
Kielin/chordin-like protein, a novel enhancer of BMP signaling, attenuates renal fibrotic disease
7
NATURE MEDICINE VOLUME 11 | NUMBER 4 | APRIL 2005 387 Kielin/chordin-like protein, a novel enhancer of BMP signaling, attenuates renal fibrotic disease Jingmei Lin 1 , Sanjeevkumar R Patel 2 , Xu Cheng 3 , Eun Ah Cho 1 , Inna Levitan 1 , Matthew Ullenbruch 1 , Sem H Phan 1 , John M Park 3 & Gregory R Dressler 1 The bone morphogenetic proteins (BMPs) profoundly affect embryonic development, differentiation and disease. BMP signaling is suppressed by cysteine-rich domain proteins, such as chordin, that sequester ligands from the BMP receptor. We describe a novel protein, KCP, with 18 cysteine-rich domains. Unlike chordin, KCP enhances BMP signaling in a paracrine manner. Smad1-dependent transcription and phosphorylated Smad1 (P-Smad1) levels are increased, as KCP binds to BMP7 and enhances binding to the type I receptor. In vivo, Kcp –/– mice are viable and fertile. Because BMPs have a pivotal role in renal disease, we examined the phenotype of Kcp –/– mice in two different models of renal injury. Kcp –/– animals show reduced levels of P-Smad1, are more susceptible to developing renal interstitial fibrosis, are more sensitive to tubular injury and show substantial pathology after recovery. The data indicate an important role for KCP in attenuating the pathology of renal fibrotic disease. In the mammalian kidney, BMP7 has an essential role in development and disease. BMP7-null mice show arrested renal development at around embryonic day (E)14.5, resulting in severe renal hypoplasia 1,2 . BMP7 is also an antifibrotic agent that can reduce interstitial fibrosis, a common pathology in a broad spectrum of chronic renal diseases. Administration of recombinant BMP7 has shown remarkable efficacy in the reduction of glomerular and interstitial fibrosis in mouse models of chronic renal disease 3,4 . BMPs bind to specific type I and type II transmembrane receptors that contain cytoplasmic serine-threonine kinase domains 5,6 . The acti- vated receptor complex then phosphorylates the intracellular Smad proteins, which translocate to the nucleus and activate ligand-respon- sive genes 7 . The regulation of BMP signaling by sequestering ligand availability is a fundamental morphogenetic mechanism during devel- opment that establishes the dorsal-ventral pattern in both invertebrates and vertebrates 8,9 . Numerous proteins such as noggin, chordin, short gastrulation (Sog), twisted gastrulation (Tsg), and their related factors caronte, cerberus, and gremlin bind BMP family ligands and prevent their contact with receptors 5,10 . Chordin and Sog are secreted pro- teins with repeated cysteine-rich domains that bind BMPs to inhibit signaling 11 . Here we describe an enhancer of BMP signaling. The protein is most homologous to Xenopus kielin 12 and contains multiple cysteine- rich domains. Thus, we have assigned the name KCP (kielin/chordin- like protein) to this protein. Unlike previously described cysteine-rich domain proteins, KCP is a potent paracrine enhancer of BMP signaling. KCP increases the affinity of ligand to receptor and/or enhances the sta- bility of the ligand-receptor complex. Given the role of BMP7 in renal disease, we analyzed the phenotypes of Kcp –/– mice in two independent models of renal injury. The data point to an important role of KCP in the enhancement of BMP signaling and attenuation of the initiation and progression of fibrotic disease after renal injury. RESULTS Identification and expression of KCP Overlapping cDNA clones were identified from an embryonic kidney library that contained multiple cysteine-rich domains. The entire coding region was similar to the Xenopus kielin protein 12 , thus we have named the protein KCP for kielin/chordin-like protein 1. The 1,254–amino acid KCP protein has a signal peptide, 18 cysteine-rich chordin repeats and a carboxyl-terminal von Willebrand factor type D domain. An alignment of the 18 cysteine-rich domains with the second cysteine- rich domain of mouse chordin shows conserved CXXCXC and CCXXC motifs (Fig. 1a). The mouse Kcp gene maps to chromosome 6, between 29.40 and 29.42 Mb. A single human homolog maps to chromosome 7q32.3, corresponding to a hypothetical kielin-like protein. Conditioned medium from the high KCP-expressing NIH 3T3 cell lines show that the protein could be secreted (Fig. 1b), although the levels are low relative to the lysate. Myc-epitope– and poly-His–tagged KCP could be isolated from conditioned medium by nickel affinity beads and detected with greater sensitivity (Fig. 1c). The Kcp expression pattern was determined by northern blot analysis (Fig. 1d) and whole-mount in situ hybridization (Fig. 2). Embryonic brain and kidney samples show a RNA band approximately 5.0 kilo- bases in size, although levels were low. Adult tissues were negative at this level of resolution. No cell lines tested showed detectable levels 1 Department of Pathology, University of Michigan, 1301 Catherine Street, Ann Arbor, Michigan 48109, USA. 2 Department of Internal Medicine, University of Michigan, 3110 Taubman, Ann Arbor, Michigan 48109, USA. 3 Department of Urology, University of Michigan, 1500 East Medical Center Drive, Ann Arbor, Michigan 48109, USA. Correspondence should be addressed to G.R.D. ([email protected]). Published online 27 March 2005; doi:10.1038/nm1217 ARTICLES © 2005 Nature Publishing Group http://www.nature.com/naturemedicine
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Kielin/chordin-like protein, a novel enhancer of BMP signaling, attenuates renal fibrotic disease

NATURE MEDICINE VOLUME 11 | NUMBER 4 | APRIL 2005 387
Kielin/chordin-like protein, a novel enhancer of BMP signaling, attenuates renal fibrotic diseaseJingmei Lin1, Sanjeevkumar R Patel2, Xu Cheng3, Eun Ah Cho1, Inna Levitan1, Matthew Ullenbruch1, Sem H Phan1, John M Park3 & Gregory R Dressler1
The bone morphogenetic proteins (BMPs) profoundly affect embryonic development, differentiation and disease. BMP signaling is suppressed by cysteine-rich domain proteins, such as chordin, that sequester ligands from the BMP receptor. We describe a novel protein, KCP, with 18 cysteine-rich domains. Unlike chordin, KCP enhances BMP signaling in a paracrine manner. Smad1-dependent transcription and phosphorylated Smad1 (P-Smad1) levels are increased, as KCP binds to BMP7 and enhances binding to the type I receptor. In vivo, Kcp–/– mice are viable and fertile. Because BMPs have a pivotal role in renal disease, we examined the phenotype of Kcp–/– mice in two different models of renal injury. Kcp–/– animals show reduced levels of P-Smad1, are more susceptible to developing renal interstitial fibrosis, are more sensitive to tubular injury and show substantial pathology after recovery. The data indicate an important role for KCP in attenuating the pathology of renal fibrotic disease.
