
Phylogenetics of sees plants: An analysis of nucleotide sequences from the plastid gene rbcL
Upload
jamescookuniversityCategory
view
4download
0
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_522File # 08TQ
522
Systematic Botany (2002), 27(3): pp. 522–533q Copyright 2002 by the American Society of Plant Taxonomists
Is the Genus Cissus (Vitaceae) Monophyletic? Evidence from Plastid andNuclear Ribosomal DNA
MAURIZIO ROSSETTO,1,3 BETSY R. JACKES,2 KIRSTEN D. SCOTT,1 and ROBERT J. HENRY1
1Centre for Plant Conservation Genetics, Southern Cross University, PO Box 157 Lismore,New South Wales 2480 Australia;
2Tropical Plant Sciences, James Cook University, Townsville, Queensland 4811 Australia3Author for correspondence ([email protected])
Communicating Editor: Aaron Liston
ABSTRACT. The trnL (UAA) intron and ITS1 region were sequenced to investigate relationships among the five generaof Vitaceae present in Australia relative to Vitis. Congruent results were obtained between separate and combined data sets,with all major clades being shared among trees. All bootstrap consensus trees obtained from single sequences or combinedanalysis strongly suggest that Cissus is polyphyletic, corroborating the morphological inconsistencies reported previously.Cissus opaca and Clematicissus angustissima consistently grouped in a common clade. A further four taxa (C. antarctica, C.hypoglauca, C. oblonga, and C. sterculiifolia) also grouped within a clade disjunct from the main Cissus clade. Our resultssuggest that these five species currently classified as Cissus should be segregated from the genus. Of further interest is theclose relationship between Cayratia and Tetrastigma. Overall, the results presented provide new insights into the relationshipswithin a number of Vitaceae genera and suggest directions for future studies.
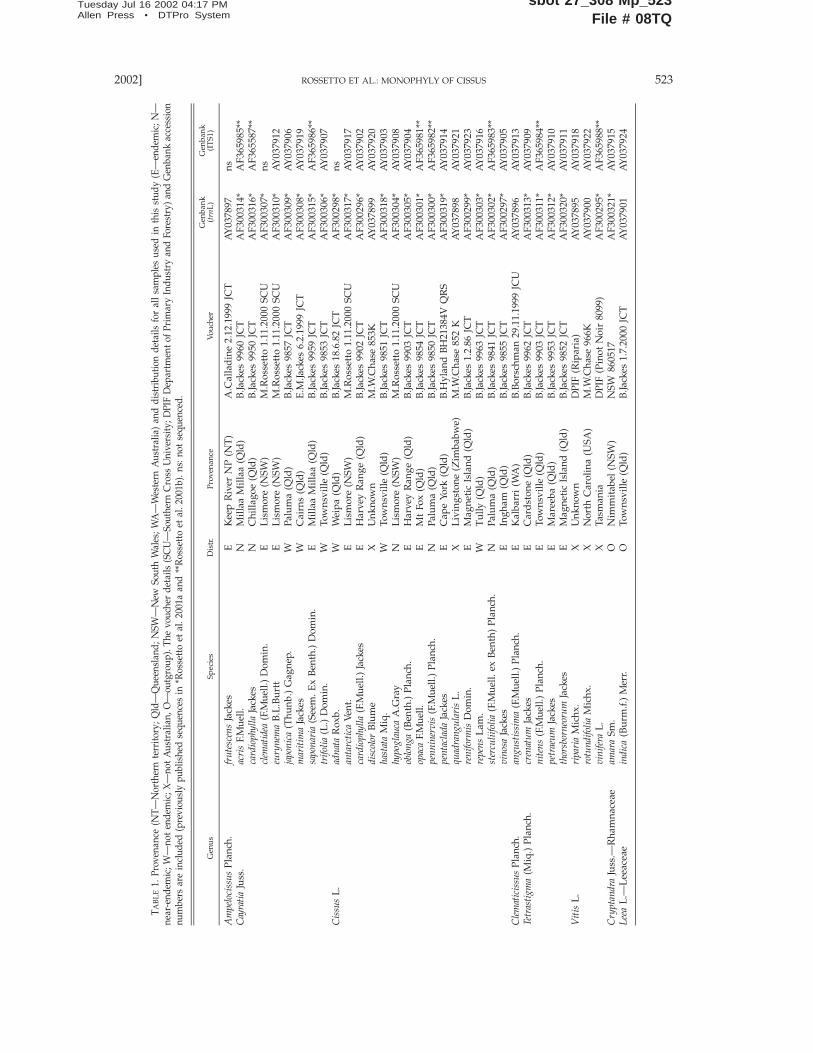
Vitaceae are woody climbers comprising 13 generaand about 700 species worldwide (Mabberley 1995).The family is characterized by leaf-opposed tendrils,which may be modified to form an inflorescence. Thesmall regular flowers are bisexual in most genera, andthe size and shape of the leaves can be variable evenwithin a single individual. The fruit is a berry withone to four seeds; it can be edible but is usually as-tringent. Vitis, Ampelopsis Michx. and ParthenocissusPlanch. are mostly restricted to temperate areas. Theremaining genera have predominantly tropical andsubtropical distributions mainly from Africa throughAsia and into the Pacific islands. In these latter regions,Vitaceae often play an important role in rainforest hab-itats and vine thickets. Originally, only Vitis was rec-ognized within Australia by Bentham (1863), but afterlater revisions, this genus is now considered to be ab-sent. Five other genera are represented in the Austra-lian flora with a total of 30 named species. These areAmpelocissus (three species, Jackes 1984), Cayratia (eightspecies, Jackes 1987), Cissus (14 species, Jackes 1988),Clematicissus (a monotypic genus, Jackes 1989a) and Te-trastigma (four species, Jackes 1989b). A total of 17 spe-cies are endemic and a further six are limited to Aus-tralia and New Guinea (Table 1).
Despite recent revisions, some uncertainty still ex-ists about the classification of Australian Vitaceae, par-ticularly Cissus for which remarkable morphologicaldiversity has been recorded. This is not surprising be-cause generic limits have been problematic within thefamily. In 1810, Brown combined what were the onlythree known genera, Vitis, Ampelopsis, and Cissus, intothe genus Vitis. By the end of the 19th century, Plan-chon (1887) resurrected these three genera and de-scribed a further six. Within Cissus, the most species-
rich genus, Planchon (1887) recognized three subdivi-sions. These subdivisions are now recognized as thegenera Cissus, Cyphostemma (Planch.) Alston, and Cayr-atia (Gagnepain 1911). The distinguishing characters,described in detail by Descoings (1960), are mainlyseed morphology and the nature of the endosperm.Generally Cissus is characterized by simple leaves andone-seeded fruits, Cyphostemma by compound leavesand one-seeded fruits, and Cayratia by compoundleaves and multiple-seeded fruits. The number ofnamed Cissus species varies according to the research-er, from 200 (Mabberley 1995), to 250 (Lavie 1990), toca. 350 (Galet 1967). The corresponding figures for theAfrican genus Cyphostemma are 150, 250, and ca. 150respectively. Such uncertainty is mainly due to the factthat within Cissus there are species with compoundleaves and more than one seed per fruit. For instance,within the Australian species of Cissus, Jackes (1988)identified three groups based on distinct morphologi-cal features, but decided to adopt a conservative ap-proach to taxonomic changes until relationships couldbe further evaluated.
