
Isoforms of the human PDZ-73 protein exhibit differential tissue expression
14
Isoforms of the human PDZ-73 protein exhibit di¡erential tissue expression Matthew J. Scanlan a ; *, Barbara Williamson a , Achim Jungbluth a , Elisabeth Stockert a , Karen C. Arden b , Carrie S. Viars b , Ali O. Gure a , John D. Gordan a , Yao-Tseng Chen a;c , Lloyd J. Old a a Ludwig Institute for Cancer Research, New York Branch at Memorial Sloan-Kettering Cancer Center, 1275 York Avenue, New York, NY 10021, USA b Ludwig Institute for Cancer Research, San Diego Branch, La Jolla, CA 92093, USA c Cornell University Medical College, 1300 York Avenue, New York, NY 10021, USA Received 5 February 1999; accepted 8 February 1999 Abstract Patients with renal and colon cancer frequently develop IgG autoantibodies toward the NY-CO-38/PDZ-73 antigen, a protein of 652 amino acids (73 kDa) which contains three copies of the PDZ protein^protein interaction domain. The gene encoding PDZ-73 mapped to chromosome 11p15.4^p15.1. Additional tissue-specific isoforms were identified : PDZ-45, which lacks the third PDZ domain and the putative PEST protein degradation motif, is expressed in kidney, colon, small intestine, brain and testis; PDZ-54 and PDZ-59, which also lack the third PDZ domains, have unique carboxyl terminal amino acids and are expressed in brain, kidney, bladder, colon cancer and renal cancer; and a putative PDZ-37 isoform, containing only the third PDZ domain, that is expressed in the central nervous system. Immunohistochemical staining with anti-PDZ 73 monoclonal antibodies showed strong cytoplasmic reactivity in epithelial cells of the small intestine, colon and kidney tubules, with a prominent apical staining pattern in cells of the small intestine. The reactivity pattern of the antibodies with various tissues correlated with the mRNA expression pattern of the PDZ-45 isoform. The existence of multiple PDZ-73 isoforms with variations in tissue distribution, PDZ domains, protein degradation sequences and carboxyl terminal structure indicate that these isoforms have distinct tissue-specific functions. ß 1999 Elsevier Science B.V. All rights reserved. Keywords : PDZ domain; NY-CO-38 antigen; Isoform expression; Serological expression cloning; 11p15.4^p15.1; Colon cancer 0167-4781 / 99 / $ ^ see front matter ß 1999 Elsevier Science B.V. All rights reserved. PII:S0167-4781(99)00033-0 Abbreviations : SEREX, serological expression cloning ; PDZ, domains in PSD-95, h Dlg and ZO-1 ; PSD-95, Post-synaptic density protein 95; hDlg, human homologue to Drosophila discs-large tumor suppressor protein; ZO-1, zonula occludens protein 1; FISH, £uorescence in situ hybridization ; BAC, bacterial arti¢cial chromosome vector ; RT^PCR, reverse transcriptase^polymerase chain reac- tion; RACE, rapid ampli¢cation of cDNA ends; mAbs, monoclonal antibodies; ABC method, avidin^biotin^complex method; APAAP, alkaline phosphatase/anti-alkaline phosphatase detection system ; NMDA, N-methyl-D^aspartate ; CAR, Coxsackie and adenovirus re- ceptor ; ELISA, enzyme-linked immunosorbent assay * Corresponding author. Fax : +1-212-717-3100 ; E-mail : [email protected] Biochimica et Biophysica Acta 1445 (1999) 39^52
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Isoforms of the human PDZ-73 protein exhibit differential tissue expression

Isoforms of the human PDZ-73 protein exhibit di¡erential tissueexpression
Matthew J. Scanlan a;*, Barbara Williamson a, Achim Jungbluth a, Elisabeth Stockert a,Karen C. Arden b, Carrie S. Viars b, Ali O. Gure a, John D. Gordan a,
Yao-Tseng Chen a;c, Lloyd J. Old a
a Ludwig Institute for Cancer Research, New York Branch at Memorial Sloan-Kettering Cancer Center, 1275 York Avenue,New York, NY 10021, USA
b Ludwig Institute for Cancer Research, San Diego Branch, La Jolla, CA 92093, USAc Cornell University Medical College, 1300 York Avenue, New York, NY 10021, USA
Received 5 February 1999; accepted 8 February 1999
Abstract
Patients with renal and colon cancer frequently develop IgG autoantibodies toward the NY-CO-38/PDZ-73 antigen, aprotein of 652 amino acids (73 kDa) which contains three copies of the PDZ protein^protein interaction domain. The geneencoding PDZ-73 mapped to chromosome 11p15.4^p15.1. Additional tissue-specific isoforms were identified: PDZ-45,which lacks the third PDZ domain and the putative PEST protein degradation motif, is expressed in kidney, colon, smallintestine, brain and testis; PDZ-54 and PDZ-59, which also lack the third PDZ domains, have unique carboxyl terminalamino acids and are expressed in brain, kidney, bladder, colon cancer and renal cancer; and a putative PDZ-37 isoform,containing only the third PDZ domain, that is expressed in the central nervous system. Immunohistochemical staining withanti-PDZ 73 monoclonal antibodies showed strong cytoplasmic reactivity in epithelial cells of the small intestine, colon andkidney tubules, with a prominent apical staining pattern in cells of the small intestine. The reactivity pattern of the antibodieswith various tissues correlated with the mRNA expression pattern of the PDZ-45 isoform. The existence of multiple PDZ-73isoforms with variations in tissue distribution, PDZ domains, protein degradation sequences and carboxyl terminal structureindicate that these isoforms have distinct tissue-specific functions. ß 1999 Elsevier Science B.V. All rights reserved.
Keywords: PDZ domain; NY-CO-38 antigen; Isoform expression; Serological expression cloning; 11p15.4^p15.1; Colon cancer
0167-4781 / 99 / $ ^ see front matter ß 1999 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 7 8 1 ( 9 9 ) 0 0 0 3 3 - 0
Abbreviations: SEREX, serological expression cloning; PDZ, domains in PSD-95, hDlg and ZO-1; PSD-95, Post-synaptic densityprotein 95; hDlg, human homologue to Drosophila discs-large tumor suppressor protein; ZO-1, zonula occludens protein 1; FISH,£uorescence in situ hybridization; BAC, bacterial arti¢cial chromosome vector; RT^PCR, reverse transcriptase^polymerase chain reac-tion; RACE, rapid ampli¢cation of cDNA ends; mAbs, monoclonal antibodies; ABC method, avidin^biotin^complex method; APAAP,alkaline phosphatase/anti-alkaline phosphatase detection system; NMDA, N-methyl-D^aspartate; CAR, Coxsackie and adenovirus re-ceptor; ELISA, enzyme-linked immunosorbent assay
* Corresponding author. Fax: +1-212-717-3100; E-mail : [email protected]
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
Biochimica et Biophysica Acta 1445 (1999) 39^52

1. Introduction
Serological expression cloning (SEREX) is a novelapproach to the identi¢cation of human tumor anti-gens recognized by the autologous host and involvesthe immunoscreening of cDNA expression librariesderived from human tumors with autologous patientsera [1]. In an initial SEREX analysis of colon can-cer, forty-eight antigens were de¢ned [2], one ofwhich, NY-CO-38 is a novel gene product encodingseveral copies of the PDZ protein^protein interactiondomain [3]. Translation of the NY-CO-38 cDNAgives a putative protein of 652 amino acids (73 337Da) with three PDZ domains as well as a possiblePEST protein degradation motif, a coiled-coil do-main, and amino terminal and carboxyl terminal do-mains with no known similarities. This protein hasbeen designated PDZ-73.
