
Jones 2009
11
929 New Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43: 929–939 0028–8330/09/4304–0929 © The Royal Society of New Zealand 2009 M08017; Online publication date 31 July 2009 Received 21 April 2008; accepted 12 June 2009 Diets of eight fish species from the upper slope off the Wairarapa coast, North Island, New Zealand, with notes on the diets of others M. R. L. JoNeS earth and oceanic Sciences Research Institute Auckland University of Technology Private Bag 92 006 Auckland, New Zealand email: [email protected] Abstract The diets of eight demersal fish species from the upper continental slope (c. 240–450 m) were determined from samples taken during late January and early February 2004, off the Wairarapa coast, North Island, New Zealand. Diets were from a combination of benthopelagic and benthic sources, with most species exhibiting ontogenetic shifts in diet, in that larger-bodied food (fish and/or natant decapod prawns) was more important in the diets of larger fish. Javelinfish (Lepidorhynchus denticulatus ), silver roughy ( Hoplostethus mediterraneus), and capro dory ( Capromimus abbreviatus) had predominantly benthopelagic diets, whereas the diets of sea perch (Helicolenus percoides), Bollons’ rattail (Caelorinchus bollonsi), two-banded rattail (C. biclinozonalis), and silverside (Argentina elongata) were predominantly benthic, with the crab Carcinoplax victoriensis an important food item. The diet of oliver’s rattail (C. oliverianus) was a mix of benthopelagic and benthic organisms. Generally, levels of dietary overlap between the eight species were low. overall, the diets of these fish probably reflect regional and seasonal levels of food availability. Notes on food observed in the stomachs of a further 18 species, for which there were fewer than 10 stomach samples per species are also provided. Keywords fish diet; scampi; javelinfish; silver roughy; capro dory; sea perch; rattails INTRODUCTION The Wairarapa region in the southeast of New Zealand’s North Island (Fig. 1) is an area with a relatively steep continental slope (Pantin 1963), on which there is a commercial trawl fishery for scampi, Metanephrops challengeri Balss, 1914, a small-clawed lobster abundant around New Zealand at depths of 200–500 m (Cryer & Stotter 1999; Smith 1999). Trawling for scampi produces considerable bycatch (non-target species), in amounts consistent with prawn trawl fisheries elsewhere (Andrew & Pepperell 1992; evans et al. 1994; Castro et al. 2005). The non-commercial fraction is not usually retained (Cryer et al. 2002; Hartill et al. 2006), so that the biology and hence the feeding relationships between bycatch species are poorly known. The benthos has been shown to be an important source of food for shallow water and upper slope fish elsewhere (e.g., Bulman et al. 2001; Fosså et al. 2002; Carbines et al. 2004). In Australian waters, Bulman et al. (2001) investigated the trophic ecology of shelf and upper slope fish species off New South Wales, reporting over half the species to be predominantly benthic feeders. Carbines et al. (2004) found that blue cod (Parapercis colias) from Foveaux Strait, South Island, New Zealand were 20% larger in areas with complex biogenic reefs than areas which had been previously trawled. In the Northern Hemisphere in Norwegian waters, Fosså et al. (2002) found that between 30 and 50% of the reefs of the deep water coral Lophelia pertusa had been impacted by trawling and local fishers now avoided these areas, perceiving them to be less productive fishing grounds. In an era when ecosystem-based fisheries management is becoming more prevalent, understanding the interactions between fish species and the benthic environment has become more important (Pikitch et al. 2004; Aydin & Mueter 2007; McShane et al. 2007; Smith et al. 2007). Research on non-commercial upper slope fish assemblages in New Zealand waters is mostly limited to interpretations of historical presence/ absence trawl survey data (e.g., Anderson et al. 1998;
Transcript of Jones 2009

929Jones—Diets of upper-slope fish from Wairarapa, New ZealandNew Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43: 929–9390028–8330/09/4304–0929 © The Royal Society of New Zealand 2009
M08017; Online publication date 31 July 2009 Received 21 April 2008; accepted 12 June 2009
Diets of eight fish species from the upper slope off the Wairarapa coast, North Island, New Zealand, with notes on the diets of others
M. R. L. JoNeSearth and oceanic Sciences Research InstituteAuckland University of TechnologyPrivate Bag 92 006Auckland, New Zealandemail: [email protected]
Abstract The diets of eight demersal fish species from the upper continental slope (c. 240–450 m) were determined from samples taken during late January and early February 2004, off the Wairarapa coast, North Island, New Zealand. Diets were from a combination of benthopelagic and benthic sources, with most species exhibiting ontogenetic shifts in diet, in that larger-bodied food (fish and/or natant decapod prawns) was more important in the diets of larger fish. Javelinfish (Lepidorhynchus denticulatus), silver roughy (Hoplostethus mediterraneus), and capro dory (Capromimus abbreviatus) had predominantly benthopelagic diets, whereas the diets of sea perch (Helicolenus percoides), Bollons’ rattail (Caelorinchus bollonsi), two-banded rattail (C. biclinozonalis), and silverside (Argentina elongata) were predominantly benthic, with the crab Carcinoplax victoriensis an important food item. The diet of oliver’s rattail (C. oliverianus) was a mix of benthopelagic and benthic organisms. Generally, levels of dietary overlap between the eight species were low. overall, the diets of these fish probably reflect regional and seasonal levels of food availability. Notes on food observed in the stomachs of a further 18 species, for which there were fewer than 10 stomach samples per species are also provided.
