
2012 - A família Orchidaceae no município de Morro do Chapéu, Bahia, Brasil

Phylogenetic analysis of Chloraeinae (Orchidaceae)based on plastid and nuclear DNA sequences
MAURICIO A. CISTERNAS1,3*, GERARDO A. SALAZAR2, GABRIELA VERDUGO1,PATRICIO NOVOA3, XIMENA CALDERÓN4 and MARÍA A. NEGRITTO5
1Escuela de Agronomía, Pontificia Universidad Católica de Valparaíso, calle San Francisco s/n, LaPalma, Quillota, Chile2Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México,Apartado Postal 70-367, 04510, México DF, México3Jardín Botánico Nacional, camino El Olivar 305, El Salto, Viña del Mar, Chile4Instituto de Ciencias e Investigación, Universidad Arturo Prat, Ejército 443, Puerto Montt, Chile5Departamento de Botánica, Facultad de Ciencias naturales y oceanográficas, Universidad deConcepción, casilla 160-c, Concepción, Chile
Received 22 November 2010; revised 8 September 2011; accepted for publication 20 October 2011
The phylogenetic relationships of subtribe Chloraeinae, a group of terrestrial orchids endemic to southern SouthAmerica, have not been satisfactorily investigated. A previous molecular phylogenetic analysis based on plastidDNA supported the monophyly of Chloraeinae and Gavilea, but showed that Chloraea is non-monophyletic and thatthe sole species of Bipinnula analysed is sister to Geoblasta. However, that analysis included only 18 of the 73species belonging to this subtribe. Here, the phylogenetic relationships of Chloraeinae were assessed by analysingaproximately 7500 bp of nucleotide sequences from nuclear ribosomal internal transcribed spacer (ITS) and plastidDNA (rbcL, matK, trnL-trnF, rpoB-trnC) for 42 species representing all four currently accepted genera ofChloraeinae and appropriate outgroups. Nuclear and plastid data were analysed separately and in combinationusing two different methods, namely parsimony and Bayesian inference. Our analyses support the monophyly ofChloraeinae and their inclusion in an expanded concept of Cranichideae, but none of the genera of Chloraeinae thatincludes more than one species is monophyletic. Gavilea and Bipinnula are paraphyletic, with Chloraea chicanested in Gavilea and Geoblasta penicillata in Bipinnula. As currently delimited, Chloraea is polyphyletic. Thetaxonomic changes proposed recently are for the most part not justifiable on phylogenetic grounds, except forrecognition of the monotypic genus Correorchis. The lack of resolution for the relationships among species of ‘core’Chloraea suggests a relatively recent diversification of this group. The current generic classification is in need orrevision, but additional study is advisable before carrying out further taxonomic changes. © 2011 The LinneanSociety of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••.
ADDITIONAL KEYWORDS: Bipinnula – Chloraea – Gavilea – Geoblasta – molecular phylogeny – SouthAmerica.
INTRODUCTION
As delimited in the most recent classification systemof Orchidaceae (Chase et al., 2003; Pridgeon et al.,2003), subtribe Chloraeinae comprises four genera oforchids endemic to South America: Bipinnula Comm.
ex Juss., Chloraea Lindl., Gavilea Poepp. and Geo-blasta Barb.Rodr. Chloraea is the largest genus, withc. 48 species located in three disjunct areas (Hauman,1922; Correa, 1969: (1) a northern group that includes16 species from Bolivia, Peru and northern Argentina;(2) an eastern group with two species from easternArgentina, Brazil and Uruguay; and (3) a westerngroup with c. 30 species from Chile and Argentina.*Corresponding author. E-mail: [email protected]
1
2
3
4
567
8
9
10
11
12
13
14
15
16
17
1819202122232425262728293031323334
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
boj_1200_
Botanical Journal of the Linnean Society, 2011, ••, ••–••. With 6 figures
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–•• 1

Bipinnula occurs in two disjunct areas; one of thesecomprises southern Brazil, Uruguay and easternArgentina (six species) and the other is Chile (fivespecies). Gavilea encompasses 10–13 species found inChile and Argentina (including the Juan Fernandezand Falkland Islands) and Geoblasta is monospecificand restricted to southern Brazil, Uruguay andeastern Argentina (Correa, 1956, 1969; Izaguirre,1973; Correa & Sánchez, 2003; Novoa et al., 2006).