PDZ domains are protein modules of 70^90 aminoacids, originally de¢ned by the presence of a GLGFrepeat consensus, and are found in a diverse group ofproteins, including protein tyrosine phosphatase L1[4], erythrocyte p55 protein [5] and members of themembrane associated guanylate kinase family of pro-teins, e.g., postsynaptic density protein 95 [6], Discs-large [7] and Zona occludens protein 1 [8]. Such do-mains interact with the cytoplasmic tails of targetproteins, generally by binding to carboxyl terminalpeptides with the (S/T)XV consensus sequence [5,9].Proteins with the PDZ motif often function in intra-cellular signal transduction and cytoskeletal organi-zation [3].
In the present study we further characterize thePDZ-73 protein in terms of its tissue expressionand localization, the occurrence of related isoforms,and the chromosomal location of the PDZ-73 codinggene.
2. Materials and methods
2.1. Serological expression cloning (SEREX) andDNA sequence analysis of PDZ-73 and itsisoforms
SEREX analysis of a metastatic colon cancercDNA expression library was described previously[1,2]. Brie£y, RNA was extracted from a surgical
specimen of a colon cancer metastasis to the lungof a 54-year-old female by the guanidinium thiocya-nate method and puri¢ed to poly(A)� RNA usingthe FAST Track kit (Invitrogen, San Diego, CA).Synthesis of cDNA and ligation into the lambdaZAP express vector was undertaken as per the man-ufacturer's protocol (Stratagene, La Jolla, CA). Us-ing an ampli¢ed portion of the metastatic colon can-cer cDNA library, 5U105 recombinants wereimmunoscreened as follows: recombinant phage ata concentration of 4U103 per 15-cm plate were am-pli¢ed for 6 h; transferred to nitrocellulose mem-branes for 15 h at 37³C; membranes were blockedwith 5% non-fat dried milk for 1 h; and then incu-bated in pre-absorbed autologous patient sera (1:100dilution) for 15 h at room temperature. Sera werepre-absorbed by repeated passage through columnsof Sepharose 4B coupled to lysates from Escherichiacoli Y1090 and bacteriophage-infected E. coliBNN97 (5 Prime 3 Prime, Boulder, CO) in orderto remove antibodies reactive with antigens relatedto the vector system. After serum exposure, ¢lterswere incubated in a 1:3000 dilution of alkaline phos-phatase conjugated, Fc fragment-speci¢c, goat anti-human IgG (Jackson Immunoresearch Laboratories,West Grove, PA) for 1 h at room temperature andprocessed for 4-nitro blue tetrazolium chloride/5-bro-mo-4-chloro-3-indolyl-phosphate color development.Serum-positive clones were subcloned and retestedfor serum reactivity as above.
Serum-positive cDNA clones (NY-CO-38 and NY-CO-37) were converted to plasmid forms (pBKCMV)by in vivo excision as per the manufacturer's proto-col (Stratagene). Plasmid DNA was puri¢ed onQiaprep spin columns (Qiagen, Chatsworth, CA).Automated DNA sequencing was performed by theDNA services facility at Cornell University (Ithaca,NY) using an ABI Prism (Perkin Elmer, Foster City,CA) automated DNA sequencer. In order to isolatePDZ-73 gene products from normal tissues, 5P and 3PRACE reactions were undertaken using KlenTaq-1DNA polymerase and Marathon-Ready normal co-lon and brain cDNA (Clontech, Palo Alto, CA). ThePDZ-73 gene-speci¢c primers utilized in the RACEreactions were located at nucleotide positions 603^627 (3P RACE primer 1, GATCCCCGTGAAA-AGCTCTCCTGAT), 943^962 (3P RACE primer 2,AGCCTGACCATCTCCATTGT), 1533^1553 (5P
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5240

RACE primer 1, TCATACACAGCAGAAACG-ACC), and 1461^1483 (5P RACE primer 2, ATC-CCTCCTTCTTGATGCGTAGG) and used in con-junction with Marathon-Ready adapter primers(Clontech). First round RACE reactions (primers 1)and second round reactions (primers 2) consisted of30 cycles and 23 cycles, respectively, at a denatura-tion temperature of 95³C (1 min/cycle) followed byan annealing/extension temperature of 68³C (3 min/cycle). Products were T/A ligated into pCR2.1 plas-mid vector (Invitrogen). Multiple RACE clones weresubjected to DNA sequencing.
2.2. Fluorescence in situ hybridization (FISH)
A PDZ-73 genomic clone was isolated by screeninga human female lymphocyte genomic library consist-ing of 2.2U105 clones in the pBeloBAC II bacterialarti¢cial chromosome vector (BAC, Genome Sys-tems, St. Louis, MO) with a 32P-labeled PDZ-73cDNA probe (bp 1^1553). The identity of resultantPDZ-73 BAC clones were veri¢ed by PCR analysisand DNA sequencing. For FISH analysis, the entirePDZ-73 BAC clone (100 kb) was nick translated us-ing Biotin-14 dATP and Biotin-14 dCTP (GibcoBRL, Grand Island, NY). Chromosome preparationswere prepared from phytohemagglutinin-stimulatednormal peripheral blood lymphocytes cultured for72 h. To induce R-banding, some of the cultureswere synchronized with thymidine after 48 h, incu-bated for 18 h at 37³C, followed by 5-bromodeoxy-uridine treatment for 6 h. The slides were stored at380³C before use. FISH of metaphase chromosomeswas undertaken as follows. The biotin-labeled probe(50^100 ng) was dissolved in hybridization mixture(50% formamide, 10% dextran sulfate, 2USSC,COT-1 DNA, 10 Wg sheared salmon sperm DNA)and incubated for 60 min at 37³C. The probe mixturewas then applied to the slide and co-denatured for 10min at 80³C on a slide warmer. Hybridization wasallowed to proceed for a minimum of 24 h in a hu-midi¢ed chamber at 37³C. The slides were thenwashed using the formamide-wash procedure as perthe FITC^biotin detection kit (Oncor, Gaithersburg,MD). Biotin-labeled probe detection was accom-plished by incubation with the FITC^avidin conju-gate. Chromosome identi¢cation was performed bysimultaneous hybridization with a chromosome 11-
speci¢c K-satellite repeat probe (Oncor) or by R-banding. Slides were examined using a Zeiss Axio-phot microscope with appropriate UV-¢lter combi-nations.
