
Is there “neural efficiency” during the processing of visuo-spatial information in male humans?...
7
Author's personal copy Behavioural Brain Research 205 (2009) 468–474 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Research report Is there “neural efficiency” during the processing of visuo-spatial information in male humans? An EEG study Paolo Capotosto a , M. Gianni Perrucci a , Marcella Brunetti a , Cosimo Del Gratta a , Michael Doppelmayr b , Roland H. Grabner c , Wolfgang Klimesch b , Aljoscha Neubauer d , Christa Neuper d , Gert Pfurtscheller d , Gian Luca Romani a , Claudio Babiloni e,f,∗ a ITAB e Dipartimento di Scienze Cliniche e Bioimmagini, Università G. D’Annunzio, Chieti, Italy b University of Salzburg, Salzburg, Austria c Institute for Behavioral Sciences, Swiss Federal Institute of Techology (ETH) Zurich, Switzerland d University of Graz, Graz, Austria e Department of Biomedical Sciences, University of Foggia, Foggia, Italy f Casa di Cura San Raffaele Cassino, Italy article info Article history: Received 20 July 2009 Received in revised form 27 July 2009 Accepted 30 July 2009 Available online 7 August 2009 Keywords: Event-related desynchronization/synchronization (ERD/ERS) Alpha rhythms Theta rhythms Neural efficiency Encoding and retrieval abstract More intelligent persons (high IQ) typically present a higher cortical activity during tasks requiring the encoding of visuo-spatial information, namely higher alpha (about 10 Hz) event-related desynchroniza- tion (ERD; Doppelmayr et al., 2005 [23]). The opposite is true (“neural efficiency”) during the retrieval of the encoded information, as revealed by both lower alpha ERD and/or lower theta (about 5 Hz) event- related synchronization (ERS; Grabner et al., 2004 [19]). To reconcile these contrasting results, here we evaluated the working hypothesis that more intelligent male subjects are characterized by a high cortical activity during the encoding phase. This deep encoding would explain the relatively low cortical activity for the retrieval of the encoded information. To test this hypothesis, electroencephalographic (EEG) data were recorded in 22 healthy young male volunteers during visuo-spatial information processing (encod- ing) and short-term retrieval of the encoded information. Cortical activity was indexed by theta ERS and alpha ERD. It was found that the higher the subjects’ total IQ, the stronger the frontal theta ERS during the encoding task. Furthermore, the higher the subjects’ total IQ, the lower the frontal high-frequency alpha ERD (about 10–12 Hz) during the retrieval task. This was not true for parietal counterpart of these EEG rhythms. These results reconcile previous contrasting evidence confirming that more intelligent per- sons do not ever show event-related cortical responses compatible with “neural efficiency” hypothesis. Rather, their cortical activity would depend on flexible and task-adapting features of frontal activation. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Previous neuroimaging studies using positron emission tomog- raphy (PET), single photon emission computed tomography (SPECT), and functional magnetic resonance imaging (fMRI) have shown that more intelligent subjects present a weaker fronto- parietal activation during cognitive tasks [1–9]. To explain these data, it has been proposed the so-called “neural efficiency” hypoth- esis, which postulates a more efficient brain function in brighter as compared to less intelligent individuals [1,10]. Other neuroimaging evidence challenged this hypothesis, showing that fronto-parietal ∗ Corresponding author at: Department of Biomedical Sciences, University of Fog- gia, V.le Pinto, 1, 71100 Foggia, Italy. Tel.: +39 0881 713276; fax: +39 0881 711716. E-mail address: [email protected] (C. Babiloni). cortical activation during cognitive tasks was stronger in more intelligent subjects [11,12]. To shed light on the physiological mechanism at the basis of the “neural efficiency” hypothesis, event-related changes in power of electroencephalographic (EEG) rhythms at about 4–7 Hz (theta) and 8–12 Hz (alpha) were used. Cortical activity accompanying cog- nitive and motor tasks is typically associated with a percentage increase of power of theta rhythms (theta event-related synchro- nization, ERS) and with a percentage decrease of power of resting alpha rhythms (alpha event-related desynchronization, ERD: [13]). In line with the “neural efficiency” hypothesis, it has been shown that more intelligent persons (high IQ) present a lower alpha ERD and/or a lower theta ERS during different types of cogni- tive tasks [14–21]. A negative relationship between the alpha ERD and intelligence has been also observed in individuals with high domain-specific skills such as chess players, etc. [22]. 0166-4328/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2009.07.032
Transcript of Is there “neural efficiency” during the processing of visuo-spatial information in male humans?...

Author's personal copy
Behavioural Brain Research 205 (2009) 468–474
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
Research report
Is there “neural efficiency” during the processing of visuo-spatial information inmale humans? An EEG study
Paolo Capotostoa, M. Gianni Perruccia, Marcella Brunetti a, Cosimo Del Grattaa,Michael Doppelmayrb, Roland H. Grabnerc, Wolfgang Klimeschb, Aljoscha Neubauerd,Christa Neuperd, Gert Pfurtschellerd, Gian Luca Romania, Claudio Babilonie,f,∗
a ITAB e Dipartimento di Scienze Cliniche e Bioimmagini, Università G. D’Annunzio, Chieti, Italyb University of Salzburg, Salzburg, Austriac Institute for Behavioral Sciences, Swiss Federal Institute of Techology (ETH) Zurich, Switzerlandd University of Graz, Graz, Austriae Department of Biomedical Sciences, University of Foggia, Foggia, Italyf Casa di Cura San Raffaele Cassino, Italy
a r t i c l e i n f o
Article history:Received 20 July 2009Received in revised form 27 July 2009Accepted 30 July 2009Available online 7 August 2009
Keywords:Event-relateddesynchronization/synchronization(ERD/ERS)Alpha rhythmsTheta rhythmsNeural efficiencyEncoding and retrieval
a b s t r a c t
More intelligent persons (high IQ) typically present a higher cortical activity during tasks requiring theencoding of visuo-spatial information, namely higher alpha (about 10 Hz) event-related desynchroniza-tion (ERD; Doppelmayr et al., 2005 [23]). The opposite is true (“neural efficiency”) during the retrievalof the encoded information, as revealed by both lower alpha ERD and/or lower theta (about 5 Hz) event-related synchronization (ERS; Grabner et al., 2004 [19]). To reconcile these contrasting results, here weevaluated the working hypothesis that more intelligent male subjects are characterized by a high corticalactivity during the encoding phase. This deep encoding would explain the relatively low cortical activityfor the retrieval of the encoded information. To test this hypothesis, electroencephalographic (EEG) datawere recorded in 22 healthy young male volunteers during visuo-spatial information processing (encod-ing) and short-term retrieval of the encoded information. Cortical activity was indexed by theta ERS andalpha ERD. It was found that the higher the subjects’ total IQ, the stronger the frontal theta ERS duringthe encoding task. Furthermore, the higher the subjects’ total IQ, the lower the frontal high-frequencyalpha ERD (about 10–12 Hz) during the retrieval task. This was not true for parietal counterpart of theseEEG rhythms. These results reconcile previous contrasting evidence confirming that more intelligent per-sons do not ever show event-related cortical responses compatible with “neural efficiency” hypothesis.Rather, their cortical activity would depend on flexible and task-adapting features of frontal activation.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Previous neuroimaging studies using positron emission tomog-raphy (PET), single photon emission computed tomography(SPECT), and functional magnetic resonance imaging (fMRI) haveshown that more intelligent subjects present a weaker fronto-parietal activation during cognitive tasks [1–9]. To explain thesedata, it has been proposed the so-called “neural efficiency” hypoth-esis, which postulates a more efficient brain function in brighter ascompared to less intelligent individuals [1,10]. Other neuroimagingevidence challenged this hypothesis, showing that fronto-parietal
∗ Corresponding author at: Department of Biomedical Sciences, University of Fog-gia, V.le Pinto, 1, 71100 Foggia, Italy. Tel.: +39 0881 713276; fax: +39 0881 711716.