The comparative analysis of homologous DNA se-quences represents an increasingly useful and recog-nized approach for examining taxonomic relationships.Molecular data provide new information for phyloge-netic investigations and consequently often clarifybroad relationships among taxa. Numerous phyloge-netic studies have been based on plastid DNA sequenc-es, with the photosynthesis-related rbcL being one ofthe most widely used genes to infer plant phylogenies(Chase et al. 1993, Soltis et al. 1999). However the num-ber of variable sites for this conserved gene is generallynot sufficient for defining relationships at low taxo-nomic levels and non-coding regions of plastid DNA
2002] 523ROSSETTO ET AL.: MONOPHYLY OF CISSUS
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_523File # 08TQ
TAB
LE
1.Pr
oven
ance
(NT—
Nor
ther
nte
rrito
ry;Q
ld—
Que
ensl
and;
NSW
—N
ewSo
uth
Wal
es;W
A—
Wes
tern
Aus
tral
ia)
and
dist
ribu
tion
deta
ilsfo
ral
lsam
ples
used
inth
isst
udy
(E—
ende
mic
;N—
near
-end
emic
;W—
not
ende
mic
;X—
notA
ustr
alia
n,O
—ou
tgro
up).
The
vouc
her
deta
ils(S
CU
—So
uthe
rnC
ross
Uni
vers
ity;D
PIF
Dep
artm
ento
fPri
mar
yIn
dust
ryan
dFo
rest
ry)a
ndG
enba
nkac
cess
ion
num
bers
are
incl
uded
(pre
viou
sly
publ
ishe
dse
quen
ces
in*R
osse
tto
etal
.200
1aan
d**
Ros
sett
oet
al.2
001b
).ns
:not
sequ
ence
d.
Gen
usSp
ecie
sD
istr
.Pr
oven
ance
Vouc
her
Gen
bank
(trn
L)
Gen
bank
(IT
S1)
Am
pelo
ciss
usPl
anch
.C
ayra
tia
Juss
.fr
utes
cens
Jack
esac
ris
F.M
uell.
card
ioph
ylla
Jack
escl
emat
idea
(F.M
uell.
)D
omin
.eu
ryne
ma
B.L
.Bur
tt
E N N E E
Kee
pR
iver
NP
(NT
)M
illaa
Mill
aa(Q
ld)
Chi
llago
e(Q
ld)
Lis
mor
e(N
SW)
Lis
mor
e(N
SW)
A.C
alla
din
e2.
12.1
999
JCT
B.Ja
ckes
9960
JCT
B.Ja
ckes
9950
JCT
M.R
osse
tto
1.11
.200
0SC
UM
.Ros
sett
o1.
11.2
000
SCU
AY
0378
97A
F300
314*
AF3
0031
6*A
F300
307*
AF3
0031
0*
ns AF3
6598
5**
AF3
6558
7**
ns AY
0379
12
Cis
sus
L.
japo
nica
(Thu
nb.)
Gag
nep.
mar
itim
aJa
ckes
sapo
nari
a(S
eem
.Ex
Ben
th.)
Dom
in.
trifo
lia(L
.)D
omin
.ad
nata
Rox
b.
W W E W W
Palu
ma
(Qld
)C
airn
s(Q
ld)
Mill
aaM
illaa
(Qld
)To
wns
ville
(Qld
)W
eipa
(Qld
)
B.Ja
ckes
9857
JCT
E.M
.Jack
es6.
2.19
99JC
TB
.Jack
es99
59JC
TB
.Jack
es98
53JC
TB
.Jack
es18
.6.8
2JC
T
AF3
0030
9*A
F300
308*
AF3
0031
5*A
F300
306*
AF3
0029
8*
AY
0379
06A
Y03
7919
AF3
6598
6**
AY
0379
07ns
anta
rcti
caVe
nt.
card
ioph
ylla
(F.M
uell.
)Ja
ckes
disc
olor
Blu
me
hast
ata
Miq
.hy
pogl
auca
A.G
ray
E E X W N
Lis
mor
e(N
SW)
Har
vey
Ran
ge(Q
ld)
Unk
now
nTo
wns
ville
(Qld
)L
ism
ore
(NSW
)
M.R
osse
tto
1.11
.200
0SC
UB
.Jack
es99
02JC
TM
.W.C
hase
853K
B.Ja
ckes
9851
JCT
M.R
osse
tto
1.11
.200
0SC
U
AF3
0031
7*A
F300
296*
AY
0378
99A
F300
318*
AF3
0030
4*
AY
0379
17A
Y03
7902
AY
0379
20A
Y03
7903
AY
0379
08ob
long
a(B
enth
.)Pl
anch
.op
aca
F.M
uell.
penn
iner
vis
(F.M
uell.
)Pl
anch
.pe
ntac
lada
Jack
esqu
adra
ngul
aris
L.
E E N E X
Har
vey
Ran
ge(Q
ld)
Mt
Fox
(Qld
)Pa
lum
a(Q
ld)
Cap
eYo
rk(Q
ld)
Liv
ings
tone
(Zim
babw
e)
B.Ja
ckes
9903
JCT
B.Ja
ckes
9854
JCT
B.Ja
ckes
9850
JCT
B.H
ylan
dBH
2138
4VQ
RS
M.W
.Cha
se85
2K
AF3
0030
5*A
F300
301*
AF3
0030
0*A
F300
319*
AY
0378
98
AY
0379
04A
F365
981*
*A
F365
982*
*A
Y03
7914
AY
0379
21
Cle
mat
icis
sus
Plan
ch.
reni
form
isD
omin
.re
pens
Lam
.st
ercu
liifo
lia(F
.Mue
ll.ex
Ben
th)
Plan
ch.
vino
saJa
ckes
angu
stis
sim
a(F
.Mue
ll.)
Plan
ch.
E W N E E
Mag
netic
Isla
nd(Q
ld)
Tul
ly(Q
ld)
Palu
ma
(Qld
)In
gham
(Qld
)K
alba
rri
(WA
)
B.Ja
ckes
1.2.
86JC
TB
.Jack
es99
63JC
TB
.Jack
es98
41JC
TB
.Jack
es98
55JC
TB
.Bor
schm
an29
.11.
1999
JCU
AF3
0029
9*A
F300
303*
AF3
0030
2*A
F300
297*
AY
0378
96
AY
0379
23A
Y03
7916
AF3
6598
3**
AY
0379
05A
Y03
7913
Tetr
asti
gma
(Miq
.)Pl
anch
.
Vit
isL
.
cren
atum
Jack
esni
tens
(F.M
uell.