2.3. mRNA expression analysis
The mRNA expression pattern of PDZ-73 and re-lated isoforms was determined by reverse transcrip-tase PCR (RT^PCR) using a panel of normal andmalignant tissue RNA. Tumor RNA was extractedfrom surgical specimens by the guanidinium thiocy-anate method. Normal tissue RNA was purchasedcommercially (Clontech). The cDNA preparationsused in the RT^PCR reactions were synthesized byincubating total RNA template (2 Wg), random hex-amers (1.66 Wg, Boehringer Mannheim, Indianapolis,IN) and MuLV reverse transcriptase (200 U, GibcoBRL), in a total reaction volume of 25 Wl, at 42³Cfor a period of 1 h. MuLV synthesized cDNA (2.5 Wl/PCR reaction) was then ampli¢ed using gene-speci¢cprimers (0.2 WM) and AmpliTaq Gold DNA polym-erase (2.5 U, PE Applied Biosystems, Branchburg,NJ) in 25-Wl PCR reactions consisting of 30 cyclesat a denaturation temperature of 94³C (1 min/cycle);an annealing temperature of 60³C (1 min/cycle); andan extension temperature of 72³C (2 min/cycle). As acontrol for genomic DNA contamination in RT^PCR reactions, duplicate cDNA templates were pre-pared as above without MuLV reverse transcriptaseand used in equivalent PCR reactions. In addition toRT^PCR product identi¢cation on the basis of size,selected products were subcloned into pCR2.1 andsubjected to DNA sequencing as described above.Gene-speci¢c PCR primers (Gibco BRL) consistedof the following sequences: PDZ-73 forward, AAG-GAACGGGTCGTGCAACG; PDZ-73 reverse,TGGGAGATGAAGAGCCCACAGC; PDZ-45 for-ward, CCTGACCATCTCCATTGTAGC; PDZ-45reverse, CGGAAATCACTTTGGCTTGC; PDZ-54forward, PDZ-59 forward, and nested forward #1,GGCTCAAAGGAACAGCTACTCTTGC; PDZ-54reverse, GATAAACCACCATCCTCTTC; PDZ-59reverse, ATTTCACAGCTGGCCTGTAG; nestedforward #2, AGCCTGACCATCTCCATTGTA;and nested reverse, GATCCCTCCTTCTTGATGC-GTAGG.
Poly(A)� RNA was prepared using the FAST
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^52 41

Track kit (Invitrogen) and isolated from the follow-ing cell lines obtained from the cell bank of the Lud-wig Institute for Cancer Research, New YorkBranch: Colo 205 (colon tumor), HT 29 (colon tu-mor), HCT 15 (colon tumor), MCF-7 (breast tumor),LA15S (neuroblastoma), MFSH (histiocytoma), SK-OV-6 (ovarian tumor) and HS27 (newborn ¢bro-blast) cell lines. Northern blots containing poly(A)�
mRNA (2 Wg/lane) from the various cell lines wereprepared using published procedures [10]. Northernblots containing normal tissue poly(A)� RNA (2 Wg/lane) were obtained commercially (Clontech). Ran-dom-primed 32P-labeled probes prepared from PCRproducts of PDZ-73 cDNA (3P probe, bp 1480^2019;5P probe, bp 54^443) were hybridized to Northernblots for 1.5 h in Expresshyb (Clontech) at 68³C,washed at high stringency (three times, 20 mineach, 0.1USSC/0.1% SDS at 68³C), and used to ex-pose Biomax MS autoradiography ¢lms (EastmanKodak, Rochester, NY).
2.4. PDZ-73 protein expression: the generation ofmonoclonal antibodies, immunohistochemistryand Western blotting
The nucleotide sequence encoding amino acids 1^493 of the PDZ-73 antigen was expressed in E. coliusing the pQE-30 expression system (Qiagen). Theinduction of recombinant protein synthesis and sub-sequent puri¢cation by Ni2� a¤nity chromatographywere performed as per the manufacturer's protocol.Recombinant PDZ-73 protein (50 Wg/dose) in adju-
vant was used to immunize BALB/c mice, ¢ve timessubcutaneously, at 2- to 3-week intervals. CompleteFreund's adjuvant was used for the ¢rst injection.Thereafter, incomplete Freund's adjuvant (DifcoLaboratories, Detroit, MI) was used. Spleen cellsfrom immunized mice were fused with the mousemyeloma cell line SP2/O and supernatants from se-quentially cloned cell populations were screenedagainst the immunizing protein and appropriate con-trol fusion proteins by ELISA [11]. Speci¢city of theantibodies was further evaluated histologically andby Western blotting.
For immunohistochemical studies, frozen tissueswere used. The Tissue Procurement Service of theDepartment of Pathology at Memorial Sloan-Ketter-ing Cancer Center provided the tissues. The tissueswere snap-frozen in isopentane, pre-cooled in dry ice,and stored at 375³C. Cuts of 5 Wm were generatedand ¢xed in 3% formaldehyde solution for 3 min.Endogenous peroxidase activity was suppressed byincubation in 0.3% H2O2+0.2% NaN3 for 20 min.The slides were then blocked with 4% BSA/PBSand the primary antibodies were diluted to 1:25 inimmunohistochemistry diluent (Fisher Scienti¢c,Pittsburgh, PA) and applied overnight at 4³C. Thedetection of the primary antibodies was performedwith a biotinylated horse anti-mouse-secondary anti-body (1:200, Vector Labs, Burlingame, CA) and anavidin^biotin^complex system (ABC method) usingdiaminobenzidine tetrahydrochloride (BioGenex,San Ramon, CA) as a chromogen. For liver andkidney tissues an alkaline phosphatase/anti-alkaline
C
Fig. 1. The nucleotide sequence, deduced amino acid sequence and domain structure of PDZ-73 and its isoforms. (A) The nucleotidesequence was derived from bi-directional sequencing of PDZ-73 and its isoforms isolated from normal colon, brain and kidney as wellas colon and renal cancer. Various structural characteristics are noted: the PDZ domains are underlined; the putative PEST domain/PDZ-45 deletion is shown in lower case letters; inequality symbols (G f) encompass the coiled coil domain; brackets denote the possi-ble PDZ-interacting module. The 1.3 kb PDZ-73 transcript encoding the putative PDZ-37 isoform is highlighted and the underlined/bold nucleotide (bp 1380) is the site of the PDZ-54 and PDZ-59 insertion. PDZ-73/NY-CO-38 and PDZ-45/NY-CO-37 GenBank ac-cession numbers are AF039700 and AF039699, respectively. (B) The domain structure of PDZ-73 and its isoforms, PDZ-45, PDZ-54and PDZ-59. The putative protein encoded by the 1.3 kb PDZ-73 transcript is also shown as PDZ-37. PDZ-73 isoforms are character-ized by di¡erences in the expressed PDZ domains, the presence of a putative PEST protein degradation sequence and the compositionof the carboxyl terminal residues. Empty circles represent carboxyl terminal residues unique to PDZ-54 and PDZ-59, and the shadedcircle represents carboxyl terminal residues unique to PDZ-59. N, amino terminal domain; 1, PDZ domain 1; 2, PDZ domain 2; 3,PDZ domain 3; C, coiled-coil domain; P, PEST degradation domain; CO, carboxyl terminal domain. (C) The nucleotide sequence en-coding the carboxyl terminal residues of the PDZ-54 and PDZ-59 isoforms. The 173 nt insertion of the PDZ-59 isoform and its trans-lation is highlighted. The 53 nt insertion of the PDZ-54 isoform is underlined. In both cases, the nucleotides are inserted betweenbase pair position 1380^1381 of PDZ-73, altering its reading frame (bold: nucleotides; italic: amino acids) and creating a truncatedprotein with a unique carboxyl terminus.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5242

BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^52 43

phosphatase (APAAP) detection system with a rab-bit anti-mouse-secondary antibody (DAKO, Carpin-tera, CA) and new fuchsin was used. Slides werecounterstained with Mayer's hematoxylin, dehy-drated in a series of graded alcohols and coveredwith Permount. On the basis of preliminary reactivitywith normal colon, kidney and colonic adenocarci-noma, monoclonal antibodies (mAbs) Co25 andCo137 (both IgG2A) were chosen for further studyon a wider tissue panel.