Species of Chloraeinae are terrestrial, only rarelybeing found living epiphytically or on rocks. Plants
have fleshy roots that are fasciculate or sometimesspaced along a rhizome (Fig. 1), and their leaves forma basal rosette or, uncommonly, are spirally arrangedalong the stem. The inflorescence is terminal, produc-ing one to many spirally disposed flowers subtendedby prominent bracts. The flowers are resupinate, withfree sepals and petals with or without conspicuouslongitudinal or reticulate veining. The apices of thelateral sepal can be fleshy, membranous or providedwith wart-like outgrowths; they often bear osmo-phores and sometimes have a fimbriate–pectinate
Figure 1. Roots of Chloraeinae. A, Chloraea chica. B, Gavilea araucana. C, Chloraea crispa. D, Bipinnula fimbriata.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
boj_1200_
2 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

extension, as in most species of Bipinnula (Fig. 2).The labellum is free, sessile or clawed, dissimilar orsimilar to the other perianth parts in size, shape andcolouration; the lip blade is membranaceous or fleshy,entire or three-lobed, smooth or more commonlyadorned with warts, calluses or crests. The columnranges from short to elongated, straight to arcuate,wingless or narrowly winged and with or without apair of nectaries between the column and the label-lum. Swollen nectaries are found in most Gavilea spp.and nectariferous channels are found in Chloraea andsome Bipinnula spp. The anther is terminal, erect orslightly incumbent, bilocular and produces fourpowdery pollinia in two pairs (Correa, 1956, 1969;Izaguirre, 1973; Dressler, 1993; Correa & Sánchez,2003; Novoa et al., 2006). Pollen is arranged intetrads (Ackerman & Williams, 1981). The stigma isventral, concave and entire. The rostellum is ovate ortriangular and blunt and it does not have a distinctviscidium, but a viscarium (Dressler, 1993) or ‘diffuseviscidium’ sensu Rasmussen (1982) is present (cf.Szlachetko & Rutkowski, 2000; Fig. 3).
The genera of Chloraeinae are separated by variousfloral traits, but it should be noted that probably noneof these traits is constant or unique to a genus, exceptfor the insect-like labellum of the monotypic Geo-blasta, which is related to its specialized pollinationmechanism involving pseudocopulation by scoliidwasps (Ciotek et al., 2006). In Bipinnula, the apices ofthe lateral sepals are usually fimbriate–pectinate,except in B. apinnula Gosewijn, in which they areentire. Gavilea usually has an abbreviated columnand swollen nectaries, whereas Chloraea is distin-guished by a combination of characters or by theabsence of a particular character. For instance, thelabellum in Chloraea has nectariferous channels, butthese are shared with multi-flowered Bipinnula spp.and an elongate column allows for its separation frommost Gavilea spp. but not from Geoblasta or Bipin-nula. Often the floral characters have been usedinconsistently, as in the key to the genera of Chlo-raeinae in Correa (2003), in which Gavilea is sepa-rated from Chloraea, among others, by the geniculateovary. Nevertheless, the generic description ofGavilea in the same work indicates that the ovary isstraight or geniculate.
Historically, the systematic position of Chloraeinaehas been controversial. Most authors have includedthem in various versions of tribe Diurideae because ofsimilarities in overall flower organization, columnstructure, exine morphology, pollen organization andthe presence of an erect anther attached to thecolumn apex via a short filament, as in many Austra-lian genera of Diurideae (Brieger, 1974–1975;Dressler, 1974, 1981, 1993; Ackerman & Williams,1981; Rasmussen, 1982, 1985). Brieger (1974–1975),
based on pollen organization and the presence of aviscidium, divided Chloraeinae into two groups:Aviscidia, including the South American generaBipinnula, Chloraea, Gavilea and Geoblasta, and Vis-cidifera, that included Megastylis Schltr., RimacolaRupp (both Diurideae according to Chase et al., 2003and references cited therein) and PachyplectronSchltr. (now considered a member of Goodyerinae,tribe Cranichideae; Chase et al., 2003; Salazar et al.,2003). Burns-Balogh & Funk (1986) included thegenera of Chloraeinae in tribe Geoblasteae, subfamilyNeottioideae, based on the possession of broad stami-nodes fused to the sides of the column (= columnwings), soft, mealy pollinia, elongated column,reduced rostellum and solid to semi-solid viscidium.However, most of these characters are widely distrib-uted in different combinations in Cranichideae andDiurideae sensu Pridgeon et al. (2003) and genuineviscidia appear to be absent from Chloraeinae s.s. (seeabove). Szlachetko & Rutkowski (2000) followed asimilar scheme to that of Burns-Balogh & Funk(1986), placing Geoblasteae in subfamily Thelymitroi-deae (Szlachetko, 1991), a synonym of Orchidoideae.
Chloraeinae have also been linked to the diuridsbecause the southern South American genus Codo-norchis Lindl., traditionally placed among Chlo-raeinae, produces root tubers (the so-called ‘root-stemtuberoids’) similar to those of various Australiangenera of Diurideae (see Pridgeon & Chase, 1995).Dressler (1993) suggested that the absence of tube-roids in Bipinnula, Chloraea, Gavilea and Geoblastamay represent a secondary loss. However, phylogeneticanalyses based on plastid (Kores et al., 2001) andnuclear DNA (Clements et al., 2002) have shown thatCodonorchis is not closely related to Chloraeinae s.s.,having been instead placed in a tribe on its own, Codo-norchideae (Cribb & Kores, 2000) or even as a distinctsubfamily, Codonorchidoideae (Jones et al., 2002).