)Pl
anch
.pe
trae
umJa
ckes
thor
sbor
neor
umJa
ckes
ripa
ria
Mic
hx.
E E E E X
Car
dst
one
(Qld
)To
wns
ville
(Qld
)M
aree
ba(Q
ld)
Mag
netic
Isla
nd(Q
ld)
Unk
now
n
B.Ja
ckes
9962
JCT
B.Ja
ckes
9903
JCT
B.Ja
ckes
9953
JCT
B.Ja
ckes
9852
JCT
DPI
F(R
ipar
ia)
AF3
0031
3*A
F300
311*
AF3
0031
2*A
F300
320*
AY
0378
95
AY
0379
09A
F365
984*
*A
Y03
7910
AY
0379
11A
Y03
7918
Cry
ptan
dra
Juss
.—R
ham
nace
aeLe
eaL
.—L
eeac
eae
rotu
ndifo
liaM
ichx
.vi
nife
raL
.am
ara
Sm.
indi
ca(B
urm
.f.)
Mer
r.
X X O O
Nor
thC
arol
ina
(USA
)Ta
sman
iaN
imm
itab
el(N
SW)
Tow
nsvi
lle(Q
ld)
M.W
.Cha
se96
6KD
PIF
(Pin
otN
oir
8099
)N
SW86
0517
B.Ja
ckes
1.7.
2000
JCT
AY
0379
00A
F300
295*
AF3
0032
1*A
Y03
7901
AY
0379
22A
F365
988*
*A
Y03
7915
AY
0379
24

524 [Volume 27SYSTEMATIC BOTANY
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_524File # 08TQ
have been found to be more informative (Gielly andTaberlet 1994). Taberlet et al. (1991) developed a seriesof universal primers for a number of these non-codingplastid regions. The trnL (UAA) intron has been suc-cessfully used as a phylogenetic tool at the inter andintra-generic levels (van Ham et al. 1994; Gielly andTaberlet 1996; Hopper et al. 1999; McDade and Moody1999). A preliminary investigation of the Australian Vi-taceae detected sufficient trnL (UAA) variability withinthis group (Rossetto et al. 2001a). As plastid DNA of-ten accumulates nucleotide substitutions more slowlythan nuclear DNA (especially when recently divergedspecies are being assessed; Small et al. 1998), phylo-genetic relationships can be inferred more readily bycomparative studies of the nucleotide sequences of ri-bosomal RNA (rRNA) genes (rDNA; Hillis and Dixon1991). Because of their faster rate of evolution (Whiteet al. 1990) a much finer intergeneric definition can beobtained by sequencing the internal transcribed spacer(ITS) region of 18S–26S nuclear ribosomal DNA(nrDNA). The two spacers, ITS1 and ITS2, can be read-ily sequenced using universal primers and have prov-en to be a good source of variable characters for lowerlevel phylogenetic studies. Informativeness is similarbetween ITS1 and ITS2 with the former being gener-ally slightly longer (Baldwin et al. 1995). Such regionsare particularly useful to complement data from usu-ally slower evolving plastid DNA regions and providedata from regions with different modes of inheritance.
At the family level, molecular studies have yet todefine the exact position of Vitaceae within the angio-sperm phylogeny. Based on morphological characters,the Vitaceae were traditionally placed in the Rham-nales (Cronquist 1988) but recent molecular work hasgenerally challenged such placement. An early rbcL-based study placed Vitaceae and Dilleniaceae as sisterfamilies within a poorly supported clade (Chase et al.1993). Sequencing of the same gene for a greater num-ber of taxa representing the majority of the eudicotfamilies (589 rbcL sequences representing 308 families)placed Vitaceae and Dilleniaceae as sister to the Car-yophyllales but still without bootstrap support (Savo-lainen et al. 2000). Further plastid and nuclear ribo-somal genes (atpB, rbcL, 18S) were added to the anal-ysis by Soltis et al. (2000) for a total of 567 taxa. Theinformation produced by this combined data set pro-vided greater resolution but still no well-supported an-swer. Dilleniaceae remained as the sister group of Car-yophyllales, well removed from Vitaceae, which werea sister group to the rosids (Soltis et al. 2000). As aresult, Vitaceae are still part of a small list of angio-sperm families for which a conclusive placement is yetto be established.
Because of the difficulty in identifying unequivocalmorphological synapomorphies within Vitaceae (par-ticularly within Cissus), this study aimed to use se-
quence information to resolve relationships across a se-lection of taxa. A number of issues that could not beclarified by the analysis of morphological characteris-tics alone gave rise to two main questions. Is the genusCissus monophyletic, and if not, what generic realign-ments are suggested? What is the relationship amongthe Australian genera of Vitaceae? The aim of thisstudy was to answer these and other questions by in-vestigating selected regions of the plastid, trnL intron,and nuclear genome, ITS1.
MATERIALS AND METHODS
Plant Material and DNA Extraction. The majority of Austra-lian Vitaceae were sampled (Table 1) except for Ampelocissus acetosa(F.Muell.) Planch., A. gardineri (F.M.Bailey) Jackes, and Cissus aris-tata Blume, for which material could not be obtained. Samples ofthe Australian Ampelocissus species were particularly difficult toobtain, and herbarium material consistently failed to yield qualityDNA. In some cases, the small amount and poor quality of her-barium material available resulted in successful amplification ofonly one region. As a result, the ITS1 sequence for A. frutescens,Cayratia clematidea, and Cissus adnata could not be obtained. Twonon-Australian Cissus species, Cissus quadrangularis and C. discolor,were investigated to provide within-genus comparison. Interge-neric relationships with Vitis were investigated by including Vitisvinifera (Pinot Noir), V. riparia, and V. rotundifolia in the analysis.As an unequivocal sister group is yet to be identified for the Vi-taceae, potential rooting was evaluated by comparing sequencesfrom various species and using the easiest to align. ConsequentlyLeea indica (Leeaceae—sometimes included in Vitaceae; Soltis et al.2000) and Cryptandra amara (Rhamnaceae) were selected as out-groups. Sampling details and distribution range of each speciesare listed in Table 1.
A DNA extraction method previously described in Rossetto etal. (2001a) was used on fresh leaf tissue or dried herbarium ma-terial to obtain DNA. Other extraction methods were not as suc-cessful across a sample of the species tested in a preliminarystudy.
trnL and ITS1 Amplification. A preliminary evaluation usingsamples from each genus showed that the trnL (UAA) intron re-gion could be amplified by PCR more efficiently and consistently(even with very poor quality material) than the intergenic spacerbetween trnL (UAA) 39 exon and trnF (GAA). Furthermore, thetrnL (UAA) intron PCR fragments exhibited size divergence be-tween these taxa. PCR amplification of the trnL region was per-formed from total DNA using primers c and d of Taberlet et al.(1991). PCR was performed in a total volume of 25 ml, containingPCR buffer (10 mM Tris-HCl pH 8.3, 50 mM KCl, 001% w/v gel-atin), 25 mM MgCl2, 1.5 units Taq (Roche), 0.2 mM of each dNTP,0.25 mM of each primer, 5 ml of betaine, 20 ng of template DNAand DNA-free water. The addition of betaine was particularly use-ful for those DNA samples that had been more difficult to extractand amplify. A Perkin Elmer 9700 thermocycler was used for aPCR protocol that included 35 cycles at 948C for 20 sec, 608C for20 sec, and 728C for 90 sec.