Keywords fish diet; scampi; javelinfish; silver roughy; capro dory; sea perch; rattails
IntroductIon
The Wairarapa region in the southeast of New Zealand’s North Island (Fig. 1) is an area with a relatively steep continental slope (Pantin 1963), on which there is a commercial trawl fishery for scampi, Metanephrops challengeri Balss, 1914, a small-clawed lobster abundant around New Zealand at depths of 200–500 m (Cryer & Stotter 1999; Smith 1999). Trawling for scampi produces considerable bycatch (non-target species), in amounts consistent with prawn trawl fisheries elsewhere (Andrew & Pepperell 1992; evans et al. 1994; Castro et al. 2005). The non-commercial fraction is not usually retained (Cryer et al. 2002; Hartill et al. 2006), so that the biology and hence the feeding relationships between bycatch species are poorly known. The benthos has been shown to be an important source of food for shallow water and upper slope fish elsewhere (e.g., Bulman et al. 2001; Fosså et al. 2002; Carbines et al. 2004). In Australian waters, Bulman et al. (2001) investigated the trophic ecology of shelf and upper slope fish species off New South Wales, reporting over half the species to be predominantly benthic feeders. Carbines et al. (2004) found that blue cod (Parapercis colias) from Foveaux Strait, South Island, New Zealand were 20% larger in areas with complex biogenic reefs than areas which had been previously trawled. In the Northern Hemisphere in Norwegian waters, Fosså et al. (2002) found that between 30 and 50% of the reefs of the deep water coral Lophelia pertusa had been impacted by trawling and local fishers now avoided these areas, perceiving them to be less productive fishing grounds. In an era when ecosystem-based fisheries management is becoming more prevalent, understanding the interactions between fish species and the benthic environment has become more important (Pikitch et al. 2004; Aydin & Mueter 2007; McShane et al. 2007; Smith et al. 2007). Research on non-commercial upper slope fish assemblages in New Zealand waters is mostly limited to interpretations of historical presence/absence trawl survey data (e.g., Anderson et al. 1998;

930 New Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43
(Richardson, 1842) (Scorpaenidae), Bollons’ rattail, Caelorinchus bollonsi, McCann & McKnight, 1980 (Macrouridae), oliver’s rattail, C. oliverianus Phillips, 1927, capro dory, Capromimus abbreviatus (Hector, 1875) (Zenionidae), silverside, Argentina elongata Hutton, 1879 (Argentinidae), and two-banded rattail, C. biclinozonalis Arai & McMillan, 1982 (Macrouridae), and also provides notes on the diets of 18 others.
MAterIAls And Methods
Fish were retained as bycatch from nine scampi trawls from between c. 240 and 450 m water depth off the Wairarapa, east coast, North Island, New Zealand, as part of a trawl survey to determine scampi abundance by the RV Kaharoa (KAH0401), during late January and early February 2004 (Jones & Leschen 2008) (Fig. 1). Trawls were undertaken using a Florida Flyer net with a 35 mm cod-end, and Bison doors, between 0600 and 1800 h (New Zealand daylight time), and for two nautical miles (c. 3.7 km) (NIWA unpubl. data). Fish were retained from a sub-set of these trawls, frozen, and then defrosted, weighed (± 1 g), and measured (± 1 mm). Pre-anal fin length (PAL) was used for rattails (Macrouridae), pre-caudal fin length (PCL) for dogfishes (etmopteridae) and catsharks (Scyliorhinidae), snout-vent length (SV) was used for eels (Congridae), and standard length (SL) was used for all other fish species. Fish were assigned a gender where possible, but no attempt was made to stage them (Table 1).
dietary analysisStomachs were removed and dissected, and their contents (when present) were blotted on tissue paper to remove excess moisture, weighed (± 0.01 g) and counted. No fish were found to have been obviously net-feeding. Index of relative importance, niche overlap, niche breadth, and Spearman rank correlations calculations were applied for species with >10 stomach content samples. Diets were interpreted using an index of relative importance (IRI) (Pinkas et al. 1971) as %IRI = F(W + N), where F is the frequency of occurrence percentage, expressed as the total number of occurrences of a food item as a percentage of the total number of stomachs; W is the percentage weight, expressed as the total weight of a food item as a percentage of the total weight of all food items; and N is the numerical percentage, expressed as the number of a food item as a percentage of the total number of that food item
Fig. 1 Survey area and the locations and mean depths (m) of scampi (Metanephrops challengeri) trawls from which the fish in this study were sourced. Trawls are represented by rectangles which represent the length of the trawl based upon start–finish coordinates, and were undertaken in a north to south direction (the “foul haul” trawl was of a markedly shorter duration than all other trawls).
Bull et al. 2001; Francis et al. 2002). Reports on feeding relationships are limited to Clark (1985a,b), who described the food and feeding of seven fish species from Campbell Plateau. Kerstan (1989) described the diet of silver roughy (Hoplostethus mediterraneus) from the upper slope of the northern half of North Island, and Graham (1938, 1953), described the diets of several shelf and upper-slope fish species from off the coast of otago, South Island, New Zealand. Neither Kerstan (1989) nor Graham (1938, 1953) investigated feeding relationships between fish species. Given the dearth of information on diets of New Zealand upper-slope fish communities, the current study provides information on aspects of the food and feeding of eight such species, javelinfish, Lepidorhynchus denticulatus (Richardson, 1846) (Macrouridae), silver roughy, H. mediterraneus Cuvier, 1829 (Trachichthyidae), sea perch, Helicolenus percoides

931Jones—Diets of upper-slope fish from Wairarapa, New Zealand
found in stomachs. Graphical representation of the IRI equation as an area chart followed Anderson (2005), Madurell & Cartes (2006), and Jones (2008a, b). Food taxa comprising less than 1% IRI were rounded to 1% and pooled. Spearman rank correlation (Spearman 1904) was applied to all samples per species to assess the relationship between the size of fish, and the number and weight of their food items. Species with >50 stomach content samples were split into arbitrary size classes to assess any ontogenetic change in diet. Niche overlap was calculated using a similarity index (Schoener 1974):
C p pxy xi yii
n
= − −=∑1 0 5
1
.
where Cxy is the similarity between diets of species x and y, pxi is the proportion of food category i in the diet of species x, and pyi is the proportion of food category i in the diet of species y. Dietary overlap increases from zero to one, and is generally considered high when it exceeds 0.6 (Keast 1978; Wallace 1981; Bulman et al. 2002). Cumulative food item curves were used to provide a broad assessment of the efficacy of the size of samples to describe the diets. Niche breadths were calculated using Levins’ (1968) formula:
B pi= ( )∑ −2 1
(where pi is the proportion of each food category), and standardised using Hespeheide’s (1975) method; Bs = (B−1)/(n−1). Input data to ontogenetic, niche overlap, and niche breadth calculations were based on %W, as mass has been considered to be a viable proxy for energy (Bulman et al. 2002; Madurell & Cartes 2005a, 2006). For species with <10 individuals with stomach contents, analysis was limited to recording the number of food items.
table 1 Total numbers examined, the number of stomachs containing food (SCF) and the weights and lengths of eight demersal fish species from off the Wairarapa coast, New Zealand. (SL, standard length; PAL, pre-anal fin length; PCL, pre-caudal fin length; SV, snout-vent length.)