E-mail address: [email protected] (C. Babiloni).
cortical activation during cognitive tasks was stronger in moreintelligent subjects [11,12].
To shed light on the physiological mechanism at the basis ofthe “neural efficiency” hypothesis, event-related changes in powerof electroencephalographic (EEG) rhythms at about 4–7 Hz (theta)and 8–12 Hz (alpha) were used. Cortical activity accompanying cog-nitive and motor tasks is typically associated with a percentageincrease of power of theta rhythms (theta event-related synchro-nization, ERS) and with a percentage decrease of power of restingalpha rhythms (alpha event-related desynchronization, ERD: [13]).In line with the “neural efficiency” hypothesis, it has been shownthat more intelligent persons (high IQ) present a lower alphaERD and/or a lower theta ERS during different types of cogni-tive tasks [14–21]. A negative relationship between the alpha ERDand intelligence has been also observed in individuals with highdomain-specific skills such as chess players, etc. [22].
0166-4328/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.bbr.2009.07.032

Author's personal copy
P. Capotosto et al. / Behavioural Brain Research 205 (2009) 468–474 469
Several EEG studies have challenged the “neural efficiency”hypothesis. During a difficult verbal-semantic task (no short-termmemory demands), an increase of the theta ERS has been shown inintelligent persons engaged in encoding tasks [23,24].
The contrasting results regarding the “neural efficiency”hypothesis could be due to different reasons. The mentionedstudies employed tasks with marked differences in the stimu-lus features, task structure, and cognitive processes (i.e. encoding,imagery, and retrieval). Furthermore, more intelligent persons arecharacterized by high cortical activity when they perceive and workon the external stimuli to be memorized (encoding) as revealedby theta ERS (at odds with “neural efficiency” hypothesis), thusallowing an easy retrieval of encoded information by a low corticalactivity, as revealed by alpha ERD (in line with “neural efficiency”hypothesis). To reconcile this contrasting evidence, EEG data werecollected during a working memory task in which we focusedon encoding and retrieval phases. We evaluated the relationshipsamong the theta ERS, the alpha ERD, and the subjects’ IQ in bothphases. Specifically, the encoding phase pertained to a match tosample task, whereas the retrieval phase included a delayed matchto sample, with a possible requirement for mental spatial rotationin both phases. As a novelty, this design minimized the differencesas graphical features and motor outputs of the two phases. Further-more, it used the same male subjects for the encoding and retrievaltasks, thus reducing the inter-subjects variability of the EEG resultsand the effects of cyclic female sexual hormones on brain responsesand cognitive processes.
2. Methods
2.1. Subjects
Previous reference field investigations have shown that more intelligent persons(high IQ) typically present higher “encoding” theta ERS and lower “retrieval” alphaERD in large populations (N = about 50) of healthy subjects [19,23]. Based on theseprevious findings on large populations, here we could test clear working hypothesesabout the expected effects on encoding theta ERS and alpha ERD. As a consequence,
we recruited just 22 right-handed (Edinburgh Inventory) healthy adult volunteers(aged 20–34, mean 25.9 ± 4.5 S.D.). Specifically, all subjects recruited were male,to avoid effects of different phases of the menstrual cycle in females. They hadno previous psychiatric or neurological history, and their sight was normal or cor-rected to normal. The subjects received a full administration of the official Italianversion of Wechsler Adult Intelligence Scale [25,26]. The total IQ score was used as aglobal index of intelligence. All experiments were conducted with the understand-ing and written consent of each participant, according to the Code of Ethics of theWorld Medical Association, and the standards established by the Author’s Institu-tional Review Board. The procedures of this study were approved by the local ethicscommittee.
2.2. Experimental procedure
All experiments were conducted at the University of Chieti-ITAB (Italy). Thesubjects kept left arm resting and the right one was positioned on a response pad.The experimental paradigm is depicted in Fig. 1. There were two tasks. The first taskrequired the encoding and processing of visuo-spatial information to be memorized.The second task required the retrieval of the encoded information. For both tasks,the same kind of stimuli was used. In the first task, a visual stimulus lasting 2.5 s(display time) consisted of two arrows at the left and right side of a computer mon-itor. The arrows were formed by two perpendicular segments and were presentedat same or different angles of rotation. One or three bars were located in one (orthree) of 12 different positions perpendicularly to one or both segments. The posi-tion of these bars was the element that allowed discriminating among arrows. Thearrow at the left side of the monitor was the sample stimulus; the arrow at the rightside was to be matched to the sample arrow. The subjects were required to press(i) the left button of the mouse if the right arrow was equal to the left arrow (sam-ple), regardless of possible plane rotations; (ii) the right button of the mouse if theright arrow was different from the left arrow (sample). No feedback was provided.Instruction focused on both speed and accuracy of the responses. A response given2.5 s after the encoding stimulus disappearance was considered as “invalid”, and thecorresponding EEG trial was discarded. After the disappearance of the visuo-spatialstimulus, there were 10 s of baseline (white screen and black central cross) and,then, the beginning of the second task with the delivery of a further visual stimuluslasting 2.5 s. Of note, the encoding activity corresponded to stimulus display time(2.5 s) and did not include any of the 10 s delay period. This visual stimulus consistedof an “X” at the left monitor side and of an arrow at the right monitor side. The arrowat the right side was to be matched to the arrow of the preceding encoding visualstimulus. The subjects were required to press: (i) the left button of the mouse if theright arrow was equal to the previous sample stimulus, even regardless of possibleplane rotations; (ii) the right button of the mouse if the right arrow was different
Fig. 1. Sketch of the experimental design. An encoding visual stimulus lasting 2.5 s was constituted by two arrows at the left and right side of a computer monitor. The arrowis formed by two perpendicular segments. One or three bars are located in one (or three) of 12 different positions perpendicularly to one or both segments. The position ofthese bars is the element which allows to discriminate among arrows. The arrow at the left side of the monitor is the sample stimulus, the arrow at the right side is to bematched to the sample arrow. Subjects were required to press the left mouse button if the right arrow is equal to the left arrow (sample) even if spatially rotated (b) or theright mouse button if the right arrow is different to the left arrow (sample) (a). After the disappearance of the encoding stimulus, there were 10 s of baseline and then thedelivery of a retrieval visual stimulus lasting 2.5 s. The retrieval visual stimulus was constituted by an “X” at the left monitor side and by an arrow at the right monitor side.The arrow at the right side is to be matched to the arrow of the preceding encoding visual stimulus. Subjects were required to press the left mouse button if the right arrowis equal to the previous sample stimulus even if spatially rotated (a), or the right mouse button if the right arrow is different to the sample stimulus (b). After the retrievalstimulus disappearance, there were 10 s of post-event time before the subsequent encoding visual stimulus.