For use in Western blots, cell extracts were pre-pared from the colon cancer cell lines, Colo 205, HT29 and HCT 15. Brie£y, 1U107 cells were lysed in 1.5ml lysis bu¡er (10 mM Tris^HCl (pH 7.3)/150 mMNaCl/10 mM MgCl2/0.5% Nonidet P-40) containingComplete protease inhibitor cocktail (BoehringerMannheim) for 30 min at room temperature. Thelysate was then centrifuged at 100 000Ug for 30min and the resultant supernatant was collected. Ly-sates from each cell line (25 Wg total protein each)were electrophoresed under denaturing conditions ina 10% polyacrylamide/Tris/glycine gel (Novex, SanDiego, CA). Proteins were then transferred by elec-trophoretic elution to PVDF membrane and ana-lyzed using the Western-light plus chemiluminescentdetection system (Tropix, Bedford, MA) in conjunc-tion with primary anti-PDZ-73 monoclonal antibod-ies, according to the manufacturer's protocol.
3. Results
3.1. Characterization of PDZ-73 and PDZ-45transcripts
The immunoscreening of a cDNA expression li-brary derived from a metastatic colon cancer withautologous serum led to the isolation of a seroreac-tive antigen, termed NY-CO-38/PDZ-73 [2]. The se-quence of the PDZ-73 cDNA (GenBank accessionnumber AF039700) is identical to the correspondingcDNA obtained from normal colon by 5P and 3PRACE procedures. The PDZ-73 nucleotide sequence(Fig. 1A) contains an open reading frame of 1956nucleotides beginning with a translation initiationconsensus sequence located at bp 97, and is followedby a 3P untranslated region of 182 bp containing apolyadenylation signal at bp 2215.
Translation of the PDZ-73 open reading framegives a protein of 652 amino acids (73 337 Da) con-taining a single N-glycosylation site, 17 sites of po-tential phosphorylation and a bipartite nuclear local-ization signal at residues 342 and 357. Notransmembrane domains or long hydrophobic re-gions are present. As shown in Fig. 1A,B, thePDZ-73 protein is organized into several domains,including a putative PEST protein degradation motif(residues 403^431), a coiled-coil domain (residues301^377), and three PDZ domains (PDZ 1, residues86^155; PDZ 2, residues 212^281; PDZ 3, residues451^525). The PDZ domains were identi¢ed by ascan of the amino acid sequence against the pro¢leentries in PROSITE, with PDZ 1, PDZ 2 and PDZ 3giving normalized scores of 16.7472, 15.9834, and14.4759, respectively, for PDZ domain pro¢lePS50106. A comparison of the PDZ domains inPDZ-73 to those in PSD-95 (accession no.U83192), hDlg (accession no. U13896) and ZO-1 (ac-cession no. L14837) shows that PDZ-73 is 36%, 36%,and 32% identical in PDZ 1, 32%, 30% and 24%identical in PDZ 2 and 13%, 15% and 28% identicalin PDZ 3, to PSD-95, hDlg and ZO-1, respectively.For comparison, the three PDZ domains of PSD-95and hDlg are nearly identical (95^97% identity),while ZO-1 is 41%, 24% and 15% identical to hDlgat PDZ 1, PDZ 2 and PDZ 3, respectively. The pro-gram used to identify the PEST motif (PEST¢nd)de¢nes these sequences as a hydrophilic stretch of12 or more amino acids, containing at least one P,one E or D and one S or T residue and £anked by K,R or H residues. Amino acids at positions 403^431of PDZ-73 gave a PEST¢nd score of +9.63, withvalues above +5.0 considered relevant. The coiled-coil domain was identi¢ed using Coils software[12]. Also, within the carboxyl terminal 9 aminoacids of PDZ-73 there is a putative PDZ domainbinding module of 7 amino acids (DSHSFPL), sim-ilar to a carboxyl terminal binding module foundin the PDZ-interacting protein SRY (YSHWTKL[13]).
SEREX screening of the metastatic colon cancercDNA library yielded an additional clone, termedNY-CO-37/PDZ-45 (GenBank accession numberAF039699), that was identical to PDZ-73 exceptfor the absence of a 74 bp segment (bp 1307^1380of PDZ-73). A stop codon is formed precisely at the
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5244

splice junction of the missing segment resulting in atruncated reading frame. This gives a protein of 403amino acids (45 839 Da) containing PDZ 1, PDZ 2and the coiled-coil domain but lacking PDZ 3 andthe carboxyl terminus (Fig. 1A,B). Furthermore, theomitted nucleotides correspond exactly to those cod-ing for the putative PEST sequence. For this reason,PDZ-45 would be expected to have a longer cellularhalf-life as compared to PDZ-73. A nested RT^PCR
strategy of normal colon cDNA, based on initialampli¢cation of the region surrounding the splicejunction followed by nested ampli¢cation internalto this region, yielded two products (data notshown). The DNA sequences of these products cor-responded to PDZ-73 and PDZ-45, indicating thatPDZ-45 represents a related isoform generated byalternate splicing, not a cloning artifact or a resultof a gene mutation in the tumor of origin.