In the mammalian kidney, BMP7 has an essential role in development and disease. BMP7-null mice show arrested renal development at around embryonic day (E)14.5, resulting in severe renal hypoplasia1,2. BMP7 is also an antifibrotic agent that can reduce interstitial fibrosis, a common pathology in a broad spectrum of chronic renal diseases. Administration of recombinant BMP7 has shown remarkable efficacy in the reduction of glomerular and interstitial fibrosis in mouse models of chronic renal disease3,4.
BMPs bind to specific type I and type II transmembrane receptors that contain cytoplasmic serine-threonine kinase domains5,6. The acti-vated receptor complex then phosphorylates the intracellular Smad proteins, which translocate to the nucleus and activate ligand-respon-sive genes7. The regulation of BMP signaling by sequestering ligand availability is a fundamental morphogenetic mechanism during devel-opment that establishes the dorsal-ventral pattern in both invertebrates and vertebrates8,9. Numerous proteins such as noggin, chordin, short gastrulation (Sog), twisted gastrulation (Tsg), and their related factors caronte, cerberus, and gremlin bind BMP family ligands and prevent their contact with receptors5,10. Chordin and Sog are secreted pro-teins with repeated cysteine-rich domains that bind BMPs to inhibit signaling11.
Here we describe an enhancer of BMP signaling. The protein is most homologous to Xenopus kielin12 and contains multiple cysteine-rich domains. Thus, we have assigned the name KCP (kielin/chordin-like protein) to this protein. Unlike previously described cysteine-rich domain proteins, KCP is a potent paracrine enhancer of BMP signaling. KCP increases the affinity of ligand to receptor and/or enhances the sta-bility of the ligand-receptor complex. Given the role of BMP7 in renal
disease, we analyzed the phenotypes of Kcp–/– mice in two independent models of renal injury. The data point to an important role of KCP in the enhancement of BMP signaling and attenuation of the initiation and progression of fibrotic disease after renal injury.
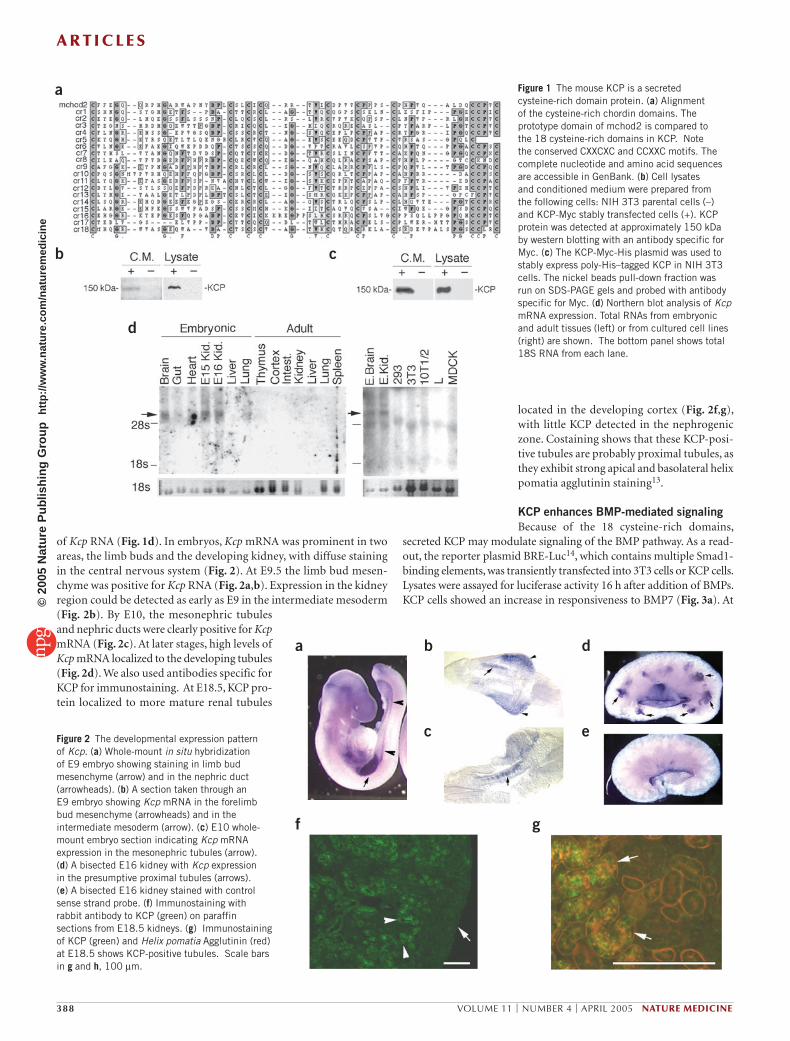
RESULTSIdentification and expression of KCPOverlapping cDNA clones were identified from an embryonic kidney library that contained multiple cysteine-rich domains. The entire coding region was similar to the Xenopus kielin protein12, thus we have named the protein KCP for kielin/chordin-like protein 1. The 1,254–amino acid KCP protein has a signal peptide, 18 cysteine-rich chordin repeats and a carboxyl-terminal von Willebrand factor type D domain. An alignment of the 18 cysteine-rich domains with the second cysteine-rich domain of mouse chordin shows conserved CXXCXC and CCXXC motifs (Fig. 1a). The mouse Kcp gene maps to chromosome 6, between 29.40 and 29.42 Mb. A single human homolog maps to chromosome 7q32.3, corresponding to a hypothetical kielin-like protein. Conditioned medium from the high KCP-expressing NIH 3T3 cell lines show that the protein could be secreted (Fig. 1b), although the levels are low relative to the lysate. Myc-epitope– and poly-His–tagged KCP could be isolated from conditioned medium by nickel affinity beads and detected with greater sensitivity (Fig. 1c).