Author's personal copy
470 P. Capotosto et al. / Behavioural Brain Research 205 (2009) 468–474
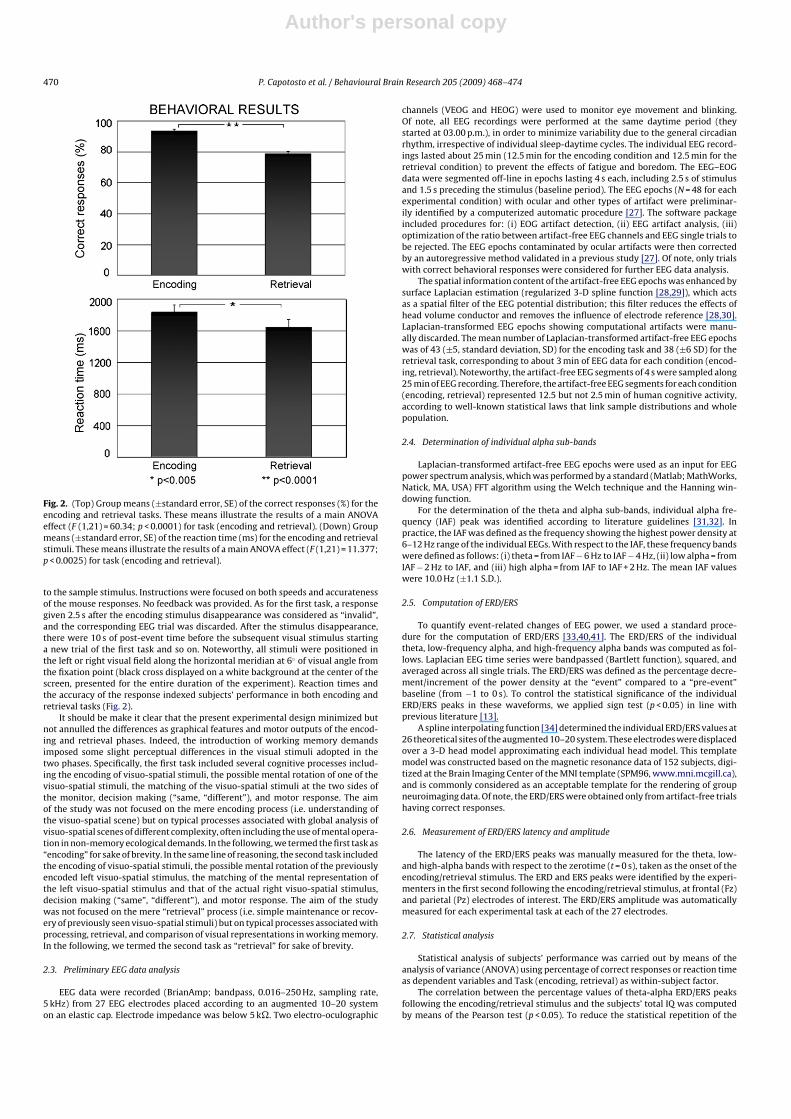
Fig. 2. (Top) Group means (±standard error, SE) of the correct responses (%) for theencoding and retrieval tasks. These means illustrate the results of a main ANOVAeffect (F (1,21) = 60.34; p < 0.0001) for task (encoding and retrieval). (Down) Groupmeans (±standard error, SE) of the reaction time (ms) for the encoding and retrievalstimuli. These means illustrate the results of a main ANOVA effect (F (1,21) = 11.377;p < 0.0025) for task (encoding and retrieval).
to the sample stimulus. Instructions were focused on both speeds and accuratenessof the mouse responses. No feedback was provided. As for the first task, a responsegiven 2.5 s after the encoding stimulus disappearance was considered as “invalid”,and the corresponding EEG trial was discarded. After the stimulus disappearance,there were 10 s of post-event time before the subsequent visual stimulus startinga new trial of the first task and so on. Noteworthy, all stimuli were positioned inthe left or right visual field along the horizontal meridian at 6◦ of visual angle fromthe fixation point (black cross displayed on a white background at the center of thescreen, presented for the entire duration of the experiment). Reaction times andthe accuracy of the response indexed subjects’ performance in both encoding andretrieval tasks (Fig. 2).
It should be make it clear that the present experimental design minimized butnot annulled the differences as graphical features and motor outputs of the encod-ing and retrieval phases. Indeed, the introduction of working memory demandsimposed some slight perceptual differences in the visual stimuli adopted in thetwo phases. Specifically, the first task included several cognitive processes includ-ing the encoding of visuo-spatial stimuli, the possible mental rotation of one of thevisuo-spatial stimuli, the matching of the visuo-spatial stimuli at the two sides ofthe monitor, decision making (“same, “different”), and motor response. The aimof the study was not focused on the mere encoding process (i.e. understanding ofthe visuo-spatial scene) but on typical processes associated with global analysis ofvisuo-spatial scenes of different complexity, often including the use of mental opera-tion in non-memory ecological demands. In the following, we termed the first task as“encoding” for sake of brevity. In the same line of reasoning, the second task includedthe encoding of visuo-spatial stimuli, the possible mental rotation of the previouslyencoded left visuo-spatial stimulus, the matching of the mental representation ofthe left visuo-spatial stimulus and that of the actual right visuo-spatial stimulus,decision making (“same”, “different”), and motor response. The aim of the studywas not focused on the mere “retrieval” process (i.e. simple maintenance or recov-ery of previously seen visuo-spatial stimuli) but on typical processes associated withprocessing, retrieval, and comparison of visual representations in working memory.In the following, we termed the second task as “retrieval” for sake of brevity.
2.3. Preliminary EEG data analysis
EEG data were recorded (BrianAmp; bandpass, 0.016–250 Hz, sampling rate,5 kHz) from 27 EEG electrodes placed according to an augmented 10–20 systemon an elastic cap. Electrode impedance was below 5 k�. Two electro-oculographic
channels (VEOG and HEOG) were used to monitor eye movement and blinking.Of note, all EEG recordings were performed at the same daytime period (theystarted at 03.00 p.m.), in order to minimize variability due to the general circadianrhythm, irrespective of individual sleep-daytime cycles. The individual EEG record-ings lasted about 25 min (12.5 min for the encoding condition and 12.5 min for theretrieval condition) to prevent the effects of fatigue and boredom. The EEG–EOGdata were segmented off-line in epochs lasting 4 s each, including 2.5 s of stimulusand 1.5 s preceding the stimulus (baseline period). The EEG epochs (N = 48 for eachexperimental condition) with ocular and other types of artifact were preliminar-ily identified by a computerized automatic procedure [27]. The software packageincluded procedures for: (i) EOG artifact detection, (ii) EEG artifact analysis, (iii)optimization of the ratio between artifact-free EEG channels and EEG single trials tobe rejected. The EEG epochs contaminated by ocular artifacts were then correctedby an autoregressive method validated in a previous study [27]. Of note, only trialswith correct behavioral responses were considered for further EEG data analysis.
