
Isolation and phylogenetic identification of a dark-septate fungus associated with the alpine plant...
7
INTERNATIONAL JOURNAL OF AGRICULTURE & BIOLOGY ISSN Print: 1560–8530; ISSN Online: 1814–9596 13–1198/2014/16–3–564–570 http://www.fspublishers.org Full Length Article To cite this paper: Roohi, A., I. Ahmed, N. Khalid, M. Iqbal and M. Jamil, 2014. Isolation and phylogenetic identification of halotolerant/halophilic bacteria from the salt mines of Karak, Pakistan, Int. J. Agric. Biol., 16: 564‒570 Isolation and Phylogenetic Identification of Halotolerant/Halophilic Bacteria from the Salt Mines of Karak, Pakistan Aneela Roohi 1,2 , Iftikhar Ahmed 1* , Nauman Khalid 3 , Muhammad Iqbal 1 and Muhammad Jamil 2* 1 National Institute for Genomics and Advanced Biotechnology (NIGAB), National Agricultural Research Centre (NARC), Park Road, Islamabad-45500, Pakistan 2 Department of Biotechnology and Genetic Engineering, Kohat University of Science and Technology, Pakistan 3 Graduate School of Agricultural and Life Sciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku 113-8657, Tokyo, Japan *For correspondences: [email protected]; [email protected] Abstract Extreme environments like salt mines are inhabited by a variety of bacteria that are well-adapted to such environments. The bacterial populations provide economic benefits in terms of enzymes synthesis. The salt mines of Karak region in Pakistan are extremely saline and the microbial communities found here have not yet been explored. In the present study, 57 halotolerant/halophilic bacterial strains were isolated from the salt mines of Karak. These strains were grown in media with 0- 35% NaCl concentration. The morphological and physiological characteristics of the isolated strains were studied to optimize the growth conditions and to classify the isolated bacterial strains into slightly halotolerant/halophilic, moderately halophilic and extreme halophilic. The phylogenetic analyses inferred from 16S rRNA gene sequence of the isolated strains demonstrated that the major population were closely related to species belonging to Planococcus, Jeotgalicoccus, Staphylococcus, Halobacillus, Halomonas, Brevibacterium, Gracilibacillus, Kocuria, Salinivibrio, Salinicoccus, Oceanobacillus and Bacillus genera. Results showed that the salt mines of Karak region are rich in halotolerant/halophilic bacterial population with diverse bacterial communities, which may be utilized in various industrial applications after proper screening and identification. © 2014 Friends Science Publishers Keywords: Extremophile; Halotolerant; Halophilic; Karak region; 16S rRNA gene sequence Introduction Bacteria play an important role in the development of viable habitats. Saline environments harbor taxonomically diverse group of bacteria that show improved structural and physiological characteristics in extreme (saline) conditions. To survive in saline environments, bacteria developed adoptive mechanisms like surface layer modifications especially in cellular phospholipid composition (Ventosa et al., 2008). An increasing interest in microorganisms from hyper-saline environments has resulted in the discovery of several new species and genera in Bacteria and Archaea domains (Woese and Fox, 1977). Extremely halophilic Archaea belong to the order Halobacteriales, which contains one family, the Halobacteriaceae (Ozcan et al., 2007). The family Halobacteriaceae belongs to domain Archaea and presently comprised of 40 genera (Euzéby, 2013). Among the halophilic microorganisms, Archaea are the most common (Antón et al., 2000; Oren, 2002). Microorganisms that form part of domain Archaea require high salt concentration for growth and are coined as halobacteria (Kamekura, 1998). These groups of microbes have been isolated from diverse environments like salterns, salt mines, salted food, saline lakes, salt marshes and saline desert soils (Ventosa et al., 2008). Halobacteria are either Gram-negative or Gram-positive and have aerobic, facultative anaerobic or obligatory anaerobic metabolism (Johnson et al., 2007). Halophilic bacteria have the ability to grow in media containing 3 to 15% (w/v) NaCl (Oren and Rodrı ́ guez-Valera, 2001; Oren, 2002). Halophilic microorganisms have many biotechnological applications like β-carotene production from fermented foods, hydrolytic enzymes or exopolysaccharides, stabilizers and other valuable compounds (Quesada et al., 1982, 2004;Ventosa et al., 2008). Bahadur Kheil and Jatta Ismail Kheil are two of the biggest and well protected salt mines in Karak District, Khyberpakhtun Khwa (KPK) Province of Pakistan, and it has an estimated salt reserve of about 52,563 tons (PMDC, 2013). Being of high quality, Karak rock salt is consumed as table salt after a simple treatment instead of complex refining. However, there are insufficient reports on exploration of bacterial diversity with respect to halophilic/ halotolerant characteristics from Pakistan (Roohi et al., 2012). Exploration of such diversity might result in considerable economic benefits in food and leather
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Isolation and phylogenetic identification of a dark-septate fungus associated with the alpine plant...

INTERNATIONAL JOURNAL OF AGRICULTURE & BIOLOGY
ISSN Print: 1560–8530; ISSN Online: 1814–9596
13–1198/2014/16–3–564–570
http://www.fspublishers.org
Full Length Article
To cite this paper: Roohi, A., I. Ahmed, N. Khalid, M. Iqbal and M. Jamil, 2014. Isolation and phylogenetic identification of halotolerant/halophilic bacteria
from the salt mines of Karak, Pakistan, Int. J. Agric. Biol., 16: 564‒570
Isolation and Phylogenetic Identification of Halotolerant/Halophilic
Bacteria from the Salt Mines of Karak, Pakistan
Aneela Roohi1,2
, Iftikhar Ahmed1*
, Nauman Khalid3, Muhammad Iqbal
1 and Muhammad Jamil
2*
1National Institute for Genomics and Advanced Biotechnology (NIGAB), National Agricultural Research Centre
(NARC), Park Road, Islamabad-45500, Pakistan 2Department of Biotechnology and Genetic Engineering, Kohat University of Science and Technology, Pakistan
3Graduate School of Agricultural and Life Sciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku 113-8657, Tokyo, Japan
*For correspondences: [email protected]; [email protected]
Abstract
Extreme environments like salt mines are inhabited by a variety of bacteria that are well-adapted to such environments. The
bacterial populations provide economic benefits in terms of enzymes synthesis. The salt mines of Karak region in Pakistan are
extremely saline and the microbial communities found here have not yet been explored. In the present study, 57
halotolerant/halophilic bacterial strains were isolated from the salt mines of Karak. These strains were grown in media with 0-
35% NaCl concentration. The morphological and physiological characteristics of the isolated strains were studied to optimize
the growth conditions and to classify the isolated bacterial strains into slightly halotolerant/halophilic, moderately halophilic
and extreme halophilic. The phylogenetic analyses inferred from 16S rRNA gene sequence of the isolated strains
demonstrated that the major population were closely related to species belonging to Planococcus, Jeotgalicoccus,
Staphylococcus, Halobacillus, Halomonas, Brevibacterium, Gracilibacillus, Kocuria, Salinivibrio, Salinicoccus,
Oceanobacillus and Bacillus genera. Results showed that the salt mines of Karak region are rich in halotolerant/halophilic
bacterial population with diverse bacterial communities, which may be utilized in various industrial applications after proper
screening and identification. © 2014 Friends Science Publishers
Keywords: Extremophile; Halotolerant; Halophilic; Karak region; 16S rRNA gene sequence
Introduction
Bacteria play an important role in the development of viable
habitats. Saline environments harbor taxonomically diverse
group of bacteria that show improved structural and
physiological characteristics in extreme (saline) conditions.