Mean Mean SL, weight PAL1, PRC2,Family Species n SCF (g) CV SV3 (mm) CV
Macrouridae Lepidorhynchus denticulatus 121 85 67.3 88.4 66.81 20.4Trachicthyidae Hoplostethus mediterraneus 82 72 15.9 78.9 76.9 22.0Scorpaenidae Helicolenus percoides 124 70 94.5 133.3 137.1 27.6Zenionidae Capromimus abbreviatus 71 39 13.9 28.9 72.9 10.0Macrouridae Caelorinchus bollonsi 58 39 67.7 236.2 67.81 41.8Macrouridae Caelorinchus oliverianus 48 37 53 44.0 78.91 14.1Argentinidae Argentina elongata 21 13 31.1 34.8 155.5 12.6Macrouridae Caelorinchus biclinozonalis 11 10 217.9 85.8 115.71 18.0
results
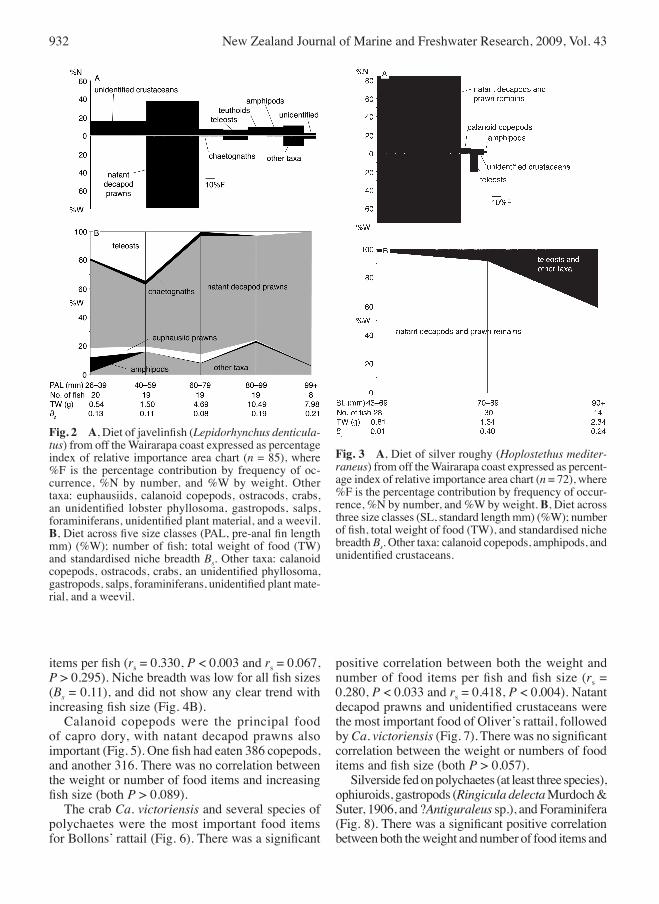
dietary analyses For javelinfish, natant decapod prawns were the most important food item (Fig. 2A), particularly Notopandalus magnoculus (Bate, 1888) and Pasiphaea aff. sivado; these two species were also abundant in the diets of larger fish (Fig. 2B). There was a significant positive correlation between both the weight and numbers of food items and fish size (Spearman rank correlation rs = 0.605, P < 0.000 and rs = 0.599, P < 0.000). Standardised niche breadth for all fish sizes was low (Bs = 0.18), and increased slightly with increasing fish size (Fig. 2B). The food of silver roughy was almost exclusively natant decapod prawns and “prawn remains” (either natant decapods, mysids or euphausiids, but too digested to be identified further) (Fig. 3A). There was a significant positive correlation between the weight of food items and fish size (rs = 0.300, P < 0.005), but not with the number of food items per fish (rs = 0.025, P > 0.418). Fish and other taxa became more prevalent in the diets of larger fish. Niche breadth for all fish sizes was low (Bs = 0.01), but increased with fish size (Fig. 3B). Natant decapod prawns (including N. magnoculus and Pontophilus acutirostratus Yaldwyn, 1960) and the crab Carcinoplax victoriensis Rathbun, 1923 were the most important food of sea perch (Fig. 4A). There was evidence for an ontogenetic shift in diet, with smaller fish feeding upon prawns, and then switching to Ca. victoriensis as they grew larger. The largest size class was dominated by the stomach contents of the largest fish recorded, which itself had eaten fish remains which weighed more than all stomach contents combined. There was a significant positive correlation between the weight of food items and fish size, but not with the number of food
932 New Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43
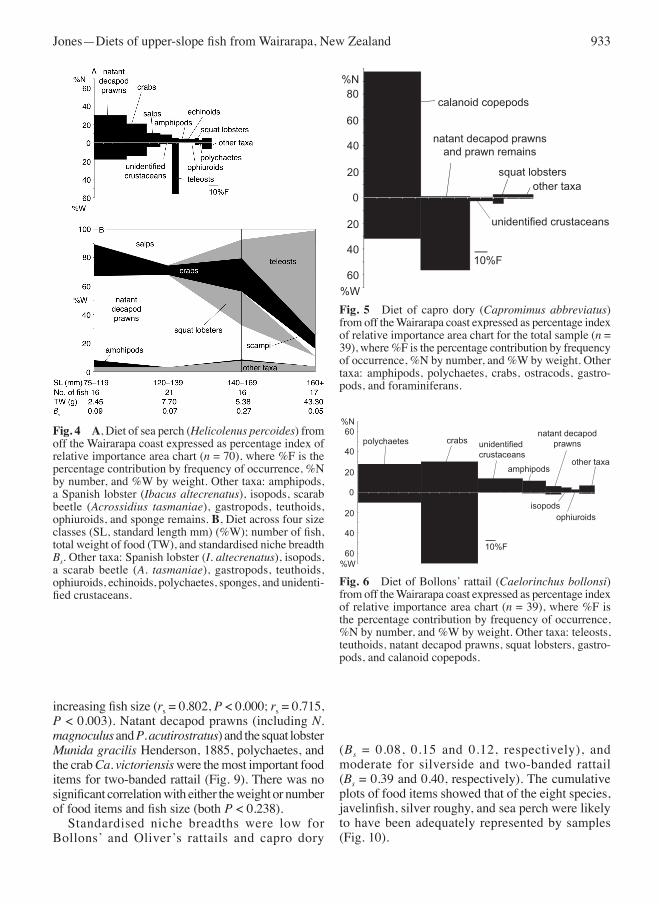
items per fish (rs = 0.330, P < 0.003 and rs = 0.067, P > 0.295). Niche breadth was low for all fish sizes (Bs = 0.11), and did not show any clear trend with increasing fish size (Fig. 4B). Calanoid copepods were the principal food of capro dory, with natant decapod prawns also important (Fig. 5). one fish had eaten 386 copepods, and another 316. There was no correlation between the weight or number of food items and increasing fish size (both P > 0.089). The crab Ca. victoriensis and several species of polychaetes were the most important food items for Bollons’ rattail (Fig. 6). There was a significant
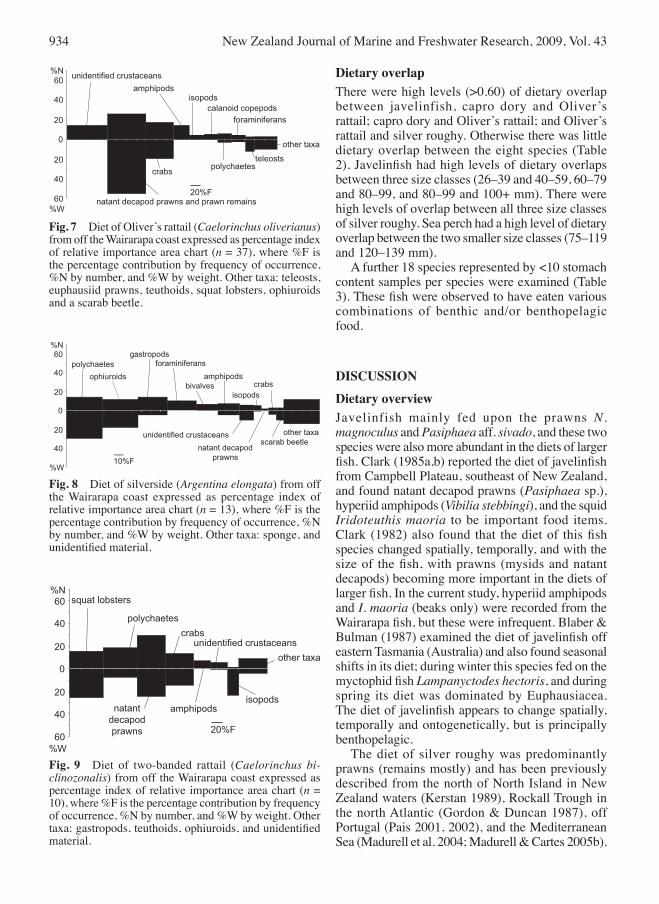