On the one hand, recent phylogenetic analysesbased on DNA sequences (Kores et al., 1997, 2000,2001; Cameron et al., 1999; Clements et al., 2002;Salazar et al., 2003) and embryological studies (Clem-ents, 1999) have shown that Chloraeinae are moreclosely related to members of tribe Cranichideaesensu Dressler (1993) than to Diurideae, supportingthe inclusion of Chloraeinae in an expanded conceptof Cranichideae (Clements et al., 2002; Chase et al.,2003; Salazar et al., 2003) or, alternatively, their rec-ognition as a tribe on their own, namely Chloraeeae(as in Pridgeon et al., 2003). However, those studieshave included only a few representatives of Chloraeaand Gavilea. On the other hand, several taxonomicchanges in Chloraeinae have been proposed recently,but none of these was backed up by phylogeneticevidence (Szlachetko & Margonska, 2001; Szlachetko& Tukałło, 2008).
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
boj_1200_
PHYLOGENY OF CHLORAEINAE 3
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
1 1
22

Figure 2. Representative species of Chloraeinae. A, Chloraea alpina. B, C. prodigiosa. C, C. disoides. D, C. cylin-drostachya. E, C. gaudichaudii. F, C. lamellata. G, C. magellanica. H, C. barbata. I, C. speciosa. J, C. philippii.K, C. nudilabia. L, Gavilea araucana. M, G. venosa. N, Chloraea chica. O, Bipinnula fimbriata. P, Gavilea odoratissima.Q, G. australis. R, Geoblasta penicillata. S, Bipinnula apinnula.
1234
boj_1200_
4 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

Chemisquy & Morrone (2010) conducted the firstphylogenetic analysis of Choraeinae, in which theyincluded 22 specimens of 18 species representing thefour currently recognized genera (ten Chloraea spp.,six Gavilea spp. and one species each of Bipinnulaand Geoblasta). They used nucleotide sequences ofthree plastid DNA regions, the gene rpoC1, the atpB-rbcL intergenic spacer and part of the trnK intron(including the matK pseudogene). Their study sup-ported the monophyly of Chloraeinae s.s. andof Gavilea, but showed that Chloraea is non-monophyletic and that the sole species of Bipinnulaanalysed is sister to Geoblasta. However, their limitedtaxonomic sampling prevented them from drawingconclusions on generic limits and relationships, and abroader sample of species and molecular characterswould contribute to a better understanding of thephylogenetic relationships in the subtribe.
In the present study, the phylogenetic relationshipsin Chloraeinae are assessed by analysing more inclu-sive samples of both taxa and characters than previ-ous analyses. The data analysed here include fiveplastid DNA regions [gene rbcL, pseudogene matKwith part of the trnK intron in which it is embedded,rpoB-trnC and trnL-trnF intergenic spacers (IGS)plus the internal transcribed region of nuclear ribo-
somal DNA (nrITS)]. These genomic regions havebeen used successfully for phylogenetic reconstructionat various taxonomic levels in several groups ofOrchidaceae (e.g. Kores et al., 1997, 2000, 2001; vanden Berg et al., 2000, 2005; Gravendeel et al., 2001;Salazar et al., 2003, 2009; Álvarez-Molina &Cameron, 2009; Chiron et al., 2009; Monteiro et al.,2010). Our aim is to gain insights into the phyloge-netic relationships within Chloraeinae s.s. by evalu-ating subtribal and generic monophyly and todiscussing the merits of recently proposed taxonomicchanges.
MATERIAL AND METHODSTAXONOMIC SAMPLING
Exemplars of 42 species of Chloraeinae representingthe four genera recognized by Pridgeon et al. (2003)were analysed for this study. Representatives of othersubtribes of Cranichideae sensu Chase et al. (2003)and of Diurideae, Codonorchideae and Orchideae,were included as outgroups following previousmolecular phylogenetic analyses (Kores et al., 1997,2000, 2001; Clements et al., 2002; Salazar et al., 2003,2009). A list of the taxa analysed with voucher
Figure 3. Gynostemium structure of Chloraeinae, side (left) and front views (right). A, B, Gavilea venosa. C, D, Chloraeacrispa. E, F, C. galeata. G, H, C. multiflora.
12
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
boj_1200_
PHYLOGENY OF CHLORAEINAE 5
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
33
44

information and GenBank accessions is provided inTable 1. The aligned matrix is available on requestfrom the first author (M.A.C.).
DNA EXTRACTION, AMPLIFICATION AND SEQUENCING
Total DNA was mainly extracted from fresh or silicagel-dried tissue, but herbarium material was used insome instances. DNA extraction was carried out withthe 2 ¥ cetyl trimethylammonium bromide (CTAB)procedure of Doyle & Doyle (1987), modified by theaddition of 2% (w/v) of polyvinylpirrolidone (PVP) tothe extraction buffer. Amplification of the target DNAregions was performed using a commercial kit (TaqPCRCore Kit; Qiagen, Hilden, Germany) followingthe manufacturers’ protocols. The primers used areindicated in Table 2.