To survive in saline environments, bacteria developed
adoptive mechanisms like surface layer modifications
especially in cellular phospholipid composition (Ventosa et
al., 2008). An increasing interest in microorganisms from
hyper-saline environments has resulted in the discovery of
several new species and genera in Bacteria and Archaea
domains (Woese and Fox, 1977). Extremely halophilic
Archaea belong to the order Halobacteriales, which contains
one family, the Halobacteriaceae (Ozcan et al., 2007). The
family Halobacteriaceae belongs to domain Archaea and
presently comprised of 40 genera (Euzéby, 2013).
Among the halophilic microorganisms, Archaea are
the most common (Antón et al., 2000; Oren, 2002).
Microorganisms that form part of domain Archaea require
high salt concentration for growth and are coined as
halobacteria (Kamekura, 1998). These groups of microbes
have been isolated from diverse environments like salterns,
salt mines, salted food, saline lakes, salt marshes and saline
desert soils (Ventosa et al., 2008). Halobacteria are either
Gram-negative or Gram-positive and have aerobic,
facultative anaerobic or obligatory anaerobic metabolism
(Johnson et al., 2007). Halophilic bacteria have the ability to
grow in media containing 3 to 15% (w/v) NaCl (Oren and
Rodrı́guez-Valera, 2001; Oren, 2002). Halophilic
microorganisms have many biotechnological applications
like β-carotene production from fermented foods, hydrolytic
enzymes or exopolysaccharides, stabilizers and other
valuable compounds (Quesada et al., 1982, 2004;Ventosa et
al., 2008).
Bahadur Kheil and Jatta Ismail Kheil are two of the
biggest and well protected salt mines in Karak District,
Khyberpakhtun Khwa (KPK) Province of Pakistan, and it
has an estimated salt reserve of about 52,563 tons (PMDC,
2013). Being of high quality, Karak rock salt is consumed as
table salt after a simple treatment instead of complex
refining. However, there are insufficient reports on
exploration of bacterial diversity with respect to halophilic/
halotolerant characteristics from Pakistan (Roohi et al.,
2012). Exploration of such diversity might result in
considerable economic benefits in food and leather

Diversity of Halotolerant/Halophilic Bacteria / Int. J. Agric. Biol., Vol. 16, No. 3, 2014
565
industries. The objective of this research was to explore the
novelty associated with extremely halotolerant/halophilic
aerobic or facultative anaerobic bacteria and to assess the
bacterial biodiversity of these extremophiles from Pakistan
using molecular techniques that has not been previously
reported.
Materials and Methods
Collection of Samples
The samples were collected from the Karak Salt Mines of
KPK Province, Pakistan. The area is situated at 32° 47 to
33° 28 North and 70° 30 to 71° 30 East. The sampling sites
were divided into two different areas according to the
difference in location of salt quarries, which are located at
Bahadur Khel and Jatta Ismail Khel. Different salt mines
were selected for the sampling (Fig. 1). The salt beds are
approximately 105 m thick in Bahadur Khel, while more
than 30 m thick in Jatta Ismail Khel. Different rock salt was
collected from 5 rock salt mines. The samples were
randomly taken in sterile plastic bags and vials. These
samples were transported to the lab for the isolation of
bacteria and processing for further analysis.
Physicochemical Analysis of Brine Samples
Physico-chemical properties of the brine samples taken
from the salt mines (Bahadur Khel and Jatta Ismail Khel)
were determined including moisture contents, water
insoluble impurities SO42–
, Cl–, HCO3
–, CO3
–2 with standard
procedures while Na+, K
+, Ca
2+, Mg
2+, and some trace
elements like Zn2+
, Cu2+
, Fe2+
, Mn2+
, Pb2+
, Cr2+
and Cd2+
were determined with atomic absorption spectrophotometer
(Z-8000, Hitachi, Japan). The pH meter (Model, 3510
Jenway) was employed for pH measurement, while
laboratory thermometer was used to determine temperature
on site.
Isolation and Enrichment of Halophilic Bacteria
To recover moderately to extremely halotolerant/halophilic
bacteria, enrichment cultures and isolation procedures were
performed by dilution plate method on tryptic soy agar
medium. 1-2 mL of diluted sample was streaked on agar
medium containing various concentrations (5-25%, w/v) of
NaCl and incubated at 28ºC. The isolated strains were sub-
cultured several times under same conditions to obtain pure
cultures of morphologically different bacteria. The purified
strains were stored at – 80ºC for further characterization.
Characterization and Identification of Halotolerant
Bacterial Strains
The purified bacterial colonies were morphologically
characterized for colony color, form, elevation, margin etc.
Bacterial strains were characterized on the basis of Gram’s
staining, cell morphology and motility using microscope
(Olympus, CX31 equipped with Digital Camera 5A). The
growth characteristics of bacterial strains were determined
at various pH range (4-11), NaCl concentrations (0-40%)
and temperatures (4, 10, 20, 28, 35, 37, 40, 45 and 50ºC).
