
ISOLEVUGLANDIN-PROTEIN CROSS-LINKING - OhioLINK ...
317
PART I: ISOLEVUGLANDIN-PROTEIN CROSS-LINKING: STRUCTURE AND MECHANISM PART II: GENERATION AND CHARACTERIZATION OF A MONOCLONAL ISOLEVUGLANDIN[4]-PROTEIN ADDUCT ANTIBODY by WENZHAO BI Submitted in partial fulfillment of the requirements For the degree of Doctor of Philosophy Thesis Advisor: Dr. Robert G. Salomon Co-Advisor: Dr. John W. Crabb Department of Chemistry CASE WESTERN RESERVE UNIVERSITY August 2014
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of ISOLEVUGLANDIN-PROTEIN CROSS-LINKING - OhioLINK ...

PART I: ISOLEVUGLANDIN-PROTEIN CROSS-LINKING:
STRUCTURE AND MECHANISM
PART II: GENERATION AND CHARACTERIZATION OF A
MONOCLONAL ISOLEVUGLANDIN[4]-PROTEIN ADDUCT
ANTIBODY
by
WENZHAO BI
Submitted in partial fulfillment of the requirements
For the degree of Doctor of Philosophy
Thesis Advisor: Dr. Robert G. Salomon
Co-Advisor: Dr. John W. Crabb
Department of Chemistry
CASE WESTERN RESERVE UNIVERSITY
August 2014

CASE WESTERN RESERVE UNIVERSITY
SCHOOL OF GRADUATE STUDIES
candidate for the Doctor of Philosophy degree *.
(signed) Dr. Michael G. Zagorski . (Chair of the committee)
Dr. James D. Burgess .
Dr. Rajesh Viswanathan .
Dr. John W. Crabb .
Dr. Robert G. Salomon .
(Date) 04 /28 /2014
*We also certify that written approval has been obtained for any proprietary materialcontained therein.
We hereby approve the dissertation of
Wenzhao Bi

This thesis is dedicated to my parents and my husband.
iii

TABLE OF CONTENTS
Table of Contents iv
List of Schemes vii
List of Tables x
List of Figures xi
Acknowledgements xvii
List of Abbreviations and Acronyms xix
Abstract xxiv
Part I: Isolevuglandin-protein Cross-linking: Structure and Mechanism
Part II: Generation and Characterization of a Monoclonal
Isolevuglandin[4]E2-protein Adduct Antibody
1. Introduction 1
1.1 Lipid Peroxidation 2
1.2 Enzymatic Pathways 2
1.3 The Free Radical Pathway 4
1.4 Biological Activities of Levuglandins and Isolevuglandins 7
1.5 References 22
2. Adduction and Cross-linking of Calpain-1 and GAPDH by Iso[4]LGE2
33
2.1 Background 34
2.2 Results and Discussion 38
2.3 Conclusions 58
iv

2.4 Experimental Procedures 60
2.5 References 71
3. Detection and Characterization of the Products from the Reaction of
Iso[4]LGE2 with Peptides: N-Acetyl-gly-lys-OMe and Amyloid
“EVHHQKL” 80
3.1 Background 81
3.2 Results and Discussion 88
3.3 Conclusions 108
3.4 Experimental Procedures 109
3.5 References 114
4. Generation of IsoLG-pyrrole by the Reaction of IsoLG with N-Acetyl-
gly-lys-OMe in the Absence of O2 and the Oxidation of IsoLG-pyrrole
to Form Pyrrole-pyrrole Cross-links and an Adduct with N-
Acetylcysteine 120
3.1 Background 121
3.2 Results and Discussion 123
3.3 Conclusions 147
3.4 Experimental Procedures 148
3.5 References 155
5. Generation of an Anti-iso[4]LGE2 Monoclonal Antibody 159
v

5.1 Background 160
5.2 Results and Discussion 162
5.3 Conclusions 169
5.4 Experimental Procedures 170
5.5 References 185
Appendix 187
Bibliography 236
vi

LIST OF SCHEMES
Chapter 1
Scheme 1.1. Cyclooxygenase oxidation of AA generates PGs and LGs via
rearrangement of PGH2
3
Scheme 1.2 Schematic representation of rearrangement of PGH2 to
levuglandin.
4
Scheme 1.3 Generation of isolevuglandins and isolevuglandin-protein adducts
during free radical oxidation of AA-esters
5
Scheme 1.4 Formation of LG-protein adducts, protein-protein and DNA-
protein cross-links
6
Chapter 2
Scheme 2.1 Postulated structures of LG and iso[4]LG protein adducts and
cross-links.
35
Scheme 2.2 Synthesis of a fluorescently labeled iso[4]LGE2 derivative. 50
Scheme 2.3 AccQ·TagTM derivatization. 54
Scheme 2.4 Chemical structure of peptides aminal cross-link. The adduct
molecular weight was 604.3.
56
Chapter 3
Scheme 3.1 Schematic representation of pyrrole cross-linking structure in
previous study: adducts of PGJ2 reaction with GSH in HCA-7
81
vii

cells.
Scheme 3.2 Schematic representation of pyrrole cross-linking structure in
previous study: adducts of nucleosides and DNA formed by
reaction with levuglandin.
82
Scheme 3.3 Schematic representation of pyrrole cross-linking structure in
previous study: formation of bis(levuglandinyl) urea.
83
Scheme 3.4 Schematic representation of pyrrole cross-linking structure in
previous study: nucleophilic substitution of alkylpyroles in the
presence of oxygen.
83
Scheme 3.5 Schematic representation of pyrrole cross-linking structure in
previous studies: (A) Pyrrole autoxidation products in 2,5-
hexanedione-treated dipeptide. (B) Formation of thiol-pyrrole
conjugates.
84
Scheme 3.6 Schematic representation of formation of iso[4]levuglandin protein
adducts and cross-links.
85
Scheme 3.7 Possible structures for products of oxidized bispyrrole (+16, +32,
+48, +64).
92
Scheme 3.8 Possible fragmentation of an iso[4]LGE2-(N-acetyl-gly-lys-OMe)
aminal cross-link at m/z 853.5 (M + H+).
96
Scheme 3.9 Possible fragmentation of iso[4]LGE2-FluoTag aminal cross-link
with N-acetyl-gly-lys-OMe at m/z 1120.6 (M + H+).
100
Chapter 4
viii

Scheme 4.1 LG/isoLG modification of proteins 121
Scheme 4.2 Synthesis of iso[4]LGE2-pyrrole. 124
Scheme 4.3 Reaction of pure iso[4]LGE2-pyrrole with N-acetylcysteine. 130
Scheme 4.4 Possible electron transfer mechanism for pure iso[4]LGE2-pyrrole
reaction with N-acetylcysteine.
134
Scheme 4.5: Reduction of TEMPO by ferrous iron. 140
Scheme 4.6: Possible electron transfer mechanism for pyrrole oxidized reaction 143
Scheme 4.7 Initiate reactions of 3-ethyl-2,4-dimethylpyrrole 144
Scheme 4.8 Preparation of 3-ethyl-1,2,4-trimethylpyrrole (MP) 145
Scheme 4.9 Initiate reactions of 3-ethyl-1,2,4-trimethylpyrrole with TEMPO
and TMAO
145
ix

LIST OF TABLES
Chapter 2
Table 2.1 Molecular weight of iso[4]LGE2-modified and unmodified
calpain-1 peptide “SVTGAKQVNY”.
42
Table 2.2 Tryptic peptides from GAPDH modified by fluorescently labeled
iso[4]LGE2
51
Chapter 3
Table 3.1 Optimized parameters for MALDI-TOF mass spectrometer. 111
Table 3.2 Optimized parameters for triple quadrupole mass spectrometer. 112
Chapter 5
Table 5.1 Sequential adaptation of hybridoma cells to serum free medium 178
x

LIST OF FIGURES
Chapter 1
Figure 1.1 Pathological effects of LGs and isoLGs. 6
Figure 1.2 Overview of monoclonal antibody production, see text for detailed
description (Copyright from wikipedia commons).
21
Chapter 2
Figure 2.1 Treatment of calpain-1 with 10 equivalents of iso[4]LGE2 for
various times.
38
Figure 2.2 Treatment of calpain-1 with various equivalents of iso[4]LGE2 for
30 min
39
Figure 2.3 MS of iso[4]LGE2-modified and unmodified calpain-1 peptide
“SVTGAKQVNY” with the modification site at K280.
42
Figure 2.4 Tandem MS characterization of the m/z 690.88 doubly charged
ion from a MS scan of chymotrypsin digested iso[4]LGE2-
modified calpain-1 showed a series of fragment ions that
unambiguously identify an iso[4]LGE2 modification on the lysyl
residue (K280).
43
Figure 2.5 Ball-and-stick and cartoon 3D model structures for the catalytic
subunit of calpain-1(pdb: 2ary).
45
Figure 2.6 The active sites of human minicalpain 1 are shown in stick
representation and labeled (left panel).
46
Figure 2.7 Western analysis of iso[4]LGE2 (50 equivalents) modified calpain- 47
xi

1 after reaction for 1h, 3h, 12h and 24h. Left panel: developed
with anti-calpain-1 catalytic subunit primary antibody.
Figure 2.8 A ball-and-stick and cartoon 3D model structure for catalytic
subunit of GAPDH (pdb: 1j0x).
52
Figure 2.9 Coomassie blue staining of 10 equivalents fluorescently labeled
iso[4]LGE2 modified GAPDH after incubation for various times
prior to addition of excess glycine.
53
Chapter 3
Figure 3.1 MALDI-TOF analysis of the reaction mixture from N-
acetylglycyllysine methyl ester and iso[4]LGE2 (molar ratio =
200:1) at 25°C for 4 days.
90
Figure 3.2 LC-ESI for the product mixture from the reaction of N-
acetylglycyllysine methyl ester with iso[4]LGE2 (molar ratio =
200:1 at 25 °C for 4 days) with selected ion recording for aminal
cross-link, lactam-H2O and bispyrrole. A) TIC, B) aminal cross-
link m/z 854.1 (+ H+), C) lactam-H2O m/z 574.7 (+ H+), D)
bispyrrole m/z 1150.5 (+ H+).
91
Figure 3.3 MALDI-TOF spectra: panel A and B for 36 min HPLC fractions,
panel C for 30 min HPLC fraction, panel D for 41 min HPLC
fraction of products from the reaction of N-acetylglycyllysine
methyl ester with iso[4]LGE2 (molar ratio = 200:1 at 25 °C for 4
and 15 days).
93
xii

Figure 3.4 Top: Structures of A2E (1), the bisoxirane (2) and nonaoxirane
(3). Bottom: ESI mass spectrum of A2E after illumination with
blue light (430 nm; 10 min) to yield a mixture of various
oxygenated derivatives resulting in a series of ions that differ in
m/z by 16 owing to the addition of oxygen atoms to A2E. The
addition of oxygen presumably to form epoxides, occurred at
carbon-carbon double bonds of A2E, measured by Micromass
ESI-Q-TOF.
94
Figure 3.5 ESI-MSMS of molecular ions at A) m/z 835 (+ H+), and B) m/z
853 (+ H+) corresponding to an iso[4]LGE2-(N-acetyl-gly-lys-
OMe) aminal cross-link and its dehydrated form.
96
Figure 3.6 MALDI-TOF analysis of the product mixture from the reaction of
N-acetylglysyllysine methyl ester with iso[4]LGE2-FluoTag
(molar ratio = 200:1) at 25 °C for 15 days.
98
Figure 3.7 ESI-MSMS of molecular ions at A) m/z 1102.5 and B) m/z 1120.6
corresponding to a iso[4]LGE2-FluoTag-(N-acetyl-gly-lys-OMe)
aminal cross-link adduct and its dehydrated form.
100
Figure 3.8 MALDI-TOF analysis of the product mixture from the reaction of
β-amyloid(11-17) peptide “EVHHQKL” with iso[4]LGE2 (molar
ratio = 200:1) 25 °C for 7 days.
101
Figure 3.9 LC-ESI for the reaction mixture from β-amyloid(11-17) peptide
and iso[4]LGE2 (molar ratio = 200:1 at 25 °C for 7 days) with
selected ion recording for bispyrrole and aminal cross-links. A)
103
xiii

TIC, B) bispyrrole m/z 804.6 (+3), m/z 603.7 (+4), C) aminal
cross-link m/z 1058.2 (+2).
Figure 3.10 MALDI-TOF spectra: panel A and B for 32 min HPLC fractions,
panel C for 22 min HPLC fraction of products from reaction of the
β-amyloid(11-17) peptide “EVHHQKL” with iso[4]LGE2 reaction
(molar ratio = 200:1 at 25 °C for 7 and 15 days).
105
Figure 3.11 MALDI-TOF analysis of the product mixture from the reaction β-
amyloid (EVHHQKL) peptide with iso[4]LGE2-FluoTag (molar
ratio = 200:1) at 25 °C for 15 days.
107
Chapter 4
Figure 4.1 MALDI-TOF analysis of iso[4]LGE2-pyrrole and oxidation
products generated upon short term exposure of the reaction
mixture from iso[4]LGE2 with N-acetylglycyllysine methyl ester
in the presence of dithionite to air or longer term aeration with
oxygen before or after removal of dithionite.
125
Figure 4.2 LC-ESI for the purification of iso[4]LGE2-pyrrole with selected
ion recording for iso[4]LGE2-pyrrole.
126
Figure 4.3 Time course of the oxidative reactions of pure iso[4]LGE2-pyrrole
under air at 25 °C for 8 days.
128
Figure 4.4 MALDI-TOF of pure iso[4]LGE2-pyrrole and its oxidation
products: lower trace = pure iso[4]LGE2 -pyrrole, upper trace =
after incubation under air at 25 °C for 8 days.
129
xiv

Figure 4.5 MALDI-TOF spectra of pure iso[4]LGE2-pyrrole reaction with N-
acetylcysteine under air.
131
Figure 4.6 MALDI-TOF of pure iso[4]LGE2-pyrrole and its reaction with N-
acetylcysteine.
132
Figure 4.7 MALDI-TOF MS of pure iso[4]LGE2-pyrrole reaction with N-
acetylcysteine under air at 25 °C for 8 days.
133
Figure 4.8 MALDI-TOF MS of pure iso[4]LGE2-pyrrole and its oxidation
products: lower trace = pure iso[4]LGE2-pyrrole, upper trace =
after exposure to 2 mol% APS and 1 mol% TEMED at 37 °C for
2 h.
139
Figure 4.9 MALDI-TOF MS of pure iso[4]LGE2-pyrrole and its oxidation
products: lower trace = pure iso[4]LGE2-pyrrole, upper trace =
after exposure to 10 mol% TEMPO at 37 °C for 3 h.
141
Chapter 5
Figure 5.1 Mouse serum immune responses (1:10,000 dilutions) comparison
with pre-immune serum.
162
Figure 5.2 SDS-PAGE analysis of purified anti-iso[4]LGE2 mAb with protein
G column.
165
Figure 5.3 Binding inhibition of protein G purified anti-iso[4]LGE2 mAb
from serum free medium by iso[4]LGE2-HSA, iso[4]LGE2-KLH,
LGE2-BSA, LGE2-KLH, CEP-BSA, CEP-HSA, BSA and HSA.
166
Figure 5.4 Comparative Western analyses using anti-iso[4]LGE2 mAb against 167
xv

iso[4]LGE2-BSA, a standard protein containing the iso[4]LGE2
modification.
xvi

ACKNOWLEDGEMENTS
I wish to express my deepest sense of gratitude and respect to my research advisor Dr.
Robert G. Salomon for his expert guidance, constant encouragement, and the amount of
time he spent to help me to become a professional researcher over the years. He provided
me with the best possible facilities, collaborations, opportunities and challenging tasks,
which enable me to enhance my research aptitude and build a concrete foundation for my
future goals.
I also wish to express my deepest thanks to Dr. John W. Crabb, Cole Eye Institute,
Cleveland Clinic Foundation, who co-supervised me in last three years of my graduate
education. His invaluable guidance in protein chemistry and mass spectrometry, and
active participation in the research projects enabled me to achieve many of the results in
this thesis. Thanks to people in his research group, Lei Zhang and Jack Crabb for
valuable training and many insightful discussions. Special thanks to Dr. Geeng-Fu Jang,
he was there throughout my studying and always willing to help whenever I had
questions or problems. He has often helped me come out of one or more of the tight
corners in my projects. Above all I thank them for their friendship and guidance.
I would like to thank my committee members, Dr. Michael G. Zagorski, Dr. James
Burgess, Dr. Rajesh Viswanathan, Dr. John Crabb for their valuable time, efforts and
suggestions in my candidate qualification and thesis.
I would like to thank Dr. Salomon’s group, former and current members: Dr. James
Laird, Dr. Mikhail D. Linetsky, Dr. Xiaodong Gu, Dr. Jaewoo Choi, Dr. Li Hong, Dr.
Yunfeng Xu, Dr. Hua Wang, Dr. Xiaoxia Z. West, Dr. Wenyuan Yu, Dr. Yu Zhang, Dr.
xvii

Yalun Cui, and Junhong Guo, Liang Xin, Yuanyuan Qian, Guangyin Wang, Nicholas D.
Tomko, Yu-Shiuan Cheng for their thoughtful discussions and their friendship.
I would like to thank Dr. Irina Pikuleva of Department of Ophthalmology and Visual
Sciences for giving me the opportunity to work in her lab, and Dr. Casey Charvet for
valuable training of protein modification at the beginning of my research. I also wish to
thank Denice Major of Hybridoma Cole Facility of Visual Sciences Research Center
(VSRC) on the project of monoclonal antibody development.
I would like to thank Dr. Jim Faulk and other staff members in Chemistry Department
in Case Western Reserve University for their help in numerous occasions.
Finally, this thesis is dedicated to my family, especially my mom, my husband, for
their constant support, endless love and encouragement throughout the course of my
education.
xviii

LIST OF ABBREVIATIONS AND ACRONYMS
Abbreviations and Acronyms Equivalent
AA arachidonic acid AAA amino acid analysis AA-PC 2-arachidonyl-phosphatidylcholine Aβ amyloid β ABTS 2,2-azino-bis (3-ethylbenzthiazoline-6-
sulphonic acid diammonium salt AcOH acetic acid AMD age-related macular degeneration APS ammonium persulfate BSA bovine serum albumin CDCl3 deuterated chloroform CEP carboxy ethylpyrrole 13C-NMR carbon magnetic resonance CDI 1,1'-Carbonyldiimidazole CID collision-induced dissociation COX cyclooxygenase Cys cysteine DBD-ED 4-(N,N-Dimethylaminosulfonyl)-7-(2-
aminoethylamino)-2,1,3-benzoxadiazole kDa or kD kilodalton DCC dicyclohexylcarbodiimide
DMEM dulbecco’s modified eagle’s medium
xix

DMSO dimethyl sulfoxide DNA deoxyribonucleic acid DPDS 2,2'-Dipyridyldisulfide DTT dithiothreitol EDCI N-(3-Dimethylaminopropyl)-N′-
ethylcarbodiimide hydrochloride EDTA ethylenediaminetetraacetic acid EGTA ethylene glycol tetraacetic acid ELISA enyzeme-linked immunosorbant assay ESI electrospray ionization ETN ethanolamine FBS fetal bovine serum FTICR fourier transform ion cyclotron resonance GAPDH glyceraldehyde-3-phosphate dehydrogenase Gly glycine GSH reduced L-glutathione HAT hypoxanthine-aminopterin-thymidine 1H-NMR Proton magnetic resonance HNE 4-hydroxy-2-nonenal HoBt 1-hydroxybenzotriazole HPLC high performance liquid chromatography HRP horseraddish peroxidase HSA human serum albumin HUVEC human umbilical vein endothelial cell
xx

Hz hertz IC50 inhibitor concentration at the 50% absorbance
value IgG immunoglobin G iso[4]LGE2 iso[4]levuglandin E2 isoLG isolevuglandin isoLGE2 isolevuglandin E2
isoLGD2 isolevuglandin D2 KLH keyhole limpet hemocyanine
LC liquid chromatography LC/MS-MS liquid chromatography-coupled electrospray
tandem mass spectrometry LDL low-density lipoprotein LG levuglandin LGD2 levuglandin D2 LGE2 levuglandin E2 Lys lysine mAb monoclonal antibody
MALDI-TOF matrix assisted laser desorption ionization-
time of flight
NAPE-PLD N-acyl phosphatidylethanolamine-hydrolyzing phospholipase D
MeOH methanol MRM multiple reaction monitoring MS/MS tandem mass spectroscopy
xxi

NEM N-ethyl morpholine
NMR nuclear magnetic resonance OHdiA 4-oxo-heptanedioic amide ox oxidized pAb polyclonal antibody PAGE polyacrylamide gel electrophoresis PBS phosphate buffered saline PE phosphatidylethanolamine PEG polyethylene glycol PGs prostaglandins PGD2 prostaglandin D2 PGE2 prostaglandin E2 PGG2 prostaglandin endoperoxide G2 PGH2 prostaglandin endoperoxide H2 PM pyridoxamine ppm parts per million PUFAs polyunsaturated fatty acid(s) PVDF polyvinylidene fluoride QTof2 quadrupole-time of flight Rf retention factor RBC red blood cell ROS reactive oxygen species RPE retinal pigmented epithelium SET single electron transfer
xxii

SDS sodium dodecyl sulfate TEMED tetramethylethylenediamine TEMPO tetramethylpyrrolidine-N-oxide TFA trifluoroacetic acid TLC thin layer chromatography
TM trabecular meshwork TMAO trimethylamine-N-oxide TMS trimethyl silyl UV ultraviolet
xxiii

Part I: Isolevuglandin-protein Cross-linking: Structure and Mechanism
Part II: Generation and Characterization of a Monoclonal
Isolevuglandin[4]E2-protein Adduct Antibody
Abstract
By
WENZHAO BI
IsoLG modification of proteins is associated with loss-of-function, cross-linking and
aggregation. The sites of 1:1 adduct formation and the types and extent of cross-linking
between subunits of the multi subunit proteins calpain-1 and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) by iso[4]LGE2 were characterized mass spectroscopically and
by SDS-PAGE. Modification of one or two lysyl residues calpain-1 suffices to abolish
activity. LC-MS/MS analysis established that Lys280 is a target of isoLG adduction to
calpain-1, Lys3 and Lys249 are targets of fluorescently labeled iso[4]LGE2 adduction to
GAPDH. Hetero- and homo-oligomers of calpain-1 catalytic and regulatory subunits
were detected by SDS PAGE.
N-Acetyl-glycyl-lysine methyl ester and β-amyloid(11-17) peptide “EVHHQKL”
were used as models for characterizing the cross-linking of protein lysyl residues
resulting from adduction of iso[4]LGE2. Aminal, bispyrrole and trispyrrole cross-links of
two model peptides, were identified and fully characterized by mass spectrometry, and a
large variety of derivatives of the bispyrrole cross-link containing one or more additional
atoms of oxygen were discovered. In sharp contrast with the dipeptide, the reaction of
xxiv

“EVHHQKL” with iso[4]LGE2 strongly favored bispyrrole versus aminal crosslink
formation. It is tempting to speculate that the “EVHHQKL” peptide-pyrrole modification
forms aggregates that favor the dimerization to produce bispyrrole since β-amyloid tends
to spontaneously oligomerize.
I discovered that the pure LG/isoLG-pyrrole derivative of the N-acetyl-lys-gly methyl
ester dipeptide could be produced in the presence of dithionite, an oxygen scavenger. The
oxidation and dimerization of the pure pyrrole is promoted by oxygen but exhibits an
induction period that can be eliminated by various catalysts, e.g., the stable free radical
tetramethylpyrrolidine-N-oxide or by trimethylamine-N-oxide.
In the presence of N-acetylcysteine, incubation of iso[4]LGE2-pyrrole at neutral pH
under air showed no pyrrole-to-pyrrole cross-linking. Instead a series of new ions was
produced corresponding to pyrrole cysteine adducts and a series derivatives incorporating
one or more atoms of oxygen. It is suggested that the thiol inhibits the pyrrole
dimerization by intercepting pyrrole cation radical generated by electron transfer to
oxygen.
An anti-iso[4]LGE2 mAb was prepared that exhibits high structural specificity for
detecting iso[4]LGE2–protein adduct epitope, and no significant cross-reactivity with
analogs including carboxyethylpyrrole (CEP) or LGE2-protein adduct epitopes.
xxv

CHAPTER 1
INTRODUCTION

1.1 Lipid Peroxidation
Lipids are essential and major components of cell membranes, and play important
roles in signaling pathways,1-2 energy storage, and gene expression.3-4 Oxidative
degradation of lipids occurs when there is an increase in the level of reactive oxygen
species which damages the cell membrane and leads to subsequent cell death. The
oxidation of polyunsaturated fatty acids (PUFAs) by enzymatic or non-enzymatic
reactions with molecular oxygen is termed “lipid peroxidation”.
Lipid peroxidation products have been implicated in a number of disease
conditions such as amyotrophic lateral sclerosis,5 age-related macular degeneration
(AMD),6-7 end-stage renal disease (RD),8-9 Alexander’s,10 Alzheimer’s,11-13 and
Parkinson’s14 diseases. It is also suspected to involve in the pathogenesis of
atherosclerosis (AS),15-16 antiphospholipid antibody syndrome,17 rheumatoid
arthritis,18 inflammatory bowel disease19 and multiple sclerosis.20 However, our
understanding of these processes on a molecular level remains primitive. Research in
Dr. Salomon’s group is making major strides to fill this gap. Our approach aims to
understand the biologically important chemistry of oxidized lipids and to determine
the extent and consequences of this chemistry in the pathogenesis of human diseases.
1.2 Enzymatic Pathways
Arachidonyl phospholipids, e.g., phosphatidyl choline with a palmityl or steryl
ester on the C1 hydroxyl and a arachidonyl ester on the C2 hydroxyl (AA-PC), a
linear twenty carbon methylene interrupted polyunsaturated fatty acid (C20:4ω6)
ester, is one of the most abundant polyunsaturated phospholipids in human low
2

density lipoprotein (LDL). It undergoes lipid peroxidation by two different pathways,
enzymatic (cyclooxygenase or lipoxygenase) and free radical mediated processes,
generating a vast array of biologically active oxidized products. The free fatty acid is
the substrate for enzyme-mediated peroxidation, while the free radical pathway
operates predominantly on esters.21
(CH2)3COOH
C5H11
710
13
AA
2O2 cyclooxygenase
(CH2)3COOHC5H11
OH
8
PGH2
OHC
(CH2)3COOHC5H11
OH
8
OO
(CH2)3COOHC5H11
OH
8
O
HO
1512
1
PGE2
O
LGE2
1
129
(CH2)3COOHC5H11
OH
8OHC
LGD2
1
129
O
(CH2)3COOHC5H11
OH
8HO
O
1512
1
PGD2
Scheme 1.1 Cyclooxygenase oxidation of AA generates PGs and LGs via
rearrangement of PGH2.
In the cyclooxygenase pathway, arachidonic acid (AA) is oxidized in a
stereospecific manner leading to the formation of the prostaglandin endoperoxide
3

PGH2. Besides enzyme-mediated transformations, a nonenzymatic spontaneous
rearrangement of PGH2 generates prostaglandins PGD2 and PGE2 (scheme 1.1).
Dr. Salomon made the discovery22-24 that PGH2 undergoes rearrangement via an
alternative pathway to form two γ-ketoaldehydes or levuglandins named LGE2 and
LGD2 (Scheme 1.1). The word levuglandin comes from the combination of
levulinaldehyde and prostaglandin because they are derivatives of levulinaldehyde
with prostanoid side chains.25 This rearrangement involves the intramolecular
migration of a bridgehead hydride to an electron deficient incipient methyl group with
concomitant cleavage of a C-C and O-O bond as shown in Scheme 1.2.26-27
Scheme 1.2 Schematic representation of rearrangement of PGH2 to levuglandin.
1.3 The Free Radical Pathway
Unlike the enzymatic pathway that converts free AA to an endoperoxide
intermediate, the free radical pathway mainly oxidizes AA-PC directly, rather than
free AA.25 It produces stereo and structural isomers of prostaglandins28 which were
named isoprostanes.29 Consequently, the free radical pathway is also called the
isoprostane pathway. This pathway also generates racemic mixtures of 16 LG
diastereomers, which a referred to collectively as isoLGE2 and isoLGD2 (Scheme 1.3).
Free radical-induced cyclooxygenation also generates 8 diastereomers of each of 6
4

structurally isomeric analogues that we designate collectively as iso[n]LGs. They are
structurally unique and give protein adducts that can only be products of free radical-
induced lipid oxidation.
(CH2)3COOR
C5H11
OH
OHC
(CH2)3COOR
C5H11
OH
O
OO
O
OO
OC
(CH2)nCH3
OP
O N(CH3)3O O
C5H11
(CH2)3COOR
OH
OO
OO
OO
OH
(CH2)3COOR (CH2)3COOR
OH
C5H11
(CH2)3COOR
OHOHC OHC OHC
OH
(CH2)3COOR (CH2)3COOR
OH
C5H11
O OO
C5H11
C5H11
(CH2)3COOH
C5H11
OH
N
H3C
(CH2)3COOH
OH
OH
(CH2)3COOH(CH2)3COOH
OH
C5H11
C5H11
NNN
H3CH3C
H3C
NH2 NH2NH2 NH2
AA-PC
8-epi-PGH2-PC
12
8-epi-LGE2-PC
iso[4]-PGH2-PC
LGE2-pyrrole
-2 H2O protein -2 H2O protein-2 H2O protein -2 H2O protein
proteinprotein
proteinprotein
8
710
13
8
iso[10]-PGH2-PC iso[7]-PGH2-PC
iso[4]-LGE2-PC iso[10]-LGE2-PC iso[7]-LGE2-PC
iso[4]-LGE2-pyrrole iso[10]-LGE2-pyrrole iso[7]-LGE2-pyrrole
2 O22 O2 2 O2
2 O2
OHC (CH2)3COOR
C5H11
OH
(CH2)3COOR
OH
OHCOHC OHC
OH
(CH2)3COOR (CH2)3COOR
OH
C5H11
C5H11
(CH2)3COOH
C5H11
OH
N(CH2)3COOH
OH
OH
(CH2)3COOH(CH2)3COOH
OH
C5H11
C5H11
NNN
NH2NH2
NH2 NH2
O OO O
H3C H3CH3C
H3C
12
8-epi-LGD2-PC
LGD2-pyrrole
-2 H2O protein-2 H2O protein
-2 H2O protein -2 H2O protein
proteinprotein
proteinprotein
8
iso[4]-LGD2-PC iso[10]-LGD2-PC iso[7]-LGD2-PC
iso[4]-LGD2-pyrrole iso[10]-LGD2-pyrrole iso[7]-LGD2-pyrrole
Scheme 1.3 Generation of isolevuglandins and isolevuglandin-protein adducts during
free radical oxidation of AA-esters
The studies described in this thesis focused on iso[4]LGE2, an isomer formed
exclusively through the free radical oxidation of AA-PC. LGs and isoLGs are highly
reactive aldehydes and initially modify the ε-amino group of lysyl residues in proteins
within seconds30 to form Schiff base intermediates that subsequently afford pyrroles
5

by dehydration.31 The pyrroles are readily oxidized producing lactams and
hydroxylactams.32-34 LGs/isoLGs covalent binding with proteins is accompanied by
cross-linking35 and hence polymerization of proteins orders of magnitude more
rapidly than other reactive arachidonate metabolites such as malondialdehyde and 4-
hydroxynon-2-enal. LGs/isoLGs also cause DNA-protein cross-links (DPCs) which
are repair-resistant and were shown to be relevant to cell killing (Scheme 1.4).36
O
O
R2
DNA
R1
NH2
Levuglandin
ON
R2Schiff base
Protein
R1
Protein NH2
N
H3C OH
NH
Protein-protein Crosslink
Protein
Protein
R1
R2
ON
R2Protein
R1
NHProtein
H
N
H3C OH
NH
DNA-protein Crosslink
Protein
DNA
R1
R2
ON
R2Protein
R1
NHDNA
H
N
H3C
Pyrrole
Bispyrrole Crosslink
Protein
O2 ROO
N
H3C
Protein
OH
OHydroxylactam
+
N
H3C
Protein
OLactam
R1
R2
R1
R2
R1
R2
N
N
R1
R2
R2
R1
Protein
Protein
H2O
Scheme 1.4 Formation of LG-protein adducts, protein-protein and DNA-protein
cross-links.
6

LGs and isoLGs have since been implicated in a number of diseases such as age-
related macular degeneration (AMD), atherosclerosis (AS) and cardiovascular disease
(CVD).25, 37-39 The possible pathological effects of LGs and isoLGs are summarized in
Figure 1.1.
Arachidonyl Phospholipids
Prostaglandin Endoperoxide
PGH2
Endoperoxides from phosphlipids
LGs
isoLGs
PGD2
PGE2
Proteins Covalent Adducts
Protein-protein Cosslinks
DNA-protein Cosslinks
Malfunction of protein and DNA
Cell death and tissue damage
Aggregation & proteasomal malfunction
Figure 1.1 Pathological effects of LGs and isoLGs.
1.4 Biological Activities of Levuglandins and Isolevuglandins
1.4.1 The effects of LGs/isoLGs on protein activity
IsoLG-mediated loss of protein activity has been reported.40 IsoLG, generated in
the epicardial border zone of the canine healing infarct, potentiated inactivation of
cardiac Na+ channels in human embryonic kidney (HEK)-293 cells and cultured atrial
(HL-1) myocytes, suggesting Na+ channel dysfunction evoked by lipid peroxidation
was a candidate mechanism for ischemia-related conduction abnormalities and
arrhythmias.41 In addition to the sodium channel, addition of synthetic isoLG/LG to
an atrial tumor myocyte cell line, AT-1, led to a pronounced dose-dependent
inhibition of the inward rectifying potassium current induced by a -40 mV voltage
step (IC50 = 2.2 μm).42 Oxidative stress reduces the activity of isolated Na+, K+-
ATPase enzymes. In the pioneering study of Shattock et al.,43 research was performed
to determine the effects of oxidant stress on the Na(+)-K+ pump current with isolated
rabbit ventricular myocytes using the whole-cell voltage-clamp technique. Through
7

photoactivation of rose bengal, singlet oxygen and superoxide were generated and
isolated cells were exposed to the induced oxidant stress. Results showed that the
Na+-K+ pump current was inhibited by oxidant stress at all voltages and this inhibition
may contribute to ischemia/reperfusion injury.43 The enzyme activity of purified
RNase A and glutathione reductase was found to decrease following a treatment with
isoLGE2 in vitro. An isoLG scavenger, pyridoxamine, protects enzymes against loss
of activity caused adduction by isoLGE2.44
The first direct evidence linking isoLG modification of a specific protein and loss
of enzyme activity in vivo involved the protease activity of calpain-1. Elevated levels
of the enzyme were detected in glaucomatous trabecular meshwork (TM)45 compared
to nonglaucomatous control (calpain-1 extracted from normal eyes), but calpain-1
activity in the glaucomatous trabecular meshwork was greatly reduced, about 50%
less activity than the same amount of protein from normal TM. Elevated levels of
iso[4]LGE2 protein adduct were also observed in glaucomatous astrocytes isolated
from human optic nerve.46
In an in vitro study, iso[4]LGE2 modified-calpain-1 was ubiquitinated by a HeLa
cell-derived system and the ubiquitinated calpain-1 could be efficiently recognized by
HeLa cell-derived constituted proteasome 26S. Dynamic light scattering studies that
were performed to monitor the size of aggregates confirmed the presence of large
aggregates that correspond to ubiquitinated iso[4]LGE2-modified calpain-1.
Furthermore, these aggregates were refractory toward degradation. Human eyes
possibly lack the capacity to degrade iso[4]LGE2-modified proteins, resulting in their
8

accumulation.45 Glaucoma is thought to be caused in part by an increase in intraocular
pressure (IOP), however normal IOP glaucoma is also common. Pressure increases
either when too much aqueous humor fluid is produced or by decreased aqueous
humor outflow. The trabecular meshwork is responsible for most of the outflow of
aqueous humor. The inhibition of proteasome activity using epoxomicin on cultured
human primary TM cells indicated increased calpain-1 accumulation, owing to
protein modification by isoLGs. This could contribute to glaucoma pathophysiology
by decreasing the ability of the TM to modulate outflow resistance.45 Given the
importance of ubiquitination for proteasomal clearance of LG/isoLGs modified
proteins, it is noteworthy that ubiquitin was shown to form adducts with LGE2.45
The mitochondrial enzyme cytochrome P450 27A1 (CYP27A1) plays a very
important role in the maintenance of cholesterol homeostasis, bile acid biosynthesis,
and activation of vitamin D347 by catalyzing the C27-hydroxylation of cholesterol and
other sterols. Recently CYP27A1 was found to be expressed in the retina48-49 and
shown to be the major contributor to enzymatic degradation of cholesterol.50
Treatment of CYP27A1 with iso[4]LGE2 in vitro diminished enzyme activity in a
time- and phospholipid-dependent manner.51 To model membrane insertion,
CYP27A1 was reconstituted into various PLs. It was found that the enzyme is
modified even when other biological amines are present and iso[4]LGE2
concentrations are relatively low (2-folder molar excess). Liposomes of various
composition were as follows: bovine retinal mitochondrial PLs, a DLPC (1,2-
dilauroyl-snglycero-3-phosphocholine) and DOPE (1,2-dioleoyl-snglycero-3-
9

phosphoethanolamine) mixture (4:3, w/w) that approximates the ratio of
phosphatidylcholine to phosphatidylethanolamine found in human hepatic
mitochondria, or DLPC that contains no free amines and is a standard model PL. In
this set of experiments, the ratio of iso[4]LGE2 was a 2-fold molar excess. Enzyme
activity was decreased sharply by 75% at 5 min and the P450 content decreased only
by 25%, indicating that modification impaired activity without completely denaturing
the protein. Most modification and loss of activity occurred within 15 min of
incubation of a 2-fold molar excess of iso[4]LGE2 with the protein. In contrast,
incorporation of the protein into liposomes containing ethanolamine phospholipids
protected against modification owing to interception of isoLGs by the primary amino
groups of the ethanolamine phospholipids.
Lys 358-isoLG adduct in CYP27A1 was found in human AMD retinal sample. To
study the effect of Lys 358 modification on the enzyme activity, mutations were
introduced to the wild-type (WT) CYP27A1. CYP27A1 Lys 358 was mutated to
either hydrophobic Leu to mimic removal of the positive charge or to Arg to conserve
the charge but abolish iso[4]LGE2 modification because Arg is at least 1,000-fold less
susceptible than Lys to modification by isoLGs. The catalytic activity of the Lys 358
CYP27A1 mutant was characterized before and after isoLG treatment. It was
confirmed that modification of Lys 358 by isoLG is the major contributor to isoLG-
associated loss of CYP27A1 activity. This was first study to conduct the measurement
of enzyme activity and simultaneously correlate the isoLG modification to specific
locations in the primary sequence. The Lys 358-isoLG adduct in CYP27A1 was found
10

in human retina from individuals afflicted with age-related macular degeneration,52
providing direct evidence that isoLG adduction impairs enzyme activity and
supporting the hypothesis that isoLG modification of CYP27A1, potentially impairs
cholesterol homeostasis in the retina.52
1.4.2 LGs/isoLGs effect on the interaction of histones with DNA
DNA-protein cross-linking by a levuglandin was first observed upon treatment of
V79 Chinese hamster lung fibroblasts or nuclei with LGE2.36 DNA in the cell lysate
was retained on a nitrocellulose filter, presumably owing to covalent DNA-protein
cross-linking. This was confirmed by proteolysis with proteinase K that allowed the
DNA to pass through the filter. DNA tightly coils around histones in cells. The lysine-
rich histones from calf thymus could form adducts with isoLG/LG. It was postulated
that this protein is a likely candidate for DNA-isoLG-protein cross-linking.53 Recently,
the formation of LG-lysine lactam adducts on histones were identified in RAW264.7
mouse macrophages, A549 cultured lung epithelial cells and rat liver, that is
dependent on COX-2 activity or COX-2 expression. With an internal LG-lysyl
standard, highest measurable amounts of adduct was detected on the H4 histone.
Adduction was blocked by a γ-ketoaldehyde scavenger (4-ethylsalicylamine), which
had no effect on COX-2 activity as measured by PGE2 production. Formation of LG-
histone adduct was associated with an increased histone solubility in aqueous NaCl
and after LG-histone adduct formation, histone-DNA binding was disrupted with an
increased DNA extraction in a salt solution. Chromatin access and transcription was
regulated by a “histone code comprised with complex patterns of lysyl acetylation and
11

methylation.54 It is reasonable to conclude that irreversible adduction of lysyl residues
could disrupt this code, or directly alter the access of DNA-interacting proteins.
Changes in histone modifications are known to result in altered DNA methylation,
deregulation of oncogenes, genomic instability, impaired DNA repair, and defects in
cell cycle checkpoints.55-56 Changes in lysyl modifications of H4 in particular are a
common hallmark of human cancers, and are associated with a global loss of DNA
methylation.57 Further elucidation of the effects of LG-histone adduction on histone
modification, DNA-histone interactions, and transcription are expected to increase the
understanding of the molecular mechanisms whereby COX-2 contributes to cancer
development and progression.
1.4.3 LGs/isoLGs covalently modify PE
Besides the ε-amino groups of protein lysyl residues, iso[4]LGE2 covalently binds
ethanolamine phospholipids in vitro to form covalent pyrrole adducts that were
oxidized in air to deliver stable lactam and hydroxylactam (HL) adducts.58 In lipid
extracts from human plasma and mouse liver, isoLG-lysoPE-HL derivatives of
isoLGs was detected through phospholipase A2 (PLA2)-catalyzed hydrolysis of
isoLG-diacyl-PE precursors.58 The formation of isoLG/LG-PE is of interest since
oxidatively modified phospholipids have been implicated in certain autoimmune
diseases. Treatment both of human embryonic kidney (HEK293) and human
umbilical vein endothelial cells (HUVEC) with isoLGE2 (15-E2-isoketal, isoK)
generated more modified PE (isoLGE2-PE) than modified protein. As internal
standard, isoLGE2-[2H4] ethanolamine was incubated with HEK293 cells and
12

isoLGE2-PE was quantified by LC/MS/MS after hydrolysis to isoLGE2-ethanolamine
by Streptomyces chromofuscus phospholipase D. Analysis of isoLGE2-protein adducts
as isoLGE2-lysyl-lactam adduct after protease digestion was performed on the protein
pellet using isoLGE2-[13C615N2]lysyl-lactam as the internal standard. Human
umbilical vein endothelial cells (HUVEC) were treated the same way. IsoLGE2-PE
induced cytotoxicity towards human umbilical vein endothelial cells (HUVEC) with
LC50 as 2.2 µM. These observations indicate that cellular PE is a significant target of
isoLGs.59
Another study showed that PE modified by isoLGE2 (isoLGE2-PE) induced THP-
1 monocyte adhesion to human umbilical cord endothelial cells. IsoLGE2-PE also
induced expression of adhesion molecules and increased MCP-1 and IL-8 mRNA in
human umbilical cord endothelial cells and this pyrrole-PE markedly altered
membrane curvature.60 To test if cells could degrade isolevuglandin modified
phosphatidylethanolamine (isoLG-PE), the stability of isoLG-PE in human embryonic
kidney 293 (HEK293) cells and in human umbilical cord endothelial cells (HUVEC)
was measured over time. HEK293 cells were incubated with isoLGE2 to generate
isoLG-PE and excess isoLG was removed by washing with DMEM. With isoLGE2-
[2H4] ethanolamine as internal standard, isoLGE2-PE levels were measured by LC-MS
after conversion to its phosphoglycero-ethanolamine derivative by base hydrolysis.
The stability of isoLGE2-PE in endothelial cells was assessed the same way. Cellular
isoLGE2-PE levels in HEK293 cells decreased more than 75% after 6 h. While in
13

HUVEC cells, the isoLGE2-PE levels rapidly dropped with less than 5% of isoLGE2-
PE present after 24 h.63
N-Acyl phosphatidylethanolamine-hydrolyzing phospholipase D (NAPE-PLD) is
a key enzyme for the hydrolysis of N-acyl phosphatidylethanoamine (NAPE)
including both NAPE and isoLGE2-PE with large aliphatic headgroups. It hydrolyzes
isoLGE2-PE to isoLGE2-ethanolamine. These results demonstrate that NAPE-PLD
contributes to the degradation of isoLGE2-PE and suggest that a major physiological
role of NAPE-PLD may be to degrade aldehyde-modified PE, thereby preventing the
accumulation of these harmful compounds.61
1.4.4 LG/isoLG scavengers
Pyridoxamine (PM) is a B6 vitamin which reacts with isoLGs 2000-fold faster
than the ε-amino groups of protein lysyl residues, making it an efficient scavenger of
this type of lipid peroxidation products.62 Therefore, PM and its analogues can be
used as isoLG/LG scavengers to protect proteins from modification and inactivation
by isoLGs/LGs and to determine the impact on pathophysiology of specifically
reducing the levels of isoLG/LG protein adduction. Treatment with PM and its
analogs, salicylamine and 5’-O-pentylpyridoxamine, had been shown to prevent
H2O2-mediated cytotoxicity in HepG2 cell culture and decrease isoLG-protein adduct
formation of ovalbumin exposed to isoLG. PM also prevented inhibition of RNase A
and glutathione reductase activity by the γ-ketoaldehyde isoLGE2. RNase activity and
glutathione reductase (GR) activity were determined by measuring the formation of
14

acid-soluble oligonucleotide63 and the initial rate of NADPH consumption,64
respectively. 44, 65
Recent research showed that exposure of mice to bright light leads to isoLG-
adduct formation in the retina.66 Pre-treatment of mice with PM greatly decreased
retinal isoLG adduct levels resulting from light exposure, and morphological changes
in photoreceptor mitochondria were not as pronounced as in untreated animals. This
study demonstrated a novel concept in vision research, i.e., that preventing damage to
biomolecules by lipid peroxidation products is a viable strategy to combat oxidative
stress in the retina. This novel therapeutic activity of PM may be applicable to
humans as PM has good drug-like properties and an excellent safety profile.66
1.4.5 Detection of biological adducts of LGs/isoLGs
Detecting free LG/isoLG adducts in vivo is complicated by their proclivity to
behave like superglue, adhering to normal physiological proteins within seconds. On
the other hand, there are two complimentary methods for quantifying LG/isoLG
adducts in vitro/vivo. One method utilizes antibody based approaches such as ELISA,
Western blotting, or immunohistochemistry. The other uses mass spectroscopic
detection of (1) LG/isoLG modified lysine12 or peptides51 derived from LG/isoLG
modified proteins or of (2) LG/isoLG modified ethanolamine phospholipids.58 The
first detection of LG/isoLG adducted proteins in vivo was in patients with
atherosclerosis or end-stage renal disease using ELISA measurements.39
Immunoassays with antibodies raised against protein adducts of LGs generated by
unambiguous chemical synthesis provided evidence for their presence in human and
15

mouse blood and tissues and enabled the discovery that free radical-induced oxidation
of arachidonyl phospholipids in low-density lipoprotein generates LGE225 as well as
stereo and structural isomers referred to collectively as isolevuglandins.38 A single-
chain antibody from a phage displayed recombinant ScFv library that bound a model
peptide adducted with synthetic isoLGE2 was isolated and characterized. Recognition
of isoLGE2-protein adducts by this anti-isoLGE2 adduct single-chain antibody is
essentially independent of the amino acid sequence of the adducted peptides or
proteins, and no cross-reactivity was detected with 4-hydroxynonenal or 4-
oxononanal adducts or with 15-F2t-isoprostane (8-iso-prostaglandin F2α).67 However,
cross reactivity with structurally isomeric isoLGs was not determined.
The second method for detection of isoLGE2-modified proteins utilizes the
sensitivity and specificity of liquid chromatography-mass spectrometry quantitation of
the excised LG-modified lysine following exhaustive proteolysis of LG/isoLG-
adducts in tissues. A heavy isotope labeled internal standard is added to the sample
for quantification.68 MS analysis revealed that the majority of LG/isoLG-protein
modifications are lactams and hydroxylactams generated by oxidation of the initial
pyrrole modifications.32 LC–MS/MS analysis also detects isoLG-
phosphatidylethanolamine adducts in human blood and mouse liver.58 Site-specific
post-translational protein modifications can be detected and quantified using mass
spectrometry-based multiple reaction monitoring (MRM) assay.69 With this method,
the isoLG-modified tryptic peptide AVLKETLR in a mitochondrial protein, Cyp27A1,
16

extracted from human retina was detected and shown to be an adduct of the lysyl ε–
amine group.51
1.5 Monoclonal Antibody Development
1.5.1 Antigens and antibodies
Substances that are recognized by B-cell receptors are termed antigens.
Classically, an antigen is defined as any substance that elicits an immune response in
a susceptible animal and is capable of binding with the specific antibodies generated.
Antigens are usually of a high molecular weight and are commonly proteins or
polysaccharides, although nucleic acids, lipids and peptides have also been reported to
function as antigens.70 Antibodies are produced by plasma B-lymphocytes and
function as a part of an immune system in mammals in the battle against disease.
Antibodies have a common structure of four peptide chains, consisting of two
identical light chains, of about 25 KDa and two identical heavy chains of about 50
KDa. Each light chain is bound to the heavy chain by a disulfide bond and a
combination of hydrogen bonds, salt linkages and hydrophobic bonds. Similarly
noncovalent interactions and disulfide bridges link two identical heavy and light chain
combinations to each other to form a basic four chain immunoglobulin structure.71
Antibodies (Ab)s play an important role when foreign antigens (Ag)s, such as
pathogens and toxins, are introduced into an organism and start a complex immune
response. Antibodies label the foreign molecule or pathogen for destruction and
removal in a lock-and-key type of mode. The basic principle of any immunological
technique is that an antibody will combine with its specific antigen to give a unique
17

Ab/Ag complex. For an efficient Ag/Ab interaction to occur the epitope must be
exposed and available for binding, alterations in the conformation of epitopes through
tissue processing, fixation, reduction and pH changes may affect the binding. In
modern medical diagnostics and basic research, antibodies have become crucial
molecules since it is possible to develop specific antibodies against almost any
component, from small drug molecules to intact cells.
1.5.2 Polyclonal antibodies
Prior to 1975, polyclonal serum was the only antibody option available.
Polyclonal antibodies refer to antibodies present in the crude serum of an immunized
animal, such as a mouse, rabbit, goat or hen, capable of recognizing one or several
different immunogenic epitopes of the administered immunogen. These antibodies
may be of different subclasses, but isolation of a single subclass, usually IgG, is
readily achieved. Polyclonal antibodies are often used since they are easily purified
from the blood of the mammal by chromatographic techniques. Their utilization is
quick and inexpensive, requiring little skill or technical expertise. They are
particularly useful when amplification of a signal from a target protein with low
expression is required as they recognize multiple epitopes on one protein. However,
despite these advantages a pAb raised against an antigen can bind several different
epitopes on the target. This increases the likelihood that pAbs cross-react with
biomolecules containing similar epitopes. Anomalous results and variability between
batches often leads to inconsistencies in results. Furthermore, the supply of pAbs is
limited for research purposes as the mammal is eventually killed.
18

1.5.3 Monoclonal antibodies and hybridoma technology
A major milestone in immunological research was the development of
monoclonal antibodies by Köhler and Milstein in 1975.72 This Nobel Prize winning
work (1984, physiology and medicine) revolutionized antibody production and today
it forms the basis of many diagnostic applications, disease therapies and basic
research.70, 73 Production of monoclonal antibodies (mAbs), in contrast with pAbs, is
more time consuming and expensive from pAb production and requires high
technology and extensive technical skills. Monoclonal antibodies are only one
subclass, usually IgG, allowing for selection of an appropriate secondary antibody for
detection. Large quantities of specific antibodies can be produced and their specificity
ensures that only one epitope is recognized on an antigen. This is extremely useful
when studying subtle protein alterations, and the antibodies are less likely to cross
react with other proteins and generally produce less background cross reaction.
Hybridoma technology involves fusion of a mouse myeloma cell line with mouse
spleen cells from an immunized donor in the presence of polyethylene glycol (PEG).74
The resultant cells are grown in a selective medium in which only hybridoma cells
can proliferate.75 The resultant hybridoma cell line possesses the immortal growth
properties of the cancerous plasma cells and the antibody secreting properties of the
normal B cells, thus creating an indefinitely growing cell line that produces large
quantities of antibodies with identical specificity.
The fusion is random and fused hybridoma cells need to be selected and isolated
from unfused B-lymphocytes and myeloma cells. The cells are cultured in
19

hypoxanthine-aminopterin-thymidine (HAT) medium. Aminopterin (A) blocks the de
novo biosynthesis of purines and pyrimidines which are essential for DNA synthesis.
When this pathway is blocked, cells use the salvage pathway utilizing hypoxanthine
(H) and thymidine (T), and this requires the activity of the enzymes thymidine kinase
(TK) and hypoxanthine-guanine phosphoribosyl transferase (HGPRT). The myelomas
used for the fusion lack HGPRT, so that unfused myeloma cells and myeloma cells
fused to other myeloma cells cannot proliferate in HAT medium. The unfused
splenocytes do possess HGPRT but have a limited lifetime and will die within two
weeks. The hybridoma cells grow effectively in the HAT medium. Many different
hybridomas are created during the fusion and every cell type produces a specific
antibody towards a wide range of antigens and not only the antigen used to immunize
the mouse. To identify the hybridomas that are specific for the antigen, cells are
distributed in 96-well plates and hybridoma supernatant is used in an enzyme-linked
immunosorbent assay (ELISA) to detect the wells containing antibodies that bind with
the antigen (positive wells) for subsequent cloning by limiting dilution.76 The method
essentially consists of diluting the cells and growing them at very low densities, often
in the presence of feeder cells, which supply growth factors. After screening, positive
wells containing only one visible clone are selected for another round of cloning.
After at least three rounds of cloning, the cell line is per definition monoclonal.
Cloning continues until a stable hybridoma, i.e., that does not lose the mAb-producing
ability, is achieved. The stable cloned hybridomas can then be expanded for large-
scale mAb production. Hybridomas can be frozen for storage and in this way an
20

unlimited supply of mAb’s can be maintained (Figure 1.2). mAb may also be stored
frozen but may be come nonspecific inactive with long term storage.
Figure 1.2 Overview of monoclonal antibody production, see text for detailed
description (Copyright from wikipedia commons).
21

1.6 References
1. Alberts, B., Molecular Biology of the Cell. 3rd ed.; Garland Pub: New York,
1994.
2. Cooper, G. M., The Cell: A Molecular Approach. . ASM Press; Sinauer
Associates: Washington, D. C., 1997.
3. Clarke, S. D. (2001) Polyunsaturated fatty acid regulation of gene
transcription: a molecular mechanism to improve the metabolic syndrome. J Nutr 131
(4), 1129-1132.
4. Ntambi, J. M.; Bene, H. (2001) Polyunsaturated fatty acid regulation of gene
expression. J Mol Neurosci 16 (2-3), 273-278; discussion 279-284.
5. Davies, K. J. (1995) Oxidative stress: the paradox of aerobic life. Biochem Soc
Symp 61, 1-31.
6. Gu, X. R.; Meer, S. G.; Miyagi, M.; Rayborn, M. E.; Hollyfield, J. G.; Crabb,
J. W.; Salomon, R. G. (2003) Carboxyethylpyrrole protein adducts and
autoantibodies, biomarkers for age-related macular degeneration. J Biol Chem 278
(43), 42027-42035.
7. Crabb, J. W.; Miyagi, M.; Gu, X. R.; Shadrach, K.; West, K. A.; Sakaguchi,
H.; Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G.
(2002) Drusen proteome analysis: An approach to the etiology of age-related macular
degeneration. Proc Natl Acad Sci U S A 99 (23), 14682-14687.
8. Salomon, R. G.; Batyreva, E.; Kaur, K.; Sprecher, D. L.; Schreiber, M. J.;
Crabb, J. W.; Penn, M. S.; DiCorletoe, A. M.; Hazen, S. L.; Podrez, E. A. (2000)
22

Isolevuglandin-protein adducts in humans: products of free radical-induced lipid
oxidation through the isoprostane pathway. Biochim Biophys Acta 1485 (2-3), 225-
235.
9. Lindner, A.; Charra, B.; Sherrard, D. J.; Scribner, B. H. (1974) Accelerated
atherosclerosis in prolonged maintenance hemodialysis. N Engl J Med 290 (13), 697-
701.
10. Castellani, R. J.; Perry, G.; Harris, P. L.; Cohen, M. L.; Sayre, L. M.;
Salomon, R. G.; Smith, M. A. (1998) Advanced lipid peroxidation end-products in
Alexander's disease. Brain Res 787 (1), 15-18.
11. Sayre, L. M.; Zelasko, D. A.; Harris, P. L.; Perry, G.; Salomon, R. G.; Smith,
M. A. (1997) 4-Hydroxynonenal-derived advanced lipid peroxidation end products
are increased in Alzheimer's disease. J Neurochem 68 (5), 2092-2097.
12. Boutaud, O.; Ou, J. J.; Chaurand, P.; Caprioli, R. M.; Montine, T. J.; Oates, J.
A. (2002) Prostaglandin H2 (PGH2) accelerates formation of amyloid beta1-42
oligomers. J Neurochem 82 (4), 1003-1006.
13. Zagol-Ikapitte, I.; Masterson, T. S.; Amarnath, V.; Montine, T. J.; Andreasson,
K. I.; Boutaud, O.; Oates, J. A. (2005) Prostaglandin H2-derived adducts of proteins
correlate with Alzheimer's disease severity. J Neurochem 94 (4), 1140-1145.
14. Clausen, J. (1984) Demential syndromes and the lipid metabolism. Acta
Neurol Scand 70 (5), 345-355.
15. Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P.
J.; Shan, L.; Febbraio, M.; Hajjar, D. P.; Silverstein, R. L.; Hoff, H. F.; Salomon, R.
23