PCR profiles for rbcL and the matK-trnK, trnL-trnFand nrITS regions were as in Salazar et al. (2003).For the rpoB-trnC IGS, an initial ‘pre-melt’ (94 °C for2 min) was followed by 28–32 cycles of 94 °C for 30 s,53 °C for 40 s and 72 °C for 40 s, concluding with afinal extension at 72 °C for 7 min. All PCR productswere cleaned with QIAquick silica columns (Qiagen)and used in cycle sequencing reactions with the ABIPrism Big Dye® Terminator Cycle Sequencing ReadyReaction kit with AmpliTaq® DNA polymerase, ver-sion 3.1 (Applied Biosystems Inc., Warrington, UK).Cycle sequencing products were cleaned with Centri-Sep sephadex columns (Princeton Separations, Inc.,Adelphia, NJ, USA) and sequenced in a 3100 GeneticAnalyzer (Applied Biosystems). Both forward andreverse sequence DNA strands were assembled andedited with the software Sequencher 4.8 (GeneCodes,Ann Arbor, MI, USA).
SEQUENCE ALIGNMENT AND INDEL CODING
Sequences of the length-conserved rbcL gene werealigned unambiguously by visual inspection, but thesequences of the nrITS, trnL-trnF, rpoB-trnC andmatK-trnK regions, which show length variation,were aligned using the E-INS-i iterative strategy(Katoh et al., 2005) of the online submission versionof the program MAFFT version 6 (Katoh, Asimenos& toh, 2009), with minor subsequent manualadjustment.
One, five, five, three and three sequences werepartially or completely missing from the rbcL, matK-trnK, trnL-trnF, nrITS, and rpoB-trnC data sets,respectively; together, the missing data amount toc 4% of the data cells in the aligned matrix.
PHYLOGENETIC ANALYSES
Previous phylogenetic analyses of Cranichideae basedon plastid and nuclear DNA used here have shown
that the different regions produce similar patterns ofrelationship and that, when they are analysed incombination, both resolution and internal cladesupport are maximized (e.g. Salazar et al., 2003,2009). Here, we conducted parsimony analyses ofthree data matrices: (1) nrITS; (2) all plastid regions;and (3) all the data combined. The analyseswere carried out using the program PAUP*version 4.0b10 for Macintosh (Swofford, 2002) andeach consisted of a heuristic search with 1000replicates of random sequence addition with treebisection–reconnection (TBR) branch swapping andthe MULTREES option activated, saving up to 20most parsimonious trees (MPTs) from each replicate.All characters were considered as unordered andequally weighted (Fitch, 1971). Individual gappositions were treated as missing data. Internalsupport for clades was assessed by non-parametricbootstraping (Felsenstein, 1985), performing 500bootstrap replicates, each with 20 replicates ofrandom sequence addition and TBR branch swapping,saving up to 20 trees per replicate. Clades obtaininga bootstrap percentage (BP) � 50 were considered asunsupported, 51–70% as weakly supported, 71–85%as moderately supported, and 86–100% as stronglysupported. In all analyses, Ophrys apifera Huds.(Orchideae) was used as prime outgroup (Barriel &Tassy, 1998).
Additionally, we conducted a model-based phyloge-netic analysis of the combined matrix using BayesianMarkov chain Monte Carlo (MCMC) inference, asimplemented in the program MrBayes version 3.1.2(Ronquist, Huelsenbeck & Van der Mark, 2005), togenerate an independent phylogenetic hypothesis forcontrast with the parsimony trees. The best-fittingmodels of sequence evolution were determined sepa-rately for rbcL, matK, trnK intron (excluding matK),trnL intron, trnL-trnF IGS, rpoB-trnC IGS and nrITSusing Modeltest 3.7 (Posada & Crandall, 1998). In allcases, a six-parameter model with among-site rateheterogeneity modelled according to a gamma distri-bution was selected, and for rbcL, matK and nrITSthere was also a proportion of invariant sites. Theappropriate character partitions were stipulated inMrBayes and all model parameters were unlinkedamong the partitions, such that each group of char-acters was allowed to have its own set of parameters(Ronquist et al., 2005). Two simultaneous analyseswere run for 1 000 000 generations using the defaultconditions of MrBayes for the Markov chains. Thetrees were sampled every hundredth generation andthe first 250 000 generations (2500 trees) of each runwere discarded as burn-in. Inferences about relation-ships and posterior probabilities of clades (PP) werebased on a majority-rule summary tree constructedby pooling the remaining 15 000 trees. Posterior prob
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
boj_1200_
6 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

Table 1. Taxa studied, voucher information and GenBank accessions
Taxon Voucher
GenBank accession
rpoB-trnC rbcL trnL-trnF matK nrITS
Tribe CodonorchideaeP.J.Cribb
Codonorchis lessonii(Brongn.) Lindl.
South America, Kores& Molvray 332, OKL
FR831995 – – AJ310008 –
Chile, Ryan 002, K(spirit)
– AJ542399 AJ544508 – AF348005
Tribe CranichideaeEndl.
Subtribe AchlydosinaeM.A.Clem. &D.L.Jones
Achlydosa glandulosa(Schltr.) M.A.Clem. &D.L.Jones
New Caledonia,Clements D-285,CANB
FR832001 AJ542401 AJ544506 AJ543950 AJ539525
Subtribe ChloraeinaeRchb.f.
Bipinnula apinnulaGosewijn
Chile, Cisternas 110,CONC
FR832003 FR831957 – FR832063 FR832100
Chile, Knees 4438, K FR832136Bipinnula fimbriata
(Phil.) Johnst.Chile, Cisternas 111,
CONCFR832004 FR831958 FR832135 FR832064 FR832101
Bipinnula montanaArechav.