PCR Amplification and Sequencing of 16S rRNA Gene
Genomic DNA extraction for amplification of 16S rRNA
gene was performed as described by Roohi et al. (2012)
by suspending few isolated colonies in TE buffer in a
micro-centrifuge tube. These cells were heated for 10 min at
95ºC and were centrifuged at 6,000 rpm for 5 min.
Table 1: Chemical analysis of rock salt samples collected
from different location from Bahadur Khel and Jatta Ismail
Khel salt mines
Element (%)
Sulfur (SO3-2) 1.31
Calcium (CaO) 1.73
Potassium (K+) 1.65
Magnesium (Mg+2) 0.11
Chloride (Cl-1) 53.91
Sodium (Na+) 34.70
Moisture 0.13
Water insoluble impurities 7.98
Trace elements (mg/Kg)
Zinc (Zn) 0.15
Copper (Cu) 0.02
Iron (Fe) 0.57
Manganese (Mn) 0.00
Lead (Pb) 0.02
Chromium (Cr) 0.37
Cadmium (Cd) 0.00
Fig. 1: Location of sample collection sites in Karak Salt
Mines, Khyber Pakhtunkhwa Province, Pakistan

Roohi et al., / Int. J. Agric. Biol., Vol. 16, No. 3, 2014
566
The supernatant was used as template DNA for the
amplification of 16S rRNA gene. The 16S rRNA gene of all
the isolated strains was amplified by polymerase chain
reaction using primers 9F (5'-
GAGTTTGATCCTGGCTCAG-3') and 1510R (5'-
GGCTACCTTGTTACGA-3') using Premix ExTaq
(Takara, Japan) following protocol described previously
(Ahmed et al., 2007). The PCR was carried out in ABI
Veriti PCR Machine (Applied Biosystems, USA) using
optimized PCR Program: initial denaturation at 94ºC for 2
min; 30 cycles of denaturation at 94ºC for 1 min, annealing
at 50ºC for 1 min and extension at 72ºC for 1:30 min. The
final extension was performed at 72ºC for 5 min. The
amplified PCR products of 16S rRNA gene of bacterial
strains were purified and sequenced using the primers 27F
(5' AGAGTTTGATCMTGGCTCAG-3') and 1492R (5'-
ACCTTGTTACGACTT-3') using commercial service of
Macrogen Inc. Korea (http://dna.macrogen.com/eng/).
Phylogenetic Analysis of Bacterial Isolates
BioEdit software (Hall, 1999) was used to assemble the
fragment sequences of 16S rRNA gene. The 16S rRNA
gene sequences were submitted to DDBJ
(http://www.ddbj.nig.ac.jp). Using 16S rRNA gene
sequences, the strains were identified by BLAST search on
EzBiocloud Server). Closely related type species sequences
were retrieved to assess the molecular evolutionary relations
and to construct phylogenetic tree using MEGA version 5
(Tamura et al., 2011). A phylogenetic tree was constructed
from unambiguously aligned nucleotides using the
neighbour-joining (NJ) algorithm. The relationship stability
was evaluated by boot strap analysis performed using 1000
re-samplings of the neighbour-joining data for the tree
topology.
Results
Physical and Chemical Properties of Samples
The rock salt samples named M1, M2, M3, M4 and M5 were
collected from 5 different sites. The results indicated that the
mineral content, pH and temperature of samples were
suitable for the growth of halophilic bacteria. The average
temperature of all samples ranged from 23.8 to 34.7°C. The
above temperature varied according to sampling sites and
changes in air temperature. The chemical analysis showed
that major components in samples were Cl- (53.91%) and
Na+ (34.70%). Small amount of Mg
2+ (0.11%) and trace
elements were also detected from rock salt. Cd2+
was absent
from observed salt samples (Table 1). The average pH of
samples ranged between 7.8 to 7.95.
Colony, Cell Morphology and Density of
Halotolerant/Halophilic Bacteria
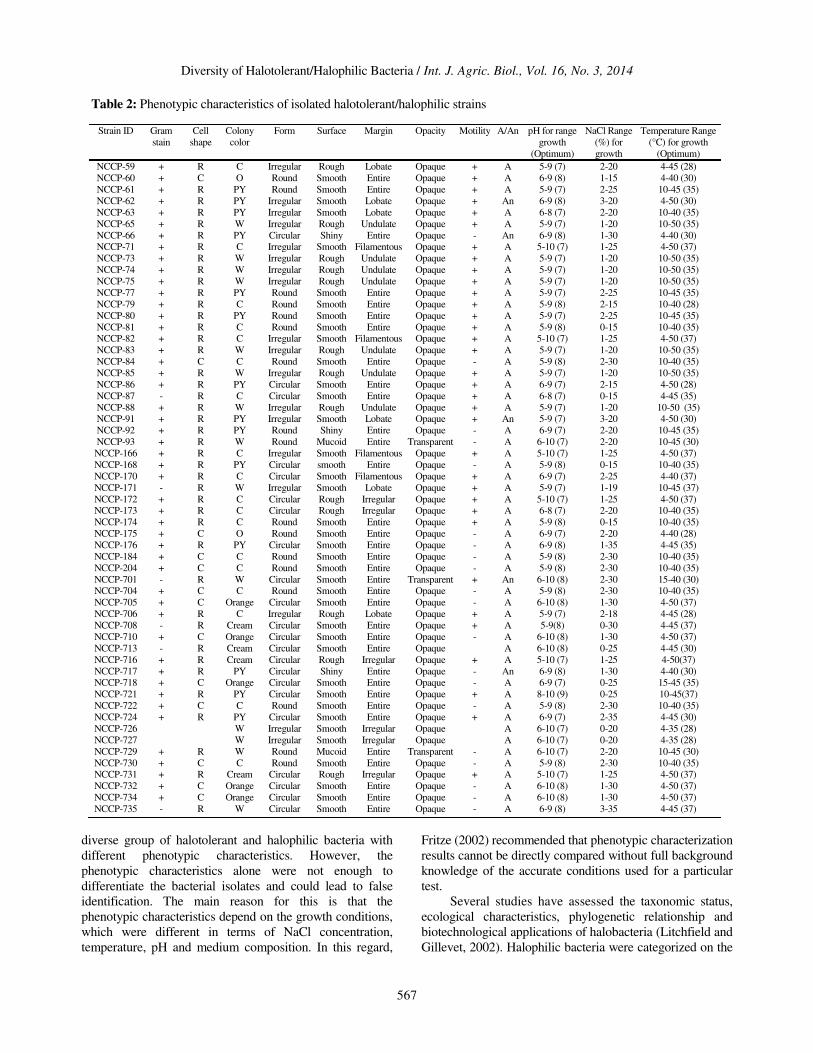
Fifty seven strains were isolated and purified on the basis
of morphology. Pigmentation of the strains included pale
yellow, creamy and whitish colonies. Most of the colonies
had entire margin, while some had undulate and wavy
filamentous margins (Table 2). Few of them also had lobate
margins. The growth conditions of all the isolated strains
were optimized for pH, NaCl tolerance and temperature.