positive correlation between both the weight and number of food items per fish and fish size (rs = 0.280, P < 0.033 and rs = 0.418, P < 0.004). Natant decapod prawns and unidentified crustaceans were the most important food of oliver’s rattail, followed by Ca. victoriensis (Fig. 7). There was no significant correlation between the weight or numbers of food items and fish size (both P > 0.057). Silverside fed on polychaetes (at least three species), ophiuroids, gastropods (Ringicula delecta Murdoch & Suter, 1906, and ?Antiguraleus sp.), and Foraminifera (Fig. 8). There was a significant positive correlation between both the weight and number of food items and
Fig. 2 A, Diet of javelinfish (Lepidorhynchus denticula-tus) from off the Wairarapa coast expressed as percentage index of relative importance area chart (n = 85), where %F is the percentage contribution by frequency of oc-currence, %N by number, and %W by weight. other taxa: euphausiids, calanoid copepods, ostracods, crabs, an unidentified lobster phyllosoma, gastropods, salps, foraminiferans, unidentified plant material, and a weevil. B, Diet across five size classes (PAL, pre-anal fin length mm) (%W); number of fish; total weight of food (TW) and standardised niche breadth Bs. other taxa: calanoid copepods, ostracods, crabs, an unidentified phyllosoma, gastropods, salps, foraminiferans, unidentified plant mate-rial, and a weevil.
Fig. 3 A, Diet of silver roughy (Hoplostethus mediter-raneus) from off the Wairarapa coast expressed as percent-age index of relative importance area chart (n = 72), where %F is the percentage contribution by frequency of occur-rence, %N by number, and %W by weight. B, Diet across three size classes (SL, standard length mm) (%W); number of fish, total weight of food (TW), and standardised niche breadth Bs. other taxa: calanoid copepods, amphipods, and unidentified crustaceans.
933Jones—Diets of upper-slope fish from Wairarapa, New Zealand
increasing fish size (rs = 0.802, P < 0.000; rs = 0.715, P < 0.003). Natant decapod prawns (including N. magnoculus and P. acutirostratus) and the squat lobster Munida gracilis Henderson, 1885, polychaetes, and the crab Ca. victoriensis were the most important food items for two-banded rattail (Fig. 9). There was no significant correlation with either the weight or number of food items and fish size (both P < 0.238). Standardised niche breadths were low for Bollons’ and oliver’s rattails and capro dory
Fig. 4 A, Diet of sea perch (Helicolenus percoides) from off the Wairarapa coast expressed as percentage index of relative importance area chart (n = 70), where %F is the percentage contribution by frequency of occurrence, %N by number, and %W by weight. other taxa: amphipods, a Spanish lobster (Ibacus altecrenatus), isopods, scarab beetle (Acrossidius tasmaniae), gastropods, teuthoids, ophiuroids, and sponge remains. B, Diet across four size classes (SL, standard length mm) (%W); number of fish, total weight of food (TW), and standardised niche breadth Bs. other taxa: Spanish lobster (I. altecrenatus), isopods, a scarab beetle (A. tasmaniae), gastropods, teuthoids, ophiuroids, echinoids, polychaetes, sponges, and unidenti-fied crustaceans.
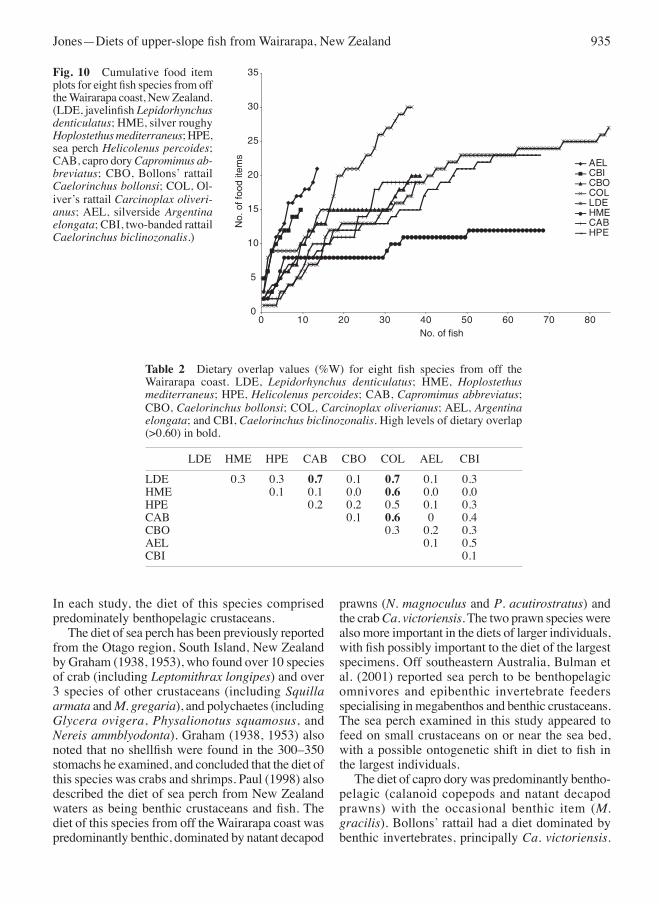
(Bs = 0.08, 0.15 and 0.12, respectively), and moderate for silverside and two-banded rattail (Bs = 0.39 and 0.40, respectively). The cumulative plots of food items showed that of the eight species, javelinfish, silver roughy, and sea perch were likely to have been adequately represented by samples (Fig. 10).