Uruguay, Cisternas112, CONC
FR832005 FR831959 FR832137 FR832065 FR832102
Bipinnula volkmanniKraenzl.
Chile, Rodríguez &Marticorena 2259,CONC
FR832002 FR831956 – FR832062 FR832099
Chloraea alpina Poepp. Chile, Saavedra &Pauchard 296,CONC
Pending FR831974 FR832151 FR832079 FR832114
Chloraea barbataLindl.
Chile, Cisternas 106,CONC
FR832012 FR831965 FR832142 FR832070 –
Chloraea bicallosa Phil.ex Kraenzl.
Chile, Cisternas 113,CONC
FR832013 FR831966 FR832143 FR832071 FR832108
Chloraea bidentata(Poepp.) M.N.Correa
Chile, Cisternas 109,CONC
FR832014 FR831967 FR832144 FR832072 –
Chloraea bletioidesLindl.
Chile, Cisternas 104,CONC
FR832017 FR831970 FR832147 FR832075 FR832111
Chloraea chica Speg. &Kraenzl.
Chile, Tellier &Márquez 5328,CONC
FR832023 FR831977 FR832153 FR832082 FR832117
Chloraea chrysanthaPoepp.
Chile, Novoa s.n.,CONC
FR832020 FR831973 FR832150 FR832078 FR832113
Chloraea crispa Lindl. Chile (cultivatedspecimen), Cisternas103, CONC
FR832015 FR831968 FR832145 FR832073 FR832109
Chloraea cristata Lindl. Chile, Novoa 177,CONC
FR832016 FR831969 FR832146 FR832074 FR832110
Chloraea cuneataLindl.
Chile, Espejo s.n.,CONC
FR832018 FR831971 FR832148 FR832076 –
ChloraeacylindrostachyaPoepp.
Chile, Cisternas 123,CONC
FR832024 FR831978 FR832154 FR832083 FR832118
123456789
10111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455565758
boj_1200_
PHYLOGENY OF CHLORAEINAE 7
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

Table 1. Continued
Taxon Voucher
GenBank accession
rpoB-trnC rbcL trnL-trnF matK nrITS
Chloraea disoidesLindl.
Chile, Cisternas 122,CONC
FR832021 FR831975 FR832152 FR832080 FR832115
Chloraea gaudichaudiiBrongn.
Chile, Cisternas 120,CONC
FR832022 FR831976 pending FR832081 FR832116
Chloraea gavilu Lindl. Chile (cultivatedspecimen), Cisternas102, CONC
FR832027 FR831981 FR832157 FR832086 FR832121
Chloraea grandifloraPoepp.
Chile, Espejo 21,CONC
FR832007 FR831960 – FR832061 FR832103
Chloraea heteroglossaReichb. f.
Chile (cultivatedspecimen), Cisternas105, CONC
FR832011 FR831964 FR832141 FR832069 FR832107
Chloraea incisa Poepp. Chile (cultivatedspecimen), Cisternass.n., CONC
FR832031 FR831984 FR832159 FR832089 FR832125
Chloraea lamellataLindl.
Chile, Cisternas 116,CONC
FR832026 FR831980 FR832156 FR832085 FR832120
Chloraea lechleri Lindl.ex Kraenzl.
Chile, Cisternas 107,CONC
FR832030 FR831983 FR832158 FR832088 FR832124
Chloraea longipetalaLindl.
Chile, Cisternas 115,CONC
FR832019 FR831972 FR832149 FR832077 FR832112
Chloraea magellanicaHook.f.
Chile, Ryan 1, K(spirit)
FR832006 AJ542403 AJ544504 AJ543948 AJ539523
Chloraea membranaceaLindl.
Chile, Cisternas 108,CONC (photographand dissected flower)
FR832033 FR831986 FR832161 FR832091 FR832126
Chloraea multifloraLindl.
Chile, Novoa 126,CONC
FR832025 FR831979 FR832155 FR832084 FR832119
Chloraea nudilabiaPoepp.
Chile, Cisternas 114,CONC
FR832032 FR831985 FR832160 FR832090 FR832126
Chloraea philippiiReichb. f.
Chile, Cisternas 119,CONC
FR832010 FR831963 FR832140 FR832068 FR832106
Chloraea prodigiosaReichb. f.
Chile (cultivatedspecimen), Cisternas101, CONC
FR832029 FR831982 – – FR832123
Chloraea reticulataSchltr.
Peru, Weigend2000-361, NY
– FJ571318 FJ571268 – FJ473314
Chloraea speciosaPoepp.
Chile, Cisternas 121,CONC
FR832009 FR831962 FR832139 FR832067 FR832105
Chloraea virescens(Willd.) Lindl.
Chile, Cisternas 117,CONC
FR832028 – – FR832087 FR832122
Chloraea volkmanniPhil. ex Kraenzl.