Our results showed that the bacterial strains grew best at pH
7 to 9 and at a temperature range of 28-37oC. The isolated
strains toleratedNaCl up to 0-30% and based on
optimization conditions the isolated strains were classified
as slightly halotolerant (0-5% NaCl), moderately halophilic
(5-15% NaCl) and extremely halophilic (15-35% NaCl).
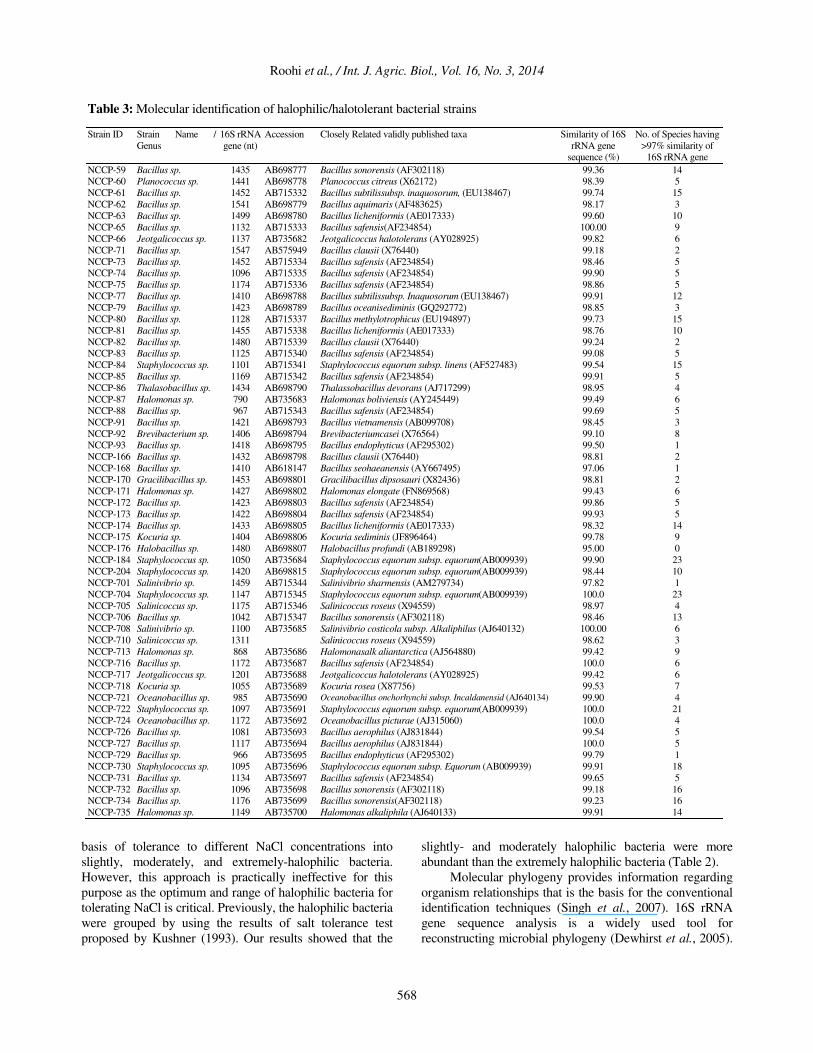
Phylogenetic Analysis
Phylogenetic analysis of the halophilic isolated strains
were performed by constructing a phylogenetic tree based
on the 16S rRNA gene sequences (Fig. 2). Phylogenetic
analysis indicated that the majority of isolated strains
belonged to the genera, Planococcus, Jeotgalicoccus,
Staphylococcus, Thalassobacillus, Halobacillus,
Halomonas, Brevibacterium, Gracilibacillus, Kocuria,
Salinivibrio, Salinicoccus ,Oceanobacillus and Bacillus. All
strains shared more than 97% identity with their closest
phylogenetic relatives except the strain NCCP-176. The
phylogenetic analyses showed that eight strains (NCCP-71,
91, 93, 168, 176, 701, 705, 710) can further be studied
taxonomically to delineate as novel species. The
phylogenetic analysis reflected the evolutionary
relationships among halophilic bacteria (Table 3).
Discussion
In the present study, we isolated and characterized
extremely halotolerant/halophilic aerobic or facultative
anaerobic bacteria from Pakistan using molecular
techniques that has not been previously reported. The above
growth condition are in line with several other studies that
concluded that most species show optimum growth at 3.5 to
4.5 M NaCl and pH 7.0 to 7.5 (Kushner, 1985; Oren and
Rodrı́guez-Valera, 2001). Similarly, our results of chemical
analysis of rock salt showed that these contain proper
amounts of essential ions to support the growth of extremely
halophilic microorganisms (Mancinelli and Hochstein,
1986; Oren, 1993; Birbir et al., 2007).
The colonial morphology of the isolated strains were
described using standard microbiological criteria. These
standards are based on colonial pigmentation, opacity,
diameter, consistency and elevation (Oren et al., 1997). In
these studies, we observed polymorphism with respect to
colonial morphology of the isolated strains as reported by
Castillo et al. (2007). Halobacterial members exhibited
various tones pigmentation ranging from red to pink-orange
due to high rate of carotenoid pigments in their cell
membrane. In addition, appeared opaque, transparent or
translucent, mucoid or non mucoid, with entire edges and
convex (Oren et al., 1997; Castillo et al., 2006). The
bacteria isolated from the various samples belonged to
Diversity of Halotolerant/Halophilic Bacteria / Int. J. Agric. Biol., Vol. 16, No. 3, 2014
567
diverse group of halotolerant and halophilic bacteria with
different phenotypic characteristics. However, the
phenotypic characteristics alone were not enough to
differentiate the bacterial isolates and could lead to false
identification. The main reason for this is that the
phenotypic characteristics depend on the growth conditions,
which were different in terms of NaCl concentration,
temperature, pH and medium composition. In this regard,
Fritze (2002) recommended that phenotypic characterization
results cannot be directly compared without full background
knowledge of the accurate conditions used for a particular
test.