20
40
60
0
20
40
%N
%W
calanoid copepods
10%F
natant decapod prawnsand prawn remains
other taxasquat lobsters
80
60
unidentifi ed crustaceans
Fig. 5 Diet of capro dory (Capromimus abbreviatus) from off the Wairarapa coast expressed as percentage index of relative importance area chart for the total sample (n = 39), where %F is the percentage contribution by frequency of occurrence, %N by number, and %W by weight. other taxa: amphipods, polychaetes, crabs, ostracods, gastro-pods, and foraminiferans.
crabs unidentifi ed crustaceans
polychaetesnatant decapod
prawns
other taxaamphipods
isopodsophiuroids
20
40
60
0
20
40
%N
%W
10%F60
Fig. 6 Diet of Bollons’ rattail (Caelorinchus bollonsi) from off the Wairarapa coast expressed as percentage index of relative importance area chart (n = 39), where %F is the percentage contribution by frequency of occurrence, %N by number, and %W by weight. other taxa: teleosts, teuthoids, natant decapod prawns, squat lobsters, gastro-pods, and calanoid copepods.
934 New Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43
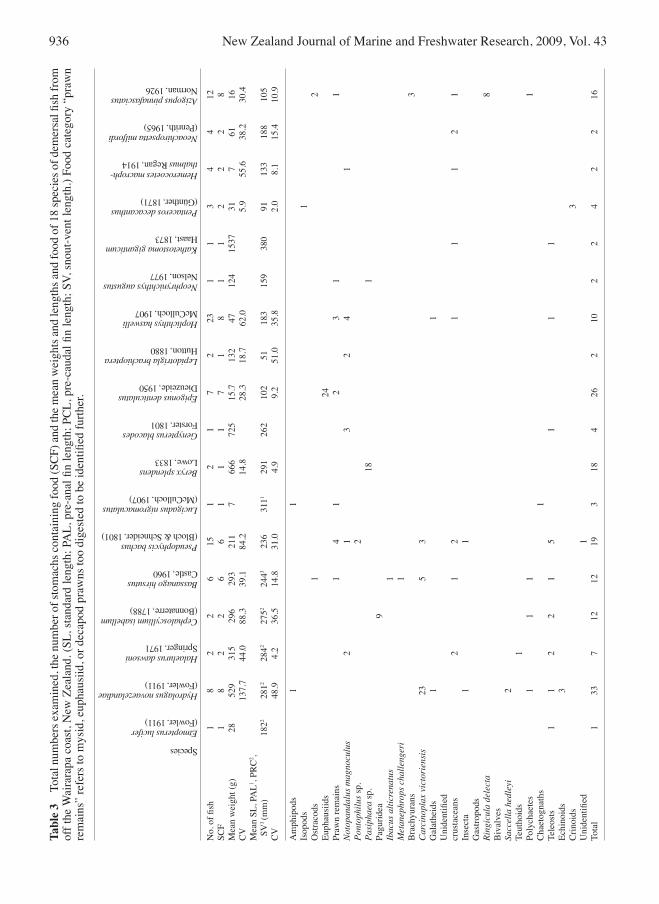
dietary overlapThere were high levels (>0.60) of dietary overlap between javelinfish, capro dory and oliver’s rattail; capro dory and oliver’s rattail; and oliver’s rattail and silver roughy. otherwise there was little dietary overlap between the eight species (Table 2). Javelinfish had high levels of dietary overlaps between three size classes (26–39 and 40–59, 60–79 and 80–99, and 80–99 and 100+ mm). There were high levels of overlap between all three size classes of silver roughy. Sea perch had a high level of dietary overlap between the two smaller size classes (75–119 and 120–139 mm). A further 18 species represented by <10 stomach content samples per species were examined (Table 3). These fish were observed to have eaten various combinations of benthic and/or benthopelagic food.
dIscussIon
dietary overview Javelinfish mainly fed upon the prawns N. magnoculus and Pasiphaea aff. sivado, and these two species were also more abundant in the diets of larger fish. Clark (1985a,b) reported the diet of javelinfish from Campbell Plateau, southeast of New Zealand, and found natant decapod prawns (Pasiphaea sp.), hyperiid amphipods (Vibilia stebbingi), and the squid Iridoteuthis maoria to be important food items. Clark (1982) also found that the diet of this fish species changed spatially, temporally, and with the size of the fish, with prawns (mysids and natant decapods) becoming more important in the diets of larger fish. In the current study, hyperiid amphipods and I. maoria (beaks only) were recorded from the Wairarapa fish, but these were infrequent. Blaber & Bulman (1987) examined the diet of javelinfish off eastern Tasmania (Australia) and also found seasonal shifts in its diet; during winter this species fed on the myctophid fish Lampanyctodes hectoris, and during spring its diet was dominated by euphausiacea. The diet of javelinfish appears to change spatially, temporally and ontogenetically, but is principally benthopelagic. The diet of silver roughy was predominantly prawns (remains mostly) and has been previously described from the north of North Island in New Zealand waters (Kerstan 1989), Rockall Trough in the north Atlantic (Gordon & Duncan 1987), off Portugal (Pais 2001, 2002), and the Mediterranean Sea (Madurell et al. 2004; Madurell & Cartes 2005b).
unidentifi ed crustaceans
natant decapod prawns and prawn remains
crabspolychaetes
amphipodsisopods
calanoid copepodsforaminiferans
other taxa
teleosts
20
40
60
0
20
40
%N
%W
20%F60
polychaetes
bivalvesophiuroids
other taxa
gastropods
natant decapodprawns
foraminiferans
amphipods
isopods
unidentifi ed crustaceans
crabs
scarab beetle
20
40
60
0
20
40
%N
%W10%F
20
40
60
0
20
40
%N
%W
20%F60
isopods
polychaetes
natant decapod prawns
other taxa
crabs
squat lobsters
amphipods
unidentifi ed crustaceans
Fig. 7 Diet of oliver’s rattail (Caelorinchus oliverianus) from off the Wairarapa coast expressed as percentage index of relative importance area chart (n = 37), where %F is the percentage contribution by frequency of occurrence, %N by number, and %W by weight. other taxa: teleosts, euphausiid prawns, teuthoids, squat lobsters, ophiuroids and a scarab beetle.