Chile, Cisternas 118,CONC
FR832008 FR831961 FR832138 FR832066 FR832104
Gavilea araucana(Phil.) M.N.Correa
Chile, Cisternas 124,CONC
FR832036 FR831988 FR832162 FR832093 FR832129
Gavilea australis(Skottsberg)M.N.Correa
Chile, Cisternas 125,CONC
FR832034 FR831987 pending FR832092 FR832128
Gavilea glandulifera(Poepp.) M.N.Correa
Chile, Cisternas 126,CONC
FR832038 FR831990 FR832164 FR832095 FR832131
Gavilea leucanthaPoepp. et Endl.
Chile, Novoa 259,CONC
FR832037 FR831989 FR832163 FR832094 FR832130
123456789
10111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455565758
boj_1200_
8 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
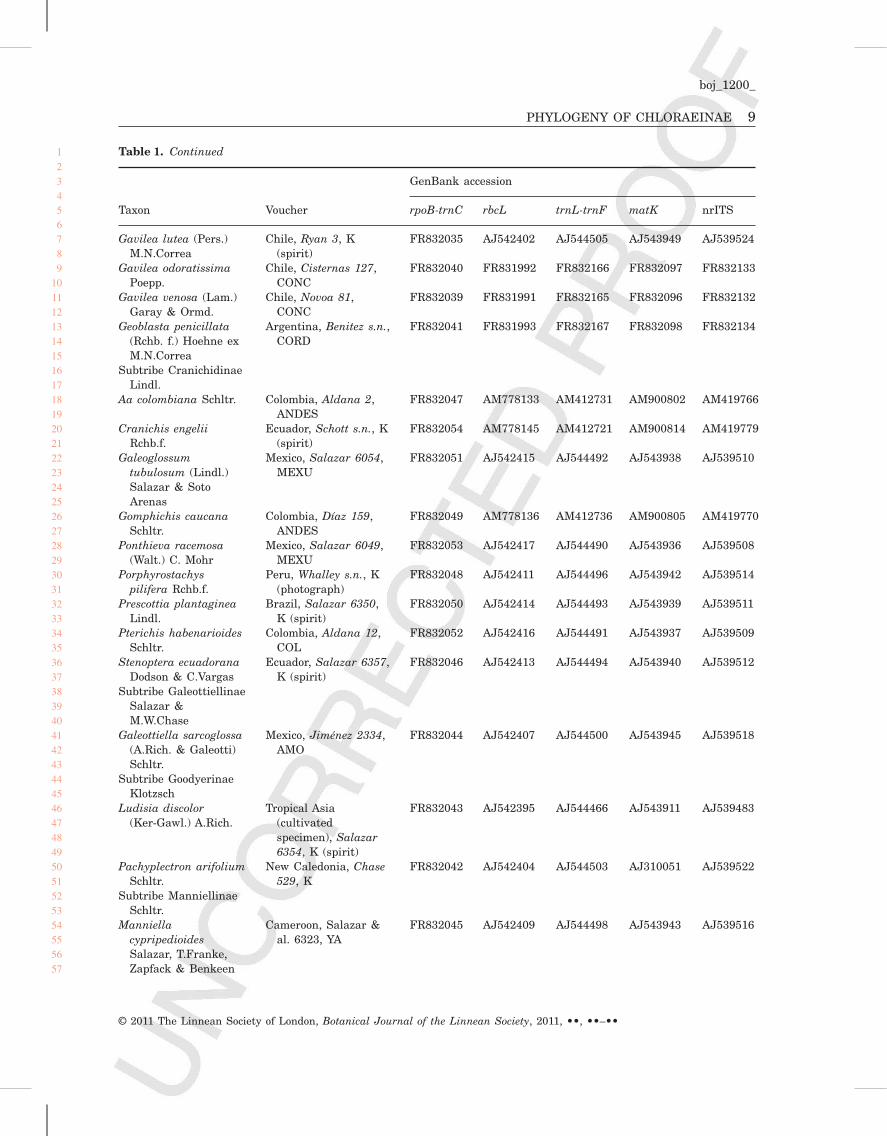
Table 1. Continued
Taxon Voucher
GenBank accession
rpoB-trnC rbcL trnL-trnF matK nrITS
Gavilea lutea (Pers.)M.N.Correa
Chile, Ryan 3, K(spirit)
FR832035 AJ542402 AJ544505 AJ543949 AJ539524
Gavilea odoratissimaPoepp.
Chile, Cisternas 127,CONC
FR832040 FR831992 FR832166 FR832097 FR832133
Gavilea venosa (Lam.)Garay & Ormd.
Chile, Novoa 81,CONC
FR832039 FR831991 FR832165 FR832096 FR832132
Geoblasta penicillata(Rchb. f.) Hoehne exM.N.Correa
Argentina, Benitez s.n.,CORD
FR832041 FR831993 FR832167 FR832098 FR832134
Subtribe CranichidinaeLindl.
Aa colombiana Schltr. Colombia, Aldana 2,ANDES
FR832047 AM778133 AM412731 AM900802 AM419766
Cranichis engeliiRchb.f.
Ecuador, Schott s.n., K(spirit)
FR832054 AM778145 AM412721 AM900814 AM419779
Galeoglossumtubulosum (Lindl.)Salazar & SotoArenas
Mexico, Salazar 6054,MEXU
FR832051 AJ542415 AJ544492 AJ543938 AJ539510
Gomphichis caucanaSchltr.
