ D. triradiatus
acme
i , - r - - _ _ ,
Site 677 Base D. triradiatus
aolle
1 - " I
Site 709
I
1.85 1.9
Base D. triradiatus
acme
-! 1.95 2 2.05 2.1 2.15 2.2
= - . T - , ~ " T , = ; 2.25 2.3 2.35 2.4
Age (Ma) Fig. 7. Rela t ive a b u n d a n c e of D. triradialux. The onset of" the acme, where D. triradiams represents 20% or more or the total Dixcoa.~ter popuht t ion , is indicated for each record.
M.R. Chapman. A.J. Chepstow-Lusty / Palaeogeography, Palaeoclimatology. Palaeoecology 134 (1997) 109 125 I I 9
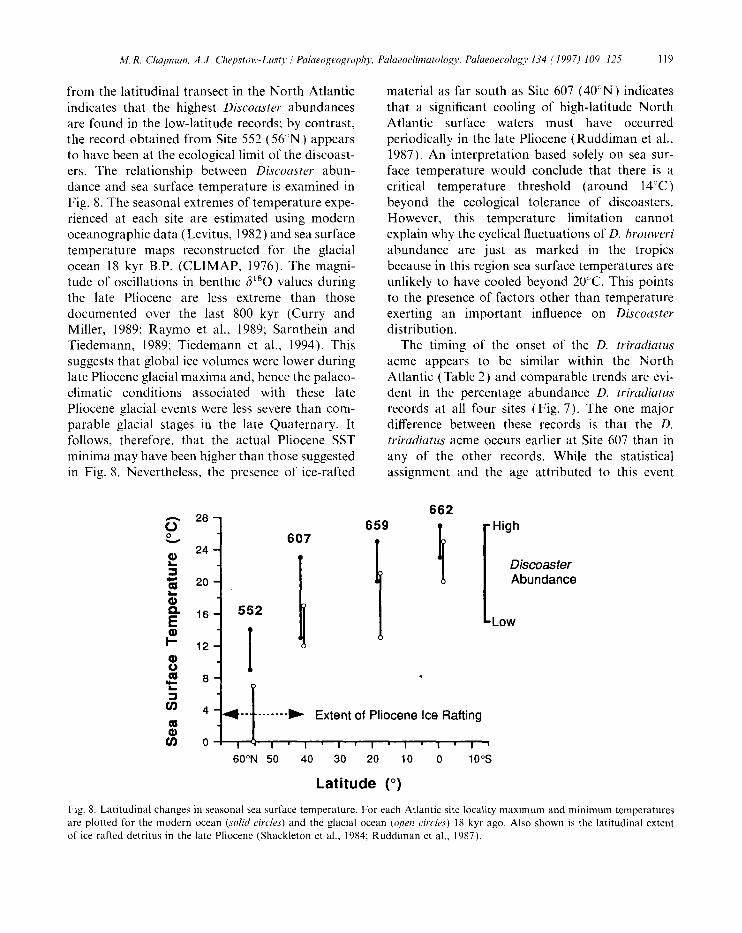
tYom the latitudinal transect in the North Atlantic indicates that the highest Discoaster abundances are found in the low-latitude records; by contrast, the record obtained from Site 552 (56°N) appears to have been at the ecological limit of the discoast- ers. The relationship between Discoaster abun- dance and sea surface temperature is examined in Fig. 8. The seasonal extremes of temperature expe- rienced at each site are estimated using modern oceanographic data (Levitus, 1982) and sea surface temperature maps reconstructed for the glacial ocean 18 kyr B.P. (CLIMAP, 1976). The magni- tude of oscillations in benthic 6180 values during the late Pliocene are less extreme than those documented over the last 800 kyr (Curry and Miller, 1989; Raymo et al., 1989; Sarnthein and Tiedemann, 1989; Tiedemann et al., 1994). This suggests that global ice volumes were lower during late Pliocene glacial maxima and, hence the palaeo- climatic conditions associated with these late Pliocene glacial events were less severe than com- parable glacial stages in the late Quaternary. It follows, therefore, that the actual Pliocene SST minima may have been higher than those suggested in Fig. 8. Nevertheless, the presence of ice-rafted
material as far south as Site 607 (40°N) indicates that a significant cooling of high-latitude North Atlantic surface waters must have occurred periodically in the late Pliocene (Ruddiman et al., 1987). An interpretation based solely on sea sur- face temperature would conclude that there is a critical temperature threshold (around 14°C) beyond the ecological tolerance of discoasters. However, this temperature limitation cannot explain why the cyclical fluctuations of D. brouweri abundance are just as marked in the tropics because in this region sea surface temperatures are unlikely to have cooled beyond 20°C. This points to the presence of factors other than temperature exerting an important influence on Discoaster distribution.