Fig. 8 Diet of silverside (Argentina elongata) from off the Wairarapa coast expressed as percentage index of relative importance area chart (n = 13), where %F is the percentage contribution by frequency of occurrence, %N by number, and %W by weight. other taxa: sponge, and unidentified material.
Fig. 9 Diet of two-banded rattail (Caelorinchus bi-clinozonalis) from off the Wairarapa coast expressed as percentage index of relative importance area chart (n = 10), where %F is the percentage contribution by frequency of occurrence, %N by number, and %W by weight. other taxa: gastropods, teuthoids, ophiuroids, and unidentified material.
935Jones—Diets of upper-slope fish from Wairarapa, New Zealand
In each study, the diet of this species comprised predominately benthopelagic crustaceans. The diet of sea perch has been previously reported from the otago region, South Island, New Zealand by Graham (1938, 1953), who found over 10 species of crab (including Leptomithrax longipes) and over 3 species of other crustaceans (including Squilla armata and M. gregaria), and polychaetes (including Glycera ovigera, Physalionotus squamosus, and Nereis ammblyodonta). Graham (1938, 1953) also noted that no shellfish were found in the 300–350 stomachs he examined, and concluded that the diet of this species was crabs and shrimps. Paul (1998) also described the diet of sea perch from New Zealand waters as being benthic crustaceans and fish. The diet of this species from off the Wairarapa coast was predominantly benthic, dominated by natant decapod
Fig. 10 Cumulative food item plots for eight fish species from off the Wairarapa coast, New Zealand. (LDe, javelinfish Lepidorhynchus denticulatus; HMe, silver roughy Hoplostethus mediterraneus; HPe, sea perch Helicolenus percoides; CAB, capro dory Capromimus ab-breviatus; CBo, Bollons’ rattail Caelorinchus bollonsi; CoL, ol-iver’s rattail Carcinoplax oliveri-anus; AeL, silverside Argentina elongata; CBI, two-banded rattail Caelorinchus biclinozonalis.)
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70 80
AELCBICBOCOLLDEHMECABHPE
No. of fish
No.
of f
ood
item
s
table 2 Dietary overlap values (%W) for eight fish species from off the Wairarapa coast. LDe, Lepidorhynchus denticulatus; HMe, Hoplostethus mediterraneus; HPe, Helicolenus percoides; CAB, Capromimus abbreviatus; CBo, Caelorinchus bollonsi; CoL, Carcinoplax oliverianus; AeL, Argentina elongata; and CBI, Caelorinchus biclinozonalis. High levels of dietary overlap (>0.60) in bold.
LDe HMe HPe CAB CBo CoL AeL CBI
LDe 0.3 0.3 0.7 0.1 0.7 0.1 0.3HMe 0.1 0.1 0.0 0.6 0.0 0.0HPe 0.2 0.2 0.5 0.1 0.3CAB 0.1 0.6 0 0.4CBo 0.3 0.2 0.3AeL 0.1 0.5CBI 0.1
prawns (N. magnoculus and P. acutirostratus) and the crab Ca. victoriensis. The two prawn species were also more important in the diets of larger individuals, with fish possibly important to the diet of the largest specimens. off southeastern Australia, Bulman et al. (2001) reported sea perch to be benthopelagic omnivores and epibenthic invertebrate feeders specialising in megabenthos and benthic crustaceans. The sea perch examined in this study appeared to feed on small crustaceans on or near the sea bed, with a possible ontogenetic shift in diet to fish in the largest individuals. The diet of capro dory was predominantly bentho-pelagic (calanoid copepods and natant decapod prawns) with the occasional benthic item (M. gracilis). Bollons’ rattail had a diet dominated by benthic invertebrates, principally Ca. victoriensis.
936 New Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43ta
ble 3
To
tal n
umbe
rs e
xam
ined
, the
num
ber o
f sto
mac
hs c
onta
inin
g fo
od (S
CF)
and
the
mea
n w
eigh
ts a
nd le
ngth
s and
food
of 1
8 sp
ecie
s of d
emer
sal fi
sh fr
om
off t
he W
aira
rapa
coa
st, N
ew Z
eala
nd. (
SL, s
tand
ard
leng
th; P
AL,
pre
-ana
l fin
leng
th; P
CL,
pre
-cau
dal fi
n le
ngth
; SV,
sno
ut-v
ent l
engt
h.) F
ood
cate
gory
“pr
awn
rem
ains
” re
fers
to m
ysid
, eup
haus
iid, o
r dec
apod
pra
wns
too
dige
sted
to b
e id
entifi
ed fu
rther
.
j No.
of fi
sh
1 8
2 2
6 15
1
2 1
7 2
23
1 1
3 4
4 12
SCF
1 8
2 2
6 6
1 1
1 7
1 8
1 1
2 2
2 8
Mea
n w
eigh
t (g)
28
52
9 31
5 29
6 29
3 21
1 7
666
725
15.7
13
2 47
12
4 15
37
31
7 61
16
CV
137.
7 44
.0
88.3
39
.1
84.2
14.8
28.3
18
.7
62.0
5.
9 55
.6
38.2
30
.4M
ean
SL, P
AL1 ,
PRC
2 , S
V3 (m
m)
1822
2812
2842
2752
2443
236
3111
291
262
102
51
183
159
380
91
133
188
105
CV
48.9
4.
2 36
.5
14.8
31
.0
4.
9
9.2
51.0
35
.8
2.0
8.1
15.4
10
.9
Am
phip
ods
1
1
Is
opod
s
1
ost
raco
ds
1
2
euph
ausi
ids
24
Pr
awn
rem
ains
1
4 1
2
3 1
1N
otop
anda
lus m
agno
culu
s
2
1
3
2
4
1 Po
ntop
hilu
s sp.
2
Pasi
phae
a sp
.