G.; Hazen, S. L. (2002) A novel family of atherogenic oxidized phospholipids
promotes macrophage foam cell formation via the scavenger receptor CD36 and is
enriched in atherosclerotic lesions. J Biol Chem 277 (41), 38517-38523.
16. Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P.
J.; Shan, L.; Gugiu, B.; Fox, P. L.; Hoff, H. F.; Salomon, R. G.; Hazen, S. L. (2002)
Identification of a novel family of oxidized phospholipids that serve as ligands for the
macrophage scavenger receptor CD36. J Biol Chem 277 (41), 38503-38516.
17. Horkko, S.; Miller, E.; Dudl, E.; Reaven, P.; Curtiss, L. K.; Zvaifler, N. J.;
Terkeltaub, R.; Pierangeli, S. S.; Branch, D. W.; Palinski, W.; Witztum, J. L. (1996)
Antiphospholipid antibodies are directed against epitopes of oxidized phospholipids.
Recognition of cardiolipin by monoclonal antibodies to epitopes of oxidized low
density lipoprotein. J Clin Invest 98 (3), 815-825.
18. Mapp, P. I.; Grootveld, M. C.; Blake, D. R. (1995) Hypoxia, oxidative stress
and rheumatoid arthritis. Br Med Bull 51 (2), 419-436.
19. Grisham, M. B. (1994) Oxidants and free radicals in inflammatory bowel
disease. Lancet 344 (8926), 859-861.
20. Toshniwal, P. K.; Zarling, E. J. (1992) Evidence for increased lipid
peroxidation in multiple sclerosis. Neurochem Res 17 (2), 205-207.
21. Salomon, R. G. (2005) Distinguishing levuglandins produced through the
cyclooxygenase and isoprostane pathways. Chem Phys Lipids 134 (1), 1-20.
22. Salomon, R. G.; Miller, D. B. (1985) Levuglandins: isolation,
characterization, and total synthesis of new secoprostanoid products from
24

prostaglandin endoperoxides. Adv Prostaglandin Thromboxane Leukot Res 15, 323-
326.
23. Salomon, R. G.; Salomon, M. F. (1977) 2,3-Dioxabicyclo [2.2.1] heptane. The
strained bicyclic peroxide nucleus of prostaglandin endoperoxides. J Am Chem Soc 99
(10), 3501-3503.
24. Coughlin, D. J.; Salomon, R. G. (1977) Synthesis of thermal reactivity of
some 2,3-dioxabicyclo[2.2.1]heptane models of prostaglandin endoperoxides. J Am
Chem Soc 99 (2), 655-657.
25. Salomon, R. G.; Subbanagounder, G.; Singh, U.; O'Neil, J.; Hoff, H. F. (1997)
Oxidation of low-density lipoproteins produces levuglandin-protein adducts. Chem
Res Toxicol 10 (7), 750-759.
26. Zagorski, M. G.; Salomon, R. G. (1980) Prostaglandin endoperoxides. 11.
Mechanism of amine-catalyzed fragmentation of 2,3-dioxabicyclo[2.2.1]heptane. J
Am Chem Soc 102 (7), 2501-2503.
27. Zagorski, M. G.; Salomon, R. G. (1982) Prostaglandin endoperoxides. 12.
Carboxylate catalysis and the effects of proton donors on the decomposition of 2,3-
dioxabicyclo[2.2.1]heptane. J Am Chem Soc 104 (12), 3498-3503.
28. Nugteren, D. H.; Vonkeman, H.; Van Dorp, D. A. Non‐enzymic conversion
of all‐cis 8,11,14‐eicosatrienoic acid into prostaglandin E1. Recl Trav Chim Pays-
Bas 86 (11), 1237-1245.
29. Morrow, J. D.; Hill, K. E.; Burk, R. F.; Nammour, T. M.; Badr, K. F.; Roberts,
L. J., 2nd (1990) A series of prostaglandin F2-like compounds are produced in vivo in
25

humans by a non-cyclooxygenase, free radical-catalyzed mechanism. Proc Natl Acad
Sci U S A 87 (23), 9383-9387.
30. Salomon, R. G.; Jirousek, M. R.; Ghosh, S.; Sharma, R. B. (1987)
Prostaglandin endoperoxides .21. covalent binding of levuglandin-E2 with proteins.
Prostaglandins 34 (5), 643-656.
31. Boutaud, O.; Brame, C. J.; Salomon, R. G.; Roberts, L. J., 2nd; Oates, J. A.
(1999) Characterization of the lysyl adducts formed from prostaglandin H2 via the
levuglandin pathway. Biochemistry 38 (29), 9389-9396.
32. Brame, C. J.; Salomon, R. G.; Morrow, J. D.; Roberts, L. J. (1999)
Identification of extremely reactive gamma-ketoaldehydes (isolevuglandins) as
products of the isoprostane pathway and characterization of their lysyl protein
adducts. J Biol Chem 274 (19), 13139-13146.
33. Roberts, L. J.; Salomon, R. G.; Morrow, J. D.; Brame, C. J. (1999) New
developments in the isoprostane pathway: identification of novel highly reactive γ-
ketoaldehydes (isolevuglandins) and characterization of their protein adducts. FASEB
J 13 (10), 1157-1168.
34. Roberts, L. J., 2nd; Brame, C. J.; Chen, Y.; Morrow, J. D.; Salomon, R. G.
(1999) Formation of reactive products of the isoprostane pathway: isolevuglandins
and cyclopentenone isoprostanes. Adv Exp Med Biol 469, 335-341.
35. Iyer, R. S.; Ghosh, S.; Salomon, R. G. (1989) Levuglandin-E2 cross-links
proteins. Prostaglandins 37 (4), 471-480.
26

36. Murthi, K. K.; Friedman, L. R.; Oleinick, N. L.; Salomon, R. G. (1993)
Formation of DNA-protein cross-links in mammalian cells by levuglandin E2.
Biochemistry 32 (15), 4090-4097.
37. Salomon, R. G. (2005) Levuglandins and isolevuglandins: Stealthy toxins of
oxidative injury. Antioxid Redox Signal 7 (1-2), 185-201.
38. Salomon, R. G.; Kaur, K.; Batyreva, E. (2000) Isolevuglandin-protein Adducts
in Oxidized Low Density Lipoprotein and Human Plasma: A Strong Connection with
Cardiovascular Disease. Trends Cardiovasc Med 10 (2), 53-59.
39. Salomon, R. G.; Subbanagounder, G.; O'Neil, J.; Kaur, K.; Smith, M. A.;
Hoff, H. F.; Perry, G.; Monnier, V. M. (1997) Levuglandin E2-protein adducts in
human plasma and vasculature. Chem Res Toxicol 10 (5), 536-545.
40. Stavrovskaya, I. G.; Baranov, S. V.; Guo, X.; Davies, S. S.; Roberts, L. J.,
2nd; Kristal, B. S. (2010) Reactive gamma-ketoaldehydes formed via the isoprostane
pathway disrupt mitochondrial respiration and calcium homeostasis. Free Radic Biol
Med 49 (4), 567-579.
41. Fukuda, K.; Davies, S. S.; Nakajima, T.; Ong, B. H.; Kupershmidt, S.; Fessel,
J.; Amarnath, V.; Anderson, M. E.; Boyden, P. A.; Viswanathan, P. C.; Roberts, L. J.;
Balser, J. R. (2005) Oxidative Mediated Lipid Peroxidation Recapitulates
Proarrhythmic Effects on Cardiac Sodium Channels. Circ Res 97 (12), 1262-1269.
42. Brame, C. J.; Boutaud, O.; Davies, S. S.; Yang, T.; Oates, J. A.; Roden, D.;
Roberts, L. J., 2nd (2004) Modification of proteins by isoketal-containing oxidized
phospholipids. J Biol Chem 279 (14), 13447-13451.
27

43. Shattock, M. J.; Matsuura, H. (1993) Measurement of Na+-K+ pump current in
isolated rabbit ventricular myocytes using the whole-cell voltage-clamp technique.
Inhibition of the pump by oxidant stress. Circ Res 72 (1), 91-101.
44. Davies, S. S.; Brantley, E. J.; Voziyan, P. A.; Amarnath, V.; Zagol-Ikapitte, I.;
Boutaud, O.; Hudson, B. G.; Oates, J. A.; Roberts, L. J., 2nd (2006) Pyridoxamine
analogues scavenge lipid-derived gamma-ketoaldehydes and protect against H2O2-
mediated cytotoxicity. Biochemistry 45 (51), 15756-15767.
45. Govindarajan, B.; Laird, J.; Salornon, R. G.; Bhattacharya, S. K. (2008)
Isolevuglaridin-modified proteins, including elevated levels of inactive calpain-1,
accumulate in glaucomatous trabecular meshwork. Biochemistry 47 (2), 817-825.
46. Govindarajan, B.; Junk, A.; Algeciras, M.; Salomon, R. G.; Bhattacharya, S.
K. (2009) Increased isolevuglandin-modified proteins in glaucomatous astrocytes.
Mol Vis 15 (114-15), 1079-1091.
47. Pikuleva, I. A. (2006) Cytochrome P450s and cholesterol homeostasis.
Pharmacol Ther 112 (3), 761-773.
48. Lee, J. W.; Fuda, H.; Javitt, N. B.; Strott, C. A.; Rodriguez, I. R. (2006)
Expression and localization of sterol 27-hydroxylase (CYP27A1) in monkey retina.
Exp Eye Res 83 (2), 465-469.
49. Liao, W. L.; Heo, G. Y.; Dodder, N. G.; Reem, R. E.; Mast, N.; Huang, S.;
Dipatre, P. L.; Turko, I. V.; Pikuleva, I. A. (2011) Quantification of cholesterol-
metabolizing P450s CYP27A1 and CYP46A1 in neural tissues reveals a lack of
28

enzyme-product correlations in human retina but not human brain. J Proteome Res 10
(1), 241-248.
50. Mast, N.; Reem, R.; Bederman, I.; Huang, S.; DiPatre, P. L.; Bjorkhem, I.;
Pikuleva, I. A. (2011) Cholestenoic Acid is an important elimination product of
cholesterol in the retina: comparison of retinal cholesterol metabolism with that in the
brain. Invest Ophthalmol Vis Sci 52 (1), 594-603.
51. Charvet, C. D.; Liao, W. L.; Heo, G. Y.; Laird, J.; Salomon, R. G.; Turko, I.
V.; Pikuleva, I. A. (2011) Isolevuglandins and Mitochondrial Enzymes in the Retina
mass spectrometry detection of post-translational modification of sterol-metabolizing
CYP27A1. J Biol Chem 286 (23), 20413-20422.
52. Charvet, C. D.; Laird, J.; Xu, Y.; Salomon, R. G.; Pikuleva, I. A. (2013)
Posttranslational modification by an isolevuglandin diminishes activity of the
mitochondrial cytochrome P450 27A1. J Lipid Res 54 (5), 1421-1429.
53. Boutaud, O.; Brame, C. J.; Chaurand, P.; Li, J. Y.; Rowlinson, S. W.; Crews,
B. C.; Ji, C.; Marnett, L. J.; Caprioli, R. M.; Roberts, L. J.; Oates, J. A. (2001)
Characterization of the lysyl adducts of prostaglandin H-synthases that are derived
from oxygenation of arachidonic acid. Biochemistry 40 (23), 6948-6955.
54. Jenuwein, T.; Allis, C. D. (2001) Translating the histone code. Science 293
(5532), 1074-1080.
55. Shogren-Knaak, M.; Ishii, H.; Sun, J. M.; Pazin, M. J.; Davie, J. R.; Peterson,
C. L. (2006) Histone H4-K16 acetylation controls chromatin structure and protein
interactions. Science 311 (5762), 844-847.
29

56. Fuks, F. (2005) DNA methylation and histone modifications: teaming up to
silence genes. Curr Opin Genet Dev 15 (5), 490-495.
57. Fraga, M. F.; Ballestar, E.; Villar-Garea, A.; Boix-Chornet, M.; Espada, J.;
Schotta, G.; Bonaldi, T.; Haydon, C.; Ropero, S.; Petrie, K.; Iyer, N. G.; Perez-
Rosado, A.; Calvo, E.; Lopez, J. A.; Cano, A.; Calasanz, M. J.; Colomer, D.; Piris, M.
A.; Ahn, N.; Imhof, A.; Caldas, C.; Jenuwein, T.; Esteller, M. (2005) Loss of
acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark
of human cancer. Nat Genet 37 (4), 391-400.
58. Li, W.; Laird, J. M.; Lu, L.; Roychowdhury, S.; Nagy, L. E.; Zhou, R.; Crabb,
J. W.; Salomon, R. G. (2009) Isolevuglandins covalently modify
phosphatidylethanolamines in vivo: detection and quantitative analysis of
hydroxylactam adducts. Free Radic Biol Med 47 (11), 1539-1552.
59. Sullivan, C. B.; Matafonova, E.; Roberts, L. J., II; Amarnath, V.; Davies, S. S.
(2010) Isoketals form cytotoxic phosphatidylethanolamine adducts in cells. J Lipid
Res 51 (5), 999-1009.
60. Guo, L.; Chen, Z.; Cox, B. E.; Amarnath, V.; Epand, R. F.; Epand, R. M.;
Davies, S. S. (2011) Phosphatidylethanolamines modified by gamma-ketoaldehyde
(gammaKA) induce endoplasmic reticulum stress and endothelial activation. J Biol
Chem 286 (20), 18170-18180.
61. Guo, L.; Gragg, S. D.; Chen, Z.; Zhang, Y.; Amarnath, V.; Davies, S. S.
(2013) Isolevuglandin-modified phosphatidylethanolamine is metabolized by NAPE-
hydrolyzing phospholipase D. J Lipid Res 54 (11), 3151-3157.
30

62. Caldes, C.; Vilanova, B.; Adrover, M.; Munoz, F.; Donoso, J. (2011) Phenol
group in pyridoxamine acts as a stabilizing element for its carbinolamines and Schiff
bases. Chem Biodivers 8 (7), 1318-1332.
63. Kalnitsky, G.; Hummel, J. P.; Dierks, C. (1959) [Some factors which affect
the enzymatic digestion of ribonucleic acid]. J Biol Chem 234 (6), 1512-1516.
64. Williams, C. H., Jr, Chemistry and biochemistry of flavoenzymes. Müller, F.,
Ed. CRC Press: 1991; pp 121-211.
65. Onorato, J. M.; Jenkins, A. J.; Thorpe, S. R.; Baynes, J. W. (2000)
Pyridoxamine, an inhibitor of advanced glycation reactions, also inhibits advanced
lipoxidation reactions. Mechanism of action of pyridoxamine. J Biol Chem 275 (28),
21177-21184.
66. Charvet, C. D.; Saadane, A.; Wang, M.; Salomon, R. G.; Brunengraber, H.;
Turko, I. V.; Pikuleva, I. A. (2013) Pretreatment with pyridoxamine mitigates
isolevuglandin-associated retinal effects in mice exposed to bright light. J Biol Chem
288 (41), 29267-29280.
67. Davies, S. S.; Talati, M.; Wang, X.; Mernaugh, R. L.; Amarnath, V.; Fessel,
J.; Meyrick, B. O.; Sheller, J.; Roberts, L. J., 2nd (2004) Localization of isoketal
adducts in vivo using a single-chain antibody. Free Radic Biol Med 36 (9), 1163-
1174.
68. Davies, S. S.; Amarnath, V.; Brame, C. J.; Boutaud, O.; Roberts, L. J., 2nd
(2007) Measurement of chronic oxidative and inflammatory stress by quantification
31

of isoketal/levuglandin gamma-ketoaldehyde protein adducts using liquid
chromatography tandem mass spectrometry. Nat Protoc 2 (9), 2079-2091.
69. Kitteringham, N. R.; Jenkins, R. E.; Lane, C. S.; Elliott, V. L.; Park, B. K.
(2009) Multiple reaction monitoring for quantitative biomarker analysis in proteomics
and metabolomics. J Chromatogr B Analyt Technol Biomed Life Sci 877 (13), 1229-
1239.
70. Goldsby, R. A.; Kindt, T. J.; Osborne, B. A.; Kiuby, J., Immunology. Fifth
Edition ed.; New York, 2003.
71. Capra, J. D.; Tung, A. S.; Nisonoff, A. (1975) Structural studies on induced
antibodies with defined idiotypic specificities. II. The light chains of anti-p-
azophenylarsonate antibodies from A/J mice bearing a cross-reactive idiotype. J
Immunol 115 (2), 414-418.
72. Kohler, G.; Milstein, C. (1975) Continuous cultures of fused cells secreting
antibody of predefined specificity. Nature 256 (5517), 495-497.
73. Borrebaeck, C. A. (2000) Antibodies in diagnostics - from immunoassays to
protein chips. Immunol Today 21 (8), 379-382.
74. Harlow, E.; Lane, D., Antibodies- A Laboratory Manual. Cold Spring Harbor
Laboratory Press: Cold Spring Harbor, New York, 1988.
75. Little, M.; Kipriyanov, S. M.; Le Gall, F.; Moldenhauer, G. (2000) Of mice
and men: hybridoma and recombinant antibodies. Immunol Today 21 (8), 364-370.
76. Lefkovits, I.; Waldeman, H., Limiting dilution analysis of cells in the immune
system. Cambridge University Press: Cambridge, 1979.
32

CHAPTER 2
ADDUCTION AND CROSS-LINKING OF CALPAIN-1 AND
GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE
(GAPDH) BY ISO[4]LGE2

2.1 Background
2.1.1 Purpose
The first characterization of the site of in vivo isoLG post-translational modification
was of the monomeric mitochondrial protein Cyp27A1. To elucidate isoLG-mediated
protein-protein cross-linking, the reaction of iso[4]LGE2 with multi subunit proteins,
calpain-1 and GAPDH, was investigated. In this study, we characterized sites of
modification and detected cross-linking between subunits of the multi subunit proteins
calpain-1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
2.1.2 Isolevuglandin modification of proteins.
The eye and brain contain high levels of polyunsaturated fatty acids (PUFAs), of
which arachidonic acid is one of the most abundant.1 Arachidonyl phospholipids (PLs)
undergo cyclooxygenase catalyzed oxidation to produce γ-ketoaldehyde levuglandins and
free radical-induced oxidation in vivo to form levuglandins (LGs) and their stereo and
structural isomers referred to collectively as isolevuglandins (isoLGs). LG and isoLG-
protein adducts in human serum have been fully characterized.2 Some isoLG isomers
such as iso[4]levuglandin E2 (iso[4]LGE2) are not produced through the enzymatic
cyclooxygenase pathway, but rather are only generated through free radical-mediated
oxidation.3 On the other hand, a few diastereomers of two isoLG structural isomers, e.g.,
levuglandins E2 and D2, are not only generated through free radical-initiated lipid
oxidation, but are also formed through the cyclooxygenase pathway.4 IsoLGs are a highly
reactive family of γ-keto aldehydes5 that react with free primary amines such as the ε-
amino group of lysine residues in proteins. They form covalent adducts with orders of
magnitude greater avidity than most other lipid oxidation products such as 4-
34

hydroxynonenal.6-8 These adducts incorporate the primary amino group in a pyrrole ring
that is readily oxidized to lactam and hydroxylactam derivatives. In the human eye,
isoLG adducted calpain-1 accumulates in the trabecular meshwork, and isoLG adduction
abolishes the enzyme’s activity.9 LG/isoLG-protein adduction has biological
consequences. For example, preferential adduction of isoLG to a specific Lys in the
mitochondrial enzyme Cyp27A1 abolishes its ability to metabolize cholesterol leading to
cholesterol accumulation and retinal pathology.10-11 Lysyl-LG/isoLG-protein adduction
also results in intermolecular protein-protein cross-linking. Pyrrole-pyrrole cross-link and
aminal cross-link structures depicted in Scheme 2.1 have been postulated for isoLG-
protein modifications, but direct molecular level evidence for the cross-link structure(s) is
lacking.12
C5H11
COO-PC
O
O
OH
OHC C5H11
(CH2)3COO-PCO
OH
isoPGH2-PC
isoLGE2-PC
COO-PC
PAPCR1
R2
OHN
CH3
hemiaminal
R1
R2
N
H3C
O2
N
H3C X
Opyrrole
isoLGE2-lactam, X = HisoLGE2-hydroxylactam, X = OH
(CH2)3COOH
C5H11
OH
O2O
O
OHiso[4]PGH2-PC
COO-PC
C5H11
OH
COO-PC
C5H11OHC
O
iso[4]LGE2-PC
PLA2
N
H3C X
O
iso[4]LGE2-lactam, X = Hiso[4]LGE2-hydroxylactam, X = OH
OH
COOH
C5H11
protein NH2
protein
protein
protein
protein
R1
R2N
H3C
pyrrole-pyrrole crosslink
protein
R1
R2N
H3C
protein
O2
OH
protein NH2
R1
R2
HO
N
CH3
aminal crosslink
protein
protein
R1
R2
OHN
CH3
protein
NH
-H2O
protein NH
Scheme 2.1 Postulated structures of LG and iso[4]LG protein adducts and cross-links.
35

2.1.3 Calpain-1.
Calpains are ubiquitous calcium-dependent cysteine proteases that mainly exist in two
forms in all tissues, µ-calpain and m-calpain. They belong to a superfamily of 14 cysteine
proteases. Activation of calpains in vitro requires the presence of calcium at micromolar
quantities for µ-calpain and millimolar quantities for m-calpain at neutral pH.13-14 Both
forms comprise a large subunit of about 80 kDa and a small subunit of about 30 kDa.
Increased calpain activity has been discovered in several neurological disorders, such as
spinal cord injury, ischemic brain injuries, stroke, and calpain activity has been
implicated in several neurodegenerative disorders such as Alzheimer’s disease and
multiple sclerosis.14 IsoLG-protein adducts accumulate in trabecular meshwork (TM)
tissue in individuals with primary open angle glaucoma. Inactive calpain-1 accumulates
in the TM in conjunction with massive levels of isoLG-modified protein. Treatment with
isoLG inactivates calpain-1. There are elevated levels of the protease calpain-1 in
glaucomatous TM based on western analysis. However, calpain-1 activity in
glaucomatous TM is only about 50% of that in controls. This paradox is explicable by the
fact that modification by isoLGs renders calpain-1 inactive. In addition, isoLG-
modification of calpain-1 promotes its ubiquitination and the formation of aggregates that
resist proteasomal processing9, 159, 159, 159, 15,9, 15 and presumably contribute to the impeded
aqueous humor outflow through the TM that results in elevated intraocular pressure. It
appears that calpain-1 is expressed to promote proteolysis of damaged proteins. However,
this function is defeated by isoLG-modification that renders it inactive. In addition,
isoLG modification can also make proteins resistant to proteolysis, e.g., by the 20S
36

proteasome.16 Thus, isoLG adduction presumably contributes to accumulation and
inactivation of calpain in the TM owing to aggregation and/or resistance to proteolysis.
2.1.4 GAPDH
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) plays a key role in the
glycolytic pathway.17 GAPDH catalyzes the oxidative phosphorylation of
glyceraldehyde-3-phosphate (G3P) in the presence of NAD+ and inorganic phosphate.
GAPDH is well conserved during evolution, being a protein with a native molecular
weight in the range of 140-150 kDa. It is composed of four identical of 35-37 kDa
subunits.18 The ubiquity and evolutionary conservation of GAPDH indicate a highly
important physiological function.19 Besides its traditional role in glycolysis, newly
recognized activities of GAPDH include regulation of the cytoskeleton,20 membrane
fusion and transport,21 glutamate accumulation into presynaptic vesicles,22 and activation
of transcription in neurons.23 Elevated levels of nuclear GAPDH are observed in
postmortem samples from patients with Parkinson's disease, Alzheimer's disease,
Huntington's disease and glaucoma, among others.24 4-Hydroxy-2-nonenal (HNE), a
major lipid peroxidation-derived reactive aldehyde, exerts an inhibitory effect on
GAPDH due to the modification of the cysteine residue (Cys-149) at the catalytic site
where it forms an HNE-cysteine Michael addition type adduct.25 Elevated iso[4]LGE2
protein adduct immunoreactivity is also found in glaucomatous astrocytes derived from
the glaucomatous optic nerve head.26
37
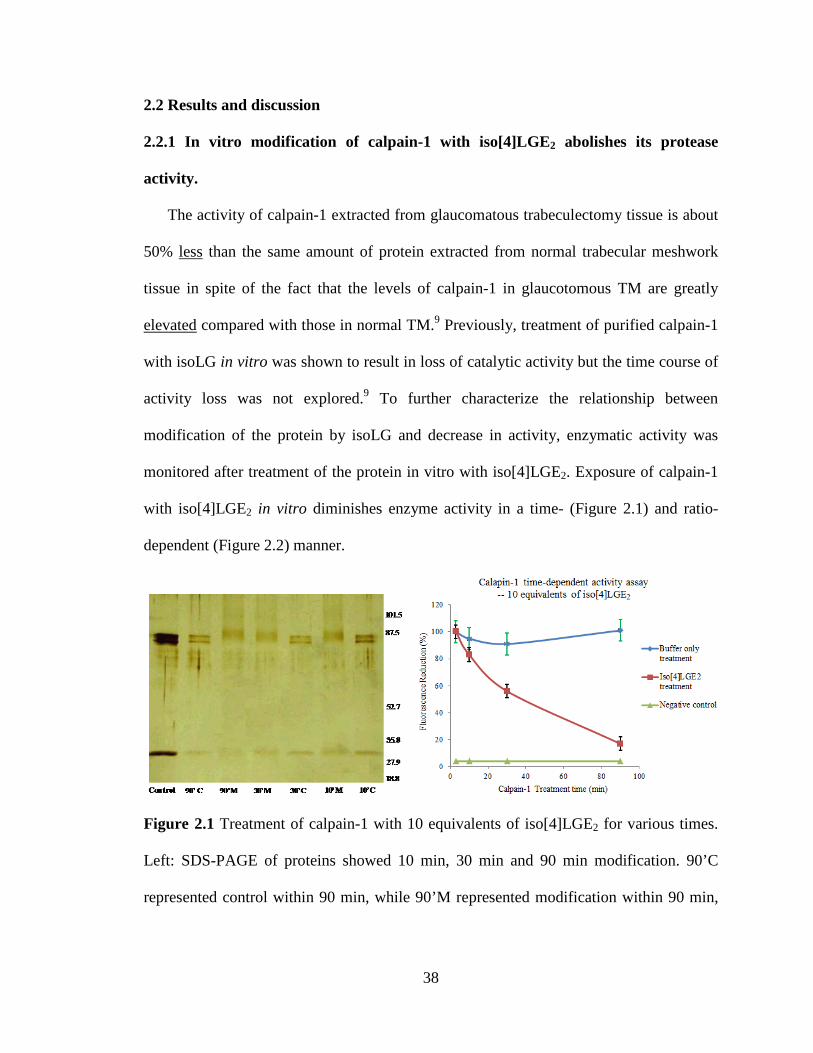
2.2 Results and discussion
2.2.1 In vitro modification of calpain-1 with iso[4]LGE2 abolishes its protease
activity.
The activity of calpain-1 extracted from glaucomatous trabeculectomy tissue is about
50% less than the same amount of protein extracted from normal trabecular meshwork
tissue in spite of the fact that the levels of calpain-1 in glaucotomous TM are greatly
elevated compared with those in normal TM.9 Previously, treatment of purified calpain-1
with isoLG in vitro was shown to result in loss of catalytic activity but the time course of
activity loss was not explored.9 To further characterize the relationship between
modification of the protein by isoLG and decrease in activity, enzymatic activity was
monitored after treatment of the protein in vitro with iso[4]LGE2. Exposure of calpain-1
with iso[4]LGE2 in vitro diminishes enzyme activity in a time- (Figure 2.1) and ratio-
dependent (Figure 2.2) manner.
Figure 2.1 Treatment of calpain-1 with 10 equivalents of iso[4]LGE2 for various times.
Left: SDS-PAGE of proteins showed 10 min, 30 min and 90 min modification. 90’C
represented control within 90 min, while 90’M represented modification within 90 min,
38

same for 30 and 10 min. Right: calpain-1 activity assay with different modification time
treatment.
Notably, the inhibitory effect was not linearly related to isoLG-protein ratio. Two
equivalents of isoLG had little additional inhibitory effect over a 1:1 molar ratio and the
inhibition caused by a 10 equivalents of isoLG to protein was less than twice that of 2
equivalents. This behavior suggests that a low level of modification by one or two
molecules of isoLG has a profound effect on activity, and that additional modifications
only provide minor additional inhibitory effects. The time dependence of the effect of
exposure of the protein to 10 equivalents of iso[4]LGE2 was examined. Enzyme activity
decreased by 50% by 30 min and gradually decreased to 10% by 90 min while the
activity of the control, treated with buffer alone, was unchanged. Analysis by
electrophoresis showed that within 10 min after treatment, both the calpain-1 catalytic
and small subunits migrated at slightly higher mass than the untreated protein presumably
because they had been covalently modified.
Control2 x 5 x 10 x2025
37
50
75
100
150
250
Figure 2.2 Treatment of calpain-1 with various equivalents of iso[4]LGE2 for 30 min.
Control2 x 5 x 10 x2025
37
50
75
100
150
250
KDa
equivalent of iso[4]LGE2
39

Left: SDS-PAGE of calpain-1 after treatment with 2, 5 or 10 equivalents of iso[4]LGE2.
Right: calpain-1 activity after treatment with 2, 5 or 10 equivalents of iso[4]LGE2.
The ratio dependence of the effect of exposure of the protein to a 2, 5 10 equivalents
of iso[4]LGE2 was also examined. Enzyme activity decreased sharply by 50% within 30
min. Analysis by electrophoresis showed that 30 min after treatment, both the calpain-1
catalytic and small subunits migrated slightly less than the untreated protein presumably
because they had been covalently modified.
In the pioneering study of Govindarajan et al.,9 isoLG modified calpain-1 was
precipitated by cold acetone, and resuspended in PBS for enzyme activity test. Our study
aims to find specific sites of modification by isoLG and to test enzyme activity at the
same time, so calpain-1 modification was performed in sodium acetate solution. The
reaction was quenched by addition of glycine and the mixture used directly for enzyme
activity assay and MS analysis.
2.2.2 Calpain-1 Lys280 is modified upon exposure to isoLGs.
IsoLG modification of proteins, e.g., calpain-1, is associated with loss-of-function,
cross-linking and aggregation. To elucidate isoLG-mediated protein-protein cross-linking,
the reaction of iso[4]LGE2 with calpain-1 was examined in detail. A site of isoLG
modification of calpain-1 was identified by peptide mapping and sequencing. Using LC-
MS and Mascot analysis one lysine residue in calpain-1 was identified that was converted
by iso[4]LGE2 into lactam derivatives. A full MS comparison of the iso[4]LGE2 modified
and unmodified calpain-1 chymotryptic peptide “SVTGAKQVNY” shows that K280 is
the modification site (Figure 2.3).
40

Iso[4]LGE2-treated calpain-1 was proteolyzed in-gel with chymotrypsin followed by
FTICR MS/MS. Peak lists from the acquired MS/MS spectra were submitted to the
Mascot data base search engine. Evaluation criteria for identification of the modified
peptides were as follows: first, only hits with a significant peptide score as reported by
Mascot’s scoring algorithm27 were considered. Second, the tandem MS spectra were
manually inspected for the candidates to confirm the existence of y and b series ions
upstream and downstream of the modified lysine. Third, the presence of multiple adduct
dehydration and oxidation states was sought (Table 2.1S), e.g., observation of both the
lactam and the monodehydrated lactam for a given residue. Mascot analysis of the spectra
from the in-gel tryptic digest (31% sequence coverage, encompassing 13% of the Lys
residues) did not identify any modified peptides with a significant score. In contrast,
analysis of the in-gel chymotryptic digest (67% sequence, 28% Lys coverage) identified
one distinct peptide, SVTGAK280QVNY, modified at Lys280 (Table 2.2S). In this
peptide, the modified lysine residue was within the peptide and not at the C terminus,
indicating that modification blocks tryptic cleavage. Thus, based on our evaluation
criteria, we identified one lysine residue in calpain-1 as modified by iso[4]LGE2.
Figure 2.3 shows a full MS comparison of iso[4]LGE2 unmodified and modified
calpain-1 peptide “SVTGAKQVNY” with modification at K280. The mass spectrum of
the modified peptide revealed two forms of adducts, a lactam and a monodehydrated
lactam, both as doubly charged ions (Table 2.1). In contrast, the spectrum of the
unmodified peptide did not exhibit peaks for either of these modifications (Table 2.1).
Due to their hydrophobicity, modified peptides eluted later than the unmodified peptide.
41

It is likely that the loss of water to form the monodehydrated lactam occurred in the mass
spectrometer.
Figure 2.3 MS of iso[4]LGE2-modified and unmodified calpain-1 peptide
“SVTGAKQVNY” with the modification site at K280. Panel A: Detection of the doubly
charged ions m/z 699.88 and 690.88 corresponding to the lactam and monodehydrated
lactam modifications of SVTGAKQVNY. Panel B: Detection of the doubly charged ion
m/z 533.78 corresponding to the unmodified peptide. See table 2.1 for calculated
molecular weights.
Table 2.1 Molecular weight of iso[4]LGE2-modified and unmodified calpain-1 peptide
“SVTGAKQVNY”.
Peptide Sequence Formula of
MW of
ΔMass Found in
Elution
Notes
SVTGAKQVNY No Mod 1065.66 Da Yes 33.18' No Modification
SVTGAKQVNY C20H28O3 1381.20 Da 316.20 Da No ND Pyrrole
SVTGAKQVNY C20H26O2 1363.86 Da 298.20 Da No ND Pyrrole Monodehydrated
42

SVTGAKQVNY C20H28O4 1397.76 332.10 Da Yes 61.54' Lactam
SVTGAKQVNY C20H26O3 1379.76 Da 314.10 Da Yes 61.54' Lactam Monodehydrated
To unambiguously establish the site of modification, MS2 analysis was performed to
study the fragmentation patterns of the modified versus the unmodified chymotryptic
peptide. A series of fragment y-ions and b-ions was detected that identify an iso[4]LGE2
modification on the Lys residue of the calpain-1 peptide SVTGAKQVNY (Figure 2.4).
S V T G A K280(IsoLG Lactam-H2O) Q V N Y
y
b
123456789
y*(2)y(2)
y(3)b(5)
y(4)
b*(9)++, b0(9)++
b(9)++
b0(6)
b(6)
y(5)
b(7)
b0(8)
b(8)
b*(9)
b(9)
m/z
Rel
ativ
e A
bund
ance
m/z of Lys=128.09m/z of lactam dehydrated=314.19128.09 + 314.19=442.28m/z from y4 to y5=442.28
Figure 2.4 Tandem MS characterization of the m/z 690.88 doubly charged ion from a
MS scan of chymotrypsin digested iso[4]LGE2-modified calpain-1 showed a series of
fragment ions that unambiguously identify an iso[4]LGE2 modification on the lysyl
residue (K280). Asterisks denote fragment ions with a modified lysyl residue.
The iso[4]LGE2-modified chymotryptic peptides that we identified enabled
determination of the specific lysyl group that was altered in various samples of
iso[4]LGE2-modified calpain-1. MS/MS analysis consistently detected modification of
the SVTGAKQVNY chymotryptic peptide from the catalytic subunit of iso[4]LGE2-
43

calpain-1 in samples of iso[4]LG-modified calpain-1 that were generated with various
iso[4]LGE2 to protein ratios, showing that this sequence was readily accessed and
covalently altered by this γ-ketoaldehyde electrophile. Figure 2.5 shows ball-and-stick
and cartoon models that provide 3D visualization of calpain-1 using PyMOL v1.4.1. All
lysines of calpain-1 are shown as space-filling models in the left panel while only the
lysine that became incorporated into the iso[4]LGE2-modification and three active site
catalytic triad residues (Cys 115, His 272 and Asn 296) are shown in the right panel
cartoon model. The 3D model shows that the modified lysine is located at and protrudes
from the surface of calpain-1. The physical environment and chemical characteristics of
this specific lysyl residue apparently facilitates access by iso[4]LGE2 and covalent
adduction through Schiff base formation with its aldehyde group followed by cyclization
and pyrrole formation. In vivo, phospholipid derivatives containing the iso[4]LG
esterified to the sn2-hydroxyl protrude like whiskers28 from membrane bilayers and the
surface of lipoprotein particles. These isoLGs esterified to phospholipids adduct to
proteins to initially form Schiff basses and then cyclized to pyrroles. At some point,
phospholipolysis releases the free carboxyl of the isoLG.
44

Figure 2.5 Ball-and-stick and cartoon 3D model models for the catalytic subunit of
calpain-1(pdb: 2ary). All the lysines were shown as space-filling models in the left panel.
Only lysine (280) that was modified by iso[4]LGE2 and three active site amino acid
residues (Cys 115, His 272 and Asn 296) were shown in the right panel.29-31
The architecture of calpain-1 consists of a short N-terminal extension (domain I),32
the catalytic core (dIIa and dIIb), the C2-like domain III,33 and a penta-EF hand domain
IV,31 which interacts with the small subunit domains dV34 and dVI (Figure 2.1S, left
panel).35-36 Subdomain IIa and domain IIb, with a substrate-binding cleft in between,
comprises domain II, also name as minicalpain. The catalytic triad residue Cys 115 is on
subdomain IIa, whereas His 272 and Asn 296 are part of subdomain IIb. In the absence of
calcium, the distance between the catalytic Cys 115 and His 272 (10 Å) is too great to
form a functional catalytic triad. It is suggested that calcium-induced conformational
45

changes draw subdomain IIa and IIb together29, 37-3829, 37-3829, 37-3829, 37-38 (Figure 2.1S, right
panel).29, 37-38 As described above, calpain-1 loses protease activity when modified with
iso[4]LGE2. The modified lysine 280 is located on β strand 11 (amino acid sequence 274-
284) which is close to but not in the calpain-1 active site (His 272). The loss of activity
consequent to modification of this lysine may result from a conformational change
caused by the modification (Figure 2.6).
Iso[4]LGE2
Lys280
β12 β11
β13
Figure 2.6 The active sites of human minicalpain 1 are shown in stick representation and
labeled (left panel).30 Modified lysine 280 and the active site (Cys 115, His 272 and Asn
296) and related β strands are shown in the cartoon 3D model for the catalytic subunit of
calpain-1 (right panel).
2.2.3 In vitro modification of calpain-1 with iso[4]LGE2 generates cross-links.
Human serum albumin (HSA) binds ten equivalents of LGE2 within 1 minute.39 The
initial lysyl-LG adducts with proteins are also reactive, eventually undergoing
intermolecular protein-protein cross-linking, e.g., LGE2 also causes extensive
intermolecular cross-linking of ovalbumin.40
46

The time course of intermolecular cross-linking of calpain-1 was monitored upon
incubation with 2 to 460 equivalents of iso[4]LGE2. The oligomerizations were
terminated after various time intervals by the addition of excess glycine, and the
oligomers were fractionated by SDS-PAGE and Western analysis (see Figure 2.7 and
appendix Figure 2.2S-2.6S). Dimers and higher oligomers were already prominent after 1
h. Primary antibodies against the catalytic and small regulatory subunits as well as anti-
iso[4] monoclonal antibody were used for Western analysis to detect both calpain-1 and
iso[4]LGE2 modified calpain-1. All the images of the gels include the loading well. Only
the incubation with 460 equivalents of isoLG showed an appreciable accumulation of
high molecular weight oligomer in the loading well.
Use Anti-Calpain 1 catalyticsubunit primary antibody
2025
37
50
75
100
150
250
12 h1 h 3 h24 hControl
24 h
240 kDa160 kDa
320 kDa
120 kDa110 kDaCatalytic subunit--80kDa
Modification
Use Anti-Calpain 1 smallsubunit primary antibody
2025
37
50
75
100
150
250
12 h1 h 3 h24 hControl
24 h
140 kDa120 kDa110 kDa
Small Subunit--30kDa
Catalytic subunit--80kDa
Modification
Catalytic subunit--80kDa
Use Anti-Iso[4]LGE2 monoclonal antibody
12 h1 h 3 h 24 h2025
37
50
75
100
150
250
140 kDa120 kDa110 kDa
160 kDa240 kDa
320 kDa
Small Subunit--30kDa
Modification
Figure 2.7 Western analysis of iso[4]LGE2 (50 equivalents) modified calpain-1 after
reaction for 1h, 3h, 12h and 24h. Left panel: developed with anti-calpain-1 catalytic
subunit primary antibody. Middle panel: developed with anti-calpain-1 small subunit
primary antibody. Right panel: developed with anti-iso[4]LGE2 monoclonal primary
antibody.
47

Notably, intense bands for the catalytic subunit, its dimer and trimer at 80, 160 and
240 kDa were detected with anti-calpain-1 catalytic subunit antibody but not with the
regulatory subunit antibody. The dimer and trimer, in contrast with the monomer, were
also not prominently stained by the anti-iso[4]LG antibody suggesting that the antibody
does not detect isoLG involved in cross-linking and that the cross-linked catalytic
subunits do not have extensive additional modification that is not involved in the cross-
link. For example, with 50 equivalents of iso[4]LGE2 (Figure 2.7), Western analysis
revealed that within in 1 h, a dimer, higher oligomers and even higher multimers
appeared. By contrast, the control, calpain-1 in the absence of iso[4]LGE2, exhibited no
oligomerization after incubation for 24 h. The anti-calpain-1 catalytic subunit (80 kDa)
primary antibody detected a band at 110 kDa corresponding to a hetero dimer of one
catalytic subunit with one regulatory small subunit (30 kDa). It also detected bands at 160
kDa, 240 kDa and 320 kDa corresponding to a dimer, trimer and tetramer of the catalytic
subunit. The anti-calpain-1 small subunit primary antibody (30 kDa) also detected a band
at 110 kDa corresponding to a hetero dimer of one catalytic subunit and one small
regulatory subunit. It also revealed a band at 120 kDa corresponding to a tetramer of the
small regulatory subunit, and a band at 140 kDa corresponding to a heterotrimer
consisting of two small regulatory subunits cross-linked with one catalytic subunit. Both
the anti-catalytic subunit and small subunit primary antibodies showed some, albeit low,
cross-reactivity. The anti-iso[4]LGE2 monoclonal antibody detected virtually all oligomer
bands resulting from modification most likely due to non-crosslinked modification.
Although the molecular structure of the protein cross-link was unknown,40-41 the
48

oligomers are resistant to dissociation by SDS and heat under reducing conditions,
indicating a high probability that the cross-link involves covalent bonding.
2.2.4 Preparation of fluorescently labeled iso[4]LGE2.
The identification and quantitation of protein cross-linking have proved to be a
formidable analytical challenge in proteomics. Fluorescence detection-HPLC is an
important method for trace analysis and in combination with mass spectrometry may
provide an approach to analyze isoLG modified protein cross-linking. For this purpose,
stable fluorescently labeled iso[4]LGE2 was developed and tagged to protein through
modification. After digestion, tagged samples were subjected to LC/MS with
fluorescence detection for further analysis.
Preparation of fluorescently labeled iso[4]LGE2 2.4 was achieved in two steps
(Scheme 2.2). Iso[4]LGE2 precursor 2.1 was synthesized as described.3 (4-(N,N-
Dimethylaminosulfonyl)-7-(2-aminoethylamino)-2,1,3-benzoxadiazole) (DBD-ED)42-44 is
a widely used fluorescence labeling reagent which emits strong fluorescence at long
wavelengths. Its 2-aminoethylamine group reacts in the presence of a condensing agent
with carboxylic acids at room temperature to yield a stable amide. The EDCI-HoBt45-49
system was chosen as condensing agent after testing triphenylphosphine (TPP), 2,2'-
Dipyridyldisulfide (DPDS)50-54 and 1,1'-Carbonyldiimidazole (CDI).55 Fluorescently
labeled iso[4]LGE2 intermediate 2.3 was prepared by treating precursor 2.1 with EDCI-
HoBt first and then adding DBD-ED at 25 °C in the dark. After overnight reaction, one
product was formed and unreacted materials could be easily removed by flash silica gel
chromatography. Stirring the acetal 2.3 in dilute acetic acid overnight followed by
49

evaporation of the solvent under high vacuum and flash chromatography accomplished
hydrolysis of the dimethyl acetal to cleanly deliver pure 2.4. MS spectra were shown in
appendix Figure 2.7S.
CO2H
O
(MeO)2HCOH
C5H11
NHCH2CH2NH2
SO2N
NO
N
CH3CH3
EDCI-HOBt
C
O
(MeO)2HCOH
C5H11
ONHCH2CH2NH
NO
N
SO2NCH3
CH3
AcOHH2O
C
O
OH
C5H11
ONHCH2CH2NH
NON
SO2NCH3
CH3
O
2.1
2.2
2.3
2.4
Scheme 2.2 Synthesis of a fluorescently labeled iso[4]LGE2 derivative.
2.2.5 GAPDH Lys3 and Lys249 are modified by fluorescently labeled isoLG.
Fluorescently labeled isoLG was first used to detect the sites of 1:1 lysine-isoLG
adducts, e.g., pyrroles, lactams, hydroxylactams and their derivatives, on a homotetramer
protein glyceraldehyde 3-phosphate dehydrogenase. Fluorescently labeled iso[4]LGE2-
treated GAPDH was trypsinyzed in-solution followed by triple quadrapole MS and Q-Tof
MS/MS analysis of tryptic peptides. Peak lists from the acquired MS/MS spectra were
analyzed with the Mascot data base search engine. Evaluation criteria for identification of
the modified peptides were the same as normal iso[4]LGE2 modified peptides. Lysine
modifications and chemical structures of lys-fluorescently labeled iso[4]LGE2 adducts
were presented in Table 2.4S and Scheme 2.1S. This analysis identified two lysine
residues in GAPDH that were modified by fluorescently labeled iso[4]LGE2 as a
monodehydrated lactam and a bisdehydrated hydroxylactam. MS/MS characterization of
50

the doubly charged ion m/z 620.86 (LEKAAK) and triply charged ion m/z 537.96
(VKVGVNGFGR) showed a series of fragment ions sufficient to unambiguously identify
a fluorescently labeled iso[4]LGE2 modification on the GAPDH lysyl residues K3 and
K249 (Table 2.2 and Table 2.5S).
Table 2.2 Tryptic peptides from GAPDH modified by fluorescently labeled iso[4]LGE2.
Sequence Observed Mr(expt) Ion score Variable Modification
LEK (-C
30H
39N
5O
5S)AAK 620.8596(2+) 1239.7046 18 Lactam Monodehydrated
VK (-C
30H
37N
5O
5S)VGVNGFGR 537.9638(3+) 1610.8696 23 Hydroxylactam Bisdehydrated
MS/MS analysis consistently detected modification on the LEKAAK and
VKVGVNGFGR sequences of fluorescently labeled iso[4]LGE2-GAPDH in preparations
generated with various equivalents of fluorescently labeled iso[4]LGE2, implying that
these sequences were readily accessed and covalently altered by this aldehyde
electrophile. Figure 2.8 showed ball-and-stick and cartoon models providing 3D
visualization of GAPDH using PyMOL v1.4.1. All lysines of GAPDH are shown as
space-filling models in the left panel while only the lysines that becoming adducted by
fluorescently labeled iso[4]LGE2 and active site are shown in the cartoon model in the
right panel. The 3D model shows that these lysines are located at and protrude from the
surface of GAPDH. The physical environment and chemical characteristics of these
specific lysyl residues probably facilitated access by iso[4]LGE2 and covalent adduction
through Schiff base formation with its terminal aldehyde group leading to pyrrole
formation. This is similar to the preference noted above for isoLG modification of
calpain-1.
51

Figure 2.8 A ball-and-stick and cartoon 3D model structure for catalytic subunit of
GAPDH (pdb: 1j0x). All the lysines were shown as space-filling models in the left panel.
Only lysines that became modified by fluorescently labeled iso[4]LGE2 and active site
(Cys 149) were shown in the right panel.55-56
2.2.6 In vitro modification of GAPDH with fluorescently labeled iso[4]LGE2 forms
cross-links.
The time course of intermolecular cross-linking of GAPDH upon incubation with 10
equivalents of fluorescently labeled iso[4]LGE2 was determined by quenching the
oligomerization after various time intervals by the addition of excess glycine, and
fractionating the oligomers by SDS-PAGE. Only the 24 h incubation showed an
accumulation of high molecular weight oligomer in the loading well (Figure 2.9). Within
1 h, a dimer, trimer and even higher multimers appeared. By contrast, GAPDH in the
absence of fluorescently labeled iso[4]LGE2 exhibited little or no oligomerization after
incubation for 24 h. The traces of oligomers that were barely detectable in the control
probably represent minor alternative oligomerization mechanisms.
52

1h 4h2h 8h 24h 24 hControl
monomer
tetramer
trimerdimer
oligomer
263443567295
130
170
Modification
Figure 2.9 Coomassie blue staining of 10 equivalents fluorescently labeled iso[4]LGE2
modified GAPDH after incubation for various times prior to addition of excess glycine.
2.2.7 Pilot studies on cross-linking during modification of GAPDH with
fluorescently labeled iso[4]LGE2.
2.2.7.1 Amino acid analysis GAPDH dimer, trimer and tetramer formation upon
modification with fluorescently labeled iso[4]LGE2.
Amino acid analysis was performed to quantify the percentage of modified GAPDH
cross-linked as dimer, trimer and tetramer contained in a sample. The identity and
quantity of each amino acid was determined by comparing retention time and
chromatographic peak areas detected from the sample with those of a calibration file that
was generated from an amino acid standard mixture (standard H, Pierce).
Dimer, trimer and tetramer fractions of fluorescently labeled iso[4]LGE2 modified
GAPDH were cut from a gel (Figure 2.9) and individually trypsin in gel-digested.
53

AccQ·TagTM amino acid analysis was used to quantify each cross-linked fraction in a
particular sample of GAPDH. First the tryptic peptides from the sample were hydrolyzed
into free amino acids under HCl vapor. Then the amino acids were derivatized using a
Waters AccQ·TagTM ultra derivatization kit. AccQ·TagTM ultra reagent reacts rapidly
with primary amino groups to yield highly stable urea. The structure of the UV active
derivatizing group is the same for all amino acids. Excess reagent hydrolyzes to yield 6-
aminoquinoline (AMQ), a non-interfering by-product.
N
HN O
NO
O
O
NHR1 R2Amino Acid
t 1/2<<1s N
HN N
O
R1
R2
Derivalized Amino Acid
+ N
O
O
HO
N-HydroxysuccinimideH2O t 1/2<<15s
N
NH2+ N
O
O
HO
N-Hydroxysuccinimide
+ CO2
6-aminoquinoline
Scheme 2.3 AccQ·TagTM derivatization.65-66
Finally, the derivatized hydrolysate was chromatographically analyzed. The identity
and quantity of each amino acid was determined by comparing retention time and
chromatographic peak areas with the peak area of the quantitative standard amino acids
(Standard H, 50 pmol, appendix Figure 2.8S). The percent of total protein sample that
corresponds to the fraction analyzed was determined by dividing the amount determined
by the quantitative amino acid analysis of the fraction by the total amount of protein
(GAPDH after 24 h modification and for SDS analysis) in the sample times 100. After 24
h incubation, about 2.5%, 2.5% and 1.5% of GAPDH was present in the sample in the
form of dimer, trimer and tetramer, respectively (appendix Table 2.6S).
54

2.2.7.2 LC/MSMS analysis of fluorescently tagged tryptic peptides
A combination of fluorescence detection-HPLC and mass spectrometry was applied
to analysis of tryptic peptides from an isoLG-modified protein (GAPDH) in an effort to
identify isoLG-cross-linked peptides. To enable detection and facilitate isolation of
isoLG-modified peptides, a fluorescent label was incorporated into the isoLG. Thus,
homo-tetrameric protein GAPDH was modified with stable fluorescently labeled
iso[4]LGE2, trypsinized, and the tryptic peptides were first separated on a reverse phase
column, fractions detected with an Acquity® fluorescence (FLR) detector were analyzed
with a triple quadrupole electrospray mass spectrometer (Applied Biosystems Inc.).
Fluorescence channels of tryptophan peptides (three digested peptides containing
tryptophan) and iso[4]LGE2 tag were both monitored. The fluorescence signal of
tryptophan peptides (280 nm for excitation, 340 nm for emission) and mass spectrometry
(ESI-triple quadrupole) were employed to evaluate the chromatographic separation of
peptides on various LC columns. A column was identified that gave sharp peaks. Then
using that column, the fluorescence signal of iso[4]LGE2 tagged peptides (450 nm for
excitation, 560 nm for emission) was monitored to detect the iso[4]LGE2 modified
peptides (see appendix Figure 2.9S and 2.10S). Peptides with iso[4]LGE2 tag
fluorescence signals were collected as one minute fractions and quickly dried using a
high-speed vacuum evaporator and stored at -20 °C until used. These peptides were re-
suspended in 0.1% formic acid/2% acetonitrile and further analyzed by LC-MS/MS using
a quadrupole-time of flight (QTof2) mass spectrometer. To identify 1:1 isoLG-peptide
adducts, peak lists from the acquired MS/MS spectra were analyzed with the Mascot data
base search engine.
55

To identify 2:1 isoLG-peptide aminal cross-links (Scheme 2.4), peptides with
fluorescence signal were collected as one minute fractions that were analyzed by MS
analysis on QTOF. The masses of all possible tryptic GAPDH peptides with one missing
cleavage were calculated (Table 2.7S). The masses of cross-linked peptide [M] were then
calculated by summing the mass of peptide #1 + mass of peptide #2 + 601.2934 (the mass
increment for the cross-link, Scheme 2.4). The exact masses calculated for the [M+H]+,
[M+2H]2+, [M+3H]3+, [M+4H]4+ ions are shown in Table 2.8S. Since the peptides
“LEKAAK” and “MVKVGVNGFGR” were found to form lysine 1:1 adducts, we
postulated that these two peptides are particularly reactive toward iso[4]LGE2.
Consequently, we considered them to be high probability candidates for forming aminal
cross-links. The molecular weights of aminal cross-links between “MVKVGVNGFGR”
+ “MVKVGVNGFGR”, “LEKAAK”+ “LEKAAK”, “LEKAAK”+ “MVKVGVNGFGR”
for ions from +1 to +4 charges were calculated and compared with all precursor ions
exacted from QTOF MS of each HPLC fraction.
N CC5H11
NHCH2CH2NHO
OH
OH
NH
NO
N
SO2Npeptide #1
peptide #2
= 604. 3169
Scheme 2.4 Chemical structure of peptides aminal cross-link. Mass of cross-linked
peptide = mass of peptide #1 + mass of peptide #2 + 601.2935 (= 604.3169 – 3.0234).
An observed ion of mass 640.9626 corresponds approximately with the m/z 640.
3732 expected for a +3 charged cross-link of the peptide “LEKAAK”+ “LEKAAK”. To
56

test the hypothesis that this ion corresponds to the above mentioned ion, MS2 daughter
ions were examined for various expected fragmentation involving loss of b2 (243), a2
(215) or “AK” (199), “AAK” (270). However, none of the expected fragments were
detected. Therefore, it is unlikely that the parent ion at 640.9626 corresponds to the
proposed cross-link (Figure 2.11S).
To supplement the approach described above, in future studies, modification of
proteins with a 1:1 mixture of d6-iso[4]LGE2 and d0-iso[4]LGE2, can provide additional
evidence that will facilitate the detection and characterization of protein cross-links. If
peptides were modified by both d6-iso[4]LGE2 and d0-iso[4]LGE2, the observation of
characteristic pairs of peaks would provide convincing evidence for the presence of an
isoLG in the ion being observed. The pair of peaks would be separated by +6 amu for
singly charged ions, +3 amu for doubly charged ions, and +2 amu for triply charged ions.
57

2.3 Conclusions
IsoLG modification of proteins is associated with loss-of-function, cross-linking and
aggregation. The sites of 1:1 adduct formation and the types and extent of cross-linking
between subunits of the multi subunit proteins calpain-1 and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) by iso[4]LGE2 were characterized mass spectroscopically and
by SDS-PAGE. Calpain-1 lost its protease activity with t1/2 = ~40 min upon incubation
with 10 equivalents of iso[4]LGE2, and as little as 2 equivalents was almost as effective.
This suggests that modification of one or two lysyl residues suffices to abolish activity.
LC-MS/MS analysis established that Lys280 is a target of isoLG adduction to calpain-1.
Since this lysine residue is not located in the active site, a conformational change induced
by the modification may account for the loss of activity. Covalently cross-linked hetero
and homo oligomers of calpain-1 catalytic and regulatory subunits were detected by SDS
PAGE. The dimer and trimer, in contrast with the monomer, of the catalytic subunit were
prominently stained by an antibody against the catalytic subunit but not by an antibody
against the regulatory subunit or by the anti-iso[4]LG antibody suggesting that the
iso[4]LG antibody does not detect isoLG involved in cross-linking and that the cross-
linked catalytic subunits do not have extensive additional modification that is not
involved in the cross-link.
The amounts of dimer, trimer and tetramer forms of GAPDH produced after 24 h
incubation with 10 equivalents of fluorescently labeled iso[4]LGE2 was determined by
amino acid analysis to be about 2.5%, 2.5% and 1.5% respectively. LC-MS/MS analysis
established that Lys3 and Lys249 are targets of fluorescently labeled iso[4]LGE2
adduction to GAPDH. These lysine residues are located in the tryptic peptides
58

“LEKAAK” and “VKVGVNGFGR”. Since the peptides “LEKAAK” and
“MVKVGVNGFGR” were found to form lysine 1:1 adducts, we postulated that these
two peptides are presumably particularly reactive toward iso[4]LGE2. Consequently, we
considered them to be high probability candidates for forming aminal cross-links.
However, no ions were detected that could be confirmed by MS/MS analysis to
correspond to these cross-linked peptides.
59