The timing of the onset of the D. triradiatus acme appears to be similar within the North Atlantic (Table 2) and comparable trends are evi- dent in the percentage abundance D. triradiatus records at all four sites (Fig. 7). The one major difference between these records is that the D. triradiatus acme occurs earlier at Site 607 than in any of the other records. While the statistical assignment and the age attributed to this event
• ~ 28 - 0 o.. 24
L _
20 L -
Q" 16 E I - 12 0
8 N = -
(/) 4 ¢g O) 0
607
552 I
I 60°N 50
662 659
Extent of Pliocene Ice Rafting
I ' I I I ' I '
40 30 20 10 0 10°S
Lati tude (o)
High
Discoaster Abundance
Low
Fig. 8. Latitudinal changes in seasonal sea surface temperature. For each Atlantic site locality maximum and minimum temperatures are plotted for the modern ocean (solid circles) and the glacial ocean (open circles) 18 kyr ago. Also shown is the latitudinal extent of ice rafted detritus in the late Pliocene (Shackleton et al., 1984; Ruddiman et al., 1987).

120 M.R. (']icqmtalT. A.J. C7wt),~tow-Lu,s'ty Pa[aeo,¢,eogreq~h.v. Pa[aeoc/hnalolo,~,,.t'. Pa]aeoecolog)" 134 ; 1997) 109 125
Table 2 Age estimates of Dixcoa,sler datums and corresponding isotope stages
Site 552 607 659 662 677 709
Base D. triradiams acme (Ma) 2.14 2.17 2.15 2.14 2.11 2.11 Oxygen isotope stage 81 82 81 81 80 80 Extinction D. hromreri and
and D. lriradiatus (Ma) 1.95 1.95 1.95 1.94 1.95 1.95 Oxygen isotope stage 72 72 72 71.72 72 72 Duration D. triradiatus acme (kyr) 190 22(1 200 2(10 160 160
(2.17 Ma) are correct, the acme does not mark a major increase in the abundance of D. triradiatus. This age discrepancy therefore appears to be due to the very low abundances of D. brouweri and D. triradiatus during stage 82 affecting the assemblage composition rather than a true increase in D. triradiatus abundance, which clearly occurs during stage 81 (2.14 Ma) at the same point as in the other D. triradiatus records (Fig. 6).
The distinctive double abundance peaks of D. brouweri found prior to the extinction level at three sites (552, 607 and 659) provide further evidence supporting the inter-site correlations made on the basis of fi~sO stratigraphy. These prominent abundance peaks occur after an interval of suppressed abundance and are considered to be genuine palaeoecological events because they are found across a large part of the North Atlantic (Fig. 6). The subsequent decline in Discoaster abundances during glacial isotope stage 72 also assists in the reliable identification of the extraction datum at 1.95 Ma. The estimated ages for the extinction of both D, hrouweri and D. triradialus are summarised in Table 2. Overall the Discoasler abundance records are remarkably consistent, although the Site 662 record suggests that discoast- ers might have survived slightly longer in the equatorial Atlantic than elsewhere.