18
1
Pa
gurid
ea
9
Ibac
us a
lticr
enat
us
1
Met
anep
hrop
s cha
lleng
eri
1
B
rach
yura
ns
3
Car
cino
plax
vic
tori
ensi
s
23
5 3
Gal
athe
ids
1
1
Uni
dent
ified
cr
usta
cean
s
2
1
2
1
1
1 2
1In
sect
a
1
1
Gas
tropo
ds
Ring
icul
a de
lect
a
8B
ival
ves
Sacc
ella
hed
leyi
2
Te
utho
ids
1
Po
lych
aete
s
1
1 1
1C
haet
ogna
ths
1
Tele
osts
1
1 2
2 1
5
1
1
1
echi
noid
s
3
C
rinoi
ds
3
U
nide
ntifi
ed
1
Tota
l 1
33
7 12
12
19
3
18
4 26
2
10
2 2
4 2
2 16
Species
Etmopterus lucifer (Fowler, 1911)
Hydrolagus novaezelandiae (Fowler, 1911)
Halaelurus dawsoni Springer, 1971
Cephaloscyllium isabellum (Bonnaterre, 1788)
Bassanago hirsutus Castle, 1960
Pseudophycis bachus (Bloch & Schneider, 1801)
Lucigadus nigromaculatus (McCulloch, 1907)
Beryx splendens Lowe, 1833
Genypterus blacodes Forster, 1801
Epigonus denticulatus Dieuzeide, 1950
Lepidotrigla brachioptera Hutton, 1880
Hoplichthys haswelli McCulloch, 1907
Neophrynichthys augustus Nelson, 1977
Kathetostoma giganticum Haast, 1873
Pentaceros decacanthus (Günther, 1871)
Hemerocoetes macroph- thalmus Regan, 1914
Neoachiropsetta milfordi (Penrith, 1965)
Azigopus pinnafasciatus Norman, 1926

937Jones—Diets of upper-slope fish from Wairarapa, New Zealand
Two-banded rattail had also had a predominately benthic diet feeding on benthic prawns (N. magnoculus and P. acutirostratus), squat lobsters (M. gracilis), and crabs (Ca. victoriensis), although the sample was small. The diets of Bollons’ and two-banded rattails and of capro dory have not been reported previously. Natant decapod prawns, unidentified crustaceans, and Ca. victoriensis were also important in the diet of oliver’s rattail. Clark et al. (1989) previously reported the diet of oliver’s rattail from southwestern Chatham Rise as comprising calanoid copepods and hyperiid amphipods as the most important food, suggesting that the overall diet of this species could be a mix of benthic and benthopelagic organisms, which is consistent with findings reported here. The silverside examined in this study appeared to have a benthic diet. Clark (1985a) reported the diet of silverside from Campbell Plateau as being almost exclusively salps (Iasis zonaria and Salpa thompsoni), although he also recorded gastropods and polychaetes. Graham (1953) described the diet of this species from the otago region and found primarily benthic fauna in its diet: the crab Cyclograpsis lavauxi (likely a misidentification of Ca. victoriensis given the almost supralittoral boulder-shore distribution of the former (Morton & Miller 1968)), squat lobsters (M. gregaria), isopods, and polychaetes (especially G. ovigera). Clark (1985a) suggested that salps consumed by fish from Campbell Plateau were evenly distributed throughout the water column and the feeding of silverside were reflecting the lower range of this distribution. It is possible that this species was picking these usually pelagic organisms, dead, from the sea bed, so the diet of this species could be predominantly benthic in origin. The diets of 10 of the remaining 18 species are reported for the first time for New Zealand waters, and although samples were too few to provide detailed dietary accounts, they provide some indication of the diets of fish species for which previously nothing was known. These dietary data represent only the second account from New Zealand waters for an upper-slope fish assemblage, and the first account for fin-fish bycatch species recovered from a scampi fishery. These data complement other recent research on the diets of deep-sea fish species (to depths of c. 1000 m) from northeastern Chatham Rise and the Puysegur region (Jones 2008a,b), providing baseline data for any future ecosystem based analysis, and building on our knowledge of trophic relationships between fish and invertebrates in a hitherto unstudied area.
AcKnowledgMents
Thanks to those persons within the New Zealand Ministry of Fisheries and National Institute of Water and Atmospheric Research Limited (NIWA) who assisted with the procurement and retention of deep-sea finfish bycatch samples from the KAH0401 fisheries trawl survey: particularly Martin Cryer, Bruce Hartill, and Dean Stotter. Thanks also to Andrew Stewart, Museum of New Zealand Te Papa Tongarewa (fish identifications), and to Steve o’Shea (Auckland University of Technology) and two anonymous reviewers for their helpful comments on earlier drafts of this manuscript.
reFerences
Anderson Me 2005. Food habits of some deep-sea fish off South Africa’s west coast and Agulhas Bank. 1. The grenadiers (Teleostei: Macrouridae). South African Journal of Marine Science 27: 409–426.
Anderson oF, Bagley NW, Hurst RJ, Francis MP, Clark MR, McMillan PJ 1998. Atlas of New Zealand fish and squid distributions from research bottom trawls. Technical Report 42. National Institute of Water and Atmospheric Research Ltd, Wellington, New Zealand.
Andrew NL, Pepperell 1992. The by-catch of shrimp trawl fisheries. oceanography and Marine Biology: an Annual Review 30: 527–565.
Aydin K, Mueter F 2007. The Bering Sea—a dynamic food web perspective. Deep-Sea Research Part II 54: 2501–2525.
Blaber SJM, Bulman CM 1987. Diets of fishes of the upper continental slope of eastern Tasmania: content, calorific values, dietary overlap and trophic relationships. Marine Biology 95: 345–356.
Bull B, Livingston Me, Hurst R, Bagley N 2001. Upper-slope fish communities on the Chatham Rise, New Zealand, 1992–99. New Zealand Journal of Marine and Freshwater Research 35: 795–815.
Bulman CM, Althaus F, He X, Bax NJ, Williams A 2001. Diets and trophic guilds of demersal fishes of the south-eastern Australian shelf. Marine and Freshwater Research 52: 537–548.
Bulman CM, He X, Koslow JA 2002. Trophic ecology of the mid-slope demersal fish community off southern Tasmania, Australia. Marine and Freshwater Research 53: 59–72.
Carbines G, Jiang W, Beentjes MP 2004. The impact of oyster dredging on blue cod Parapercis colias, in Foveaux Strait, New Zealand. Aquatic Conservation: Marine and Freshwater ecosystems 14: 491–504.

938 New Zealand Journal of Marine and Freshwater Research, 2009, Vol. 43
Castro M, Araújo A, Monteiro P 2005. Fate of discards from deep-water crustacean trawl fishery off the south coast of Portugal. New Zealand Journal of Marine and Freshwater Research 39: 437–446.
Clark MR 1982. The food and feeding relationships of fish species from the Campbell Plateau, New Zealand. Unpublished PhD thesis, Victoria University of Wellington, Wellington, New Zealand.
Clark MR 1985a. Food and feeding of seven fish species from the Campbell Plateau, New Zealand. New Zealand Journal of Marine and Freshwater Research 19: 339–363.
Clark MR 1985b. Feeding relationships of seven fish species from the Campbell Plateau, New Zealand. New Zealand Journal of Marine and Freshwater Research 19: 365–374.