The Kcp expression pattern was determined by northern blot analysis (Fig. 1d) and whole-mount in situ hybridization (Fig. 2). Embryonic brain and kidney samples show a RNA band approximately 5.0 kilo-bases in size, although levels were low. Adult tissues were negative at this level of resolution. No cell lines tested showed detectable levels
1Department of Pathology, University of Michigan, 1301 Catherine Street, Ann Arbor, Michigan 48109, USA. 2Department of Internal Medicine, University of Michigan, 3110 Taubman, Ann Arbor, Michigan 48109, USA. 3Department of Urology, University of Michigan, 1500 East Medical Center Drive, Ann Arbor, Michigan 48109, USA. Correspondence should be addressed to G.R.D. ([email protected]).
Published online 27 March 2005; doi:10.1038/nm1217
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e
388 VOLUME 11 | NUMBER 4 | APRIL 2005 NATURE MEDICINE
of Kcp RNA (Fig. 1d). In embryos, Kcp mRNA was prominent in two areas, the limb buds and the developing kidney, with diffuse staining in the central nervous system (Fig. 2). At E9.5 the limb bud mesen-chyme was positive for Kcp RNA (Fig. 2a,b). Expression in the kidney region could be detected as early as E9 in the intermediate mesoderm (Fig. 2b). By E10, the mesonephric tubules and nephric ducts were clearly positive for Kcp mRNA (Fig. 2c). At later stages, high levels of Kcp mRNA localized to the developing tubules (Fig. 2d). We also used antibodies specific for KCP for immunostaining. At E18.5, KCP pro-tein localized to more mature renal tubules
located in the developing cortex (Fig. 2f,g), with little KCP detected in the nephrogenic zone. Costaining shows that these KCP-posi-tive tubules are probably proximal tubules, as they exhibit strong apical and basolateral helix pomatia agglutinin staining13.
KCP enhances BMP-mediated signalingBecause of the 18 cysteine-rich domains,
secreted KCP may modulate signaling of the BMP pathway. As a read-out, the reporter plasmid BRE-Luc14, which contains multiple Smad1-binding elements, was transiently transfected into 3T3 cells or KCP cells. Lysates were assayed for luciferase activity 16 h after addition of BMPs. KCP cells showed an increase in responsiveness to BMP7 (Fig. 3a). At
a
b
d
c
Figure 1 The mouse KCP is a secreted cysteine-rich domain protein. (a) Alignment of the cysteine-rich chordin domains. The prototype domain of mchod2 is compared to the 18 cysteine-rich domains in KCP. Note the conserved CXXCXC and CCXXC motifs. The complete nucleotide and amino acid sequences are accessible in GenBank. (b) Cell lysates and conditioned medium were prepared from the following cells: NIH 3T3 parental cells (–) and KCP-Myc stably transfected cells (+). KCP protein was detected at approximately 150 kDa by western blotting with an antibody specific for Myc. (c) The KCP-Myc-His plasmid was used to stably express poly-His–tagged KCP in NIH 3T3 cells. The nickel beads pull-down fraction was run on SDS-PAGE gels and probed with antibody specific for Myc. (d) Northern blot analysis of Kcp mRNA expression. Total RNAs from embryonic and adult tissues (left) or from cultured cell lines (right) are shown. The bottom panel shows total 18S RNA from each lane.
a
f g
b
c
d
eFigure 2 The developmental expression pattern of Kcp. (a) Whole-mount in situ hybridization of E9 embryo showing staining in limb bud mesenchyme (arrow) and in the nephric duct (arrowheads). (b) A section taken through an E9 embryo showing Kcp mRNA in the forelimb bud mesenchyme (arrowheads) and in the intermediate mesoderm (arrow). (c) E10 whole-mount embryo section indicating Kcp mRNA expression in the mesonephric tubules (arrow). (d) A bisected E16 kidney with Kcp expression in the presumptive proximal tubules (arrows). (e) A bisected E16 kidney stained with control sense strand probe. (f) Immunostaining with rabbit antibody to KCP (green) on paraffin sections from E18.5 kidneys. (g) Immunostaining of KCP (green) and Helix pomatia Agglutinin (red) at E18.5 shows KCP-positive tubules. Scale bars in g and h, 100 µm.
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e

NATURE MEDICINE VOLUME 11 | NUMBER 4 | APRIL 2005 389
50 ng/ml BMP7, 3T3 cells showed 10-fold activation of BRE-Luc, whereas KCP-expressing cells activated BRE-Luc more than 150-fold. At 100 ng/ml BMP7, KCP-expressing cells activated BRE-Luc 500-fold, compared to 50-fold for the parental 3T3 cells. Similar results were obtained with BMP4 (Fig. 3b).
We developed two paracrine systems to examine KCP function. First, BRE-Luc was transfected into 3T3 cells, then cells were seeded onto either 3T3 cells or KCP cells. After coculture for 24 h, increas-ing amounts of BMP7 were added and luciferase activity measured (Fig. 3c). The BRE-Luc cells cocultured with KCP cells showed a three-fold enhanced response to BMP7 at 50 ng/ml, compared to coculture with 3T3 cells. Thus, KCP function is mediated, at least in part, by secreted protein. Conditioned medium from KCP cells also enhanced BMP-dependent signaling approximately fivefold (Fig. 3d). These experiments negate any potential differences resulting from clonal variation in the reporter cells.
The enhanced response of KCP expression to BMP7 was confirmed by western blotting with antibodies for P-Smad1 (Fig. 3e). One hour after BMP addition, KCP cells showed higher amounts of P-Smad1 at 10 ng/ml BMP7 (Fig. 3e). At 50 and 100 ng/ml of BMP7, higher amounts of P-Smad1 were also observed in KCP cells compared to con-trols. Twenty-four hours after BMP7 addition, increased P-Smad1 levels were maintained in KCP cells, at high concentrations of BMP7 (Fig. 3e). Thus, KCP sensitizes cells to BMPs and maintains the response.