5. Comparison of equatorial records
The records from the Indo-Pacific cores docu- ment a much more marked reduction in D. brouw- eri abundance around 2.15 Ma (stage 82) than at Site 662 in the Atlantic Ocean (Fig. 5). Discoaster populations are especially suppressed alter
2.15 Ma at Site 677 and, although there is a brief abundance peak at the end of stage 79 (2.08 Ma), the abundances over the following 200 kyr remain low right up to the extinction datum (Chepstow- Lusty and Chapman, 1995). The base of the D. triradiatus acme occurs in stage 79 (2.11 Ma) at Site 677, appreciably later than this event in the Site 662 record (2.14 Ma). In spite of the low abundances of D. bl'ouweri and D. triradiatus, the D. triradkltus acme is clearly identifiable as is the extinction of both these species in stage 72 (1.95 Ma). Rafli et al. (1993) did not consider these low abundances significant enough to iden- tify the extinction event. However. we believe that the observed fluctuations in Did'coaster popnlations are unlikely to be due to reworking in a core with such a high sedimentation rate and subdued Discoasler abundances.
The late Pliocene sediments from the western equatorial Indian Ocean are characterised by very high Discoaster abundances. The D. hrouweri record at Site 709 is similar to the Site 677 record in that abundances are much greater prior to stage 82. After this cold glacial event, the D. hrouweri abundances are much reduced up to the extinction event. Although relatively low for Site 709, the D. brouweri abundances do remain higher than any of the other sites ( Fig. 5). At 2.17 Ma, D. lriradia- tus abundances suddenly increased to around 10% of the total Dis'coaster population ( Figs. 6 and 7), but this level is below the recognised definition of the D. lriradialus acme (20%). This 10% represen- tation is maintained l\)r 60 kyr until stage 80 (2.11 Ma), when the relative abundance of D. triradiatus linally exceeds 20%. The extinction of both D. hrouweri and D. lriradiatus occurs within

M.R. Chapman, A.J. Chepstow-Lusty / Palaeogeography, Palaeoclimatology, Palaeoecology 134 (1997) 109 125 121
isotope stage 72 (1.95 Ma) in agreement with the other records.
The dramatic reduction in Discoaster abundance at Site 677 over the last 200 kyr of their strati- graphic range is interesting as it may signify a marked change in the planktonic ecology and surface water chemistry of the Pliocene oceans. In the modern eastern equatorial Pacific, the phyto- plankton assemblages are dominated by siliceous diatoms which thrive in the cool, upwelled, nutri- ent-rich surface waters (Dymond and Lyle, 1985). Fig. 9 shows that the decline in Discoaster abun- dance over the interval 2.2-1.9 Ma was also coin- cident with an increase in diatom abundance (Chepstow-Lusty, 1990). Large, parallel cyclic fluctuations in the production of biogenic C a C O 3 and opal during the late Pliocene are clearly inversely related (Murray et al., 1995). In the late Pliocene to early Pleistocene, Kemp (1995) docu- mented a significant decrease in the CaCO3 content of the eastern equatorial Pacific sediments which he attributed to a rapid increase in diatom abun- dance. This change is also evident as a prominent increase in the opal mass accumulation rate at ODP Site 847 and implies that the relative flux of siliceous microplankton was much higher between 2.1 and 1.7 Ma (Murray et al., 1995).