Table 1The expression pattern of PDZ-73 isoforms in normal tissues, renal cancer, colon cancer and tumor cell lines
PDZ-73 PDZ-45 PDZ-54 PDZ-59 Northern blot Immunohistochemistry
Normal tissueKidney + + + + ++++ Positive (proximal and distal tubules)Colon mucosa + + Negative Negative +++ Positive (epithelium)Small intestine + + Negative Negative +++ Positive (epithelium)Brain + + + Negative +++* Positive (neuronal nuclei)Testis + + Negative Negative + NegativePancreas + Negative Negative Negative +++ NegativeLung + Negative Negative Negative Negative NegativeLiver + Negative Negative Negative ++ NegativeHeart + Negative Negative Negative Negative NegativeBreast + Negative Negative Negative NT NegativeBladder + Negative Negative + NT NegativeFetal brain + Negative Negative Negative NT NTAdrenal gland + Negative Negative Negative NT NTUterus Negative Negative Negative Negative NT NTSkeletal muscle Negative Negative Negative Negative Negative NTPBL Negative Negative Negative Negative Negative NTPlacenta Negative Negative Negative Negative Negative NTSpleen Negative Negative Negative Negative Negative NTThymus Negative Negative Negative Negative Negative NTEsophagus Negative Negative Negative Negative NT NTProstate NT NT NT NT Negative NTOvary NT NT NT NT Negative NT
Tumor tissueRenal cancer (4) +(2/4) +(2/4) +(2/4) +(2/4) NT Weak focal staining patternColon cancer (14) +(14/14) +(14/14) +(14/14) Negative NT Positive (neoplastic epithelium)
Colon cancer cell linesHCT 15 Negative Negative Negative Negative Negative NTColo 205 + + + Negative +++ NTHT 29 + + Negative Negative ++ NT
The data are presented for RT^PCR expression analysis of four PDZ-73 isoforms. Results from Northern blotting experiments arealso shown, as is the immunohistochemical analysis of PDZ-73 protein expression. These data illustrate the restricted tissue distribu-tion of the various isoforms with PDZ-73 being the most widely expressed, while PDZ-45 demonstrates greater tissue restriction andPDZ-54 and PDZ-59 demonstrate highly restricted expression. The signal detected in Northern blots of brain mRNA (*) re£ects ex-pression of the putative PDZ-37 isoform. A comparison of RT^PCR, Northern blot and immunohistochemistry indicates that anti-PDZ-73 monoclonal antibody staining re£ects expression of the more stable PDZ-45 isoform. Scoring of Northern blot signals (levelof mRNA expression): +, weak; ++, moderate; +++, strong; ++++, intense. NT, not tested.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^52 45

3.2. Tissue mRNA expression analysis of the PDZ-73and PDZ-45 isoforms: identi¢cation of additionalisoforms
As shown in Table 1, the mRNA expression pat-tern of the PDZ-73 and of PDZ-45 isoforms wasinvestigated by RT^PCR using an RNA panel de-rived from various normal and malignant tissues.PDZ-73 mRNA expression was detected in 13/20normal tissues as well as in all colon tumors (14/14cases) and in renal cancer (2/4 cases). Its expressionin normal lung and heart, not previously shown byNorthern blot (see below), probably re£ects the abil-ity of RT^PCR to amplify trace amounts of tem-
plate. Expression of the PDZ-45 isoform was deter-mined by RT^PCR using a 3P primer complementaryto the sequence of the splice junction. Comparedwith PDZ-73, this isoform displayed a more re-stricted expression pattern, being detected only incolon, small intestine, stomach, testis, brain and kid-ney (5/20 normal tissues), as well as in all PDZ-73positive tumors of the colon and kidney.
As shown in Fig. 2, hybridization of a PDZ-73probe (bp 1480^2019) to Northern blots containingmRNA from various human tissues and tumor celllines revealed a major hybridization signal of 2.4 kb.Similar signals were seen in 6/15 tissues tested (Fig. 3,small intestine, colon, pancreas, kidney, brain and
Fig. 2. The cDNA encoding PDZ-73 identi¢es a 2.4 kb and 1.2 kb RNA. (A) Northern blot analysis of mRNA derived from varioushuman tissues was undertaken using a 32P-labeled PDZ-73 cDNA probe as described in Section 2. Arrowhead, region identi¢ed byprobe. (B) Samples of mRNA isolated from various human tumor cell lines were analyzed by Northern blotting as above. The celllines analyzed include the Colo 205 colon tumor, HT 29 colon tumor, HCT 15 colon tumor, MCF-7 breast tumor, LA15S neuroblas-toma, MFSH histiocytoma, SK-OV-6 ovarian tumor and HS27 newborn ¢broblast cell lines. Arrowhead, region identi¢ed by probe.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5246
liver). This 2.4 kb signal was also detected in testisbut required extended autoradiographic exposure(data not shown). Two of eight cell lines (Fig. 2B,Colo 205 and HT29 colon tumor cell lines) also gavea 2.4 kb hybridization band. Normal human HS27¢broblasts failed to hybridize with this probe (Fig.2B). In brain tissue, and more speci¢cally, spinalcord and medulla, the major PDZ-73 hybridizationsignal corresponded to a size of 1.2 kb with a minorsignal at 2.4 kb (Fig. 2A). Northern blots using aprobe corresponding to the 5P end of PDZ-73 (bp54^443) failed to detect the 1.2 kb transcript (datanot shown). Using human brain cDNA as the tem-plate, a set of 5P RACE products were cloned whichshowed a uniform size ( þ 30 bp) and complete se-
quence identity with PDZ-73. The sequence of thelargest of these RACE products has a 5P end equiv-alent to bp 931 of PDZ-73, corresponding to a pu-tative transcript composed of 1304 nucleotides and isin general agreement with the size of the major hy-bridizing species present in Northern blots of humanbrain mRNA (Fig. 2A). Several methionine codonsare present near the 5P end of this putative transcript,two of which conform to the translation initiationconsensus (bp 136 and 172 equivalent to bp 1066and bp 1102 of PDZ-73, Fig. 1A). These data suggestthe existence of a lower molecular mass PDZ-73 iso-form resulting from translation beginning at alterna-tive start sites. This 1.3 kb PDZ-73 transcript en-codes a putative protein of 328 amino acids (37 195