Several studies have assessed the taxonomic status,
ecological characteristics, phylogenetic relationship and
biotechnological applications of halobacteria (Litchfield and
Gillevet, 2002). Halophilic bacteria were categorized on the
Table 2: Phenotypic characteristics of isolated halotolerant/halophilic strains
Strain ID Gram
stain
Cell
shape
Colony
color
Form Surface Margin Opacity Motility A/An pH for range
growth
(Optimum)
NaCl Range
(%) for
growth
Temperature Range
(°C) for growth
(Optimum)
NCCP-59 + R C Irregular Rough Lobate Opaque + A 5-9 (7) 2-20 4-45 (28)
NCCP-60 + C O Round Smooth Entire Opaque + A 6-9 (8) 1-15 4-40 (30)
NCCP-61 + R PY Round Smooth Entire Opaque + A 5-9 (7) 2-25 10-45 (35)
NCCP-62 + R PY Irregular Smooth Lobate Opaque + An 6-9 (8) 3-20 4-50 (30)
NCCP-63 + R PY Irregular Smooth Lobate Opaque + A 6-8 (7) 2-20 10-40 (35)
NCCP-65 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35)
NCCP-66 + R PY Circular Shiny Entire Opaque - An 6-9 (8) 1-30 4-40 (30) NCCP-71 + R C Irregular Smooth Filamentous Opaque + A 5-10 (7) 1-25 4-50 (37)
NCCP-73 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35)
NCCP-74 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35)
NCCP-75 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35)
NCCP-77 + R PY Round Smooth Entire Opaque + A 5-9 (7) 2-25 10-45 (35)
NCCP-79 + R C Round Smooth Entire Opaque + A 5-9 (8) 2-15 10-40 (28)
NCCP-80 + R PY Round Smooth Entire Opaque + A 5-9 (7) 2-25 10-45 (35)
NCCP-81 + R C Round Smooth Entire Opaque + A 5-9 (8) 0-15 10-40 (35) NCCP-82 + R C Irregular Smooth Filamentous Opaque + A 5-10 (7) 1-25 4-50 (37)
NCCP-83 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35)
NCCP-84 + C C Round Smooth Entire Opaque - A 5-9 (8) 2-30 10-40 (35)
NCCP-85 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35)
NCCP-86 + R PY Circular Smooth Entire Opaque + A 6-9 (7) 2-15 4-50 (28)
NCCP-87 - R C Circular Smooth Entire Opaque + A 6-8 (7) 0-15 4-45 (35)
NCCP-88 + R W Irregular Rough Undulate Opaque + A 5-9 (7) 1-20 10-50 (35) NCCP-91 + R PY Irregular Smooth Lobate Opaque + An 5-9 (7) 3-20 4-50 (30)
NCCP-92 + R PY Round Shiny Entire Opaque - A 6-9 (7) 2-20 10-45 (35)
NCCP-93 + R W Round Mucoid Entire Transparent - A 6-10 (7) 2-20 10-45 (30)
NCCP-166 + R C Irregular Smooth Filamentous Opaque + A 5-10 (7) 1-25 4-50 (37)
NCCP-168 + R PY Circular smooth Entire Opaque - A 5-9 (8) 0-15 10-40 (35)
NCCP-170 + R C Circular Smooth Filamentous Opaque + A 6-9 (7) 2-25 4-40 (37)
NCCP-171 - R W Irregular Smooth Lobate Opaque + A 5-9 (7) 1-19 10-45 (37)
NCCP-172 + R C Circular Rough Irregular Opaque + A 5-10 (7) 1-25 4-50 (37) NCCP-173 + R C Circular Rough Irregular Opaque + A 6-8 (7) 2-20 10-40 (35)
NCCP-174 + R C Round Smooth Entire Opaque + A 5-9 (8) 0-15 10-40 (35)
NCCP-175 + C O Round Smooth Entire Opaque - A 6-9 (7) 2-20 4-40 (28)
NCCP-176 + R PY Circular Smooth Entire Opaque - A 6-9 (8) 1-35 4-45 (35)
NCCP-184 + C C Round Smooth Entire Opaque - A 5-9 (8) 2-30 10-40 (35)
NCCP-204 + C C Round Smooth Entire Opaque - A 5-9 (8) 2-30 10-40 (35)
NCCP-701 - R W Circular Smooth Entire Transparent + An 6-10 (8) 2-30 15-40 (30) NCCP-704 + C C Round Smooth Entire Opaque - A 5-9 (8) 2-30 10-40 (35)
NCCP-705 + C Orange Circular Smooth Entire Opaque - A 6-10 (8) 1-30 4-50 (37)
NCCP-706 + R C Irregular Rough Lobate Opaque + A 5-9 (7) 2-18 4-45 (28)
NCCP-708 - R Cream Circular Smooth Entire Opaque + A 5-9(8) 0-30 4-45 (37)
NCCP-710 + C Orange Circular Smooth Entire Opaque - A 6-10 (8) 1-30 4-50 (37)
NCCP-713 - R Cream Circular Smooth Entire Opaque A 6-10 (8) 0-25 4-45 (30)
NCCP-716 + R Cream Circular Rough Irregular Opaque + A 5-10 (7) 1-25 4-50(37)
NCCP-717 + R PY Circular Shiny Entire Opaque - An 6-9 (8) 1-30 4-40 (30) NCCP-718 + C Orange Circular Smooth Entire Opaque - A 6-9 (7) 0-25 15-45 (35)
NCCP-721 + R PY Circular Smooth Entire Opaque + A 8-10 (9) 0-25 10-45(37)
NCCP-722 + C C Round Smooth Entire Opaque - A 5-9 (8) 2-30 10-40 (35)
NCCP-724 + R PY Circular Smooth Entire Opaque + A 6-9 (7) 2-35 4-45 (30)
NCCP-726 W Irregular Smooth Irregular Opaque A 6-10 (7) 0-20 4-35 (28)
NCCP-727 W Irregular Smooth Irregular Opaque A 6-10 (7) 0-20 4-35 (28)
NCCP-729 + R W Round Mucoid Entire Transparent - A 6-10 (7) 2-20 10-45 (30)
NCCP-730 + C C Round Smooth Entire Opaque - A 5-9 (8) 2-30 10-40 (35) NCCP-731 + R Cream Circular Rough Irregular Opaque + A 5-10 (7) 1-25 4-50 (37)
NCCP-732 + C Orange Circular Smooth Entire Opaque - A 6-10 (8) 1-30 4-50 (37)
NCCP-734 + C Orange Circular Smooth Entire Opaque - A 6-10 (8) 1-30 4-50 (37)
NCCP-735 - R W Circular Smooth Entire Opaque - A 6-9 (8) 3-35 4-45 (37)
Roohi et al., / Int. J. Agric. Biol., Vol. 16, No. 3, 2014
568
basis of tolerance to different NaCl concentrations into
slightly, moderately, and extremely-halophilic bacteria.