The timing of the onset of this high-productivity period appears to have been linked to the expan- sion of Northern Hemisphere glaciation around 2.5 Ma (Shackleton et al., 1984; Sikes et al., 1991; Tiedemann et al., 1994) and the closure of the Panamanian seaway (Keller et al., 1989). It is possible that enhanced upwelling of silica into the surface waters of the Peru Current South Equatorial Current system preferentially favoured diatom floras, which have high nutrient and silica demands (Codispoti, 1983), at the expense of calcareous phytoplankton including the discoast- ers. Consequently, variations in the late Pliocene Discoaster records from Site 677 may be a response to fluctuations in the proximity and intensity of upwelling and the effect that this had on surface water characteristics, such as nutrient availability and temperature. Quantitative examination of the total nannofossil assemblages at Sites 849 and 852 indicates that productivity rather than dissolution is the main factor controlling the relative abun- dance of discoasters in the eastern equatorial Pacific (Flores et al., 1995). Furthermore, Discoaster abundances were found to be relatively low in the equatorial divergence region because of the higher production rates of the very small reticulofenestrids (VSR), a group of dominant
~ ' 75 ' u
E 50
E 25 0 ¢a
, i
0
~iii~i i i~ i 500
. . . . . . . . ; l 0
'E E ¢/) s . _
m o u o~
i n
1.9 2 2.1 2.2 2.3 2.4 2.5 A g e ( M a )
Fig. 9. Variations in the abundance of discoasters (solid circles) and diatoms (open circles) between 2.5 and 1.9 Ma at Site 677.

122 M.R. Chapman. A.J. ('hepstow-Lus O' / Palaeogeograph.v, Palaeoclimato/oX~v, Palaeoecology 134 (1997) 109 125
opportunistic taxa that are believed to thrive in nutrient-rich conditions (Flores et al., 1995).
Site 662 in the eastern tropical Atlantic is also situated in an equatorial upwelling region but the modern surface production, and hence the under- lying sediments, are predominantly biogenic car- bonate (Wefer and Fischer, 1993) and the fertility of the surface waters and rates of nutrient influx are lower than in the eastern equatorial Pacific. Although Dis'coaster abundances did decline during the latest Pliocene it seems that the sea surface conditions at Site 662 were more favoura- ble than those at Site 677. Differences between equatorial records have also been reported from the western and eastern equatorial Pacific (Chepstow-Lusty and Chapman, 1995) which demonstrate that Dis'coaster abundance cannot simply be explained as a function of latitude. This is further exemplified by the much higher Discoaster abundances recorded at Site 709. Modern sea surface temperatures at Site 7(19 exhibit relatively little seasonal variability ( 1 2 C ) and the mean sedimentation rates indicate that carbonate productivity is much lower here than at the other two low-latitude sites. It is possible to conclude that the consistently high sea surface temperatures and low nutrient levels maintained at Site 709 were advantageous oceanographic con- ditions for D. brouweri and D. triradiatus. In spite of the strongly contrasting oceanographic regimes, the estimated age for the base of the D. triradiatus acme appears to have been consistent between Site 677 and Site 709, as was the timing of the extinction of D. brouweri and D. triradiatus, but the timing of the acme event at these two sites differs from the tropical Atlantic record at Site 662 (Table 2).
6. Dis'coaster abundance and global climate: a palaeoceanographic scenario
Even though the sites considered here are widely distributed and affected by very different oceano- graphic conditions, the global abundance patterns exhibit some broad similarities. Fluctuations in D. brouweri and D. triradiatus abundance appear to relate directly to glacial interglacial climatic oscil- lations and the intensity of Northern Hemisphere
glaciation. In particular, major decreases in abun- dance are coincident with some of the more intense glacial isotope stages (72, 82, 96, 98 and 100) and these minima can be correlated between the different sites. This relationship is confirmed by spectral analysis of the Dis'coaster abundance records which reveal periodicities that correspond to precessional and obliquity orbital forcing fre- quencies (Chepstow-Lusty et al., 1989). Changes in the surface water hydrography brought about by the more vigorous circulation patterns during glacial climatic states appear to have suppressed Discoaster numbers on a global scale. The fact that abundances drop to such low values at all sites apart from Site 709 suggests that the warm, oligotrophic subtropical gyres where surface condi- tions remained relatively stable might have acted as refugia for discoasters in glacial times. The reduced intensity of ocean circulation and decreased latitudinal gradients during interglacial episodes, and perhaps changes in the carbon- ate silica budget of the surface oceans, could have been major factors controlling the more wide- spread distribution of discoasters when climatic states were more favourable.