Colombia, Díaz 159,ANDES
FR832049 AM778136 AM412736 AM900805 AM419770
Ponthieva racemosa(Walt.) C. Mohr
Mexico, Salazar 6049,MEXU
FR832053 AJ542417 AJ544490 AJ543936 AJ539508
Porphyrostachyspilifera Rchb.f.
Peru, Whalley s.n., K(photograph)
FR832048 AJ542411 AJ544496 AJ543942 AJ539514
Prescottia plantagineaLindl.
Brazil, Salazar 6350,K (spirit)
FR832050 AJ542414 AJ544493 AJ543939 AJ539511
Pterichis habenarioidesSchltr.
Colombia, Aldana 12,COL
FR832052 AJ542416 AJ544491 AJ543937 AJ539509
Stenoptera ecuadoranaDodson & C.Vargas
Ecuador, Salazar 6357,K (spirit)
FR832046 AJ542413 AJ544494 AJ543940 AJ539512
Subtribe GaleottiellinaeSalazar &M.W.Chase
Galeottiella sarcoglossa(A.Rich. & Galeotti)Schltr.
Mexico, Jiménez 2334,AMO
FR832044 AJ542407 AJ544500 AJ543945 AJ539518
Subtribe GoodyerinaeKlotzsch
Ludisia discolor(Ker-Gawl.) A.Rich.
Tropical Asia(cultivatedspecimen), Salazar6354, K (spirit)
FR832043 AJ542395 AJ544466 AJ543911 AJ539483
Pachyplectron arifoliumSchltr.
New Caledonia, Chase529, K
FR832042 AJ542404 AJ544503 AJ310051 AJ539522
Subtribe ManniellinaeSchltr.
ManniellacypripedioidesSalazar, T.Franke,Zapfack & Benkeen
Cameroon, Salazar &al. 6323, YA
FR832045 AJ542409 AJ544498 AJ543943 AJ539516
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
boj_1200_
PHYLOGENY OF CHLORAEINAE 9
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

Table 1. Continued
Taxon Voucher
GenBank accession
rpoB-trnC rbcL trnL-trnF matK nrITS
SubtribePterostylidinae Pfitz.
Pterostylis curta R.Br. Australia, Chase 572,K
FR832000 AJ542400 AJ544507 AJ543951 AJ539526
Subtribe SpiranthinaeLindl.
Cyclopogon epiphyticus(Dodson) Dodson
Ecuador, Salazar 6355,K
FR832057 AJ542425 AJ544482 AJ543927 AJ539499
Dichromanthuscinnabarinus (LaLlave & Lex.) Garay
Mexico, Linares 4469,MEXU
FR832060 AJ542438 AJ544469 AJ543914 AJ539486
Mesadenus lucayanus(Britton) Schltr.
Mexico, Salazar 6043,MEXU
FR832059 AJ542436 AJ544471 AJ543916 AJ539488
Sarcoglottis acaulis(J.E.Sm.) Schltr.
Trinidad, Salazar6356, K (spirit)
FR832056 AJ542424 AJ544483 AJ543928 AJ539500
Spiranthes cernua (L.)Rich.
USA, Nickrent 4188,MEXU
FR832058 AJ542435 AJ544472 AJ543916 AJ539489
StenorrhynchosglicensteiniiChristenson
Mexico, Salazar 6090,MEXU
FR832055 AJ542420 AJ544487 AJ543532 AJ539505
Tribe Diurideae Endl.Subtribe Acianthinae
(Lindl.) Schltr.Acianthus caudatus
R.Br.n.a. – – – – AF347976
Acianthus exsertusR.Br.
Australia, Chase 565,K
FR831999 AF074101 AJ409373 AJ309993 –
Subtribe CaladeniinaePfitzer
Microtis parvifloraR.Br.
Australia, Chase 553,K
FR831997 AF074194 AJ409428 AJ310045 –
Australia, ‘MA21’,CANB
– – – – DQ104550
Subtribe DiuridinaeLindl.
Diuris sulphurea R.Br. Australia, Chase 554,K
FR831998 AJ542398 AJ544509 AJ543952 AJ539527
SubtribeCryptostylidinaeSchltr.
Cryptostylis subulata(Labill.) Rchb.f.
Australia, Chase 332,K
FR831996 AF074140 AJ409395 AJ310015 AF348015
Tribe OrchideaeSubtribe Orchidinae
Dressler & DodsonOphrys apifera Huds. UK, Chase 536, K FR831994 AJ542396 AJ544511 AJ543953 AJ539529
nrITS, nuclear ribosomal internal transcribed spacer.
123456789
1011121314151617181920212223242526272829303132333435363738394041424344454647484950515253
54
boj_1200_
10 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

abilities (PP) � 0.95 were considered as strongsupport, 0.90–0.94 as moderate support and < 90 asweak support.