The spatial information content of the artifact-free EEG epochs was enhanced bysurface Laplacian estimation (regularized 3-D spline function [28,29]), which actsas a spatial filter of the EEG potential distribution; this filter reduces the effects ofhead volume conductor and removes the influence of electrode reference [28,30].Laplacian-transformed EEG epochs showing computational artifacts were manu-ally discarded. The mean number of Laplacian-transformed artifact-free EEG epochswas of 43 (±5, standard deviation, SD) for the encoding task and 38 (±6 SD) for theretrieval task, corresponding to about 3 min of EEG data for each condition (encod-ing, retrieval). Noteworthy, the artifact-free EEG segments of 4 s were sampled along25 min of EEG recording. Therefore, the artifact-free EEG segments for each condition(encoding, retrieval) represented 12.5 but not 2.5 min of human cognitive activity,according to well-known statistical laws that link sample distributions and wholepopulation.
2.4. Determination of individual alpha sub-bands
Laplacian-transformed artifact-free EEG epochs were used as an input for EEGpower spectrum analysis, which was performed by a standard (Matlab; MathWorks,Natick, MA, USA) FFT algorithm using the Welch technique and the Hanning win-dowing function.
For the determination of the theta and alpha sub-bands, individual alpha fre-quency (IAF) peak was identified according to literature guidelines [31,32]. Inpractice, the IAF was defined as the frequency showing the highest power density at6–12 Hz range of the individual EEGs. With respect to the IAF, these frequency bandswere defined as follows: (i) theta = from IAF − 6 Hz to IAF − 4 Hz, (ii) low alpha = fromIAF − 2 Hz to IAF, and (iii) high alpha = from IAF to IAF + 2 Hz. The mean IAF valueswere 10.0 Hz (±1.1 S.D.).
2.5. Computation of ERD/ERS
To quantify event-related changes of EEG power, we used a standard proce-dure for the computation of ERD/ERS [33,40,41]. The ERD/ERS of the individualtheta, low-frequency alpha, and high-frequency alpha bands was computed as fol-lows. Laplacian EEG time series were bandpassed (Bartlett function), squared, andaveraged across all single trials. The ERD/ERS was defined as the percentage decre-ment/increment of the power density at the “event” compared to a “pre-event”baseline (from −1 to 0 s). To control the statistical significance of the individualERD/ERS peaks in these waveforms, we applied sign test (p < 0.05) in line withprevious literature [13].
A spline interpolating function [34] determined the individual ERD/ERS values at26 theoretical sites of the augmented 10–20 system. These electrodes were displacedover a 3-D head model approximating each individual head model. This templatemodel was constructed based on the magnetic resonance data of 152 subjects, digi-tized at the Brain Imaging Center of the MNI template (SPM96, www.mni.mcgill.ca),and is commonly considered as an acceptable template for the rendering of groupneuroimaging data. Of note, the ERD/ERS were obtained only from artifact-free trialshaving correct responses.
2.6. Measurement of ERD/ERS latency and amplitude
The latency of the ERD/ERS peaks was manually measured for the theta, low-and high-alpha bands with respect to the zerotime (t = 0 s), taken as the onset of theencoding/retrieval stimulus. The ERD and ERS peaks were identified by the experi-menters in the first second following the encoding/retrieval stimulus, at frontal (Fz)and parietal (Pz) electrodes of interest. The ERD/ERS amplitude was automaticallymeasured for each experimental task at each of the 27 electrodes.
2.7. Statistical analysis
Statistical analysis of subjects’ performance was carried out by means of theanalysis of variance (ANOVA) using percentage of correct responses or reaction timeas dependent variables and Task (encoding, retrieval) as within-subject factor.
The correlation between the percentage values of theta-alpha ERD/ERS peaksfollowing the encoding/retrieval stimulus and the subjects’ total IQ was computedby means of the Pearson test (p < 0.05). To reduce the statistical repetition of the
Author's personal copy
P. Capotosto et al. / Behavioural Brain Research 205 (2009) 468–474 471
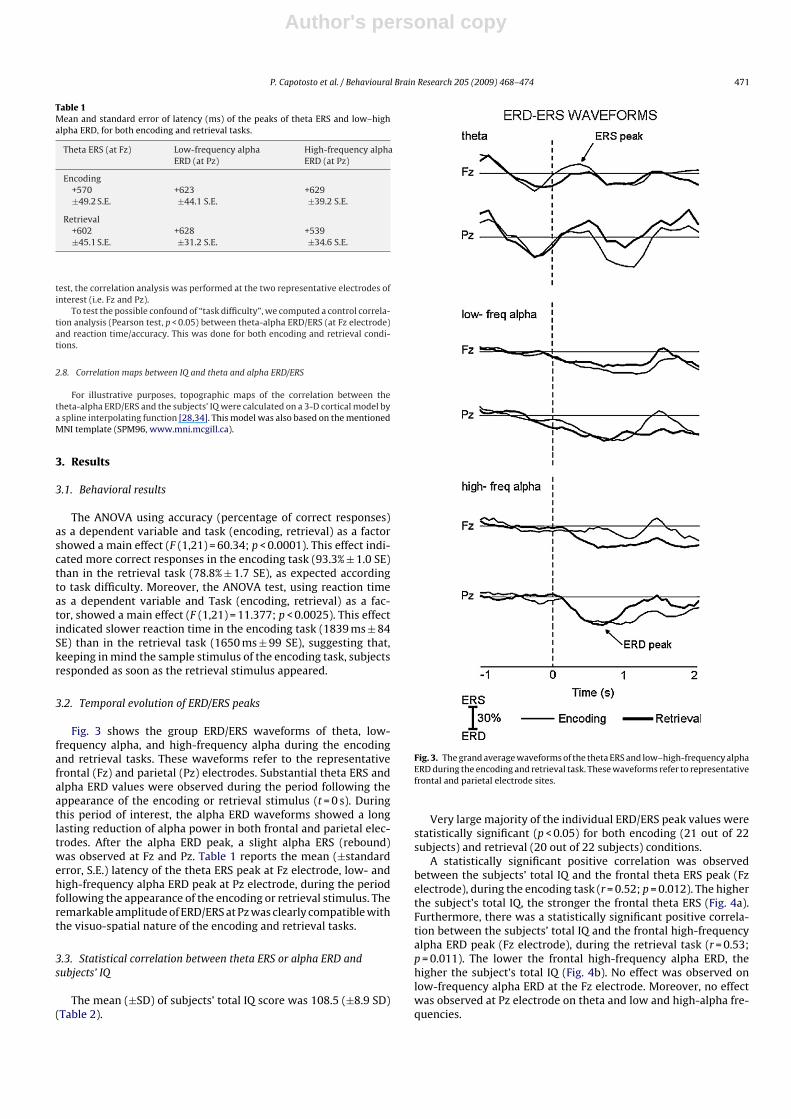
Table 1Mean and standard error of latency (ms) of the peaks of theta ERS and low–highalpha ERD, for both encoding and retrieval tasks.