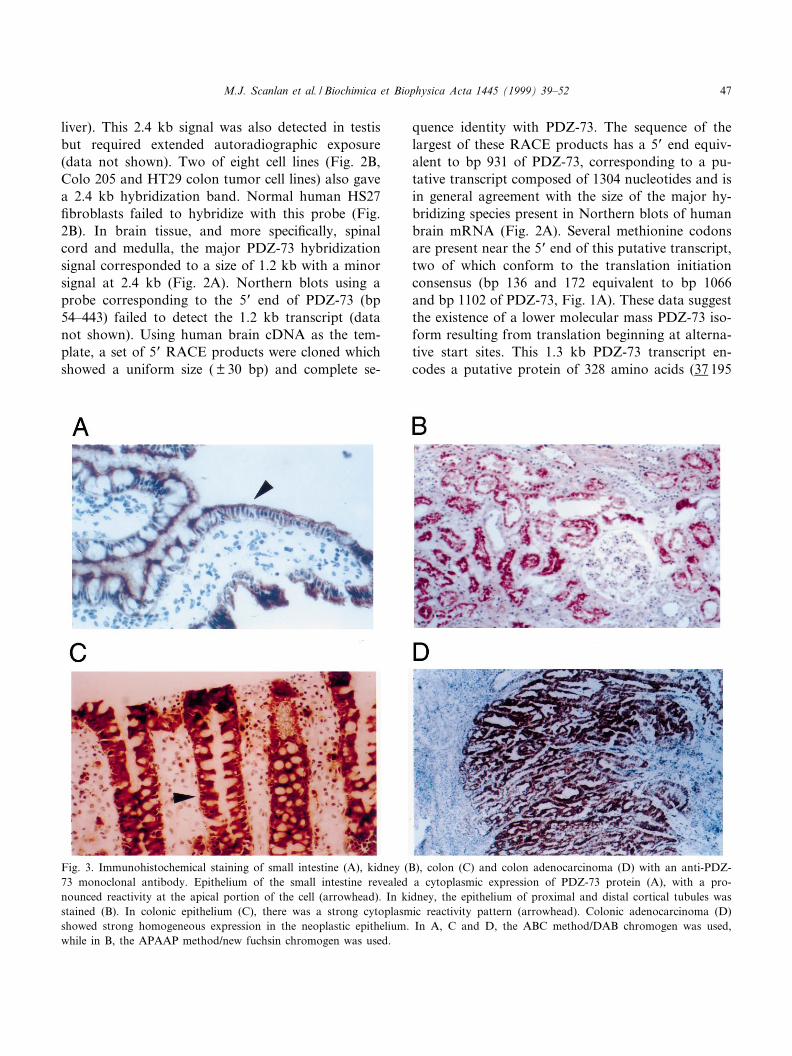
Fig. 3. Immunohistochemical staining of small intestine (A), kidney (B), colon (C) and colon adenocarcinoma (D) with an anti-PDZ-73 monoclonal antibody. Epithelium of the small intestine revealed a cytoplasmic expression of PDZ-73 protein (A), with a pro-nounced reactivity at the apical portion of the cell (arrowhead). In kidney, the epithelium of proximal and distal cortical tubules wasstained (B). In colonic epithelium (C), there was a strong cytoplasmic reactivity pattern (arrowhead). Colonic adenocarcinoma (D)showed strong homogeneous expression in the neoplastic epithelium. In A, C and D, the ABC method/DAB chromogen was used,while in B, the APAAP method/new fuchsin chromogen was used.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^52 47

Da), and has been termed the PDZ-37 isoform. Asshown in Fig. 1A,B, it may contain a portion of thecoiled-coil domain, the PDZ 3 domain, the putativePEST domain, and the COOH terminus but lacksthe PDZ 1 and PDZ 2 domains.
RT^PCR reactions designed to amplify the regionsurrounding the PDZ-45 splice junction yielded addi-tional products from normal kidney, bladder andbrain RNA not detected in other normal tissues.The cloning and sequencing of these RT^PCR prod-ucts showed they represent additional PDZ-73 var-iants, referred to as PDZ-54 and PDZ-59. The PDZ-54 isoform contains a 53 bp insertion at the splicesite previously de¢ned for PDZ-45 (bp 1380, Fig.1A,C). This insertion alters the reading frame, creat-ing a truncated protein of 481 amino acids (54 631Da) which contains the PDZ 1, PDZ 2, coiled-coiland putative PEST domain but lacks the PDZ 3domain and the carboxyl terminal domain (Fig.1B). It also contains an additional carboxyl terminal`tail' of 53 amino acids, not present in PDZ-73,which contains two serine/threonine phosphorylationmotifs. As shown in Table 1, a restricted mRNAexpression pattern was observed for PDZ-54, withtranscripts detected only in normal kidney and brain,as well as in PDZ-73 positive colon and renal tu-mors. The PDZ-59 isoform (521 amino acids,59 006 Da) contains a 173 bp insertion, also at thePDZ-45 splice site (bp 1380, Fig. 1A,C), and includesthe PDZ-54 insertion plus an additional 120 bp im-mediately 5P to it (Fig. 1C), which encodes a carbox-yl terminal tail of 93 amino acids. The PDZ-59 iso-form was detected only in normal kidney andbladder and also in renal cancer. The carboxyl ter-mini of PDZ-54 and PDZ-59 lack potential PDZinteracting modules. The putative PDZ-54 andPDZ-59 proteins are similar to PDZ-45 but di¡erby the presence of a possible PEST sequence andby their carboxyl terminal amino acid sequences(Fig. 1B,C).
3.3. Immunochemical relativity with anti-PDZ-73monoclonal antibodies
Table 1 lists 11 tissues that were tested for PDZ-73protein expression by immunohistochemistry withanti-PDZ 73 monoclonal antibodies (mAbs Co25and Co137). The most intense staining in normal
tissues was observed in the epithelium of the smallintestine, kidney and colon. The epithelium of thesmall intestine revealed cytoplasmic expression ofPDZ-73, with a pronounced reactivity at the apicalcell border (Fig. 3A). In the kidney, epithelial cells ofthe proximal and distal cortical tubules were stained(Fig. 3B). The renal medulla lacked PDZ-73 proteinexpression. In colonic epithelium, strong cytoplasmicreactivity was observed (Fig. 3C). In colonic adeno-carcinoma, neoplastic cells showed homogenous ex-pression of PDZ-73 (Fig. 3D). In addition to thetissues listed in Table 1, the following tissues werenegative by immunohistochemistry: skin, peripheralnerve, lymph node, prostatic adenocarcinoma, malig-nant melanoma, ductal breast carcinoma, ovariancarcinoma (serous^papillary) and squamous cell car-cinoma of the lung.
As shown in Table 1, all tissues, with the exceptionof testis, which are both PDZ-73- and PDZ-45-pos-itive by RT^PCR are positive for protein expressionby immunohistochemistry. Tissues that do not ex-press PDZ-45, but are PDZ-73-positive (pancreas,lung, liver, heart and breast), give weak focal stain-ing or are negative by immunohistochemistry. In par-ticular, pancreatic tissue shows a high level of PDZ-73 expression as seen in Northern blots (Fig. 2A),but is negative for PDZ-45 expression by RT^PCRand lacks immunohistochemical reactivity with anti-PDZ-73 mAbs. Together, these results suggest thatimmunohistochemical staining re£ects speci¢c ex-pression of the PDZ-45 isoform or the rate ofPDZ-73 protein turnover, i.e., the presence of thePEST protein degradation sequence in PDZ-73 con-fers upon it a shorter cellular half-life and it is there-fore not detected by immunohistochemistry.
Western blots were prepared from lysates of Colo205, HT 29 and HCT 15 cells using anti-PDZ 73monoclonal antibodies (Co25, Co36, Co137, andCo425). All antibodies yielded similar results withCo36 providing the strongest signal, results of whichare shown in Fig. 4. Western blots detected a proteinof 48 kDa in Colo 205 and HT 29 cell lysates thatwas absent in lysates from HCT 15 cells. As shownin Table 1 and Fig. 2B, both Colo 205 and HT 29cells express both PDZ-73 and PDZ-45 transcriptswhile HCT 15 is negative for their expression. Nohigher molecular mass bands unique to Colo 205and HT 29 were detected. If possible glycosylation
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5248

events are taken into account, these data suggest thatthe 48 kDa band unique to Colo 205 and HT 29represents the PDZ-45 protein.