Primers that make use of conserved regions in the 18S and 5.8Scoding regions of the rRNA cistron have been developed for theamplification of the non-coding ITS1 region (White et al. 1990). Inthis study, PCR amplification of ITS1 was performed using theforward primers ITS5, GN1 and the reverse primers ITS2, C1(White et al. 1990, Scott and Playford 1996). In a small number ofcases in which the above-mentioned primers failed to produceclear PCR products, an additional primer was successfully used(59 CCT GCG GAA GGA TCA TTG 39). PCR was performed in atotal volume of 25 ml. The PCR protocol used included 35 cyclesat 948C for 20 sec, 50 to 558C for 20 sec (depending on amplifi-cation success) and 728C for 90 sec. The ITS1 region was preferred

2002] 525ROSSETTO ET AL.: MONOPHYLY OF CISSUS
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_525File # 08TQ
TABLE 2. Summary of the molecular data and their analysis. The data reported for the combined analysis, does not always representthe summation of the two following columns as different numbers of taxa were used.
Comparison Combined trnL (UAA) ITS1
Number of taxaTotal charactersFragment size range (bp)Variable charactersInformative substitutions
31939—
422289
34601
478/53714860
31338
197/291278217
Informative indelsMPTsMPT lengthConsistency IndexRetention IndexAverage GC content
152
10970.590.67
—
157407198
0.820.87
33.5%
02
8720.560.69
64.7%
to the ITS2 region as it consistently produced better amplificationproducts.
Sequencing, Alignment and Data Analysis. Cycle-sequencingwas carried out using the Big-Dye terminator (Perkin Elmer) fol-lowing Perkin-Elmer’s protocols on an ABI 3700 thermocycler. Se-quences were visualised on an ABI 377 sequencer at the AustralianGenome Research Facility (Brisbane, Australia). Sequencing withthe forward primer produced sufficiently reliable sequences for theentire product, but the reverse strand was also sequenced for mostPCR products as a control. Alignment of the trnL intron and ITS1sequences was performed using Clustal W (Thompson et al. 1994)with slight manual modifications necessary to optimise the mul-tiple alignments. Sequences were submitted to GenBank and theirrespective accession numbers are listed in Table 1.
The sequences obtained were evaluated by maximum parsimo-ny. The two regions were analysed separately and as a combineddata set. Information on parsimony informative indels (insertion/deletions shared by two or more taxa) was added to the sequencedata as a presence/absence matrix, which carried equal weight assingle nucleotide substitutions. A total of 15 indels were selectedfor trnL (UAA) but, because of the variability of the region, noinformative indels could be used for ITS1. Phylogenetic relation-ships among the 34 species for trnL (UAA) and the 31 species forITS1 and the combined analysis were inferred with 1000 randomsequence additions and tree bisection-reconnection (TBR) branchswapping, permitting five trees to be held at each step thenswapped to completion. Tree searches were conducted via Fitchparsimony (equal weight unordered character states, Fitch 1971)using PAUP* 4.0b4a (Swofford 1998). Clade robustness was mea-sured by the bootstrap method (Felsenstein 1985) with 1000 rep-licates, and using 1000 random sequence addition and tree bisec-tion-reconnection (TBR) branch swapping, but again permittingonly five trees to be held at each step.
Kishino-Hasegawa and non-parametric tests (PAUP*, Swofford1998) were used to assess the significance of the differences inplastid and nrDNA tree topologies and length. Various constraintsrepresenting a number of species pairs (listed in the results) wereimposed on the ITS1 and trnL data to assess if the tree topologywas significantly different from the most parsimonious tree. Re-cently the validity of the Kishino-Hasegawa test for most popularapplications has been questioned (Goldman et al. 2000). In thisstudy the tests performed were selected to offer the potential tospecifically identify the source of conflict between trees.
RESULTS
Comparison Between Sequences. The trnL (UAA)aligned matrix was 601 characters long and includeda total of 75 informative characters, including the in-dels (Table 2). The plastid DNA fragment ranged from478 bp (Tetrastigma thorsborneorum) to 537 bp (Cissusadnata). Sequencing of the trnL (UAA) intron success-
fully distinguished the majority of the taxa studied,with the exception of three species pairs: Cayratia acrisand C. saponaria, T. nitens and T. petraeum, Vitis viniferaand V. riparia. The trnL (UAA) intron produced a totalof 7407 MPTs, 198 steps long. The strict consensus treecomprises three major bootstrap-supported clades di-vided into two sister groups (Fig. 1a). Clade A, con-taining Cissus opaca and Clematicissus angustissima, isstrongly supported and distinct from the rest of theAustralian Vitaceae analysed. Its sister group containstwo major clades and a polytomy (Fig. 1a). Within thismajor group, clade B includes three subclades contain-ing respectively four Cayratia, all the Tetrastigma, andthe remaining Cayratia species as well as the single spe-cied of Ampelocissus analysed. Clade E contains a num-ber of Australian and the two non-Australian Cissustested. The remaining Cissus and Vitis species are notresolved within distinct, supported clades (Fig. 1a).
Amplification products could not be obtained for alltaxa with the ITS1 primers and as a result only 31species could be analysed. The aligned ITS1 matrixwas 338 characters long (Table 2) with fragments rang-ing from 197 bp (Cissus quadrangularis) to 291 bp (T.thorsborneorum). Towards the end of ITS1 a region ofapproximately 20 bp was more variable in length,making unambiguous alignment impossible becauseof independent and overlapping length mutations thatoccurred in different taxa. As a result, the region waseliminated from the final analysis. ITS1 successfullydistinguished between all samples, but the differencebetween Cayratia acris and C. saponaria was a singledeletion and the distinction between Tetrastigma nitensand T. petraeum was limited to two transitions. A totalof 217 parsimony informative characters produced twomost parsimonious trees 872 steps long (CI50.56 andRI50.69; Fig. 1b). The strict consensus tree producedby the ITS1 data is illustrated in Fig. 1b. The threemajor clades present within the plastid DNA tree arepreserved with some minor inconsistency (i.e. Cissusrepens is included in clade E and no distinct Tetrastigmasubclade was obtained—Fig. 1b). Furthermore, due to

526 [Volume 27SYSTEMATIC BOTANY
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_526File # 08TQ
FIG. 1. Strict consensus trees based on the plastid and nrDNA matrices. Bootstrap percentages (.50%) are listed above thebranches. The letters representing the major clades (as discussed within the text) are indicated on the side of the figure. a) trnL(UAA) intron, 198 steps long, CI 5 0.81, RI 5 0.89 b) ITS1, 872 steps long, CI 5 0.56, RI 5 0.69.