D. brouweri abundances fluctuate markedly over the entire 2.6 1.95-Ma interval, but peak abun- dance values are generally lower after 2.15 Ma (Fig. 5). This prolonged interval of reduced abun- dance which precedes the extinction event coin- cided with an increase in abundance of the D. triradiatus form (considered by many to be merely a variant of D. brouweri). In general, the relation- ship between D. brouweri and D. triradiatus over the time interval 2.15 to 1.95 Ma is consistent (Figs. 6 and 7) and both species have distinctive abundance peaks prior to the extinction level. The expansion of D. triradiatus' could reflect the last adaptive response of the discoasters to the accentu- ation of glacial interglacial climatic variability during the late Pliocene. If this were the case, the switch to D. triradiatus morphology, which may represent the limits of the available genetic flexibil- ity, was insufficient to prevent the extinction of this long-lived lineage.
Although the complex interplay of factors such as productivity pressure and surface temperatures may explain global shifts in Dis'coaster popula-

M.R. Chapman. A.J. Chepstow-Lusty / Palaeoy;eography. Pahwoclimatology, Pakwoecology 134 (1997) 109 125 123
tions, it is more difficult to envisage how these processes could account for the global pattern of the D. triradiatus acme and the global synchroneity of the extinction of both D. brouweri and D. triradiatus. Mechanisms that explain these rapid global biological events solely in terms of climatic factors appear to be insufficient, especially because populations of D. brouweri had previously managed to survive glacial events of similar magnitude (glacial isotope stages 96, 98 and 100).
In the biosphere, climatic pressure can certainly lead to increased competitive pressure from other species, especially if they are rivals competing for a niche that is no longer optimal for the original species. However, if this was the case our palaeon- tological records indicate that these processes must have operated extremely rapidly on a global scale. In fact, the synchroneity of the extinction events and the absence of vestigial populations is some- what surprising because the shoaling of the Panama Isthmus during the late Pliocene (Keller et al., 1989) would have led to the effective genetic isolation of low-latitude planktonic populations within the Atlantic and Indo-Pacific biogeographic provinces. If the refugial hypothesis outlined earlier is correct, a marked change in the depth of the thermocline or nutricline characteristics of the subtropical gyres could account for the extinction of the discoasters. Alternatively, the demise of Discoaster populations could reflect enhanced competition pressures within the phytoplankton community resulting from the encroachment and expansion of a new species or adaptationally advantaged taxa into the subtropical ocean. In the latest Pliocene, there is some evidence that the disappearance of the discoasters was accompanied by an increase in the importance of other gephyro- capsids and VSR nannofossil floras (Raffi et al., 1993; Flores et al., 1995), but the mechanism for any replacement is not straight forward. It is therefore difficult to accurately assess the relative contribution of these various factors in the extinc- tion of the long-lived Discoaster genus.
7. Conclusions
Variations in the relative abundance records of discoasters show that this extinct group of
microfossils was affected by enhanced glacial interglacial climatic variability and environmental changes following the accentuation of Northern Hemisphere glaciations during the late Pliocene. Variations in D. brouweri and D. triradiatus abun- dance records indicate that total Discoaster pop- ulations were greatly reduced in times of intense glacial conditions. The diverse oceanographic set- tings and wide geographical coverage of the D. brouweri and D. triradiatus records considered here also clearly demonstrate that the D. triradiatus acme and the Discoaster extinction events are useful biostratigraphic datums.
Age estimates for the base of the D. triradiatus acme differ by around 40 to 60 kyr (2.17 to 2.11 Ma). Consistent age estimates obtained from the North Atlantic (2.15 Ma) suggest that the D. triradiatus acme occurs about 40 kyr (one obliq- uity cycle) earlier in the North Atlantic than in the Indo-Pacific ocean. For approximately 200 kyr, D. brouweri and D. triradiatus covary in abundance before both species disappear simultaneously at 1.95 Ma (isotope stage 72)just above the base of the Olduvai subchron (Chepstow-Lusty et al., 1989; Raffi et al., 1993). It should be noted that the ages reported here are based on the SBP90 time scale and hence are older than those found elsewhere in the literature (e.g., Chepstow-Lusty et al., 1989).