2.4 Experimental Procedures
2.4.1 General methods. All chemicals used were high purity analytical grade. The
following commercially available materials were used as received: 4-(N,N-
dimethylaminosulfonyl)-7-(2-aminoethylamino)-2,1,3-benzoxadiazole (DBD-ED) and
2,2'-dipyridyldisulfide (DPDS) were from TCI America (Portland, OR). Calpain-1 (from
human erythrocytes) was from Calbiochem (La Jolla, CA). 1-Hydroxybenzotriazole
(HoBt), iodoacetamide, N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride
(EDCI), 1,1'-carbonyldiimidazole (CDI), and GAPDH (from rabbit muscle) were from
Sigma (St. Louis, MO). Dithiothreitol was from Denville Scientific (South Plainfield NJ).
Chymotrypsin and trypsin were from Promega (Madison, WI). Sodium acetate,
ammonium bicarbonate, disodium hydrogen phosphate and monosodium phosphate were
from Fisher Scientific Co. Iso[4]LGE2 was prepared by Dr. Jim Laird and iso[4]LGE2-
FluoTag was prepared as discussed below.
Chromatography was performed with ACS grade solvent. Thin layer chromatography
(TLC) was performed on glass plates precoated with silica gel (Kieselgel 60 F254, E.
Merck, Darmstadt, West Germany). Rf values are quoted for plates of thickness 0.25 mm.
The plates were visualized by viewing the developed plates under short or long
wavelength UV light, iodine, or by heating the plates after dipping in 20% solution of
phosphomolybdic acid in ethanol. Flash chromatography was performed according to
established methods57 on ICN SiliTech 32-63 D 60A silica gel from ICN Biomedicals
Gmbh, Eschwege, Germany.
60

Software: DeepView from Swiss-PdbViewer (http://spdbv.vital-it.ch/), SWISS-
MODEL Workspace (http://swissmodel.expasy.org/) and PyMOL v1.4.1
(http://pymol.org/)
2.4.2 Synthesis of fluorescently labeled iso[4]LGE2. HOBt (1.48 mg, 11 µmol in 40 µl
DMF), EDCI (2.12 mg, 11 µmol in 56 µl CH2Cl2), and precursor 2.1 (4 mg, 10 µmol in
50 µl CH2Cl2) were stirred at room temperature for 2.5 h. A solution of DBD-ED (11
µmol, 2.84 mg) dissolved in 140 µl of acetonitrile was added to the reaction mixture.
After overnight reaction in the dark, TLC showed only one major product formed and
complete disappearance of precursor 2.1. The reaction mixture was concentrated and the
residue was purified by flash chromatography on a silica gel column (ethyl acetate:
methanol=20:1, TLC: Rf = 0.7) to give the fluorescently tagged iso[4]LGE2
dimethylacetal derivative 2.3 (4.8 mg, 7 µmol, yellow oil). ESI-MS: m/z calcd for
C32H51N5O8S (M), 665.35; found, 664.13 (M-H) 700.00 (M-H+Cl) for negative mode,
and 688.33 (M+Na) for positive mode.
Hydrolysis of the dimethyl acetal in 2.3 was accomplished by dissolving it (4 mg, 6
µmol) in 3:1 acetic acid/water (1.3 mL) and incubating overnight at room temperature in
the dark, followed by rotary evaporation of solvents. The residue was purified by flash
chromatography on a silica gel column (ethyl acetate: methanol=40:1, TLC: Rf = 0.6) and
placing under high vacuum through a dry ice-acetone cooled trap for 2 days to give
compound 2.4. (2.3 mg, 3.8 µmol, yellow solid). ESI-MS: m/z calcd for C30H45N5O7S
(M), 601.28; found, 600.13 (M-H), 635.87 (M-H+Cl) for negative mode, and 624.20
(M+Na) for positive mode.
61

2.4.3 In vitro modification of calpain-1 by iso[4]LGE2. Calpain-1 was modified with
10 equivalents of iso[4]LGE2. Calpain-1 (100 µg, 8.9 nmol) in 190 µl of a solution
containing 20 mM imidazole, 5 mM β–mercaptoethanol, 1 mM EDTA, 1 mM EGTA, 30 %
glycerol, pH 6.8 was mixed with 310 μl of sodium acetate solution (10 mM, pH 6.8).
First 11 µl of iso[4]LGE2 stock (1 µg/µl in methanol, 11 µg, 30 nmol) was added to
calpain-1 and the mixture reacted on a shaker (IKA MTS 2/4 digital microtiter shaker,
300 rpm) for 40 min. Then another 11 µl of iso[4]LGE2 stock (1 µg/µl, 11 µg, 30 nmol)
was added and reacted for 40 min, then the last 11 µl of iso[4]LGE2 stock (1 µg/µl, 11 µg,
30 nmol) was added and reacted for 40 min. The reaction mixture (total 533 µl) was
quenched by addition of glycine (2 µl, 100 mM, 178 nmol, 2 molar equivalent to
iso[4]LGE2) with brief mixing. The mixture was then flash-frozen in liquid nitrogen and
stored at -80 °C until analysis by MS.
Modified calpain-1 (20 µg) was subjected to 4-12% SDS-PAGE. The gel was stained
with Coomassie blue, and a region corresponding to proteins with molecular mass
between 80-90 and 30-40 kDa was excised respectively. Destained, reduced in 20 mM
DTT for 30 min at room temperature, and alkylated in 100 mM iodoacetamide in the dark
for 30 min at room temperature. For LC/MSMS analysis, the protein was first in gel
digested with chymotrypsin. Chymotrypsin stock solution was prepared by dissolving
chymotrypsin (25 µg) in NH4HCO3 (1000 µl 50 mM), to provide a final concentration of
25 µg/ml. Chymotrypsin solution (16 µl, 0.4 µg) was added to modified calpain (20 µg)
for 22 h at 37 °C using a protease to calpain-1 ratio of 1:50 (w/w) in 50 mM NH4HCO3.
Peptides were extracted with 50% acetonitrile and 5% formic acid, dried in a vacuum
concentrator, and stored at -20 °C. Peptide separations were performed on an Ultimate
62

3000 LC system with a C18 Acclaim PepMap 100 column (0.075 x 150 mm, Dionex,
Sunnyvale, CA). Peptides were eluted over a 50-min gradient from 0 to 80% acetonitrile
in water, containing 0.1% formic acid, at a flow rate of 300 nl/min. The column effluent
was continuously directed into the nanospray source of the mass spectrometer, a hybrid
Fourier transform ion cyclotron resonance (FTICR)/linear ion trap mass spectrometer
(LTQ FT Ultra, Thermo Scientific, West Palm Beach, FL). The following parameters
were used for all acquisition methods on the LTQ FT Ultra MS: an ion spray voltage of
2400 V and an interface capillary heating temperature of 200 °C. Full mass spectra were
acquired from the FTICR, and the tandem mass spectra (MS/MS) of the eight most
intense ions were recorded by the linear ion trap in data-dependent mode with normalized
collision energy of 35 eV, isolation width of 2.5 Da, and activation Q of 0.25.
2.4.4 In vitro modification of GAPDH by fluorescently labeled iso[4]LGE2. GAPDH
was modified with 10 equivalents of fluorescently labeled iso[4]LGE2. Fluorescently
labeled iso[4]LGE2 (168 µg, 280 nmol) (21 µl of 8 mg/ml in methanol) was added to into
GAPDH (28 nmol) (1 mg in 479 µl of 50 mM sodium phosphorus buffer, pH 7.0) and the
mixture was under air on an IKA MTS 2/4 digital microtiter shaker at 300 rpm for a total
8 h at 25 °C. Then the reaction mixture (total 500 µl) was quenched by addition of
glycine (6 µl, 100 mM, 560 nmol, 2 molar equivalent to fluorescently labeled iso[4]LGE2)
with brief mixing. The mixture was flash-frozen in liquid nitrogen and stored at -80 °C
for further analysis by MS and amino acid analysis.
For LC/MSMS analysis, in solution digestion was performed using trypsin. The
protein was reduced with 20 mM DTT for 30 min at room temperature, and alkylated in
63

200 mM iodoacetamide in the dark for 1 h at room temperature. The protein was then
digested by trypsin (20 µg) for 24 h at 37 °C with a protease to GAPDH ratio of 1:50
(w/w) in 50 mM of NEM·OAc (pH 8.6). Peptides were dried in a vacuum concentrator,
and the residue stored at -20 °C.
The tryptic peptides were first separated on a reverse phase column (2.1x100 mm i.d.
C18 1.8 µm column from Waters Acquity UPLC HSS). Chromatography was carried out
by elution with a linear gradient (elute A, 0.1% formic acid/H2O; elute B, 0.1% formic
acid/ACN). The sample was loaded onto the column with 98% A and 2% B at a flow rate
of 70 µl/min for 10 min. For the analysis, the gradient was as follows: 2-10% B over 5
min, to 70 % within 30 min at a flow rate of 70 µl/min, to 98 % within 0.1 min and hold
for 5 min at a flow rate of 140 µl/min.
The analyses of digested peptides were performed by Acquity® fluorescence (FLR)
detector and API-3000 triple quadrapole electrospray mass spectrometer (Applied
Biosystems Inc.) operated in the positive ion mode and a MS scan of m/z 220-2000.
Fluorescence channels of tryptophan peptides (three digested peptides containing
tryptophan) and iso[4]LGE2 tag were both monitored. The fluorescence of tryptophan
peptides (280 nm for excitation, 340 nm for emission) and mass spectrometry were
monitored to confirm the chromatographic separation. The fluorescence signal of
iso[4]LGE2 tag (450 nm for excitation, 560 nm for emission) was monitored to confirm
the elution time of fluorescently labeled iso[4]LGE2 modified peptides. Peptides with
iso[4]LGE2 tag fluorescence signals were collected as one minute fraction, quickly dried
using a high-speed vacuum evaporator and then stored at -20 °C until for further analysis.
64

The tryptic peptides with fluorescence signals were re-suspended in 0.1% formic
acid/2% acetonitrile and further analyzed by LC-MS/MS using a quadrupole-time of
flight (QTof2) mass spectrometer equipped with a CapLC system (Waters, Millford, MA),
ProteinLynx Global Server acquisition and processing software. Peptide digests were
trapped on a precolumn (0.3 x 1mm, 5µC18, LC Packing) with 0.1% formic acid in 2%
acetonitrile as loading solvent then eluted onto a capillary column (PicoFrit 0.050 x 75
mm, 15µ tip ID; New Objective Inc., Woburn, MA) over a 60-min gradient from 0 to 80%
acetonitrile in water, containing 0.1% formic acid, at a flow rate of 250 nl/min. The
column effluent was continuously directed into the nanospray source of the mass
spectrometer which was operated in standard MS/MS switching mode with the three most
intense ions in each survey scan subjected to MS/MS analysis. Protein identifications
from MS/MS data utilized ProteinLynxTM Global Server (Waters Corporation) and
Mascot (Matrix Science) search engines and the Swiss-Protein and NCBI protein
sequence databases.
2.4.5 Electrophoresis. According to a published protocol,58 modified proteins (~ 5-10 µg)
were suspended in Laemmli sample buffer,59 fractionated on a 4-12% SDS-PAGE, and
then staind with coomassie blue (Bio-Rad coomassie stain #161-0786) or silver stain (GE
Healthcare, PlusOne silver staining kit, protein, instructions 71-7177-00AL). The bands
were quantified using a GS-710 scanner and QuantityOne® software from BioradTM
(Hercules, CA).
65

2.4.6 Western analysis. According to published protocols,58, 60 modified proteins (~ 5-10
µg) were suspended Laemmli sample buffer,59 fractionated on a 4-12% SDS-PAGE, and
electroblotted onto a polynivyl difluoride (PVDF) membrane (Millipore, Bedford, MA).
The PVDF membranes were treated with Odyssey blocking buffer (Li-Cor Cat# 927-
40000) and probed using various antibodies: anti-calpain-1 small subunit (Fisher
Cat#01674303) mAb, anti-calpain-1 catalytic subunit (Fisher Cat#01674304) mAb, anti-
iso[4]LGE2 mAb (see Chapter 5). Goat anti-mouse IRDye® 680 (Odyssey Cat#926-
32220) was used as secondary antibody. The immunoreactive bands were detected and
quantified using a Li-Cor infrared imaging system.
2.4.7 Calpain-1 activity assay. Calpain-1 assay was performed on aliquots of modified
isoLG protein using a calpain activity assay kit (Biovision Inc., Mountainview, CA)
following recommended protocols. The fluorometric assay was based on detecting the
cleavage of a calpain substrate, Ac-LLY-AFC, that fluoresces at 505 nm, whereas the
cleaved substrate fluoresces at 400 nm. Thus, calpain-1 (26 μl of 2.286 μM, 6.6 μg,
0.0595 nmol) was added to iso[4]LGE2 (0.21 μg 0.595 nmol) in 74 μl of aqueous sodium
acetate (10 mM), mixed well and reacted under air on shaker (IKA MTS 2/4 digital
microtiter shaker, 300 rpm) at 25 °C. At each time point (3, 10, 30 and 90 min), one
aliquot (25 μl) was taken and quenched with 1.2 μl 1 mM glycine (1.2 nmol) before
activity assay. The aliquot (26.2 μl) was added extraction buffer (58.8 μl), 10x reaction
buffer (Ready-to-Use reagent from Biovision calpain activity assay kit) (10 μl) and
calpain substrate (5 μl). Then the mixture was incubated at 37 °C for 1 hour in the dark
and fluorescence was detected using Spectra Max Gemini spectrofluorometer (excitation
66

400 nm, emission 505 nm) (Molecular Devices, Sunnyvale, CA). Negative control was
performed the same method with calpain inhibitor Z-LLY-FMK from Biovision calpain
activity assay kit.
2.4.8 MS/MS identification of isoLG-modified sites in calpain-1 and GAPDH.
Peptides were identified from LTQ FT Ultra MS/MS and quadrupole-time of flight (Q-
Tof-2) experimental data by generating peak lists with Mascot Daemon and submitting
these to the Mascot search engine, version 2.3.0 (Matrix Science, Boston). S-
Carbamidomethylation of cysteine was set as a fixed modification, whereas oxidation of
methionine (methionine sulfoxide) was set as a variable modification. Formulas
corresponding to the various oxidation and dehydration states of iso[4]LGE2 adducts61
and fluorescently labeled iso[4]LGE2 adducts were manually entered into the Mascot
search parameters and selected as variable modifications for lysine (appendix Table 2.1S
and 2,4S, structures shown in Scheme 2.1S). Mass tolerances were ±15 ppm for precursor
ions and ± 0.8 Da for fragment ions. One missed cleavage site was allowed for trypsin
(cleaves at residues Lys and Arg but not at Pro) and three missed cleavage sites were
allowed for chymotrypsin (cleaves at Phe, Leu, Trp, and Tyr but not at Pro). Searches
were restricted to a sequence data base containing only human calpain-1 and rabbit
GAPDH because purified protein was used. Only peptides with a significane score
greater than 15 (p<0.05) according to Mascot’s scoring algorithm were considered.
2.4.9 Amino acid analysis. IsoLG modified GAPDH (400 µg) was subjected to 4-12%
SDS-PAGE. The gel was stained with Coomassie blue, and a region corresponding to
67

proteins with molecular mass between 70-80, 110-120 and 140-160 kDa were excised.
Each excised region was destained, reduced in 20 mM DTT for 30 min at room
temperature, and then alkylated in 200 mM iodoacetamide in the dark for 1 h at room
temperature. The protein was then digested in gel with trypsin (8 µg) for 24 h at 37 °C
with a protease to GAPDH ratio of 1:50 (w/w) in 50 mM of NEM·OAc (pH 8.6).
Peptides were extracted from the gel in 0.1%TFA/20%ACN, dried in a vacuum
concentrator, and stored at -20 °C for later analysis. AccQ·TagTM amino acid analysis
was used to quantify cross-linked GAPDH. The trypsin digested peptides were
hydrolyzed to amino acids with HCl, then with derivatized AccQ·Tag reagent and
analyzed chromatographically as described below.
2.4.9.1 Vapor phase HCl Hydrolysis: Digested dimer, trimer and tetramer peptides
were treated separately. Each sample was added 50 µl 0.1%TFA/20%ACN and then 150
µl 0.1%TFA. 20 µl aliquot was taken out from each tube and hydrolyzed to free amino
acids under HCl vapor. Peptide solutions (20 µl) were transferred to 6×50 mm hydrolysis
tubes and dried under vacuum. After drying, the glass tubes were placed into a screw-cap
vial containing 300 μl of 6M HCl and a few crystals of phenol (~1-2 mg). The vial was
evacuated briefly then flushed with argon and this alternating process was repeated three
times. The vial was finally sealed under vacuum and was heated at 150 °C in an oven for
1 h. After hydrolysis, sample tubes were dried using a high-speed vacuum evaporator and
stored at -20 °C until derivatization.
2.4.9.2 AccQ·Tag Derivatization and chromatography: AccQ·TagTM ultra
derivatization kit (Waters) was used. AccQ·TagTM ultra reagent reacts rapidly with
primary amino groups to yield highly stable urea. The structure of the derivatizing group
68

was the same for all amino acids.62-63 Peptide hydrolysates were transferred to reaction
vials and AccQ·TagTM borate buffer (30 µl) was added to adjust the pH to above 9.5.
After borate buffer was mixed well with the peptide hydrolysates by vortexing. To a 24
µl aliquot was added AccQ·TagTM reagent (6 µl, Ready-to-Use reagent from Waters
AccQ·TagTM ultra derivatization kit). The contents of the vial were mixed by vortexing
and then heated for 10 min at 55 °C. Then deionized water (30 µl) was added and the
mixture was vortexed. All 60 µl of the sample solution was transferred to a UPLC sample
vial for testing. The amino acid standard was prepared in the same manner.
The derivatized samples and H-standard were analyzed with a Waters Acquity ultra
performance LC system using 2 µl injections for optimum resolution and sensitivity (100
x 2.1 mm column). For the separation, sodium acetate (140 mM) containing 17 mM
triethylamine (pH 5) was used as buffer A and 60 % acetonitrile/H2O as buffer B. The
gradient was as follows with a flow rate of 700 μl/min: time 0-0.54 min, 0.1% B; time
5.74 min, 9.1% B; time 7.74 min, 21.2 % B; time 8.04 min, 59.6% B and time 8.05 min,
90% B. AccQ·Tag amino acids were detected with the Acquity UPLC tunable UV
detector at 260 nm.
2.4.9.3 Amino acid analysis and data interpretation: Amino acid analysis was
performed to quantify the concentrations of digested dimer, trimer and tetramer peptides. The
amount of dimer, trimer and tetramer were determined by (i) dividing the amount of each
individual residue by the known residue value, (ii) calculating a mean amount analyzed,
(iii) discarding all individual values that deviated more than 15% from the mean, and (iv)
recalculating the mean amount analyzed from the remaining relevant residues.64 The
identity and quantity of each amino acid was established by comparing retention time and
69

chromatographic peak areas with the peak area of a quantitative standard mixture of amino
acids (Standard H, 50 pmol) that was derivatized and analyzed the same way as the
digested peptides with the same amount of reagents and instruments.
Standard response (standard H) per pmol for each amino acid is equal to standard
signal (HPLC peak area) divided by pmol amino acid for analysis. Sample signal (peak
area of GAPDH derivatized dimer sample) divided by the standard response will get
analyzed pmol of each amino acid. Then analyzed sample pmol of each amino acid is
divided by the known number of amino acid per molecule protein (GAPDH) to obtain
pmol protein. Then the mean pmol protein (within 15% deviation) is calculated and
divided by sample dilution ratio to get pmol of the derivatized protein (GAPDH dimer).
Speed vac the extraction buffer, each sample was dissolved in 50 µl 0.1% TFA, 20% ACN, then added 150 µl 0.1% TFA. Total volume of each sample was 200 µl
Running gel with 400 µg (11200 pmol) of modified GAPDH (24 h)
Cut and accumulate all the dimer, trimer, tetramer bands for digestion
From each 200 µl, take out 20 µl for amino acid analysis
Dimer1.34 pmol/µl (0.05 µg/µl)
total: 0.05 µg/µl× 200 µl=10 µg 1.34 pmol/µl× 200 µl=268 pmol
modified percentage: 10µg/400µg=2.5%
Trimer1.5 pmol/ul (0.05 µg/µll)
total: 0.05 µg/µl× 200 µl=10 µg 1.5 pmol/µl× 200 µl=300 pmol
percentage: 10µg/400µg=2.5%
Tetramer0.93 pmol/ul (0.03 µg/µl)
total: 0.03 µg/µl× 200 µl=6 µg 0.93 pmol/µl× 200 µl=186 pmol
percentage: 6µg/400µg=1.5%
70

2.5 References
1. Gulcan, H. G.; Alvarez, R. A.; Maude, M. B.; Anderson, R. E. (1993) Lipids of
human retina, retinal-pigment epithelium, and bruch membrane choroid - comparison of
macular and peripheral regions. Invest Ophthalmol Vis Sci 34 (11), 3187-3193.
2. Poliakov, E.; Meer, S. G.; Roy, S. C.; Mesaros, C.; Salomon, R. G. (2004) Iso 7
LGD(2) - Protein adducts are abundant in vivo and free radical-induced oxidation of an
arachidonyl phospholipid generates this D series isolevuglandin in vitro. Chem Res
Toxicol 17 (5), 613-622.
3. Subbanagounder, G.; Salomon, R. G.; Murthi, K. K.; Brame, C.; Roberts, L. J.
(1997) Total synthesis of Iso 4 -levuglandin E-2. J Org Chem 62 (22), 7658-7666.
4. Salomon, R. G.; Miller, D. B. (1985) Levuglandins: isolation, characterization,
and total synthesis of new secoprostanoid products from prostaglandin endoperoxides.
Adv Prostaglandin Thromboxane Leukot Res 15, 323-326.
5. Salomon, R. G. (2005) Levuglandins and isolevuglandins: Stealthy toxins of
oxidative injury. Antioxid Redox Signal 7 (1-2), 185-201.
6. Kobierski, M. E.; Kim, S. C.; Murthi, K. K.; Iyer, R. S.; Salomon, R. G. (1994)
Synthesis of a pyrazole isostere of pyrroles formed by the reaction of the epsilon-amino
groups of protein lysyl residues with levuglandin E(2). J Org Chem 59 (20), 6044-6050.
7. Difranco, E.; Subbanagounder, G.; Kim, S.; Murthi, K.; Taneda, S.; Monnier, V.
M.; Salomon, R. G. (1995) Formation and stability of pyrrole adducts in the reaction of
levuglandin E(2) with proteins. Chem Res Toxicol 8 (1), 61-67.
71

8. Salomon, R. G.; Sha, W.; Brame, C.; Kaur, K.; Subbanagounder, G.; O'Neil, J.;
Hoff, H. F.; Roberts, L. J. (1999) Protein adducts of iso 4 levuglandin E-2, a product of
the isoprostane pathway, in oxidized low density lipoprotein. J Biol Chem 274 (29),
20271-20280.
9. Govindarajan, B.; Laird, J.; Salornon, R. G.; Bhattacharya, S. K. (2008)
Isolevuglaridin-modified proteins, including elevated levels of inactive calpain-1,
accumulate in glaucomatous trabecular meshwork. Biochemistry 47 (2), 817-825.
10. Charvet, C. D.; Laird, J.; Xu, Y.; Salomon, R. G.; Pikuleva, I. A. (2013)
Posttranslational modification by an isolevuglandin diminishes activity of the
mitochondrial cytochrome P450 27A1. J Lipid Res 54 (5), 1421-1429.
11. Charvet, C. D.; Liao, W. L.; Heo, G. Y.; Laird, J.; Salomon, R. G.; Turko, I. V.;
Pikuleva, I. A. (2011) Isolevuglandins and Mitochondrial Enzymes in the Retina mass
spectrometry detection of post-translational modification of sterol-metabolizing
CYP27A1. J Biol Chem 286 (23), 20413-20422.
12. Boutaud, O.; Brame, C. J.; Chaurand, P.; Li, J. Y.; Rowlinson, S. W.; Crews, B.
C.; Ji, C.; Marnett, L. J.; Caprioli, R. M.; Roberts, L. J.; Oates, J. A. (2001)
Characterization of the lysyl adducts of prostaglandin H-synthases that are derived from
oxygenation of arachidonic acid. Biochemistry 40 (23), 6948-6955.
13. Sazontova, T. G.; Arkhipenko, I. V. (2005) The role of free radical processes and
redox-signalization in adaptation of the organism to changes in oxygen level. Ross Fiziol
Zh Im I M Sechenova 91 (6), 636-655.
72

14. Ray, S. K.; Shields, D. C.; Saido, T. C.; Matzelle, D. C.; Wilford, G. G.; Hogan, E.
L.; Banik, N. L. (1999) Calpain activity and translational expression increased in spinal
cord injury. Brain Res 816 (2), 375-380.
15. Govindarajan, B.; Laird, J. M.; Sherman, R.; Salomon, R. G.; Bhattacharya, S. K.
(2008) Neuroprotection in Glaucoma Using Calpain-1 Inhibitors: Regional Differences in
Calpain-1 Activity in the Trabecular Meshwork, Optic Nerve and Implications for
Therapeutics. CNS Neurol Disord Drug Targets 7 (3), 295-304.
16. Davies, S. S.; Amarnath, V.; Montine, K. S.; Bernoud-Hubac, N.; Boutaud, O.;
Montine, T. J.; Roberts, L. J. (2002) Effects of reactive gamma-ketoaldehydes formed by
the isoprostane pathway (isoketals) and cyclooxygenase pathway (levuglandins) on
proteasome function. FASEB J 16 (3), 715-717.
17. Harris, J. I.; Waters, M., The enzymes. 3rd edn (Boyer, P. D., ed.) ed.; Academic
Press: New York, 1976; Vol. 13, 1-49.
18. Fothergill-Gilmore, L. A.; Michels, P. A. (1993) Evolution of glycolysis. Prog
Biophys Mol Biol 59 (2), 105-235.
19. Mountassif, D.; Baibai, T.; Fourrat, L.; Moutaouakkil, A.; Iddar, A.; El Kebbaj, M.
S.; Soukri, A. (2009) Immunoaffinity purification and characterization of glyceraldehyde-
3-phosphate dehydrogenase from human erythrocytes. Acta Biochim Biophys Sin
(Shanghai) 41 (5), 399-406.
20. Huitorel, P.; Pantaloni, D. (1985) Bundling of microtubules by glyceraldehyde-3-
phosphate dehydrogenase and its modulation by ATP. Eur J Biochem 150 (2), 265-269.
21. Tisdale, E. J. (2001) Glyceraldehyde-3-phosphate dehydrogenase is required for
vesicular transport in the early secretory pathway. J Biol Chem 276 (4), 2480-2486.
73

22. Ikemoto, A.; Bole, D. G.; Ueda, T. (2003) Glycolysis and glutamate accumulation
into synaptic vesicles. Role of glyceraldehyde phosphate dehydrogenase and 3-
phosphoglycerate kinase. J Biol Chem 278 (8), 5929-5940.
23. Doucet, J. P.; Tuana, B. S. (1991) Identification of low molecular weight GTP-
binding proteins and their sites of interaction in subcellular fractions from skeletal muscle.
J Biol Chem 266 (26), 17613-17620.
24. Berry, M. D. (2004) Glyceraldehyde-3-phosphate dehydrogenase as a target for
small-molecule disease-modifying therapies in human neuro degenerative disorders. J
Psychiatry Neurosci 29 (5), 337-345.
25. Uchida, K.; Stadtman, E. R. (1993) Covalent attachment of 4-hydroxynonenal to
glyceraldehyde-3-phosphate dehydrogenase. A possible involvement of intra- and
intermolecular cross-linking reaction. J Biol Chem 268 (9), 6388-6393.
26. Govindarajan, B.; Junk, A.; Algeciras, M.; Salomon, R. G.; Bhattacharya, S. K.
(2009) Increased isolevuglandin-modified proteins in glaucomatous astrocytes. Mol Vis
15 (114-15), 1079-1091.
27. Pappin, D. J. C.; Hojrup, P.; Bleasby, A. J. (1993) Rapid identification of proteins
by peptide-mass fingerprinting. Curr Biol 3 (6), 327-332.
28. Greenberg, M. E.; Li, X. M.; Gugiu, B. G.; Gu, X.; Qin, J.; Salomon, R. G.;
Hazen, S. L. (2008) The lipid whisker model of the structure of oxidized cell membranes.
J Biol Chem 283 (4), 2385-2396.
29. Huang, Y.; Wang, K. K. (2001) The calpain family and human disease. Trends
Mol Med 7 (8), 355-362.
74

30. Davis, T. L.; Walker, J. R.; Finerty, P. J., Jr.; Mackenzie, F.; Newman, E. M.;
Dhe-Paganon, S. (2007) The crystal structures of human calpains 1 and 9 imply diverse
mechanisms of action and auto-inhibition. J Mol Biol 366 (1), 216-229.
31. Sorimachi, H.; Ishiura, S.; Suzuki, K. (1997) Structure and physiological function
of calpains. Biochem J 328, 721-732.
32. Shao, X. G.; Fernandez, I.; Sudhof, T. C.; Rizo, J. (1998) Solution structures of
the Ca2+-free and Ca2+-bound C(2)A domain of synaptotagmin I: Does Ca2+ induce a
conformational change? Biochemistry 37 (46), 16106-16115.
33. Sorimachi, H.; Suzuki, K. (2001) The structure of calpain. J Biochem 129 (5),
653-664.
34. Croall, D. E.; Ersfeld, K. (2007) The calpains: modular designs and functional
diversity. Genome Biol 8 (6), 218.
35. Azuma, M.; Shearer, T. R. (2008) The role of calcium-activated protease calpain
in experimental retinal pathology. Surv Ophthalmol 53 (2), 150-163.
36. Lametsch, R.; Lonergan, S.; Huff-Lonergan, E. (2008) Disulfide bond within mu-
calpain active site inhibits activity and autolysis. Biochim Biophys Acta 1784 (9), 1215-
1221.
37. Reverter, D.; Strobl, S.; Fernandez-Catalan, C.; Sorimachi, H.; Suzuki, K.; Bode,
W. (2001) Structural basis for possible calcium-induced activation mechanisms of
calpains. Biol Chem 382 (5), 753-766.
38. Moldoveanu, T.; Hosfield, C. M.; Lim, D.; Elce, J. S.; Jia, Z. C.; Davies, P. L.
(2002) A Ca2+ switch aligns the active site of calpain. Cell 108 (5), 649-660.
75

39. Salomon, R. G.; Jirousek, M. R.; Ghosh, S.; Sharma, R. B. (1987) Prostaglandin
endoperoxides .21. covalent binding of levuglandin-E2 with proteins. Prostaglandins 34
(5), 643-656.
40. Iyer, R. S.; Ghosh, S.; Salomon, R. G. (1989) Levuglandin-E2 cross-links proteins.
Prostaglandins 37 (4), 471-480.
41. Iyer, R. S.; Kobierski, M. E.; Salomon, R. G. (1994) Generation of pyrroles in the
reaction of levuglandin E(2) with proteins. J Org Chem 59 (20), 6038-6043.
42. Prados, P.; Fukushima, T.; Santa, T.; Homma, H.; Tsunoda, M.; AlKindy, S.;
Mori, S.; Yokosu, H.; Imai, K. (1997) 4-N,N-Dimethylaminosulfonyl-7-N-(2-
aminoethyl)amino-benzofurazan as a new precolumn fluorescence derivatization reagent
for carboxylic acids (fatty acids and drugs containing a carboxyl moiety) in liquid
chromatography. Anal Chim Acta 344 (3), 227-232.
43. Fukushima, T.; Usui, N.; Santa, T.; Imai, K. (2003) Recent progress in
derivatization methods for LC and CE analysis. J Pharm Biomed Anal 30 (6), 1655-1687.
44. Nakanishi, S.; Tanaka, H.; Hioki, K.; Yamada, K.; Kunishima, M. (2010)
Labeling study of avidin by modular method for affinity labeling (MoAL). Bioorg Med
Chem Lett 20 (23), 7050-7053.
45. Fukushima, T.; Arai, K.; Tomiya, M.; Mitsuhashi, S.; Sasaki, T.; Santa, T.; Imai,
K.; Toyo'oka, T. (2008) Fluorescence determination of N-acetylaspartic acid in the rat
cerebrum homogenate using high-performance liquid chromatography with pre-column
fluorescence derivatization. Biomed Chromatogr 22 (1), 100-105.
76

46. Collier, H. O. J.; Roy, A. C. (1974) Hypothesis - inhibition of prostaglandin-e-
sensitive adenyl-cyclase as mechanism of morphine analgesia. Prostaglandins 7 (5), 361-
376.
47. Chan, L. C.; Cox, B. G. (2007) Kinetics of amide formation through
Carbodiimide/N-Hydroxybenzotriazole (HOBt) couplings. J Org Chem 72 (23), 8863-
8869.
48. Sambarkar, P. B.; Patil, A. C. (2012) Synthesis of amides from acid and amine
using coupling reagents. J Curr Pharm Res 10 (1), 22-24.
49. Bailen, M. A.; Chinchilla, R.; Dodsworth, D. J.; Najera, C. (1999) 2-
mercaptopyridone 1-oxide-based uronium salts: New peptide coupling reagents. J Org
Chem 64 (24), 8936-8939.
50. Kitson, T. M. (1979) 2,2'-Dithiodipyridine activates aldehyde dehydrogenase and
protects the enzyme against inactivation by disulfiram. Biochem J 183 (3), 751-753.
51. Loader, C.; Anderson, H. J. (1971) Pyrrole Chemistry. Part XI. Some Reactions
of the Pyrrole Grignard Reagent with Alkyl Carbonates and Alkyl Thiocarbonates. Can J
Chem 49 (1), 45-48.
52. Nicolaou, K. C.; Claremon, D. A.; Papahatjis, D. P. (1981) A mild method for the
synthesis of 2-ketopyrroles from carboxylic-acids. Tetrahedron Lett 22 (46), 4647-4650.
53. Nakahara, Y.; Fujita, A.; Ogawa, T. (1985) An efficient synthesis of 2-
acylpyrroles. Agric Biol Chem 49 (5), 1491-1495.
54. Min, J. Z.; Toyo'oka, T.; Kawanishi, H.; Fukushima, T.; Kato, M. (2005) Novel
fluorescent asparaginyl-N-acetyl-D-glucosamines (Asn-GlcNAc) for the resolution of
77

oligosaccharides in glycopeptides, based on enzyme transglycosylation reaction. Anal
Chim Acta 550 (1-2), 173-181.
55. Cowan-Jacob, S. W.; Kaufmann, M.; Anselmo, A. N.; Stark, W.; Grutter, M. G.
(2003) Structure of rabbit-muscle glyceraldehyde-3-phosphate dehydrogenase. Acta
Crystallogr Sect D-Biol Crystallogr 59, 2218-2227.
56. Nakajima, H.; Amano, W.; Fujita, A.; Fukuhara, A.; Azuma, Y. T.; Hata, F.; Inui,
T.; Takeuchi, T. (2007) The active site cysteine of the proapoptotic protein
glyceraldehyde-3-phosphate dehydrogenase is essential in oxidative stress-induced
aggregation and cell death. J Biol Chem 282 (36), 26562-26574.
57. Still, W. C.; Kahn, M.; Mitra, A. (1978) Rapid chromatographic technique for
preparative separations with moderate resolution. J Org Chem 43 (14), 2923-2925.
58. Crabb, J. W.; Miyagi, M.; Gu, X. R.; Shadrach, K.; West, K. A.; Sakaguchi, H.;
Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G. (2002)
Drusen proteome analysis: An approach to the etiology of age-related macular
degeneration. Proc Natl Acad Sci U S A 99 (23), 14682-14687.
59. Laemmli, U. K. (1970) Cleavage of structural proteins during assembly of head of
bacteriophage-T4. Nature 227 (5259), 680-685.
60. Bhattacharya, S. K.; Rockwood, E. J.; Smith, S. D.; Bonilha, V. L.; Crabb, J. S.;
Kuchtey, R. W.; Robertson, N. G.; Peachey, N. S.; Morton, C. C.; Crabb, J. W. (2005)
Proteomics reveal cochlin deposits associated with glaucomatous trabecular meshwork. J
Biol Chem 280 (7), 6080-6084.
61. Brame, C. J.; Salomon, R. G.; Morrow, J. D.; Roberts, L. J. (1999) Identification
of extremely reactive gamma-ketoaldehydes (isolevuglandins) as products of the
78

isoprostane pathway and characterization of their lysyl protein adducts. J Biol Chem 274
(19), 13139-13146.
62. Cohen, S. A.; Michaud, D. P. (1993) Synthesis of a fluorescent derivatizing
reagent, 6-aminoquinolyl-n-hydroxysuccinimidyl carbamate, and its application for the
analysis of hydrolysate amino-acids via high-performance liquid-chromatography. Anal
Biochem 211 (2), 279-287.
63. Ward, M. (1996) Amino acid analysis using precolumn derivatization with 6-
aminoquinolyl-n-hydroxysuccinimidyl carbamate. The protein protocols handbook, 461-
465.
64. Crabb, J. W.; Ericsson, L.; Atherton, D.; Smith, A. J.; Kutny, R., A collaborative
amino acid analysis study from the association of biomolecular resource facilities. In
Current research in protein chemistry, Villafranca, J. J., Ed. Elsevier Science: Academic
Press, Inc, 1990; pp 49-61.
79

CHAPTER 3
DETECTION AND CHARACTERIZATION OF THE PRODUCTS
FROM THE REACTION OF ISO[4]LGE2 WITH PEPTIDES: N-
ACETYL-GLY-LYS-OMe AND AMYLOID “EVHHQKL”

3.1 Background
3.1.1 Purpose
Levuglandins (LGs) and isolevuglandins (isoLGs) react with protein lysyl residues to
form covalent adducts that include pyrroles and their more oxidized derivatives, lactams
and hydroxylactams, as well as protein-protein cross-links. Two alternative structures
have been proposed for the protein-protein cross-links, a pyrrole-pyrrole dimer derived
from two molecules of protein and two molecules of LG/isoLG or an aminal derived
from two molecules of protein and one molecule of LG/isoLG. In the study, to obtain
direct evidence for the molecular structure of lysyl-LG/isoLG cross-links, two simple
model systems were investigated, the reaction iso[4]LGE2 with N-acetylglycyllysine
methyl ester and with the β-amyloid (11-17) peptide EVHHQKL.
3.1.2 Isolevuglandin modification of proteins
A variety of products are formed through oxidation of fatty acids by enzymatic or free
radical induced oxidation pathways. The enzymatic oxidation of arachidonic acid (AA)
by cyclooxygenase (COX) leads to the formation of levuglandins, whereas oxidation of
phosphoesters of arachidonic acid leads to the production of isolevuglandins (isoLGs).1-3
Iso[4]LGE2, can only be generated through free radical oxidation of arachidonic acid
phospholipids. Iso[4]LGE2 is a highly reactive γ-ketoaldehyde which modifies the ε-
amino groups of lysine residues forming protein-bound pyrroles lactams and
hydroxylactams,4 as well as protein-protein cross-links for which aminal5 and pyrrole-
pyrrole6-7 structures have been postulated, as well as DNA-protein cross-links.8
Incubation of salmon sperm DNA with LGE2 causes DNA-protein cross-linking in
cultured cells (10 mM MOPS buffer), suggesting that levuglandins can directly react with
81

DNA. Cyclopentenone PGs contain α,β-unsaturated carbonyl groups that readily adduct
various biomolecules such as glutathione (GSH) in vitro (Scheme 3.1).9
O OH
COOH
PGJ2
+
NHCH2OCH2CH2CHCOOHNH2
CHCONHCH2COOH
CH2SH
GSH transferase
NHCH2OCH2CH2CHCOOHNH2
CHCONHCH2COOH
CH2S
O OH
COOH
+ 2H ·
NHCH2OCH2CH2CHCOOHNH2
CHCONHCH2COOH
CH2S
HO OH
COOH
GSH
Scheme 3.1 Schematic representation of pyrrole cross-linking structure in previous study:
adducts of PGJ2 reaction with GSH in HCA-7 cells.9
LGE2 also reacts with nucleotides in vitro to form covalent adducts with a
dihydroxypyrrolidine structure (Scheme 3.2).10
82

O O
OH
HOO
N
N
O
NH2
dR
N
N
O
NH
dR
OHR'R
O
HemiaminalH2O
ProteinsDNA
N
N
O
N
dR
R'R
O
N
N
O
N
dR
OH
R'R
HO
2 H2O
N
N
O
N
dR
R'R
Pyrrole
+ O2
ProteinsN
N
O
N
R'R
lysine
O
Lactam
Dihydroxypyroolidine Schiff base
Scheme 3.2 Schematic representation of pyrrole cross-linking structure in previous study:
adducts of nucleosides and DNA formed by reaction with levuglandin.10
PGH2 reacts with arginine to yield covalent adducts (Scheme 3.3).11 These structures
would be appropriate candidates for isoLG/LG-protein cross-link.
83

O OH
COOH
LGE2
H
OH2N CHCCH2
OHO
CH2CH2NHCNH2
NH
Arginine
+
H2N CHCCH2
OHO
CH2CH2N
CN N LGE2LGE2
hydrolysis
COOH
N
HOOC
N
O
O
O
H2N CHCCH2
OHO
CH2CH2NH2
Ornithine
+
Scheme 3.3 Schematic representation of pyrrole cross-linking structure in previous study:
formation of bis(levuglandinyl) urea.11
IsoLGs initially form reversible Schiff base adducts12-13 with proteins which can
dehydrate to pyrroles. These adducts can be further oxidized to form stable lactams and
hydroxylactams. A possible cross-linking mechanism identifies the LG-protein Schiff
base adduct as the reactive electrophile that binds with a primary amino group in a
second molecule of protein resulting in aminal formation.7 IsoLG/LG cross-link forms
also include pyrrole further reaction with nucleophiles like unoxidized pyrroles (Scheme
3.4)4 or thiols (Scheme 3.5).14-15
N
R R
N
R R
N
R R
O2 NH
O2NH
NH
Scheme 3.4 Schematic representation of pyrrole cross-linking structure in previous study:
nucleophilic substitution of alkylpyrroles in the presence of oxygen.4
84

O
O
NH
O HN
OOR
O
H
NH2
NH
O HN
OOR1
O
H
N
N
NH
OR2
O
OHN
O
H
+
R= H, CH3
R1 = R2 = HR1 = H, R2 = CH3
(A)
O
O
HN
O
COOH
H CH2SH
HN
O
COOH
H CH2
NH2
HN
O
COOH
S
N
CH2HNH
COOH
O
(B)
Scheme 3.5 Schematic representation of pyrrole cross-linking structures in previous
studies. (A) Pyrrole autoxidation products in 2,5-hexanedione-treated dipeptide.14 (B)
Formation of thiol-pyrrole conjugates.15
The different ways in which isoLGs have been shown or postulated to modify
proteins is depicted in Scheme 3.6. IsoLGs modify proteins in a matter of seconds and
cross-link them within minutes, which is much faster than the rates at which HNE or
MDA form cross-links.6 Modification of the proteasome and other intracellular proteins
targeted by isoLGs can lead to impaired degradation.16-18 For example, isoLG-adducted
ovalbumin and Aβ1-40 aggregates significantly inhibit the proteasome activity of
neuroglial cultured cells and induce cell death, suggesting that isoLG/LGs may have
85

relevance to the pathogenesis of neurotoxicity19 and neurodegenerative diseases.17 Also
these adducts have been shown to hinder the activity of the 20S proteasome, a component
of cytosolic protein, degradation machinery in a dose dependent manner.17 Accumulation
of iso[4]LGE2-modified proteins has been observed in many diseases such as
artherosclerosis20 and rheumatoid arthiritis.21
Scheme 3.6 Schematic representation of formation of iso[4]levuglandin protein adducts
and cross-links.
OO
CH3
Protein
ON
CH3
Schiff base
Protein
HO-PC
N
H3C
Pyrrole
Bispyrrole Crosslink
N
H3C OH
NH
Protein-protein Aminal Crosslink
Protein
Protein
Protein
O2 ROO
N
H3C
Protein
OH
O
Hydroxylactam+
N
H3C
Protein
O
Lactam
CC5H11
OH
OO-PC
CO
O-PCC5H11
OH
CC5H11
OOH
OH
CC5H11
OOH
OH
CC5H11
OH
CC5H11
OH
OOH
OOH
N
N
COOH C5H11
OH
C5H11COOH
HOProtein
Protein
Lys NH2
Protein Lys NH2
C5H11
AA-PC
(CH2)3COO
Iso[4]LGE2
-H2O
O2
O2
H2O
OPO(CH2)2NMe3
RCOO
O
O
86

3.1.3 β-Amyloid and isolevuglandins
Alzheimer’s disease (AD) brain is under intense oxidative stress, manifested by
increased protein oxidation, lipid peroxidation, free radical formation, DNA/RNA
oxidation, nitrotyrosine levels, and advanced glycation end products.22-23 Cerebral
accumulation of soluble oligomers of (Aβ) 1-42 that are cytotoxic to neurons is central to
the pathogenesis of Alzheimer’s disease (AD).24-28 Addition of Aβ (1-42) to neurons
leads to increased protein carbonyls and decreased cell survival compared to controls.29-34
Protein oxidation is increased in AD brain in regions rich in Aβ (1-42). In parallel,
evidence has indicated long-term use of inhibitors of the cyclooxygenases is associated
with a decrease in the severity of Alzheimer’s disease. Meantime PGH2 accelerates the
formation of oligomers of Aβ1-42 in conjunction with lipid modification of the protein. It
provides a molecular basis for the hypothetical mechanism that links cyclooxygenase
activity to the formation of oligomers of Aβ.16 Aggregation of amyloid β (Aβ) 1-42
peptide forming neurotoxic oligomers may be a consequence of the LG-protein-protein
cross-linking that was observed upon treatment of Aβ 1-42 with LGE2.35
87

3.2 Results and discussion
3.2.1 Reactions of model peptides with iso[4]LGE2.
Iso[4]LGE2 or iso[4]LGE2-FluoTag was incubated with N-acetylglycyllysine methyl
ester or β-amyloid (11-17) peptide “EVHHQKL” individually in NEM·OAc (pH 8.6) at
25°C for several days. Reactions were also conducted in both acidic (50 mM phosphate
buffer, pH 5.8) and neutral (50 mM phosphate buffer, pH 7.0) solutions. The results were
similar as in basic buffer. The reaction product profile exhibited no relationship with
buffer pH (vide infra). The resulting mixtures were dried on a speed vacuum, and re-
suspended in 10% acetonitrile/water with a trace of formic acid (0.1%) and analyzed by
mass spectrometry.
3.2.2 Mass spectrometric analysis of the iso[4]LGE2-peptide adducts.
N-Acetyl-glycyl-lysine methyl ester (N-acetyl-Gly-Lys-OMe) and β-amyloid(11-17)
peptide “EVHHQKL” were used as models for characterizing the modification of protein
lysyl residues resulting from adduction of iso[4]LGE2. A preliminary analysis of the
reaction product mixtures using MALDI-TOF mass spectrometry revealed the presence
of numerous products of peptide modification and indicated that both β-amyloid(11-17)
and N-acetylglycyllysine methyl ester showed similar products of adduction. Further
analysis was performed using triple quadrupole by ESI-MS and Q-TOF by ESI-MS/MS
to characterize the products.
(i) Iso[4]LGE2 modification of N-acetyl-gly-lys methyl ester.
MALDI-TOF MS analysis of the reaction product mixture from iso[4]LGE2 with a
large excess of N-acetylglycyllysine methyl ester showed major ions at m/z 576, 592,
88

608, 853, and 1149 corresponding to protonated pyrrole, lactam, hydroxylactam, aminal
cross-link and bispyrrole in addition to ions resulting from the loss of one or two
molecules of water from those adducts. Figure 3.1 shows a representative spectrum
revealing the presence of for various adducts in the reaction product mixture (see
appendix Table 3.1S for the molecular weights of various iso[4]LGE2-(N-acetyl-gly-lys-
OMe) adducts).
The reaction products for various reactant molar ratios were recorded (see appendix
Figure 3.1S (1) for N-acetyl-glycyl-lysine methyl ester: iso[4]LGE2 = 2000:1, (2) for
1000:1 and (3) for 200:1 in 1, 4, 7, 15, 21 and 50 day reactions). The initial concentration
of N-acetyl-glycyl-lysine methyl ester was constant while various ratios of iso[4]LGE2
were added. At various incubation times, identical aliquots of reaction mixture were
purified by Ziptip and then analyzed.
One-to-one adducts, i.e., pyrrole, lactam and hydroxylactam and their dehydration
products were generated very quickly (within one day reaction) in the reaction of
iso[4]LGE2 with N-acetyl-glycyl-lysine methyl ester (see appendix Figures 3.1S 1A).
Cross-link adducts included aminals, bispyrroles, trispyrroles and their dehydrated
forms. With higher iso[4]LGE2 ratio and longer incubation times, a higher percentage
(comparison of cross-link peak intensity with one-to-one adducts) of bispyrrole adducts
was generated. Specifically, bispyrrole peak intensity was higher when the molar ratio of
N-acetyl-glycyl-lysine methyl ester to iso[4]LGE2 was 200:1 than 2000:1 (compare
appendix Figures 3.1S 3A-F). This suggests that the dipeptide interferes with pyrrole-
pyrrole cross-link formation. This interpretation is also consistent with the results of later
experiments (see Chapter 4, section 4.2.4) testing the effect of dipeptide on the oxygen-
89

promoted dimerization and adduct formation reactions of pure pyrrole. Bispyrrole peak
intensity was also higher when N-acetyl-glycyl-lysine methyl ester reaction time with
iso[4]LGE2 was 15 days versus 1 day.
Derivatives of the polypyrroles, i.e., bispyrrole and trispyrrole showed a sequence of
ions corresponding to further addition of oxygen atoms (+16). Each polypyrrole parent
ion is accompanied by a series of ions corresponding to the addition of one or more atoms
of oxygen to the polypyrrole. The details are discussed below.
Figure 3.1 MALDI-TOF MS analysis of the reaction mixture from N-acetylglycyllysine
methyl ester and iso[4]LGE2 (molar ratio = 200:1) (50 mM NEM∙OAc aqueous solution
with 4% methanol, v/v, pH 8.6) at 25 °C for 4 days.
90

The product mixture from reaction of iso[4]LGE2 with N-acetyl-gly-lys-OMe was
separated by reverse phase HPLC using an acetonitrile/water gradient and monitored by
LC-ESI extracted ion channels (Figure 3.2). The major products corresponding to lactam
(trace 3.2C), aminal cross-link (trace 3.2B) and bispyrrole (trace 3.2D) eluted in order of
decreasing polarity at about 12, 30, and 36 minutes.
A
TIC
BispyrroleD
B
C
AminalCrosslink
Lactam-H2O
m/z 854
m/z 1150
m/z 574
6.8e8
1.3e6
5.8e6
3.7e6
Figure 3.2: LC-ESI for the product mixture from the reaction of N-acetylglycyllysine
methyl ester with iso[4]LGE2 (molar ratio = 200:1 at 25 °C for 4 days) with selected ion
recording for aminal cross-link, lactam-H2O and bispyrrole. A) TIC, B) aminal cross-link
m/z 854.1 (+ H+), C) lactam-H2O m/z 574.7 (+ H+), D) bispyrrole m/z 1150.5 (+ H+).
MALDI-TOF MS analysis of the fractions eluting at about 12, 30 and 36 minutes
confirmed the presence of the lactam, aminal and bispyrrole cross-links. Thus, MALDI-
TOF MS analysis of the 30 minute elution showed major ions at m/z 853.5, 835.5 and
817.5 corresponding to protonated aminal cross-link in addition to ions resulting from the
loss of one and two molecules of water. MALDI-TOF MS analysis of the 36 minute
91

elution showed major ions at m/z 1149.7, 1131.7 and 1113.7 corresponding to protonated
bispyrrole in addition to ions resulting from the loss of one and two molecules of water.
The MALDI-TOF mass spectra of these principle products are shown in Figure 3.3. A
series of ions are present in the mass spectrum of the isoLG-N-acetylglylys-OMe
bispyrrole cross-link chromatographic fraction corresponding to the addition of one or
more atoms of oxygen to the bispyrrole, and the relative amount of these products of
further oxidation is greater after incubation for 15 days compared to 4 days (Figure 3.3
panels A and B, respectively). The peak intensity of further oxidized bispyrrole (products
with addition of oxygen atoms) was higher compared with bispyrrole and its dehydrated
forms after 15 days modification than after 4 days. The pattern of further addition of
oxygen atoms (+16) was also observed for the trispyrrole products. Thus, pyrrole
oligomerizes to form polypyrroles, and all of these products undergo further oxidation.
Possible structures for products of oxidized bispyrrole (+16, +32, +48, +64) were shown
in Scheme 3.7.
N
N
OR2
R1
Peptide
Peptide
R1
R2
+16
N
O
N
OR2
R1
Peptide
Peptide
R1
R2
+32
N
O
N
OR2
R1
R1
R2
+48 +64
O
N
O
N
OR2
R1
R1
R2
O
O
Peptide
Peptide
Peptide
Peptide
Scheme 3.7 Possible structures for products of oxidized bispyrrole (+16, +32, +48, +64).
Previously, singlet oxygen, a major intermediate in the photodynamic effect,
generated by the combined action of light, a photosensitizing dye and oxygen, was
believed to insert oxygen atoms into biological substrates.36 For example, singlet oxygen
92

reacts with double bonds giving epoxides.37 The pyridinium bisretinoid A2E, a major
hydrophobic fluorophore of retinal pigment epithelium lipofuscin undergoes such
addition of multiple atoms of oxygen. Thus, a series of oxidized derivatives is generated
upon 10 min of blue light irradiation, each of which represents the addition of an oxygen
atom at a carbon-carbon double bond. A2E self-generates singlet oxygen with the singlet
oxygen in turn reacting with A2E to generate epoxides at carbon-carbon double bonds
(Figure 3.4). Epoxidation occurs at all nine double bonds in the hydrocarbon side chains
without modifying the pyridinium ring.38-39
A B
C D
+O
+O+O
+O
+O-18
-18
-18
-18
+O+O
+O
+O
+O
+O
-18
-18
-18-18
-18
Figure 3.3 MALDI-TOF MS spectra: panel A (4 days reaction) and B (15 days reaction)
for 36 min HPLC fractions, panel C (15 days reaction) for 30 min HPLC fraction, panel
93

D (15 days reaction) for 41 min HPLC fraction of products from the reaction of N-
acetylglycyllysine methyl ester with iso[4]LGE2 (molar ratio = 200:1 at 25 °C for 4 and
15 days).
A2E 1
Figure 3.4 Top: Structures of A2E (1), the bisoxirane (2) and nonaoxirane (3). Bottom:
ESI mass spectrum of A2E after illumination with blue light (430 nm; 10 min) to yield a
mixture of various oxygenated derivatives resulting in a series of ions that differ in m/z
by 16 owing to the addition of oxygen atoms to A2E. The addition of oxygen,
94

presumably to form epoxides, occurred at carbon-carbon double bonds of A2E, measured
by Micromass ESI-Q-TOF.38-39
To study their fragmentation patterns, tandem MS/MS analyses of pyrrole, lactam,
hydroxylactam and aminal cross-link ions were carried by Q-TOF mass spectrometry.
ESI-MS/MS analysis of Zip-Tip purified reaction mixture from N-acetylglycyllysine
methyl ester and iso[4]LGE2 (molar ratio = 200:1 at 25 °C for 4 days) provided additional
support for the structure postulated for the aminal cross-link. Previously, it was
postulated that an initial iso[4]LGE2 Schiff base adduct with the lysine residue of N-
acetyl-gly-lys-OMe is an electrophilic intermediate that undergoes aminal cross-linking
with another nucleophilic side chain residue of a second molecule of N-
acetylglycyllysine methyl ester.5 The MS/MS spectrum of the parent ion from the
putative aminal cross-link (m/z 853.5) shows peaks at m/z 853.5 and 835.5 corresponding
to the [M+H]+ ion from iso[4]LGE2-(N-acetyl-gly-lys-OMe) aminal cross-link and its
dehydrated form (Figure 3.5B). The MS/MS spectrum of m/z 835.5 exhibits two
diagnostic daughter ions at m/z 558.4 and 436.3. The ion at m/z 558.4, which is isobaric
with protonated dehydrated pyrrole, corresponds to loss of one molecule of N-acetyl-gly-
lys-OMe (-259) and one molecule of H2O (-18). The ion at m/z 436.3 corresponds to loss
of non-3-en-1-yne (C9H14) (-122). The proposed structure of each daughter ion is shown
in Figure 3.5 and Scheme 3.8. See appendix Figure 3.2S and Scheme 3.1S for spectra and
fragmentation structures of the pyrrole, lactam and hydroxylactam one-to-one adducts.
95

m/z
TOF MSMS 835.55ES+
TOF MSMS 853.58ES+
A
B
Relative Abundance
Relative Abundance
m/z
N CC5H11
OHO
HN
ONH
O COOCH3
OH
OH
NH
HN
NH
O
O COOCH3
H+N C
C5H11
OHO
HN
ONH
O COOCH3OH
NH
HN
NH
O
O COOCH3
H+
N CC5H11
OHO
HN
ONH
O COOCH3H+
N C OHO
HN
ONH
O COOCH3
H+
Figure 3.5 ESI-MSMS of molecular ions at A) m/z 835 (+ H+), and B) m/z 853 (+ H+)
corresponding to an iso[4]LGE2-(N-acetyl-gly-lys-OMe) aminal cross-link and its
dehydrated form.
N CC5H11
OHO
HN
ONH
O COOCH3
OH
OH
NH
HN
NH
O
O COOCH3835.5 (M - H2O + H+)
558.4 (835.5 - NGL - H2O)
558.4 (835.5 - NGL - H2O)
436.3 (558.4 - C9H14)
Scheme 3.8 Possible fragmentation of an iso[4]LGE2-(N-acetyl-gly-lys-OMe) aminal
cross-link at m/z 853.5 (M + H+).
96