2002] 527ROSSETTO ET AL.: MONOPHYLY OF CISSUS
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_527File # 08TQ
FIG. 1. Continued.

528 [Volume 27SYSTEMATIC BOTANY
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_528File # 08TQ
the greater number of variable sites within ITS1, theremainder of the taxa analyzed are also included with-in smaller but distinct clades. Vitis vinifera and V. ri-paria are included within clade D and the four remain-ing Cissus species (C. antarctica, C. hypoglauca, C. oblon-ga, and C. sterculiifolia) are grouped within clade C(Fig. 1b).
Combined Data. The combined aligned matrix was939 characters long including a total of 304 informativecharacters. The Fitch analysis resulted in two mostparsimonious trees 1097 steps long (Fig. 2). The strictconsensus tree obtained with the combined data setwas similar to those produced by analyzing the dataseparately (Fig. 1a and 1b). Three major and two minorclades were distinguished. Clade A has high bootstrapsupport and unequivocally groups Cissus opaca and theonly currently recognized Clematicissus species. Thesecond major clade (clade B) includes all Cayratia andTetrastigma and is also supported by a high bootstrappercentage. Within clade B two well supported subcla-des are found, one containing all the Tetrastigma andtwo Cayratia species, the other containing three Cayra-tia. Clade E (100% bootstrap) comprises nine Cissusspecies, including the two non-Australian species test-ed. Finally, the last two clades include Cissus antarctica,C. hypoglauca, C. oblonga, C. sterculiifolia, and V. rotun-difolia (clade C2 55% bootstrap) and the two remain-ing Vitis species (clade D2 71% bootstrap).
Minor differences between the cpDNA and nrDNAtrees were detected (Fig. 1). In particular, inconsisten-cies between a number of species pairings were notedamong the data sets. For instance, four species pairs(Cayratia maritima and C. trifolia; Cissus discolor and C.hastata; C. quadrangularis and C. reniformis; T. nitens,and T. petraeum) were highly supported by plastidDNA analysis (Fig. 1a) but were not observed on theITS1 tree (Fig. 1b). Similarly, three species pairs (Cissuscardiophylla and C. penninervis, C. reniformis and C. re-pens, C. antarctica and C. oblonga) were supported bynuclear DNA but not by plastid DNA analysis. Para-metric and non-parametric pair-wise tests (PAUP*,Swofford 1998) suggested that the difference betweentrnL and ITS1 tree topologies was significant(P,0.001). However, most of the conflict was likelycaused by contrasting data across one species pair,Cayratia trifolia and C. maritima, as imposing this to-pology constraint on the ITS data resulted in highlysignificant differences with the most parsimonioustrees (P,0.001). Some conflict was also caused by im-posing the Cissus reniformis/C. repens pair as a con-straint on the trnL data (P,0.05). In relation to themain scope of this study, this incongruence was notconsidered as sufficient to prevent the combination ofthe data.
DISCUSSION
Congruence Between the Two DNA Regions. Thesize range of the trnL (UAA) intron in the AustralianVitaceae was within the range described by Gielly andTaberlet (1994) for 16 very diverse taxa. Previous re-ports suggested that the trnL (UAA) intron had aslightly lower resolution than other non-coding plastidDNA regions (Gielly and Taberlet 1994, 1996). McDadeand Moody (1999) found that in Acanthaceae the spac-er region between trnL (UAA) 39 and trnF (GAA) of-fered more parsimony informative variation than thetrnL (UAA) intron. Nevertheless, these authors foundthat data from both regions were remarkably congru-ent despite slight differences in resolution. Within Vi-taceae, sequencing of trnL produced a number of var-iable sites sufficient for distinguishing between mostof the taxa analyzed. ITS1 was also useful and madean even greater contribution to the study by producingover 70% of all characters used.
In our study the results from the two separate ma-trices were found to be mostly congruent with thecombined data and all major clades were present ineach tree. Some discrepancies were found, with a num-ber of species pairs supported by plastid but not nu-clear DNA and vice-versa (Fig. 1). In particular, im-posing the Cayratia trifolia/C. maritima pair with theITS data produced a significantly less parsimonious re-construction. Thus while the more basal nodes of thetrees were in agreement, some incongruence wasfound within the major clades. DNA transfer attribut-able to introgressive hybridization between relatedspecies has been suggested as a possible explanationfor similar inconsistencies (Cros et al. 1998). This op-tion cannot be excluded for these two species as theyare morphologically very similar and sympatric (Jackes1987). Coalescence failure and other types of noise inthe ITS data (including allelic polymorphism due torecent hybridization events) are other mechanisms thatcould complicate reconstructions between close rela-tives. Slowinsky and Page (1999) suggest that combin-ing genes with different evolutionary history and num-ber of nucleotides can potentially produce trees thatare different from the true species trees. However, themain focus of this study was to define broad relation-ships between groups of taxa (which were respectedin all trees obtained) rather than of individual species.
As they represent different evolutionary rates, thecombined analysis of the two data sets (Fig. 2) shouldprovide the greatest explanatory power for phyloge-netic reconstructions and is therefore justified for thepurposes of this study. Overall, all trees obtainedagreed in suggesting that Cissus is not a monophyleticgenus, that Cayratia and Tetrastigma are closely relatedand that Clematicissus is unlikely to remain as a mono-typic genus.

2002] 529ROSSETTO ET AL.: MONOPHYLY OF CISSUS
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_529File # 08TQ
Relationships Amongst the Australian Vita-ceae. CLADE A: CLEMATICISSUS ANGUSTISSIMA AND CISSUS
OPACA. Overall, the results presented here stronglysupport Cissus opaca and Clematicissus angustissima be-longing to a single genus. Together with AcareospermaGagnep. and Nothocissus (Miq.) Latiff from south-east-ern Asia and Pterocissus Urb. & Ekman from the WestIndies, Clematicissus was traditionally considered asone of the four monotypic genera within Vitaceae (La-tiff 1982). However, C. angustissima and C. opaca con-sistently paired in the analysis of the individual datasets and in Fig. 2 they form a highly supported cladedistinct from all other Vitaceae investigated. Jackes(1989) suggested a superficial resemblance betweenthese two species. They both share an unusual inflo-rescence arrangement (the normally unbranched ten-drils divide and one or both branches are terminatedby 1–2 umbels) and great variability between juvenileand adult leaf morphology within single individuals(Jackes 1988, 1989). Similarities in pollen morphologyhave also been reported (Lavie pers. com.), and bothspecies are distributed within comparable habitats,preferring drier environments than most of the otherAustralian Vitaceae. The findings suggest that this ge-nus is likely to have evolved within Australia beforecontact with the Asian flora.