The disappearance of the discoasters appears to have been a truly synchronous event, both in the different ocean basins and also between equatorial and mid-latitude regions within the Atlantic. The extinction event occurs nearly 600 kyr after the expansion of Northern Hemisphere glaciation (Tiedemann et al., 1994) and during the final stages of the closure of the Panamanian seaway ( Keller et al., 1989). The actual mechanism respon- sible for the extinction of the discoasters is intri- guing as it must operate at a global level over a geologically short time scale. Large-scale climatic changes at the end of the Pliocene, altering the nutrient and temperature characteristics of the global ocean, and increasing competition pressures within the phytoplankton communities are both viable mechanisms for reducing Discoaster popula- tion sizes beyond a sustainable threshold.

124 M.R. ('hapman, A.J. CTwl)Stow-Lusly / Palaeogeograplty. Pahwoe/hnalo/ogSv, Palaeoeeolog~V 134 (1997) 109 125
Acknowledgements
M.R.C. was supported by NERC, A.C.-L. was funded by NERC, BP, Montgomery Trust and the Henry Giles Fellowship. We are grateful for the work of shipboard scientists and ODP who pro- vided the material used in this study. We especially thank Nick Shackleton and Jan Backman for fruitful discussion over the years. We are grateful to Jose Flores, Isabella Raffi, and an anonymous reviewer for their comments and suggestions to improve the original manuscript. The light micro- scope photographs were taken with the assistance of Kate Conway.
References
Backman, J., Pestiaux, P., 1987. Pliocene Diaeoaster abundance wlriations from DSDP Site 606: biochronology and paleo- enviromnental implications, lnit. Rep. DSDP 94, 903 910.
Backman, J., Shackleton, N.J., 1983. Quantitative biochronol- ogy of Pliocene and early Pleistocene calcareous nannofossils from the Atlantic, Indian and Pacific oceans. Mar. Micro- paleontol. 8, 141-170.
Backman, J., Duncan, R.A., et al. (Eds.), 1988. lnit. Rep. ODP, 115. Ocean Drill. Prog., College Station, TX.
Becker, K., Sakai, H., et al. (Eds.), 1988. Init. Rep. ODP, 111. Ocean Drill. Prog., College Station, TX.
Bukry, D., 1971. Diseoa.~ter evolutionary trends. M icropaleon- tology 17, 43 52.
Chapman, M.R., Funnell, B.M., Weaver. P.P.E.. 1996. High resolution Pliocene planktonic foraminiferal biozonation of the tropical North Atlantic. In: Moguilevsky, A., Whatley, R. (Eds.), Microfossils and Oceanic Environments. Univer- sity of Wales, Aberystwyth-Press, Aberystwyth, pp. 307 316.
Cbepstow-Lusty, A., 1990. Nannoplankton as indicators of cli- matic variability in the Upper Pliocene. Ph.D. Thesis, Cam- bridge University, Cambridge, 325 pp. (unpubl.).
Chepstow-Lusty, A., 1996. The last million years of the dis- coasters: a global synthesis for the Upper Pliocene. In: Moguilevsky, A., Whatley, R. (Eds.), Microfossils and Oce- anic Environments. University of Wales, Aberystwyth-Press, Aberystwyth, pp. 165 175.
Chepstow-Lusty, A., Chapman, M., 1995. The decline and extinction of upper Pliocene discoasters: A comparison of two equatorial Pacific Ocean records. J. Nannoplankton Res. 17, 15-19.