3.4. Mapping of the PDZ-73 gene
A 100 kb BAC genomic clone was isolated byhybridization screening and shown by DNA sequenc-ing to contain the PDZ-73 cDNA. A portion of thecDNA sequence, as well as an intron sequence, wasidentical to two di¡erent sequence tagged sites fromchromosome 11 (GenBank accession numbersB03396 and B04920, respectively). Fine mapping ofPDZ-73 by FISH (Fig. 5A,B) showed the PDZ-73 tobe a single copy gene located at chromosomal band11p15.4^p15.1.
4. Discussion
The PDZ-73 protein was previously de¢ned as anovel antigen (NY-CO-38) recognized by serum IgGautoantibodies present in 23% and 17% of the pa-tients with renal and colon cancer, respectively. Itwas not reactive with sera from normal controls.The current study indicates that expression of thePDZ-73 gene produces ¢ve transcripts that di¡er bytheir tissue mRNA expression pattern. These tran-scripts encode putative protein isoforms that varywith respect to the number of PDZ domains, carbox-
yl terminal amino acid sequence, and predicted cel-lular half-lives. Immunohistochemistry with anti-PDZ-73 mAbs demonstrated high levels of proteinexpression in gastrointestinal epithelium, kidney tu-bules and colon cancer that correlated with mRNAexpression of the PDZ-45 isoform. DNA sequenceand FISH analysis of a PDZ-73 genomic clone re-vealed that this protein is encoded by a single copygene which maps to distal end of the short arm ofchromosome 11 at position 11p15.4^p15.1. Interest-ingly, deletions in this region have been associatedwith ovarian [14,15] and bladder cancer [16].
PDZ-73 can be considered a member of a diversefamily of proteins characterized by the presence ofmultiple PDZ domains. Such protein domains arebelieved to function as speci¢c sca¡olding sites forthe organization of cytoskeletal elements and signal-ing molecules into multiprotein complexes [3]. Forexample, the ¢rst and second PDZ domains of post-synaptic density protein 95 (PSD-95) interact withPDZ domain binding motifs in both Shaker typeK� channels [17] and the N-methyl-D^aspartate(NMDA) receptor [18,19], leading to co-clusteringof these ion channels. Speci¢c homophilic interac-tions between PDZ domains also take place. This isthe case with the PDZ^PDZ domain interactionsthat occur between PSD-95 and nitric oxide synthe-tase (nNOS) which leads to the formation of a tri-meric complex consisting of PSD-95, nNOS and theNMDA receptor at neuronal synapses [20]. The abil-ity of PDZ domains to link membrane-bound pro-teins to the cytoskeleton is exempli¢ed in erythro-cytes, whereby the band 4.1 actin binding proteinbinds to p55 which concurrently interacts, via itsPDZ domain, with glycophorin C [5,21]. PDZ-73may have similar functional characteristics, beingcapable of binding one or more associated proteinsby means of its three PDZ domains, as well as pos-sibly form homophilic PDZ interactions and bind toother PDZ domain proteins via its putative carboxylterminal PDZ binding module. The identi¢cation ofmolecules that interact with PDZ-73 may be possibleby using the anti-PDZ-73 mAbs generated in thisstudy in immunoprecipitation assays. Recently, ayeast 2 hybrid screen of proteins that interact withthe Coxsackie and adenovirus receptor (CAR [22])identi¢ed PDZ-73 as an interacting protein (R. Fin-berg, personal communication). CAR, a transmem-
Fig. 4. PDZ-45 protein expression in colon cancer cell lines.The Co36 monoclonal antibody detected a 48 kDa protein (ar-rowhead) in Western blots of Colo 205 and HT 29 lysates(lanes 1 and 2, respectively). This protein was absent in HCT15 lysates (lane 3), a cell line that lacks detectable PDZ-73 andPDZ-45 transcripts (Table 1 and Fig. 3B). Based on size andimmunoreactivity, this 48 kDa band may represent the PDZ-45protein.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^52 49

Fig. 5. Localization of the PDZ-73 gene to chromosome position 11p15.4^p15.1 by £uorescence in situ hybridization. Slides were pre-pared as described in Section 2. (A). Partial metaphase spread counter-stained with PI showing co-hybridization of both a chromo-some 11-speci¢c K-satellite probe at the centromere (white arrow) and the PDZ-73 BAC probe (blue arrow), demonstrating speci¢csignals in the distal region of chromosome 11. (B) Partial R-banded metaphase spread counter-stained with PI showing co-hybridiza-tion of both a chromosome 11-speci¢c K-satellite probe at the centromere (white arrow) and the PDZ-73 BAC probe (blue arrow)demonstrating speci¢c signals at 11p15.4^p15.1.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5250

brane protein, contains a carboxyl terminal PDZbinding motif suggesting it may bind to one of thePDZ domains of PDZ-73 or its other isoforms.
Expression of PDZ-73 gene products is character-ized by variations in the number of PDZ domains,the presence/absence of a putative PEST protein deg-radation motif and the composition of the carboxylterminal residues. The PDZ-45 isoform, presentmainly in tissues of the gastrointestinal tract as wellas kidney, brain and testis, possesses PDZ domains 1and 2 but lacks the putative PEST sequence, PDZdomain 3 and the possible carboxyl terminal PDZbinding motif present in PDZ-73. This protein wouldhave a longer half-life and could not be involved inprotein^protein interactions requiring the third PDZdomain or the putative carboxyl terminal PDZ bind-ing motif. Based upon their similar structure butshorter predicted cellular half-life, the highly tissuerestricted variants, PDZ-59 and PDZ-54, may func-tion in a similar manner as PDZ-45, but more tran-siently. The putative central nervous system isoform,PDZ-37, contains only PDZ domain 3 and the car-boxyl terminal PDZ binding motif and thus, cannotmake the PDZ domain 1 and domain 2 interactionsassociated with PDZ-73, PDZ-45, PDZ-54 and PDZ-59. Overall, di¡erential expression of PDZ-73 iso-forms re£ects variations in the number of expressedPDZ domains. Such a phenomenon may restrict, in atissue-speci¢c manner, the number of potential bind-ing partners and suggests a possible regulatory rolefor PDZ-73 isoforms.
In terms of the number of PDZ domains and se-quence similarities within these domains, PDZ-73 ismost similar to members of the membrane associatedguanylate kinase family of proteins (MAGUK pro-tein family; PSD-95, hDlg and ZO-1). However, itlacks the SH3 and guanylate kinase domains presentin this family of junctional complex proteins. Thestrong apical expression of PDZ-73/45 in epithelialcells of the small intestine is consistent with its local-ization to submembranous sites, a characteristic ofmembers of the MAGUK family of proteins [3].With the exception of neuronal nuclei in the brain,no nuclear staining was observed with anti-PDZ-73mAbs in other tissues, although it does possess pu-tative nuclear localization signals. It is possible thatPDZ-73 could translocate to the nucleus under cer-tain conditions, as does Zo-1 [23].