Theta ERS (at Fz) Low-frequency alphaERD (at Pz)
High-frequency alphaERD (at Pz)
Encoding+570 +623 +629±49.2 S.E. ±44.1 S.E. ±39.2 S.E.
Retrieval+602 +628 +539±45.1 S.E. ±31.2 S.E. ±34.6 S.E.
test, the correlation analysis was performed at the two representative electrodes ofinterest (i.e. Fz and Pz).
To test the possible confound of “task difficulty”, we computed a control correla-tion analysis (Pearson test, p < 0.05) between theta-alpha ERD/ERS (at Fz electrode)and reaction time/accuracy. This was done for both encoding and retrieval condi-tions.
2.8. Correlation maps between IQ and theta and alpha ERD/ERS
For illustrative purposes, topographic maps of the correlation between thetheta-alpha ERD/ERS and the subjects’ IQ were calculated on a 3-D cortical model bya spline interpolating function [28,34]. This model was also based on the mentionedMNI template (SPM96, www.mni.mcgill.ca).
3. Results
3.1. Behavioral results
The ANOVA using accuracy (percentage of correct responses)as a dependent variable and task (encoding, retrieval) as a factorshowed a main effect (F (1,21) = 60.34; p < 0.0001). This effect indi-cated more correct responses in the encoding task (93.3% ± 1.0 SE)than in the retrieval task (78.8% ± 1.7 SE), as expected accordingto task difficulty. Moreover, the ANOVA test, using reaction timeas a dependent variable and Task (encoding, retrieval) as a fac-tor, showed a main effect (F (1,21) = 11.377; p < 0.0025). This effectindicated slower reaction time in the encoding task (1839 ms ± 84SE) than in the retrieval task (1650 ms ± 99 SE), suggesting that,keeping in mind the sample stimulus of the encoding task, subjectsresponded as soon as the retrieval stimulus appeared.
3.2. Temporal evolution of ERD/ERS peaks
Fig. 3 shows the group ERD/ERS waveforms of theta, low-frequency alpha, and high-frequency alpha during the encodingand retrieval tasks. These waveforms refer to the representativefrontal (Fz) and parietal (Pz) electrodes. Substantial theta ERS andalpha ERD values were observed during the period following theappearance of the encoding or retrieval stimulus (t = 0 s). Duringthis period of interest, the alpha ERD waveforms showed a longlasting reduction of alpha power in both frontal and parietal elec-trodes. After the alpha ERD peak, a slight alpha ERS (rebound)was observed at Fz and Pz. Table 1 reports the mean (±standarderror, S.E.) latency of the theta ERS peak at Fz electrode, low- andhigh-frequency alpha ERD peak at Pz electrode, during the periodfollowing the appearance of the encoding or retrieval stimulus. Theremarkable amplitude of ERD/ERS at Pz was clearly compatible withthe visuo-spatial nature of the encoding and retrieval tasks.
3.3. Statistical correlation between theta ERS or alpha ERD andsubjects’ IQ
The mean (±SD) of subjects’ total IQ score was 108.5 (±8.9 SD)(Table 2).
Fig. 3. The grand average waveforms of the theta ERS and low–high-frequency alphaERD during the encoding and retrieval task. These waveforms refer to representativefrontal and parietal electrode sites.
Very large majority of the individual ERD/ERS peak values werestatistically significant (p < 0.05) for both encoding (21 out of 22subjects) and retrieval (20 out of 22 subjects) conditions.
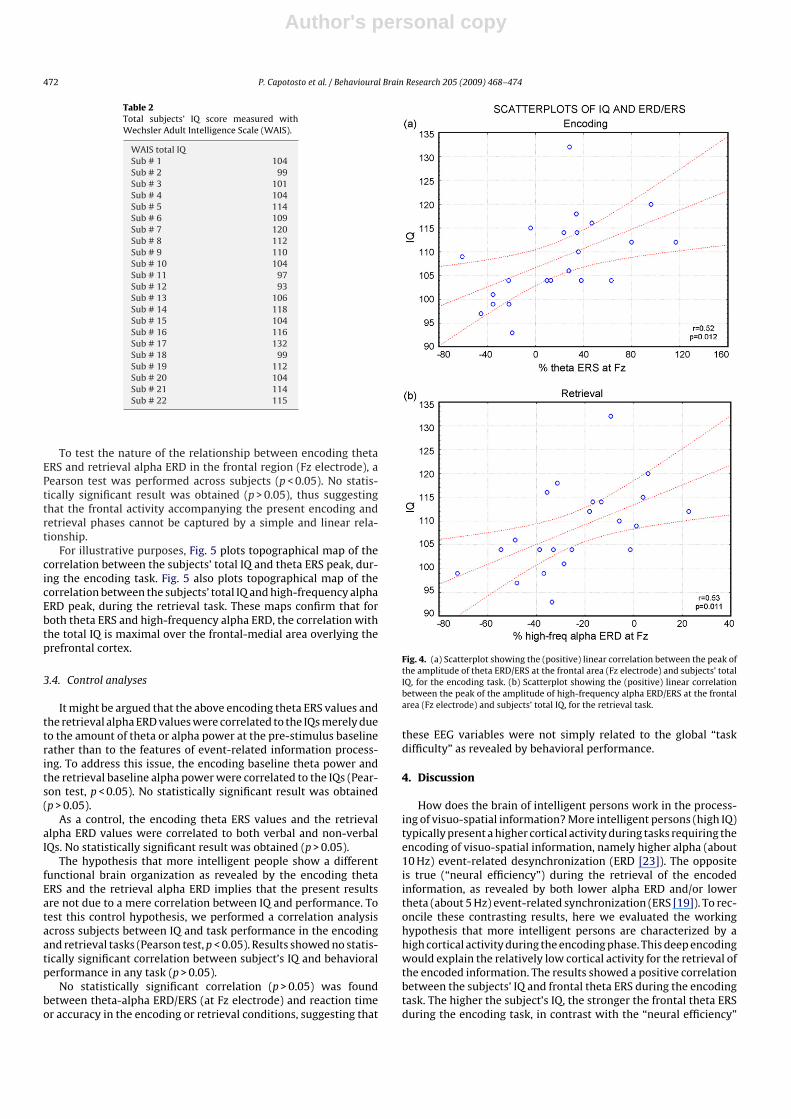
A statistically significant positive correlation was observedbetween the subjects’ total IQ and the frontal theta ERS peak (Fzelectrode), during the encoding task (r = 0.52; p = 0.012). The higherthe subject’s total IQ, the stronger the frontal theta ERS (Fig. 4a).Furthermore, there was a statistically significant positive correla-tion between the subjects’ total IQ and the frontal high-frequencyalpha ERD peak (Fz electrode), during the retrieval task (r = 0.53;p = 0.011). The lower the frontal high-frequency alpha ERD, thehigher the subject’s total IQ (Fig. 4b). No effect was observed onlow-frequency alpha ERD at the Fz electrode. Moreover, no effectwas observed at Pz electrode on theta and low and high-alpha fre-quencies.
Author's personal copy
472 P. Capotosto et al. / Behavioural Brain Research 205 (2009) 468–474
Table 2Total subjects’ IQ score measured withWechsler Adult Intelligence Scale (WAIS).