KCP might form a complex with the secreted ligands and alter the affinity to receptors. But the size and cysteine-rich nature of KCP made it difficult to generate soluble protein in bacteria. Alternatively, conditioned medium from KCP and control cells was incubated with nickel-affinity beads, washed, and incubated with BMP7 (Fig. 3f). The bound proteins were analyzed for KCP and BMP7 by western blotting. BMP7 was bound to beads after preincubation with KCP-conditioned medium, but not with control medium. Furthermore, BMPR-IA, fused to the Fc portion of the human IgG, was incubated with BMP7 in the presence or absence of KCP-conditioned or control medium (Fig. 3g). The BMPR-IA protein was immobilized on protein A agarose and bound BMP7 and KCP were analyzed by western blotting (Fig. 3g). The results show a clear increase in the amounts of BMP7 bound to the recombinant receptor in KCP-conditioned medium. Furthermore, KCP
could be detected in the bound fraction. Thus, enhancement of BMP signaling may be the result of an increased receptor-ligand interactions mediated by KCP.
Renal fibrosis in Kcp mutantsTo understand the in vivo function of KCP, we deleted exons 2–21 (encoding amino acids 67–774) of the Kcp locus by homologous recom-bination in the mouse germline (Fig. 4a). Homozygous Kcp–/– mutants were viable and fertile and had no gross phenotypic abnormalities. The lack of phenotype in the hybrid genetic background (C57bl6:129) sug-gests that BMP signaling levels are sufficient such that any potential enhancement by KCP during development is redundant.
BMPs are also known to function in the response to renal injury3,4,15,16. We utilized both wild-type and Kcp–/– littermates in the unilateral ure-teral obstruction (UUO) model of fibrosis17,18. Obstruction by surgical ligation of the ureter leads to a rapid development of interstitial fibro-sis within 4–7 d. The contralateral, or unobstructed, kidney does not develop considerable pathology. Kidneys from mice at 1, 2 and 4 weeks after UUO were examined for Kcp expression, BMP signaling and inter-stitial fibrosis. RNA analysis showed Kcp activation in both obstructed and contralateral kidneys from normal mice, whereas untreated and homozygous null animals did not express Kcp mRNA (Fig. 4b,c). We observed a considerable reduction of P-Smad1 levels in Kcp–/– mutants (Fig. 4d). The contralateral kidneys from wild-type mice had the high-est levels of P-Smad1, indicating a response to stress in this model. We also observed P-Smad1 was in the obstructed kidneys of wild-type mice, yet it was undetectable in the obstructed kidneys of Kcp–/– mice. These data are consistent with the ability of KCP to enhance BMP signaling through Smad1.
a b
c
e f
g
dFigure 3 KCP enhances BMP-mediated gene expression. (a) Dose response of BMP7 on activation of the BRE-Luc reporter in parental NIH 3T3 control cells (open squares) and KCP stable-transfected NIH 3T3 cells (filled squares). (b) Similar experiment as in a using BMP4 as an activator of BRE-Luc. (c) Paracrine assay for KCP activity. Control NIH 3T3 cells were transiently transfected with BRE-Luc reporter and seeded onto either the NIH 3T3 parental control cells (open squares) or KCP stably transfected cells (filled squares). BMP7 was added to the medium as indicated. (d) NIH 3T3 cells were transfected with BRE-Luc and incubated with either control cell (open squares) or KCP conditioned medium (filled squares) before stimulation with increasing amounts of BMP7. The results are average, normalized values from triplicates. The error bars represent one standard deviation from the mean (*P < 0.05). (e) Western blots of lysates from parental NIH 3T3 cells or KCP stably transfected cells treated with increasing amounts of BMP7 for 1 h or 24 h. Blots were probed with antibodies against P-Smad1, Smad1 and tubulin. (f) Nickel-affinity pull-down experiment using conditioned media from control cells or KCP-Myc-His–tagged-expressing cells. Affinity beads were preincubated with conditioned medium, washed and incubated with (+) or without (–) BMP7. BMP7 and KCP bound to the affinity beads were analyzed by western blotting. (g) BMP7 and recombinant BMP type I receptor (ALK3) were mixed in control cell or KCP-conditioned medium. Proteins were pulled down with Protein-A agarose and analyzed by western blotting. Lane 1, control lane with 10 ng BMP7; lanes 2–4, pull-down fractions.
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e

390 VOLUME 11 | NUMBER 4 | APRIL 2005 NATURE MEDICINE
The obstructed and contralateral kidneys were assessed for interstitial fibrosis. Smooth muscle actin (SMA) is a marker of interstitial area. The fibroblast-specific marker FSP1 (ref. 19) was also used to quan-tify the increased number of fibroblasts in response to obstruction. After 7 or 13 d, Kcp–/– obstructed kidneys showed a twofold increase in FSP1-positive cells compared to the wild-type obstructed kidneys at similar stages (Fig. 5a,d). Contralateral kidneys showed no differ-ence at 7 d, but Kcp–/– contralateral kidneys did show a considerable twofold increase in FSP1-positive cells after 13 d (Fig. 5b,d). We cal-culated the area and intensity of SMA staining (Fig. 5e). After 7 d, obstructed Kcp–/– kidneys showed a twofold increase in interstitial area (10.6 ± 2.9%) compared to the wild-type kidneys (5.0 ± 1.7%). After 13 d, the obstructed Kcp–/– kidneys had, on average, 67% more inter-stitial area (13.4 ± 1.6%) compared the wild-type kidneys (8.0 ± 1.4%). The intensity of SMA staining at 13 d after obstruction was significantly (P < 0.05) higher in Kcp–/– kidneys (205.6 ± 10) compared to wild-type kidneys (142 ± 19.8). The expression of the mRNA encoding collagen I was increased in Kcp–/– obstructed kidneys at 7 and 28 d (Fig. 5f) and in the contralateral Kcp–/– kidneys after 28 d (Fig. 5f). Total collagen con-tent was measured by hydroxyproline assays (Fig. 5g), which indicated a 30% increase after 28 d in the Kcp–/– obstructed kidneys compared to the wild-type kidneys.