Clark MR, King KJ, McMillan PJ, 1989. The food and feeding relationships of black oreo, Allocyttus niger, smooth oreo Pseudocyttus maculatus and eight other fish species from the continental slope of the south-west Chatham Rise, New Zealand. Journal of Fish Biology 35: 465–484.
Cryer M, Stotter DR 1999. Movement and growth rates of scampi inferred from tagging, Alderman Islands, western Bay of Plenty. Technical Report 49. National Institute of Water and Atmospheric Research Ltd, Wellington, New Zealand.
Cryer M, Hartill B, o’Shea S 2002. Modification of marine benthos by trawling: towards a generalization for the deep ocean? ecological Applications 12: 1824−1839.
evans SM, Hunter Je, elizal, Wahju RI 1994. Composition and fate of the catch and bycatch in the Farne Deep (North Sea) Nephrops fishery. ICeS Journal of Marine Science 51: 155–168.
Fosså JH, Mortensen PB, Furevik DM 2002. The deep-water coral Lophelia pertusa in Norwegian waters: distribution and fishery impacts. Hydrobiologia 471: 1–12.
Francis MP, Hurst RJ, McArdle BH, Bagley NW, Anderson oF 2002. New Zealand demersal fish assemblages. environmental Biology of Fishes 65: 215–234.
Gordon JDM, Duncan JAR 1987. Aspects of the biology of Hoplostethus atlanticus and H. mediterraneus (Pisces: Berycomorphi) from the slopes of the Rockall Trough and the Porcupine Sea Bight (north-eastern Atlantic). Journal of the Marine Biological Association of the United Kingdom 67: 119–133.
Graham DH 1938. Food of fishes from otago Harbour and adjacent sea. Transactions and Proceedings of the Royal Society of New Zealand 68: 421–436.
Graham DH 1953. A treasury of New Zealand fishes. Wellington, New Zealand, AH & AW Reed. 424 p.
Hartill BW, Cryer M, MacDiarmid AB 2006. Reducing bycatch in New Zealand’s scampi trawl fisheries. New Zealand aquatic environment and biodiversity report No. 4. New Zealand Ministry of Fisheries, Wellington, New Zealand.
Hespenheide HA 1975. Prey characteristics and predator niche width. In: Cody ML, Diamond JM ed. ecology and evolution of communities. Cambridge, Massachusetts, Belknap Press. Pp. 158–180.
Jones MRL 2008a. Dietary analysis of Coryphaenoides serrulatus, C. subserrulatus and several other species of macrourid fish (Pisces: Macrouridae) from northeastern Chatham Rise, New Zealand. New Zealand Journal of Marine and Freshwater Research 42: 73–84.
Jones MRL 2008b. Biology and diet of Coryphaenoides subserrulatus and Etmopterus baxteri from the Puysegur region, southern New Zealand. New Zealand Journal of Marine and Freshwater Research 42: 333–337.
Jones MRL, Leschen RAB 2008. Beetles in the diet of six species of deep-sea fish. New Zealand Journal of Zoology 35: 251–253.
Keast A 1978. Feeding interrelations between age-groups of pumpkinseed (Lepomis gibbosus) and comparisons with bluegill (L. macrochiris). Fisheries Research Board of Canada 35: 12–27.
Kerstan SL 1989. The food of silver roughy (Hoplostethus mediterraneus, Beryciformes, Trachicthyidae) from New Zealand waters. Kieler Meeresforschungen 32: 241–247.
Levins R 1968. evolution in changing environments. New Jersey, Princeton University Press. 132 p.
McShane Pe, Broadhurst MK, Williams A 2007. Keeping watch on the unwatchable: technological solutions for the problems generated by ecosystem-based management. Fish and Fisheries 8: 153–161.
Madurell T, Cartes Je 2005a. Trophodynamics of a deep-sea demersal fish assemblage from the bathyal eastern Ionian Sea (Mediterranean Sea). Deep-Sea Research Part I 52: 2049–2064.
Madurell T, Cartes Je 2005b. Temporal changes in feeding habits and daily rations of Hoplostethus mediterraneus, Cuvier, 1829 in the eastern Mediterranean (Ionian Sea). Marine Biology 146 (5): 951–962.
Madurell T, Cartes Je 2006. Trophic relationships and food consumption of slope dwelling macrourids from the bathyal Ionian Sea (eastern Mediterranean). Marine Biology 148: 1325–1338.

939Jones—Diets of upper-slope fish from Wairarapa, New Zealand
Madurell T, Cartes Je, Labropoulou M 2004. Changes in the structure of fish assemblages in a bathyal site of the Ionian Sea (eastern Mediterranean). Fisheries Research 66: 245–260.
Morton Je, Miller MC 1968. The New Zealand sea shore. Auckland, New Zealand, Collins. 638 p.
Pais C 2001. Aspects of the biology of Hoplostethus mediterraneus from the south coast of Portugal. Journal of the Marine Biological Association of the United Kingdom 81: 711–712.
Pais C 2002. Diet of a deep-sea fish, Hoplostethus mediterraneus, from the south coast of Portugal. Journal of the Marine Biological Association of the United Kingdom 82: 351–352.
Pantin HM 1963. Submarine morphology east of the North Island, New Zealand. DSIR Bulletin 149. Department of Scientific and Industrial Research, Wellington, New Zealand.
Paul LJ 1998. A summary of biology and commercial landings, and a stock assessment of the sea perches, Helicolenus spp. (Scorpaenidae) in New Zealand waters. New Zealand Fisheries Assessment Research Document. 1998/29.
Pikitch eK, Santora C, Babcock A and others 2004. ecosystem-based fisheries management. Science 305: 346–347.
Pinkas L, oliphant MS, Iverson ILK 1971. Food habits of albacore, blue fin tuna, and bonito in California waters. Fish Bulletin 152, State of California, Department of Fish and Game, United States.
Schoener TW 1974. Resource partitioning in ecological communities. Science 185: 27–39.
Smith ADM, Fulton eJ, Hobday AJ, Smith DC, Shoulder P 2007. Scientific tools to support the practical implementation of ecosystem-based fisheries management. ICeS Journal of Marine Science 64: 633–639.
Smith PJ 1999. Allozyme variation in scampi (Metanephrops challengeri) fisheries around New Zealand. New Zealand Journal of Marine and Freshwater Research 33: 491–497.
Spearman C 1904. The proof and measurement of association between two things. American Journal of Psychology 15: 72–101.
Wallace RK 1981. An assessment of diet-overlap indexes. Transactions of the American Fisheries Society 110: 72–76.




