CLADE B: CAYRATIA AND TETRASTIGMA. These twogenera comprise a single clade in the separate andcombined analyses. There are some discrepancies be-tween the results obtained from the two data sets.Plastid DNA sequences distinguished a monophyleticTetrastigma and two Cayratia clades, one of the latteralso including the single species of Ampelocissus suc-cessfully analyzed (it would however be premature tocome to any conclusion about Ampelocissus based onthese results alone). Within the nrDNA tree, Tetrastig-ma is paraphyletic relative to Cayratia japonica and C.trifolia. In a numerical study based on 46 morpholog-ical characters from 36 species in eight Malay genera,Latiff (1983) presented a tree divided into five majorgroups. These groups contained respectively: 1- Am-pelopsis and Cayratia; 2- Parthenocissus and Tetrastigma;3- Ampelocissus; 4- Pterisanthes Blume; 5- Nothocissusand Cissus. The first four groups were joined in a largecluster that was sister to the fifth group (Latiff 1983).Within these groups, Latiff found that Cayratia japonicaand C. trifolia (section Discypharia) were quite distinctfrom all other Cayratia and closer to Ampelopsis. Suchfindings are in some agreement with our moleculardata (in particular the nrDNA data), which defined amajor clade including Cayratia and Tetrastigma distinctfrom the Cissus clade. Both studies also agreed in dis-tinguishing Cayratia japonica and C. trifolia from mostof the other members of the genus. In the combinedanalysis these two species are part of a strongly sup-ported subclade that includes all the Tetrastigma spe-
cies analyzed (Fig. 2). Another small but strongly sup-ported sub-clade includes three Cayratia species (C. ac-ris, C. cardiophylla, and C. saponaria) sharing distinctivefloral characteristics and a green/yellow fruit colora-tion that distinguish them from all other Australianmembers of the genus. The molecular data corroboratethe close relationship between Cayratia and Tetrastigma,which are traditionally distinguished on the basis oflimited morphological characters (e.g. flowers are bi-sexual in Cayratia and unisexual in Tetrastigma, leavesare usually pubescent in Cayratia but glabrous in Te-trastigma). Further analysis incorporating a greaternumber of taxa will be needed to establish more pre-cisely the boundaries (if any) between these two generaand eventually modify the current classification.
A number of taxa within clade B are apparentlyclosely related. The Cayratia acris and C. saponaria in-dividuals tested only differed in a single cytosine de-letion at position 213 in the ITS1 sequence of C. sapon-aria. These two taxa have been separated because ofdifferences in overall leaf shape, degree of pubescence,and habitat preferences. Cayratia saponaria is restrictedto the rainforests of north-eastern Australia, whereasC. acris is also found in semi-deciduous vine thicketsand riverine gullies from northern New South Walesto New Guinea. Similarly, the results obtained for T.nitens and T. petraeum suggest a close relationship. Asfor the previous pair, these taxa are mainly distin-guished by their habitat preferences, T. petraeum (main-ly restricted to rocky habitats) has a narrower distri-bution within the northern parts of Australia than T.nitens.
CLADES C AND D: FOUR CISSUS SPECIES AND VITIS.The molecular data presented suggest that a furtherfour taxa should be removed from the genus Cissus. Inthe combined analysis four Cissus species fell within aclade distinct from all other Cissus, clade C, which alsoincorporated V. rotundifolia (Fig. 2). It has been previ-ously suggested that a number of these taxa, especiallythose with compound leaves or multiple seeds, shouldhave been removed from Cissus and possibly includedwithin Nothocissus (a simple-leaved monotypic genus;Latiff pers. com.). However, these characters alone donot justify the segregation from Cissus, and floral mor-phology does not support this artificial grouping sincethe distinctive fasciculate inflorescences of Nothocissusspicifera (Griff.) Latiff (Latiff 1982) are not present inany of these taxa. There are however, a number of mor-phological characteristics shared between most of theCissus species in clade C. In all four species the twocotyledons are similar in shape and have entire mar-gins. All other Cissus species examined and belongingto clade E have dissimilar cotyledons, one reniformand the other cordiform. Three species (C. antarctica,C. oblonga, and C. hypoglauca) share the development oflarge pubescent stipules that form a pocket around the

530 [Volume 27SYSTEMATIC BOTANY
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_530File # 08TQ

2002] 531ROSSETTO ET AL.: MONOPHYLY OF CISSUS
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_531File # 08TQ
←
FIG. 2. Representative MPT from the combined analysis with proportionally scaled branches. This tree is 1097 steps longwith CI 5 0.59 and RI 5 0.68. Bootstrap percentages (.50%) are listed above the branches and the letters representing themajor clades (as discussed within the text) are indicated on the side of the figure (the * marks a branch that is collapsed inthe strict consensus tree).
developing leaves, a feature not recorded in any otherCissus investigated. Domatia can be found on the lowerleaf surfaces of C. antarctica, C. oblonga and C. stercu-liifolia, also a feature not present within any of the oth-er Cissus. Further, a karyological study by Lavie (1970)found that C. antarctica is a polyploid like the Ameri-can taxa but unlike the African and Asian Cissus. Aninvestigation on the ploidy of the other species withinClade C is highly desirable. Finally, from the biogeo-graphical perspective, all four species are either en-demic, or near-endemic (i.e., limited to Australia andNew Guinea—Table 1) with the only potentially close-ly related taxon being Cissus acrantha Lauterb. fromNew Guinea (it shares a number of characteristics ex-clusive to this group). Such findings suggest that thisgroup of taxa is distinct from all other Cissus sampledand that the ancestor of this group may have evolvedwithin Australia.
Of particular interest is the relationship between thisnew group and Vitis. Identifying the Australian nativetaxa most closely related to Vitis was one of the aimsof this study, as it represented the basis for investigat-ing the interspecific genetic diversity potentially avail-able to grape improvement. The fact that these fournative species are found within a clade including V.rotundifolia and supported by low bootstrap value(55%—Fig. 2) can be interpreted in a number of ways.The molecular data could be interpreted as supportingthe reinstatement of the genus Vitis within Australia.However, it is worth noting that the Australian speciesform a weakly supported yet distinct subclade (Fig. 2)and that both regions also distinguished between V.rotundifolia and the other Vitis analysed (Fig. 1). Plan-chon (1887) originally classified V. rotundifolia within adistinct section, Muscadinia. Vitis rotundifolia has beencalled Muscadinia rotundifolia Small, based on variousmorphological characteristics, reproductive barriersand chromosome numbers (Bouquet 1983). Thus theplacement of the four Australian Cissus species withinthe genus Muscadinia is another possible alternative.Finally, in a study of morphological and anatomicaldevelopment in the Vitaceae, Gerrath and Posluszny(1994) selected C. antarctica as a genus representativeand found differences in flower development in com-parison to the representative of three other genera,Ampelopsis, Parthenocissus, and Vitis. They concludedthat the developmental features of C. antarctica wereintermediate between those of Vitis and Ampelopsis(Gerrath and Posluszny 1994). Thus overall, a more ac-curate hypothesis might be to consider C. antarctica, C.
hypoglauca, C. oblonga, and C. sterculiifolia as part of anew genus distinct from Vitis and Muscadinia.