Fibrosis in the contralateral, unobstructed kidneys was also seen in Kcp–/– mutants. After 13 d, the Kcp–/– contralateral kidneys have more interstitial myofibroblasts and show localized areas of SMA within the tubular interstitium (Fig. 5b) compared to wild-type kidneys. After 28 d, the contralateral kidneys of Kcp–/– mice show evidence of localized fibrosis in cortical tubular interstitium and in periglomerular regions (Fig. 5c). No evidence of fibrosis was observed in the contralateral kid-neys of wild-type mice. The data indicate that Kcp–/– mice show an enhanced fibrotic response in obstructed kidneys and a novel fibrotic response in the unobstructed, contralateral kidneys. Both obstructed and contralateral kidneys from Kcp–/– mice show reduced levels of
Smad1 phosphorylation, consistent with the ability of KCP protein to enhance BMP signaling.
Recovery from acute tubular injuryThe role of BMP7 in recovery from acute tubular necrosis (ATN) has been well documented16,20. Thus, we examined the ability of Kcp–/– mice to recover from toxic injury using the folic acid model of ATN. Mice were given a single dose of folic acid, observed over time and killed at 7, 14 or 28 d after injection. Within 24 h of injection, mice underwent an acute phase of renal failure characterized by oligourea, lethargy and fur ruffling. At the prescribed doses of folic acid (250 mg/kg), renal func-tion generally recovered within 7 d as proximal tubule cells regenerated. In wild-type mice, mortality after folic acid injection was less than 2% (n = 56). But in Kcp–/– mutants more than 23% (n = 82) of the injected animals died within the first 2 d. By 7 d after injection, wild-type kidneys had normal histological appearance, indicating near-complete recovery, whereas Kcp–/– mutants showed cortical cysts and extracellular matrix deposition (Fig. 6a,b). After 28 d of recovery, substantial scarring was seen in Kcp–/– mutants (Fig. 6c,d), with interstitial collagen deposi-tion around tubules and in focal regions devoid of tubules. Interstitial SMA-positive myofibroblasts and FSP1-positive fibroblasts were found throughout all Kcp–/– kidneys 28 d after injection, whereas little evidence of interstitial volume increase or fibrosis was observed in wild-type ani-mals (Fig. 6e,f). These data indicate a role for KCP protein in mediating the response to acute renal injury.
DISCUSSIONMammalian KCP is a secreted cysteine-rich domain protein that binds BMPs to enhance, rather than suppress, signaling. In both vertebrates and flies, secreted cysteine-rich domain proteins are known to regulate growth factor signaling in the extracellular space10. Like other cyste-ine-rich proteins11,21, KCP has similar spacing of the cysteines in the CXXCXC and CCXXC motifs and can physically interact with the active ligands. Most similar to KCP is Xenopus kielin12, which contains a signal peptide and 27 repeated cysteine-rich domains. In oocytes, injection of mRNA encoding kielin activates some dorsal markers. But unlike chordin, kielin activity is not sufficient to neuralize the ectoderm in the animal cap assay, suggesting that it is not simply a BMP inhibitor12.
KCP activity is paracrine and enhances expression of Smad1-depen-dent luciferase 10-fold. KCP-expressing cells were more responsive at low BMP concentrations and maintained the response over longer peri-ods of time. The CTGF protein binds to both BMPs and transform-ing growth factor (TGF)-βs, but it inhibits BMP-mediated signaling while enhancing the TGF-β response21. The vertebrate homologs of Drosophila cross-veinless-2, called BMPER in mouse or Cv-2 in chick-ens, contain 5 cysteine-rich domains and a carboxyl-terminal von Willebrand factor type D domain, similar to KCP. BMPER antagonizes
a
b
c
d
Figure 4 Generation and analysis of Kcp–/– mutants. (a) Schematic diagram of the Kcp targeting vector that was designed to delete exons 2–21, spanning amino acids 67–774 of the coding region. The lacZ gene was inserted in frame after amino acid 67 in exon 2. (b) Northern blot of poly-A RNA selected from 40 µg total RNA and probed with a Kcp-specific fragment. Controls are from untreated age-matched animals. E17K, embryonic kidney RNA; C, RNA from contralateral kidneys; U, RNA from the obstructed kidneys. After 7 d exposure, blots were stripped and hybridized with GAPDH. (c) RT-PCR using primers specific for Kcp and Gapd. Kcp was expressed in all wild-type contralateral (C) and obstructed (U) kidneys, but not in mutant and untreated animals. (d) Western blot analysis of P-Smad1 in wild-type and Kcp–/– mutant kidneys from obstructed (UUO) and contralateral (C) sides. Duplicate gels were probed with antibodies specific for P-Smad1 and Smad1.
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e

NATURE MEDICINE VOLUME 11 | NUMBER 4 | APRIL 2005 391
BMP signaling22, whereas chick Cv-2 antago-nizes or enhances BMP signaling, depending on the assay23. Thus, cysteine-rich domain proteins can differentially modulate signals received from the TGF-β superfamily.
In the developing kidney, BMP4 and BMP7 regulate branching mor-phogenesis and growth1,2,24. BMP7 is also abundant in adult tubular epithelial cells, collecting ducts, cortical interstitial cells and podo-cytes16,25. Despite the essential role of BMP7 development, we did not see effects of Kcp loss of function in the genetic background used. Whether different genetic backgrounds or a reduction in BMP7 gene dosage may unmask a phenotype in development remains to be deter-mined. In any event, the induction of Kcp expression in ATN and UUO models and the phenotypes of Kcp–/– mutants argues for a function in adult kidney epithelia, particularly in response to stress.