However, this approach is practically ineffective for this
purpose as the optimum and range of halophilic bacteria for
tolerating NaCl is critical. Previously, the halophilic bacteria
were grouped by using the results of salt tolerance test
proposed by Kushner (1993). Our results showed that the
slightly- and moderately halophilic bacteria were more
abundant than the extremely halophilic bacteria (Table 2).
Molecular phylogeny provides information regarding
organism relationships that is the basis for the conventional
identification techniques (Singh et al., 2007). 16S rRNA
gene sequence analysis is a widely used tool for
reconstructing microbial phylogeny (Dewhirst et al., 2005).
Table 3: Molecular identification of halophilic/halotolerant bacterial strains
Strain ID Strain Name /
Genus
16S rRNA
gene (nt)
Accession Closely Related validly published taxa Similarity of 16S
rRNA gene sequence (%)
No. of Species having
>97% similarity of 16S rRNA gene
NCCP-59 Bacillus sp. 1435 AB698777 Bacillus sonorensis (AF302118) 99.36 14 NCCP-60 Planococcus sp. 1441 AB698778 Planococcus citreus (X62172) 98.39 5
NCCP-61 Bacillus sp. 1452 AB715332 Bacillus subtilissubsp. inaquosorum, (EU138467) 99.74 15
NCCP-62 Bacillus sp. 1541 AB698779 Bacillus aquimaris (AF483625) 98.17 3
NCCP-63 Bacillus sp. 1499 AB698780 Bacillus licheniformis (AE017333) 99.60 10
NCCP-65 Bacillus sp. 1132 AB715333 Bacillus safensis(AF234854) 100.00 9
NCCP-66 Jeotgalicoccus sp. 1137 AB735682 Jeotgalicoccus halotolerans (AY028925) 99.82 6
NCCP-71 Bacillus sp. 1547 AB575949 Bacillus clausii (X76440) 99.18 2
NCCP-73 Bacillus sp. 1452 AB715334 Bacillus safensis (AF234854) 98.46 5 NCCP-74 Bacillus sp. 1096 AB715335 Bacillus safensis (AF234854) 99.90 5
NCCP-75 Bacillus sp. 1174 AB715336 Bacillus safensis (AF234854) 98.86 5
NCCP-77 Bacillus sp. 1410 AB698788 Bacillus subtilissubsp. Inaquosorum (EU138467) 99.91 12
NCCP-79 Bacillus sp. 1423 AB698789 Bacillus oceanisediminis (GQ292772) 98.85 3
NCCP-80 Bacillus sp. 1128 AB715337 Bacillus methylotrophicus (EU194897) 99.73 15
NCCP-81 Bacillus sp. 1455 AB715338 Bacillus licheniformis (AE017333) 98.76 10
NCCP-82 Bacillus sp. 1480 AB715339 Bacillus clausii (X76440) 99.24 2 NCCP-83 Bacillus sp. 1125 AB715340 Bacillus safensis (AF234854) 99.08 5
NCCP-84 Staphylococcus sp. 1101 AB715341 Staphylococcus equorum subsp. linens (AF527483) 99.54 15
NCCP-85 Bacillus sp. 1169 AB715342 Bacillus safensis (AF234854) 99.91 5
NCCP-86 Thalasobacillus sp. 1434 AB698790 Thalassobacillus devorans (AJ717299) 98.95 4
NCCP-87 Halomonas sp. 790 AB735683 Halomonas boliviensis (AY245449) 99.49 6
NCCP-88 Bacillus sp. 967 AB715343 Bacillus safensis (AF234854) 99.69 5
NCCP-91 Bacillus sp. 1421 AB698793 Bacillus vietnamensis (AB099708) 98.45 3
NCCP-92 Brevibacterium sp. 1406 AB698794 Brevibacteriumcasei (X76564) 99.10 8 NCCP-93 Bacillus sp. 1418 AB698795 Bacillus endophyticus (AF295302) 99.50 1
NCCP-166 Bacillus sp. 1432 AB698798 Bacillus clausii (X76440) 98.81 2
NCCP-168 Bacillus sp. 1410 AB618147 Bacillus seohaeanensis (AY667495) 97.06 1
NCCP-170 Gracilibacillus sp. 1453 AB698801 Gracilibacillus dipsosauri (X82436) 98.81 2
NCCP-171 Halomonas sp. 1427 AB698802 Halomonas elongate (FN869568) 99.43 6
NCCP-172 Bacillus sp. 1423 AB698803 Bacillus safensis (AF234854) 99.86 5
NCCP-173 Bacillus sp. 1422 AB698804 Bacillus safensis (AF234854) 99.93 5
NCCP-174 Bacillus sp. 1433 AB698805 Bacillus licheniformis (AE017333) 98.32 14 NCCP-175 Kocuria sp. 1404 AB698806 Kocuria sediminis (JF896464) 99.78 9
NCCP-176 Halobacillus sp. 1480 AB698807 Halobacillus profundi (AB189298) 95.00 0
NCCP-184 Staphylococcus sp. 1050 AB735684 Staphylococcus equorum subsp. equorum(AB009939) 99.90 23
NCCP-204 Staphylococcus sp. 1420 AB698815 Staphylococcus equorum subsp. equorum(AB009939) 98.44 10
NCCP-701 Salinivibrio sp. 1459 AB715344 Salinivibrio sharmensis (AM279734) 97.82 1
NCCP-704 Staphylococcus sp. 1147 AB715345 Staphylococcus equorum subsp. equorum(AB009939) 100.0 23
NCCP-705 Salinicoccus sp. 1175 AB715346 Salinicoccus roseus (X94559) 98.97 4 NCCP-706 Bacillus sp. 1042 AB715347 Bacillus sonorensis (AF302118) 98.46 13
NCCP-708 Salinivibrio sp. 1100 AB735685 Salinivibrio costicola subsp. Alkaliphilus (AJ640132) 100.00 6
NCCP-710 Salinicoccus sp. 1311 Salinicoccus roseus (X94559) 98.62 3
NCCP-713 Halomonas sp. 868 AB735686 Halomonasalk aliantarctica (AJ564880) 99.42 9
NCCP-716 Bacillus sp. 1172 AB735687 Bacillus safensis (AF234854) 100.0 6
NCCP-717 Jeotgalicoccus sp. 1201 AB735688 Jeotgalicoccus halotolerans (AY028925) 99.42 6
NCCP-718 Kocuria sp. 1055 AB735689 Kocuria rosea (X87756) 99.53 7