Of further interest within clade C is the close rela-tionship between C. antarctica and C. oblonga, not re-solved by trnL but distinguishable by a number of mu-tations in the ITS1 sequence. Such relatedness confirmsthe observation of Jackes (1988) that, based on mor-phological evidence, C. oblonga (found in drier environ-mental conditions) could be considered as the decid-uous counterpart of C. antarctica.
CLADE E: CISSUS. One of the main aims of thisstudy was to assess the robustness of the genus Cissus.A number of characters have been previously suggest-ed as the basis for the segregation of species groups.For instance, it was suggested that C. hypoglauca, C.sterculiifolia, and C. penninervis should not be includedwithin the genus because of the presence of compoundleaves and the nature of the inflorescence. However,this does not appear to be a good character as thischaracter is also found in a number of other non-Aus-tralasian Cissus (e.g., C. aralioides (Welw. ex Baker)Planch. from Senegal, Angola, and Mozambique, C.gardneri Thwaites and C. trilobata Lam. from Sri Lanka,and C. tweediana from Argentina). Seed number was amain characteristic on which the Cayratia/Cissus divi-sion was based (Gagnepain 1911), with Cissus repre-senting the one-seeded taxa and Cayratia the multiple-seeded ones. Whereas multiple-seededness is in agree-ment with the exclusion of the five previously men-tioned taxa from clade E, it also sporadically holds truefor a number of other Australasian Cissus (e.g., C. ar-istata, C. penninervis, C. reniformis and C. vinosa; Jackes1988).
Based on the sequence data presented in this study,Cissus is not a monophyletic group. As discussed inthe previous sections, five species (C. antarctica, C. hy-poglauca, C. oblonga, C. opaca, C. sterculiifolia) do not fallwithin the main Cissus clade (clade E—Fig. 2). CladeE contains all non-Australian and non-endemic Cissusincluded, has strong bootstrap support, and exhibitsthe morphological characteristics traditionally associ-ated with this genus (such as simple leaves, leaf-op-posed umbellate cymes, four-merous flowers, mono-spermic fruits). Three recently described species ex-hibiting these characters (C. cardiophylla, C. pentaclada,and C. vinosa; Jackes 1988) also fall within this clade.
The combined phylogenetic analysis of plastid andnrDNA indicates that the Australian representatives ofthe genus Cissus do not form a clade. Cissus opaca andClematicissus angustissima belong to the same clade and

532 [Volume 27SYSTEMATIC BOTANY
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_532File # 08TQ
four other previously named Cissus species form a dis-tinct clade likely to represent a new genus. Some un-certainty remains regarding the relationship betweenTetrastigma and Cayratia, but these two genera appearto be closely related. Nevertheless, it is premature topropose a phylogenetic-based nomenclature of all theAustralian Vitaceae, as in most cases the type speciesfor the genera (Cayratia pedata (Lam.) Juss. & Gagnep.,Cissus vitiginea L. and Tetrastigma lanceolarium Planch.)were not included in this analysis. Overall, the rela-tionships among clades B, C, D, E could not be re-solved in this study, not an uncommon occurrence atthis taxonomic level. As a result, further sequencingand a greater number of species (including the typespecies for the genera investigated) will be needed toincrease the resolving power of future studies.
ACKNOWLEDGEMENTS. This project was funded in part by theAustralian Agriculture Research Institute. The authors are indebt-ed to Mark W. Chase and two anonymous referees for critical re-views of the manuscript. The authors also wish to thank the fol-lowing people for contributing to collection of samples: R. Borsch-man, K. Downs (RBGS), B. Hyland, E. M. Jackes, S. Wilson (DPIF,Tasmania).
LITERATURE CITED
BAKKER, F. T., D. HELLBRUGGE, A. CULHAM, and M. GIBBY. 1998.Phylogenetic relationships within Pelargonium sect. Peristera(Geraniaceae) inferred from nrDNA and cpDNA sequencecomparisons. Plant Systematics and Evolution 211: 273–287.
BALDWIN, B. G., M. J. SANDERSON, J. M. PORTER, M. F. WOJCIE-CHOWSKI, C. S. CAMPBELL, and M. J. DONOGHUE. 1995. TheITS region of the ribosomal DNA: a valuable source of evi-dence on Angiosperm phylogeny. Annals of the Missouri Bo-tanical Garden 82: 247–277.
BENTHAM, G. 1863. Flora Australiensis—Vol. I. London: Reeve & Co.BOUQUET, A. 1983. Contribution a l’etude de l’espece Muscadinia ro-
tundifolia et de ses hybrides avec Vitis vinifera. Applications enselection. PhD Thesis: University of Bordeaux. France.
BROWN, R. 1810. Promodus Florae Novae Hollandiae et Insulae Van-Damien. London: Taylor.
CHASE, M. W., D. SOLTIS, R. G. OLMSTEAD, D. MORGAN, D. H. LES,B. MISHLER, M. R. DUVALL, R. A. PRICE, H. G. HILLS, Y-L.QIU, K. A. KRON, J. H. RETTIG, E. CONTI, J. D. PALMER, J. R.MANHART, K. J. SYTSMA, H. J. MICHAELS, W. J. KRESS, K. G.KAROL, W. D. CLARK, M. HEDREN, B. S. GAUT, R. K. JANSEN,K-J. KIM, C. F. WIMPEE, J. F. SMITH, G. R. FURNIER, S. H.STRAUS, Q-Y. XIANG, G. M. PLUNKETT, P. S. SOLTIS, S. M.SWENSEN, S. E. WILLIAMS, P. A. GADEK, C. J. QUINN, L.EGUIARTE, E. GOLENBERG, G. H. LEARN, S. W. GRAHAM, S. C.H. BARRETT, S. DAYANANDAN, and V. A. ALBERT. 1993. Phy-logenetics of seed plants: an analysis of nucleotide sequencesfrom the plastid gene rbcL. Annals of the Missouri BotanicalGarden 80: 528–580.
CRONQUIST, A. 1988. The evolution and classification of floweringplants, 2nd ed. New York: The New York Botanical Garden.
CROS, J., M. C. COMBES, P. TROUSLOT, F. ANTHONY, S. HAMON, A.CHARRIER, and P. LASHERMES. 1998. Phylogenetic analysis ofchloroplast DNA variation in Coffea L. Molecular Phylogenet-ics and Evolution 9: 109–117.
DESCOINGS, B. 1960. Un genre de Vitacees: comprehension et dis-tinction des genres Cissus L., et Cyphostemma (Planch.) Alston.Notulae Systematicae 16: 113–125.
FELSENSTEIN, J. 1985. Confidence limits on phylogenies: an ap-proach using the bootstrap. Evolution 39: 783–791.
FITCH, W. M. 1971. Toward defining the course of evolution: min-imum change for a specific tree topology. Systematic Zoology20: 406–416.
GAGNEPAIN, F. 1911. Classification des Cissus et Cayratia. NotulaeSystematicae 1: 339–343.
GALET, P. 1967. Recherches sur les methodes d’identification et de clas-sification des Vitacees des zones temperees. PhD Thesis: Univer-sity of Monpellier. France.