Tubular interstitial fibrosis is a common denominator of chronic renal disease. Renal fibrosis can be stimulated by obstruction, genetic mutations, chronic hypertension or diabetes. TGF-β is thought to be a profibrotic stimulus26,27, whereas BMP7 can attenuate the degree of fibrosis in experimental animal models4,15. TGF-β may stimulate the epithelial-to-mesenchyme transition, as has been observed in vitro and in vivo28–30, whereas BMP7 can reverse the this transition3. The antifibrotic functions of BMP7 could be compounded by the suppres-sion of proinflammatory cytokines31. In the UUO model, activation of Smad1 phosphorylation is indicative of BMP signaling. Thus, reduc-ing the effects of BMP signaling and limiting Smad1 activity should enhance the fibrotic response. This is precisely what we observe in the obstructed kidneys of Kcp–/– mutant mice. Even more surprising is the onset of a fibrotic response in the unobstructed, contralateral kid-neys of Kcp–/– mutant mice after 2 weeks, as no such phenotype had been reported to date. That P-Smad1 levels rise substantially in the contralateral kidneys indicates a level of stress that may be the result of increased glomerular filtration, as only one kidney remains functional, or circulating cytokines generated by an accompanying inflammatory response in the obstructed kidney.
BMP7 is also thought to be involved in regeneration after ischemic or toxic injury16,20. In the folic acid model of ATN, Kcp mutants are more susceptible to injury, have higher mortality, and show consider-able scarring after recovery. Recovery from ATN is complex and begins within 8 h of the insult32,33. In the ischemia model of ATN, BMP7 expression decreases in damaged tubules20 and recombinant BMP7 enhances recovery16. The effects of BMP7 in both the UUO and ATN models may be protective such that fewer epithelial cells die within the first 24-h period. Alternatively, BMP signaling may help to stimulate the regeneration of epithelial cells in the ATN models, recapitulating a development program, while preventing the epithelial-mesenchy-mal transition in the UUO model. In either case, the ability of KCP to enhance BMP activity may be an important mechanism to attenuate the effects of such renal injuries.
The secreted KCP protein is a potent, paracrine enhancer of BMP signaling. KCP is activated in response to renal stresses, such as obstruction or nephrotoxic shock, where it may function as a posi-tive mediator of BMP signaling and impact the ability of renal epithe-lial cells to recover from injury. Because KCP is a large, cysteine-rich protein, we have not been able to purify soluble, recombinant KCP to test its efficacy as an antifibrotic agent directly in experimental models. Given the sensitivity of Kcp–/– mutants to fibrosis and its novel BMP enhancing effects, KCP’s potential as a therapeutic agent should be explored.
METHODSDNA clones. The full-length coding sequence of KCP was inserted into a pCMV plasmid containing a Myc epitope tag at the carboxyl-terminal end and the neo-mycin resistance gene. To increase the secretion efficiency for conditioned media, the full-length KCP was also inserted into the PsecTag B plasmid (Invitrogen).
a
c d e
f g
bFigure 5 Analysis of obstructed and contralateral kidneys. Obstructed kidney sections (a) and contralateral kidney sections (b) were immunostained with antibodies to FSP1 (green) and SMA (red). Genotypes and number of days after obstruction are indicated. (c) Trichrome staining of sections from contralateral kidneys 28 d after obstruction. Genotypes and number of days after obstruction are indicated. Cross-linked collagen in the cortical interstitium and in the periglomerular regions of KCP mutant kidneys appears blue. Scale bars, 100 µm. Representative sections are shown with all exposures manually set at equal times. (d) Quantification of FSP1-positive interstitial fibroblasts. C, contralateral, unobstructed kidneys; UUO, obstructed kidneys. (e) Morphometric analysis of interstitial area as determined by the relative area of SMA staining. Averages of ten sections per data set are shown. (f) Levels of mRNA encoding collagen were measured by northern blotting with a ColA1A and GAPDH probes. The ratio of ColA1A to GAPDH is expressed in relative units. (g) Hydroxyproline content was determined by a colorimetric assay after acid hydrolysis and expressed as a percentage of dry tissue weight. All statistically significant differences (P < 0.05) are marked (*).
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e

392 VOLUME 11 | NUMBER 4 | APRIL 2005 NATURE MEDICINE
Expression analysis. Whole-mount in situ hybridization was performed as described34. For immunostaining, antibodies specific for KCP were made against the carboxyl-terminal amino acids 1,201–1,254. We performed immunostaining using TSA Plus Fluorescence Enhancement (Perkin Elmer) according to the man-ufacturer’s protocol. Paraffin sections were unmasked with VectaLabs Antigen retrival system. We calculated the areas and intensity of staining with antibodies specific for SMA from ten images at fixed exposure times using Metamorph 5.0 (Universal Imaging Corp.).
RNAs were prepared with TRIZOL (Gibco BRL) following the manufacturer’s instructions. We performed electrophoresis on 10 µg total RNA in 1% agarose formaldehyde gels, blotted to Hybond-N membrane overnight and probed with a Kcp cDNA. RNA expression was quantified using a phosphoimager and normal-ized to levels of mouse Gapdh expression on the same filters.
Cell culture and activation assays. NIH 3T3 cells were cultured in Dulbecco Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum and penicillin/streptomycin. FuGene 6 transfections were performed as described in manufacturer’s protocols (Roche Molecular Biochemicals). NIH3T3 and KCP stably transfected cells were cotransfected with BRE-luc14 and CMV-β-gal to monitor transfection efficiency. We added BMP7 or BMP4 (R & D Systems) 24 h after transfection. We harvested cells 24 h after BMP addition and measured luciferase activity (Promega). Experiments were done in triplicate and the means ± s.d. are shown.
For stable transformants, we transfected cells with KCP expression plasmid, which carried the gene whose product confers neomycin resistance. Expressing clones were expanded; nonexpressing clones were kept as negative controls. For conditioned media, KCP stably transfected monolayers were washed with phos-phate-buffered saline and the medium was changed to serum-free DMEM with 1× Insulin-Transferrin-Selenium-X (ITS, Invitrogen Corp.) for 48 h. We centri-fuged the conditioned medium to remove debris and concentrated it 10-fold using Amicon Ultra-15 centrifugal filter. Control medium was collected from NIH 3T3 cells.
For paracrine assays, NIH 3T3 cells were transfected with BRE-luc and a β-gal plasmid, then 4 × 104 cells were plated into 12-well dishes containing 4 × 104 NIH 3T3 or KCP cells. After coculture, we added BMP7 and measured luciferase 24 h later. For conditioned media, NIH 3T3 cells were transfected with BRE-luc and β-gal plasmids and conditioned medium added 8 h before BMP7.