WAIS total IQSub # 1 104Sub # 2 99Sub # 3 101Sub # 4 104Sub # 5 114Sub # 6 109Sub # 7 120Sub # 8 112Sub # 9 110Sub # 10 104Sub # 11 97Sub # 12 93Sub # 13 106Sub # 14 118Sub # 15 104Sub # 16 116Sub # 17 132Sub # 18 99Sub # 19 112Sub # 20 104Sub # 21 114Sub # 22 115
To test the nature of the relationship between encoding thetaERS and retrieval alpha ERD in the frontal region (Fz electrode), aPearson test was performed across subjects (p < 0.05). No statis-tically significant result was obtained (p > 0.05), thus suggestingthat the frontal activity accompanying the present encoding andretrieval phases cannot be captured by a simple and linear rela-tionship.
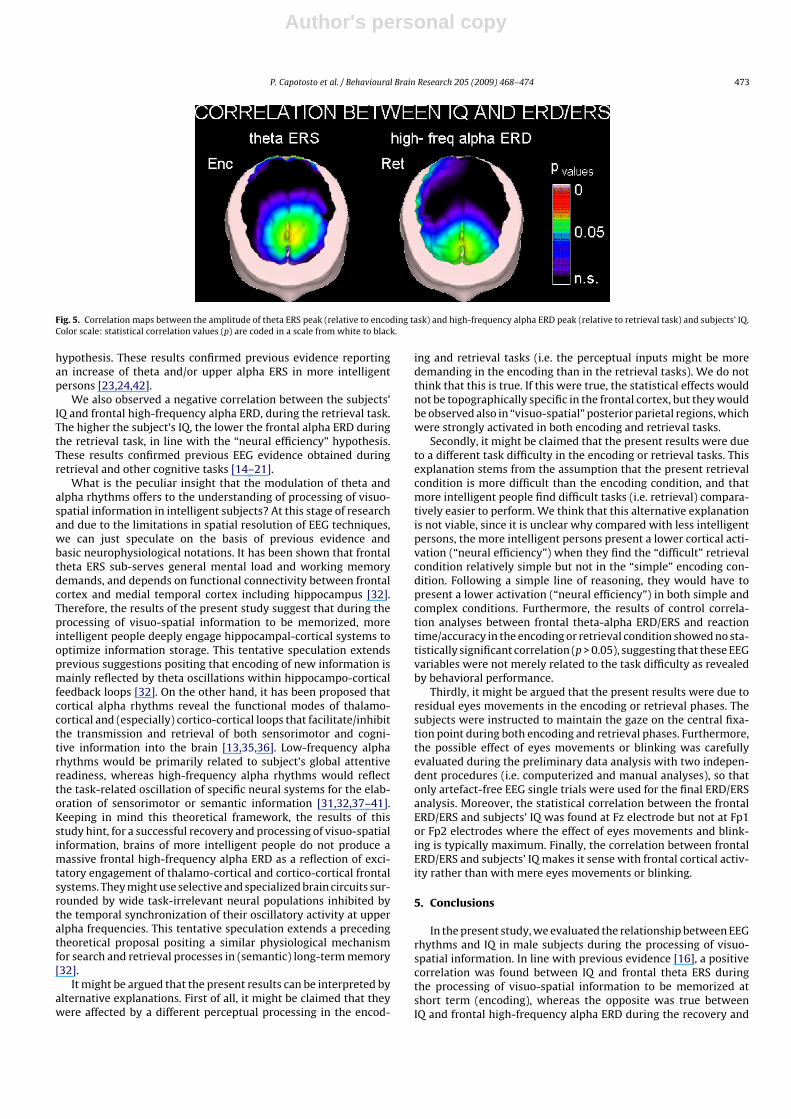
For illustrative purposes, Fig. 5 plots topographical map of thecorrelation between the subjects’ total IQ and theta ERS peak, dur-ing the encoding task. Fig. 5 also plots topographical map of thecorrelation between the subjects’ total IQ and high-frequency alphaERD peak, during the retrieval task. These maps confirm that forboth theta ERS and high-frequency alpha ERD, the correlation withthe total IQ is maximal over the frontal-medial area overlying theprefrontal cortex.
3.4. Control analyses
It might be argued that the above encoding theta ERS values andthe retrieval alpha ERD values were correlated to the IQs merely dueto the amount of theta or alpha power at the pre-stimulus baselinerather than to the features of event-related information process-ing. To address this issue, the encoding baseline theta power andthe retrieval baseline alpha power were correlated to the IQs (Pear-son test, p < 0.05). No statistically significant result was obtained(p > 0.05).
As a control, the encoding theta ERS values and the retrievalalpha ERD values were correlated to both verbal and non-verbalIQs. No statistically significant result was obtained (p > 0.05).
The hypothesis that more intelligent people show a differentfunctional brain organization as revealed by the encoding thetaERS and the retrieval alpha ERD implies that the present resultsare not due to a mere correlation between IQ and performance. Totest this control hypothesis, we performed a correlation analysisacross subjects between IQ and task performance in the encodingand retrieval tasks (Pearson test, p < 0.05). Results showed no statis-tically significant correlation between subject’s IQ and behavioralperformance in any task (p > 0.05).
No statistically significant correlation (p > 0.05) was foundbetween theta-alpha ERD/ERS (at Fz electrode) and reaction timeor accuracy in the encoding or retrieval conditions, suggesting that
Fig. 4. (a) Scatterplot showing the (positive) linear correlation between the peak ofthe amplitude of theta ERD/ERS at the frontal area (Fz electrode) and subjects’ totalIQ, for the encoding task. (b) Scatterplot showing the (positive) linear correlationbetween the peak of the amplitude of high-frequency alpha ERD/ERS at the frontalarea (Fz electrode) and subjects’ total IQ, for the retrieval task.
these EEG variables were not simply related to the global “taskdifficulty” as revealed by behavioral performance.
4. Discussion
How does the brain of intelligent persons work in the process-ing of visuo-spatial information? More intelligent persons (high IQ)typically present a higher cortical activity during tasks requiring theencoding of visuo-spatial information, namely higher alpha (about10 Hz) event-related desynchronization (ERD [23]). The oppositeis true (“neural efficiency”) during the retrieval of the encodedinformation, as revealed by both lower alpha ERD and/or lowertheta (about 5 Hz) event-related synchronization (ERS [19]). To rec-oncile these contrasting results, here we evaluated the workinghypothesis that more intelligent persons are characterized by ahigh cortical activity during the encoding phase. This deep encodingwould explain the relatively low cortical activity for the retrieval ofthe encoded information. The results showed a positive correlationbetween the subjects’ IQ and frontal theta ERS during the encodingtask. The higher the subject’s IQ, the stronger the frontal theta ERSduring the encoding task, in contrast with the “neural efficiency”
Author's personal copy
P. Capotosto et al. / Behavioural Brain Research 205 (2009) 468–474 473
Fig. 5. Correlation maps between the amplitude of theta ERS peak (relative to encoding task) and high-frequency alpha ERD peak (relative to retrieval task) and subjects’ IQ.Color scale: statistical correlation values (p) are coded in a scale from white to black.
hypothesis. These results confirmed previous evidence reportingan increase of theta and/or upper alpha ERS in more intelligentpersons [23,24,42].