GERRATH, J. M. and U. POSLUSZNY. 1994. Morphological and ana-tomical development in the Vitaceae. VI. Cissus antarctica. Ca-nadian Journal of Botany 72: 635–643.
GIELLY, L. and P. TABERLET. 1994. The use of chloroplast DNA toresolve plant phylogenies: noncoding versus rbcL sequences.Molecular Biology and Evolution 11: 769–777.
——— and ———. 1996. A phylogeny of the European gentiansinferred from chloroplast trnL (UAA) intron sequences. Bo-tanical Journal of the Linnean Society 120: 57–75.
GOLDMAN, N., J. P. ANDERSON, and A. G. RODRIGO. 2000. Likeli-hood-based tests of topologies in phylogenetics. SystematicBotany. 49: 652–670.
HILLIS, D. M. and M. T. DIXON. 1991. Ribosomal DNA: molecularevolution and phylogenetic inference. The Quarterly Reviewof Biology 66: 411–453.
HOPPER, S. D., M. F. FAY, M. ROSSETTO, and M. W. CHASE. 1999.A molecular phylogenetic analysis of the bloodroot and kan-garoo paw family, Haemodoraceae: taxonomic, biogeograph-ic and conservation implications. Botanical Journal of the Lin-nean Society 131: 285–299.
JACKES, B. R. 1984. Revision of the Australian Vitaceae, 1. Ampe-locissus Planchon. Austrobaileya 2: 81–86.
———. 1987. Revision of the Australian Vitaceae, 2. Cayratia Juss.Austrobaileya 2: 365–379.
———. 1988. Revision of the Australian Vitaceae, 3. Cissus L. Aus-trobaileya 2: 481–505.
———. 1989a. Revision of the Australian Vitaceae, 4. ClematicissusPlanchon. Austrobaileya 3: 11–102.
———. 1989b. Revision of the Australian Vitaceae, 5. Tetrastigma(MIQ.) Planchon. Austrobaileya 3: 149–158.
LATIFF, A. 1983. Numerical analysis and classification of MalesianVitaceae. Malays 12: 7–13.
———. 1982. Studies in Malesian Vitaceae, II. Nothocissus: a newMalesian genus. Federation Museums Journal 27: 70–74.
LAVIE, P. 1970. Contribution a l’etude caryostematique des Vitacees.PhD Thesis: University of Montpellier. France.
———. 1990. Vitacees de l’ouest Africain. Especes senegalaises. Sene-gal: Ministere de la cooperation et du developpement.
MABBERLEY, D. J. 1995. Vitaceae. Pp. 446–482 in A revised handbookto the flora of Ceylon, ed. M.D. Dassanayake. Amerind Publish-ing Co.
MCDADE, L. A. and M. L. MOODY. 1999. Phylogenetic relation-ships among Acanthaceae: evidence from noncoding trnL-trnF chloroplast DNA sequences. American Journal of Botany86: 70–80.
PLANCHON, J. E. 1887. Monographie des Ampelidees vraies. Pp:306–654 in Monog. Phan. 5, ed. A. de Candol. Paris: G. Mas-son.
ROSSETTO, M., B. R. JACKES, K. D. SCOTT, and R. J. HENRY. 2001a.Intergeneric relationships in the Australian Vitaceae: new ev-idence from cpDNA analysis. Genetic Research and CropEvolution 48: 307–314.
———, J. MCNALLY, and R. J. HENRY. 2001b. Evaluating the po-tential of SSR flanking regions for examining taxonomic re-lationships in Vitaceae. Theoretical and Applied Genetics 103:
SAVOLAINEN, V., M. F. FAY, D. C. ALBACH, A. BACKLUND, M. VAN
DER BANK, K. M. CAMERON, S. A. JOHNSON, M. D. LLEDO, J.
?1
?2

2002] 533ROSSETTO ET AL.: MONOPHYLY OF CISSUS
Tuesday Jul 16 2002 04:17 PMAllen Press • DTPro System
sbot 27_308 Mp_533File # 08TQ
C. PINTAUD, M. POWELL, M. C. SHEAHAN, D. E. SOLTIS, P. S.SOLTIS, P. WESTON, W. M. WHITTEN, K. J. WURDACK, and M.W. CHASE. 2000. Phylogeny of the eudicots: a nearly completefamilial analysis based on rbcL gene sequences. Kew Bulletin55: 257–309.
SLOWINSKI, J. B. and R. D. M. PAGE. 1999. How should speciesphylogenies be inferred from sequence data? Systematic Bi-ology 48: 814–825.
SMALL, R. L., J. A. RYBURN, R. C. CRONN, T. SEELANAN, and J. F.WENDEL. 1998. The tortoise and the hare: choosing betweennoncoding plastome and nuclear ADH sequences for phylog-eny reconstruction in a recently diverged plant group. Amer-ican Journal of Botany 85: 1301–1315.
SOLTIS, P. S., D. E. SOLTIS, and M. W. CHASE. 1999. Angiospermphylogeny inferred from multiple genes as a tool for com-parative biology. Nature 402: 402–404.
SOLTIS, D. E., P. S. SOLTIS, M. W. CHASE, M. E. MORT, D. C. AL-BACH, M. ZANIS, V. SAVOLAINEN, W. H. HAHN, S. B. HOOT,M. F. FAY, M. AXTELL, S. M. SWENSEN, L. M. PRINCE, W. J.KRESS, K. C. NIXON, and J. S. FARRIS. 2000. Angiosperm phy-
logeny inferred from 18S rDNA, rbcL, and atpB sequences.Botanical Journal of The Linnean Society 133: 381–461.
SWOFFORD, D. L. 1998. PAUP*. Phylogenetic Analysis using Parsimony(* and other methods). Version 4. Sunderland: Sinauer Associ-ates.
TABERLET, P., L. GIELLY, G. PAUTOU, and J. BOUVET. 1991. Universalprimers for amplification of three non-coding regions of chlo-roplast DNA. Plant Molecular Biology 17: 1105–1109.
THOMPSON, J. D., D. G. HIGGINS, and T. J. GIBSON. 1994. CLUSTALW: improving the sensitivity of progressive multiple se-quence alignment through sequence weighting, position spe-cific penalties and weight matrix choice. Nucleic Acids Re-search 22: 4673–4680.
VAN HAM, C. H. J., H. HART, T. H. M. MES, and J. M. SANDBRINK.1994. Molecular evolution of noncoding regions of the chlo-roplast genome in the Crassulaceae and related species. Cur-rent Genetics 25: 558–566.
WHITE, T. J., T. BRUNS, S. LEE, and J. TAYLOR. 1990. Amplificationand direct sequencing of fungal ribosomal RNA genes forphylogenetics. Pp. 315–322 in PCR protocols: A guide to methodsand applications, eds. M. Innis, D. Gelfand, J. Sninsky. and T.White. San Diego: Academic Press.
Copyright © 2022 FDOKUMEN




