The basis for the immunogenicity of PDZ-73 incancer patients is not known. The fact that thePDZ-54 and PDZ-59 isoforms are not expressed bynormal colonic epithelium but are expressed in coloncancer suggests that aberrant isoform expressioncould be the immunogenic event. Other possibilitiesinclude ampli¢ed expression of PDZ-73 isoforms oran unidenti¢ed mutation in the PDZ-73 gene.
References
[1] U. Sahin, Oë . Tu«reci, H. Schmitt, B. Cochlovius, T. Jo-hannes, R. Schmits, F. Stenner, G. Luo, I. Schobert, M.Pfreundschuh, Human neoplasms elicit multiple speci¢c im-mune responses in the autologous host, Proc. Natl. Acad.Sci. USA 92 (1995) 11810^11813.
[2] M.J. Scanlan, Y.-T. Chen, B. Williamson, A.O. Gure, E.Stockert, J.D. Gordan, Oë . Tureci, U. Sahin, M. Pfreund-schuh, L.J. Old, Characterization of human colon cancerantigens recognized by autologous antibodies, Int. J. Cancer76 (1998) 652^658.
[3] C.P. Ponting, C. Phillips, K.E. Davies, D.J. Blake, PDZdomains: targeting signalling molecules to sub-membrane-ous sites, BioEssays 19 (1997) 469^479.
[4] J. Saras, L. Claesson-Welsh, C.-H. Heldin, L.J. Gonez,Cloning and characterization of PTPL1, a protein tyrosinephosphatase with similarities to cytoskeletal-associated pro-teins, J. Biol. Chem. 269 (1994) 24082^24089.
[5] S.M. Marfatia, J.H. Morais-Cabral, A.C. Kim, O. Byro-nand, A.H. Chishti, The PDZ domain of human erythrocytep55 mediates its binding to the cytoplasmic carboxyl termi-nus of glycophorin C, J. Biol. Chem. 272 (1997) 24191^24197.
[6] K.-O. Cho, C.A. Hunt, M.B. Kennedy, The rat postsynapticdensity fraction contains a homolog of the Drosophila discs-large tumor suppressor protein, Neuron 9 (1992) 929^942.
[7] R.E. Lue, S.M. Marfatia, D. Branton, A.H. Chishti, Cloningand characterization of hhDlg: the human homologue of theDrosophila discs large tumor suppressor binds to protein4.1, Proc. Natl. Acad. Sci. USA 91 (1994) 9818^9822.
[8] E. Willott, M.-S. Balda, A.S. Fanning, B. Jameson, C. VanItallie, J.M. Anderson, The tight junction protein ZO-1 ishomologous to the Drosophila discs^large tumor suppressorprotein of septate junctions, Proc. Natl. Acad. Sci. USA 90(1993) 7834^7838.
[9] Z. Songyang, A.S. Fanning, C. Fu, J. Xu, S.M. Marfatia,A.H. Chishti, A. Crompton, A.C. Chan, J.M. Anderson,L.C. Cantley, Recognition of unique carboxyl-terminal mo-tifs by distinct PDZ domains, Science 275 (1997) 73^77.
[10] P. Chomczynski, One-hour downward alkaline capillarytransfer for blotting of DNA and RNA, Anal. Biochem.201 (1992) 134^139.
[11] Y.-T. Chen, E. Stockert, Y. Chen, P. Garin-Chesa, W.J.Rettig, P. van der Bruggen, T. Boon, L.J. Old, Identi¢cation
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^52 51

of the MAGE-1 gene product by monoclonal and polyclonalantibodies, Proc. Natl. Acad. Sci. USA 91 (1994) 1004^1008.
[12] A. Lupas, M. Van Dyke, J. Stock, Predicting coiled coilsfrom protein sequences, Science 252 (1991) 1162^1164.
[13] F. Poulat, P. de Santa Barbara, M. Desclozeaux, S. Soullier,B. Moniot, N. Bonneaud, B. Boizet, P. Berta, The humantestis determining factor SRY binds a nuclear factor contain-ing PDZ protein interaction domains, J. Biol. Chem. 272(1997) 7167^7172.
[14] K.H. Lu, J.N. Weitzel, S. Kodali, W.R. Welch, R.S. Berko-witz, S.C. Mok, A novel4-cM minimally deleted region onchromosome 11p15.1 associated with high grade nonmuci-nous epithelial ovarian carcinomas, Cancer Res. 57 (1997)387^390.
[15] D.M. Eccles, L. Gruber, M. Stewart, C.M. Steel, R.C. Leo-nard, Allele loss on chromosome 11p is associated with poorsurvival in ovarian cancer, Dis. Markers 10 (1992) 95^99.
[16] M.E. Shaw, M.A. Knowles, Deletion mapping of chromo-some 11 in carcinoma of the bladder, Genes ChromosomesCancer 13 (1995) 1^8.
[17] H.C. Kornau, L.T. Schenker, M.B. Kennedy, P.H. Seeburg,Domain interaction between NMDA receptor subunits andthe postsynaptic density protein PSD-95, Science 269 (1995)1737^1740.
[18] E. Kim, M. Niethammer, A. Rothschild, Y.N. Jan, M.Sheng, Clustering of Shaker-type K� channels by interaction
with a family of membrane-associated guanylate kinases,Nature 378 (1995) 85^88.
[19] M. Niethammer, E. Kim, M. Sheng, Interaction between theC terminus of NMDA receptor subunits and multiple mem-bers of the PSD-95 family of membrane-associated guanylatekinases, J. Neurosci. 16 (1996) 2157^2163.
[20] J.E. Brenman, D.S. Chao, S.H. Gee, A.W. McGee, S.E.Craven, D.R. Santillano, F. Huang, H. Xia, M.F. Peters,S.C. Froehner, D.S. Bredt, Interaction of nitric oxide syn-thase with the postsynaptic density protein PSD-95 andalpha1-syntrophin mediated by PDZ domains, Cell 84(1996) 757^767.
[21] S.M. Marfatia, R.A. Lue, D. Branton, A.H. Chishti, In vitrobinding studies suggest a membrane-associated complex be-tween erythroid p55, protein 4.1, and glycophorin C, J. Biol.Chem. 269 (1994) 8631^8634.
[22] J.M. Bergelson, J.A. Cunningham, G. Droguett, E.A. Kurt-Jones, A. Krithivas, J.S. Hong, M.S. Horwitz, R.L. Crowell,R.W. Finberg, Isolation of a common receptor for Coxsack-ie B viruses and adenoviruses 2 and 5, Science 275 (1997)1320^1323.
[23] C.J. Gottardi, M. Arpin, A.S. Fanning, D. Louvard, Thejunction-associated protein, zonula occludens-1, localizes tothe nucleus before the maturation and during the remodelingof cell^cell contacts, Proc. Natl. Acad. Sci USA 93 (1996)10779^10784.
BBAEXP 93258 1-4-99 Cyaan Magenta Geel Zwart
M.J. Scanlan et al. / Biochimica et Biophysica Acta 1445 (1999) 39^5252




