(ii) Modification of N-acetyl-gly-lys methyl ester with fluorescently labeled iso[4]LGE2.
MALDI-TOF MS analysis of the reaction product mixture from the reaction of
fluorescently labeled iso[4]LGE2 (iso[4]LGE2-FluoTag) with N-acetylglycyllysine
methyl ester showed major ions at m/z 843, 859, 875 and 1120 corresponding to
protonated pyrrole, lactam, hydroxylactam and aminal cross-link, respectively, in
addition to ions resulting from the loss of one or two molecules of water from those
adducts. Figure 3.6 shows a representative spectrum showing the presence of various
adducts (see appendix Table 3.1S for molecular weight of adducts of N-acetyl-gly-lys-
OMe with iso[4]LGE2-FluoTag).
The reaction products were recorded (see appendix Figure 3.3S (1) for two different
reactant molar ratios of N-acetyl-glycyl-lysine methyl ester: iso[4]LGE2-FluoTag =
2000:1 and (2) for 200:1 measured after incubation for 1, 4, 15, 21 and 50 days). The
initial concentration of N-acetyl-glycyl-lysine methyl ester was constant while two
different ratios of iso[4]LGE2-FluoTag were added. At various incubation times, identical
aliquots of reaction mixture were purified by Ziptip and analyzed.
The 1:1 adducts, i.e., pyrrole, lactam and hydroxylactam and their dehydrated forms
were generated very quickly (within one day reaction) in the reaction of iso[4]LGE2-
FluoTag with N-acetyl-glycyl-lysine methyl ester (see appendix Figure 3.3S 1A). For the
aminal cross-link and its dehydrated forms, with longer incubation times, a higher
percentage of aminal cross-link adducts versus 1:1 adducts was generated. Specifically,
the aminal cross-link peak intensity was higher when N-acetyl-glycyl-lysine methyl ester
reaction time with iso[4]LGE2-FluoTag was 15 days than just 1 day (see appendix
Figures 3.3S 2A-E).
97

MALDI-TOF MS analysis was performed on 100 fmol of Zip-Tip purified reaction
mixture. Bispyrrole couldn’t be detected. Higher loading amount (200-500 fmol) was
also tested, but no bispyrrole was detected either. Thus, whereas bispyrrole is a minor but
significant product from the reaction of iso[4]LGE2 with N-acetylglysyllysine methyl
ester, it is not even a detectable product from the reaction of iso[4]LGE2-FluoTag with
this peptide. The difference between these adducts is isoLG ester versus carboxylate. One
possibility is that the carboxylate interacts with a putative cation radical intermediate and
alters its proclivity for combination with O2 versus a second molecule of pyrrole.
Figure 3.6 MALDI-TOF MS analysis of the product mixture from the reaction of N-
acetylglysyllysine methyl ester with iso[4]LGE2-FluoTag (molar ratio = 200:1) (50 mM
NEM∙OAc aqueous solution with 6% methanol, v/v, pH 8.6) at 25 °C for 15 days.
98

An investigation of the fragmentation patterns from tandem MS/MS analysis of
aminal cross-link ions by Q-TOF mass spectrometry provided additional support for the
structure postulated for this type of cross-link. The initial Schiff base of iso[4]LGE2-
FluoTag with the lysine residue of N-acetylglycyllysine methyl ester is an electrophilic
intermediate that undergoes aminal cross-linking with another nucleophilic side chain
residue of a second molecule of N-acetyl-gly-lys-OMe. The MS/MS spectrum of the
parent ion from the putative aminal cross-link (m/z 1120.6) shows peaks at m/z 1120.6
and 1102.5 corresponding to the [M+H]+ ion from a iso[4]LGE2-FluoTag-(N-acetyl-gly-
lys-OMe) aminal cross-link and its dehydrated form (Figure 3.7B). The MS/MS spectrum
of m/z 1102.5 exhibits two diagnostic daughter ions at m/z 825.5 and 703.4. The ion at
m/z 825.5, which is isobaric with protonated dehydrated pyrrole, corresponds to loss of
one molecule of N-acetyl-gly-lys-OMe (-259) and one molecule of H2O (-18). The ion at
m/z 703.4 corresponds to loss of non-3-en-1-yne (C9H14) (-122). The proposed structure
of each daughter ion is shown in Figure 3.7 and Scheme 3.9. See appendix Figure 3.4S
and Scheme 3.2S for spectra and fragmentation structures of one-to-one adducts: pyrrole,
lactam and hydroxylactam.
99

Figure 3.7 ESI-MSMS of molecular ions at A) m/z 1102.5 and B) m/z 1120.6
corresponding to a iso[4]LGE2-FluoTag-(N-acetyl-gly-lys-OMe) aminal cross-link
adduct and its dehydrated form.
N CC5H11
NHCH2CH2NHO
HN
ONH
O COOCH3
OH
OH
NH
HN
NH
O
O COOCH31102.6 (M - H2O + H+)
825.5 (1102.6 - NGL - H2O)
825.5 (1102.6 - NGL - H2O)
703.4 (825.5 - C9H14)
NO
N
SO2N
Scheme 3.9 Possible fragmentation of iso[4]LGE2-FluoTag aminal cross-link with N-
acetyl-gly-lys-OMe at m/z 1120.6 (M + H+).
TOF MSMS 1102.5ES+
TOF MSMS 1120.6ES+
A
B
m/z
100

(iii) Modification of the β-amyloid(11-17) peptide “EVHHQKL” by iso[4]LGE2.
MALDI-TOF MS analysis of the reaction product mixture from iso[4]LGE2 with β-
amyloid(11-17) peptide “EVHHQKL” showed major ions at m/z 1206, 1222, 1238, 2114
and 2410 corresponding to protonated pyrrole, lactam, hydroxylactam, aminal cross-link
and bispyrrole, respectively, in addition to ions resulting from the loss of one or two
molecules of water at m/z 1188, 1204, 1220, 1202, 2096, 2078, 2392, 2374. Figure 3.8
shows a representative spectrum that shows ions corresponding to various adducts (see
appendix Table 3.1S for the molecular weights of various iso[4]LGE2-β-amyloid
(EVHHQKL) peptide adducts).
Figure 3.8 MALDI-TOF MS analysis of the product mixture from the reaction of β-
amyloid(11-17) peptide “EVHHQKL” with iso[4]LGE2 (molar ratio = 200:1) (50 mM
NEM∙OAc aqueous solution with 4% methanol, v/v, pH 8.6) 25 °C for 7 days.
101

Reaction products were recorded for two reactant molar ratios (see appendix Figure
3.5S (1) for β-amyloid (EVHHQKL) peptide: iso[4]LGE2 = 2000:1 and (2) 200:1
measured after incubation for 1, 4, 7, 15, 21 and 50 days). The initial concentration of β-
amyloid (EVHHQKL) peptide was the same for both reactions while various ratios of
iso[4]LGE2 were added. At various incubation times, identical aliquots of the reaction
mixture were purified by Ziptip and then analyzed.
The 1:1 adducts, i.e., pyrrole, lactam and hydroxylactam and their dehydrated forms
were generated very quickly (within one day) in the reaction of iso[4]LGE2 with β-
amyloid (EVHHQKL) peptide (see appendix Figure 3.5S 1A). Cross-link adducts
included aminals, bispyrroles, trispyrroles and their dehydrated forms. With higher
iso[4]LGE2 ratio and longer incubation times, a higher percentage (comparison of cross-
link peak intensity with one-to-one adducts) of bispyrrole adducts generated. Specificity,
bispyrrole peak intensity was higher when the molar ratio of β-amyloid (EVHHQKL)
peptide to iso[4]LGE2 was 200:1 than 2000:1 (see appendix Figure 3.5S 2A-F).
Bispyrrole peak intensity was also higher (5 % more) when β-amyloid (EVHHQKL)
peptide reaction time with iso[4]LGE2 was 15 days than just 1 day. A similar trend was
observed with the dipeptide. It agrees with the conclusion the dipeptide or “EVHHQKL”
peptide interferes with bispyrrole formation. Presumably for the same reason, i.e. that
these peptides compete with a second molecule of pyrrole for a putative cation radical
intermediate.
More cross-link product versus bispyrrole formed during dipeptide reaction with
iso[4]LGE2. In contrast, more bispyrrole versus aminal cross-link product formed during
the reaction of “EVHHQKL” with iso[4]LGE2. It is tempting to speculate that the
102

“EVHHQKL” peptide-pyrrole modification forms aggregates that favor the dimerization
to produce bispyrrole since β-amyloid tends to spontaneously oligomerize. Future studies
might seek evidence for noncovalent aggregation of “EVHHQKL” peptide-pyrroles
under oxygen free conditions where oxidative transformations are precouded.
Polypyrroles, i.e., bispyrrole and trispyrrole showed a sequence of ions corresponding
to the further addition of oxygen atoms (+16). A series of peaks corresponding ions for
polypyrroles that had undergone the addition of one or more atoms of were observed. The
details will be discussed below.
The product mixture from reaction of iso[4]LGE2 with β-amyloid(11-17) peptide
“EVHHQKL” was separated by reverse phase HPLC using an acetonitrile/water gradient
and monitored by LC-ESI extracted ion channels (Figure 3.9). The major products
corresponding to aminal cross-link (trace 3.2C) and bispyrrole (trace 3.2B) eluted in
order of decreasing polarity at about 22 and 32 minutes.
Bispyrrole
Aminal crosslink
A
B
C
TIC
m/z 804 (+3)
m/z 1058(+2)
7e6
5.2e8
3e6
Figure 3.9 LC-ESI for the reaction mixture from β-amyloid(11-17) peptide and
iso[4]LGE2 (molar ratio = 200:1 at 25 °C for 7 days) with selected ion recording for
103

bispyrrole and aminal cross-links. A) TIC, B) bispyrrole m/z 804.6 (+3), m/z 603.7 (+4),
C) aminal cross-link m/z 1058.2 (+2).
MALDI-TOF MS analysis of the fractions eluted at 22 and 32 minutes confirmed the
presence of the aminal and bispyrrole cross-links. MALDI-TOF MS analysis of the
aminal fraction showed major ions at m/z 2114.2, 2096.2 and 2078.2 corresponding to
protonated aminal cross-link in addition to ions resulting from the loss of one and two
molecules of water. MALDI-TOF MS analysis of the bispyrrole fraction showed major
ions at m/z 2410.3, 2392.3 and 2374.3 corresponding to protonated bispyrrole in addition
to ions resulting from the loss of one and two molecules of water. The mass spectra of the
principle products are shown in Figure 3.10. A series of peaks are present in the mass
spectrum of the isoLG-β-amyloid(11-17) peptide chromatographic fraction corresponding
to the addition of one or more atoms of oxygen to the bispyrrole, and the relative amount
of these products of further oxidation as indicated by the relative peak intensity of further
oxidized bispyrrole (products with addition of oxygen atoms) compared with bispyrrole
and its dehydrated forms after 15 days modification than after 7 days (Figure 3.10 panels
A and B, respectively).
A series of products of further addition of oxygen (multiple +16 m/z increments)
pattern was also observed for the trispyrrole adduct. Thus, pyrrole oligomerizes form
polypyrroles, and all of these products undergo further oxidation. Similar behavior was
observed for the iso[4]LGE2-(N-acetyl-gly-lys-OMe) bispyrrole cross-link and discussed
in section “(i) Iso[4]LGE2 modification of N-acetyl-gly-lys methyl ester”.
104

A B
C
-18
-18
-18
-18
-18
-18
D
Figure 3.10 MALDI-TOF MS spectra: panel A (7 days reaction) and B (15 days
reaction) for 32 min HPLC fractions, panel C and D (7 days reaction) for 22 min, 15 min
HPLC fraction of products from reaction of the β-amyloid(11-17) peptide “EVHHQKL”
with iso[4]LGE2 reaction (molar ratio = 200:1 at 25 °C).
(iv) Modification of β-amyloid(11-17) peptide “EVHHQKL” with fluorescently labeled
iso[4]LGE2.
MALDI-TOF MS analysis of the reaction product mixture from iso[4]LGE2-FluoTag
with the β-amyloid (EVHHQKL) peptide showed major ions at m/z 1473, 1489 and 1505
corresponding to protonated pyrrole, lactam and hydroxylactam in addition to ions
105

resulting from the loss of one or two molecules of water from those adducts. Figure 3.11
is a representative spectrum that shows ions corresponding to various adducts (see
appendix Table 3.1S for molecular weight of adducts of β-amyloid (EVHHQKL) peptide
with iso[4]LGE2-FluoTag).
Reaction products were recorded for two reactant molar ratios (see appendix Figure
3.6S (1) for β-amyloid (EVHHQKL) peptide: iso[4]LGE2-FluoTag = 2000:1 and (2)
200:1 measured after incubation for 1, 4, 15, 21 and 50 days. The initial concentration of
β-amyloid (EVHHQKL) peptide was the same for both reactions while various ratios of
iso[4]LGE2-FluoTag were added. At various incubation times, identical aliquots of
reaction mixture were purified by Ziptip and analyzed.
The 1:1 adducts, i.e., pyrrole, lactam and hydroxylactam and their dehydrated forms
were generated very quickly (within one day) in the reaction of iso[4]LGE2-FluoTag with
β-amyloid (EVHHQKL) peptide (appendix Figure 3.6S 1A). MALDI-TOF analysis was
performed on 100 fmol of Zip-Tip purified reaction mixture. Compared with 1:1 adducts,
aminal cross-link and bispyrrole were barely be detected.
106

Figure 3.11 MALDI-TOF MS analysis of the product mixture from the reaction β-
amyloid (EVHHQKL) peptide with iso[4]LGE2-FluoTag (molar ratio = 200:1) (50 mM
NEM∙OAc aqueous solution with 6% methanol, v/v, pH 8.6) at 25 °C for 15 days.
107

3.3 Conclusions
IsoLG modification of proteins is associated with loss-of-function, cross-linking and
aggregation. N-Acetyl-glycyl-lysine methyl ester (N-acetyl-Gly-Lys-OMe) and β-
amyloid(11-17) peptide “EVHHQKL” were used as models for characterizing the cross-
linking of protein lysyl residues resulting from adduction of iso[4]LGE2. Aminal,
bispyrrole and trispyrrole cross-links of two model peptides, were identified and fully
characterized by mass spectrometry, and a large variety of derivatives of the bispyrrole
cross-link containing one or more additional atoms of oxygen were discovered. More
aminal cross-link product versus bispyrrole is formed during dipeptide reaction with
iso[4]LGE2, and bispyrrole is almost undetectable in the reaction of a fluorescently
labeled iso[4]LGE2 with dipeptide. In contrast, more bispyrrole versus aminal cross-link
product formed during the reaction of “EVHHQKL” with iso[4]LGE2. It is tempting to
speculate that the “EVHHQKL” peptide-pyrrole modification forms aggregates that favor
the dimerization to produce bispyrrole since β-amyloid tends to spontaneously
oligomerize. Future studies might seek evidence for noncovalent aggregation of
“EVHHQKL” peptide-pyrroles under oxygen free conditions where oxidative
transformations are precouded.
With higher iso[4]LGE2 ratio:peptide, a higher percentage of bispyrrole versus 1:1
adducts was generated from both N-acetyl-glycyl-lysine methyl ester and β-amyloid
(EVHHQKL) peptide. This agrees with the conclusion that these peptides interfere with
bispyrrole formation. Presumably for the same reason, i.e. that these peptides compete
with a second molecule of pyrrole for a cation radical intermediate that will be discussed
in the following chapter.
108

3.4 Experimental Procedures
3.4.1 General methods. All chemicals used were high purity analytical grade. The
following commercially available materials were used as received: N-acetylglycyllysine
methyl ester was from Bachem (Torrance, CA), β-Amyloid (11-17) peptide
“EVHHQKL” was from Peptide 2.0 Inc (Chantilly, VA), N-ethylmorpholine was from
Aldrich (Milwaukee, WI), and all other chemicals were obtained from Fisher Scientific
Co (Chicago, IL). Iso[4]LGE2 was prepared by Dr. Jim Laird and iso[4]LGE2-FluoTag
was prepared as described in Chapter 2. Chromatography was performed with ACS grade
solvents.
3.4.2 Peptide modification with iso[4]LGE2 or iso[4]LGE2-FluoTag. Reactant stocks
were prepared as follows: 25 mM N-acetylglycyllysine methyl ester and β-amyloid (11-
17) peptide “EVHHQKL” in NEM∙OAc (50 mM, pH 8.6), 1 mg/ml iso[4]LGE2 (without
or with fluorescence tag) in MeOH.
4.4 µl of iso[4]LGE2 (12.5 nmol, 4.4 µg) or 7.7 µl of iso[4]LGE2-FluoTag (12.5
nmol, 7.7 µg) was added to N-acetylglycyllysine methyl ester (2.5 µmol, 647.5 µg) in 50
mM pH 8.6 NEM∙OAc (100 µl) (molar ratio of iso4: dipeptide = 1:200) and reacted
under air on shaker (IKA MTS 2/4 digital microtiter shaker, 500 rpm) at 25 °C. At
various times (1, 4, 7, 15, 21 and 50 days), an aliquot (~1.2 nmol, 10 µl) of reaction
mixture was quickly dried using a high-speed vacuum evaporator and then stored at -20
°C until analysis. The aliquot was analyzed with a MALDI-TOF, triple quadrupole or Q-
TOF mass spectrometer. Two different molar ratios (iso4: peptide = 1:2000 and 1:200)
were studied. β-amyloid (11-17) peptide “EVHHQKL” was treated the same way.
109

3.4.3 Analysis of adducts using MALDI-TOF. Initial diagnostic analysis of the samples
was performed by MALDI-TOF mass spectrometry on an AB Sciex4800 Plus MALDI
TOF/TOF™ Analyzer equipped with a UV laser (355 nm) and reflector mode with a
matrix of α-cyano-4-hydroxycinnamic acid (CHCA, 10 mg/ml in acetonitrile/water/3%
formic acid, 5:4:1, v/v/v). A peptide mass standards kit for calibration of AB Sciex
MALDI-TOFTM instruments was used to calibrate the plate. Each of the reaction
aliquots was resuspended in 10% aqueous acetonitrile solution and vortexed to mix well
before C18 Ziptip purification (Millipore, ZipTip with 0.6 µL C18 resin). To wet the C18
Ziptip pipettor plunger was depressed to a dead stop using the maximum volume setting
of 20 µl. Acetonitrile (wetting solution) was aspirated into tip then dispensed to waste
and the process repeated twice. The tip was equilibrated for binding by washing three
times with the 0.1% formic acid/water (equilibration solution). Samples were bound to
ZipTip by fully depressing the pipettor plunger to a dead stop then aspirating and
dispense the sample 3 to 7 cycles. The tip was then washed with at least 3 cycles of 0.1%
formic acid/water (wash solution) dispensing to waste using. The samples were eluted
with 5 µl of 80% acetonitrile/0.1% formic acid (elution solution) into a clean vial to give
a final concentration of 100 fmol/µl by aspirating and dispensing eluant through the
ZipTip at least 3 times without introducing air. Two microliters of the sample was then
mixed with 1 µl of matrix, and then 1.5 µl were applied to the plate spot and allowed to
dry. The data was processed with Data Explorer 4.0 (AB Sciex) software.
110

Table 3.1: Optimized parameters for MALDI-TOF mass spectrometer.
Acquisition
Method
Parameters Data Parameters Data Shots/sub-spectrums 50 Bin size (ns) 0.5 Total shots/spectrum 1000 Input bandwidth (MHZ) 500 Laser intensity 3000 Detector voltage multiplier 0.92 Vertical scale (v) 0.5 Final detector voltage (KV) 2.015
Processing
Method
Calibration Parameters Data Peak Detection Parameters Data Min S/N 20 Min S/N 10 Mass tolerance (m/z) 2 Local noise window width (m/z) 250 Min peaks to match 4 Min peak width (bins) 2.9 Max error (ppm) 100 Mass resolution 22000
3.4.4 LC-MS (triple quadrupole) analysis. Iso[4]LGE2-modified β-amyloid(11-17)
peptide was separated by HPLC with a 300 µm x 250 mm i.d. C18 5 µm column
(DIONEX AcclaimTM). Chromatography was carried out with linear elution gradient
(elute A, 0.1% TFA/H2O; elute B, 0.1% TFA/ACN) at a flow rate of 20 µl/min. The
samples were loaded onto the column with 2% B for 10 min. The gradient was as
follows: 2-12% B over 5 min, to 60 % over 48 min, to 98% B over 2 min, and hold for 10
min. Iso[4]LGE2 modified acetyl-gly-lys-O-methyl ester was seperated by HPLC with a
2.1 x 100 mm i.d. C18 1.8 µm column (Waters Acquity UPLC HSS). Chromatography
was carried out with linear elution gradient (elute A, 0.1% FA/H2O; elute B, 0.1%
FA/ACN) at a flow rate of 70 µl/min. Sample was loaded onto the column with 2% B for
10 min. The gradient was as follows: 2-10% B over 2 min, to 98 % within 25 min, and
hold for 18 min. The effluent was monitored by ESI-MS/MS with a API-3000 triple
quadrapole electrospray mass spectrometer (Applied Biosystems Inc.) to determine the
retention time of different oxidized adducts. The instrument was operated in the positive
mode and high-pressure nitrogen was used as source gas and scanning from m/z 220-
2000 using the parameters listed in table 3.2.
111

Table 3.2: Optimized parameters for triple quadrupole mass spectrometer.
Parameters Data Declustering Potential (DP) 30 Focus Potential (FP) 250 Entrance Potential (EP) 10 Nebulizer Gas (NEB) 10 Curtain Gas (CUR) 8 Ion Spray Voltage (IS) 4000 Temperature (TEM) 200
3.4.5 LC- MS/MS (Q-TOF) analysis. The samples were analyzed by a quadrupole-time
of flight (Q-TOF-2) mass spectrometer (Micromass) equipped with MassLynxTM
acquisition and processing software. 2 pmol iso[4]LGE2 or iso[4]LGE2-FluoTag modified
N-acetylglycyllysine methyl ester (molar ratio of N-acetylglycyllysine methyl ester:
iso[4]LGE2 = 200:1, 25 °C for 4 days), prepared as described in Section 3.4.2, was
analyzed by Q-TOF. For sample preparation, 62.5 pmol of reaction mixture from the
reaction of iso[4]LGE2 with N-acetylglycyllysine methyl ester was resuspended in 250 µl
10% acetonitrile. An aliquot (8 µl, 2 pmol) of this solution was purified with a Zip-Tip as
described in section 3.4.3 and eluted in 10 µl of 80% acetonitrile/0.1% formic acid. This
purified eluate was quickly dried using a high-speed vacuum evaporator and then
dissolved in 15 µl of 5% acetonitrile/2% formic acid and injected into the Q-TOF. The
samples were infused using a 100 µl Hamilton syringe onto a capillary column
(PicoFritTM 0.075 x 70 mm, 15 µ tip ID; New Objective Inc., Woburn, MA) at 0.5 µl/min
using a syringe pump (Harvard apparatus, Pump 11) and ionized immediately using an
electrosprayer designed in-house. The mass spectrometer was operated in standard MS
and MS/MS switching mode. The MS data was collected for the mass range of 50-1500
m/z and MS/MS data was analysed with Micromass software (MassLynx) Version 3.5.
112

The machine was calibrated using a solution of 2 fmol of [Glu1]-Fibrinopeptide B
(Sigma) in 50% aqueous acetonitrile (+0.1% formic acid) infused using glass
micropipette with 5 µ tip. The intensity of the peak from the MS/MS spectrum at 785.4
m/z was used as a reference for calibration. The final mass measurement accuracy of ≤ 10
ppm was achieved before the use of the machine.
113

3.5 References
1. Kahn, M. G.; Giblin, F. J.; Epstein, D. L. (1983) Glutathione in calf trabecular
meshwork and its relation to aqueous humor outflow facility. Invest Ophthalmol Vis Sci
24 (9), 1283-1287.
2. Salomon, R. G. (2005) Distinguishing levuglandins produced through the
cyclooxygenase and isoprostane pathways. Chem Phys Lipids 134 (1), 1-20.
3. Morrow, J. D.; Harris, T. M.; Roberts, L. J., 2nd (1990) Noncyclooxygenase
oxidative formation of a series of novel prostaglandins: analytical ramifications for
measurement of eicosanoids. Anal Biochem 184 (1), 1-10.
4. Amarnath, V.; Valentine, W. M.; Amarnath, K.; Eng, M. A.; Graham, D. G.
(1994) The mechanism of nucleophilic substitution of alkylpyrroles in the presence of
oxygen. Chem Res Toxicol 7 (1), 56-61.
5. Difranco, E.; Subbanagounder, G.; Kim, S.; Murthi, K.; Taneda, S.; Monnier, V.
M.; Salomon, R. G. (1995) Formation and stability of pyrrole adducts in the reaction of
levuglandin E(2) with proteins. Chem Res Toxicol 8 (1), 61-67.
6. Iyer, R. S.; Ghosh, S.; Salomon, R. G. (1989) Levuglandin-E2 cross-links
proteins. Prostaglandins 37 (4), 471-480.
7. Jirousek, M. R.; Murthi, K. K.; Salomon, R. G. (1990) Electrophilic levuglandin
E2-protein adducts bind glycine: A model for protein cross-linking. Prostaglandins 40
(2), 187-203.
8. Murthi, K. K.; Friedman, L. R.; Oleinick, N. L.; Salomon, R. G. (1993) Formation
of DNA-protein cross-links in mammalian cells by levuglandin E2. Biochemistry 32 (15),
4090-4097.
114

9. Cox, B.; Murphey, L. J.; Zackert, W. E.; Chinery, R.; Graves-Deal, R.; Boutaud,
O.; Oates, J. A.; Coffey, R. J.; Morrow, J. D. (2002) Human colorectal cancer cells
efficiently conjugate the cyclopentenone prostaglandin, prostaglandin J(2), to glutathione.
Biochim Biophys Acta 1584 (1), 37-45.
10. Carrier, E. J.; Amarnath, V.; Oates, J. A.; Boutaud, O. (2009) Characterization of
covalent adducts of nucleosides and DNA formed by reaction with levuglandin.
Biochemistry 48 (45), 10775-10781.
11. Zagol-Ikapitte, I.; Bernoud-Hubac, N.; Amarnath, V.; Roberts, L. J., 2nd;
Boutaud, O.; Oates, J. A. (2004) Characterization of bis(levuglandinyl) urea derivatives
as products of the reaction between prostaglandin H2 and arginine. Biochemistry 43 (18),
5503-5510.
12. Salomon, R. G.; Jirousek, M. R.; Ghosh, S.; Sharma, R. B. (1987) Prostaglandin
endoperoxides .21. covalent binding of levuglandin-E2 with proteins. Prostaglandins 34
(5), 643-656.
13. Boutaud, O.; Brame, C. J.; Salomon, R. G.; Roberts, L. J., 2nd; Oates, J. A.
(1999) Characterization of the lysyl adducts formed from prostaglandin H2 via the
levuglandin pathway. Biochemistry 38 (29), 9389-9396.
14. Zhu, M.; Spink, D. C.; Yan, B.; Bank, S.; DeCaprio, A. P. (1994) Formation and
structure of cross-linking and monomeric pyrrole autoxidation products in 2,5-
hexanedione-treated amino acids, peptides, and protein. Chem Res Toxicol 7 (4), 551-
558.
115

15. Zhu, M.; Spink, D. C.; Yan, B.; Bank, S.; DeCaprio, A. P. (1995) Inhibition of
2,5-hexanedione-induced protein cross-linking by biological thiols: chemical mechanisms
and toxicological implications. Chem Res Toxicol 8 (5), 764-771.
16. Boutaud, O.; Ou, J. J.; Chaurand, P.; Caprioli, R. M.; Montine, T. J.; Oates, J. A.
(2002) Prostaglandin H2 (PGH2) accelerates formation of amyloid beta1-42 oligomers. J
Neurochem 82 (4), 1003-1006.
17. Davies, S. S.; Amarnath, V.; Montine, K. S.; Bernoud-Hubac, N.; Boutaud, O.;
Montine, T. J.; Roberts, L. J. (2002) Effects of reactive gamma-ketoaldehydes formed by
the isoprostane pathway (isoketals) and cyclooxygenase pathway (levuglandins) on
proteasome function. FASEB J 16 (3), 715-717.
18. Murthi, K. K.; Salomon, R. G.; Sternlicht, H. (1990) Levuglandin E2 inhibits
mitosis and microtubule assembly. Prostaglandins 39 (6), 611-622.
19. Boutaud, O.; Andreasson, K. I.; Zagol-Ikapitte, I.; Oates, J. A. (2005)
Cyclooxygenase-dependent lipid-modification of brain proteins. Brain Pathol 15 (2),
139-142.
20. Tak, P. P.; Zvaifler, N. J.; Green, D. R.; Firestein, G. S. (2000) Rheumatoid
arthritis and p53: how oxidative stress might alter the course of inflammatory diseases.
Immunol Today 21 (2), 78-82.
21. Murphy, M. E.; Kehrer, J. P. (1989) Oxidative stress and muscular dystrophy.
Chem Biol Interact 69 (2-3), 101-173.
22. Butterfield, D. A.; Drake, J.; Pocernich, C.; Castegna, A. (2001) Evidence of
oxidative damage in Alzheimer's disease brain: central role for amyloid β-peptide. Trends
Mol Med 7 (12), 548-554.
116

23. Butterfield, D. A.; Lauderback, C. M. (2002) Lipid peroxidation and protein
oxidation in Alzheimer's disease brain: potential causes and consequences involving
amyloid beta-peptide-associated free radical oxidative stress. Free Radic Biol Med 32
(11), 1050-1060.
24. Glenner, G. G.; Wong, C. W. (1984) Alzheimer's disease: initial report of the
purification and characterization of a novel cerebrovascular amyloid protein. Biochem
Biophys Res Commun 120 (3), 885-890.
25. Selkoe, D. J. (2001) Alzheimer's disease: genes, proteins, and therapy. Physiol
Rev 81 (2), 741-766.
26. Lambert, M. P.; Barlow, A. K.; Chromy, B. A.; Edwards, C.; Freed, R.; Liosatos,
M.; Morgan, T. E.; Rozovsky, I.; Trommer, B.; Viola, K. L.; Wals, P.; Zhang, C.; Finch,
C. E.; Krafft, G. A.; Klein, W. L. (1998) Diffusible, nonfibrillar ligands derived from
Abeta1-42 are potent central nervous system neurotoxins. Proc Natl Acad Sci U S A 95
(11), 6448-6453.
27. Santiard-Baron, D.; Lacoste, A.; Ellouk-Achard, S.; Soulie, C.; Nicole, A.;
Sarasin, A.; Ceballos-Picot, I. (2001) The amyloid peptide induces early genotoxic
damage in human preneuron NT2. Mutat Res 479 (1-2), 113-120.
28. Tong, L.; Thornton, P. L.; Balazs, R.; Cotman, C. W. (2001) Beta -amyloid-(1-42)
impairs activity-dependent cAMP-response element-binding protein signaling in neurons
at concentrations in which cell survival Is not compromised. J Biol Chem 276 (20),
17301-17306.
117

29. Varadarajan, S.; Yatin, S.; Aksenova, M.; Butterfield, D. A. (2000) Review:
Alzheimer's amyloid beta-peptide-associated free radical oxidative stress and
neurotoxicity. J Struct Biol 130 (2-3), 184-208.
30. Yatin, S. M.; Varadarajan, S.; Link, C. D.; Butterfield, D. A. (1999) In vitro and
in vivo oxidative stress associated with Alzheimer's amyloid beta-peptide (1-42).
Neurobiol Aging 20 (3), 325-330; discussion 339-342.
31. Yatin, S. M.; Yatin, M.; Aulick, T.; Ain, K. B.; Butterfield, D. A. (1999)
Alzheimer's amyloid beta-peptide associated free radicals increase rat embryonic
neuronal polyamine uptake and ornithine decarboxylase activity: protective effect of
vitamin E. Neurosci Lett 263 (1), 17-20.
32. Yatin, S. M.; Yatin, M.; Varadarajan, S.; Ain, K. B.; Butterfield, D. A. (2001)
Role of spermine in amyloid beta-peptide-associated free radical-induced neurotoxicity. J
Neurosci Res 63 (5), 395-401.
33. Varadarajan, S.; Kanski, J.; Aksenova, M.; Lauderback, C.; Butterfield, D. A.
(2001) Different mechanisms of oxidative stress and neurotoxicity for Alzheimer's A
beta(1--42) and A beta(25--35). J Am Chem Soc 123 (24), 5625-5631.
34. Yatin, S. M.; Varadarajan, S.; Butterfield, D. A. (2000) Vitamin E Prevents
Alzheimer's Amyloid beta-Peptide (1-42)-Induced Neuronal Protein Oxidation and
Reactive Oxygen Species Production. J Alzheimers Dis 2 (2), 123-131.
35. Boutaud, O.; Montine, T. J.; Chang, L.; Klein, W. L.; Oates, J. A. (2006) PGH2-
derived levuglandin adducts increase the neurotoxicity of amyloid beta1-42. J
Neurochem 96 (4), 917-923.
118

36. Straight, R. C.; Spikes, J. D., Singlet Oxygen. CRC: Boca Raton, FL, 1985; Vol.
4, 91.
37. Schulte-Elte, K. H.; Gadola, M.; Ohloff, G. (1973) Oxydative Umsetzungen in der
Damascon-Reihe. 1ΔgO2- und SeO2-Oxydation von β-Damascenon. Helv Chim Acta 56
(6), 2028-2035.
38. Ben-Shabat, S.; Itagaki, Y.; Jockusch, S.; Sparrow, J. R.; Turro, N. J.; Nakanishi,
K. (2002) Formation of a nonaoxirane from A2E, a lipofuscin fluorophore related to
macular degeneration, and evidence of singlet oxygen involvement. Angew Chem Int Ed
Engl 41 (5), 814-817.
39. Sparrow, J. R.; Vollmer-Snarr, H. R.; Zhou, J.; Jang, Y. P.; Jockusch, S.; Itagaki,
Y.; Nakanishi, K. (2003) A2E-epoxides damage DNA in retinal pigment epithelial cells.
Vitamin E and other antioxidants inhibit A2E-epoxide formation. J Biol Chem 278 (20),
18207-18213.
119

CHAPTER 4
GENERATION OF ISOLG-PYRROLE BY THE REACTION OF
ISOLG WITH N-ACETYL-GLY-LYS-OMe IN THE ABSENCE OF O2
AND THE OXIDATION OF ISOLG-PYRROLE TO FORM
PYRROLE-PYRROLE CROSS-LINKS AND AN ADDUCT WITH N-
ACETYLCYSTEINE

4.1 Background
4.1.1 Purpose
Presumably, pyrrole-pyrrole cross-links are produced by oxidative coupling of
LG/isoLG-pyrroles. To enable examination of the mechanism of isoLG-pyrrole-pyrrole
cross-linking, it is desirable to synthesize pure isoLG-pyrroles. In this chapter, the
preparation of pure isoLG-pyrroles and a study of their chemistry will be described.
Evidence is presented that oxidation and cross-linking of isoLG-derived pyrroles is
autocatalytic the initiation of which can be catalyzed by tetramethylpyrrolidine-N-oxide,
a stable free radical.
4.1.2 LGs and isoLGs modification with proteins
Previously, we discovered that the prostaglandin endoperoxide intermediate of the
cyclooxygenase pathway, and its stereo and structural isomers (isoprostane
endoperoxides), which are generated through free radical-induced oxidation of
arachidonyl phospholipids, undergo rearrangements that produce γ-ketoaldehydes,
levuglandins (LGs) and isoLGs, respectively. Adduction of LGs/isoLGs to proteins
initially produces pyrroles through hemiacetals within seconds. The pyrroles are readily
oxidized to lactams and hydroxylactams that are the major lysyl derivatives present in
proteins in vivo. LG/isoLG adduction also generates protein-protein cross-links within
minutes.1 Pyrrole-pyrrole cross-link and aminal cross-link structures shown in Scheme
4.1 have been postulated, but no direct molecular level evidence for the cross-link
structure(s) has been reported.
121

Scheme 4.1 LG/isoLG modification of proteins
122

4.2 Results and discussion
4.2.1 Preparation of iso[4]LGE2-pyrrole under anerobic conditions.
As reported in chapter 3, the formation of pyrrole-pyrrole cross-links was detected
upon adduction of isoLG with N-acetyl-gly-lys methyl ester. Pyrrole-pyrrole dimer
formation is an oxidative process. Therefore, cross-linking is not expected to occur under
anerobic conditions. To test this hypothesis, the treatment of acetyl-gly-lys methyl ester
was conducted in an oxygen-free aqueous solution containing sodium dithionite, an
oxygen scavenger. As predicted, cross-linking was completely prevented. However,
immediately after removing Na2S2O4 (as describe below), the iso[4]LGE2-pyrrole
(Scheme 4.2) derivative underwent oxidation and dimerization that was extensive after
the solution was aerated with oxygen overnight (Figure 4.1). It should be noted that this
preparation contained a mixture of iso[4]LGE2-pyrrole unreacted iso[4]LGE2 and
dipeptide as well as byproducts derived from them, possibly including peracid generated
through oxidation of the aldehyde functional group in iso[4]LGE2.
Incubation of N-acetyl-gly-lys methyl ester with iso[4]LGE2 under an atmosphere of
argon, including bubbling argon gas before and during reaction, produced iso[4]LGE2-
pyrrole (4.3) as well as products of further oxidation, i.e., lactam, hydroxylactam. Sodium
dithionite (Na2S2O4) is an oxygen scavenger that can maintain the oxygen concentration
at extremely low levels.2 Only iso[4]LGE2-pyrrole (4.3) was produced if the adduction
reaction was carried out in water solution in the presence sodium dithionite under an
atmosphere of argon at 25 °C. An excess of acetyl-gly-lys methyl ester (20 equivalents)
was incubated with iso[4]LGE2 to favor complete consumption of the γ-ketoaldehyde.
Since sodium dithionite prevented oxidative transformations of the pyrrole when exposed
123

to air, it was essential to completely remove dithionite from the sample in order to study
further oxidative reactions. The reaction solution was concentrated under high vacuum to
remove water and methanol was added to dissolve the organic product. Then the mixture
was loaded onto a column of silica gel packed with a slurry in EtOAc and eluted with
methanol to remove most of sodium dithionite. After concentration under reduced
pressure, reagent grade chloroform that had been sparged with argon was added to
dissolve iso[4]LGE2-pyrrole and the solution was filtered to remove sodium dithionite
and any silical gel that had dissolved in the methanol. The crude iso[4]LGE2-pyrrole
solution was concentrated by evaporation with argon gas and re-suspended in 10%
acetonitrile/water for further use.
NH
O HN COOCH3
NH2O+
C
O
C5H11
OO
OH
OH Ar GasNa2S2O4
H2O as solvent
4.1 4.2
N CC5H11
OHO
HN
ONH
O COOCH3
OH4.3
Scheme 4.2 Synthesis of iso[4]LGE2-pyrrole.
Before and immediately after removing the sodium dithionite, no oxidation products
were detected by MALDI initially (Figures 4.1A and B). Upon exposure to air, within
one hour, it formed various oxidized derivatives, e.g., lactam, hydroxylactam, and
bispyrrole (Figure 4.1C). After bubbling O2 overnight, more oxidized products had been
generated, including aminal cross-link including bispyrrole as the major product (Figure
4.1D). Because this sample of iso[4]LGE2-pyrrole contained unreacted acetyl-gly-lys
methyl ester, the formation of peptide-peptide aminal cross-link presumably was a
consequence of the presence of excess N-acetyl-gly-lys methyl ester in the sample,
perhaps involving a reaction between the pyrrole and the nucleophilic side chain residue
of a second molecule of dipeptide.
124

Removal of dithionite
Oxidation
under Air1 h
Aeration overnight
Figure 4.1 MALDI-TOF MS analysis of iso[4]LGE2-pyrrole and oxidation products
generated upon short term exposure of the reaction mixture from iso[4]LGE2 with N-
acetylglycyllysine methyl ester in the presence of dithionite to air or longer term aeration
with oxygen before or after removal of dithionite.
Panel A: Reaction mixture from N-acetylglycyllysine methyl ester and iso[4]LGE2
(molar ratio = 20:1) in the presence sodium dithionite under an atmosphere of argon at 25
°C overnight. Panel B: Iso[4]LGE2-pyrrole immediately after removing the sodium
dithionite. Panel C: Reaction mixture of iso[4]LGE2-pyrrole after 1 h exposure to air.
Panel D: Reaction mixture of iso[4]LGE2-pyrrole after bubbling O2 overnight.
125

4.2.2 The oxidation of HPLC-purified iso[4]LGE2-pyrrole with air exhibits an
induction period.
A sample of iso[4]LGE2-pyrrole contaminated with excess N-acetyl-gly-lys methyl
ester and unreacted iso[4]LGE2 was purified by HPLC using an acetonitrile/water
gradient using LC-ESI to detect the components of interest by monitoring the appropriate
molecular weights in the extracted ion channels (Figure 4.2). The fraction eluting at 12
min contained pure iso[4]LGE2-pyrrole. The structural integrity of HPLC-purified
iso[4]LGE2-pyrrole was confirmed by MALDI-TOF MS (See appendix Figure 4.1S).
Iso[4]LGE2-pyrrole-H2O ion Extraction -- 558 (+1)
Iso[4]LGE2-pyrrole-H2O (558, +1)
Acetyl-Gly-Lys-O-Methyl Ester ion Extraction -- 260 (+1)
A
TIC
B
C
Iso[4]LGE2-pyrrole
Acetyl-gly-lys-O-methyl ester
LC monitoring for pyrrole
LC monitoring for dipeptide
MS of pyrrole
Figure 4.2 LC-ESI for the purification of iso[4]LGE2-pyrrole with selected ion recording
for iso[4]LGE2-pyrrole. A) TIC, B) LC monitoring and MS analysis for iso[4]LGE2-
pyrrole-H2O (m/z 558 (+ H+)), C) LC monitoring for N-acetyl-gly-lys methyl ester (m/z
260 (+ H+)).
MALDI and triple quadrupole mass spectrometry was used to confirm that purified
iso[4]LGE2-pyrrole was not contaminated with N-acetyl-gly-lys methyl ester or
126

iso[4]LGE2. The background of matrix for MALDI-TOF and solvent used in triple
quadrupole mass spectrometry was determined to not exhibit any peaks corresponding to
N-acetyl-gly-lys methyl ester or iso[4]LGE2 in the matrix or solvent (no m/z around 260
(+1 for N-acetyl-gly-lys methyl ester) or 353 (+1 for iso[4]LGE2)). Peaks not
corresponding to the matrix or solvent peaks could be detected upon analysis by MALDI
or triple quadrupole mass spectrometry of N-acetyl-gly-lys methyl ester or iso[4]LGE2
(see appendix Figure 4.2S).
The reaction time course (Figure 4.3) for incubation of pure iso[4]LGE2-pyrrole
under air at 25 °C in aqueous acetonitrile solution (10% acetonitrile/H2O) was monitored
for eight days (see appendix Figure 4.3S for details). MALDI-TOF mass spectra of pure
iso[4]LGE2-pyrrole and the reaction product mixture after exposure to air for eight days
are shown in Figure 4.4. Compared to iso[4]LGE2-pyrrole with un-reacted N-acetyl-gly-
lys methyl ester and iso[4]LGE2, pure iso[4]LGE2-pyrrole was much more stable under
air. During the first two days, almost no oxidized product was formed. At longer reaction
times, 1:1 adducts, i.e., lactam, hydroxylactam and bispyrrole became prominent. But, as
expected, no peptide-peptide aminal cross-link formed since excess N-acetyl-gly-lys
methyl ester and iso[4]LGE2 were not present.
The pure pyrrole obtained by preparative HPLC (Figure 4.2: LC-ESI for the
purification of iso[4]LGE2-pyrrole) was no longer prone toward oxidation or dimerization
when exposed to oxygen. The product evolution time course for the reaction of this
HPLC-purified iso[4]LGE2-pyrrole with oxygen exhibited an induction period followed
by rapid increase in the generation of all products (Figure 4.3).
127

0 day 2 days 4 days 6 days 8 days0
20
40
60
80
100Pe
rcen
t of T
otal
Time
Bispyrrole-H2O Lactam-H2O Hydroxylactam-2H2O Pyrrole-H2O
Time Course of Pure Iso[4]LGE2-pyrrole Reaction under Air
Figure 4.3 Time course of the oxidative reactions of pure iso[4]LGE2-pyrrole under air at
25 °C for 8 days. As shown in appendix Figure 4.3S, the observed peak intensity for
pyrrole-H2O is 100 (for each time point). Peak intensity of bispyrrole-H2O, lactam-H2O
and hydroxylactam-2H2O was zero for the first two days, and then gradually increased.
The total intensity is the sum of pyrrole-H2O, bispyrrole-H2O, lactam-H2O and
hydroxylactam-2H2O. The peak intensity of each product is divided by the total intensity
to provide the percent of total shown in the figure.
128

Figure 4.4 MALDI-TOF MS of pure iso[4]LGE2-pyrrole and its oxidation products:
lower trace = pure iso[4]LGE2 -pyrrole, upper trace = after incubation under air at 25 °C
for 8 days.
4.2.3 Reaction of pure iso[4]LGE2-pyrrole with Nα-acetylcysteine.
The possible role of nucleophilic thiols and amines in pyrrole-mediated cross-linking
of proteins was investigated in a model system. To test the possibility that nucleophilic
thiols can participate in isoLG-derived pyrrole-mediated cross-linking, pure iso[4]LGE2-
pyrrole was incubated in the presence of N-acetylcysteine under air. N-acetylcysteine
(4.4) was added to pure iso[4]LGE2-pyrrole (4.3) and incubated under air at 25 °C for
several days (Scheme 4.3).
129

N CC5H11
OHO
HN
ONH
O COOCH3
OH
HN COOH
O CH2SH
+
4.3 4.4
N CC5H11
OHO
HN
ONH
O COOCH3
OHSHN
COOHO
Air25oC
4.5
Scheme 4.3 Reaction of pure iso[4]LGE2-pyrrole with N-acetylcysteine.
Representative MALDI-TOF spectra of the reaction mixture are shown in Figure 4.5.
In the presence of N-acetylcysteine, incubation of iso[4]LGE2-pyrrole at neutral pH under
air showed no pyrrole-to-pyrrole cross-linking. Thus, the thiol-containing compound
inhibited the cross-linking of iso[4]LGE2-pyrrole (Figure 4.6). Instead a series of new
ions was produced. They corresponded to pyrrole cysteine adducts and a series
derivatives produced from them by the addition of one or more atoms of oxygen (Figure
4.7). The amount of these products increased with longer incubation. A thiol-pyrrole
conjugate was detected by MALDI-TOF MS that presumably incorporates a bond
between the pyrrole ring at C-2 to the sulfur atom of the N-acetylcysteine.3-4 Thus, thiols
inhibit the pyrrole dimerization by intercepting pyrrole cation radicals generated by
electron transfer from the pyrrole to oxygen (Scheme 4.4).
130

A B
Figure 4.5 MALDI-TOF MS spectra of pure iso[4]LGE2-pyrrole reaction with N-
acetylcysteine under air. Panel A: Reaction mixture of pure iso[4]LGE2-pyrrole and N-
acetylcysteine (molar ratio = 1:10) at 25 °C for 2 days. Panel B: Reaction mixture of pure
iso[4]LGE2-pyrrole and N-acetylcysteine (molar ratio = 1:10) at 25 °C for 8 days.
131

Figure 4.6 MALDI-TOF MS of pure iso[4]LGE2-pyrrole and its reaction with N-
acetylcysteine. Trace A = pure iso[4]LGE2-pyrrole incubation under air at 25 °C for 8
days. Trace B = pure iso[4]LGE2-pyrrole reaction with N-acetylcysteine under argon at
25 °C for 8 days. Trace C = pure iso[4]LGE2-pyrrole reaction with N-acetylcysteine
under argon in the presence of sodium dithionite at 25 °C for 8 days. Trace D = pure
iso[4]LGE2-pyrrole reaction with N-acetylcysteine under air at 25 °C for 8 days.
132
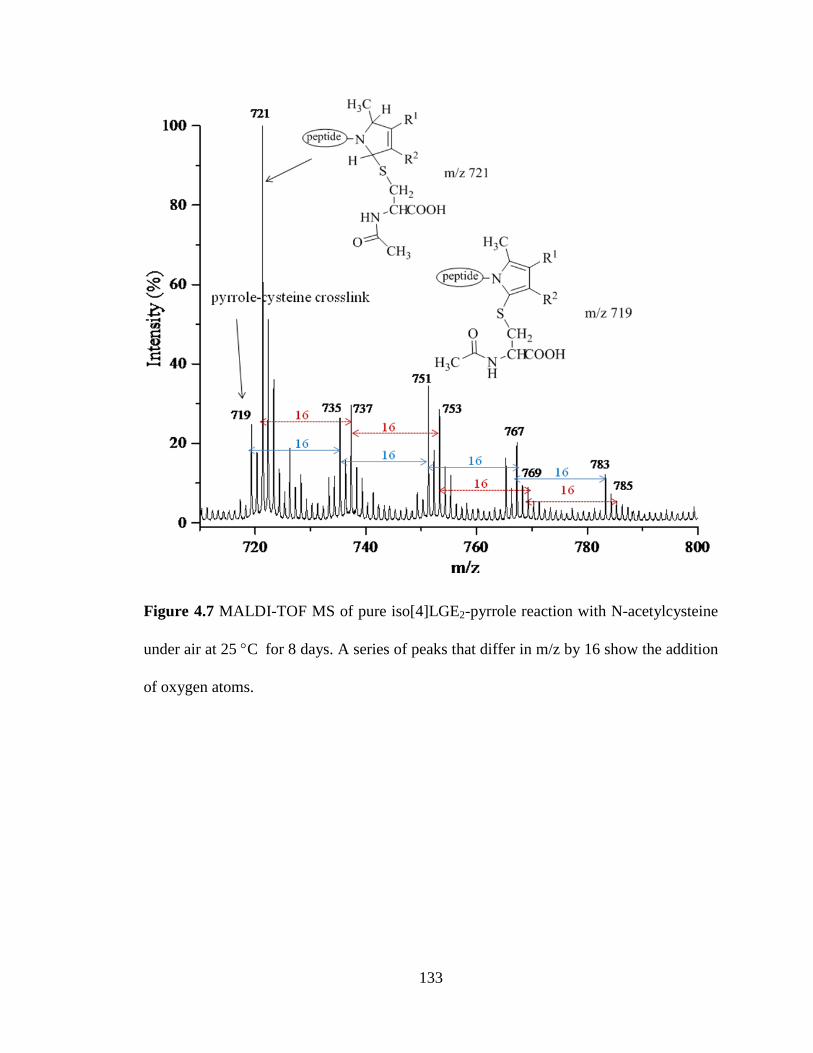
Figure 4.7 MALDI-TOF MS of pure iso[4]LGE2-pyrrole reaction with N-acetylcysteine
under air at 25 °C for 8 days. A series of peaks that differ in m/z by 16 show the addition
of oxygen atoms.
133

R1
R2
N
H3C
pyrrole
protein
R1
R2N
H3C
pyrrole-pyrrole crosslink
protein
R1
R2N
H3C
protein
O O +R1
R2
N
H3C
pyrrole
protein
O O
R1
R2N
H3C
proteinR1
R2N
H3C
protein
R1
R2
N
H3C
H
H
HO OH
pyrrole-pyrrole
HNCH
C
CH2
O
O
SH
O CH3R1
R2
N
H3C
protein
OO
HN CHCOO
CH2
S
OCH3
H
H
R1
R2
N
H3C
protein
NH
CHCOO
CH2
SO
H3C
pyrrole-cysteine crosslink
N-acetylcysteine
HO OH
protein
OO
pyrrole-cysteine
R1
R2
N
H3C
HN CHCOOH
CH2
S
OCH3
H
protein
H
HO OH
O O
m/z 721 (+H+)
m/z 719 (+H+)m/z 1149 (+H+)
Scheme 4.4 Possible electron transfer mechanism for pure iso[4]LGE2-pyrrole reaction
with N-acetylcysteine.
4.2.4 Initiation of the oxidative transformations of iso[4]LGE2-pyrrole.
Because the oxidative transformations of iso[4]LGE2-pyrrole (from which excess
dipeptide and isoLG had been removed by HPLC) exhibited a reaction time course
consistent with an autocatalytic mechanism, a search was initiated to discover possible
initiators. Pure iso[4]LGE2-pyrrole was incubated under air in the presence of various
134

potential initiators. The mass spectra of the resulting reaction product mixtures are
presented in the appendix.
(i) Stability of iso[4]LGE2-pyrrole in air in the presence of Nε-acetyl-L-lysine,
ethanolamine, L-lysine or β-amyloid peptide
Known components of the reaction mixture for the synthesis if isoLG-pyrroles were
tested first as well as analogues including Nε-acetyl-L-lysine, ethanolamine, L-lysine and
β-amyloid peptide. Pure iso[4]LGE2-pyrrole was comparably stable upon exposure to
these compounds without decomposing or forming oxidized adduct. Thus, Nε-acetyl-L-
lysine was added to pure iso[4]LGE2-pyrrole under air. After 2 days incubation, no
reaction was detected either with one equivalent of Ac-lysine or with a large excess
relative to pyrrole. Also the results were no different at pH=3 or 7. A representative mass
spectrum of the reaction product mixture is shown in appendix Figure 4.4S. Similarly,
after incubation for 2 days in the presence of 10 equivalents of L-lysine, pure
iso[4]LGE2-pyrrole underwent no reaction. Additional incubation of iso[4]LGE2-pyrrole
resulted in gradual consumption of the pyrrole, but no identifiable products were detected
(see appendix Figure 4.5S). Similarly, no reaction was detected after 2 days incubation of
pure iso[4]LGE2-pyrrole in the presence of 10 equivalents of β-amyloid peptide (see
appendix Figure 4.6S).
Incubation in the presence of ethanolamine resulted in decomposition, especially with
a large molar excess. Thus, a solution of 100 mM ethanolamine in sodium phosphate
buffer was adjusted pH to 7. Incubation of iso[4]LGE2-pyrrole under air in the presence
of one equivalent of ethanolamine resulted in gradual consumption of the pyrrole, but
some remained un-decomposed after 2 days (Figure 4.7S left). However, in the presence
135

of 10 equivalents of ethanolamine, the pyrrole decomposed completely within 2 h (Figure
4.7S right).
(ii) Stability of iso[4]LGE2-pyrrole in air in the presence of N-acetyl-gly-lys methyl
ester
Pure iso[4]LGE2-pyrrole was incubated under air in the presence of N-acetyl-gly-lys
methyl ester. Various conditions (N-acetyl-gly-lys methyl ester in H2O, pH 7; N-acetyl-
gly-lys methyl ester in phosphate buffer, pH 8.6; and N-acetyl-gly-lys methyl ester
acetate salt in H2O, pH 5.8; molar ratio from 0.1 to 10 equivalents to iso[4]LGE2-pyrrole)
were tested. The results were quite similar under all conditions. The oxidized pyrrole
product dehydrated hydroxylactam formed within 1 h and percentage increased with
continued incubation. But no aminal cross-link or bispyrrole was detected (see appendix
Figure 4.8S). These results contrasted with the lack of reaction observed upon incubation
of the pyrrole in the presence of Nε-acetyl-L-lysine, ethanolamine, L-lysine or β-amyloid
peptide. It was considered that commercial N-acetyl-gly-lys methyl ester may contain
some impurities that account for this discrepancy. To test this hypothesis, the commercial
sample of N-acetyl-gly-lys methyl ester was purified by HPLC using an
acetonitrile/water gradient. A fraction containing pure N-acetyl-gly-lys methyl ester was
isolated and confirmed with MALDI-TOF MS. Purified iso[4]LGE2-pyrrole was then
incubated under air at 25 °C for 6 days in the presence of HPLC-purified N-acetyl-gly-lys
methyl ester (Figure 4.9S). After incubation for several days, no oxidized products
formed.
(iii) The effects of vitamin C, vitamin E, H atom donor ((R)-(-)-2-(2,5-
dihydrophenyl)glycine), free radical initiator (2,2'-azobis(2-methylpropionamidine)
136

dihydrochloride), UV irradiation, rose bengal, methylene blue, Co2(CO)8,
H2O2/K2CO3, dimethyldioxirane, microwave irradiation, di-tert-butyl peroxide,
riboflavin (VB2), K3Fe(CN)6, FeCl3, K2S2O8 (persulfate) on the air stability of pure
iso[4]LGE2-pyrrole
The purified isoLG-pyrrole is unchanged after incubation in aqueous solution under
air for 2 days. In contrast, about half of the crude, unpurified pyrrole was transformed
into lactam, hydroxylactam, and bispyrrole under these same conditions. Furthermore, the
oxidative transformations of pure pyrrole exhibited a reaction time course suggestive of
autocatalysis (Figure 4.1 above). Therefore, various free radical initiators, H atom donors
and potential initiators of pyrrole oxidation were tested for their ability to trigger the
oxidation of purified iso[4]LGE2-pyrrole by air. In the presence of large amounts (0.1
equivalents) of vitamin C, vitamin E and 2,2'-azobis(2-methylpropionamidine)dihydro-
chloride, the purified pyrrole was completely consumed but no products could be
detected in the mass range of m/z 400-2000 after incubation overnight. In the presence of
smaller amounts (0.01 equivalents) of these additives, some pyrrole (about 20%)
remained un-reacted, but no products could be detected in the mass range of m/z 400-
2000 after incubation overnight (appendix Table 4.1S). Traces of metal ions are
especially likely to be present in aqueous buffers. Therefore, catalysis by traces of redox
active metal ions was investigated to test their ability to trigger the oxidation of purified
iso[4]LGE2-pyrrole by air. Fe(III) is known to promote polymerization of pyrroles
through single electron transfer (SET) that generates a pyrrole radical cation. Reoxidation
of the resulting Fe(II) by oxygen allows Fe(III) to function as a catalyst.5-7 Similar
reaction was performed by adding catalytic amount of metallic salt free radical initiators
137

to iso[4]LGE2-pyrrole. All of these potential catalysts caused rapid consumption of the
pyrrole but no traces of lactam, hydroxylactam, and bispyrrole were detected under the
reaction tested. Reaction details were summarized in appendix Table 4.1S (e).
To test the hypothesis that the product distribution from oxidative transformations of
LG/isoLG-derived pyrrole can be influenced by a hydrogen atom donor ((R)-(-)-2-(2,5-
dihydrophenyl)glycine), the pyrrole was incubated for 2 h in the presence of 5
equivalents of this hydrogen atom donor. The pyrrole was completely consumed but no
traces of lactam, hydroxylactam or bispyrrole were detected. With 1 equivalent, the
pyrrole was partially consumed in 2 h, and completely consumed after incubation over
night, but no traces of lactam, hydroxylactam or bispyrrole were detected. Incubation of
the pyrrole for 2 days in the presence of 0.2 equivalents of this hydrogen atom donor
resulted in no apparent reaction. The pyrrole was stable and no traces of lactam,
hydroxylactam or bispyrrole were detected.
4.2.5 Autoxidation of iso[4]LGE2-pyrrole promoted by APS/TEMED, TEMPO and
TMAO.
Potential catalysts that could promote pyrrole-pyrrole oxidative coupling and
oxidation to lactams and hydroxylactones were tested. Persulfate ion was reported to
promote pyrrole polymerization by SET that generates a pyrrole radical cation.6
Incubation of the isoLG-pyrrole derivative of N-acetyl-gly-lys methyl ester with 1-2
mol% ammonium persulfate (APS) and 0.5-1 mol% tetramethylethylenediamine
(TEMED) generated a product mixture containing small amounts of pyrole-pyrole dimer
138

and much greater quantities of lactam and hydroxylactam (Figure 4.8), but these products
were decomposed by treatment with more persulfate.
Since it was a free radical reaction, it was quite hard to control reaction to form one
major product. If a little more of the APS (over 5 mol% equivalent) and TEMED (over 2
mol% equivalent) were added, the pyrrole was completely consumed but no lactam,
hydroxylactam or bispyrrole were detected. The products were similar for reacting at 25
°C and 37 °C. Representative spectra are shown in appendix Figure 4.10-11S.
Figure 4.8 MALDI-TOF MS of pure iso[4]LGE2-pyrrole and its oxidation products:
lower trace = pure iso[4]LGE2-pyrrole, upper trace = after exposure to 2 mol% APS and
1 mol% TEMED at 37 °C for 2 h.
139