Protein assays. Equal amounts of protein in 150 mM sodium chloride, 1% NP-40, 50 mM Tris pH 8.0, and protease inhibitors cocktail (Roche Molecular Biochemicals) were separated on 8% sodium dodecyl sulfate gels, transferred to polyvinylidene fluoride membranes and immunoblotted by enhanced chemilu-minescence (Amersham Biosciences). We used antibodies specific for: Myc (9E10, BabCO, 1:2,000), BMP7 (R & D Systems, 1:500), phosphorylated Smad1 (Cell signaling, 1:500), Smad1 (Santa Cruz, 1:1,000) and tubulin (Sigma, 1:10,000).
Pull-down assays. The conditioned media were passed through the ProBond Ni-NTA agarose resin (Qiagen) and washed under native conditions. The beads were incubated with 5 ng/µl of BMP7 in phosphate-buffered saline (PBS) con-taining 1.0 mM Mg2+ and 1.3 mM Ca2+ overnight, washed with PBS three times, and eluted in 2× SDS-PAGE sample buffer. For the in vitro receptor-ligand assays, we incubated 100 µl of the concentrated conditioned medium with 4 ng/µl of BMP7 in PBS containing 1.0 mM Mg2+ and 1.3 mM Ca2+ overnight, then incu-bated with 10 ng/µl of BMPR-1A-Fc (R & D Systems) at room temperature for 1 h. The protein-A agarose was preblocked by serum-free DMEM with 1 mg/ml bovine serum albumin and then used to pull down the BMPR1A-Fc.
Mice. We introduced the targeting vector into R1 embryonic stem (ES) cells using electrophoresis and then performed positive and negative selection35. We screened approximately 1,000 clones by Southern blotting and identified more than 20 positive clones. After karyotyping, three ES cell lines were injected into host blastocysts and germline chimeric mice obtained. All animal protocols were approved by the University of Michigan Committee on Use and Care and of Animals.
UUO model. We anesthetized mice with an intraperitoneal injection of ketamine and xylozine. Through a midline abdominal incision, the right ureter was exposed and tied off at the mid-ureteral level with fine suture (4-0 silk) to induce a com-plete obstruction. We analyzed at least 4 wild-type and Kcp–/– mice at 7, 13 and 28 d. Kidneys were fixed in 4% paraformaldehyde and embedded in paraffin blocks. For RNA and hydroxyproline analysis, kidney samples were fresh frozen. We prepared RNAs in Trizol and performed poly-A selection using oligo-dT cellulose. Hydroxyproline assays were performed as described36.
ATN model. Age-matched mice were injected intraperitoneally with 250 mg/kg folic acid37 and killed at 7, 14 or 28 d after injury. We observed mice daily after injection and collected serum at the time of death. At least seven animals of each genotype were analyzed at each time point.
Accession and identification numbers. The GenBank accession number for KCP protein is AY884211, and the LocusLink identification number for a human homolog corresponding to a hypothetical kielin-like protein is 375616.
a b
c d
e f
Figure 6 Recovery from acute tubular injury. Wild-type (a,c,e) and Kcp–/– mutants (b,d,f) were examined 7 (a, b) or 28 d (c–f) after folic acid injection. (a,b) Trichrome staining at 7 d after injury typically show cortical cysts (arrows) in Kcp–/– mutant kidneys and multiple focal areas of collagen deposition (arrowheads), whereas wild-type kidneys have near-normal histology. (c,d) Trichrome staining at 28 d after injury. Areas of tubular effacement with extensive collagen deposition were observed in Kcp–/– mutants extending from the cortex to the periphery of the kidney (arrows). Interstitial collagen deposition (arrowheads) was widespread from the cortex to the cortico-medullary boundary in Kcp–/– mutants. Some remaining cysts (*) at this time are devoid of epithelial cells, leaving only the basement membrane. (e,f) Immunostaining with antibodies specific for FSP1 (green) and SMA (red) also indicate extensive accumulation of myofibroblasts (arrows) and FSP1-positive fibroblasts (arrowheads) within the interstitium of Kcp–/– mutants 28 d after injury. Little evidence of fibrosis was observed in wild-type kidneys at this time. Nuclei were stained with DAPI (blue, e, f only) and scale bars, 100 µm.
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e

NATURE MEDICINE VOLUME 11 | NUMBER 4 | APRIL 2005 393
ACKNOWLEDGMENTSWe thank P. ten Dijke for the BRE-Luc reporter, E. Neilson for the anti-FSP1 antibody, A. Eddy for advice with the UUO model, and R. Wiggins, E. Fearon and K. Cadigan for reading and discussions. Gene targeting was done in the University of Michigan Transgenic Animal Model Core with the assistance of E. Hughes and T. Saunders. This work is supported by US National Institutes of Health grants DK062914 and PKD Grant-in-Aid to G.R.D., DK02803 to S.R.P., and HL52285 to S.H.P.
COMPETING INTERESTS STATEMENTThe authors declare that they have no competing financial interests.
Received 18 October 2004; accepted 31 January 2005Published online at http://www.nature.com/naturemedicine/
1. Dudley, A.T., Lyons, K.M. & Robertson, E.J. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 9, 2795–2807 (1995).
2. Luo, G. et al. BMP-7 is an inducer of nephrogenesis, and is also required for eye development and skeletal patterning. Genes Dev. 9, 2808–2820 (1995).
3. Zeisberg, M. et al. BMP-7 counteracts TGF-beta1-induced epithelial-to-mesenchymal transition and reverses chronic renal injury. Nat. Med. 9, 964–968 (2003).
4. Zeisberg, M. et al. Bone morphogenic protein-7 inhibits progression of chronic renal fibrosis associated with two genetic mouse models. Am. J. Physiol. Renal. Physiol. 285, F1060–F1067 (2003).
5. Shi, Y. & Massague, J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell 113, 685–700 (2003).
6. Zwijsen, A., Verschueren, K. & Huylebroeck, D. New intracellular components of bone morphogenetic protein/Smad signaling cascades. FEBS Lett. 546, 133–139 (2003).