Chepstow-Lusty, A., Backman, J., Shackleton. N.J., 1989. Comparison of Upper Pliocene Discoaster abundance varia- tions fiom North Atlantic Sites 552, 607, 659, 658 and 662: Further evidence for marine plankton responding to orbital
forcing. Proc. ODP, 108. Ocean Drill. Prog., College Station, TX, pp. 121 141.
Chepstow-Lusty, A., Backman, J., Shackleton, N.J., 1991. Palaeoclimatic control of Upper Pliocene Di~eoaster assemblages in the North Atlantic. J. Micropalaeontol. 9 (2), 133 143.
Chepstow-Lusty, A., Backman, J., Shackleton, N.J., 1992. Comparison of Upper Pliocene Diseoaster abundance varia- tions from the Atlantic, Pacific and Indian Oceans: The signi- ficance of productivity pressure at low latitudes. Mere. Sci. Geol. 44, 357 373.
CLIMAP Project Members, 1976. The surface of the ice-age earth. Science, 191, 1131 1137.
Codispoti, L.A., 1983. On nutrient availability and sediments in upwelling areas. In: Suess, E., Thiede. J. (Eds.), Coastal Upwelling, Part A: Responses of the Sedimentary Regime to Present Coastal Upwelling. Plenum, New York, NY, pp. 125 145.
Curry. W.B., Miller, K.G., 1989. Oxygen and carbon isotopic variation in Pliocene benthic foraminifers of the Equatorial Atlantic. Proc. ODP, Sci. Results, 108. Ocean Drill. Prog., College Station, TX, pp. 157 166.
Dwyer, G.S., Cronin, T.M., Baker, P.A., Raymo, M.E., Buzas, J.S.. Correge, T., 1995. North Atlantic deepwater temper- ature change during late Pliocene and late Quaternary climatic cycles. Science 270, 1347 1351.
Dymond, J., Lyle, M., 1985. Flux comparisons between sedi- ments and sediment traps in the eastern tropical Pacific: implications for atmospheric CO, variations during the Pleistocene. Limnol. Oceanogr. 3(I, 699 712.
Flores, J,A., Sierro, F.J., Rafli, 1., 1995. Evolution of the calcar- eous nannoli)ssil assemblage as a response to the paleoceano- graphic changes in the eastern equatorial Pacific Ocean from 4 to 2 Ma (Leg 138, Sites 849 and 852. Proc. ODP, Sci. Results, 138. Ocean Drill. Prog., College Station, TX, pp. 163 176.
Jansen, E.. Sjoholm, J., 1991. Reconstruction of glaciation over the past 6 Myr from ice-borne deposits in the Norwegian Sea. Nature (London) 349, 600 603.
Karlin, K., Ruddiman, W.F. Mclntyre. A., 1989. Comparison of Late Pliocene and Late Pleistocene sea-surl:ace temper- atures in the Equatorial Atlantic divergence. Proc. ODP, Sci. Results, 108. Ocean Drill. Prog., College Station, TX, pp. 187 210.
Keller. G., Zenker, C.E.. Stone, S.M., 1989. Late Neogene his- tory of the Pacific Caribbean gateway. J. S.Am. Earth Sci. 2, 73 108.
Kemp, A.E.S.. 1995. Neogene and Quaternary pelagic sedi- ments and depositional history of the eastern equatorial Pacific Ocean ( Leg 138). Proc,ODP. Sci. Results, 138. Ocean Drill. Prog., College Station. TX, pp. 627 64(I.
Levitus, S., 1982. Climatological Atlas of the World Ocean. U.S. Dep. Commer., Natl. Ocean. Atmos. Adm., Washing- ton, DC, xiii + 173 pp.
Moore, T.C., Heath, R.C., Konsman, R.O., 1973. Biogenic sed- iments of the Panama Basin. J. Geol. 81,458 472.