Nitroxide, also known as amino-N-oxide, is a functional group which contains an N-
O bond and two C-N bonds. Nitroxide radicals, especially (2,2,6,6-tetramethylpiperidin-
1-yl)oxyl, have been used7 as stable persistent free radical initiators. It is the most widely
used nitroxide radical owing to its remarkable persistence.8-10
Since tetramethylpyrrolidine-N-oxide (TEMPO) could function as a SET oxidant11,
we tested its ability to promote isoLG-pyrrole dimerization. Treatment of dimer-free
pyrrole with 10 mol % of TEMPO at 37 °C for 3 h cleanly generated a product mixture
containing similar amounts of lactam and pyrrole-pyrrole dimer, and the products were
not degraded by longer incubation or use of larger mol% of TEMPO. Thus, pyrrole-
pyrrole cross-linking of isoLG-pyrrole was also facilitated by the stable free radical
tetramethylpyrrolidine-N-oxide (TEMPO) (Figure 4.9, see next page).
TEMPO hydroxylamine (TEMPOH) is formed by one electron reduction of TEMPO.
In air, TEMPOH can be readily oxidized back to TEMPO. The applications of TEMPO in
biochemistry often relate to its reduction to TEMPOH. In such systems, reduction of
TEMPO usually involves metal ions as shown in Scheme 4.5.12
NO
+ Fe (II) NOH
+ Fe (III)
Scheme 4.5 Reduction of TEMPO by ferrous iron.
TEMPO can be reduced to TEMPOH by metal ions. To neutralize any traces of metal
ions that might be present, EDTA was added to the reaction mixture. The same
autoxidation of iso[4]LGE2-pyrrole occurred in the presence of EDTA (one equivalent
relative to TEMPO) as in its absence. Apparently metal ion catalysis is not required for
140

TEMPO to promote oxidative reactions of iso[4]LGE2-pyrrole. Represented spectra are
shown in appendix Figure 4.14-15S.
Figure 4.9 MALDI-TOF MS of pure iso[4]LGE2-pyrrole and its oxidation products:
lower trace = pure iso[4]LGE2-pyrrole, upper trace = after exposure to 10 mol% TEMPO
at 37 °C for 3 h.
Although trimethylamine-N-oxide (TMAO) is not a stable free radical analogue of
TEMPO, it has a nitrogen bound oxygen atom and can act as a prooxidant in the
autoxidation of methyl linoleate.12 TMAO can also react with Cu(I) and Cu(II)
derivatives to generate transient oxo-Cu(III) (cupryl) intermediates in oxidations such as
141

ortho hydroxylation of N-benzoyl-2-methylanaline.12-13 TMAO decomposes to aminium
radical TMA·+ then give the form of TMA (trimethylamine).14
Levels of TMAO are strongly positively correlated with cardiovascular disease risk.15
Understanding the molecular mechanisms of its ability to promote oxidative
transformations of biomolecules is likely to provide important insights into its
pathological involvements.
Remarkably, TMAO proved similarly effective to TEMPO in promoting oxidation of
the isoLG-pyrrole derivative of N-acetyl-gly-lys methyl ester to lactam and
hydroxylactam and oxidative pyrrole-pyrrole coupling. It is especially interesting that
TMAO can promote the oxidative transformations of isoLG-pyrroles. Treatment of
dimer-free pyrrole with 10 mol% of TMAO at 37 °C for 3 h cleanly generated a product
mixture containing similar amounts of lactam and pyrrole-pyrrole dimer, and the products
were not degraded by longer incubation or use of larger mol% of TMAO. All reactions
were done at 37 °C which was closed to human biological environment. We also
characterized this reaction including examining the role of trace metal ions through
adding EDTA (EDTA was equivalent to TMAO). Result indicated no direct effect to the
pyrrole initial reaction. Represented spectrums were shown in appendix Figure 4.12-13S.
It is conceivable that this chemistry contributes to the pathological consequences of
TMAO biosynthesis that involves gut microbial metabolism of phosphatidylcholines,16-17
and L-carnitine.18 Furthermore, exploration of the mechanisms of LG/isoLG pyrrole
oxidation reactions may provide insights into chemistry involved in pathologies
associated with elevated levels of TMAO.
142

R1
R2N
H3C
pyrrole
protein
R1
R2N
H3C
pyrrole-pyrrole crosslink
protein
R1
R2N
H3C
protein
O O +R1
R2N
H3C
pyrrole
protein
O O+
R1
R2N
H3C
proteinR1
R2N
H3C
protein
R1
R2N
H3C
proteinH
pyrrole
R1
R2N
H3C
protein
HL H
L
H
O OHO OH
pyrrole-pyrroleR1
R2N
H3C
protein
H
OHO H
R1
R2N
H3C
protein
HO
- H2O
R1
R2N
H3C
protein
H
Olactam
OO
NproteinR2
R1
R1
R2N
H3C
protein
OH
Ohydroxylactam
endoperoxide
O O
O
hydroperoxide
R1
R2N
H3C
protein
H
OO HL H
L
2H-pyrrol-1-ium
H
Scheme 4.6 Possible electron transfer mechanism for pyrrole oxidized reaction
The study results described above indicating the reaction of pure LG/isoLG-pyrroles
with oxygen is slow in the absence of catalysts. Nevertheless, it is likely that the first step
in the reaction of oxygen with these pyrroles generates a pyrrole cation radical-
superoxide ion pair. Hypothetical mechanistic pathways are presented in Scheme 4.6 for
the partitioning of this intermediate to generate pyrrole-pyrrole dimers, hydroxylactams
and lactams. Dimer formation involves addition of the electrophilic cation radical to the
electron rich pyrrole ring of a second molecule of pyrrole followed by further oxidation
and proton loss. Hydrogen peroxide is a likely byproduct of oxidative pyrrole-pyrrole
143

coupling. The tendency of isoLG pyrroles to undergo spontaneous oxidation may be the
result of a hydrogen peroxide promoted autocatalytic pyrrole oxidative coupling.
4.2.6 Pilot studies of bispyrrole synthesis.
Reaction conditions that promote bispyrrole formation were also probed with two
simple model systems, 3-ethyl-2,4-dimethylpyrrole and 3-ethyl-1,2,4-trimethylpyrrole.19
Pilot studies showed that both of these compounds formed dimers and trimers when
incubated under air in organic solvents. The details were discussed below and related
mass spectra of the reaction product mixtures were presented in appendix Figure 4.16-
18S.
(i) Initiate reactions of 3-ethyl-2,4-dimethylpyrrole with TEMPO and TMAO.
TEMPOor TMAO
NH
H3C CH3
CH3
HN
H3C
HN
CH3
CH3
CH3CH3
CH3
4.6 4.7
Scheme 4.7 Initiate reactions of 3-ethyl-2,4-dimethylpyrrole
Commercial available 3-ethyl-2,4-dimethylpyrrole (4.6) was resuspended in organic
solvent and tested its polymerization reactions in different conditions at 25 °C or 37 °C.
Pyrrole self-reaction in acetonitrile was tested first and result showed 3-ethyl-2,4-
dimethylpyrrole could form dimer and trimer spontaneously having no relationship with
adding TEMPO or TMAO (TEMPO or TMAO was 1:1 equivalent to pyrrole) (related
spectrums were shown in appendix Figure 4.16S). Methanol or acetone as solvents were
144

also tried and the results were quite similar. A possible way to avoid 3-ethyl-2,4-
dimethylpyrrole polemerization itself was pyrrole methylation with CH3I.
(ii) Preparation of 3-ethyl-1,2,4-trimethylpyrrole (MP)
NH
CH3I
KOH, DMSO N
4.6 4.8
Scheme 4.8 Preparation of 3-ethyl-1,2,4-trimethylpyrrole (MP)
Preparation of 3-ethyl-1,2,4-trimethylpyrrole 4.8 was carried out with the general
method Heaney and Ley developed.20 After reaction, combined organic extracts removed
solvent under reduced pressure to give pure 4.8 with silica gel column purification with
ethyl acetate: hexane=1:8 .
(iii) Initiate reactions of 3-ethyl-1,2,4-trimethylpyrrole with TEMPO and TMAO.
TEMPOor TMAO
N
H3C CH3
CH3
N
H3C
N
CH3
CH3
CH3CH3
CH3
4.8 4.9
Scheme 4.9 Initiate reactions of 3-ethyl-1,2,4-trimethylpyrrole with TEMPO and TMAO
3-Ethyl-1,2,4-trimethylpyrrole (4.8) was incubated under air at 25 °C in the presence
or absence of TEMPO (1 equivalent) in acetonitrile solution. 3-Ethyl-1,2,4-
trimethylpyrrole underwent dimerization and trimerization under both conditions (spectra
are shown in appendix Figure 4.18S), i.e, TEMPO had no significant effect on increasing
145

dimer formation. The relatively greater ease of autoxidation and pyrrole-pyrrole cross-
linking of pyrrole 4.8 compared to iso[4]LGE2-pyrrole could simply be a consequence of
the greater concentration of oxygen in organic versus aqueous medium. The
concentration O2 in air equilibrated H2O (0.28 mM) is about 7 fold lower than in air
equilibrated CHCl3 (2.05 mM), acetone (2.4 mM), acetonitrile (2.42 mM), or ethanol
(1.94 mM).21-27 Because the autoxidation of isoLG pyrrole was performed in the presence
of a lower concentration of O2 than autoxidation of 4.8, initiators, e.g., TEMPO or
TMAO exerted a greater influence on the autoxidation. Thus, autoxidation of isoLG
pyrrole occurs less readily than oxidation of 3-ethyl-1,2,4-trimethylpyrrole in air
saturated organic solvents in which the concentration of O2 is much higher than in water.
146

4.3 Conclusions
We discovered that the pure LG/isoLG-pyrrole derivative of the N-acetyl-lys-gly
methyl ester dipeptide can be produced in the presence of dithionite, an oxygen scavenger.
The oxidation and dimerization of the pure pyrrole is promoted by oxygen and exhibits
an induction period that can be eliminated by various catalysts, e.g., the stable free radical
TEMPO as well as by TMAO. More vigorous oxidative decomposition was promoted by
vitamin C, vitamin E and 2,2'-azobis(2-methylpropionamidine)dihydrochloride.
Previously, the reaction of phosphatidylethanolamine (PE) with an isoLG was reported to
produce a stable pyrrole dervative that resists further oxidation to lactams.28 This stability
was ascribed to lower concentrations of O2 in organic solvents such as CHCl3 (in which
the reaction of PE with isoLGs was conducted) than in H2O (the solvent used for the
reaction of proteins and the N-acetyl-lys-gly methyl ester dipeptide with isoLGs).
However, the opposite is the case, the concentration of oxygen in CHCl3 is seven times
higher than in air saturated H2O. Given the susceptibility of the autoxidation of the N-
acetyl-lys-gly methyl ester dipeptide-derived isoLG pyrrole to catalysis, the stability of
PE-isoLG pyrrole may be a consequence of its purity, i.e., the absence of catalytic
impurities that promote its autoxidation.
In the presence of N-acetylcysteine, incubation of iso[4]LGE2-pyrrole at neutral pH
under air showed no pyrrole-to-pyrrole cross-linking. Instead a series of new ions was
produced corresponding to pyrrole cysteine adducts and a series derivatives produced
from them by the addition of one or more atoms of oxygen. It is suggested that the thiol
inhibits the pyrrole dimerization by intercepting pyrrole cation radical generated by
electron transfer to oxygen.
147

4.4 Experimental Procedures
4.4.1 General methods. All chemicals used were high purity analytical grade. The
following commercially available materials were used as received: acetyl-gly-lys-O-
methyl ester was from Bachem (Torrance, CA), 3-ethyl-2,4-dimethylpyrrole was from
Aldrich (Milwaukee, WI), all other chemicals were obtained from Fisher Scientific Co
(Chicago, IL). Iso[4]LGE2 was prepared by Dr. Jim Laird. Chromatography was
performed with ACS grade solvents.
4.4.2 Iso[4]LGE2-pyrrole. N-Acetyl-gly-lys methyl ester (2 mg, 7.71 µmol) was added
to 1 ml ddH2O and the solution was sparged with bubbled argon for several minutes to
remove air. Then Na2S2O4 (1 mg, 5.75 µmol) was added. Iso[4]LGE2 in methanol (136
µg, 0.386 µmol, 5 µg/µl) was added and the mixture was incubated overnight at room
temperature. TLC showed the presence of unreacted dipeptide, only one major product
and complete disappearance of iso[4]LGE2. The solution was concentrated under high
vacuum to remove water into a Dry Ice-acetone cooled trap, and then methanol (1 mL)
was added to dissolve the organic product. Then the mixture was loaded onto a 6 cm
column of silica gel packed with a slurry in EtOAc in a disposable pipette, and eluted
with methanol (2 mL) (Rf = 0.5) to remove most of sodium dithionite. After concentration
under reduced pressure, reagent grade chloroform (1 mL) that had been sparged with
argon was added to dissolve iso[4]LGE2-pyrrole and the solution was filtered to remove
sodium dithionite and any silical gel that had dissolved in the methanol. The crude
iso[4]LGE2-pyrrole (~222 µg, 0.39 µmol) solution was concentrated by evaporation with
argon gas and re-suspended in 500 µl 10% acetonitrile/water.
148

Iso[4]LGE2-pyrrole was purified by HPLC with a 2.1 x 100 mm i.d. C18 1.8 µm
column (Waters Acquity UPLC HSS). Chromatography was carried out with linear
elution gradient (elute A, 0.1% FA/H2O; elute B, 0.1% FA/ACN) at a flow rate of 150
µl/min. For determining the retention time of iso[4]LGE2-pyrrole (~222 µg, 0.39 µmol)
that had been freed of Na2S2O4 was dissolved in 10% acetonitrile (ACN) 0.1% formic
acid (FA) (50 µl) and then loaded to the column with 20% B for 10 min, and eluted with
the following gradient: 20-60% B over 20 min, to 98 % within 0.1 min and hold for 5 min.
The effluent was monitored by ESI-MS/MS with an API-3000 triple quadrupole
electrospray mass spectrometer (Applied Biosystems Inc.) The instrument was operated
in the positive mode and high-pressure nitrogen was used as source gas and scanning
from m/z 220-2000 using the parameters listed in Table 3.2. The pyrrole eluted between
12.8 and 14 minutes (Figure 4.2). Another aliquot of ~222 mg was injected and the eluent
was collected in 1 minute fractions from 11 minutes to 31 minutes and the fractions were
analyzed for purity by MALDI-TOF MS. Fractions containing the pure pyrrole were
combined. Optimized parameters for triple quadrupole mass spectrometer were noted in
section 3.4.4.
4.4.3 Autoxidation of crude iso[4]LGE2-pyrrole. Crude iso[4]LGE2-pyrrole that
contained excess dipeptide (~222 µg, 0.39 µmol in 500 µl 10% acetonitrile/water) was
incubated under air 1h on a shaker (IKA MTS 2/4 digital microtiter shaker, 500 rpm).
Then 100 µl of the reaction mixture (0.077 µmol) was quickly dried using a high-speed
vacuum evaporator and analyzed by MALDI-TOF MS. The remaining reaction mixture
149

(0.31 µmol) was aerated with oxygen overnight on a shaker (IKA MTS 2/4 digital
microtiter shaker, 500 rpm) at room temperature, and then analyzed by MALDI-TOF MS.
4.4.4 Autoxidation of iso[4]LGE2-pyrrole. Purified iso[4]LGE2-pyrrole (44 µg, 0.077
µmol in 450 µl 10% acetonitrile/water) was incubated under air at 25 °C on shaker (IKA
MTS 2/4 digital microtiter shaker, 500 rpm). At different time point (2, 6, and 8 days),
same amount of reaction mixture (15 µg, 0.026 µmol in 150 µl 10% acetonitrile/water)
was quickly dried using a high-speed vacuum evaporator and analyzed MALDI-TOF MS.
4.4.5 Autoxidation of iso[4]LGE2-pyrrole in the presence of Nα-acetylcysteine. 7.7 µl
Nα-acetylcysteine (0.77 µmol, 126 µg) (100 mM in H2O, pH 3) was added to purified
iso[4]LGE2-pyrrole (44.36 µg, 0.077 µmol in 500 µl 10% acetonitrile/water) and reacted
under air at 25 °C on shaker (IKA MTS 2/4 digital microtiter shaker, 500 rpm). At
different time point (2, 4, 6 and 8 days), same amount of reaction mixture (0.019 µmol,
125 µl) was quickly dried using a high-speed vacuum evaporator and analyzed by
MALDI-TOF MS.
Control experiments were performed. 7.7 µl Nα-acetylcysteine (0.77 µmol, 126 µg)
(100 mM in H2O, pH 3) was added to purified iso[4]LGE2-pyrrole (44.36 µg, 0.077 µmol
in 500 µl 10% acetonitrile/water) and reacted under argon gas protection at 25 °C on
shaker (IKA MTS 2/4 digital microtiter shaker, 500 rpm). At different time point (2, 4, 6
and 8 days), same amount of reaction mixture (0.019 µmol, 125 µl) was quickly dried
using a high-speed vacuum evaporator and analyzed by MALDI-TOF MS.
150

7.7 µl Nα-acetylcysteine (0.77 µmol, 126 µg) (100 mM in H2O, pH 3) was added to
purified iso[4]LGE2-pyrrole (44.36 µg, 0.077 µmol in 500 µl 10% acetonitrile/water) and
reacted under argon in the presence of dithionite at 25 °C on shaker (IKA MTS 2/4
digital microtiter shaker, 500 rpm). At different time point (2, 4, 6 and 8 days), same
amount of reaction mixture (0.019 µmol, 125 µl) was quickly dried using a high-speed
vacuum evaporator and analyzed by MALDI-TOF MS.
4.4.6 Autoxidation of iso[4]LGE2-pyrrole in the presence of acetyl-gly-lys methyl
ester. 7.7 µl N-acetyl-gly-lys methyl ester (0.77 µmol, 200 µg) (100 mM in H2O, pH 7)
was added to purified iso[4]LGE2-pyrrole (44 µg, 0.077 µmol in 500 µl 10%
acetonitrile/water) and reacted under air on shaker (IKA MTS 2/4 digital microtiter
shaker, 500 rpm). At different time point (1 h, 8 h, 1 and 2 days), same amount of
reaction mixture (0.019 µmol, 125 µl) was quickly dried using a high-speed vacuum
evaporator and analyzed by MALDI-TOF MS.
N-Acetyl-gly-lys methyl ester was purified by HPLC on a 2.1 x 100 mm i.d. C18 1.8
µm column (Waters Acquity UPLC HSS). N-Acetyl-gly-lys methyl ester (10 µl, 10
µmol, 259 µg) (100 mM in H2O, pH 7) was dissolved in 10 µl 2% ACN 0.1% FA. This
solution (18 µl) was injected onto the column and eluted with a linear gradient (elute A,
0.1% FA/H2O; elute B, 0.1% FA/ACN) at a flow rate of 100 µl/min. The gradient was 2-
8% B over 8 min. Fractions were collected every minute and analyzed for dipeptide by
MALDI-TOF MS. The fraction eluting during the 4th min contained purified N-acetyl-
gly-lys methyl ester. It was quickly dried using a high-speed vacuum evaporator and re-
suspended in 10% acetonitrile/water.
151

A mixture of purified N-acetyl-gly-lys methyl ester (0.77 µmol, 200 µg) and purified
iso[4]LGE2-pyrrole (44 µg, 0.077 µmol in 500 µl 10% acetonitrile/water) was incubated
under air at 25 °C for several days on shaker (IKA MTS 2/4 digital microtiter shaker, 500
rpm). At various time points (2 h, 6 h, 2 and 6 days) aliquots of the reaction mixture
(0.019 µmol, 125 µl) were quickly dried using a high-speed vacuum evaporator and
analyzed by MALDI-TOF MS.
4.4.7 Autoxidation of iso[4]LGE2-pyrrole in the presence of free radical initiators or
hydrogen atom donors. Vitamin C (7.7 µl of 1 mM in pH 7 H2O, 7.72 nmol, 1.36 µg)
was added to the purified iso[4]LGE2-pyrrole (44.36 µg, 0.077 µmol in 450 µl 10%
acetonitrile/water) and the mixture was incubated under air on shaker (IKA MTS 2/4
digital microtiter shaker, 500 rpm). At various time points (2 h, 24 h, 7 days), aliquots of
the reaction mixture (150 µl) were quickly dried using a high-speed vacuum evaporator
and analyzed by MALDI-TOF MS. Similar incubations were performed with 2,2'-
azobis(2-methylpropionamidine) dihydrochloride (1 mM in H2O, pH 6) or vitamin E (5
mM in EtOH, pH 6) in place of vitamin C.
(R)-(-)-2-(2,5-Dihydrophenyl)glycine (7.7 µl of 10 mM in 200 mM (pH 2) HCl,
0.077 µmol, 12 µg) was added to purified iso[4]LGE2-pyrrole (44 µg, 0.077 µmol in 450
µl 10% acetonitrile/water) and the mixture was incubated under air on shaker (IKA MTS
2/4 digital microtiter shaker, 500 rpm). At various time points (2 h, 24 h, 7 days), aliquots
of the reaction mixture (150 µl) were quickly dried using a high-speed vacuum
evaporator and analyzed by MALDI-TOF MS.
152

4.4.8 Autoxidation of iso[4]LGE2-pyrrole in the presence of APS/TEMED, TMAO
or TEMPO. Trimethylamine-N-oxide (TMAO, 7.7 µl of 1 mM in pH 7 H2O, 7.7 nmol,
5.8 µg) was added to purified iso[4]LGE2-pyrrole (44 µg, 0.077 µmol in 500 µl 10%
acetonitrile/water) and mixture was incubated under air on a shaker (IKA MTS 2/4 digital
microtiter shaker, 500 rpm). At various time points (1 h, 2 h, 3 h and 1 day), aliquots of
the reaction mixture (0.019 µmol, 125 µl) were quickly dried using a high-speed vacuum
evaporator and analyzed by MALDI-TOF MS. The same experiment was performed with
(2,2,6,6-tetramethylpiperidin-1-yl)oxyl (TEMPO, 1 mM in 10% ACN, pH 6) in place of
TMAO.
Ammonium persulfate (APS, 1.6 µl of 1 mM in H2O, 1.544 nmol, 352.4 ng) and
tetramethylethylenediamine (TEMED, 0.7 µl of 1 mM in H2O, 0.772 nmol, 90 ng) were
added to iso[4]LGE2-pyrrole (44 µg, 0.077 µmol in 500 µl 10% acetonitrile/water) and
incubated under air on a shaker (IKA MTS 2/4 digital microtiter shaker, 500 rpm). At
various time points (1 h, 2 h, 3 h and 1 day) an aliquot of the reaction mixture (0.019
µmol, 125 µl) was quickly dried using a high-speed vacuum evaporator and analyzed by
MALDI-TOF MS.
4.4.9 3-Ethyl-1,2,4-trimethylpyrrole (4.8). Anhydrous DMSO (20 ml) was added to
crushed KOH pellets (2.24 g, 0.04 mol) and the mixture was stirred for 5 min. 2,4-
dimethyl-3-ethylpyrrole (4.6, 1.23 g, 0.01 mol) was added and the mixture was stirred for
45 min. The mixture was cooled in ice and CH3I (2.84 g, 0.02 mol) was added dropwise
with stirring and the stirring was continued for 45 min at room temperature. H2O (20 ml)
was added and the mixture was extracted with Et2O (3 × 50 ml). The combined organic
153

extracts were washed with H2O (3 × 25 ml), dried (MgSO4), and the solvent was removed
under reduced pressure. The residue was purified by flash chromatography (ethyl acetate:
hexane = 1:8, Rf = 0.8, yield 50%) to give 4.8 (0.68 g).29
TEMPO (10 µl of 100 mM in acetonitrile, 1 µmol, 156 µg) was added to 10 µl 3-
ethyl-1,2,4-trimethylpyrrole (1 µmol, 137 µg) (100 mM in acetonitrile) and the mixture
was incubated under air on a shaker (IKA MTS 2/4 digital microtiter shaker, 500 rpm) for
3 h. The reaction mixture was quickly dried using a high-speed vacuum evaporator and
analyzed by MALDI-TOF MS.
4.4.10 Analysis of adducts using MALDI-TOF MS. Initial diagnostic analysis of the
samples was performed by MALDI-TOF mass spectrometry (as described in section
3.4.3).
154

4.5 References
1. Salomon, R. G. (2005) Levuglandins and isolevuglandins: Stealthy toxins of
oxidative injury. Antioxid Redox Signal 7 (1-2), 185-201.
2. Englander, S. W.; Calhoun, D. B.; Englander, J. J. (1987) Biochemistry without
oxygen. Anal Biochem 161 (2), 300-306.
3. Zhu, M.; Spink, D. C.; Yan, B.; Bank, S.; DeCaprio, A. P. (1995) Inhibition of
2,5-hexanedione-induced protein cross-linking by biological thiols: chemical mechanisms
and toxicological implications. Chem Res Toxicol 8 (5), 764-771.
4. Zhu, M.; Spink, D. C.; Yan, B.; Bank, S.; DeCaprio, A. P. (1994) Formation and
structure of cross-linking and monomeric pyrrole autoxidation products in 2,5-
hexanedione-treated amino acids, peptides, and protein. Chem Res Toxicol 7 (4), 551-558.
5. Liao, Y.; Wang, X.; Qian, W.; Li, Y.; Li, X.; Yu, D. G. (2012) Bulk synthesis,
optimization, and characterization of highly dispersible polypyrrole nanoparticles toward
protein separation using nanocomposite membranes. J Colloid Interface Sci 386 (1), 148-
157.
6. Mezhuev, Y. O.; Korshak, Y. V.; Shtilman, M. I.; Piskareva, A. I. (2012)
Activation parameters of single-electron transfer from pyrrole molecule to persulfate ion.
(English). Izv V.U.Z. Khim Khim Tekhnol 55 (10), 42-45.
7. Shi, H. C.; Li, Y. (2007) Formation of nitroxide radicals from secondary amines
and peracids: A peroxyl radical oxidation pathway derived from electron spin resonance
detection and density functional theory calculation. J Mol Catal A Chem 271 (1–2), 32-41.
8. Barriga, S. (2001) 2,2,6,6-Tetramethylpiperidin-1-oxyl (TEMPO). Synlett 2001
(04), 0563.
155

9. Lebedev, O. L.; Kazarnovskii, S. N. (1960) Catalytic oxidation of aliphatic
amines with hydrogen peroxide. Zh Obshch Khim 30, 1631-1635.
10. Novak, I.; Harrison, L. J.; Kovac, B.; Pratt, L. M. (2004) Electronic structure of
persistent radicals: nitroxides. J Org Chem 69 (22), 7628-7634.
11. Samuni, A.; Goldstein, S.; Russo, A.; Mitchell, J. B.; Krishna, M. C.; Neta, P.
(2002) Kinetics and mechanism of hydroxyl radical and OH-adduct radical reactions with
nitroxides and with their hydroxylamines. J Am Chem Soc 124 (29), 8719-8724.
12. Ishikawa, Y.; Yuki, E. (1974) Effect of trimethylamine oxide on the oxidation of
natural tocopherol mixtures. Yukagaku 23, 710-713.
13. Reinaud, O.; Capdevielle, P.; Maumy, M. (1991) ortho-Aryloxylation of N-
substituted benzamides: a new oxidizing process induced by the
copper(II)/trimethylamine N-oxide system. J Chem Soc Perkin Trans I (9), 2129-2134.
14. Reinaud, O.; Capdevielle, P.; Maumy, M. (1990) Copper(II) mediated aromatic
hydroxylation by trimethylamine N-oxide. J Chem Soc Chem Commun (7), 566-568.
15. Bennett, B. J.; de Aguiar Vallim, T. Q.; Wang, Z.; Shih, D. M.; Meng, Y.;
Gregory, J.; Allayee, H.; Lee, R.; Graham, M.; Crooke, R.; Edwards, P. A.; Hazen, S. L.;
Lusis, A. J. (2013) Trimethylamine-N-oxide, a metabolite associated with atherosclerosis,
exhibits complex genetic and dietary regulation. Cell Metab 17 (1), 49-60.
16. Wang, Z.; Klipfell, E.; Bennett, B. J.; Koeth, R.; Levison, B. S.; Dugar, B.;
Feldstein, A. E.; Britt, E. B.; Fu, X.; Chung, Y. M.; Wu, Y.; Schauer, P.; Smith, J. D.;
Allayee, H.; Tang, W. H.; DiDonato, J. A.; Lusis, A. J.; Hazen, S. L. (2011) Gut flora
metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 472 (7341),
57-63.
156

17. Tang, W. H.; Wang, Z.; Levison, B. S.; Koeth, R. A.; Britt, E. B.; Fu, X.; Wu, Y.;
Hazen, S. L. (2013) Intestinal microbial metabolism of phosphatidylcholine and
cardiovascular risk. N Engl J Med 368 (17), 1575-1584.
18. Koeth, R. A.; Wang, Z.; Levison, B. S.; Buffa, J. A.; Org, E.; Sheehy, B. T.; Britt,
E. B.; Fu, X.; Wu, Y.; Li, L.; Smith, J. D.; DiDonato, J. A.; Chen, J.; Li, H.; Wu, G. D.;
Lewis, J. D.; Warrier, M.; Brown, J. M.; Krauss, R. M.; Tang, W. H.; Bushman, F. D.;
Lusis, A. J.; Hazen, S. L. (2013) Intestinal microbiota metabolism of L-carnitine, a
nutrient in red meat, promotes atherosclerosis. Nat Med 19 (5), 576-585.
19. Hansen, G. H.; Henriksen, R. M.; Kamounah, F. S.; Lund, T.; Hammerich, O.
(2005) Oxidative coupling and polymerization of pyrroles: Part I. The electrochemical
oxidation of 2,4-dimethyl-3-ethylpyrrole in acetonitrile. Electrochim Acta 50 (25–26),
4936-4955.
20. Heaney, H.; Ley, S. V. (1974) 1-Benzylindole. Org Synth 54, 58.
21. Franco, C.; Olmsted, J., 3rd (1990) Photochemical determination of the solubility
of oxygen in various media. Talanta 37 (9), 905-909.
22. Battino, R., Oxygen and ozone. Pergamon: Oxford; New York, 1981; Vol. 7.
23. Murov, S. L., Handbook of Photochemistry. Dekker: New York, 1973; p. 89. .
24. Achord, J. M.; Hussey, C. L. (1980) Determination of dissolved oxygen in
nonaqueous electrochemical solvents. Anal Biochem 52 (3), 601-602.
25. Tokunaga, J. (1975) Solubilities of oxygen, nitrogen, and carbon dioxide in
aqueous alcohol solutions. J Chem Eng Data 20 (1), 41-46.
26. Monroe, B. M. (1982) Photochemical estimation of oxygen solubility. Photochem
Photobiol 35 (6), 863-865.
157

27. Evans, D. H.; Claiborne, J. B., The Physiology of Fishes. CRC Press: 2006; p. 88.
28. Bernoud-Hubac, N.; Fay, L. B.; Armarnath, V.; Guichardant, M.; Bacot, S.;
Davies, S. S.; Roberts, L. J., 2nd; Lagarde, M. (2004) Covalent binding of isoketals to
ethanolamine phospholipids. Free Radic Biol Med 37 (10), 1604-1611.
29. Hunter, G. A.; McNab, H.; Withell, K. (2010) The Meldrum’s Acid Route to
Prodigiosin Analogues. Synthesis (10), 1707-1711.
158

CHAPTER 5
GENERATION OF AN ANTI-ISO[4]LGE2 MONOCLONAL ANTIBODY

5.1 Background
IsoLGs are generated through free radical-induced oxidation of arachidonates (see
chapter 1). They are extraordinarily reactive; forming covalent adducts incorporating
protein lysyl ε-amino groups within seconds. Since proteins so efficiently trap them, also
many plasma proteins have half-lives of several weeks, and modified proteins and
protein-protein cross-links are resistant to proteolysis,1 isoLGE2 adducted plasma proteins
are expected to accumulate in plasma which could provide a unique marker and effective
dosimeter for the oxidative injuries in the brain or retina.
Development and validation of isolevuglandin (isoLG) biomarkers of oxidative stress
could be used to assess the antioxidant effects of dietary supplements in vivo and to
examine their mechanisms of action, efficacy and effectiveness with respect to human
health. However, our understanding of these processes remains primitive, in part because
of the lack of sensitive and specific tools for detecting cumulative damage resulting from
oxidative stress. An enzyme-linked immunoassay (ELISA) with polyclonal antibodies
against isoLG-protein adducts reveals disease-related elevations in these biomarkers in
blood from patients with end-stage renal disease, cardiovascular disease, age-related
macular degeneration, and children born prematurely, as well as in brain tissue from
individuals with autism spectrum disease and liver from a murine model of alcoholic
liver disease.2 Therefore, detection of blood plasma isoLGE2 may provide a convenient,
minimally invasive method for monitoring such pathologies.
Previously, we generated anti-iso[4]LGE2 and anti-LGE2 polyclonal antibodies (pAb)
that have been useful for ELISA or immunohistochemistry for detection of iso[4]LGE2-
and LGE2-protein adducts.3 These antibodies are structurally specific showing low cross
160

reactivity. They allow the confirmation that a variety of structural LG isomers are
produced in vivo. Specially, isoLGE2-KLH pAb bind isoLGE2-human serum albumin
(HSA) 200 times more strongly than they bind iso[4]LGE2-HSA, while the iso[4]LGE2-
KLH pAb bind iso[4]LGE2-human serum albumin (HSA) at least 2000 times more
strongly than they bind LGE2-HSA.4
However, pAbs have shortcomings that complicate standardization and accuracy.
They vary from batch to batch, resulting in variable responses, background can be high
and amounts are limited by the life-time of the animal source. In contrast, monoclonal
antibodies can be generated in unlimited quantities, and their specificity ensures that only
one epitope is recognized on an antigen. This is extremely useful when observing subtle
protein alterations, and the antibodies can be selected to exhibit low background cross
reaction, e.g., with abundant blood proteins such as serum albumin. Therefore, the
generation of a highly specific monoclonal anti-iso[4]LGE2 antibody was undertaken.
161

5.2 Results and discussion
5.2.1. Antibody production
Monoclonal anti-iso[4]LGE2 antibody was produced at the Hybridoma Cole Facility
of Visual Sciences Research Center (VSRC), Case Western Reserve University. Five
Balb/c mice were immunized with iso[4]LGE2-KLH. After 60 days, blood was draw from
each mouse, and tested for immune response. High anti-iso[4]LGE2 responses were
present in all 5 mice. Blood from mouse #972 showed high anti-iso[4]LGE2 response and
the lowest cross-reactivity to LGE2-BSA (Figure 5.1). Competitive antibody binding
inhibition studies with serum from all mice confirmed this conclusion (appendix Figure
5.1S). Therefore, mouse #972 was chosen for monoclonal antibody production.
Figure 5.1 Mouse serum immune responses (1:10,000 dilutions) comparison with pre-
immune serum.
Splenocytes from mouse 972 were fused with myeloma cells and clone screening was
done by both indirect and competitive ELISAs. Single cell clones from each round of
162

subcloning that produced the anti-iso[4]LGE2 monoclonal antibody were identified, and
the clone exhibiting the strongest apparent immune-reactivity was grown in larger
amounts. Anti-iso[4]LGE2 mAb from this clone was purified as described below using
protein G column and the protein concentration of the purified mAbs was determined at
OD 280 nm.
From fusion attempted on 1920 samples (wells on 20 plates) 725 wells were found to
contain hybridoma cells (appendix Figure 5.2S). The antigens iso[4]LGE2-BSA and KLH
were used for the first screening tests. Of 371 large and very large clones (clones with
many cells), three showed strong response (clones number 9G8, 12G4, 18G7) against
iso[4]LGE2-BSA, and two others showed lower response (clones number 2E8, 15A5). Of
354 small and medium clones (clones with less cells), six (clones number 4G5, 5B7, 8B3,
10C11, 16E4, 19A4) showed lower response against iso[4]LGE2-BSA compared to the
above five clones from large clones (9G8, 12G4, 18G7, 2E8, 15A5). More antigens
(iso[4]LGE2-BSA, iso[4]LGE2-HSA, BSA, HSA, LGE2-BSA, LGE2-KLH, CEP-HSA)
were tested by indirect and competitive ELISAs of the supernatant of each positive clone
(appendix Figure 5.3S). Clones 9G8, 12G4, 18G7 showed the best responses. Clones 9G8
and 12G4 were chosen for the first round of subcloning. The isotype of these was
determined. Clone 9G8 was IgM and 12G4 was IgG1.
After the first screening, the chosen clones were selected by another two rounds of
subcloning to test clone cells’ productivity and consistency, and to confirm that the clone
was derived from a single cell. Since the isotype of 9G8 was IgM, we mainly focused on
clone 12G4 that was IgG1. Clone 12G4 had 34 first-round subclones and all 34 clones
showed strong response. It showed strong specificity to iso[4]LGE2-BSA and
163

iso[4]LGE2-HSA and almost no response to LGE2-BSA (appendix Figure 5.4S). To
determine an optimal clone from 12G4 for the next round of subcloning, 10 clones were
chosen from the 34 (first subclones) for test again. Both the original antibody supernatant
concentration and a 10 fold dilution were tested. Both showed strong binding ability.
With more cell supernatant, competitive ELISA of these ten 12G4 clones was tested,
12G4 F3 was chosen for next round subcloning (appendix Figure 5.5S).
The supernatant and a 10 fold dilution from each of 16 secondary subclones of 12G4
F3 was tested by indirect ELISA (appendix Figure 5.6S). The final three candidates from
12G4 F3--C1 D1 G4 were chosen and fully tested by competitive ELISA (appendix
Figure 5.7S). Clone 12G4 F3 D1 was selected for adapting to serum free medium
(appendix Figure 5.8S).
Anti-iso[4]LGE2 mAb in serum free medium was diluted with binding buffer (20 mM
sodium phosphate buffer, pH 7) and loaded onto a protein-G column using commercially
available protein-G beads. Non-IgG proteins were washed away by eluting the column
with neutral binding buffer (20 mM sodium phosphate buffer, pH 7) and the
decomplexation is accomplished by changing the pH of the eluting buffer (glycine-HCl,
pH 2.6). The purified mAb solution was neutralized and dialyzed immediately against 10
mM PBS (pH 7.4) and then lyophilized and stored at -80 ℃. No non-specific binding was
observed in the purified mAb sample analyzed by sodium dodecyl sulfate polyacrylamide
gel electrophoresis (SDS-PAGE) as shown in Figure 5.2. Purification of IgG was
confirmed since only signal bands (heavy and light chain) were detected in SDS-PAGE.
164

Heavy Chain
Light Chain
kD
Figure 5.2 SDS-PAGE analysis of purified anti-iso[4]LGE2 mAb with protein G column.
5.2.2. Specificity of anti-iso[4]LGE2 mAb.
To assess the structural specificity and selectivity of the monoclonal antibody,
competitive inhibition of antibody binding to iso[4]LGE2-modified proteins by various
haptens was examined. For all cross-reactivity studies, iso[4]LGE2-BSA was used as
coating agent and iso[4]LGE2-HSA was used as a standard. The concentration at 50%
inhibition (IC50) for iso[4]LGE2-HSA was defined as 100% cross-reactivity. Duplicates
of serial dilutions of all inhibitors were used and final curves were constructed using
mean absorbance values (Figure 5.3).
165

Figure 5.3 Binding inhibition of protein G purified anti-iso[4]LGE2 mAb from serum
free medium by iso[4]LGE2-HSA, iso[4]LGE2-KLH, LGE2-BSA, LGE2-KLH, CEP-
BSA, CEP-HSA, BSA and HSA.
The IC50 values refer to the concentration of antigen that is required to inhibit 50%
binding of the antibody to the coating agent. The IC50 for iso[4]LGE2-HSA and
iso[4]LGE2-KLH using iso[4]LGE2-BSA as coating agent are 88.2 µM and 106.6 µM,
respectively. For a variety lysyl-ε-amino pyrrole protein modifications that have a
carboxylic acid group and are derived from lipid oxidation products (LGE2-BSA, LGE2-
KLH and CEP-BSA, CEP-HSA), anti-iso[4]LGE2 mAb showed remarkable selectivity.
All of them showed no significant cross-reactivity. Equally important, BSA and HSA
were not recognized at all by this antibody.
166

5.2.3. Western analysis of anti-iso[4]LGE2 mAb.
The anti-iso[4]LGE2 mAb which exhibited high specificity towards the iso[4]LGE2
epitope, allow the development of immunochemical assays for the detection and
quantification of iso[4]LGE2 adducts generated under oxidative stress. It was of special
importance to employ these methods for diagnosing or screening for diseases in which
elevated levels of iso[4]LGE2 adducts were present at the disease site, or at remote sites,
as in the blood.
We did pilot western blot analysis against iso[4]LGE2-BSA, a standard protein
containing the iso[4]LGE2 modification plus BSA control. Different sample loading
amounts and primary concentrations were tested (Figure 5.4). Anti-iso[4]LGE2 mAb
showed both high selectivity and sensitivity.
Marker 10µg 5µg 1µg 0.1µg10ngIso[4]LGE2-BSA
5 µgBSA
1Ab: 10 µg/ml in PBS2Ab: 1:2000
Marker 5µg 1µg 0.1µg 10 ng
1Ab: 5 µg/ml in PBS2Ab: 1:2000
170130
95
72
55
43
34
26
17
Iso[4]LGE2-BSA
Marker5µg1µg0.1µg10 ng
1Ab: 1 µg/ml in PBS2Ab: 1:2000
Iso[4]LGE2-BSA
Figure 5.4 Comparative Western analyses using anti-iso[4]LGE2 mAb against
iso[4]LGE2-BSA, a standard protein containing the iso[4]LGE2 modification. Left panel:
developed with anti-iso[4]LGE2 mAb concentration as 10 µg/ml in PBS. Middle panel:
developed with anti-iso[4]LGE2 mAb concentration as 5 µg/ml. Right panel: developed
167

with anti-iso[4]LGE2 mAb concentration as 1 µg/ml. Goat anti-mouse secondary
antibody was diluted 2000 times for all panel treatments.
A single-chain antibody from a phage displayed recombinant ScFv library that bound
a model peptide adducted with synthetic isoLGE2 was developed by Roberts et al.5
Recognition of isoLGE2–lysyl adducts isolated from oxidized arachidonic acid by this
anti-isoLGE2 adduct single-chain antibody was independent of the amino acid sequence
of the adducted peptides or proteins, and no cross-reactivity was detected with 4-
hydroxynonenal or 4-oxononanal adducts. But no test was performed to determine its
cross-reactivity with iso[n]LGE2-deived protein epitopes.5
The EC50 for oxidized arachidonyl–lysine adduct and isoLGE2-lysine adduct were
470 and 310 nM, respectively. HNE- or ONA-adducted peptides, and nonreactive F2-
isoprostane product, 15-F2t-IsoP (8-iso-PGF2a), showed no binding at peptide
concentrations up to 5 µM. These results indicated that single-chain antibody would
specifically recognize isoLGE2-lysine adducts, but not those formed by other pyrrole-
forming lipid aldehydes formed during free radical-mediated lipid peroxidation.5
Importantly, the binding of this antibody with iso[n]LGs was not determined.
Iso[4]LGE2 is an isomer formed exclusively through the free radical oxidation of AA-
PC, unlike isoLGE2 which can also be produced through the COX pathway.6 The anti-
iso[4]LGE2 mAb produced in this thesis research allows unambiguous assessment of the
formation of iso-LGs from the isoprostane pathway.
168

5.3 Conclusions
An anti-iso[4]LGE2 mAb was prepared that exhibits high sensitivity and structural
specificity for detecting iso[4]LGE2–protein adduct epitope, and no significant cross-
reactivity with analogs including carboxyethylpyrrole (CEP) or LGE2-protein adduct
epitope that have carboxyalkyl side chains. The detection range of anti-iso[4]LGE2 mAb
for ELISA and western analysis was 5 µg/ml. Pilot Western blot analysis against
iso[4]LGE2-BSA, a standard protein containing the iso[4]LGE2 modification plus BSA
control, showed high specificity and selectivity with our anti-iso[4]LGE2 mAb.
169

5.4 Experimental Procedures
5.4.1 Materials. The following commercially available materials were used as received:
DMEM-high glucose (no pyruvate #10-017) was from Mediumtech. Oxaloacetic acid
(#O7753), insulin (#I4011), Na pyruvate (#P3662), 8-azaguanine (#A5284), hybridoma
growth supplement (NCTC-109) (#N1140), 50 x HAT (#H0262), 50 x HT (#H0137),
naphthol blue black (N3393) were from Sigma (St. Louis, MO). PEG (#783641) was
from Roche (Madison, WI). Fatal bovine serum (#16000-044), trypan blue (0.4%),
penicillin/streptomycin (PS #12800) and gentamicin (#15750-060) were from Gibco
(Grand Island, NY). CellTM mAb medium, quantum yield and animal component free
grade were from Becton&Dickinson (Franklin Lakes, NJ). HL-1TM chemically defined
serum-free medium was from LONZA. Protein G resin was from GenScript (Piscataway,
NJ). Glycine (161-0724) and Quick Start™ Bradford dye reagent (1x) were from Bio-
Rad (Hercules, CA). ABTS Ready to USE substrate solution was from Invitrogen.
Immunization stock iso[4]LGE2-KLH (3.604 μg/μl, pyrrole = 721.3 μM, ratio = 600.4)
was prepared by Dr. Jim Laird.
General methods:
(1) Macrophage extraction:
Materials used for macrophage extraction included 10 mL sterile syringes (one for
each mouse), 18 1x1/2 gauge needles (one for each mouse), 70% ethanol for
spraying, 2 autoclaved surgical scissors, forceps, curled fine forceps, 50 mL sterilized
conical tubes and an ice bucket. Macrophage medium, also called super HAT
medium, was prepared by reconstituting the contents of a commercial vial (medium
supplement (50×) Hybri-Max™ Sigma H0262, γ-irradiated, lyophilized powder) with
170

10 mL of sterile cell culture medium (DMEM). The final working concentration of
each component was 100 μM hypoxanthine, 0.4 μM aminopterin and 16 μM
thymidine.
(2) SP2/0 myeloma cells growth:
Preparation of 8-azaguanine was achieved by reconstituting a commercial vial with
10 mL DMEM, to make a 50x stock; that then aliquoted as 2 mL per tube and stored
at -20 ℃. The final concentration was 130 µM. 100x Solution I stock was prepared by
adding 1320 mg oxaloacetic acid, 71 mg insulin (powder at 28 U/mg) and 550 mg Na
pyruvate to 100 mL PBS, then aliquoting as 6 mL per tube and stored at -20 ℃. The
final concentration of each component was 0.2 Units/ml insulin, 0.5 mM Na pyruvate
and 1 mM oxaloacetic acid. Solution I was a good supplement to DMEM medium to
provide the proper pH and enhance growth of the hybridoma cells. SP2/0 passage
medium contained 87 mL DMEM, 10 mL FBS, 1 mL 50x 8-azaguanine, 1 mL 100x
Solution I, 1 mL L-glutamine and 100 µl gentamicin. SP2/0 growth medium was
prepared by adding 20 mL FBS, 1 mL 100x Solution I, 2 mL NCTC-109, 1 mL L-
glutamine and 100 µl gentamicin to 78 mL DMEM.
(3) Fusion:
Materials used for fusion included sterile scissors, forceps, scalpels and sterile glass
slides with frosted end. Super DMEM medium was prepared by adding 120 mL FBS,
60 mL NCTC-109, 6 mL 100x Solution I, 12 mL 50x HAT or HT medium, 6 mL L-
glutamine and 600 µl gentamicin to 400 mL DMEM. Hybridoma medium (20%)
included 78 mL DMEM, 20 mL FBS, 6 mL NCTC-109, 1 mL 100x Solution I, 1 mL
171

L-glutamine and 100 µl gentamicin. Hybridoma medium (15%) contained 83 mL
DMEM, 15 mL FBS, 2.5 mL NCTC-109, 1 mL 100x Solution I, 1 mL L-glutamine
and 100 µl gentamicin.
(4) Antibody purification:
Binding and elution buffers were 20 mM sodium phosphate buffer (pH 7) and 0.1 M
glycine, (pH 2.6) (375.35 mg/50 ml sterilized water, make fresh), respectively.
5.4.2 Immunization. Five ear tagged and numbered Balb/c mice were given
intraperitoneal (IP) injections of the immunogen (iso[4]LGE2-KLH). For the first IP
injection, 140 μl the antigenic stock solution (containing 500 μg of immunogen) was
emulsified with 360 μl of PBS and 500 μl FCA. Each mouse received 0.2 ml of the
emulsion utilizing a 25 gauge needle. The initial injection contained immunogen (100 μg
per mouse) in PBS and an equivalent volume of Fruend’s complete adjuvant (FCA,
Sigma F-5881). For booster IP injections, 70 μl the antigenic stock (containing 250 μg of
immunogen) solution was emulsified with 430 μl of PBS and 500 μl FIA. The animals
received their second injection two weeks after the first injection. The second injections
were given utilizing the same volume (0.2 ml) as the initial injection, but with half as
much immunogen (50 μg per mouse). Fruend’s incomplete adjuvant (FIA, Sigma, F-
5506) was used for this and all subsequent injections. Three weeks latter a third injection
was given and then the fourth injection. Ten to fourteen days following the fourth
injection a blood sample was collected from each mouse. The mice were anesthetized
with halothane at the Case animal facility. When the mice showed no response to the foot
pinch test, they were restrained by hand and the retro orbital sinus was punctured with the
172

tip of a 250 μl non-heparinized capillary tube (Fisher cat# 02-668-10). Approximately
100 μl of blood was collected from each mouse. After the bleed, the mice were returned
to their respective cages to recover. Serum was prepared and tested from each blood
sample. Thus, collected blood was spun at 7000 rpm for 10 min. The serum was aspirated
and diluted 1:10,000 for ELISA analysis. Once a satisfactory immune response was
established in one of the mice, a fusion was initiated. Three days before the fusion, the
mouse with the best immune response was injected intravenously (IV. injection into the
tail vein) with 0.2 ml of the sterile antigenic solution (50 μg per mouse, 14 μl the
antigenic stock solution mixed with 186 μl of PBS) without adjuvant utilizing a 27 gauge
needle. The mouse was restrained with a small rodent restraint for this injection. For the
fusion, the mouse was euthanized via rapid cervical dislocation and the spleen aseptically
removed. The remaining four mice were kept until it was verified that antibody was
obtained that exhibited high specificity to the immunogen.
5.4.3 Cell counting. To determine the cell density and viability, cell counting was
performed using a hemocytomer. Thirty μl of the cell suspension was mixed with 30 μl
(equal volume) of 0.4% trypan blue solution to create a dilution factor of 2 and then
transferred into a fresh 1.5-mL tube followed by incubation for 5 min. With a cover-slip
in place, 10 μl of the trypan blue-stained cell suspension was loaded into a chamber on
the hemocytometer. All the cells were counted in the nine 1-mm wide center squares that
hold up to a total volume of 0.9 μl. Viable cells remained opaque, whereas non-viable
cells were stained in blue. The cell density and viability were calculated using the
following equations.
173

Viable cell density (cells per mL) = (total number of viable cells/0.9μl) x dilution factor x
1000 μl/mL
Cell viability = (number of viable cells/number of viable and dead cells) x 100%
5.4.4 Macrophage intraperitoneal lavage exaction. Macrophage extraction from ten
Balb/c mice was performed two weeks before fusion. Another five mice were used for
each of two rounds of subcloning. After assembling all materials, autoclaving instruments
and preparing super HAT medium and storing it at 37 °C, mice were euthanized by rapid
cervical dislocation to avoid excessive bleeding into the peritoneal cavity. Thus, mice
were firmly held in one hand by the nape of neck and the tail was pulled with other hand
until a pop was felt owing to a separation between cranium and backbone. Mice were
sterilized with 70% ethanol and immediately placed into tissue culture hood. A small
incision was made for laparotomy in the abdominal skin of the mouse with forceps and
scissor. Mice were opened by pulling the skin firmly with both hands to each side so that
skin was completely pulled down. Then the peritoneum was sprayed with 70% ethanol. A
needle was uncapped inside the hood and 5-6 ml 11.6% of sucrose in sterilized water
(w/w) was squirted into the peritoneum. The needle was injected, hole-side down, into
the mouse in its lower abdominal area, preferably in a region where there was fat. During
the process, needle was inserted gently and not too deeply without perforating any organs
and the needle was withdrawn slowly so that the fat didn’t clog the needle. The mouse
skin was transparent after removing fur, so it could avoid puncture the organs when
inserted needle. The mouse was massaged the abdominal area to promote the detachment
of sucrose solution to tissues (15 seconds), and then repeated twice. For macrophage
174

extraction, the needle of the same syringe was inserted horizontally into the upper part of
abdomen. Then about 4 ml sucrose solution containing macrophages was withdrawn.
This procedure was repeated twice to provide a total of about 12 ml fluid. The cell
suspension was poured into a 50 ml collection tube and the cells were centrifuged at 1000
rpm for 10 min at 20 °C. The cells were re-suspended using a 10 ml pipette and
macrophage medium was added to the pellet (10 ml per plate at a concentration of 105
cells/ml). The myeloma cells were fed with suspended macrophage cells, 100 µl per well
(104 cells/well) and then incubated at 37 °C in 5% CO2.
5.4.5 Growth of SP2/0 myeloma cells. The myeloma SP2/0 cells were grown in passage
medium (15% FBS) for about 6 days then slowly (after making sure that the cell viability
was 90% or higher before transfer to new medium, see p173 section 5.4.3) put into
growth medium (20% FBS). After centrifugation at 1200 rpm for 5 min at 4 °C, the
supernatant was removed and the pellet re-suspended in 10 ml of DMEM +
penicillin/streptomycin (PS) + 2% FBS.
5.4.6 Fusion Procedure. All fusion work was done in a cell culture laminar flow hood
under sterile conditions. The spleen was removed from the immunized mouse and placed
in a petri dish containing 10 ml of DMEM, penicillin/streptomycin (100 µl) and 2% FBS
(200 µl). The spleen was cut into 4 pieces. Each piece was gently ground between two
frosted end slides to release all cells from the connective tissue. The spleen cells were
collected in a 50 ml conical tube and put on ice. The SP2/0 myeloma cells were also
collected in a 50 ml conical tube and chilled on ice. Both sets of cells were washed twice
175

with 20 ml of cold serum free-DMEM. Both pellets were re-suspended in 5 ml of cold
serum free-DMEM and put on ice. Using a hemocytometer, the spleen cells were counted
in a 20 μl aliquot and after adding 20 μl 4% acetic acid that would lyse any RBC’s.
Fusion was done at a cell ratio of 3:1 (spleen cells: myeloma). 6.3 million spleen cells
were used for this fusion, so the number of SP2/0 myeloma cells would be 2.1 million.
Using a hemocytometer, the myeloma cells were also counted in a 20 μl aliquot. 6.3
million spleen cells with 2.1 million myeloma cells were mixed in one 50 ml conical tube
and the mixed cells were spun down. All supernatant was removed and the pellet made as
dry as possible. The pellet was loosened by briskly tapping the tube. It was important to
break up the pellet so the PEG would contact as many cells as possible. The tube was
placed in a 37 °C water bath and 1 ml of 37 ℃ PEG 1500 solution (Ready-to-Use PEG
1500 (Cat#783641, 50% PEG 1500 (w/v) in 75 mM Hepes (pH 8.0)) from Roche) was
added dropwise directly onto the pellet over a full 1 minute period. The tube was rotated
while adding the PEG and gently swirled to resuspend the cells allowing the mixture to
sit in the water bath for 1 minute. While rotating the tube, 1 ml of 37 °C, serum free-
DMEM was added over 1 minute. Then 20 ml more of 37 °C serum free-DMEM was
added dropwise over 4 minutes. Then the cells were spun down at 1000 rpm for 5
minutes. It was not unusual to see two layers of cells at this stage. The supernatant was
removed immediately and the cell pellet was re-suspended in 100 ml super HAT medium.
Cell suspension (0.1 ml) was placed in each well of a twenty 96-well plate already that
contained macrophages, and the medium was changed every 5 days. By 10-14 days,
colonies of hybridoma cells were apparent. They usually grew on the periphery of each
176

well. Culture supernatant (50 μl) could be removed for screening when 1/4 to 1/3 of the
well was covered with growing cells.
5.4.7 Clonal selection. At approximately 5 days post fusion the initial signs of viable
hybridoma colonies were observable in the wells. Concomitantly, the wells containing
unfused myeloma cells were dead or dying. By 12-14 days post fusion the hybridoma
colonies were macroscopically visible and ready for assay by ELISA for antibody
containing wells. After the first screening, viable clones were chosen and submitted to
another two rounds of subcloning to test the clone cells’ productivity and consistency,
and also to make sure that each clone was derived from a single cell.
5.4.8 Cultivation of hybridoma cells
The cultivation process was carried out inside a laminar flow hood. In order to prevent
cells from any potential contamination, all tools and containers were disinfected with
70% ethanol before using in the hood. A cryogenic vial containing 1 mL of frozen (in
liquid nitrogen) hybridoma cells (~107 cells) expressing anti-iso[4]LGE2 mAb was
thawed at 37 °C in a water bath immediately after being taken out of the liquid nitrogen.
The thawed cell suspension was transferred into 10 mL of cell culture medium DMEM in
a fresh 15-mL conical tube and spun down at 1000 rpm for 5 min at room temperature.
The supernatant was discarded after centrifugation. The cell pellet was resuspended
gently in 10 mL hybridoma medium (DMEM 8.3 mL, FBS 1.5 mL, NCTC-109 250 µl,
100x Solution I 100 µl, L-glutamine 100 µl and gentamicin 10 µl). The 10-mL cell
suspension was plated in a 100 mm cell culture dish with a seeding density of 106 viable
177

cells/mL and a cell viability of 90% or higher (see p173 section 5.4.3) and incubated at
37 °C in 5% CO2. After 2-3 days, the cells were split into 3-4 fresh culture dishes with 10
mL of hybridoma medium for each. For freezing cell samples, about 108 hybridoma cells
(90% viable or higher) of each passage were harvested by centrifugation at 1000 rpm for
5 min and resuspended in 10 mL of freezing medium consisting of fresh DMEM (80%),
FBS (10%) and DMSO (10%). The cell suspension was transferred in 1-mL aliquots into
1.7-mL cryogenic vials that were later placed into a foam box, and subsequently frozen at
-80 °C for the first 24-48 hours, then placed into liquid nitrogen in a cryogenic tank for
long-term storage.
5.4.9 Adaptation of hybridoma cells to serum free medium
The hybridoma cells cultivated in hybridoma medium (mentioned above) were
sequentially adapted to HL-1TM chemically defined serum-free medium by following the
phases as indicated in Table 5.1. To avoid decreases of cell viability, the cells were
maintained under the same conditions in each phase for a few passages and then passaged
to the next phase until the cell viability reached to 90% or higher (see p173 section 5.4.3).
Table 5.1 Sequential adaptation of hybridoma cells to serum free medium
Phase Component of cell culture medium
1 100 % hybridoma medium
2 75% hybridoma medium; 25% HL-1 serum-free medium
3 50% hybridoma medium; 50% HL-1 serum-free medium
4 25% hybridoma medium; HL-1 serum-free medium
5 100% HL-1 serum-free medium
178