NCCP-721 Oceanobacillus sp. 985 AB735690 Oceanobacillus onchorhynchi subsp. Incaldanensid (AJ640134) 99.90 4 NCCP-722 Staphylococcus sp. 1097 AB735691 Staphylococcus equorum subsp. equorum(AB009939) 100.0 21
NCCP-724 Oceanobacillus sp. 1172 AB735692 Oceanobacillus picturae (AJ315060) 100.0 4
NCCP-726 Bacillus sp. 1081 AB735693 Bacillus aerophilus (AJ831844) 99.54 5
NCCP-727 Bacillus sp. 1117 AB735694 Bacillus aerophilus (AJ831844) 100.0 5
NCCP-729 Bacillus sp. 966 AB735695 Bacillus endophyticus (AF295302) 99.79 1
NCCP-730 Staphylococcus sp. 1095 AB735696 Staphylococcus equorum subsp. Equorum (AB009939) 99.91 18
NCCP-731 Bacillus sp. 1134 AB735697 Bacillus safensis (AF234854) 99.65 5
NCCP-732 Bacillus sp. 1096 AB735698 Bacillus sonorensis (AF302118) 99.18 16 NCCP-734 Bacillus sp. 1176 AB735699 Bacillus sonorensis(AF302118) 99.23 16
NCCP-735 Halomonas sp. 1149 AB735700 Halomonas alkaliphila (AJ640133) 99.91 14

Diversity of Halotolerant/Halophilic Bacteria / Int. J. Agric. Biol., Vol. 16, No. 3, 2014
569
In a previous study, Rohban et al. (2009) also isolated
bacterial strains belonging to the genera Salicola,
Halovibrio, Halomonas, Bacillus, Oceanobacillus,
Thalassobacillus, Virgibacillus, Gracilibacillus,
Halobacillus, Piscibacillus and Salinicoccus from Saltan
lake of Iran.
Staphylococci has ability to grow in a wide range of
salt concentrations (Graham and Wilkinson, 1992; Garzoni
and Kelley, 2009; Morikawa et al., 2010) and is in
accordance with our study. Staphylococcus strains were
isolated from various salt samples during these studies and
the results agreed with previous reports. Highly diverse
nature of Bacillus genus found in this study is in accordance
with the observations made by Claus and Berkeley (1986).
Due to the ubiquity and capability to survive under adverse
conditions, heterotrophic Bacillus strains cannot be
considered species of certain specific habitats (Claus and
Berkeley, 1986). It is therefore not surprising that a large
number of the isolates recovered in the present study
belonged to the same bacterial group.
In conclusion, the strains from salt mines of Karak can
be further characterized using polyphasic taxonomic
approach and other molecular methods. This is the first
study on investigating the bacterial diversity of the Karak
salt mines of Pakistan and it provided a large number of
strains which are candidate novel species. The rich diversity
of genus Bacillus found in this study points out to the
extensive distribution and ecological relationship with other
microorganisms, e.g. halophilic bacteria demands further
comprehensive study. The microbial diversity can prove to
be a valuable future resource in various industrial and
biotechnological processes. Such microbes can also be used
as a source of gene(s) that can increase salt tolerance in
different crop species through genetic transformation.
Acknowledgements
This work was supported partly by financial assistance from
PSDP funded Project Research for Agricultural
Development Project under a sub-project (Grant No. CS-
55/RADP/PARC) entitled “Establishment of Microbial Bio-
Resource Laboratories: National Culture Collection of
Pakistan (NCCP)” from Pakistan Agricultural Research
Council, Islamabad, Pakistan
References
Ahmed, I., A. Yokota and T. Fujiwara, 2007. A novel highly boron tolerant
bacterium, bacillus boroniphilus sp. Nov., isolated from soil, that
requires boron for its growth. Extremophiles, 11: 217‒224
Antón, J., R. Rosselló‒Mora, F. Rodríguez‒Valera and R. Amann, 2000.
Extremely halophilic bacteria in crystallizer ponds from solar
salterns. Appl. Environ. Microbiol., 66: 3052‒3057
Birbir, M., B. Calli, B. Mertoglu, R. Bardavid, A. Oren, M. Ogmen and A.
Ogan, 2007. Extremely halophilic archaea from tuz lake, turkey, and
the adjacent kaldirim and kayacik salterns. World J. Microbiol.
Biotechnol. 23: 309‒316
Castillo, A.M., M.C. Gutiérrez, M. Kamekura, Y. Ma, D.A. Cowan, B.E.
Jones, W.D. Grant and A. Ventosa, 2006. Halovivax asiaticus gen.
Nov., sp. Nov., a novel extremely halophilic archaeon isolated
from inner mongolia, china. Int. J. Syst. Evol. Microbiol., 56: 765‒770
Castillo, A.M., M.C. Gutiérrez, M. Kamekura, Y. Xue, Y. Ma, D.A. Cowan,
B.E. Jones, W.D. Grant and A. Ventosa, 2007. Halorubrum
ejinorense sp. Nov., isolated from lake ejinor, inner mongolia, china.
Int. J. Syst. Evol. Microbiol., 57: 2538‒2542
Fig. 2: Phylogenetic tree showing the interrelationships of
isolated halotolerant/halophilic strains along with their
closely related taxa belonging to Firmicutes, Actinobacteria
and Proteobacteria inferred from 16SrRNA gene sequence.