We also observed a negative correlation between the subjects’IQ and frontal high-frequency alpha ERD, during the retrieval task.The higher the subject’s IQ, the lower the frontal alpha ERD duringthe retrieval task, in line with the “neural efficiency” hypothesis.These results confirmed previous EEG evidence obtained duringretrieval and other cognitive tasks [14–21].
What is the peculiar insight that the modulation of theta andalpha rhythms offers to the understanding of processing of visuo-spatial information in intelligent subjects? At this stage of researchand due to the limitations in spatial resolution of EEG techniques,we can just speculate on the basis of previous evidence andbasic neurophysiological notations. It has been shown that frontaltheta ERS sub-serves general mental load and working memorydemands, and depends on functional connectivity between frontalcortex and medial temporal cortex including hippocampus [32].Therefore, the results of the present study suggest that during theprocessing of visuo-spatial information to be memorized, moreintelligent people deeply engage hippocampal-cortical systems tooptimize information storage. This tentative speculation extendsprevious suggestions positing that encoding of new information ismainly reflected by theta oscillations within hippocampo-corticalfeedback loops [32]. On the other hand, it has been proposed thatcortical alpha rhythms reveal the functional modes of thalamo-cortical and (especially) cortico-cortical loops that facilitate/inhibitthe transmission and retrieval of both sensorimotor and cogni-tive information into the brain [13,35,36]. Low-frequency alpharhythms would be primarily related to subject’s global attentivereadiness, whereas high-frequency alpha rhythms would reflectthe task-related oscillation of specific neural systems for the elab-oration of sensorimotor or semantic information [31,32,37–41].Keeping in mind this theoretical framework, the results of thisstudy hint, for a successful recovery and processing of visuo-spatialinformation, brains of more intelligent people do not produce amassive frontal high-frequency alpha ERD as a reflection of exci-tatory engagement of thalamo-cortical and cortico-cortical frontalsystems. They might use selective and specialized brain circuits sur-rounded by wide task-irrelevant neural populations inhibited bythe temporal synchronization of their oscillatory activity at upperalpha frequencies. This tentative speculation extends a precedingtheoretical proposal positing a similar physiological mechanismfor search and retrieval processes in (semantic) long-term memory[32].
It might be argued that the present results can be interpreted byalternative explanations. First of all, it might be claimed that theywere affected by a different perceptual processing in the encod-
ing and retrieval tasks (i.e. the perceptual inputs might be moredemanding in the encoding than in the retrieval tasks). We do notthink that this is true. If this were true, the statistical effects wouldnot be topographically specific in the frontal cortex, but they wouldbe observed also in “visuo-spatial” posterior parietal regions, whichwere strongly activated in both encoding and retrieval tasks.
Secondly, it might be claimed that the present results were dueto a different task difficulty in the encoding or retrieval tasks. Thisexplanation stems from the assumption that the present retrievalcondition is more difficult than the encoding condition, and thatmore intelligent people find difficult tasks (i.e. retrieval) compara-tively easier to perform. We think that this alternative explanationis not viable, since it is unclear why compared with less intelligentpersons, the more intelligent persons present a lower cortical acti-vation (“neural efficiency”) when they find the “difficult” retrievalcondition relatively simple but not in the “simple” encoding con-dition. Following a simple line of reasoning, they would have topresent a lower activation (“neural efficiency”) in both simple andcomplex conditions. Furthermore, the results of control correla-tion analyses between frontal theta-alpha ERD/ERS and reactiontime/accuracy in the encoding or retrieval condition showed no sta-tistically significant correlation (p > 0.05), suggesting that these EEGvariables were not merely related to the task difficulty as revealedby behavioral performance.
Thirdly, it might be argued that the present results were due toresidual eyes movements in the encoding or retrieval phases. Thesubjects were instructed to maintain the gaze on the central fixa-tion point during both encoding and retrieval phases. Furthermore,the possible effect of eyes movements or blinking was carefullyevaluated during the preliminary data analysis with two indepen-dent procedures (i.e. computerized and manual analyses), so thatonly artefact-free EEG single trials were used for the final ERD/ERSanalysis. Moreover, the statistical correlation between the frontalERD/ERS and subjects’ IQ was found at Fz electrode but not at Fp1or Fp2 electrodes where the effect of eyes movements and blink-ing is typically maximum. Finally, the correlation between frontalERD/ERS and subjects’ IQ makes it sense with frontal cortical activ-ity rather than with mere eyes movements or blinking.
5. Conclusions
In the present study, we evaluated the relationship between EEGrhythms and IQ in male subjects during the processing of visuo-spatial information. In line with previous evidence [16], a positivecorrelation was found between IQ and frontal theta ERS duringthe processing of visuo-spatial information to be memorized atshort term (encoding), whereas the opposite was true betweenIQ and frontal high-frequency alpha ERD during the recovery and

Author's personal copy
474 P. Capotosto et al. / Behavioural Brain Research 205 (2009) 468–474
processing of the encoded visuo-spatial information (retrieval).These results reconcile previous contrasting evidence on “neuralefficiency” hypothesis [19,23,24]. They also suggest that more intel-ligent persons show high frontal cortical activity for the processingand short-term memorization of visuo-spatial information, and thismight allow the recovery and processing of the encoded informa-tion with a low cortical activity. Cortical activity of more intelligentmale persons would be better characterized by flexible and task-adapting features of frontal activation rather than by a fixed “neuralefficiency” operating mode.
References
[1] Haier RJ, Siegel BV, Nuechterlein KH, Hazlett E, Wu JC, Paek J, et al. Corticalglucose metabolicrate correlates of abstract reasoning and attention studiedwith positron emission tomography. Intelligence 1988;12:199–217.
[2] Haier RJ, Siegel Jr BV, MacLachlan A, Soderling E, Lottenberg S, Buchs-baum MS. Regional glucose metabolic changes after learning a complexvisuospatial/motor task: a positron emission tomographic study. Brain Res1992;570(January (1–2)):134–43.
[3] Haier RJ, Jung RE, Yeo RA, Head K, Alkire MT. Structural brain variation andgeneral intelligence. Neuroimage 2004;23(1):425–33.
[4] Charlot V, Tzourio N, Zilbovicius M, Mazoyer B, Denis M. Different mentalimagery abilities result in different regional cerebral blood flow activationpatterns during cognitive tasks. Neuropsychologia 1992;30(6):565–80.
[5] Parks RW, Loewenstein DA, Dodrill KL, Barker WW, Yoshii F, Chang JY, et al.Cerebral metabolic effects of a verbal fluency test: a PET scan study. J Clin ExpNeuropsychol 1988;10(5):565–75.
[6] Rypma B, D’Esposito M. The roles of prefrontal brain regions in components ofworking memory: effects of memory load and individual differences. Proc NatlAcad Sci USA 1999;96(May (11)):6558–63.
[7] Rypma B, Berger JS, D’Esposito M. The influence of working-memory demandand subject performance on prefrontal cortical activity. J Cogn Neurosci2002;14(5):721–31.