5.4.10 Expression of anti-iso[4]LGE2 mAb
The hybridoma cells were subcultured with a seeding density of about 106 cells/mL in
200 mL of HL-1 serum-free medium with 2 mL of the penicillin and streptomycin
solution in two 250-mL shaking flasks. To maximize the production of anti-iso[4]LGE2
mAb, each flask containing 100 mL of cell culture was incubated at 37 °C in 5% CO2
with shaking at 60 rpm (Advanced digital orbital shaker from VWR) for 5-7 days without
changing the medium. By the end of this period, most of the cells died. The cell
suspension was transferred into six 50-mL conical tubes that were then centrifuged at 4
°C for 30 min at 10,000x g. The supernatant containing anti-iso[4]LGE2 mAb was
filtered through a 0.45 μm filter membrane (MILLEX® -GP syringe driven filter unit
from Millipore) with the aid of a water-driven aspirator, put into fresh tubes and kept at 4
°C.
5.4.11 Purification of the anti-iso[4]LGE2 mAb. A protein G column was packed with
immunopure protein G beads (5.0 ml), and was then equilibrated with 10 column
volumes of binding buffer (20 mM sodium phosphate buffer). Anti-iso[4]LGE2
monoclonal supernatant in animal component free medium (250 ml) was diluted 1:1 with
binding buffer (250 ml). 2-5 ml of Tris-HCl (pH 8) was added to adjust the pH>7 (pH is
important for IgG binding to protein G beads). The diluted supernatant was loaded onto
the top of the column and allowed to flow through it with gravity. In order to increase the
binding saturation of protein G, the eluate was recycled by loading it to columns three
times. In the future, it would be advisable to re-load the eluate to a washed column to see
if any mAb remained. Then binding buffer (50 ml) was passed through the column to
179

remove unbound non-IgG proteins. IgG was then eluted with elution buffer (100 mM
glycine-HCl, pH 2.6) collected into 1.0 ml fractions containing 100 µl of 1 M Tris-HCl
(pH 9.5) buffer to neutralize the pH. The protein level of each eluted fraction was
qualitatively monitored using the Quick Start™ Bradford protein assay. Briefly, 100 μl of
an eluted fraction was added to 100 μl of Bradford dye reagent in a 96-well plate
followed by gently mixing with a pipette at room temperature. The color of the eluted
protein-containing solution (blue) was compared to the color of the reference solution
(bright green) containing a 1:1 mixture of elution buffer and Bradford dye reagent. The
more protein present in the solution, the more blue the mixture appeared to be. Collection
of fractions was terminated when the color of the dye mixture appeared nearly bright
green. The higher absorbance fractions (2-3 ml) were combined and dialyzed against 10
mM PBS (pH 7.4) 2×2 L at 4 °C for 24 h. The final concentration (1-2 mg/ml) was
measured at OD 280 nm. Several batches of mAb were purified, the concentrations
varies. The concentrations were 1.1 mg/ml, 1.13 mg/ml and 2.2 mg/ml for three different
batches. Binding buffer was used to re-equilibrate the column, then the column was
washed with 20% ethanol (15 ml) for storage at 4 °C.
5.4.12 Indirect enzyme-linked immunosorbent assay (ELISA) for anti-iso[4]LGE2
mouse serum, clone screening and purified mAb. Antigen (iso[4]LGE2-BSA,
iso[4]LGE2-HSA, BSA, HSA, LGE2-BSA, LGE2-KLH, CEP-HSA) solutions (1.2 μg in
10 mM PBS,100 μl/well, 0.012 μg/μl) was added to each well of a sterilized 96 well
ELISA plate (Cat. 9018, Corning Inc. Corning, NY). The plate was incubated overnight
at 4 °C. After the coating solution was discarded, each well was washed with PBST
180

(Tween 0.1%) (4×300 µl) and tapped onto paper towels until dry, then filled with 2%
BSA /PBS 300 µl, and incubated at 37 °C for 2 h to block the remaining active sites on
the plastic solid phase. In a pilot study, 3% chicken egg ovalbuman/PBS was tested as
blocking buffer. However, the blocking wasn’t efficient. The medium control also showed
a weak signal. Using 2% BSA/PBS as blocking buffer, the medium control showed
almost no signal, just like the untreated background. So 2% BSA/PBS was chosen as
blocking buffer. Each well was then washed 4 times with PBST (Tween 0.1%) and tapped
onto paper towels till dry. Mouse serum (100 μl diluted 10,000:1 with PBS) or clone
supernatant or protein G purified anti-iso[4]LGE2 mAb from HL-1 serum free medium (5
μg/ml in PBS) was dispensed into the ELISA plate wells and the plate was incubated
overnight at 4 °C. Pre-immune mouse serum (before injection with immunogen), diluted
as above, was employed as a negative response control if necessary. After discarding the
supernatant and washing with PBST (Tween 0.1%) (4×300 µl), 100 μl goat anti-mouse
HRP-conjugated secondary antibody (IgG specific), diluted 1:1200 with PBST (Tween
0.1%), was added to each well. Then the plate was incubated with shaking at room
temperature for 1 h using an orbital shaker (IKA MTS 2/4 digital microtiter shaker, 600
rpm). After discarding the supernatant, the wells were washed PBST (Tween 0.1%)
(4×300 µl). To each well was then added 100 µl of ABTS Ready-to-USE substrate
solution (from Invitrogen). The plate was allowed to develop at room temperature for 20
min at room temperature and all the absorbances were measured using a microplate
reader with a test wavelength of 450 nm.
5.4.13 Enzyme-linked immunosorbent assay (ELISA) for competitive antibody
181

binding inhibition studies using the anti-iso[4]LGE2 mAb. For antibody binding
inhibition studies to measure cross-reactivities, iso[4]LGE2-BSA was used as coating
reagent and iso[4]LGE2-HSA was used as a standard for purified anti-iso[4]LGE2 mAb.
LGE2-BSA, CEP-BSA, CEP-HSA, HSA and LGE2-KLH served as inhibitors. For each
inhibitor, up to eleven serial dilutions of a standard, a blank, and a positive control, were
run. Each well of a 96 well ELISA plate was coated with iso[4]LGE2-BSA solution (100
µl), prepared by diluting a solution containing 1016 μM iso[4]LGE2-BSA to 1.12 μM
(0.3 μg/well) with pH 7.4 PBS (10 mM). The plate was incubated overnight at 4 °C, then
washed with 10 mM PBST (Tween 0.1%) (4×300 µl) and then blocked by incubating 1 h
at 37 °C with 300 µl of 2% BSA in 10 mM PBS. Then the plate was rinsed with 10 mM
PBST (Tween 0.1%) (4×300 µl). Eleven serial dilutions of inhibitor or standard (120 µl
each with a dilution factor of 0.2) were pre-incubated at 37 °C for 1 h with protein G
purified anti-iso[4]LGE2 monoclonal antibody from HL-1 serum free medium (120 µl, 5
μg/ml in 10 mM PBS).The initial inhibitor and standard concentrations were iso[4]LGE2-
HSA (446.5 μM), iso[4]LGE2-KLH (446.5 μM), LGE2-BSA (446.5 μM), CEP-BSA
(446.5 μM), CEP-HSA (446.5 μM), BSA (446.5 μM) and HSA (446.5 μM) respectively.
These were prepared by diluting a iso[4]LGE2-HSA (893.3 μM), iso[4]LGE2-KLH (721.3
μM), LGE2-BSA (1974 μM), CEP-BSA (496 μM), CEP-HSA (623.6 μM), BSA (1500
μM) and HSA (1500 μM) with 10 mM PBS, respectively. Blank wells were filled with
2% BSA/PBS (100 µl). Positive control wells were filled with the diluted antibody
solution (50 µl) and PBS (50 µl). The antibody antigen complex solutions (100 µl) were
then added in duplicate to their respective halves of the plate, which was then incubated
at room temperature with gentle agitation on an orbital shaker (IKA MTS 2/4 digital
182

microtiter shaker, 600 rpm) for 1 h. After discarding the supernatant and washing with
PBST (Tween 0.1%) (4×300 µl), 100 μl goat anti-mouse HRP-conjugated secondary
antibody (IgG specific), diluted 1:1200 with PBST (Tween 0.1%), was added to each
well and then the plate was incubated with shaking at room temperature for 1 h on the
orbital shaker. After discarding the supernatant, the wells were washed PBST (Tween
0.1%) (4×300 µl). To each well was then added 100 µl of Ready-to-Use substrate
solution ABTS. The plate was allowed to develop at room temperature for 20 min at
room temperature and all the absorbances were measured using a microplate reader with
a test wavelength of 450 nm. The absorbance in each well was measured with a dual-
wavelength Bio-Rad 450 microplate reader (Hercules, CA) with detection at 405 nm
relative to 655 nm. Absorbance values for duplicate assays were averaged and scaled to
make the maximum curve fit value close to 100 percent. The averaged scaled percent
absorbance values were plotted against the log of concentration. Theoretical curves for
each plot were fit to the absorbance data with a four parameter logistic function, f(x) = (a-
d)/[1+(x/c)b]+d using Origin 9.0 (OriginLab Corporation, Northampton, MA). Parameter
a = the asymptotic maximum absorbance, b = slope at the inflection point, c = the
inhibitor concentration at the 50 % absorbance value (IC50), and d = the asymptotic
minimum absorbance.
5.4.14 Western analysis. It was carried out according to published protocols.7-8
Iso[4]LGE2-BSA (~ 10 ng -10 µg) was dissolved in Laemmli sample buffer,9 fractionated
on a 4-12% SDS-PAGE, and electro blotted onto a polyvinyl difluoride (PVDF)
membrane (Millipore, Bedford, MA). The PVDF membranes were then treated with
183

Odyssey blocking buffer (Li-Cor cat# 927-40000) and probed using anti-iso[4]LGE2
mAb at various dilutions. Goat anti-mouse IRDye® 680 (Odyssey cat# 926-32220) was
used as secondary antibody. The immunoreactive bands were tested and quantified using
a Li-Cor infrared imaging system.
184

5.5 References
1. Shringarpure, R.; Grune, T.; Sitte, N.; Davies, K. J. (2000) 4-Hydroxynonenal-
modified amyloid-beta peptide inhibits the proteasome: possible importance in
Alzheimer's disease. Cell Mol Life Sci 57 (12), 1802-1809.
2. Nagata, K.; Suzuki, H.; Sakaguchi, S. (2007) Common pathogenic mechanism in
development progression of liver injury caused by non-alcoholic or alcoholic
steatohepatitis. J Toxicol Sci 32 (5), 453-468.
3. Salomon, R. G.; Batyreva, E.; Kaur, K.; Sprecher, D. L.; Schreiber, M. J.; Crabb,
J. W.; Penn, M. S.; DiCorletoe, A. M.; Hazen, S. L.; Podrez, E. A. (2000) Isolevuglandin-
protein adducts in humans: products of free radical-induced lipid oxidation through the
isoprostane pathway. Biochim Biophys Acta 1485 (2-3), 225-235.
4. Salomon, R. G.; Sha, W.; Brame, C.; Kaur, K.; Subbanagounder, G.; O'Neil, J.;
Hoff, H. F.; Roberts, L. J. (1999) Protein adducts of iso 4 levuglandin E-2, a product of
the isoprostane pathway, in oxidized low density lipoprotein. J Biol Chem 274 (29),
20271-20280.
5. Davies, S. S.; Talati, M.; Wang, X.; Mernaugh, R. L.; Amarnath, V.; Fessel, J.;
Meyrick, B. O.; Sheller, J.; Roberts, L. J., 2nd (2004) Localization of isoketal adducts in
vivo using a single-chain antibody. Free Radic Biol Med 36 (9), 1163-1174.
6. Morrow, J. D.; Hill, K. E.; Burk, R. F.; Nammour, T. M.; Badr, K. F.; Roberts, L.
J., 2nd (1990) A series of prostaglandin F2-like compounds are produced in vivo in
humans by a non-cyclooxygenase, free radical-catalyzed mechanism. Proc Natl Acad Sci
U S A 87 (23), 9383-9387.
185

7. Crabb, J. W.; Miyagi, M.; Gu, X. R.; Shadrach, K.; West, K. A.; Sakaguchi, H.;
Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G. (2002)
Drusen proteome analysis: An approach to the etiology of age-related macular
degeneration. Proc Natl Acad Sci U S A 99 (23), 14682-14687.
8. Bhattacharya, S. K.; Rockwood, E. J.; Smith, S. D.; Bonilha, V. L.; Crabb, J. S.;
Kuchtey, R. W.; Robertson, N. G.; Peachey, N. S.; Morton, C. C.; Crabb, J. W. (2005)
Proteomics reveal cochlin deposits associated with glaucomatous trabecular meshwork. J
Biol Chem 280 (7), 6080-6084.
9. Laemmli, U. K. (1970) Cleavage of structural proteins during assembly of head of
bacteriophage-T4. Nature 227 (5259), 680-685.
186

Appendix

Adduct Form Formula Mass (Cal.) Pyrrole C20H28O3 316.20 Pyrrole monodehydrated C20H26O2 298.19 Lactam C20H28O4 332.20 Lactam monodehydrated C20H26O3 314.19 Hydroxylactam C20H26O5 348.19 Hydroxylactam monodehydrated C20H26O4 330.18 Hydroxylactam bisdehydrated C20H24O3 312.17
Table 2.1S Lysine modifications with iso[4]LGE2 Peptide Sequence m/z (obs) Mr (obs) ion score Adduct Modification form SVTGAK280QVNY 690.8754 (+2) 1379.7508 29 C20H26O3 Lactam Monodehydrated Table 2.2S Peptide of calpain-1 modified by iso[4]LGE2, as identified by Mascot analysis # b b++ b* b*++ b0 b0++ Seq. y y++ y* y*++ y0 y0++ #
1 88.0393 44.5233 70.0287 35.5180 S 10
2 187.1077 94.0575 169.0972 85.0522 V 1293.7089 647.3581 1276.6824 638.8448 1275.6984 638.3528 9
3 288.1554 144.5813 270.1448 135.5761 T 1194.6405 597.8239 1177.6140 589.3106 1176.6299 588.8186 8
4 345.1769 173.0921 327.1663 164.0868 G 1093.5928 547.3001 1076.5663 538.7868 7
5 416.2140 208.6106 398.2034 199.6053 A 1036.5714 518.7893 1019.5448 510.2760 6
6 858.4971 429.7522 841.4706 421.2389 840.4866 420.7469 K 965.5342 483.2708 948.5077 474.7575 5
7 986.5557 493.7815 969.5292 485.2682 968.5451 484.7762 Q 523.2511 262.1292 506.2245 253.6159 4
8 1085.6241 543.3157 1068.5976 534.8024 1067.6136 534.3104 V 395.1925 198.0999 378.1660 189.5866 3
9 1199.6671 600.3372 1182.6405 591.8239 1181.6565 591.3319 N 296.1241 148.5657 279.0975 140.0524 2
10 Y 182.0812 91.5442 1
Table 2.3S Mascot characterization of the doubly charged ion m/z 690.88 (SVTGAKQVNY) showed series of fragment ions sufficient to unambiguously identify a iso[4]LGE2 modification on the calpain-1 lysyl residue (K280).
Figure 2.1S Domain architecture of calpain-1 and three active sites (left panel). Minicalpain was shown as subdomain IIa and domain IIb, with catalytic triad residue Cys 115, His 272 and Asn 296 (right panel).
188

101.587.5
52.7
35.8
27.9
18.8
Catalyitc Subunit
Small Subunit
Control
Crosslink
2 h Figure 2.2S Coomassie blue staining image of calpain-1 after 2 h treatment with 460 equivalents of iso[4]LGE2
Crosslink part
Catalytic subunit--80kDa
Control one
Modified Calpain-1
Use Anti-Calpain1 catalyticsubunit primary antibody
30 min 1 h 3 h
Use Anti-Calpain 1 smallsubunit primary antibody
ControlModified Calpain-1
Small Subunit--30kDa
30 min 1 h 3 h
Crosslink part
240 kDa
320 kDa
250
150
100
75
50
37
25
20
Use Anti-Iso[4]LGE2monoclonal antibody
Modified Calpain-130 min 1 h 3 h
Crosslink part
Catalytic subunit--80kDa
Control
250
150
100
75
50
37
2520
Figure 2.3S Western blot images of calpain-1 after 30 min, 1 h and 3 h treatment with 460 equivalents of iso[4]LGE2
189

Use Anti-Calpain 1 catalyticsubunit primary antibody
overnight
1 h 3 hcontrol overnight
overnight
1 h 3 h control
Catalytic subunit--80kDa
50 times 100 times
240 kDa160 kDa
320 kDa
120 kDa110 kDa
250
150
100
75
50
37
25
20
Use Anti-Calpain 1 smallsubunit primary antibody
overnight
1 h 3 hcontrol overnight
overnight
1 h 3 h control
50 times 100 times
Catalytic subunit--80kDa
140 kDa120 kDa110 kDa
Small Subunit--30kDa
overnight
1 h 3 h overnight
overnight
1 h 3 h
50 times 100 times
control
Catalytic subunit--80kDa
140 kDa120 kDa110 kDa
Small Subunit--30kDa
160 kDa240 kDa
320 kDa
Use Anti-Iso[4]LGE2 monoclonal antibody
250
150
100
75
5037
2520
Figure 2.4S Western blot images of calpain-1 after 1 h, 3 h and overnight treatment with 50 and 100 equivalents of iso[4]LGE2
190

overnight
1 h3 h control6 hovernight
1 h3 h6 h
10 times 5 times
Catalytic subunit--80kDa
120 kDa110 kDa
160 kDa240 kDa
Use Anti-Calpain I catalyticsubunit primary antibody
overnight
1 h3 h control6 hovernight
1 h3 h6 h10 times 5 times
Catalytic subunit--80kDa
Small Subunit--30kDa
140 kDa120 kDa110 kDa
Use Anti-Calpain I smallsubunit primary antibody
Figure 2.5S Western blot images of calpain-1 after 1 h, 3 h, 6h and overnight treatment with 5 and 10 equivalents of iso[4]LGE2
Control2times 24h
2times 48h
5times 24h
5times 48h
10times 24h
10times 48h
20times 24h
20times 48h
Catalytic subunit--80kDa
120 kDa110 kDa
160 kDa240 kDa
Use Anti-Calpain I catalyticsubunit primary antibody
Control2times
24h2times 48h
5times 24h
5times 48h
10times 24h
10times 48h
20times 24h
20times 48h
Catalytic subunit--80kDa
Small Subunit--30kDa
140 kDa120 kDa110 kDa
Use Anti-Calpain I smallsubunit primary antibody
Figure 2.6S Western blot images of calpain-1 after 24 h and 48 h treatment with 2, 5, 10 and 20 equivalents of iso[4]LGE2
191

A1 A2
B1 B2
Figure 2.7S ESI-MS of synthesis fluorescence labeled iso[4]LGE2 intermediate (2.3, panel A1 and A2) and final product (2.4, panel A1 and A2) in both negative mode and positive mode. Adduct Form Formula Mass (Cal.) Pyrrole C30H41N5O5S 583.28 Pyrrole monodehydrated C30H39N5O4S 565.27 Lactam C30H41N5O6S 599.28 Lactam monodehydrated C30H39N5O5S 581.27 Hydroxylactam C30H41N5O7S 615.27 Hydroxylactam monodehydrated C30H39N5O6S 597.26 Hydroxylactam bisdehydrated C30H37N5O5S 579.25 Table 2.4S Lysine modifications with fluorescence labeled iso[4]LGE2.
192

C
OH
ONHCH2CH2NH
NO
N
SO2NCH3
CH3N
C
OH
ONHCH2CH2NH
NO
N
SO2NCH3
CH3N
O
Protein Lys
LysProtein
C
OH
ONHCH2CH2NH
NO
N
SO2NCH3
CH3N
O
LysProtein
HO
C5H11
C5H11
C5H11
H2O
CO
NHCH2CH2NH
NO
N
SO2NCH3
CH3NProtein Lys C5H11
Pyrrole with fluorescence tag
Pyrrole monodehydrated
H2O
Lactam monodehydrated
CO
NHCH2CH2NH
NO
N
SO2NCH3
CH3N
O
LysProtein C5H11
Lactam with fluorescence tag
Hydroxylactam with fluorescence tag
CO
NHCH2CH2NH
NO
N
SO2NCH3
CH3N
O
LysProtein
HO
C5H11
H2O
Hydroxylactam monodehydrated
H2O
CO
NHCH2CH2NH
NO
N
SO2NCH3
CH3N
O
LysProtein C5H11
Hydroxylactam bisdehydrated
Scheme 2.1S Chemical structures of Lys-fluorescence labeled iso[4]LGE2 adducts.
193

Table A b b++ b* Seq. y y++ y* #
1 114.0913 57.5493 L 6
2 243.1339 122.0706 E 1127.5918 564.2995 1110.5654 5
3 952.4961 476.7517 935.4695 K 998.5492 499.7782 981.5223 4
4 1023.5332 512.2702 1006.5067 A 289.1870 145.0972 272.1608 3
5 1094.5703 547.7888 1077.5438 A 218.1499 109.5786 201.1236 2
6 K 147.1128 74.0600 130.0867 1
Table b b b++ b* b*++ Seq. y y++ y* y*++ #
1 100.0757 50.5415 V 10
2 807.4222 404.2147 790.3956 395.7015 K 1512.7780 756.8926 1495.7515 748.3794 9
3 906.4906 453.7489 889.4641 445.2357 V 805.4315 403.2194 788.4050 394.7061 8
4 963.5121 482.2597 946.4855 473.7464 G 706.3631 353.6852 689.3365 345.1719 7
5 1062.5805 531.7939 1045.5539 523.2806 V 649.3416 325.1745 632.3151 316.6612 6
6 1176.6234 588.8153 1159.5969 580.3021 N 550.2732 275.6402 533.2467 267.1270 5
7 1233.6449 617.3261 1216.6183 608.8128 G 436.2303 218.6188 419.2037 210.1055 4
8 1380.7133 690.7133 1363.6867 682.3470 F 379.2088 190.1081 362.1823 181.5948 3
9 1437.7348 719.3710 1420.7082 710.8577 G 232.1404 116.5738 215.1139 108.0606 2
10 R 175.1190 88.0631 158.0924 79.5498 1
Table 2.5S Mascot characterization of the doubly charged ion m/z 620.86 (LEKAAK) and triply charged ion m/z 537.96 (VKVGVNGFGR) showed series of fragment ions sufficient to unambiguously identify a fluorescence labeled iso[4]LGE2 modification on the GAPDH lysyl residue.
194

Panel A:
Panel B:
Figure 2.8S The peak area of the amino acids was compared with the peak area of the quantitative standard amino acids (Standard H). The detection limit was tested by 25, 50, 100, 250, 400 and 500 pmol of standard amino acids. Reliable relative standard deviation (< 10%) was achieved and the detection limit was determined with standard amino acids up to 50 pmol and was plotted with all amino acids in (B).
195

Table 2.6S (1) Amino acid analysis template.
196

NAME/ID# GAPDH(Rabbit) Dimer Injection Id: 1489
VOL HYDROLYZED 20.00
RATIO APPLIED 0.01600
MOLECULAR WEIGHT 35780.00
AMINO ACID Known Composition pmol analyzed pmol protein over 15% under 15% exp comp integer comp % int err
His (H) 11.00 4.5 0.41 0.41 0.41 10.42 10.00 9.09
Asn (N) 17.00 0.00 FALSE
Ser (S) 19.00 13.28 0.70 FALSE FALSE 30.74 31.00 63.16
Gln (Q) 6.00 0.00 FALSE
Arg (R) 10.00 4.91 0.49 0.49 0.49 11.37 11.00 10.00
Gly (G) 32.00 23.94 0.75 FALSE FALSE 55.42 55.00 71.88
Asp (D) 20.00 10.7 0.54 FALSE FALSE 24.77 25.00 25.00
Glu (E) 13.00 6.31 0.49 0.49 0.49 14.61 15.00 15.38
Thr (T) 21.00 6.66 0.32 0.32 FALSE 15.42 15.00 28.57
Ala (A) 34.00 11.87 0.35 0.35 FALSE 27.48 27.00 20.59
Pro (P) 11.00 4.82 0.44 0.44 0.44 11.16 11.00 0.00
Lys (K) 26.00 6.81 0.26 0.26 FALSE 15.76 16.00 38.46
Tyrb (Y) 9.00 3.61 0.40 0.40 0.40 8.36 8.00 11.11
Met (M) 10.00 2.54 0.25 0.25 FALSE 5.88 6.00 40.00
Val (V) 35.00 9.86 0.28 0.28 FALSE 22.82 23.00 34.29
Ile (I) 20.00 6.17 0.31 0.31 FALSE 14.28 14.00 30.00
Leu (L) 18.00 9.2 0.51 FALSE FALSE 21.30 21.00 16.67
Phe (F) 14.00 5.14 0.37 0.37 0.37 11.90 12.00 14.29
TOTAL #AA 326.00 NEW ESTIMATE
PMOL PROTEIN AVERAGE %
ESTIMATED AMOUNT WITHIN 15% 0.43 ERROR 26.78
PMOL PROTEIN 0.43
15% OF PROTEIN 0.06 TOTAL PMOL AMINO ACID 130.32
-15% OF PROTEIN 0.36 TOTAL PMOL HYDROL 27.00 TOTAL UGRAMS 0.97
+15% OF PROTEIN 0.49 CONC pmol/ul 1.35 CONC ug/ul 0.05
Table 2.6S (2) Amino acid composition of dimer of modified GAPDH.
197

NAME/ID# GAPDH(Rabbit) Trimer Injection Id:
1494
VOL HYDROLYZED 20.00
RATIO APPLIED 0.01600
MOLECULAR WEIGHT 35780.00
AMINO ACID Known Composition pmol analyzed pmol protein over 15% under 15% exp comp integer comp % int err
His (H) 11.00 5.46 0.50 0.50 0.50 11.38 11.00 0.00
Asn (N) 17.00 0.00 FALSE
Ser (S) 19.00 13.29 0.70 FALSE FALSE 27.69 28.00 47.37
Gln (Q) 6.00 0.00 FALSE
Arg (R) 10.00 5.96 0.60 FALSE FALSE 12.42 12.00 20.00
Gly (G) 32.00 22.87 0.71 FALSE FALSE 47.65 48.00 50.00
Asp (D) 20.00 11.69 0.58 FALSE FALSE 24.36 24.00 20.00
Glu (E) 13.00 5.77 0.44 0.44 0.44 12.02 12.00 7.69
Thr (T) 21.00 8.72 0.42 0.42 0.42 18.17 18.00 14.29
Ala (A) 34.00 13.33 0.39 0.39 FALSE 27.78 28.00 17.65
Pro (P) 11.00 5.47 0.50 0.50 0.50 11.40 11.00 0.00
Lys (K) 26.00 7.52 0.29 0.29 FALSE 15.67 16.00 38.46
Tyrb (Y) 9.00 4.4 0.49 0.49 0.49 9.17 9.00 0.00
Met (M) 10.00 3.23 0.32 0.32 FALSE 6.73 7.00 30.00
Val (V) 35.00 11.6 0.33 0.33 FALSE 24.17 24.00 31.43
Ile (I) 20.00 6.85 0.34 0.34 FALSE 14.27 14.00 30.00
Leu (L) 18.00 9.81 0.55 0.55 0.55 20.44 20.00 11.11
Phe (F) 14.00 6.62 0.47 0.47 0.47 13.79 14.00 0.00
TOTAL #AA 326.00 NEW ESTIMATE
PMOL PROTEIN AVERAGE %
ESTIMATED AMOUNT WITHIN 15% 0.48 ERROR 19.87
PMOL PROTEIN 0.48
15% OF PROTEIN 0.07 TOTAL PMOL AMINO ACID 142.59
-15% OF PROTEIN 0.41 TOTAL PMOL HYDROL 30.00 TOTAL UGRAMS 1.07
+15% OF PROTEIN 0.55 CONC pmol/ul 1.50 CONC ug/ul 0.05
Table 2.6S (3) Amino acid composition of trimer of modified GAPDH.
198

NAME/ID# GAPDH(Rabbit)
Tetramer Injection Id: 1499
VOL HYDROLYZED 20.00
RATIO APPLIED 0.01600
MOLECULAR WEIGHT 35780.00
AMINO ACID Known Composition pmol analyzed pmol protein over 15% under 15% exp comp integer comp % int err
His (H) 11.00 3.68 0.33 0.33 0.33 12.32 12.00 9.09
Asn (N) 17.00 0.00 FALSE
Ser (S) 19.00 10.35 0.54 FALSE FALSE 34.64 35.00 84.21
Gln (Q) 6.00 0.00 FALSE
Arg (R) 10.00 3.76 0.38 FALSE FALSE 12.59 13.00 30.00
Gly (G) 32.00 16.2 0.51 FALSE FALSE 54.22 54.00 68.75
Asp (D) 20.00 6.64 0.33 0.33 0.33 22.23 22.00 10.00
Glu (E) 13.00 3.54 0.27 0.27 0.27 11.85 12.00 7.69
Thr (T) 21.00 4.94 0.24 0.24 FALSE 16.53 17.00 19.05
Ala (A) 34.00 7.87 0.23 0.23 FALSE 26.34 26.00 23.53
Pro (P) 11.00 3.33 0.30 0.30 0.30 11.15 11.00 0.00
Lys (K) 26.00 4.21 0.16 0.16 FALSE 14.09 14.00 46.15
Tyrb (Y) 9.00 2.27 0.25 0.25 0.25 7.60 8.00 11.11
Met (M) 10.00 1.6 0.16 0.16 FALSE 5.36 5.00 50.00
Val (V) 35.00 6.7 0.19 0.19 FALSE 22.43 22.00 37.14
Ile (I) 20.00 4.05 0.20 0.20 FALSE 13.56 14.00 30.00
Leu (L) 18.00 6.14 0.34 FALSE FALSE 20.55 21.00 16.67
Phe (F) 14.00 3.49 0.25 0.25 FALSE 11.68 12.00 14.29
TOTAL #AA 326.00 NEW ESTIMATE
PMOL PROTEIN AVERAGE %
ESTIMATED AMOUNT WITHIN 15% 0.30 ERROR 28.61
PMOL PROTEIN 0.29
15% OF PROTEIN 0.04 TOTAL PMOL AMINO ACID 88.77
-15% OF PROTEIN 0.25 TOTAL PMOL HYDROL 18.67 TOTAL UGRAMS 0.67
+15% OF PROTEIN 0.34 CONC pmol/ul 0.93 CONC ug/ul 0.03
Table 2.6S (4) Amino acid composition of tetramer of modified GAPDH. Table 2.6S Amino acid analysis template and amino acid composition of dimer, trimer and tetramer of modified GAPDH.
199

Tryptophan Peptide
Iso4 Fluorescence Tag
TIC Mass
Figure 2.9S Solution trypsin digested unmodified GAPDH was monitored by iso4 fluorescence tag and tryptophan peptides fluorescence channels.
Tryptophan Peptide
Iso4 Fluorescence Tag
TIC Mass
Figure 2.10S Solution trypsin digested fluorescently labeled iso[4]LGE2 modified GAPDH was monitored by iso4 fluorescence tag and tryptophan peptides fluorescence channels.
200

Sequence Molecular weight Peptides
270-321 5787.6488 GILGYTEDQVVSCDFNSATHSSTFDAGAGIALNDHFVKLISWYDNEFGYSNR 262-307 4853.2878 QASEGPLKGILGYTEDQVVSCDFNSATHSSTFDAGAGIALNDHFVK 144-184 4434.2501 IVSNASCTTNCLAPLAKVIHDHFGIVEGLMTTVHAITATQ K 19-53 4023.8811 AAFNSGKVDVVAINDPFIDLHYMVYMFQYDSTHGK 26-59 4017.9069 VDVVAINDPFIDLHYMVYMFQYDSTHGKFHGTVK 161-192 3374.7292 VIHDHFGIVEGLMTTVHAITATQKTVDGPSGK 117-143 2964.4361 VIISAPSADAPMFVMGVNHEKYDNSLK 79-105 2930.3643 DPANIKWGDAGAEYVVESTGVFTTMEK 85-111 2869.3592 WGDAGAEYVVESTGVFTTMEKAGAHLK 138-160 2539.241 YDNSLKIVSNASCTTNCLAPLAK 199-217 1723.958 GAAQNIIPASTGAAKAVGK 218-232 1660.897 VIPELNGKLTGMAFR 65-78 1600.93 LVINGKAITIFQER 322-333 1389.6632 VVDLMVHMASKE 214-225 1223.7237 AVGKVIPELNGK 185-195 1214.6407 TVDGPSGKLWR 54-64 1186.6094 FHGTVKAENGK 1-11 1178.623 MVKVGVNGFGR 259-269 1154.6659 VVKQASEGPLK 60-70 1141.6455 AENGKLVINGK 250-257 922.476 AAKYDDIK 106-115 908.5192 AGAHLKGGAK 253-258 780.4017 YDDIKK 247-252 658.4013 LEKAAK 112-116 487.2867 GGAKR 258-261 472.3373 KVVK
Table 2.7S Calculated tryptic digested GAPDH peptides with one miss cleavage. All cysteines have been treated with iodoacetamide to form carbamidomethyl-cysteine. Methionines have been oxidized to form methionine sulfoxide. Using monoisotopic masses of the occurring amino acid residues and giving peptide masses as [M].
201

202

Table 2.8S Mass of cross-linked peptides [M], including +1 to +4 charges
203

TOF MS ES+ 282
m/z
m/z
TOF MSMS 640.97 ES+
32
Figure 2.11S TOF MS and MSMS of ion 640.9626.
171

Table 3.1S Molecular weight summary of Acetyl-Gly-Lys-O-Methyl Ester/Amyloid (11-17) “EVHHQKL” peptide reactions with iso[4]LGE2/iso[4]LGE2-Fluo tag
(1) Acetyl-Gly-Lys-O-Methyl Ester reaction with iso[4]LGE2 Acetyl-Gly-Lys-O-Methyl Ester (Average molecular weight): 259.3056 H:1.008 H2O:18.0153 Adducts-- no fluorescence tag
Adduct Formula
Adduct MW
Product MW
Plus one charge (+1)
Plus two charges (+2)
Pyrrole C20H28O3 316.4405 575.7461 576.7541 288.88105 Pyrrole -H2O C20H26O2 298.4252 557.7308 558.7388 279.8734 Lactam C20H28O4 332.4399 591.7455 592.7535 296.88075 Lactam-H2O C20H26O3 314.4246 573.7302 574.7382 287.8731 Hydroxylactam C20H28O5 348.4393 607.7449 608.7529 304.88045 Hydroxylactam-H2O C20H26O4 330.4240 589.7296 590.7376 295.8728 Hydroxylactam-2H2O C20H24O3 312.4088 571.7144 572.7224 286.8652
Cross-link C20H33O4 337.4796 853.0668 854.0748 427.5414 Cross-link-H2O C20H31O3 319.4643 835.0515 836.0595 418.53375 Cross-link-2H2O C20H29O2 301.4491 817.0362 818.0442 409.5261 Bis-pyrrole C40H58O6 634.8969 1149.4761 1150.4841 575.74605 Bis-pyrrole-H2O C40H56O5 616.8816 1131.4608 1132.4688 566.7384 Bis-pyrrole-2H2O C40H54O4 598.8664 1113.4455 1114.4535 557.73075 Tris-pyrrole C60H88O9 953.3353 1725.2041 1726.2121 863.6101 Tris-pyrrole-H2O C60H86O8 935.3200 1707.1888 1708.1968 854.6024 Tris-pyrrole-2H2O C60H84O7 917.3047 1689.1735 1690.1815 845.5948 Tris-pyrrole-3H2O C60H82O6 899.2894 1671.1582 1672.1662 836.5871 Acetyl-Gly-Lys-O-Methyl Ester (Monoisotopic molecular weight): 259.1532 H:1.0078 H2O:18.0106 Adducts-- no fluorescence tag
Adduct Formula
Adduct MW
Product MW
Plus one charge (+1)
Plus two charges (+2)
Pyrrole C20H28O3 316.2038 575.3570 576.3648 288.6863 Pyrrole -H2O C20H26O2 298.1933 557.3465 558.3543 279.68105 Lactam C20H28O4 332.1988 591.3520 592.3598 296.6838 Lactam-H2O C20H26O3 314.1882 573.3414 574.3492 287.6785 Hydroxylactam C20H28O5 348.1937 607.3469 608.3547 304.68125 Hydroxylactam-H2O C20H26O4 330.1831 589.3363 590.3441 295.67595 Hydroxylactam-2H2O C20H24O3 312.1725 571.3257 572.3335 286.67065 Cross-link C20H33O4 337.2379 852.5209 853.5287 427.26825 Cross-link-H2O C20H31O3 319.2273 834.5103 835.5181 418.26295 Cross-link-2H2O C20H29O2 301.2168 816.4997 817.5075 409.25765 Bis-pyrrole C40H58O6 634.4233 1148.6985 1149.7063 575.35705 Bis-pyrrole-H2O C40H56O5 616.4128 1130.6879 1131.6957 566.35175 Bis-pyrrole-2H2O C40H54O4 598.4022 1112.6773 1113.6851 557.34645 Tris-pyrrole C60H88O9 952.6428 1724.0556 1725.0634 863.0356 Tris-pyrrole-H2O C60H86O8 934.6322 1706.0450 1707.0528 854.0303 Tris-pyrrole-2H2O C60H84O7 916.6216 1688.0344 1689.0422 845.0250 Tris-pyrrole-3H2O C60H82O6 898.6110 1670.0238 1671.0316 836.0197
172

(2) Acetyl-Gly-Lys-O-Methyl Ester reaction with iso[4]LGE2-Fluo tag Acetyl-Gly-Lys-O-Methyl Ester (Monoisotopic molecular weight): 259.1532 H:1.0078 H2O:18.0106 Adducts-- no fluorescence tag
Adduct Formula
Adduct MW
Product MW
Plus one charge (+1)
Plus two charges (+2)
Pyrrole C30H41N5O5S 583.2828 842.4360 843.4438 422.2258 Pyrrole -H2O C30H39N5O4S 565.2723 824.4255 825.4333 413.22055 Lactam C30H41N5O6S 599,2778 858.4310 859.4388 430.2233 Lactam-H2O C30H39N5O5S 581.2672 840.4204 841.4282 421.2180 Hydroxylactam C30H41N5O7S 615.2727 874.4259 875.4337 438.22075 Hydroxylactam-H2O C30H39N5O6S 597.2621 856.4153 857.4231 429.21545 Hydroxylactam-2H2O C30H37N5O5S 579.2515 838.4047 839.4125 420.21015 Cross-link C30H46N5O6S 604.3168 1119.5999 1120.6077 560.80775 Cross-link-H2O C30H44N5O5S 586.3062 1101.5893 1102.5971 551.80245 Cross-link-2H2O C30H42N5O4S 568.2956 1083.5787 1084.5865 542.79715 Bis-pyrrole C60H84N10O10S2 1168.5813 1682.8562 1683.8640 842.4359 Bis-pyrrole-H2O C60H82N10O9S2 1150.5707 1664.8456 1665.8534 833.4306 Bis-pyrrole-2H2O C60H80N10O8S2 1132.5601 1646.835 1647.8428 824.4253
(3) Amyloid (11-17) “EVHHQKL” peptide reaction with iso[4]LGE2 EVHHQKL (Average molecular weight): 889.99854, (+1) 891.00593, (+2) 446.0666 H:1.008 H2O:18.0153 Adducts-- no fluorescence tag
Adduct Formula
Adduct MW
Product MW
Plus one charge (+1)
Plus two charges (+2)
Plus three charges (+3)
Plus four charges (+4)
Pyrrole C20H28O3 316.4405 1206.4390 1207.4470 604.2275 403.15433 302.61775 Pyrrole -H2O C20H26O2 298.4252 1188.4237 1189.4317 595.21985 397.14923 298.11392 Lactam C20H28O4 332.4399 1222.4384 1223.4464 612.2272 408.48746 306.6176 Lactam-H2O C20H26O3 314.4246 1204.4231 1205.4311 603.21955 402.48236 302.11377 Hydroxylactam C20H28O5 348.4393 1238.4378 1239.4458 620.2269 413.8206 310.61745 Hydroxylactam-H2O C20H26O4 330.4240 1220.4225 1221.4305 611.21925 407.8155 306.11362 Hydroxylactam-2H2O
C20H24O3 312.4088 1202.4073 1203.4153 602.21165 401.81043 301.60982
Cross-link C20H33O4 337.4796 2114.4526 2115.4606 1058.2343 705.82553 529.62115 Cross-link-H2O C20H31O3 319.4643 2096.4373 2097.4453 1049.2266 699.82043 525.11732 Cross-link-2H2O C20H29O2 301.4491 2078.4221 2079.4301 1040.2190 693.81536 520.61352 Bis-pyrrole C40H58O6 634.8969 2410.8619 2411.8699 1206.4389 804.62863 603.72347 Bis-pyrrole-H2O C40H56O5 616.8816 2392.8466 2393.8546 1197.4313 798.62353 599.21965 Bis-pyrrole-2H2O C40H54O4 598.8664 2374.8314 2375.8394 1188.4237 792.61846 594.71585 Tris-pyrrole C60H88O9 953.3353 3617.2829 3618.2909 1809.6495 1206.7690 905.3287 Tris-pyrrole-H2O C60H86O8 935.3200 3599.2676 3600.2756 1800.6418 1200.7639 900.8249 Tris-pyrrole-2H2O C60H84O7 917.3047 3581.2523 3582.2603 1791.6342 1194.7588 896.3211 Tris-pyrrole-3H2O C60H82O6 899.2894 3563.2370 3564.2450 1782.6265 1188.7537 891.8173 Pyrrole+Lactam C40H58O7 650.8963 2426.8613 2427.8693 1214.4387 809.9618 607.7233 Pyr+Lac-H2O C40H56O6 632.8810 2408.8460 2409.8540 1205.4310 803.9567 603.2195 Pyr+Lac-2H2O C40H54O5 614.8658 2390.8307 2391.8387 1196.4234 797.9516 598.7157
173

EVHHQKL (Monoisotopic molecular weight): 889.476977, (+1) 890.454253, (+2) 445.745765 H:1.0078 H2O:18.0106 Adducts-- no fluorescence tag
Adduct Formula
Adduct MW
Product MW
Plus one charge (+1)
Plus two charges (+2)
Plus three charges (+3)
Plus four charges (+4)
Pyrrole C20H28O3 316.2038 1205.6808 1206.6886 603.8482 402.9014 302.4280 Pyrrole -H2O C20H26O2 298.1933 1187.6703 1188.6781 594.84295 396.8979 297.9254 Lactam C20H28O4 332.1988 1221.6750 1222.6828 611.8453 408.2328 306.4266 Lactam-H2O C20H26O3 314.1882 1203.6652 1204.6730 602.8404 402.2295 301.9241 Hydroxylactam C20H28O5 348.1937 1237.6707 1238.6785 619.84315 413.5647 310.4255 Hydroxylactam-H2O C20H26O4 330.1831 1219.6601 1220.6679 610.83785 407.5612 305.9228 Hydroxylactam-2H2O
C20H24O3 312.1725 1201.6495 1202.6573 601.83255 401.5576 301.4202
Cross-link C20H33O4 337.2379 2113.1685 2114.1763 1057.5921 705.3973 529.2999 Cross-link-H2O C20H31O3 319.2273 2095.1579 2096.1657 1048.5867 699.3938 524.7973 Cross-link-2H2O C20H29O2 301.2168 2077.1473 2078.1551 1039.5814 693.39023 520.2946 Bis-pyrrole C40H58O6 634.4233 2409.3461 2410.3539 1205.6808 804.12316 603.3443 Bis-pyrrole-H2O C40H56O5 616.4128 2391.3355 2392.3433 1196.6755 798.11963 598.8417 Bis-pyrrole-2H2O C40H54O4 598.4022 2373.3249 2374.3327 1187.6702 792.1161 594.3390 Tris-pyrrole C60H88O9 952.6428 3617.9588 3618.9666 1809.9872 1206.9941 905.4975 Tris-pyrrole-H2O C60H86O8 934.6322 3599.9482 3600.9560 1800.9819 1200.9905 900.9948 Tris-pyrrole-2H2O C60H84O7 916.6216 3581.9376 3582.9454 1791.9766 1194.9870 896.4922 Tris-pyrrole-3H2O C60H82O6 898.6110 3563.9270 3564.9348 1782.9713 1188.9835 891.9895 Pyrrole+Lactam C40H58O7 650.4183 2425.3411 2426.3489 1213.6783 809.4548 607.3431 Pyr+Lac-H2O C40H56O6 632.4077 2407.3305 2408.3383 1204.6730 803.4513 602.8404 Pyr+Lac-2H2O C40H54O5 614.3971 2389.3209 2390.3287 1195.6682 797.4481 598.3380
(4) Amyloid (11-17) “EVHHQKL” peptide reaction with iso[4]LGE2-Fluo tag EVHHQKL (Monoisotopic molecular weight): 889.476977, (+1) 890.454253, (+2) 445.745765 H:1.0078 H2O:18.0106
Adducts-- no fluorescence tag
Adduct Formula
Adduct MW
Product MW
Plus one charge (+1)
Plus two charges (+2)
Plus three charges (+3)
Plus four charges (+4)
Pyrrole C30H41N5O5S 583.2828 1472.7598 1473.7676 737.3877 491.9277 369.19775 Pyrrole -H2O C30H39N5O4S 565.2723 1454.7493 1455.7571 728.38245 485.9242 364.69513 Lactam C30H41N5O6S 599,2778 1488.7548 1489.7626 745.3852 497.2594 373.1965 Lactam-H2O C30H39N5O5S 581.2672 1470.7442 1471.7520 736.3799 491.2559 368.69385 Hydroxylactam C30H41N5O7S 615.2727 1504.7497 1505.7575 753.38265 502.5910 377.19523 Hydroxylactam-H2O C30H39N5O6S 597.2621 1486.7391 1487.7469 744.37735 496.5875 372.69258 Hydroxylactam-2H2O
C30H37N5O5S 579.2515 1468.7285 1469.7363 735.37502 490.5840 368.1899
Cross-link C30H46N5O6S 604.3168 3266.6997 2381.2553 1191.1315 1089.9077 817.6827 Cross-link-H2O C30H44N5O5S 586.3062 3248.6891 2363.2447 1182.1262 1083.9042 813.1801 Cross-link-2H2O C30H42N5O4S 568.2956 3230.6785 2345.2341 1173.1209 1077.9006 808.6774 Bis-pyrrole C60H84N10O10S2 1168.5813 3830.9642 2944.5119 1472.7598 1277.9959 958.7489 Bis-pyrrole-H2O C60H82N10O9S2 1150.5707 3812.9536 2926.5013 1463.7545 1271.9923 954.2462 Bis-pyrrole-2H2O C60H80N10O8S2 1132.5601 3794.9430 2908.4907 1454.7492 1265.9888 949.7436
174

Figure 3.1S Acetyl-Gly-Lys-O-Methyl ester reactions with iso[4]LGE2 in different molar ratio and time point
(1) Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (Dipeptide: Iso4 = 2000:1 molar ratio) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 8 days, 15 days, 21 days and 50 days.
Modified One Adducts
Crosslink
Bispyrrole Trispyrrole
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 1 day
Trispyrrole
-H2O-2H2O
-3H2O
A
Crosslink
Modified One Adducts
BispyrroleTrispyrrole
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 4 days
B
175

Trispyrrole
Trispyrrole
Bispyrrole
Crosslink
Modified One Adducts Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (2000:1) in
50 mM NEM OAc pH 8.6 for 8 days-H2O
-2H2O
-3H2O
C
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 15 days
Modified One Adducts
Crosslink
Bispyrrole Trispyrrole
Bispyrrole
-H2O
-2H2O
+16
+16+16
+16
D
176

TrispyrroleBispyrrole
Crosslink
Modified One Adducts Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (2000:1) in
50 mM NEM OAc pH 8.6 for 21 days
Bispyrrole
-H2O
-2H2O
+16+16
+16
+16
E
TrispyrroleBispyrroleCrosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 50 days
Trispyrrole-2H2O -H2O
-3H2O
+16
+16
+16
F
177

(2) Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (Dipeptide: Iso4 = 1000:1 molar ratio) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 8 days, 15 days, 21 days and 50 days.
TrispyrroleBispyrrole
Crosslink
Modified One Adducts Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (1000:1)
in 50 mM NEM OAc pH 8.6 for 1 day
Trispyrrole-H2O-2H2O-3H2O
A
Crosslink
BispyrroleTrispyrrole
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (1000:1) in 50 mM NEM OAc pH 8.6 for 4 days
Modified One Adducts
B
178

Crosslink
BispyrroleTrispyrrole
Trispyrrole-2H2O-H2O
-3H2O
Modified One Adducts Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (1000:1) in 50 mM
NEM OAc pH 8.6 for 8 daysC
TrispyrroleBispyrrole
Crosslink
Modified One Adducts Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (1000:1) in 50 mM
NEM OAc pH 8.6 for 15 daysBispyrrole
-H2O
-2H2O
+16
+16+16
+16
D
179

TrispyrroleBispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (1000:1) in 50 mM NEM OAc pH 8.6 for 21 days
Bispyrrole
-H2O
-2H2O
+16+16
+16
+16
+16
E
Bispyrrole
Trispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (1000:1) in 50 mM NEM OAc pH 8.6 for 50 days
Trispyrrole-H2O-2H2O
-3H2O+16
+16
+16
F
180

(3) Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (Dipeptide: Iso4 = 200:1 molar ratio) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 8 days, 15 days, 21 days and 50 days.
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 1 day
Bispyrrole Trispyrrole
Crosslink
Modified One Adducts
Trispyrrole-H2O-2H2O
-3H2O
+16 +16
A
TrispyrroleBispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 4 days Bispyrrole
-H2O
-2H2O
+16
+16
+16
+16
B
181

Trispyrrole
Trispyrrole
Bispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with Iso[4] no fluorescence tag (200:1) in 50 mM NEM OAc pH 8.6 for 8 days
-H2O
-2H2O
-3H2O
+16
+16
+16
+16
C
Trispyrrole
Bispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with Iso[4] no fluorescence tag (200:1) in 50 mM NEM OAc pH 8.6 for 15 days
Bispyrrole
-2H2O
-H2O
+16
+16
+16
+16
D
182

TrispyrroleBispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with Iso[4] no fluorescence tag (200:1) in 50 mM NEM OAc pH 8.6 for 21 days
Bispyrrole
-H2O
-2H2O
+16
+16
+16
+16
+16
E
Acetyl-Gly-Lys-O-Methyl ester reacts with Iso[4] no fluorescence tag (200:1) in 50 mM NEM OAc pH 8.6 for 50 days
Modified One Adducts
Crosslink
Bispyrrole Trispyrrole
Trispyrrole
+16
+16
+16
-H2O-2H2O
-3H2O
F
183

Crosslink
-H2O
-2H2O
Pyrrole-H2O
Lactam-H2O
HydroxyLactam-H2O
-2H2O
A B
C D
Figure 3.2S ESI-MSMS of molecular ions at A) aminal cross-link, B) pyrrole, C) lactam and D) hydroxylactam corresponding to a iso[4]LGE2–(N-acetyl-Gly-Lys-OMe) adduct.
184

NH
O HN COOCH3
NH2O
+C
O
C5H11
OO
OH
Crosslink N C
NH
C5H11
OHO
OH
P
P
-H2O
MW 853.5(+1)
MW 835.5(+1)
MW 558.4(+1)Like Pyrrole-H2O
MW 436.3(+1)
Minus MW 122
N C OHO
P C5H11+
MS/MS
MS/MSMW 853.5(+1)835.5(+1)817.5(+1)
(-H2O)(-2H2O)
MW 473.3(+1)HN C
NH
C5H11
OHO
OHP
N C
NH
C5H11
OHO
P
P
N CC5H11
O
P
N C
NH
C5H11
OH
OHO
OH
P
P
OH OH
OH OH
Pyrrole MW 576.4(+1)-H2O MW 558.4(+1)
Lactam MW 592.4(+1)N C
C5H11
OHO
P MW 475.4(+1)Minus MW 99-H2O MW 574.4(+1)
O
MW 415.4(+1)Minus MW 60
HydroxyLactam 608.4(+1)N C
C5H11
OHO
P
O-2H2O MW 572.4(+1)-H2O MW 590.4(+1)
NH
OHN COOCH3
NH2O
+ C
O
C5H11
OO
OH
MW 436.3(+1)Minus MW 122
N C OHO
PC5H11+
N CC5H11
OHO
P
OH
MW 473.4(+1)Minus MW 99 MW 413.4(+1)Minus MW 60
One Site Modification
Scheme 3.1S Possible fragmentation of (N-acetyl-Gly-Lys-OMe)-iso[4]LGE2 adducts.
185

Figure 3.3S Acetyl-Gly-Lys-O-Methyl ester reactions with iso[4]LGE2-Fluo tag in different molar ratio and time point
(1) Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (Dipeptide : Iso4 = 2000:1) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 15 days, 21 days and 50 days.
Bispyrrole
Crosslink
Modified One Adducts Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag
(2000:1) in 50 mM NEM OAc pH 8.6 for 1 day
Bispyrrole
-H2O-2H2O
A
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (2000:1) in 50 mM NEM OAc pH 8.6 for 4 days
Modified One Adducts
Bispyrrole
Crosslink
B
186

Bispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (2000:1) in 50 mM NEM OAc pH 8.6 for 15 days Crosslink
-H2O
-2H2O
C
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (2000:1) in 50 mM NEMOAc pH 8.6 for 21 days
Crosslink
-H2O
-2H2O
D
187

Modified One Adducts
Crosslink
Crosslink
-H2O
-2H2O
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (2000:1) in 50 mM NEMOAc pH 8.6 for 50 days
E
(2) Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (Dipeptide : Iso4 = 200:1) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 15 days, 21 days and 50 days.
Crosslink
Modified One Adducts
Bispyrrole
Bispyrrole-H2O-2H2O
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (200:1) in 50 mM NEMOAc pH 8.6 for 1 day
A
188

Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (200:1) in 50 mM NEMOAc pH 8.6 for 4 days
Bispyrrole
Crosslink
Modified One Adducts
B
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (200:1) in 50 mM NEMOAc pH 8.6 for 15 days
Modified One Adducts
Crosslink
Crosslink
-H2O
-2H2O
C
189

Bispyrrole
Crosslink
Modified One Adducts
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (200:1) in 50 mM NEMOAc pH 8.6 for 21 days
Crosslink
-2H2O
-H2O
D
Crosslink
Modified One Adducts Crosslink
Acetyl-Gly-Lys-O-Methyl ester reacts with iso[4]LGE2-Fluo tag (200:1) in 50 mM NEMOAc pH 8.6 for 50 days
-H2O
-2H2O
E
190

HydroxyLactam-2H2O
HydroxyLactam-H2O
HydroxyLactam
Lactam-H2O
Lactam
Pyrrole-H2O
Pyrrole
Crosslink
-H2O
-2H2O
A B
C D
Figure 3.4S: ESI-MSMS of molecular ions at A) animal cross-link, B) pyrrole, C) lactam and D) hydroxylactam corresponding to a Iso[4]LGE2-FluoTag - (N-acetyl-Gly-Lys-OMe) adduct.
191