M.R. (71apman, A.J. Chepstow-Lusty / Palaeogeography, Palaeoclimatology, Palaeoecology 134 (1997) 109 125 125
Murray, D.W., Farrell, J.W., McKenna, V., 1995. Biogenic sed- imentation at Site 847, eastern equatorial Pacific Ocean, during the past 3 m.y. Proc. ODP, Sci. Results, 138. Ocean Drill. Prog., College Station, TX, pp. 429 460.
Perch-Nielsen, K., 1985. Cenozoic calcareous nannofossils. In: Bolli, H.M., Saunders, J.B., Perch-Nielsen, K. (Eds.), Plank- ton Stratigraphy. Cambridge University Press, Cambridge, pp. 427-554.
Raffi, 1., Backman, J., Rio, D., Shackleton, N., 1993. Plio- Pleistocene nannofossil biostratigraphy and calibration to oxygen isotope stratigraphies from Deep Sea Drilling Project 607 and Ocean Drilling Program Site 677. Paleoceanography 8, 387 408.
Raymo, M.E., Ruddiman, W.F., Backman, J., Clement, B.M., Martinson, D.G., 1989. Late Pliocene variation in Northern Hemisphere ice sheets and North Atlantic deep water circula- tion. Paleoceanography 4, 413 446.
Roberts, D.G., Schnitker, D., et al. (Eds.), 1984. lnit. Rep. DSDP, 81. U.S. Gov. Print. Off., Washington, DC.
Ruddiman, W.F., Mclntyre, A., Raymo, M., 1987. Paleo- environmental results from North Atlantic Sites 607 and 609. lnit. Rep. DSDP 94, 855-878.
Ruddiman, W.F., Sarnthein, M., Baldauf, J. et al. (Eds.), 1988. lnit. Rep. ODP, 108. Ocean Drill. Prog., College Station, TX.
Sarnthein, M., Tiedemann, R., 1989. Toward a high-resolution isotope stratigraphy of the last 3.4 million years: Sites 658 and 659 off Northwest Africa. lnit. Rep. ODP 108, 167 185.
Shackleton, N.J., Hall, M.A., 1984. Oxygen and carbon isotope
stratigraphy of Deep Sea Drilling Project Hole 552A: Plio- Pleistocene glacial history, lnit. Rep. ODP 81,599 609.
Shackleton, N.J., Hall, M.A., 1990. Pliocene oxygen isotope stratigraphy of Hole 709C. Proc. ODP, Sci. Results 115, 529 538.
Shackleton, N.J. and Leg 81 Shipboard Scientific Party, 1984. Oxygen isotopic calibration of the onset of ice-rafting and history of glaciation in the North Atlantic Region. Nature (London) 307, 620-623.
Shackleton, N.J., Berger, A.L., Peltier, W.R., 1990. An alterna- tive astronomical calibration of the lower Pleistocene times- cale based on ODP Site 677. Trans. R. Soc. Edinburgh, Earth Sci. 81,251 261.
Shackleton, N.J., Hall, M.A., Pate, D., 1995. Pliocene stable isotope stratigraphy of Site 846. Proc. ODP, Sci. Results 138, 337-357.
Sikes, E.L., Keigwin, L.D., Curry, W.B., 1991. Pliocene paleo- ceanography: circulation and the oceanographic changes associated with the 2.4 Ma glacial event. Paleoceanography 6, 245 257.
Tiedemann, R., Sarnthein, M., Shackleton, N.J., 1994. Astronomic timescale for the Pliocene Atlantic 6180 and dust flux records of ODP Site 659. Paleoceanography 9, 619 638.
Wefer, G., Fischer, G., 1993. Seasonal patterns of vertical par- ticle flux in equatorial and coastal upwelling areas of the eastern Atlantic. Deep-Sea Res. 40, 1613 1645.
Wei, W., 1993. Calibration of Upper Pliocene Lower Pleisto- cene nannofossil events with oxygen isotope stratigraphy. Paleoceanography 8, 85 99.
Copyright © 2022 FDOKUMEN




