[8] Rypma B, Berger JS, Genova HM, Rebbechi D, D’Esposito M. Dissociating age-related changes in cognitive strategy and neural efficiency using event-relatedfMRI. Cortex 2005;41(4):582–94.
[9] Ruff CC, Knauff M, Fangmeier T, Spreer J. Reasoning and working memory: com-mon and distinct neuronal processes. Neuropsychologia 2003;41(9):1241–53.
[10] Neubauer AC, Fink A, Grabner RH. Sensitivity of alpha band ERD to individualdifferences in cognition. Prog Brain Res 2006;159:167–78 [review].
[11] Newman SD, Carpenter PA, Varma S, Just MA. Frontal and parietalparticipation in problem solving in the Tower of London: fMRI and compu-tational modeling of planning and high-level perception. Neuropsychologia2003;41(12):1668–82.
[12] Gray JR, Chabris CF, Braver TS. Neural mechanisms of general fluid intelligence.Nat Neurosci 2003;6(3):316–22.
[13] Pfurtscheller G, Lopes da Silva FH. Event-related EEG/MEG synchronization anddesynchronization: basic principles. Clin Neurophysiol 1999;110(11):1842–57[review].
[14] Neubauer AC, Freudenthaler HH, Pfurtscheller G. Intelligence and spatio-temporal patterns of event-related desynchronization. Intelligence1995;20:249–67.
[15] Neubauer AC, Sange G, Pfurtscheller G. Psychometric intelligence and event-related desynchronization during performance of a letter matching task. In:Pfurtscheller G, Lopes da Silva fh, editors. Handbook of electroencephalographyand clinical neurophysiology, vol. 6, revised series. Amsterdam: Elsevier; 1999.p. 219–31.
[16] Neubauer AC, Grabner RH, Freudenthaler HH, Beckmann JF, Guthke J. Intelli-gence and individual differences in becoming neurally efficient. Acta Psychol(Amst) 2004;116(1):55–74.
[17] Neubauer AC, Grabner RH, Fink A, Neuper C. Intelligence and neural efficiency:further evidence of the influence of task content and sex on the brain–IQ rela-tionship. Brain Res Cogn Brain Res 2005;25(1):217–25.
[18] Grabner RH, Stern E, Neubauer AC. When intelligence loses its impact:neural efficiency during reasoning in a familiar area. Int J Psychophysiol2003;49(2):89–98.
[19] Grabner RH, Fink A, Stipacek A, Neuper C, Neubauer AC. Intelligence and work-ing memory systems: evidence of neural efficiency in alpha band ERD. BrainRes Cogn Brain Res 2004;20(2):212–25.
[20] Neubauer AC, Fink A. Fluid intelligence and neural efficiency: effects of taskcomplexity and sex. Pers Ind Diff 2003;35:811–27.
[21] Jausovec N, Jausovec K. Spatiotemporal brain activity related to intelligence:a low resolution brain electromagnetic tomography study. Cogn Brain Res2003;16:267– 72.
[22] Grabner RH, Neubauer AC, Stern E. Superior performance and neural effi-ciency: the impact of intelligence and expertise. Brain Res Bull 2006;69(4):422–39.
[23] Doppelmayr M, Klimesch W, Sauseng P, Hodlmoser K, Stadler W,Hanslmayr S. Intelligence related differences in EEG-bandpower. Neurosci Lett2005;381(June (3)):309–13.
[24] Doppelmayr M, Klimesch W, Hödlmoser K, Sauseng P, Gruber W. Intelligencerelated upper alpha desynchronization in a semantic memory task. Brain ResBull 2005;66(July (2)):171–7.
[25] Wechsler D. The WAIS-R manual. New York: The Psychological Corporation;1981.
[26] Orsini A, Laicardi C. WAIS-R. Contributo alla taratura italiana. Firenze. Organiz-zazioni Speciali; 1997.
[27] Moretti DV, Babiloni F, Carducci F, Cincotti F, Remondini E, Rossini PM,et al. Computerized processing of EEG-EOG-EMG artifacts for multi-centricstudies in EEG oscillations and event-related potentials. Int J Psychophysiol2003;47(3):199–216.
[28] Babiloni F, Babiloni C, Carducci F, Fattorini L, Onorati P, Urbano A. SplineLaplacian estimate of EEG potentials over a realistic magnetic resonance-constructed scalp surface model. Electroencephalogr Clin Neurophysiol1996;98(4):363–73.
[29] Babiloni F, Carducci F, Babiloni C, Urbano A. Improved realistic Laplacianestimate of highly-sampled EEG potentials by regularization techniques. Elec-troencephalogr Clin Neurophysiol 1998;106(4):336–43.
[30] Nunez PL. Neocortical dynamics and human EEG rhythms. New York: OxfordUniversity Press; 1995.
[31] Klimesch W. Memory processes, brain oscillations and EEG synchronization.Int J Psychophysiol 1996;24(1–2):61–100 [review].
[32] Klimesch W. EEG alpha and theta oscillations reflect cognitive and memory per-formance: a review and analysis. Brain Res Brain Res Rev 1999;29(2–3):169–95(review).
[33] Pfurtscheller G, Neuper C. Event-related synchronization of mu rhythm in theEEG over the cortical hand area in man. Neurosci Lett 1994;174(June (1)):93–6.
[34] Babiloni F, Babiloni C, Fattorini L, Carducci F, Onorati P, Urbano A. Performancesof surface Laplacian estimators: a study of simulated and real scalp potentialdistributions. Brain Topogr 1995;8(1):35–45.
[35] Steriade M, Llinas RR. The functional states of the thalamus and the associatedneuronal interplay. Physiol Rev 1988;68(3):649–742 (review).
[36] Brunia CH. Neural aspects of anticipatory behavior. Acta Psychol (Amst)1999;101(2–3):213–42 (review).
[37] Klimesch W, Doppelmayr M, Russegger H, Pachinger T, Schwaiger J. Inducedalpha band power changes in the human EEG and attention. Neurosci Lett1998;244(2):73–6.
[38] Babiloni C, Babiloni F, Carducci F, Cincotti F, Rosciarelli F, Rossini PM, Arendt-Nielsen L, Chen ACN. Mapping of early and late human somatosensory evokedbrain potentials to phasic galvanic painful stimulation. Human Brain Mapping2001;12(3):168–79.
[39] Babiloni C, Miniussi C, Babiloni F, Carducci F, Cincotti F, Del Percio C, SirelloG, Fracassi C, Nobre AC, Rossini PM. Sub-second “temporal attention” modu-lates alpha rhythms. A high-resolution EEG study. Brain Res Cogn Brain Res2004;19(3):259–68.
[40] Pfurtscheller G, Aranibar A. Evaluation of event-related desynchronization(ERD) preceding and following voluntary self-paced movement. Electroen-cephalogr Clin Neurophysiol 1979;46(2):138–46.
[41] Pfurtscheller G, Neuper C, Flotzinger D, Pregenzer M. EEG-based discriminationbetween imagination of right and left hand movement. Electroencephalogr ClinNeurophysiol 1997;103(6):642–51.
[42] Vogt F, Klimesch W, Doppelmayr M. High-frequency components in thealpha band and memory performance. J Clin Neurophysiol 1998;15(2):167–72.




