NH
O HN COOCH3
NH2O
+ C
O
C5H11
O
NHCH2CH2NHO
NO
N
SO2N
OH
CrosslinkN C
NH
C5H11
OH
FluoTagO
OH
P
P
-H2OMW 1120.6(+1) MW 1102.6(+1)
MW 825.5(+1)Like Pyrrole-H2O 703.4(+1)
-MW 122 N C FluoTagO
P
C5H11+
MS/MS
N C
NH
C5H11
OH
FluoTagO
P
P
N CC5H11
FluoTagO
P
MS/MSMW 1120.6(+1)
1102.6(+1)
1084.6(+1)
(-H2O)
(-2H2O)
N C
NH
C5H11
OH
FluoTag
O
OH
P
P
740.5(+1)HN C
NH
C5H11
FluoTag
O
OHP
Pyrrole MW 843.4(+1)
Pyrrole-H2O MW 825.4(+1)
NH
O HN COOCH3
NH2O
+ C
O
C5H11
O
NHCH2CH2NHO
NO
N
SO2N
OH
N CC5H11
FluoTagO
P
Lactam MW 859.4(+1) N CC5H11
FluoTagO
P 742.4(+1)-MW 99
Lactam-H2O MW 841.4(+1)O
682.4(+1) -MW 60
HydroxyLactam 875.4(+1) N CC5H11
FluoTagO
P
OHydroxyLactam-2H2O 839.4(+1)HydroxyLactam-H2O 857.4(+1) 740.4(+1)
-MW 99680.4(+1)
-MW 60
703.4(+1)-MW 122 N C FluoTag
O
P
C5H11+
Scheme 3.2S Possible fragmentation of (N-acetyl-Gly-Lys-OMe)-iso[4]LGE2-FluoTag adducts.
192

Figure 3.5S Amyloid (11-17) “EVHHQKL” peptide reactions with iso[4]LGE2 in different molar ratio and time point
(1) Amyloid (11-17) “EVHHQKL” reacts with iso[4]LGE2 (Peptide: Iso4 = 2000:1) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 15 days, 21 days and 50 days.
Modified One Adducts
Crosslink Bispyrrole Trispyrrole
Bispyrrole
-2H2O-H2O
Amyloid (11-17) "EVHHQKL" reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 1 day
+16
A
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 4 days
Crosslink Bispyrrole Trispyrrole
Bispyrrole
-H2O
-2H2O+16
B
193

Bispyrrole
Bispyrrole
TrispyrroleCrosslink
Amyloid (11-17) "EVHHQKL" reacts with iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 15 days
-H2O
-2H2O+16
+16
Modified One Adducts C
Bispyrrole TrispyrroleCrosslink
Modified One Adducts Amyloid (11-17) "EVHHQKL" reacts with
iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 21 days
Bispyrrole
-H2O
-2H2O
+16
+16
+16
D
194

Crosslink
Bispyrrole
Bispyrrole Trispyrrole
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2 (2000:1) in 50 mM NEM OAc pH 8.6 for 50 days
-H2O
-2H2O
+16
+16
+16+16
E
(2) Amyloid (11-17) “EVHHQKL” reacts with iso[4]LGE2 (Peptide: Iso4 = 200:1) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 7 days, 15 days, 21 days and 50 days.
Crosslink Bispyrrole Trispyrrole
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 1 day
Bispyrrole
-H2O
-2H2O
+16
+16
A
195

TrispyrroleBispyrroleCrosslink
Modified One Adducts Amyloid (11-17) "EVHHQKL" reacts
with iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 4 days
Bispyrrole
-H2O
-2H2O
+16
B
-2H2OBispyrrole
+16
-H2OAmyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 7 days
Modified One Adducts
Bispyrrole TrispyrroleCrosslink
C
196

Crosslink Bispyrrole Trispyrrole
Bispyrrole
-H2O
-2H2O +16
Modified One Adducts Amyloid (11-17) "EVHHQKL" reacts
with iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 15 days
Bispyrrole
D
Crosslink Bispyrrole Trispyrrole
Modified One Adducts Amyloid (11-17) "EVHHQKL" reacts
with iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 21days
Bispyrrole
-2H2O
-H2O
+16
+16
E
197

-2H2O
-H2O
Bispyrrole
+16
+16
+16
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2 (200:1) in 50 mM NEM OAc pH 8.6 for 50 days
Modified One Adducts
Crosslink Bispyrrole Trispyrrole
F
Figure 3.6S Amyloid (11-17) “EVHHQKL” peptide reactions with iso[4]LGE2-Fluo tag in different molar ratio and time point
(1) Amyloid (11-17) “EVHHQKL” reacts with iso[4]LGE2-Fluo tag (Peptide: Iso4 = 2000:1) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 7 days, 15 days, 21 days and 50 days.
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (2000:1)in 50 mM NEM OAc pH 8.6 for 1 day
Modified One Adducts
No Obvious crosslink or bispyrrole peak formed
A
198

Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (2000:1)in 50 mM NEM OAc pH 8.6 for 4 days
Crosslink
Crosslink-H2OB
Crosslink
Crosslink
-H2O
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (2000:1)in 50 mM NEM OAc pH 8.6 for 7 days
Modified One Adducts
C
199

Crosslink
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (2000:1)in 50 mM NEM OAc pH 8.6 for 15 days
Crosslink
-H2O
-2H2O
D
Crosslink
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (2000:1)in 50 mM NEM OAc pH 8.6 for 21 days
Crosslink
-H2O
E
200
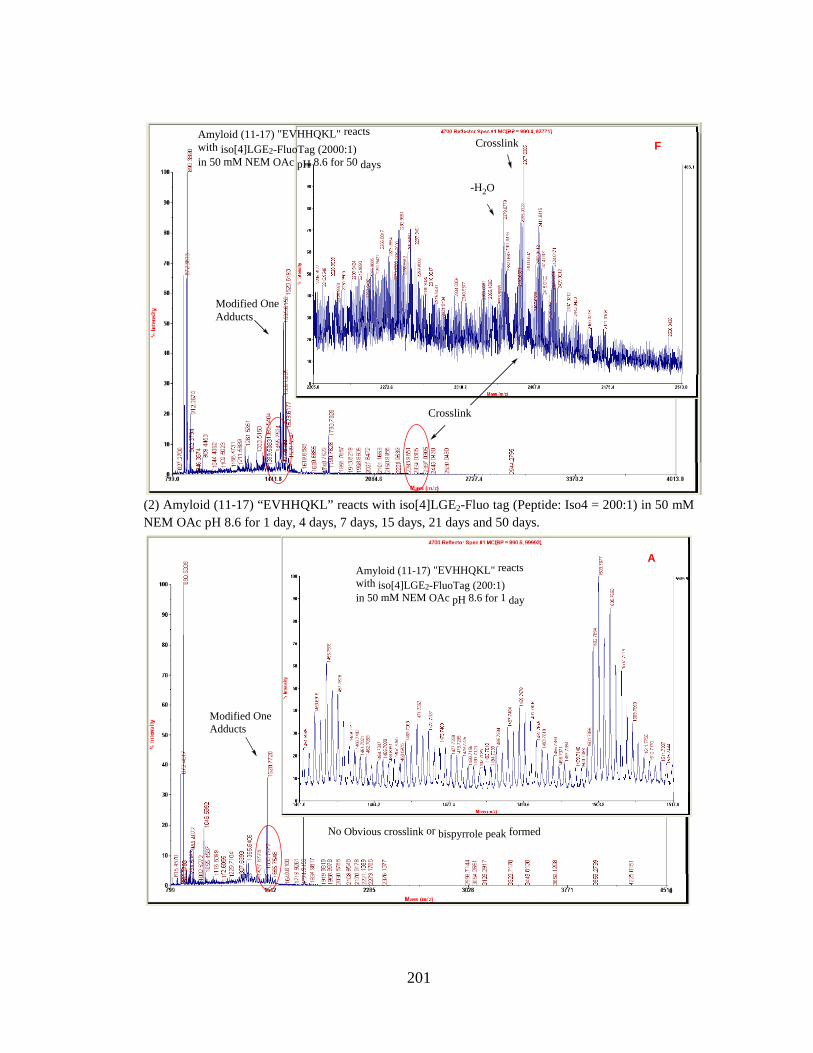
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (2000:1)in 50 mM NEM OAc pH 8.6 for 50 days
Modified One Adducts
Crosslink
Crosslink
-H2O
F
(2) Amyloid (11-17) “EVHHQKL” reacts with iso[4]LGE2-Fluo tag (Peptide: Iso4 = 200:1) in 50 mM NEM OAc pH 8.6 for 1 day, 4 days, 7 days, 15 days, 21 days and 50 days.
Modified One Adducts
No Obvious crosslink or bispyrrole peak formed
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (200:1)in 50 mM NEM OAc pH 8.6 for 1 day
A
201

Modified One Adducts
Crosslink
Crosslink
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (200:1)in 50 mM NEM OAc pH 8.6 for 4 days
B
Crosslink
-H2O
Crosslink
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (200:1)in 50 mM NEM OAc pH 8.6 for 7 days
C
202

Crosslink
Crosslink
-H2O
Modified One Adducts
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (200:1)in 50 mM NEM OAc pH 8.6 for 15 days
D
Crosslink-H2O
Modified One Adducts
Crosslink
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (200:1)in 50 mM NEM OAc pH 8.6 for 21 days
E
203
Amyloid (11-17) "EVHHQKL" reactswith iso[4]LGE2-FluoTag (200:1)in 50 mM NEM OAc pH 8.6 for 50 days
Modified One Adducts
Crosslink
-H2O
Crosslink
F
UPLC purified pyrrole
No oxidized crosslink and bispyrrole
Pyrrole-H2O
NO Acetyl-Gly-Lys-O-Methyl ester and Iso[4]
Pyrrole-H2O
Figure 4.1S MALDI-TOF spectrums of iso[4]LGE2-pyrrole
204

Iso[4]LGE2 purified with C18 toptip Acetyl-Gly-Lys-O-Methyl ester
Figure 4.2S MALDI-TOF spectrums of iso[4]LGE2 and Acetyl-Gly-Lys-O-methyl ester ---- indicating new peaks besides the matrix.
No oxidized crosslink and bispyrrole
NO GK didpeptide and Iso[4]UPLC purified pyrrolePyrrole-H2O Pyrrole-H2O After 2 days reaction
under Air
No crosslink formed, a little bispyrrole formed
Pyrrole-H2O After 6 days reaction under Air
No crosslink formed, some bispyrrole formed
More than 8 days incubationPyrrole-H2O
Bispyrrole peaks
Lactam
Hydroxylactam
Figure 4.3S MALDI-TOF spectrums of HPLC-purified iso[4]LGE2-pyrrole reaction under air within 8 days.
205

Pyrrole-H2OPure Iso[4]LGE2-pyrrole was added Ac-Lysineand reacted for 2 days.Iso[4]LGE2-pyrrole didn't oxidize through this way.It didn't relate with molar ratio and pH.
No oxidized products were formed
Figure 4.4S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with Nε-acetyl-L-lysine under air for 2 days.
Pyrrole-H2O Pure Iso[4]LGE2-pyrrole was addedL-Lysine (1:10 mlolar ratio),reacted under air for 2 days.
No oxidized products were formed
Figure 4.5S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with L-
lysine under air for 2 days.
206

Pyrrole-H2O
Amyloid peptide
Pure pyrrole was added Amyloid peptide in H2O (1:10 mlolar ratio), reacted under air for 6 h.No other oxidized peak formedNo Lactam or Hydrolactam formed
Pyrrole-H2OAmyloid peptide Pyrrole-H2O
Amyloid peptide
Pure pyrrole was added Amyloid peptide in H2O (1:10 mlolar ratio), reacted under air overnight.No other oxidized peak formedNo Lactam or Hydrolactam formed
Pure pyrrole was added Amyloid peptide in H2O (1:10 mlolar ratio), reacted under air for 2 days.No other oxidized peak formedNo Lactam or Hydrolactam formed
Figure 4.6S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with β-Amyloid peptide under air for 2 days.
Pyrrole-H2O
The peak 544 was shown up after adding Ethanolamine.
Pure pyrrole was added ethanolamine equal ratio and reacted for 2 days.Pyrrole decreased through this way.pH=7.
Pure pyrrole was added ethanolamine much more excess and reacted for 2 h.Pyrrole couldn't be observed.pH=7.
The peak 544 was shown up after adding Ethanolamine.
No pyrrole-H2O
Figure 4.7S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with ethanolamine under air.
207

Pure pyrrole was added GK Dipeptide----Reacted 1h under Air
Pyrrole-H2O
Pure pyrrole was added GK Dipeptide----Reacted 8h under Air
Hydroxylactam-2H2O
Pyrrole-H2O
Hydroxylactam-2H2OPure pyrrole was added GK Dipeptide----Reacted 1 day under Air
Pyrrole-H2O
Pyrrole-H2O
Hydroxylactam-2H2O Pure pyrrole was added GK Dipeptide----Reacted 2 days under Air
GK dipeptide in H2O GK dipeptide in H2O
GK dipeptide in H2OGK dipeptide in H2O
(From BACHEM) (From BACHEM)
(From BACHEM)(From BACHEM)
Hydroxylactam-2H2O
Figure 4.8S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with Acetyl-Gly-Lys-O-Methyl Ester under air.
Pyrrole-H2O Pyrrole-H2O
Pure pyrrole was addedUPLC purified GK dipeptide--reacted under air for 2 h.
Pure pyrrole was addedUPLC purified GK dipeptide--reacted under air for 6 h.
No other oxidized product formed No other oxidized product formedNo Lactam or Hydroxylactam formed No Lactam or Hydroxylactam formed
Pyrrole-H2O Pure pyrrole was addedUPLC purified GK dipeptide--reacted under air for 2 days.
No Lactam or Hydroxylactam formedNo other oxidized product formed
Pyrrole-H2O Pure pyrrole was addedUPLC purified GK dipeptide--reacted under air for 6 days.
No Lactam or Hydroxylactam formedNo other oxidized product formed
Formed Bispyrrole
Figure 4.9S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with HPLC-purified Acetyl-Gly-Lys-O-Methyl Ester under air.
208

Table 4.1S Summaries of HPLC-purified iso[4]LGE2-pyrrole reaction with different chemicals.
(a) HPLC-purified iso[4]LGE2-pyrrole reactions with Acetyl-Gly-Lys-O-Methyl Ester and Initiator (2,2'-Azobis(2-methylpropionamidine) dihydrochloride)
Molar ratio 2 h reaction Overnight reaction A week reaction Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =1:0.1
Pyrrole—no changes Pyrrole—decreased little bit, very little oxidized product formed
Almost no pyrrole, little oxidized product formed
Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =1:1
Pyrrole—decreased little bit, little oxidized product formed
Pyrrole—decreased little bit, more oxidized product formed
Almost no pyrrole, much oxidized products formed
Pyrrole: Initiator=1:0.1 Pyrrole—decreased, no oxidized product formed
No pyrrole or oxidized product
No pyrrole or oxidized product
Pyrrole: Initiator=1:1 No pyrrole or oxidized product
No pyrrole or oxidized product
No pyrrole or oxidized product
Molar ratio 2 h reaction Overnight reaction 2 days reaction Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =1:10
Pyrrole—decreased little bit, much oxidized product formed
Pyrrole—decreased little bit, more oxidized product formed
Pyrrole—decreased, much more oxidized products formed
Pyrrole: Initiator=1:0.01 Pyrrole—no change, no oxidized product formed
Pyrrole— decreased, no oxidized product formed
Pyrrole— decreased, no oxidized product formed
Molar ratio 4 h reaction Overnight reaction Pyrrole: Initiator=1:0.05 Very little pyrrole left and no oxidation No pyrrole left and no oxidation
(b) HPLC-purified iso[4]LGE2-pyrrole reactions with Vitamin C, Acetyl-Gly-Lys-O-Methyl Ester and Initiator (2,2'-Azobis(2-methylpropionamidine) dihydrochloride)
Molar ratio 2 h reaction Overnight reaction A week reaction Vitamin C: Pyrrole=0.1:1 Pyrrole—no changes
No oxidized product formed No pyrrole No oxidized product formed
No pyrrole and formed a peak (+670), no other oxidized products detected
Vitamin C: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =0.1:1:0.1
Pyrrole—no changes No oxidized product formed
Pyrrole—decreased little bit, very little oxidized product formed
Pyrrole—decreased little bit, very little oxidized product formed. Also formed peak (+670)
Vitamin C: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =0.1:1:1
Pyrrole—decreased little bit, little oxidized product formed
Pyrrole—decreased little bit, more oxidized product formed
Pyrrole—decreased little bit, much more oxidized product formed. Also formed peak (+670)
Vitamin C: Pyrrole: Initiator=0.1:1:0.1
Pyrrole—no changes No oxidized product formed
No pyrrole No oxidized product formed
No pyrrole, no oxidized product formed. Also formed peak (+670)
Vitamin C: Pyrrole: Initiator=0.1:1:1
Pyrrole—no changes No oxidized product formed
No pyrrole No oxidized product formed
No pyrrole, no oxidized product formed. Also formed peak (+670), and some other unknown peaks
Molar ratio 2 h reaction Overnight reaction 2 days reaction Vitamin C: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =0.01:1:10
Pyrrole—decreased little bit, much oxidized product formed
Pyrrole—decreased, more oxidized product formed
Pyrrole—decreased, much more oxidized products formed
Vitamin C: Pyrrole: Initiator=0.01:1:0.01
Pyrrole—no changes. No oxidized product formed
Pyrrole— decreased. No oxidized product formed
Pyrrole— decreased. No oxidized product formed
(c) HPLC-purified iso[4]LGE2-pyrrole reactions with Vitamin E, Acetyl-Gly-Lys-O-Methyl
Ester and Initiator (2,2'-Azobis(2-methylpropionamidine) dihydrochloride) Molar ratio 2 h reaction Overnight reaction A week reaction Vitamin E: Pyrrole=0.1:1 Pyrrole—no changes
No oxidized product formed No pyrrole or oxidized product
No pyrrole or oxidized product
209

Vitamin E: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =0.1:1:0.1
Pyrrole—no changes No oxidized product formed
No pyrrole, formed very little oxidized products
No pyrrole, some known peaks
Vitamin E: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =0.1:1:1
Pyrrole—decreased little bit, little oxidized product formed
Pyrrole—decreased, little oxidized product formed
No pyrrole, some known peaks
Vitamin E: Pyrrole: Initiator=0.1:1:0.1
Pyrrole—decreased, No oxidized product formed
No pyrrole or oxidized product
No pyrrole or oxidized product
Vitamin E: Pyrrole: Initiator=0.1:1:1
Pyrrole—decreased, No oxidized product formed
No pyrrole or oxidized product
No pyrrole or oxidized product
Molar ratio 2 h reaction Overnight reaction 2 days reaction Pyrrrole: Vitamin E=1: 0.01 Pyrrole—no changes. No
oxidized product formed Pyrrole— decreased. No oxidized product formed
No pyrrole left, no oxidized products and some unknown peaks
Molar ratio 4 h reaction Overnight reaction Pyrrole: Vitamin E:initiator =1: 0.01:0.01
No pyrrole left and no oxidation Pyrrole— decreased and no oxidation
(d) HPLC-purified iso[4]LGE2-pyrrole reactions with H atom donor ((R)-(-)-2-(2,5-
Dihydrophenyl)glycine), Acetyl-Gly-Lys-O-Methyl Ester and Initiator (2,2'-Azobis(2-methylpropionamidine) dihydrochloride)
Molar ratio 2 h reaction Overnight reaction A week reaction H atom Donor: Pyrrole=5:1 No pyrrole or oxidized
product No pyrrole or oxidized product
No pyrrole or oxidized product
H atom Donor: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =5:1:0.1
No pyrrole or oxidized product
No pyrrole or oxidized product
No pyrrole or oxidized product
H atom Donor: Pyrrole: Acetyl-Gly-Lys-O-Methyl Ester =5:1:1
No pyrrole, little oxidized products formed
No pyrrole, little oxidized products formed
No pyrrole, some known peaks
H atom Donor: Pyrrole: Iniitator=5:1:0.1
No pyrrole or oxidized product
No pyrrole or oxidized product
No pyrrole or oxidized product
H atom Donor: Pyrrole: Iniitator=5:1:1
No pyrrole or oxidized product
No pyrrole or oxidized product
No pyrrole or oxidized product
Molar ratio 2 h reaction Overnight reaction 2 days reaction Pyrrrole: H atom dondor =5:1
Pyrrole—no changes. No oxidized product formed
Pyrrole—no changes. No oxidized product formed
Pyrrole—no changes. No oxidized product formed
Molar ratio 4 h reaction Overnight reaction Pyrrole: H atom Donor =1:1 Very little pyrrole left and no oxidation No pyrrole left and no oxidation Pyrrole:H atom Donor: Acetyl-Gly-Lys-O-Methyl Ester =2:1:1
Pyrrole—decreased, some oxidized product formed similar like pyrrole with Acetyl-Gly-Lys-O-Methyl Ester
(e) HPLC-purified iso[4]LGE2-pyrrole reactions with UV, rose bengal, methylene blue,
Co2(CO)8, H2O2 K2CO3, dimethyldioirane, microwave, Di-tert-butyl peroxide, riboflavin (VB2), K3Fe(CN)6, FeCl3, K2S2O8.
Molar ratio 30 min 1 h reaction 2 h reaction Pyrrole: rose bengal=1:1, UV, air No pyrrole or oxidized
product No pyrrole or oxidized product
No pyrrole or oxidized product
Pyrrole: methylene blue =1:1, UV, air
No pyrrole or oxidized product
No pyrrole or oxidized product
No pyrrole or oxidized product
UV light and air No pyrrole or oxidized product
No pyrrole or oxidized product
No pyrrole or oxidized product
Condition 2 min 8 min UV light and air Pyrrole—decreased. No oxidized product Pyrrole—decreased. No oxidized product
210
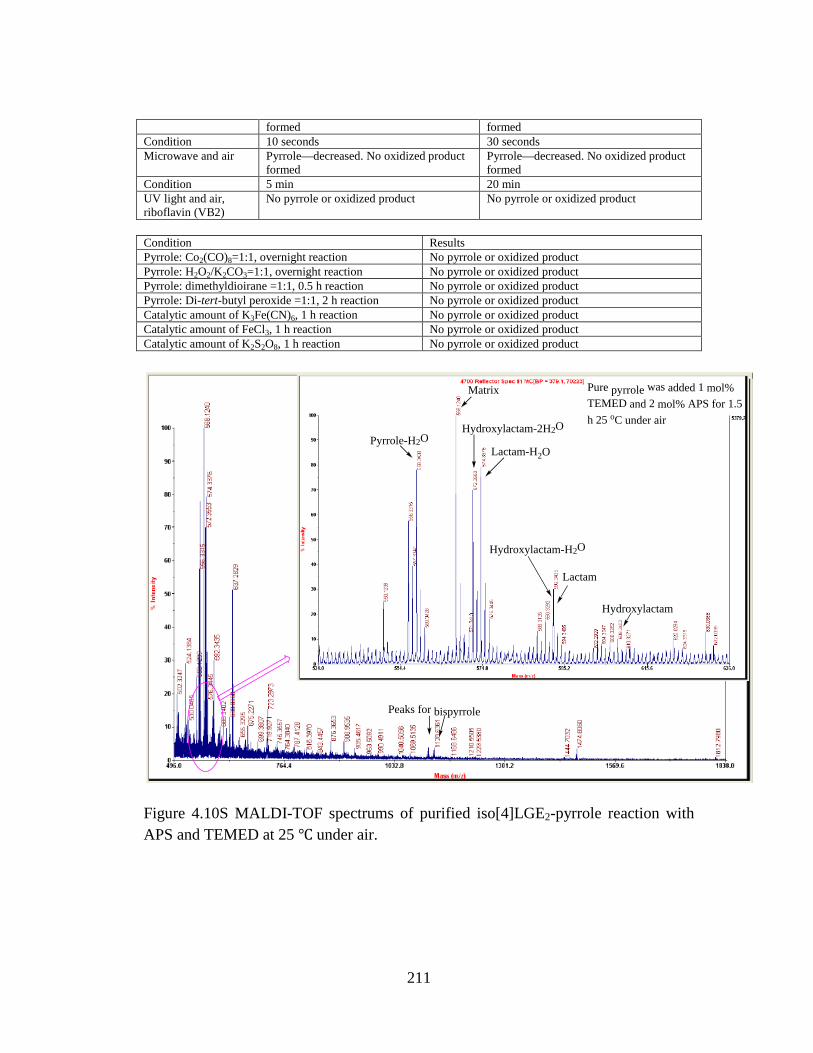
formed formed Condition 10 seconds 30 seconds Microwave and air Pyrrole—decreased. No oxidized product
formed Pyrrole—decreased. No oxidized product formed
Condition 5 min 20 min UV light and air, riboflavin (VB2)
No pyrrole or oxidized product No pyrrole or oxidized product
Condition Results Pyrrole: Co2(CO)8=1:1, overnight reaction No pyrrole or oxidized product Pyrrole: H2O2/K2CO3=1:1, overnight reaction No pyrrole or oxidized product Pyrrole: dimethyldioirane =1:1, 0.5 h reaction No pyrrole or oxidized product Pyrrole: Di-tert-butyl peroxide =1:1, 2 h reaction No pyrrole or oxidized product Catalytic amount of K3Fe(CN)6, 1 h reaction No pyrrole or oxidized product Catalytic amount of FeCl3, 1 h reaction No pyrrole or oxidized product Catalytic amount of K2S2O8, 1 h reaction No pyrrole or oxidized product
Peaks for bispyrrole
Pyrrole-H2OLactam-H2O
Hydroxylactam-2H2O
Hydroxylactam-H2O
Lactam
Hydroxylactam
Matrix Pure pyrrole was added 1 mol% TEMED and 2 mol% APS for 1.5 h 25 oC under air
Figure 4.10S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with APS and TEMED at 25 ℃ under air.
211

Bispyrrole
Hydrolactam-2H2O
Pyrrole-H2O Pyrrole-H2O
Hydrolactam-2H2O
Pure pyrrole was added 1 mol% TEMED and 1 mol% APS 37 degree for 1h
Lactam-H2O
Hydrolactam-H2OHydrolactam
Pyrrole-H2O Pyrrole-H2O
Hydrolactam-2H2O
Hydrolactam-2H2O
Lactam-H2O
Hydrolactam-H2O
Hydrolactam
Bispyrrole
Pure pyrrole was added 1 mol% TEMED and 1 mol% APS 37 degree for 2 h
Figure 4.11S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with APS and TEMED at 37 ℃ under air.
212

Bispyrrole
Pyrrole-H2O Pyrrole-H2O Pure pyrrole was added 1 mMTMAO in H2O and reacted 37 degree 3hRatio as Pyrrole:TMAO=1:1
Lactam-H2O
Hydroxylactam-2H2O
Hydroxylactam-H2O
Lactam
Hydroxylactam
Figure 4.12S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with 1 mM TMAO at 37 ℃ 3 h.
Bispyrrole
Pyrrole-H2O
Pyrrole-H2O
Pure pyrrole was added 1 mMTMAO in H2O, 1mM EDTAin H2O and reacted 37 degree 3hRatio as Pyrrole:TMAO:EDTA=1:0.1:0.1
Hydroxylactam-H2O
Lactam
Hydroxylactam
Lactam-H2O
Hydroxylactam-2H2O
Figure 4.13S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with 1 mM TMAO/EDTA at 37 ℃ 3 h.
213

Pure pyrrole was added 1mM TEMPO in 10%ACN and reacted 37 degree 2 hRatio as Pyrrole:TEMPO=1:1
Bispyrrole
Pyrrole-H2O
Lactam-H2O
Hydroxylactam-2H2O
Lactam
Hydroxylactam-H2O Hydroxylactam
Matrix
Pyrrole-H2O
Figure 4.14S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with 1 mM TEMPO at 37 ℃ 2 h.
Bispyrrole
Pyrrole-H2O
Pyrrole-H2O
Lactam-H2OHydroxylactam-2H2O
Hydroxylactam-H2O
Lactam
Hydroxylactam
Pure pyrrole was added 1 mMTEMPO in 10%ACN, 1 mM EDTAin H2O and reacted 37 degree 2hRatio as Pyrrole:TEMPO:EDTA=1:1:1
Figure 4.15S MALDI-TOF spectrums of purified iso[4]LGE2-pyrrole reaction with 1 mM TEMPO/EDTA at 37 ℃ 2 h.
214

10ul 100mM 3-Ethyl-2,4-dimethylpyrrole in ACN +10ul ACN 25 degree for 3h
Dimer
Trimer
HN
H3C
HN
CH3
CH3
CH3CH3
CH3
HN
H3C
HN
CH3
CH3CH3
CH3
HN
CH3
CH3
CH3
10ul 100mM 3-Ethyl-2,4-dimethylpyrrole in ACN +10ul 100mM TEMPO in ACN 25 degree for 3h
DimerTrimer
HN
H3C
HN
CH3
CH3
CH3CH3
CH3 HN
H3C
HN
CH3
CH3CH3
CH3
HN
CH3
CH3
CH3
215

10ul 100mM 3-Ethyl-2,4-dimethylpyrrole in ACN +10ul 100mM TMAO in ACN 25 degree for 3h
Dimer
HN
H3C
HN
CH3
CH3
CH3CH3
CH3
Trimer
HN
H3C
HN
CH3
CH3CH3
CH3
HN
CH3
CH3
CH3
Figure 4.16S MALDI-TOF spectrums of 3-ethyl-2,4-dimethylpyrrole initiate reactions.
216

Pure 3-Ethyl-2,4-dimethylPyrrole -- Staring material
Figure 4.17S: NMR spectrums of 3-ethyl-2,4-dimethylpyrrole and 3-ethyl-1,2,4-trimethylpyrrole.
217

3-ethyl-1,2,4-trimethyl-1H-pyrrolein ACN for 2 h at R T
N
H3C CH3
CH3
Dimer
Trimer
3-ethyl-1,2,4-trimethyl-1H-pyrrolein ACN for 24 h at R T
N
H3C CH3
CH3
Dimer
Trimer
218

3-ethyl-1,2,4-trimethyl-1H-pyrrolein ACN was added TEMPO for 2 h at R T
N
H3C CH3
CH3Dimer
Trimer
3-ethyl-1,2,4-trimethyl-1H-pyrrolein ACN was added TEMPO for 24 h at R T
N
H3C CH3
CH3Dimer
Trimer
Figure 4.18S MALDI-TOF spectrums of 3-ethyl-1,2,4-trimethylpyrrole initiate reactions.
219

1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 1000.0
0.5
1.0
1.5
2.0
2.5
3.0
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00429 0.00493
Adj. R-Square 0.9965 0.5946Value Standard Error
Iso4-HSA
A1 2.73774 0.03437A2 0.3593 0.03772x0 0.04611 0.00465p 1.06258 0.09919EC20 0.01251 --EC50 0.04611 0.00465EC80 0.16997 --
LGE2-BSA
A1 2.00993 0.04965A2 2.18793 0.02654x0 4.38542E-4 9.1443E11p 42.5833 1.36294E17EC20 4.24495E-4 --EC50 4.38542E-4 9.1443E11EC80 4.53053E-4 --
Iso[4]LGE2 Immunized Mouse Serum (Num 968)
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3 Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00562 0.03004
Adj. R-Square 0.99571 0.75778Value Standard Error
Iso4-HSA
A1 2.61284 0.04279A2 0.1448 0.04066x0 0.02219 0.00252p 1.02856 0.10652EC20 0.00576 --EC50 0.02219 0.00252EC80 0.0854 --
LGE2-BSA
A1 1.68854 0.07977A2 2.34502 0.09011x0 0.06851 0.05174p 1.80698 1.60749EC20 0.03181 --EC50 0.06851 0.05174EC80 0.14754 --
Iso[4]LGE2 Immunized Mouse Serum (Num 973)
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3 Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Iso[4]LGE2 Immunized Mouse Serum (Num 972)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
8.41644E-4 6.39738E-4
Adj. R-Square 0.99931 0.88766Value Standard Error
Iso4-HSA
A1 2.49696 0.01491A2 0.15149 0.01592x0 0.03961 0.00164p 1.19425 0.05244EC20 0.01241 --EC50 0.03961 0.00164EC80 0.12646 --
LGE2-BSA
A1 2.77897 0.00954A2 2.54044 0.05882x0 13.19208 8.6989p 1.14809 0.62089EC20 3.94379 2.16219EC50 13.19208 8.6989EC80 44.12786 52.61921
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 1000
1
2
3
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Iso[4]LGE2 Immunized Mouse Serum (Num 974)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr0.01547 0.00292
Adj. R-Square 0.97081 0.67058Value Standard Error
Iso4-HSA
A1 1.8093 0.07646A2 0.24286 0.06278x0 0.00981 0.0029p 1.04583 0.28089EC20 0.00261 --EC50 0.00981 0.0029EC80 0.03693 --
LGE2-BSA
A1 2.68675 0.01923A2 2.4815 0.05261x0 3.79482 187001.28669p 11.33427 9.23029E6EC20 3.35794 168996.85846EC50 3.79482 187001.28669EC80 4.28854 638493.91753
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3 Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Iso[4]LGE2 Immunized Mouse Serum (Num 975)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00157 0.00146
Adj. R-Square 0.9986 0.60859Value Standard Error
Iso4-HSA
A1 2.50431 0.02101A2 0.1267 0.02728x0 0.09768 0.00701p 0.87433 0.04871EC20 0.02001 --EC50 0.09768 0.00701EC80 0.47686 --
LGE2-BSA
A1 2.60962 0.01352A2 2.4795 0.03815x0 4.0171 325238.25016p 11.56137 7.97073E6EC20 3.56318 6072.41457EC50 4.0171 325238.25016EC80 4.52885 741060.46055
Figure 5.1S Cross-reactivity of iso[4]LGE2-HSA (■) with respect to LGE2-BSA (○) for immunized mice serum (104 dilution).
220

221

222

223
Figure 5.2S 20 plates (total 1920 wells) were done for the fusion step and 725 wells contained hybridoma cells.
224

2E8
(VL) 9G8 (VL)
12G4 (L)
15A5 (L)
18G7 (XXL)
4G5 (S)
5B7 (M)
8B3 (S)
10C11 (M)
16E4 (M)
19A4 (M)
Mice Serum
Iso4-BSA 0.336 2.826 2.557 0.271 2.528 0.123 0.12 0.112 0.136 0.12 0.119 2.825 Iso4-HSA 0.422 2.627 2.868 0.33 2.801 0.111 0.112 0.107 0.155 0.118 0.11 2.875 BSA 0.111 0.087 0.084 0.093 0.088 0.087 0.086 0.085 0.088 0.085 0.086 0.09 LGE2-BSA 0.238 0.105 0.106 0.199 0.835 0.087 0.085 0.084 0.103 0.087 0.088 0.193 LGE2-KLH 0.596 0.3 0.28 0.452 2.704 0.096 0.092 0.087 0.137 0.088 0.089 2.273 CEP-BSA 0.483 0.115 0.144 0.377 2.934 0.112 0.111 0.115 0.148 0.108 0.11 0.233
Clone 9G8(VL) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.101 0.139 0.341 1.721 2.901 2.866 LGE2-BSA 2.779 2.768 2.787 2.764 2.741 2.773 Clone 12G4(L) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.162 0.282 1.077 1.581 1.908 2.161 LGE2-BSA 1.671 1.642 1.642 1.618 1.984 1.99 18G7(XXL) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.158 0.28 0.796 1.039 1.098 1.23 LGE2-BSA 0.713 0.666 0.7 0.906 1.062 1.192 2E8 (VL) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.185 0.145 0.139 0.157 0.171 0.178 LGE2-BSA 0.141 0.132 0.135 0.154 0.165 0.183 15A4(L) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.194 0.189 0.192 0.208 0.209 0.211 LGE2-BSA 0.195 0.210 0.208 0.214 0.205 0.216 4G5(S) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.091 0.087 0.088 0.089 0.087 0.092
0
0.5
1
1.5
2
2.5
3
3.5
Iso4-BSA
Iso4-HSA
BSA
LGE2-BSA
LGE2-KLH
CEP-BSA
225

LGE2-BSA 0.086 0.085 0.086 0.085 0.085 0.089 5B7(M) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.091 0.091 0.087 0.091 0.091 0.091 LGE2-BSA 0.105 0.099 0.096 0.102 0.104 0.098 8B3(S) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.090 0.088 0.091 0.090 0.091 0.094 LGE2-BSA 0.094 0.091 0.094 0.091 0.090 0.101 10C11(M) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.114 0.113 0.109 0.112 0.114 0.117 LGE2-BSA 0.101 0.098 0.099 0.103 0.107 0.113 16E4(M) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.091 0.090 0.089 0.090 0.091 0.094 LGE2-BSA 0.093 0.095 0.094 0.094 0.094 0.095 19A4(M) 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 0.0286 μM 0.00572 μM Iso4-HSA 0.087 0.104 0.085 0.088 0.093 0.090 LGE2-BSA 0.089 0.092 0.091 0.089 0.090 0.094
1E-3 0.01 0.1 1 10 100
0
1
2
3
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
First Screening Clone 9G8 (VL)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00542 3.47E-4
Adj. R-Square 0.99695 -0.38874Value Standard Error
Iso4-HSA
A1 2.92998 0.06295A2 0.12987 0.05353x0 0.16781 0.01153p 1.87261 0.31819EC20 0.08004 --EC50 0.16781 0.01153EC80 0.35182 --
LGE2-BSA
A1 2.757 0.01347A2 2.778 0.01307x0 0.15186 24535.50489p 11.3928 3.02543E7EC20 0.13446 --EC50 0.15186 24535.50489EC80 0.17151 --
1E-3 0.01 0.1 1 10 1000
1
2
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
First Screening Clone 12G4 (L)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.01697 7.05375E-4
Adj. R-Square 0.97579 0.97782Value Standard Error
Iso4-HSA
A1 2.16432 0.17644A2 -0.03979 0.26488x0 0.58803 0.2551p 0.77428 0.27782EC20 0.09814 --EC50 0.58803 0.2551EC80 3.52351 --
LGE2-BSA
A1 1.99 0.02656A2 1.64325 0.01531x0 0.03668 117253.60997p 16.22487 2.08308E8EC20 0.03368 --EC50 0.03668 117253.60997EC80 0.03996 --
1E-3 0.01 0.1 1 10 1000.0
0.5
1.0
1.5First Screening Clone 18G7 (XXL)
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr0.00555 0.002
Adj. R-Square 0.97242 0.9578Value Standard Error
Iso4-HSA
A1 1.16183 0.05626A2 0.10589 0.11033x0 1.08726 0.35461p 1.19937 0.42082EC20 0.34226 0.13992EC50 1.08726 0.35461EC80 3.45395 2.11388
LGE2-BSA
A1 1.20755 0.07748A2 0.67638 0.03667x0 0.0899 0.04122p 1.05988 0.46226EC20 0.02431 --EC50 0.0899 0.04122EC80 0.3325 --
Figure 5.3S Indirect and competitive ELISA tests of first screening clones. 9G8, 12G4, 18G7 showed best responses. 9G8 and 12G4 were picked for first round of subclone.
226

12G4 A2 A10 A11 A12 B2 B4 B5 B6 B8 B10 B12 C2 Iso4-BSA
2.620 2.639 2.660 2.600 1.843 2.562 2.523 2.577 2.601 2.568 2.572 2.674
Iso4-HSA
2.879 2.788 2.841 2.785 2.305 2.809 2.863 2.857 2.884 2.775 2.825 2.833
LGE2-BSA
0.111 0.134 0.147 0.145 0.100 0.151 0.121 0.157 0.147 0.172 0.166 0.163
Control 0.038 0.037 0.035 0.034 0.037 0.037 0.036 0.035 0.035 0.034 0.037 0.039 12G4 C3 C4 C7 C11 D6 D7 D8 D10 D11 E3 E10 F3 Iso4-BSA
2.861 2.760 2.708 2.905 2.808 2.920 2.800 2.909 2.978 2.880 2.914 2.998
Iso4-HSA
2.817 2.842 2.876 2.809 2.848 2.873 2.813 2.828 2.828 2.860 2.863 2.966
LGE2-BSA
0.119 0.136 0.106 0.228 0.147 0.120 0.164 0.152 0.149 0.101 0.157 0.127
12G4 F4 F5 F10 F11 G1 G3 G4 G6 H2 H10 9G8 12G4 Iso4-BSA
2.773 2.676 2.793 2.812 2.805 2.857 2.790 2.904 2.688 2.795 2.877 2.802
Iso4-HSA
2.797 2.761 2.815 2.777 2.762 2.820 2.863 2.847 2.804 2.940 2.793 2.964
LGE2-BSA
0.117 0.134 0.150 0.132 0.157 0.117 0.149 0.152 0.129 0.163 0.126 0.169
Control 0.037 0.035 0.036 0.037 0.037 0.036 0.035 0.035 0.037 0.035 0.037 0.036
Figure 5.4S Indirect ELISA tests of first subclones of 12G4, and all 34 subclones showed strong specificity response.
0
0.5
1
1.5
2
2.5
3
3.5
First Subclones of 12G4
Iso4-BSA
Iso4-HSA
LGE2-BSA
227

12G4 Subclone supernatant
A2 B2 B5 C3 C7 E3 F3 F4 G3 H2 12G4 control
Iso4-BSA 2.909 2.124 2.866 2.850 2.837 2.904 2.946 2.861 2.946 2.946 2.877 0.095 Iso4-HSA 2.992 2.721 2.992 2.998 2.975 2.991 2.998 2.999 2.968 2.992 2.984 0.116 LGE2-BSA 0.139 0.098 0.136 0.121 0.119 0.114 0.156 0.129 0.117 0.135 0.162 0.089 12G4 Subclone supernatant 1 to 10 Dilution
A2 B2 B5 C3 C7 E3 F3 F4 G3 H2 12G4 control
Iso4-BSA 2.452 0.836 2.445 2.262 2.026 2.120 2.627 2.525 2.368 2.582 2.625 0.108 Iso4-HSA 2.801 1.336 2.881 2.711 2.799 2.753 2.916 2.884 2.852 2.966 2.869 0.094 LGE2-BSA 0.099 0.096 0.096 0.101 0.101 0.098 0.105 0.098 0.097 0.105 0.112 0.093
Clone F3 89.33 μM
17.86 μM
3.572 μM
0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
9.15x10-6 μM
Iso4-HSA 0.273 1.236 2.609 2.894 2.898 2.912 2.923 2.934 2.942 2.956 2.964 LGE2-BSA 2.583 2.691 2.828 2.924 2.964 2.968 2.969 2.972 2.961 2.962 2.920
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 1000
1
2
3
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
First SubClone F3 From 12G4 (IgG1)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
4.57502E-4 3.11822E-4
Adj. R-Square 0.99943 0.98211Value Standard Error
Iso4-HSA
A1 2.93245 0.00778A2 0.12189 0.03423x0 13.5591 0.38499p 1.52238 0.05758EC20 5.45454 0.196EC50 13.5591 0.38499EC80 33.70573 1.7493
LGE2-BSA
A1 2.96075 0.00707A2 2.53953 0.04421x0 8.98885 3.02324p 0.9191 0.19181EC20 1.98906 0.49487EC50 8.98885 3.02324EC80 40.62194 24.45795
00.5
11.5
22.5
33.5
A2
B2
B5
C3
C7 E3 F3 F4 G3
H2
12G
4A
2 10
times
dilu
tion
B2
10tim
es d
ilutio
nB
5 10
times
dilu
tion
C3
10tim
es d
ilutio
nC
7 10
times
dilu
tion
E3 1
0tim
es d
ilutio
nF3
10t
imes
dilu
tion
F4 1
0tim
es d
ilutio
nG
3 10
times
dilu
tion
H2
10tim
es d
ilutio
n12
G4
10tim
es d
ilutio
n
First subcloning of 12G4 (10 subclones)
Iso4-BSA
Iso4-HSA
LGE2-BSA
228

Clone F4 89.33 μM 17.86 μM 3.572 μM 0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
9.15x10-6 μM
Iso4-HSA 0.103 0.123 0.372 1.81 2.411 2.672 2.867 2.701 2.483 2.567 2.828 LGE2-BSA 1.938 2.073 1.943 2.068 2.247 2.53 2.594 2.602 2.538 2.462 2.484
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3
First SubClone F4 From 12G4 (IgG1)
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.01975 0.00443
Adj. R-Square 0.98458 0.93767Value Standard Error
Iso4-HSA
A1 2.66941 0.05594A2 0.07805 0.10931x0 1.06241 0.17855p 1.53969 0.34607EC20 0.43178 0.0931EC50 1.06241 0.17855EC80 2.61408 0.79247
LGE2-BSA
A1 2.53988 0.0297A2 1.9952 0.03776x0 0.13939 0.042p 1.71788 1.08487EC20 0.0622 --EC50 0.13939 0.042EC80 0.31239 --
Clone H2 89.33 μM 17.86 μM 3.572 μM 0.7144
μM 0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
9.15x10-6 μM
Iso4-HSA 0.152 0.528 1.923 2.821 2.823 2.935 2.978 2.894 2.799 2.908 2.97 LGE2-BSA 2.403 2.441 2.514 2.687 2.806 2.86 2.98 2.945 2.974 2.998 2.976
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3
First SubClone H2 From 12G4 (IgG1)
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00413 5.99668E-4
Adj. R-Square 0.99614 0.98878Value Standard Error
Iso4-HSA
A1 2.9075 0.0239A2 0.1184 0.07811x0 5.41547 0.46265p 1.50213 0.16144EC20 2.15194 0.199EC50 5.41547 0.46265EC80 13.62829 2.18571
LGE2-BSA
A1 2.98294 0.01396A2 2.36034 0.03844x0 0.66943 0.22011p 0.57159 0.09008EC20 0.05921 --EC50 0.66943 0.22011EC80 7.56842 --
Clone E3 17.86
μM 3.572 μM
0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
9.15x10-6 μM
Iso4-HSA 0.112 0.405 1.466 1.721 1.614 1.572 1.651 1.617 1.733 LGE2-BSA 1.852 1.672 1.657 1.699 1.888 1.907 1.953 1.867 1.9
229

1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2First SubClone E3 From 12G4 (IgG1)
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00427 0.00558
Adj. R-Square 0.9886 0.56489Value Standard Error
Iso4-HSA
A1 1.65156 0.02702A2 0.10152 0.07127x0 1.84199 0.25133p 2.14595 0.34992EC20 0.96545 0.16588EC50 1.84199 0.25133EC80 3.51435 0.60774
LGE2-BSA
A1 1.90675 0.03735A2 1.72 0.04313x0 0.03357 45925.02555p 13.68614 1.16874E8EC20 0.03034 --EC50 0.03357 45925.02555EC80 0.03715 --
Clone 12G4 89.33 μM 17.86 μM 3.572 μM 0.7144
μM 0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
9.15x10-6 μM
Iso4-HSA 0.326 1.209 2.303 2.645 2.934 2.928 2.931 2.899 2.839 2.875 2.882 LGE2-BSA 2.557 2.599 2.571 2.864 2.878 2.896 2.921 2.936 2.949 2.816 2.859
1E-6 1E-5 1E-4 1E-3 0.01 0.1 1 10 1000
1
2
3
Iso4-HSA LGE2-BSA
12G4 (IgG1)
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00316 0.002
Adj. R-Square 0.99581 0.91311Value Standard Error
Iso4-HSA
A1 2.89609 0.02195A2 -0.11517 0.18039x0 14.15572 2.45868p 0.96226 0.10066EC20 3.35167 0.37545EC50 14.15572 2.45868EC80 59.78639 18.25262
LGE2-BSA
A1 2.89357 0.0169A2 2.57567 0.03162x0 0.886 4884.16907p 10.57528 270803.30233EC20 0.77715 --EC50 0.886 4884.16907EC80 1.0101 --
Clone A2 17.86
μM 3.572 μM
0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
Iso4-HSA 0.408 1.937 2.398 2.436 2.417 2.380 2.598 2.436 2.379 LGE2-BSA 2.383 2.304 2.324 2.368 2.365 2.288 2.372 2.415 2.362 Clone B2 17.86 μM 3.572 μM 0.7144
μM 0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
Iso4-HSA 0.188 1.164 2.145 2.187 2.230 2.183 2.368 2.086 LGE2-BSA 2.117 2.043 2.071 2.076 2.105 2.101 2.309 1.648 Clone B5 17.86
μM 3.572 μM
0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
Iso4-HSA 0.178 0.972 1.992 2.039 2.116 2.018 2.208 2.223 2.023 LGE2-BSA 2.131 1.998 1.995 1.969 2.077 2.055 2.123 2.231 1.983 Clone C3 17.86
μM 3.572 μM
0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
Iso4-HSA 0.897 2.105 2.244 2.246 2.270 2.224 2.248 2.391 2.232 LGE2-BSA 2.422 2.265 2.286 2.334 2.291 2.189 2.199 2.312 2.199
230

Clone C7 17.86 μM
3.572 μM
0.7144 μM
0.1429 μM
2.86x10-2
μM 5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
Iso4-HSA 0.647 2.103 2.445 2.473 2.604 2.556 2.657 2.641 2.458 LGE2-BSA 2.451 2.464 2.469 2.484 2.555 2.573 2.632 2.475 2.559 Clone G3
17.86 μM
3.572 μM
0.7144 μM
0.1429 μM
2.86x10-
2μM 5.72x10-
3 μM 1.14x10-
3 μM 2.28x10-
4 μM 4.58x10-
5 μM 9.15x10-
6 μM 1.83x10-
6 μM 3.66x10-
7 μM Iso4-HSA
1.242 2.461 2.796 2.793 2.878 2.910 2.868 2.820 2.956 2.882 2.740 2.685
LGE2-BSA
2.520 2.408 2.455 2.641 2.567 2.611 2.644 2.743 2.642 2.602 2.460 2.566
Figure 5.5S Indirect and competitive ELISA tests of 10 clones from 12G4, and 12G4 F3 was chose for next subclone.
12G4 F3 Subclone supernatant A4 A6 B3 B6 C1 C4 C6 D1 control Iso4-BSA 2.962 2.852 2.890 2.672 2.877 2.733 2.906 2.834 0.096 Iso4-HSA 2.869 2.816 2.867 2.799 2.859 2.729 2.867 2.851 0.093 LGE2-BSA 0.203 0.116 0.173 0.106 0.144 0.106 0.201 0.178 0.087 12G4 F3 Subclone supernatant 1 to 10 Dilution
A4 A6 B3 B6 C1 C4 C6 D1 control
Iso4-BSA 2.907 2.467 2.781 1.996 2.803 2.128 2.902 2.790 0.100 Iso4-HSA 2.958 2.670 2.788 2.428 2.781 2.509 2.817 2.849 0.092 LGE2-BSA 0.123 0.100 0.107 0.093 0.111 0.095 0.117 0.114 0.091 12G4 F3 Subclone supernatant D4 E1 E4 F4 F5 G2 G4 G7 control Iso4-BSA 2.748 2.735 2.729 2.749 2.779 2.673 2.571 2.702 0.092 Iso4-HSA 2.907 2.875 2.848 2.838 2.814 2.827 2.774 2.830 0.089 LGE2-BSA 0.187 0.190 0.185 0.211 0.198 0.149 0.123 0.199 0.095 12G4 F3 Subclone supernatant 1 to 10 Dilution
D4 E1 E4 F4 F5 G2 G4 G7 control
Iso4-BSA 2.700 2.668 2.650 2.713 2.751 2.657 2.373 2.676 0.093 Iso4-HSA 2.986 2.937 2.828 2.905 2.860 2.801 2.751 2.881 0.092 LGE2-BSA 0.119 0.128 0.129 0.124 0.130 0.112 0.106 0.125 0.099
0
0.5
1
1.5
2
2.5
3
3.5
A4 A6 B3 B6 C1 C4 C6 D1 D4 E1 E4 F4 F5 G2 G4 G7
16 Secondary SubClones From 12G4 F3 (IgG)
Iso4-BSA
Iso4-HSA
LGE2-BSA
231

Figure 5.6S Indirect ELISA tests of 16 secondary subclones from 12G4 F3.
12G4 F3 SubClone
Iso4-BSA Iso4-HSA Iso4-KLH BSA KLH HSA LGE2-BSA CEP-BSA CEP-HSA
C1 2.732 2.971 2.643 0.105 0.110 0.106 0.149 0.146 0.147 D1 2.596 2.809 2.580 0.097 0.101 0.097 0.149 0.135 0.155 G4 2.533 2.815 2.386 0.093 0.098 0.093 0.110 0.113 0.112
12G4 F3 SubClone 10time Dilution
Iso4-BSA Iso4-HSA Iso4-KLH BSA KLH HSA LGE2-BSA CEP-BSA
CEP-HSA
00.5
11.5
22.5
33.5
16 Secondary SubClones From 12G4 F3 (IgG) (10 Times Dilution)
Iso4-BSA
Iso4-HSA
LGE2-BSA
0
0.5
1
1.5
2
2.5
3
3.5C1 D1 G4 Secondary Subclones from 12G4 F3
C1
C1 10timeDilutionD1
D1 10timeDilutionG4
G4 10timeDilution
232

C1 10time Dilution 2.157 2.635 1.777 0.102 0.104 0.105 0.107 0.103 0.102 D1 10time Dilution 2.370 2.839 2.057 0.096 0.098 0.094 0.106 0.102 0.105 G4 10time Dilution 1.576 2.422 1.338 0.100 0.106 0.100 0.103 0.099 0.103
Clone C1 446.65 89.33 μM 17.86 μM 3.572 μM 0.7144 μM 0.1429 μM 2.86x10-2 μM 5.72x10-3 μM 1.14x10-3 μM
Iso4-HSA 0.167 0.866 2.494 2.67 2.603 2.603 2.511 2.476 2.543 LGE2-BSA 2.725 2.727 2.725 2.662 2.679 2.629 2.573 2.545 2.569
0
0.5
1
1.5
2
2.5
3
3.5
C1 D1 G4 Secondary Subclones from 12G4 F3
C1
D1
G4
0
0.5
1
1.5
2
2.5
3
C1 D1 G4 Secondary Subclones from 12G4 F3
C1 10time Dilution
D1 10time Dilution
G4 10time Dilution
233