7. Nishimura, R. et al. The role of Smads in BMP signaling. Front. Biosci. 8, s275–s284 (2003).
8. Capdevila, J. & Belmonte, J.C. Extracellular modulation of the Hedgehog, Wnt and TGF-beta signalling pathways during embryonic development. Curr. Opin. Genet. Dev. 9, 427–433 (1999).
9. Christian, J.L. BMP, Wnt and Hedgehog signals: how far can they go? Curr. Opin. Cell Biol. 12, 244–249 (2000).
10. Garcia Abreu, J., Coffinier, C., Larrain, J., Oelgeschlager, M. & De Robertis, E.M. Chordin-like CR domains and the regulation of evolutionarily conserved extracellular signaling systems. Gene 287, 39–47 (2002).
11. Larrain, J. et al. BMP-binding modules in chordin: a model for signalling regulation in the extracellular space. Development 127, 821–830 (2000).
12. Matsui, M., Mizuseki, K., Nakatani, J., Nakanishi, S. & Sasai, Y. Xenopus kielin: A dorsalizing factor containing multiple chordin-type repeats secreted from the embry-onic midline. Proc. Natl. Acad. Sci. USA 97, 5291–5296 (2000).
13. Laitinen, L., Vitanen, I. & Saxen, L. Changes in the glycosylation pattern dur-ing embronic development of mouse kidney as revealed with lectin conjugates. J. Histochem. Cytochem. 35, 55–65 (1987).
14. Korchynskyi, O. & ten Dijke, P. Identification and functional characterization of distinct critically important bone morphogenetic protein-specific response elements in the Id1 promoter. J. Biol. Chem. 277, 4883–4891 (2002).
15. Hruska, K.A. et al. Osteogenic protein-1 prevents renal fibrogenesis associated with
ureteral obstruction. Am. J. Physiol. Renal. Physiol. 279, F130–F143 (2000).16. Vukicevic, S. et al. Osteogenic protein-1 (bone morphogenetic protein-7) reduces
severity of injury after ischemic acute renal failure in rat. J. Clin. Invest. 102, 202–214 (1998).
17. Chevalier, R.L. Molecular and cellular pathophysiology of obstructive nephropathy. Pediatr. Nephrol. 13, 612–619 (1999).
18. Klahr, S. & Morrissey, J. Obstructive nephropathy and renal fibrosis: The role of bone morphogenic protein-7 and hepatocyte growth factor. Kidney Int. Suppl. S105–S112 (2003).
19. Strutz, F. et al. Identification and characterization of a fibroblast marker: FSP1. J. Cell Biol. 130, 393–405 (1995).
20. Almanzar, M.M. et al. Osteogenic protein-1 mRNA expression is selectively modulated after acute ischemic renal injury. J. Am. Soc. Nephrol. 9, 1456–1463 (1998).
21. Abreu, J.G., Ketpura, N.I., Reversade, B. & De Robertis, E.M. Connective-tissue growth factor (CTGF) modulates cell signalling by BMP and TGF-beta. Nat. Cell Biol. 4, 599–604 (2002).
22. Moser, M. et al. BMPER, a novel endothelial cell precursor-derived protein, antagonizes bone morphogenetic protein signaling and endothelial cell differentiation. Mol. Cell. Biol. 23, 5664–5679 (2003).
23. Coles, E., Christiansen, J., Economou, A., Bronner-Fraser, M. & Wilkinson, D.G. A vertebrate crossveinless 2 homologue modulates BMP activity and neural crest cell migration. Development 131, 5309–5317 (2004).
24. Miyazaki, Y., Oshima, K., Fogo, A., Hogan, B.L. & Ichikawa, I. Bone morphogenetic protein 4 regulates the budding site and elongation of the mouse ureter. J. Clin. Invest. 105, 863–873 (2000).
25. Kopp, J.B. BMP receptors in kidney. Kidney Int. 58, 2237-2238 (2000).26. Schnaper, H.W., Hayashida, T., Hubchak, S.C. & Poncelet, A.C. TGF-beta signal
transduction and mesangial cell fibrogenesis. Am. J. Physiol. Renal. Physiol. 284, F243–F252 (2003).
27. Kopp, J.B. et al. Transgenic mice with increased plasma levels of TGF-beta 1 develop progressive renal disease. Lab. Invest. 74, 991–1003 (1996).
28. Herzlinger, D. Renal interstitial fibrosis: remembrance of things past? J. Clin. Invest. 110, 305–306 (2002).
29. Iwano, M. et al. Evidence that fibroblasts derive from epithelium during tissue fibrosis. J. Clin. Invest. 110, 341–350 (2002).
30. Kalluri, R. & Neilson, E.G. Epithelial-mesenchymal transition and its implications for fibrosis. J. Clin. Invest. 112, 1776–1784 (2003).
31. Gould, S.E., Day, M., Jones, S.S. & Dorai, H. BMP-7 regulates chemokine, cytokine, and hemodynamic gene expression in proximal tubule cells. Kidney Int. 61, 51–60 (2002).
32. Nony, P.A. & Schnellmann, R.G. Mechanisms of renal cell repair and regeneration after acute renal failure. J. Pharmacol. Exp. Ther. 304, 905–912 (2003).
33. Safirstein, R. Renal regeneration: reiterating a developmental paradigm. Kidney Int. 56, 1599–1600. (1999).
34. Brophy, P.D., Ostrom, L., Lang, K.M. & Dressler, G.R. Regulation of ureteric bud outgrowth by Pax2-dependent activation of the glial derived neurotrophic factor gene. Development 128, 4747–4756 (2001).
35. Joyner, A.L. Gene Targeting: A Practical Approach 33–61 (Oxford Univ Press, Oxford, 1993).
36. Phan, S.H., Thrall, R.S. & Ward, P.A. Bleomycin-induced pulmonary fibrosis in rats: biochemical demonstration of increased rate of collagen synthesis. Am. Rev. Respir. Dis. 121, 501–506 (1980).
37. Imgrund, M. et al. Re-expression of the developmental gene Pax-2 during experimental acute tubular necrosis in mice 1. Kidney Int. 56, 1423–1431 (1999).
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
emed
icin
e




