Data with gaps were removed after alignment by
CLUXTAL X. The rooted tree was constructed using the
neighbour-joining method contained in the MEGA5
software (Tamura et al., 2011). Bootstrap values, expressed
as percentages of 1000 replications, are given at branching
points. Closely related type species were presented in bold
letters and the 3% bar shows sequence divergence

Roohi et al., / Int. J. Agric. Biol., Vol. 16, No. 3, 2014
570
Claus, D. and R.C.W. Berkeley, 1986. Genus Bacillus 1872. In: Bergey's
Manual of Systematic Bacteriology. P.H.A. Sneath, PHA, N.S. Mair,
M.E. Sharpe and J.G. Holt (ed.). Vol. 2. pp: 1105‒1139. Williams
and Wilkins, Baltimore, Maryland, USA
Dewhirst, F.E., Z. Shen, M.S. Scimeca, L.N. Stokes, T. Boumenna, T.
Chen, B.J. Paster and J.G. Fox, 2005. Discordant 16s and 23s rrna
gene phylogenies for the genus helicobacter: implications for
phylogenetic inference and systematics. J. Bacteriol., 187: 6106‒6118
Euzéby, J.P., 2013. List of prokaryotic names with standing in
nomenclature. Accessed on 9 June 2013 http:
//www.bacterio.net/h/halobacteriaceae.html
Fritze, D., 2002. Bacillus identification‒ traditional approaches. In:
Applications and Systematics of Bacillus and Relatives. Berkeley, R.,
M. Heyndrickx, N. Logan, P. De Vos (Eds.), pp. 100‒122.
Blackwell, Oxford, UK
Garzoni, C. and W.L. Kelley, 2009. Staphylococcus aureus: New evidence
for intracellular persistence. Trends Microbiol., 17: 59‒65
Graham, J.E. and B.J. Wilkinson, 1992. Staphylococcus aureus
osmoregulation: roles for choline, glycine betaine, proline, and
taurine. J. Bacteriol., 174: 2711‒2716
Hall, T.A., 1999. Bioedit: a user‒friendly biological sequence alignment
editor and analysis program for windows 95/98/nt. Nucleic Acids
Symposium Series
Johnson, A., L. Thurlow, R. Zwenger and E. Gillock, 2007. Partial
characterization of two moderately halophilic bacteria from a kansas
salt marsh. Prairie Nat, 39: 29‒39
Kamekura, M., 1998. Diversity of extremely halophilic bacteria.
Extremophiles, 2: 289‒295
Kushner, D.J., 1985. The halobacteriaceae. Academic Press, New York,
USA
Kushner, D.J., 1993. Growth and nutrition of halophilic bacteria. CRC
Press, Boca Raton, Florida, USA
Litchfield, C.D. and P.M. Gillevet, 2002. Microbial diversity and
complexity in hypersaline environments: a preliminary assessment.
J. Ind. Microbiol. Biotechnol., 28: 48‒55
Mancinelli, R.L. and L.I. Hochstein, 1986. The occurrence of denitrification
in extremely halophilic bacteria. FEMS Microbiol. Lett., 35: 55‒58
Morikawa, K., R.L. Ohniwa, T. Ohta, Y. Tanaka, K. Takeyasu and T.
Msadek, 2010. Adaptation beyond the stress response: Cell structure
dynamics and population heterogeneity in staphylococcus aureus.
Microb. Environ., 25: 75‒82
Oren, A., 1993. Ecology of extremely halophilic microorganisms. CRC
Press, Boca Raton, Florida, USA
Oren, A., 2002. Diversity of halophilic microorganisms: environments,
phylogeny, physiology, and applications. J. Ind. Microbiol.
Biotechnol., 28: 56‒63
Oren, A. and F.s. Rodrı́guez‒Valera, 2001. The contribution of halophilic
bacteria to the red coloration of saltern crystallizer ponds1. FEMS
Microbiol. Ecol., 36: 123‒130
Oren, A., A. Ventosa and W.D. Grant, 1997. Proposed minimal standards
for description of new taxa in the order halobacteriales. Int. J. Syst.
Bacteriol., 47: 233‒238
Ozcan, B., G. Ozcengiz, A. Coleri and C. Cokmus, 2007. Diversity of
halophilic archaea from six distinct parts of turkey. J. Microbiol.
Biotechnol., 17: 985–992
PMDC, 2013. Pakistan mineral development corporation. Ministry of
Petroleum and Natural Resources, Government of Pakistan,
Islamabad
Quesada, E., V. Béjar, M. Ferrer, C. Calvo, I. Llamas, F. Martínez‒Checa,
S. Arias, C. Ruiz‒García, R. Páez, M. Martinez‒Cánovas and A. del
Moral, 2004. Moderately halophilic, exopolysaccharide‒producing
bacteria. Berlin: Springer‒Verlag, Berlin, Germany
Quesada, E., A. Ventosa, F. Rodriguez‒Valera and A. Ramos‒Cormenzana,
1982. Types and properties of some bacteria isolated from
hypersaline soils. J. Appl. Microbiol., 53: 155‒161
Rohban, R., M. Amoozegar and A. Ventosa, 2009. Screening and isolation
of halophilic bacteria producing extracellular hydrolyses from howz
soltan lake, Iran. J. Ind. Microbiol. Biotechnol., 36: 333‒340
Roohi, A., I. Ahmed, M. Iqbal and M. Jamil, 2012. Preliminary isolation
and characterization of halotolerant and halophilic bacteria from salt
mines of Karak, Pakistan. Pak. J. Bot., 44: 365‒370
Singh, S., R. Chandra, D.K. Patel and V. Rai, 2007. Isolation and
characterization of novel serratia marcescens (ay927692) for
pentachlorophenol degradation from pulp and paper mill waste.
World J. Microbiol. Biotechnol.,23: 1747‒1754
Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei and S. Kumar,
2011. Mega5: Molecular evolutionary genetics analysis using
maximum likelihood, evolutionary distance, and maximum
parsimony methods. Mol. Biol. Evol., 28: 2731‒2739
Ventosa, A., E. Mellado, C. Sánchez‒Porro and M. Marquez , 2008.
Halophilic and halotolerant micro‒organisms from soils. Berlin
Springer‒Verlag, Berlin, Germany
Woese, C.R. and G.E. Fox, 1977. Phylogenetic structure of the prokaryotic
domain: The primary kingdoms. Proc. Natl. Acad. Sci. USA, 74:
5088‒5090
(Received 30 September 2013; Accepted 24 February 2014)




