1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3
Iso4-HSA LGE2-BSA
Abs
orba
nce
(405
nm
)
Conc (mM)
Secondary SubClone C1 From 12G4 F3 (IgG1)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.00508 4.64133E-4
Adj. R-Square 0.99398 0.91179Value Standard Error
Iso4-HSA
A1 2.56737 0.02922A2 0.15447 0.07536x0 64.78795 6.16553p 2.71801 0.71018EC20 38.90331 8.28027EC50 64.78795 6.16553EC80 107.89513 9.84797
LGE2-BSA
A1 2.54534 0.02859A2 2.72758 0.01936x0 0.27338 0.2359p 0.5939 0.30393EC20 0.02649 --EC50 0.27338 0.2359EC80 2.82161 --
Clone D1 446.65 89.33 μM 17.86 μM 3.572 μM 0.7144
μM 0.1429 μM
2.86x10-2 μM
5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
Iso4-HSA 0.237 1.071 2.467 2.74 2.744 2.727 2.718 2.607 2.521 2.511 2.576 LGE2-BSA 2.768 2.757 2.674 2.709 2.762 2.721 2.661 2.618 2.581 2.548 2.583
1E-5 1E-4 1E-3 0.01 0.1 1 10 1000
1
2
3
Iso4-HSA LGE2-BSA
Secondary SubClone D1 From 12G4 F3 (IgG1)
Abs
orba
nce
(405
nm
)
Conc (mM)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.01029 0.00111
Adj. R-Square 0.98486 0.82315Value Standard Error
Iso4-HSA
A1 2.64274 0.03628A2 0.18262 0.12464x0 66.95829 7.53202p 1.99096 0.46569EC20 33.37389 6.99503EC50 66.95829 7.53202EC80 134.33893 24.92572
LGE2-BSA
A1 2.56746 0.02426A2 2.73474 0.01579x0 0.01721 0.01361p 0.9968 0.68458EC20 0.00428 --EC50 0.01721 0.01361EC80 0.06916 --
Clone G4 446.65 89.33 μM 17.86 μM 3.572 μM 0.7144
μM 0.1429 μM
2.86x10-2 μM
5.72x10-3 μM
1.14x10-3 μM
2.28x10-4 μM
4.58x10-5 μM
Iso4-HSA 0.146 0.312 1.715 2.509 2.493 2.441 2.361 2.296 2.29 2.22 2.204 LGE2-BSA 2.617 2.603 2.513 2.536 2.464 2.434 2.36 2.354 2.273 2.232 2.248
1E-5 1E-4 1E-3 0.01 0.1 1 10 100
0
1
2
3
Abs
orba
nce
(405
nm
)
Conc (mM)
Iso4-HSA LGE2-BSA
Secondary SubClone G4 From 12G4 F3 (IgG1)
Model Logistic
Equationy = A2 + (A1-A2)/(1 + (x/x0)^p)
Reduced Chi-Sqr
0.01528 6.5648E-4
Adj. R-Square 0.97925 0.96563Value Standard Error
Iso4-HSA
A1 2.35277 0.04418A2 0.17214 0.1278x0 26.115 5.41129p 2.40971 1.04015EC20 14.69069 2.18377EC50 26.115 5.41129EC80 46.42352 20.08087
LGE2-BSA
A1 2.15437 0.10782A2 2.68791 0.10761x0 0.14208 0.19755p 0.22635 0.11969EC20 3.10904E-4 --EC50 0.14208 0.19755EC80 64.92507 --
Figure 5.7S Indirect and competitive ELISA tests of final three candidates from 12G4 F3--C1 D1 G4.
Antigens Iso4-BSA
Iso4-HSA
Iso4-KLH
LGE2-BSA
BSA HSA KLH CEP-BSA
CEP-HSA
OHdiA-BSA
PBS
mAb 2.833 2.935 2.948 0.252 0.155 0.13 0.178 0.344 0.354 0.228 0.128
234
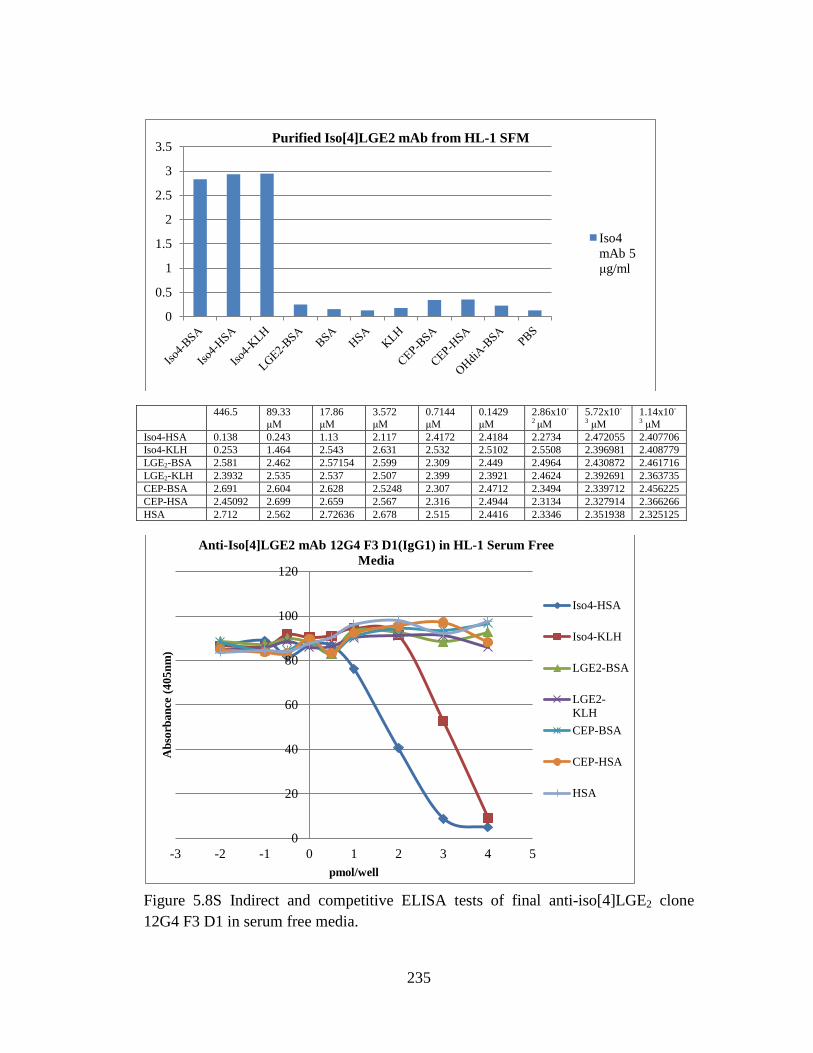
446.5 89.33
μM 17.86 μM
3.572 μM
0.7144 μM
0.1429 μM
2.86x10-
2 μM 5.72x10-
3 μM 1.14x10-
3 μM Iso4-HSA 0.138 0.243 1.13 2.117 2.4172 2.4184 2.2734 2.472055 2.407706 Iso4-KLH 0.253 1.464 2.543 2.631 2.532 2.5102 2.5508 2.396981 2.408779 LGE2-BSA 2.581 2.462 2.57154 2.599 2.309 2.449 2.4964 2.430872 2.461716 LGE2-KLH 2.3932 2.535 2.537 2.507 2.399 2.3921 2.4624 2.392691 2.363735 CEP-BSA 2.691 2.604 2.628 2.5248 2.307 2.4712 2.3494 2.339712 2.456225 CEP-HSA 2.45092 2.699 2.659 2.567 2.316 2.4944 2.3134 2.327914 2.366266 HSA 2.712 2.562 2.72636 2.678 2.515 2.4416 2.3346 2.351938 2.325125
Figure 5.8S Indirect and competitive ELISA tests of final anti-iso[4]LGE2 clone 12G4 F3 D1 in serum free media.
0
0.5
1
1.5
2
2.5
3
3.5 Purified Iso[4]LGE2 mAb from HL-1 SFM
Iso4 mAb 5 μg/ml
0
20
40
60
80
100
120
-3 -2 -1 0 1 2 3 4 5
Abs
orba
nce
(405
nm)
pmol/well
Anti-Iso[4]LGE2 mAb 12G4 F3 D1(IgG1) in HL-1 Serum Free Media
Iso4-HSA
Iso4-KLH
LGE2-BSA
LGE2-KLHCEP-BSA
CEP-HSA
HSA
235

Bibliography

1. Achord, J. M.; Hussey, C. L. (1980) Determination of dissolved oxygen in
nonaqueous electrochemical solvents. Anal. Biochem. 52 (3), 601-602.
2. Alberts, B., Molecular Biology of the Cell. 3rd ed.; Garland Pub: New York,
1994.
3. Amarnath, V.; Valentine, W. M.; Amarnath, K.; Eng, M. A.; Graham, D. G.
(1994) The mechanism of nucleophilic substitution of alkylpyrroles in the
presence of oxygen. Chem. Res. Toxicol. 7 (1), 56-61.
4. Azam, A.; Moriguchi, T.; Shinozuka, K. (2006) Modified alpha-beta chimeric
oligoDNA bearing a multi-conjugate of 2,2-bis(hydroxymethyl)propionic
acid-anthraquinone-polyamine exhibited improved and stereo-nonspecific
triplex-forming ability. Chem. Commun. (3), 335-337.
5. Azuma, M.; Shearer, T. R. (2008) The role of calcium-activated protease
calpain in experimental retinal pathology. Surv Ophthalmol 53 (2), 150-163.
6. Bailen, M. A.; Chinchilla, R.; Dodsworth, D. J.; Najera, C. (1999) 2-
mercaptopyridone 1-oxide-based uronium salts: New peptide coupling
reagents. J. Org. Chem. 64 (24), 8936-8939.
7. Barriga, S. (2001) 2,2,6,6-Tetramethylpiperidin-1-oxyl (TEMPO). Synlett
2001 (04), 0563.
8. Battino, R., Oxygen and ozone. Pergamon: Oxford; New York, 1981; Vol. 7.
9. Bennett, B. J.; de Aguiar Vallim, T. Q.; Wang, Z.; Shih, D. M.; Meng, Y.;
Gregory, J.; Allayee, H.; Lee, R.; Graham, M.; Crooke, R.; Edwards, P. A.;
Hazen, S. L.; Lusis, A. J. (2013) Trimethylamine-N-oxide, a metabolite
associated with atherosclerosis, exhibits complex genetic and dietary
regulation. Cell Metab 17 (1), 49-60.
10. Ben-Shabat, S.; Itagaki, Y.; Jockusch, S.; Sparrow, J. R.; Turro, N. J.;
Nakanishi, K. (2002) Formation of a nonaoxirane from A2E, a lipofuscin
fluorophore related to macular degeneration, and evidence of singlet oxygen
involvement. Angew. Chem. Int. Ed. Engl. 41 (5), 814-817.
11. Bernoud-Hubac, N.; Fay, L. B.; Armarnath, V.; Guichardant, M.; Bacot, S.;
237

Davies, S. S.; Roberts, L. J., 2nd; Lagarde, M. (2004) Covalent binding of
isoketals to ethanolamine phospholipids. Free Radic Biol Med 37 (10), 1604-
1611.
12. Berry, M. D. (2004) Glyceraldehyde-3-phosphate dehydrogenase as a target
for small-molecule disease-modifying therapies in human neuro degenerative
disorders. J Psychiatry Neurosci 29 (5), 337-345.
13. Bhattacharya, S. K.; Rockwood, E. J.; Smith, S. D.; Bonilha, V. L.; Crabb, J.
S.; Kuchtey, R. W.; Robertson, N. G.; Peachey, N. S.; Morton, C. C.; Crabb,
J. W. (2005) Proteomics reveal cochlin deposits associated with glaucomatous
trabecular meshwork. J. Biol. Chem. 280 (7), 6080-6084.
14. Borrebaeck, C. A. (2000) Antibodies in diagnostics - from immunoassays to
protein chips. Immunol Today 21 (8), 379-382.
15. Boutaud, O.; Andreasson, K. I.; Zagol-Ikapitte, I.; Oates, J. A. (2005)
Cyclooxygenase-dependent lipid-modification of brain proteins. Brain Pathol
15 (2), 139-142.
16. Boutaud, O.; Brame, C. J.; Chaurand, P.; Li, J. Y.; Rowlinson, S. W.; Crews,
B. C.; Ji, C.; Marnett, L. J.; Caprioli, R. M.; Roberts, L. J.; Oates, J. A. (2001)
Characterization of the lysyl adducts of prostaglandin H-synthases that are
derived from oxygenation of arachidonic acid. Biochemistry 40 (23), 6948-
6955.
17. Boutaud, O.; Brame, C. J.; Salomon, R. G.; Roberts, L. J., 2nd; Oates, J. A.
(1999) Characterization of the lysyl adducts formed from prostaglandin H2 via
the levuglandin pathway. Biochemistry 38 (29), 9389-9396.
18. Boutaud, O.; Montine, T. J.; Chang, L.; Klein, W. L.; Oates, J. A. (2006)
PGH2-derived levuglandin adducts increase the neurotoxicity of amyloid
beta1-42. J. Neurochem. 96 (4), 917-923.
19. Boutaud, O.; Ou, J. J.; Chaurand, P.; Caprioli, R. M.; Montine, T. J.; Oates, J.
A. (2002) Prostaglandin H2 (PGH2) accelerates formation of amyloid beta1-
42 oligomers. J. Neurochem. 82 (4), 1003-1006.
238

20. Brame, C. J.; Boutaud, O.; Davies, S. S.; Yang, T.; Oates, J. A.; Roden, D.;
Roberts, L. J., 2nd (2004) Modification of proteins by isoketal-containing
oxidized phospholipids. J. Biol. Chem. 279 (14), 13447-13451.
21. Brame, C. J.; Salomon, R. G.; Morrow, J. D.; Roberts, L. J. (1999)
Identification of extremely reactive gamma-ketoaldehydes (isolevuglandins)
as products of the isoprostane pathway and characterization of their lysyl
protein adducts. J. Biol. Chem. 274 (19), 13139-13146.
22. Brennan, M. L.; Hazen, S. L. (2003) Amino acid and protein oxidation in
cardiovascular disease. Amino Acids 25 (3-4), 365-374.
23. Butterfield, D. A.; Drake, J.; Pocernich, C.; Castegna, A. (2001) Evidence of
oxidative damage in Alzheimer's disease brain: central role for amyloid β-
peptide. Trends Mol. Med. 7 (12), 548-554.
24. Butterfield, D. A.; Lauderback, C. M. (2002) Lipid peroxidation and protein
oxidation in Alzheimer's disease brain: potential causes and consequences
involving amyloid beta-peptide-associated free radical oxidative stress. Free
Radic Biol Med 32 (11), 1050-1060.
25. Caldes, C.; Vilanova, B.; Adrover, M.; Munoz, F.; Donoso, J. (2011) Phenol
group in pyridoxamine acts as a stabilizing element for its carbinolamines and
Schiff bases. Chem Biodivers 8 (7), 1318-1332.
26. Capra, J. D.; Tung, A. S.; Nisonoff, A. (1975) Structural studies on induced
antibodies with defined idiotypic specificities. II. The light chains of anti-p-
azophenylarsonate antibodies from A/J mice bearing a cross-reactive idiotype.
J Immunol 115 (2), 414-418.
27. Carrier, E. J.; Amarnath, V.; Oates, J. A.; Boutaud, O. (2009) Characterization
of covalent adducts of nucleosides and DNA formed by reaction with
levuglandin. Biochemistry 48 (45), 10775-10781.
28. Castellani, R. J.; Perry, G.; Harris, P. L.; Cohen, M. L.; Sayre, L. M.;
Salomon, R. G.; Smith, M. A. (1998) Advanced lipid peroxidation end-
products in Alexander's disease. Brain Res 787 (1), 15-18.
239

29. Chan, L. C.; Cox, B. G. (2007) Kinetics of amide formation through
Carbodiimide/N-Hydroxybenzotriazole (HOBt) couplings. J. Org. Chem. 72
(23), 8863-8869.
30. Charvet, C. D.; Laird, J.; Xu, Y.; Salomon, R. G.; Pikuleva, I. A. (2013)
Posttranslational modification by an isolevuglandin diminishes activity of the
mitochondrial cytochrome P450 27A1. J. Lipid Res. 54 (5), 1421-1429.
31. Charvet, C. D.; Liao, W. L.; Heo, G. Y.; Laird, J.; Salomon, R. G.; Turko, I.
V.; Pikuleva, I. A. (2011) Isolevuglandins and Mitochondrial Enzymes in the
Retina mass spectrometry detection of post-translational modification of
sterol-metabolizing CYP27A1. J. Biol. Chem. 286 (23), 20413-20422.
32. Charvet, C. D.; Saadane, A.; Wang, M.; Salomon, R. G.; Brunengraber, H.;
Turko, I. V.; Pikuleva, I. A. (2013) Pretreatment with pyridoxamine mitigates
isolevuglandin-associated retinal effects in mice exposed to bright light. J.
Biol. Chem. 288 (41), 29267-29280.
33. Clarke, S. D. (2001) Polyunsaturated fatty acid regulation of gene
transcription: a molecular mechanism to improve the metabolic syndrome. J
Nutr 131 (4), 1129-1132.
34. Clausen, J. (1984) Demential syndromes and the lipid metabolism. Acta
Neurol Scand 70 (5), 345-355.
35. Cohen, S. A.; Michaud, D. P. (1993) Synthesis of a fluorescent derivatizing
reagent, 6-aminoquinolyl-n-hydroxysuccinimidyl carbamate, and its
application for the analysis of hydrolysate amino-acids via high-performance
liquid-chromatography. Anal. Biochem. 211 (2), 279-287.
36. Collier, H. O. J.; Roy, A. C. (1974) Hypothesis - inhibition of prostaglandin-e-
sensitive adenyl-cyclase as mechanism of morphine analgesia. Prostaglandins
7 (5), 361-376.
37. Cooper, G. M., The Cell: A Molecular Approach. . ASM Press; Sinauer
Associates: Washington, D. C., 1997.
38. Coughlin, D. J.; Salomon, R. G. (1977) Synthesis of thermal reactivity of
240

some 2,3-dioxabicyclo[2.2.1]heptane models of prostaglandin endoperoxides.
J. Am. Chem. Soc. 99 (2), 655-657.
39. Cowan-Jacob, S. W.; Kaufmann, M.; Anselmo, A. N.; Stark, W.; Grutter, M.
G. (2003) Structure of rabbit-muscle glyceraldehyde-3-phosphate
dehydrogenase. Acta Crystallogr Sect D-Biol Crystallogr 59, 2218-2227.
40. Cox, B.; Murphey, L. J.; Zackert, W. E.; Chinery, R.; Graves-Deal, R.;
Boutaud, O.; Oates, J. A.; Coffey, R. J.; Morrow, J. D. (2002) Human
colorectal cancer cells efficiently conjugate the cyclopentenone prostaglandin,
prostaglandin J(2), to glutathione. Biochim. Biophys. Acta 1584 (1), 37-45.
41. Crabb, J. W.; Ericsson, L.; Atherton, D.; Smith, A. J.; Kutny, R., A
collaborative amino acid analysis study from the association of biomolecular
resource facilities. In Current research in protein chemistry, Villafranca, J. J.,
Ed. Elsevier Science: Academic Press, Inc, 1990; pp 49-61.
42. Crabb, J. W.; Miyagi, M.; Gu, X. R.; Shadrach, K.; West, K. A.; Sakaguchi,
H.; Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.;
Hollyfield, J. G. (2002) Drusen proteome analysis: An approach to the
etiology of age-related macular degeneration. Proc Natl Acad Sci U S A 99
(23), 14682-14687.
43. Croall, D. E.; Ersfeld, K. (2007) The calpains: modular designs and functional
diversity. Genome Biol 8 (6), 218.
44. Davies, K. J. (1995) Oxidative stress: the paradox of aerobic life. Biochem.
Soc. Symp. 61, 1-31.
45. Davies, S. S.; Amarnath, V.; Brame, C. J.; Boutaud, O.; Roberts, L. J., 2nd
(2007) Measurement of chronic oxidative and inflammatory stress by
quantification of isoketal/levuglandin gamma-ketoaldehyde protein adducts
using liquid chromatography tandem mass spectrometry. Nat Protoc 2 (9),
2079-2091.
46. Davies, S. S.; Amarnath, V.; Montine, K. S.; Bernoud-Hubac, N.; Boutaud,
O.; Montine, T. J.; Roberts, L. J. (2002) Effects of reactive gamma-
241

ketoaldehydes formed by the isoprostane pathway (isoketals) and
cyclooxygenase pathway (levuglandins) on proteasome function. FASEB J. 16
(3), 715-717.
47. Davies, S. S.; Brantley, E. J.; Voziyan, P. A.; Amarnath, V.; Zagol-Ikapitte, I.;
Boutaud, O.; Hudson, B. G.; Oates, J. A.; Roberts, L. J., 2nd (2006)
Pyridoxamine analogues scavenge lipid-derived gamma-ketoaldehydes and
protect against H2O2-mediated cytotoxicity. Biochemistry 45 (51), 15756-
15767.
48. Davies, S. S.; Talati, M.; Wang, X.; Mernaugh, R. L.; Amarnath, V.; Fessel,
J.; Meyrick, B. O.; Sheller, J.; Roberts, L. J., 2nd (2004) Localization of
isoketal adducts in vivo using a single-chain antibody. Free Radic Biol Med
36 (9), 1163-1174.
49. Davis, T. L.; Walker, J. R.; Finerty, P. J., Jr.; Mackenzie, F.; Newman, E. M.;
Dhe-Paganon, S. (2007) The crystal structures of human calpains 1 and 9
imply diverse mechanisms of action and auto-inhibition. J. Mol. Biol. 366 (1),
216-229.
50. Difranco, E.; Subbanagounder, G.; Kim, S.; Murthi, K.; Taneda, S.; Monnier,
V. M.; Salomon, R. G. (1995) Formation and stability of pyrrole adducts in
the reaction of levuglandin E(2) with proteins. Chem. Res. Toxicol. 8 (1), 61-
67.
51. Dohi, T.; Ito, M.; Yamaoka, N.; Morimoto, K.; Fujioka, H.; Kita, Y. (2009)
Hypervalent iodine(III): selective and efficient single-electron-transfer (SET)
oxidizing agent. Tetrahedron 65 (52), 10797-10815.
52. Dohi, T.; Morimoto, K.; Maruyama, A.; Kita, Y. (2006) Direct synthesis of
bipyrroles using phenyliodine bis(trifluoroacetate) with bromotrimethylsilane.
Org. Lett. 8 (10), 2007-2010.
53. Doucet, J. P.; Tuana, B. S. (1991) Identification of low molecular weight
GTP-binding proteins and their sites of interaction in subcellular fractions
from skeletal muscle. J. Biol. Chem. 266 (26), 17613-17620.
242

54. Englander, S. W.; Calhoun, D. B.; Englander, J. J. (1987) Biochemistry
without oxygen. Anal. Biochem. 161 (2), 300-306.
55. Evans, D. H.; Claiborne, J. B., The Physiology of Fishes. CRC Press: 2006.
56. Fothergill-Gilmore, L. A.; Michels, P. A. (1993) Evolution of glycolysis.
Prog. Biophys. Mol. Biol. 59 (2), 105-235.
57. Fraga, M. F.; Ballestar, E.; Villar-Garea, A.; Boix-Chornet, M.; Espada, J.;
Schotta, G.; Bonaldi, T.; Haydon, C.; Ropero, S.; Petrie, K.; Iyer, N. G.;
Perez-Rosado, A.; Calvo, E.; Lopez, J. A.; Cano, A.; Calasanz, M. J.;
Colomer, D.; Piris, M. A.; Ahn, N.; Imhof, A.; Caldas, C.; Jenuwein, T.;
Esteller, M. (2005) Loss of acetylation at Lys16 and trimethylation at Lys20
of histone H4 is a common hallmark of human cancer. Nat Genet 37 (4), 391-
400.
58. Franco, C.; Olmsted, J., 3rd (1990) Photochemical determination of the
solubility of oxygen in various media. Talanta 37 (9), 905-909.
59. Fuks, F. (2005) DNA methylation and histone modifications: teaming up to
silence genes. Curr Opin Genet Dev 15 (5), 490-495.
60. Fukuda, K.; Davies, S. S.; Nakajima, T.; Ong, B. H.; Kupershmidt, S.; Fessel,
J.; Amarnath, V.; Anderson, M. E.; Boyden, P. A.; Viswanathan, P. C.;
Roberts, L. J.; Balser, J. R. (2005) Oxidative Mediated Lipid Peroxidation
Recapitulates Proarrhythmic Effects on Cardiac Sodium Channels. Circ Res
97 (12), 1262-1269.
61. Fukushima, T.; Arai, K.; Tomiya, M.; Mitsuhashi, S.; Sasaki, T.; Santa, T.;
Imai, K.; Toyo'oka, T. (2008) Fluorescence determination of N-acetylaspartic
acid in the rat cerebrum homogenate using high-performance liquid
chromatography with pre-column fluorescence derivatization. Biomed.
Chromatogr. 22 (1), 100-105.
62. Fukushima, T.; Usui, N.; Santa, T.; Imai, K. (2003) Recent progress in
derivatization methods for LC and CE analysis. J. Pharm. Biomed. Anal. 30
(6), 1655-1687.
243

63. Glenner, G. G.; Wong, C. W. (1984) Alzheimer's disease: initial report of the
purification and characterization of a novel cerebrovascular amyloid protein.
Biochem. Biophys. Res. Commun. 120 (3), 885-890.
64. Goldsby, R. A.; Kindt, T. J.; Osborne, B. A.; Kiuby, J., Immunology. Fifth
Edition ed.; New York, 2003.
65. Govindarajan, B.; Junk, A.; Algeciras, M.; Salomon, R. G.; Bhattacharya, S.
K. (2009) Increased isolevuglandin-modified proteins in glaucomatous
astrocytes. Mol Vis 15 (114-15), 1079-1091.
66. Govindarajan, B.; Laird, J.; Salornon, R. G.; Bhattacharya, S. K. (2008)
Isolevuglaridin-modified proteins, including elevated levels of inactive
calpain-1, accumulate in glaucomatous trabecular meshwork. Biochemistry 47
(2), 817-825.
67. Govindarajan, B.; Laird, J. M.; Sherman, R.; Salomon, R. G.; Bhattacharya, S.
K. (2008) Neuroprotection in Glaucoma Using Calpain-1 Inhibitors: Regional
Differences in Calpain-1 Activity in the Trabecular Meshwork, Optic Nerve
and Implications for Therapeutics. CNS Neurol Disord Drug Targets 7 (3),
295-304.
68. Greenberg, M. E.; Li, X. M.; Gugiu, B. G.; Gu, X.; Qin, J.; Salomon, R. G.;
Hazen, S. L. (2008) The lipid whisker model of the structure of oxidized cell
membranes. J. Biol. Chem. 283 (4), 2385-2396.
69. Grisham, M. B. (1994) Oxidants and free radicals in inflammatory bowel
disease. Lancet 344 (8926), 859-861.
70. Gu, J.; Pauer, G. J.; Yue, X.; Narendra, U.; Sturgill, G. M.; Bena, J.; Gu, X.;
Peachey, N. S.; Salomon, R. G.; Hagstrom, S. A.; Crabb, J. W. (2009)
Assessing susceptibility to age-related macular degeneration with proteomic
and genomic biomarkers. Mol Cell Proteomics 8 (6), 1338-1349.
71. Gu, X. R.; Meer, S. G.; Miyagi, M.; Rayborn, M. E.; Hollyfield, J. G.; Crabb,
J. W.; Salomon, R. G. (2003) Carboxyethylpyrrole protein adducts and
autoantibodies, biomarkers for age-related macular degeneration. J. Biol.
244

Chem. 278 (43), 42027-42035.
72. Guichardant, M.; Taibi-Tronche, P.; Fay, L. B.; Lagarde, M. (1998) Covalent
modifications of aminophospholipids by 4-hydroxynonenal. Free Radic Biol
Med 25 (9), 1049-1056.
73. Gulcan, H. G.; Alvarez, R. A.; Maude, M. B.; Anderson, R. E. (1993) Lipids
of human retina, retinal-pigment epithelium, and bruch membrane choroid -
comparison of macular and peripheral regions. Invest Ophthalmol Vis Sci 34
(11), 3187-3193.
74. Guo, L.; Chen, Z.; Cox, B. E.; Amarnath, V.; Epand, R. F.; Epand, R. M.;
Davies, S. S. (2011) Phosphatidylethanolamines modified by gamma-
ketoaldehyde (gammaKA) induce endoplasmic reticulum stress and
endothelial activation. J. Biol. Chem. 286 (20), 18170-18180.
75. Guo, L.; Gragg, S. D.; Chen, Z.; Zhang, Y.; Amarnath, V.; Davies, S. S.
(2013) Isolevuglandin-modified phosphatidylethanolamine is metabolized by
NAPE-hydrolyzing phospholipase D. J. Lipid Res. 54 (11), 3151-3157.
76. Hansen, G. H.; Henriksen, R. M.; Kamounah, F. S.; Lund, T.; Hammerich, O.
(2005) Oxidative coupling and polymerization of pyrroles: Part I. The
electrochemical oxidation of 2,4-dimethyl-3-ethylpyrrole in acetonitrile.
Electrochim. Acta 50 (25–26), 4936-4955.
77. Harlow, E.; Lane, D., Antibodies- A Laboratory Manual. Cold Spring Harbor
Laboratory Press: Cold Spring Harbor, New York, 1988.
78. Harris, J. I.; Waters, M., The enzymes. 3rd edn (Boyer, P. D., ed.) ed.;
Academic Press: New York, 1976; Vol. 13.
79. Heaney, H.; Ley, S. V. (1974) 1-Benzylindole. Org Synth 54, 58.
80. Horkko, S.; Miller, E.; Dudl, E.; Reaven, P.; Curtiss, L. K.; Zvaifler, N. J.;
Terkeltaub, R.; Pierangeli, S. S.; Branch, D. W.; Palinski, W.; Witztum, J. L.
(1996) Antiphospholipid antibodies are directed against epitopes of oxidized
phospholipids. Recognition of cardiolipin by monoclonal antibodies to
epitopes of oxidized low density lipoprotein. J Clin Invest 98 (3), 815-825.
245

81. Huang, Y.; Wang, K. K. (2001) The calpain family and human disease.
Trends Mol. Med. 7 (8), 355-362.
82. Huitorel, P.; Pantaloni, D. (1985) Bundling of microtubules by
glyceraldehyde-3-phosphate dehydrogenase and its modulation by ATP. Eur.
J. Biochem. 150 (2), 265-269.
83. Hunter, G. A.; McNab, H.; Withell, K. (2010) The Meldrum’s Acid Route to
Prodigiosin Analogues. Synthesis (10), 1707-1711.
84. Ikemoto, A.; Bole, D. G.; Ueda, T. (2003) Glycolysis and glutamate
accumulation into synaptic vesicles. Role of glyceraldehyde phosphate
dehydrogenase and 3-phosphoglycerate kinase. J. Biol. Chem. 278 (8), 5929-
5940.
85. Ishikawa, Y.; Yuki, E. (1974) Effect of trimethylamine oxide on the oxidation
of natural tocopherol mixtures. Yukagaku 23, 710-713.
86. Iyer, R. S.; Ghosh, S.; Salomon, R. G. (1989) Levuglandin-E2 cross-links
proteins. Prostaglandins 37 (4), 471-480.
87. Iyer, R. S.; Kobierski, M. E.; Salomon, R. G. (1994) Generation of pyrroles in
the reaction of levuglandin E(2) with proteins. J. Org. Chem. 59 (20), 6038-
6043.
88. Jenuwein, T.; Allis, C. D. (2001) Translating the histone code. Science 293
(5532), 1074-1080.
89. Jirousek, M. R.; Murthi, K. K.; Salomon, R. G. (1990) Electrophilic
levuglandin E2-protein adducts bind glycine: A model for protein cross-
linking. Prostaglandins 40 (2), 187-203.
90. Kahn, M. G.; Giblin, F. J.; Epstein, D. L. (1983) Glutathione in calf trabecular
meshwork and its relation to aqueous humor outflow facility. Invest
Ophthalmol Vis Sci 24 (9), 1283-1287.
91. Kalnitsky, G.; Hummel, J. P.; Dierks, C. (1959) Some factors which affect the
enzymatic digestion of ribonucleic acid. J. Biol. Chem. 234 (6), 1512-1516.
92. Kitson, T. M. (1979) 2,2'-Dithiodipyridine activates aldehyde dehydrogenase
246

and protects the enzyme against inactivation by disulfiram. Biochem. J 183
(3), 751-753.
93. Kitteringham, N. R.; Jenkins, R. E.; Lane, C. S.; Elliott, V. L.; Park, B. K.
(2009) Multiple reaction monitoring for quantitative biomarker analysis in
proteomics and metabolomics. J Chromatogr B Analyt Technol Biomed Life
Sci 877 (13), 1229-1239.
94. Kobierski, M. E.; Kim, S. C.; Murthi, K. K.; Iyer, R. S.; Salomon, R. G.
(1994) Synthesis of a pyrazole isostere of pyrroles formed by the reaction of
the epsilon-amino groups of protein lysyl residues with levuglandin E(2). J.
Org. Chem. 59 (20), 6044-6050.
95. Koeth, R. A.; Wang, Z.; Levison, B. S.; Buffa, J. A.; Org, E.; Sheehy, B. T.;
Britt, E. B.; Fu, X.; Wu, Y.; Li, L.; Smith, J. D.; DiDonato, J. A.; Chen, J.; Li,
H.; Wu, G. D.; Lewis, J. D.; Warrier, M.; Brown, J. M.; Krauss, R. M.; Tang,
W. H.; Bushman, F. D.; Lusis, A. J.; Hazen, S. L. (2013) Intestinal microbiota
metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis.
Nat Med 19 (5), 576-585.
96. Kohler, G.; Milstein, C. (1975) Continuous cultures of fused cells secreting
antibody of predefined specificity. Nature 256 (5517), 495-497.
97. Laemmli, U. K. (1970) Cleavage of structural proteins during assembly of
head of bacteriophage-T4. Nature 227 (5259), 680-685.
98. Lambert, M. P.; Barlow, A. K.; Chromy, B. A.; Edwards, C.; Freed, R.;
Liosatos, M.; Morgan, T. E.; Rozovsky, I.; Trommer, B.; Viola, K. L.; Wals,
P.; Zhang, C.; Finch, C. E.; Krafft, G. A.; Klein, W. L. (1998) Diffusible,
nonfibrillar ligands derived from Abeta1-42 are potent central nervous system
neurotoxins. Proc Natl Acad Sci U S A 95 (11), 6448-6453.
99. Lametsch, R.; Lonergan, S.; Huff-Lonergan, E. (2008) Disulfide bond within
mu-calpain active site inhibits activity and autolysis. Biochim. Biophys. Acta
1784 (9), 1215-1221.
100. Lebedev, O. L.; Kazarnovskii, S. N. (1960) Catalytic oxidation of aliphatic
247

amines with hydrogen peroxide. Zh. Obshch. Khim. 30, 1631-1635.
101. Lee, J. W.; Fuda, H.; Javitt, N. B.; Strott, C. A.; Rodriguez, I. R. (2006)
Expression and localization of sterol 27-hydroxylase (CYP27A1) in monkey
retina. Exp Eye Res 83 (2), 465-469.
102. Lefkovits, I.; Waldeman, H., Limiting dilution analysis of cells in the immune
system. Cambridge University Press: Cambridge, 1979.
103. Li, W.; Laird, J. M.; Lu, L.; Roychowdhury, S.; Nagy, L. E.; Zhou, R.; Crabb,
J. W.; Salomon, R. G. (2009) Isolevuglandins covalently modify
phosphatidylethanolamines in vivo: detection and quantitative analysis of
hydroxylactam adducts. Free Radic Biol Med 47 (11), 1539-1552.
104. Liao, W. L.; Heo, G. Y.; Dodder, N. G.; Reem, R. E.; Mast, N.; Huang, S.;
Dipatre, P. L.; Turko, I. V.; Pikuleva, I. A. (2011) Quantification of
cholesterol-metabolizing P450s CYP27A1 and CYP46A1 in neural tissues
reveals a lack of enzyme-product correlations in human retina but not human
brain. J Proteome Res 10 (1), 241-248.
105. Liao, Y.; Wang, X.; Qian, W.; Li, Y.; Li, X.; Yu, D. G. (2012) Bulk synthesis,
optimization, and characterization of highly dispersible polypyrrole
nanoparticles toward protein separation using nanocomposite membranes. J.
Colloid Interface Sci. 386 (1), 148-157.
106. Lindner, A.; Charra, B.; Sherrard, D. J.; Scribner, B. H. (1974) Accelerated
atherosclerosis in prolonged maintenance hemodialysis. N Engl J Med 290
(13), 697-701.
107. Little, M.; Kipriyanov, S. M.; Le Gall, F.; Moldenhauer, G. (2000) Of mice
and men: hybridoma and recombinant antibodies. Immunol Today 21 (8), 364-
370.
108. Loader, C.; Anderson, H. J. (1971) Pyrrole Chemistry. Part XI. Some
Reactions of the Pyrrole Grignard Reagent with Alkyl Carbonates and Alkyl
Thiocarbonates. Can. J. Chem. 49 (1), 45-48.
109. Ma, Y. Nitroxides in Mechanistic Studies: Ageing of Gold Nanoparticles and
248

Nitroxide Transformation in Acids. PhD thesis, University of York., 2010.
110. Mapp, P. I.; Grootveld, M. C.; Blake, D. R. (1995) Hypoxia, oxidative stress
and rheumatoid arthritis. Br Med Bull 51 (2), 419-436.
111. Mast, N.; Reem, R.; Bederman, I.; Huang, S.; DiPatre, P. L.; Bjorkhem, I.;
Pikuleva, I. A. (2011) Cholestenoic Acid is an important elimination product
of cholesterol in the retina: comparison of retinal cholesterol metabolism with
that in the brain. Invest Ophthalmol Vis Sci 52 (1), 594-603.
112. Mezhuev, Y. O.; Korshak, Y. V.; Shtilman, M. I.; Piskareva, A. I. (2012)
Activation parameters of single-electron transfer from pyrrole molecule to
persulfate ion. (English). Izv V.U.Z. Khim Khim Tekhnol 55 (10), 42-45.
113. Min, J. Z.; Toyo'oka, T.; Kawanishi, H.; Fukushima, T.; Kato, M. (2005)
Novel fluorescent asparaginyl-N-acetyl-D-glucosamines (Asn-GlcNAc) for
the resolution of oligosaccharides in glycopeptides, based on enzyme
transglycosylation reaction. Anal. Chim. Acta 550 (1-2), 173-181.
114. Moldoveanu, T.; Hosfield, C. M.; Lim, D.; Elce, J. S.; Jia, Z. C.; Davies, P. L.
(2002) A Ca2+ switch aligns the active site of calpain. Cell 108 (5), 649-660.
115. Monroe, B. M. (1982) Photochemical estimation of oxygen solubility.
Photochem. Photobiol. 35 (6), 863-865.
116. Morrow, J. D.; Harris, T. M.; Roberts, L. J., 2nd (1990) Noncyclooxygenase
oxidative formation of a series of novel prostaglandins: analytical
ramifications for measurement of eicosanoids. Anal. Biochem. 184 (1), 1-10.
117. Morrow, J. D.; Hill, K. E.; Burk, R. F.; Nammour, T. M.; Badr, K. F.;
Roberts, L. J., 2nd (1990) A series of prostaglandin F2-like compounds are
produced in vivo in humans by a non-cyclooxygenase, free radical-catalyzed
mechanism. Proc Natl Acad Sci U S A 87 (23), 9383-9387.
118. Mountassif, D.; Baibai, T.; Fourrat, L.; Moutaouakkil, A.; Iddar, A.; El
Kebbaj, M. S.; Soukri, A. (2009) Immunoaffinity purification and
characterization of glyceraldehyde-3-phosphate dehydrogenase from human
erythrocytes. Acta Biochim Biophys Sin (Shanghai) 41 (5), 399-406.
249

119. Murov, S. L., Handbook of Photochemistry. Dekker: New York, 1973.
120. Murphy, M. E.; Kehrer, J. P. (1989) Oxidative stress and muscular dystrophy.
Chem. Biol. Interact. 69 (2-3), 101-173.
121. Murthi, K. K.; Friedman, L. R.; Oleinick, N. L.; Salomon, R. G. (1993)
Formation of DNA-protein cross-links in mammalian cells by levuglandin E2.
Biochemistry 32 (15), 4090-4097.
122. Murthi, K. K.; Salomon, R. G.; Sternlicht, H. (1990) Levuglandin E2 inhibits
mitosis and microtubule assembly. Prostaglandins 39 (6), 611-622.
123. Nagata, K.; Suzuki, H.; Sakaguchi, S. (2007) Common pathogenic mechanism
in development progression of liver injury caused by non-alcoholic or
alcoholic steatohepatitis. J Toxicol Sci 32 (5), 453-468.
124. Nakahara, Y.; Fujita, A.; Ogawa, T. (1985) An efficient synthesis of 2-
acylpyrroles. Agric Biol Chem 49 (5), 1491-1495.
125. Nakajima, H.; Amano, W.; Fujita, A.; Fukuhara, A.; Azuma, Y. T.; Hata, F.;
Inui, T.; Takeuchi, T. (2007) The active site cysteine of the proapoptotic
protein glyceraldehyde-3-phosphate dehydrogenase is essential in oxidative
stress-induced aggregation and cell death. J. Biol. Chem. 282 (36), 26562-
26574.
126. Nakanishi, S.; Tanaka, H.; Hioki, K.; Yamada, K.; Kunishima, M. (2010)
Labeling study of avidin by modular method for affinity labeling (MoAL).
Bioorg. Med. Chem. Lett. 20 (23), 7050-7053.
127. Nicolaou, K. C.; Claremon, D. A.; Papahatjis, D. P. (1981) A mild method for
the synthesis of 2-ketopyrroles from carboxylic-acids. Tetrahedron Lett. 22
(46), 4647-4650.
128. Novak, I.; Harrison, L. J.; Kovac, B.; Pratt, L. M. (2004) Electronic structure
of persistent radicals: nitroxides. J. Org. Chem. 69 (22), 7628-7634.
129. Ntambi, J. M.; Bene, H. (2001) Polyunsaturated fatty acid regulation of gene
expression. J Mol Neurosci 16 (2-3), 273-278; discussion 279-284.
130. Nugteren, D. H.; Vonkeman, H.; Van Dorp, D. A. Non‐enzymic conversion of
250

all‐cis 8,11,14‐eicosatrienoic acid into prostaglandin E1. Recl. Trav. Chim.
Pays-Bas 86 (11), 1237-1245.
131. Onorato, J. M.; Jenkins, A. J.; Thorpe, S. R.; Baynes, J. W. (2000)
Pyridoxamine, an inhibitor of advanced glycation reactions, also inhibits
advanced lipoxidation reactions. Mechanism of action of pyridoxamine. J.
Biol. Chem. 275 (28), 21177-21184.
132. Pappin, D. J. C.; Hojrup, P.; Bleasby, A. J. (1993) Rapid identification of
proteins by peptide-mass fingerprinting. Curr Biol 3 (6), 327-332.
133. Pikuleva, I. A. (2006) Cytochrome P450s and cholesterol homeostasis.
Pharmacol Ther 112 (3), 761-773.
134. Podrez, E. A.; Byzova, T. V.; Febbraio, M.; Salomon, R. G.; Ma, Y.;
Valiyaveettil, M.; Poliakov, E.; Sun, M.; Finton, P. J.; Curtis, B. R.; Chen, J.;
Zhang, R.; Silverstein, R. L.; Hazen, S. L. (2007) Platelet CD36 links
hyperlipidemia, oxidant stress and a prothrombotic phenotype. Nat Med 13
(9), 1086-1095.
135. Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton,
P. J.; Shan, L.; Febbraio, M.; Hajjar, D. P.; Silverstein, R. L.; Hoff, H. F.;
Salomon, R. G.; Hazen, S. L. (2002) A novel family of atherogenic oxidized
phospholipids promotes macrophage foam cell formation via the scavenger
receptor CD36 and is enriched in atherosclerotic lesions. J. Biol. Chem. 277
(41), 38517-38523.
136. Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton,
P. J.; Shan, L.; Gugiu, B.; Fox, P. L.; Hoff, H. F.; Salomon, R. G.; Hazen, S.
L. (2002) Identification of a novel family of oxidized phospholipids that serve
as ligands for the macrophage scavenger receptor CD36. J. Biol. Chem. 277
(41), 38503-38516.
137. Poliakov, E.; Meer, S. G.; Roy, S. C.; Mesaros, C.; Salomon, R. G. (2004) Iso
7 LGD(2) - Protein adducts are abundant in vivo and free radical-induced
oxidation of an arachidonyl phospholipid generates this D series
251

isolevuglandin in vitro. Chem. Res. Toxicol. 17 (5), 613-622.
138. Prados, P.; Fukushima, T.; Santa, T.; Homma, H.; Tsunoda, M.; AlKindy, S.;
Mori, S.; Yokosu, H.; Imai, K. (1997) 4-N,N-Dimethylaminosulfonyl-7-N-(2-
aminoethyl)amino-benzofurazan as a new precolumn fluorescence
derivatization reagent for carboxylic acids (fatty acids and drugs containing a
carboxyl moiety) in liquid chromatography. Anal. Chim. Acta 344 (3), 227-
232.
139. Ray, S. K.; Shields, D. C.; Saido, T. C.; Matzelle, D. C.; Wilford, G. G.;
Hogan, E. L.; Banik, N. L. (1999) Calpain activity and translational
expression increased in spinal cord injury. Brain Res 816 (2), 375-380.
140. Reinaud, O.; Capdevielle, P.; Maumy, M. (1990) Copper(II) mediated
aromatic hydroxylation by trimethylamine N-oxide. J Chem Soc Chem
Commun (7), 566-568.
141. Reinaud, O.; Capdevielle, P.; Maumy, M. (1991) ortho-Aryloxylation of N-
substituted benzamides: a new oxidizing process induced by the
copper(II)/trimethylamine N-oxide system. J Chem Soc Perkin Trans I (9),
2129-2134.
142. Reverter, D.; Strobl, S.; Fernandez-Catalan, C.; Sorimachi, H.; Suzuki, K.;
Bode, W. (2001) Structural basis for possible calcium-induced activation
mechanisms of calpains. Biol. Chem. 382 (5), 753-766.
143. Roberts, L. J., 2nd; Brame, C. J.; Chen, Y.; Morrow, J. D.; Salomon, R. G.
(1999) Formation of reactive products of the isoprostane pathway:
isolevuglandins and cyclopentenone isoprostanes. Adv Exp Med Biol 469,
335-341.
144. Roberts, L. J.; Salomon, R. G.; Morrow, J. D.; Brame, C. J. (1999) New
developments in the isoprostane pathway: identification of novel highly
reactive γ-ketoaldehydes (isolevuglandins) and characterization of their
protein adducts. FASEB J. 13 (10), 1157-1168.
145. Salomon, R. G. (2005) Levuglandins and isolevuglandins: Stealthy toxins of
252

oxidative injury. Antioxid Redox Signal 7 (1-2), 185-201.
146. Salomon, R. G. (2005) Distinguishing levuglandins produced through the
cyclooxygenase and isoprostane pathways. Chem. Phys. Lipids 134 (1), 1-20.
147. Salomon, R. G.; Batyreva, E.; Kaur, K.; Sprecher, D. L.; Schreiber, M. J.;
Crabb, J. W.; Penn, M. S.; DiCorletoe, A. M.; Hazen, S. L.; Podrez, E. A.
(2000) Isolevuglandin-protein adducts in humans: products of free radical-
induced lipid oxidation through the isoprostane pathway. Biochim. Biophys.
Acta 1485 (2-3), 225-235.
148. Salomon, R. G.; Jirousek, M. R.; Ghosh, S.; Sharma, R. B. (1987)
Prostaglandin endoperoxides .21. covalent binding of levuglandin-E2 with
proteins. Prostaglandins 34 (5), 643-656.
149. Salomon, R. G.; Kaur, K.; Batyreva, E. (2000) Isolevuglandin-protein
Adducts in Oxidized Low Density Lipoprotein and Human Plasma: A Strong
Connection with Cardiovascular Disease. Trends Cardiovasc Med 10 (2), 53-
59.
150. Salomon, R. G.; Miller, D. B. (1985) Levuglandins: isolation,
characterization, and total synthesis of new secoprostanoid products from
prostaglandin endoperoxides. Adv Prostaglandin Thromboxane Leukot Res
15, 323-326.
151. Salomon, R. G.; Salomon, M. F. (1977) 2,3-Dioxabicyclo [2.2.1] heptane. The
strained bicyclic peroxide nucleus of prostaglandin endoperoxides. J. Am.
Chem. Soc. 99 (10), 3501-3503.
152. Salomon, R. G.; Sha, W.; Brame, C.; Kaur, K.; Subbanagounder, G.; O'Neil,
J.; Hoff, H. F.; Roberts, L. J. (1999) Protein adducts of iso 4 levuglandin E-2,
a product of the isoprostane pathway, in oxidized low density lipoprotein. J.
Biol. Chem. 274 (29), 20271-20280.
153. Salomon, R. G.; Subbanagounder, G.; O'Neil, J.; Kaur, K.; Smith, M. A.;
Hoff, H. F.; Perry, G.; Monnier, V. M. (1997) Levuglandin E2-protein adducts
in human plasma and vasculature. Chem. Res. Toxicol. 10 (5), 536-545.
253

154. Salomon, R. G.; Subbanagounder, G.; Singh, U.; O'Neil, J.; Hoff, H. F. (1997)
Oxidation of low-density lipoproteins produces levuglandin-protein adducts.
Chem. Res. Toxicol. 10 (7), 750-759.
155. Sambarkar, P. B.; Patil, A. C. (2012) Synthesis of amides from acid and amine
using coupling reagents. J Curr Pharm Res 10 (1), 22-24.
156. Samuni, A.; Goldstein, S.; Russo, A.; Mitchell, J. B.; Krishna, M. C.; Neta, P.
(2002) Kinetics and mechanism of hydroxyl radical and OH-adduct radical
reactions with nitroxides and with their hydroxylamines. J. Am. Chem. Soc.
124 (29), 8719-8724.
157. Santiard-Baron, D.; Lacoste, A.; Ellouk-Achard, S.; Soulie, C.; Nicole, A.;
Sarasin, A.; Ceballos-Picot, I. (2001) The amyloid peptide induces early
genotoxic damage in human preneuron NT2. Mutat Res 479 (1-2), 113-120.
158. Sayre, L. M.; Zelasko, D. A.; Harris, P. L.; Perry, G.; Salomon, R. G.; Smith,
M. A. (1997) 4-Hydroxynonenal-derived advanced lipid peroxidation end
products are increased in Alzheimer's disease. J. Neurochem. 68 (5), 2092-
2097.
159. Sazontova, T. G.; Arkhipenko, I. V. (2005) The role of free radical processes
and redox-signalization in adaptation of the organism to changes in oxygen
level. Ross Fiziol Zh Im I M Sechenova 91 (6), 636-655.
160. Schulte-Elte, K. H.; Gadola, M.; Ohloff, G. (1973) Oxydative Umsetzungen in
der Damascon-Reihe. 1ΔgO2- und SeO2-Oxydation von β-Damascenon. Helv.
Chim. Acta 56 (6), 2028-2035.
161. Selkoe, D. J. (2001) Alzheimer's disease: genes, proteins, and therapy. Physiol
Rev 81 (2), 741-766.
162. Shao, X. G.; Fernandez, I.; Sudhof, T. C.; Rizo, J. (1998) Solution structures
of the Ca2+-free and Ca2+-bound C(2)A domain of synaptotagmin I: Does Ca2+
induce a conformational change? Biochemistry 37 (46), 16106-16115.
163. Shattock, M. J.; Matsuura, H. (1993) Measurement of Na+-K+ pump current in
isolated rabbit ventricular myocytes using the whole-cell voltage-clamp
254

technique. Inhibition of the pump by oxidant stress. Circ Res 72 (1), 91-101.
164. Shi, H. C.; Li, Y. (2007) Formation of nitroxide radicals from secondary
amines and peracids: A peroxyl radical oxidation pathway derived from
electron spin resonance detection and density functional theory calculation. J
Mol Catal A Chem 271 (1–2), 32-41.
165. Shogren-Knaak, M.; Ishii, H.; Sun, J. M.; Pazin, M. J.; Davie, J. R.; Peterson,
C. L. (2006) Histone H4-K16 acetylation controls chromatin structure and
protein interactions. Science 311 (5762), 844-847.
166. Shringarpure, R.; Grune, T.; Sitte, N.; Davies, K. J. (2000) 4-
Hydroxynonenal-modified amyloid-beta peptide inhibits the proteasome:
possible importance in Alzheimer's disease. Cell Mol Life Sci 57 (12), 1802-
1809.
167. Sorimachi, H.; Ishiura, S.; Suzuki, K. (1997) Structure and physiological
function of calpains. Biochem. J 328, 721-732.
168. Sorimachi, H.; Suzuki, K. (2001) The structure of calpain. J. Biochem. 129
(5), 653-664.
169. Sparrow, J. R.; Vollmer-Snarr, H. R.; Zhou, J.; Jang, Y. P.; Jockusch, S.;
Itagaki, Y.; Nakanishi, K. (2003) A2E-epoxides damage DNA in retinal
pigment epithelial cells. Vitamin E and other antioxidants inhibit A2E-
epoxide formation. J. Biol. Chem. 278 (20), 18207-18213.
170. Stavrovskaya, I. G.; Baranov, S. V.; Guo, X.; Davies, S. S.; Roberts, L. J.,
2nd; Kristal, B. S. (2010) Reactive gamma-ketoaldehydes formed via the
isoprostane pathway disrupt mitochondrial respiration and calcium
homeostasis. Free Radic Biol Med 49 (4), 567-579.
171. Still, W. C.; Kahn, M.; Mitra, A. (1978) Rapid chromatographic technique for
preparative separations with moderate resolution. J. Org. Chem. 43 (14),
2923-2925.
172. Straight, R. C.; Spikes, J. D., Singlet Oxygen. CRC: Boca Raton, FL, 1985;
Vol. 4.
255

173. Subbanagounder, G.; Salomon, R. G.; Murthi, K. K.; Brame, C.; Roberts, L. J.
(1997) Total synthesis of Iso 4 -levuglandin E-2. J. Org. Chem. 62 (22), 7658-
7666.
174. Sullivan, C. B.; Matafonova, E.; Roberts, L. J., II; Amarnath, V.; Davies, S. S.
(2010) Isoketals form cytotoxic phosphatidylethanolamine adducts in cells. J.
Lipid Res. 51 (5), 999-1009.
175. Sun, M.; Finnemann, S. C.; Febbraio, M.; Shan, L.; Annangudi, S. P.; Podrez,
E. A.; Hoppe, G.; Darrow, R.; Organisciak, D. T.; Salomon, R. G.; Silverstein,
R. L.; Hazen, S. L. (2006) Light-induced oxidation of photoreceptor outer
segment phospholipids generates ligands for CD36-mediated phagocytosis by
retinal pigment epithelium: a potential mechanism for modulating outer
segment phagocytosis under oxidant stress conditions. J. Biol. Chem. 281 (7),
4222-4230.
176. Suzuki, K.; Hata, S.; Kawabata, Y.; Sorimachi, H. (2004) Structure,
activation, and biology of calpain. Diabetes 53, S12-S18.
177. Tak, P. P.; Zvaifler, N. J.; Green, D. R.; Firestein, G. S. (2000) Rheumatoid
arthritis and p53: how oxidative stress might alter the course of inflammatory
diseases. Immunol Today 21 (2), 78-82.
178. Tang, W. H.; Wang, Z.; Levison, B. S.; Koeth, R. A.; Britt, E. B.; Fu, X.; Wu,
Y.; Hazen, S. L. (2013) Intestinal microbial metabolism of
phosphatidylcholine and cardiovascular risk. N Engl J Med 368 (17), 1575-
1584.
179. Tisdale, E. J. (2001) Glyceraldehyde-3-phosphate dehydrogenase is required
for vesicular transport in the early secretory pathway. J. Biol. Chem. 276 (4),
2480-2486.
180. Tokunaga, J. (1975) Solubilities of oxygen, nitrogen, and carbon dioxide in
aqueous alcohol solutions. J. Chem. Eng. Data 20 (1), 41-46.
181. Tong, L.; Thornton, P. L.; Balazs, R.; Cotman, C. W. (2001) Beta -amyloid-
(1-42) impairs activity-dependent cAMP-response element-binding protein
256

signaling in neurons at concentrations in which cell survival Is not
compromised. J. Biol. Chem. 276 (20), 17301-17306.
182. Toshniwal, P. K.; Zarling, E. J. (1992) Evidence for increased lipid
peroxidation in multiple sclerosis. Neurochem. Res. 17 (2), 205-207.
183. Uchida, K.; Stadtman, E. R. (1993) Covalent attachment of 4-hydroxynonenal
to glyceraldehyde-3-phosphate dehydrogenase. A possible involvement of
intra- and intermolecular cross-linking reaction. J. Biol. Chem. 268 (9), 6388-
6393.
184. Varadarajan, S.; Kanski, J.; Aksenova, M.; Lauderback, C.; Butterfield, D. A.
(2001) Different mechanisms of oxidative stress and neurotoxicity for
Alzheimer's A beta(1--42) and A beta(25--35). J. Am. Chem. Soc. 123 (24),
5625-5631.
185. Varadarajan, S.; Yatin, S.; Aksenova, M.; Butterfield, D. A. (2000) Review:
Alzheimer's amyloid beta-peptide-associated free radical oxidative stress and
neurotoxicity. J Struct Biol 130 (2-3), 184-208.
186. Wang, Z.; Klipfell, E.; Bennett, B. J.; Koeth, R.; Levison, B. S.; Dugar, B.;
Feldstein, A. E.; Britt, E. B.; Fu, X.; Chung, Y. M.; Wu, Y.; Schauer, P.;
Smith, J. D.; Allayee, H.; Tang, W. H.; DiDonato, J. A.; Lusis, A. J.; Hazen,
S. L. (2011) Gut flora metabolism of phosphatidylcholine promotes
cardiovascular disease. Nature 472 (7341), 57-63.
187. Ward, M. (1996) Amino acid analysis using precolumn derivatization with 6-
aminoquinolyl-n-hydroxysuccinimidyl carbamate. The protein protocols
handbook, 461-465.
188. Williams, C. H., Jr, Chemistry and biochemistry of flavoenzymes. Müller, F.,
Ed. CRC Press: 1991; pp 121-211.
189. Yatin, S. M.; Varadarajan, S.; Butterfield, D. A. (2000) Vitamin E Prevents
Alzheimer's Amyloid beta-Peptide (1-42)-Induced Neuronal Protein
Oxidation and Reactive Oxygen Species Production. J Alzheimers Dis 2 (2),
123-131.
257

190. Yatin, S. M.; Varadarajan, S.; Link, C. D.; Butterfield, D. A. (1999) In vitro
and in vivo oxidative stress associated with Alzheimer's amyloid beta-peptide
(1-42). Neurobiol Aging 20 (3), 325-330; discussion 339-342.
191. Yatin, S. M.; Yatin, M.; Aulick, T.; Ain, K. B.; Butterfield, D. A. (1999)
Alzheimer's amyloid beta-peptide associated free radicals increase rat
embryonic neuronal polyamine uptake and ornithine decarboxylase activity:
protective effect of vitamin E. Neurosci Lett 263 (1), 17-20.
192. Yatin, S. M.; Yatin, M.; Varadarajan, S.; Ain, K. B.; Butterfield, D. A. (2001)
Role of spermine in amyloid beta-peptide-associated free radical-induced
neurotoxicity. J. Neurosci. Res. 63 (5), 395-401.
193. Zagol-Ikapitte, I.; Bernoud-Hubac, N.; Amarnath, V.; Roberts, L. J., 2nd;
Boutaud, O.; Oates, J. A. (2004) Characterization of bis(levuglandinyl) urea
derivatives as products of the reaction between prostaglandin H2 and arginine.
Biochemistry 43 (18), 5503-5510.
194. Zagol-Ikapitte, I.; Masterson, T. S.; Amarnath, V.; Montine, T. J.;
Andreasson, K. I.; Boutaud, O.; Oates, J. A. (2005) Prostaglandin H2-derived
adducts of proteins correlate with Alzheimer's disease severity. J. Neurochem.
94 (4), 1140-1145.
195. Zagorski, M. G.; Salomon, R. G. (1980) Prostaglandin endoperoxides. 11.
Mechanism of amine-catalyzed fragmentation of 2,3-
dioxabicyclo[2.2.1]heptane. J. Am. Chem. Soc. 102 (7), 2501-2503.
196. Zagorski, M. G.; Salomon, R. G. (1982) Prostaglandin endoperoxides. 12.
Carboxylate catalysis and the effects of proton donors on the decomposition
of 2,3-dioxabicyclo[2.2.1]heptane. J. Am. Chem. Soc. 104 (12), 3498-3503.
197. Zemski Berry, K. A.; Murphy, R. C. (2007) Characterization of acrolein-
glycerophosphoethanolamine lipid adducts using electrospray mass
spectrometry. Chem. Res. Toxicol. 20 (9), 1342-1351.
198. Zhu, M.; Spink, D. C.; Yan, B.; Bank, S.; DeCaprio, A. P. (1994) Formation
and structure of cross-linking and monomeric pyrrole autoxidation products in
258

2,5-hexanedione-treated amino acids, peptides, and protein. Chem. Res.
Toxicol. 7 (4), 551-558.
199. Zhu, M.; Spink, D. C.; Yan, B.; Bank, S.; DeCaprio, A. P. (1995) Inhibition of
2,5-hexanedione-induced protein cross-linking by biological thiols: chemical
mechanisms and toxicological implications. Chem. Res. Toxicol. 8 (5), 764-
771.
259




