RESULTSPARSIMONY ANALYSES
The nrITS analysis consisted of 775 aligned positions,of which 397 (51%) were potentially parsimony-informative. The analysis found 254 shortest treeswith a length of 2111 steps, consistency index exclud-ing uninformative characters (CI) = 0.42 and retentionindex (RI) = 0.67. Figure 4A shows one of the trees andFigure 4B the strict consensus of the 254 trees, onwhich the bootstrap percentages are indicated. Mono-phyly of Chloraeinae obtained strong support, as didits sister-group relationship to other Cranichideae.Within Chloraeinae, C. cylindrostachya Poepp. andC. reticulata Schltr. diverge successively, the latterbeing sister to the remaider of the subtribe, whichconsists of a polytomy consisting of a clade withC. chica Kraenzl. & Speg. sister to a monophyleticGavilea, a paraphyletic Bipinnula with Geoblastapenicillata (Rchb.f.) Hoehne embedded, C. membrana-cea Lindl. and a clade encompassing all other Chloraeaspp. (BP < 50). Among these, three main cladesobtained low to high support. The first consists ofC. gaudichaudii Brongn., C. speciosa Poepp., C. gran-diflora Poepp. and C. magellanica Hook.f. (BP 100),
the second includes C. alpina Poepp., C. nudilabiaPoepp. and C. bicallosa Phil. ex Kraenzl., and the thirdgroup encompasses C. gavilu Lindl. to C. volkmanniPhil. ex Kraenzl (BP 100). The last group, in turn,includes two clades: C. gavilu to C. longipetala Lindl.(BP 95) and C. incisa Poepp. to C. volkmanni (Fig. 4B).
The combined plastid regions included 6700 char-acters, 1057 (16%) of them potentially informative toparsimony. Analysis recovered 20 000 MPTs with alength of 4436 steps, CI = 0.49 and RI = 0.74. One ofthe shortest trees and the strict consensus of the20 000 trees (with bootstrap values added) are shownin Figure 5. Overall relationships and patterns ofsupport are similar to those of the nrITS analysisexcept that, within Chloraeinae, relationships areless resolved. Nevertheless, the successive divergenceof C. cylindrostachya and C. reticulata and a cladeincluding the rest of the subtribe recovered in thenrITS analysis were also strongly supported in theplastid analysis. Moreover, some less-inclusive cladesfound in the nrITS analysis were also supported byplastid DNA, including Bipinnula (with Geoblastaembedded; BP 78), Gavilea minus G. australis(Skottsb.) M.N.Correa (BP 97), a clade composed ofC. gaudichaudii, C. speciosa, C. grandiflora andC. magellanica (BP 100) and a group consisting ofC. bicallosa and C. nudilabia, but with C. cuneataLindl. (not included in the nrITS analysis) instead ofC. alpina. The last species, C. membranacea and the
Table 2. Primers used for PCR and/or sequencing
Primer name Primer sequence of 5′ to 3′ Reference
ITS regionITS 5 GGAAGTAAAAGTCGTAACAAGG White et al. 1990ITS 4 TCCTCCGCTTATTGATATGC White et al. 1990
trnL-trnF regionc CGAAATCGGTAGACGCTA Taberlet et al. 1991d GGGGATAGAGGGACTTGAAC Taberlet et al. 1991e GGTTCAAGTCCCTCTATCCC Taberlet et al. 1991f ATTTGAACTGGTGACACGAG Taberlet et al. 1991
rbcL1F ATGGCAGAATTACAA(A/G)GA Kores et al. 19971360R CTTCACAAGCAGCAGCTAGTTC Kores et al. 1997636F GCGTTGGAGAGATCGTTTCT Muasya et al. 1998724R TCGCATGTACCYGCAGTTGC Muasya et al. 1998
matK-trnK region-19F CGTTCTGACCATATTGCACTATG Molvray, Kores & Chase 20002R AACTAGTCGGATGGAGTAG Steele & Vilgalys 19941309F GACTTTC(G/T)TGTGCTAGAACT Molvray et al. 2000556R GAAGRAACATCTTTKATCCA Molvray et al. 2000
rpoB-trnC IGSrpoB CKA CAA AAY CCY TCR AAT TG Shaw et al. 2005trnC CAC CCR GAT TYG AAC TGG GG Shaw et al. 2005
123456789
1011121314151617181920212223242526
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
boj_1200_
PHYLOGENY OF CHLORAEINAE 11
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

Fig
ure
4.P
hyl
ogen
etic
rela
tion
ship
sof
Ch
lora
ein
aefr
omth
epa
rsim
ony
anal
ysis
ofn
ucl
ear
ribo
som
alin
tern
altr
ansc
ribe
dsp
acer
(nrI
TS
)se
quen
ces.
A,
one
ofth
e25
4m
ost
pars
imon
iou
str
ees
(MP
Ts)
wit
hbr
anch
len
gth
sdr
awn
prop
orti
onal
toth
en
um
ber
ofch
ange
s.B
,st
rict
con
sen
sus
ofth
e25
4M
PTs
(nu
mbe
rsab
ove
bran
ches
are
boot
stra
ppr
opor
tion
s,n
otsh
own
wh
en<
50%
).
12
3
boj_1200_
12 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

Fig
ure
5.P
hyl
ogen
etic
rela
tion
ship
sof
Ch
lora
ein
aefr
omth
epa
rsim
ony
anal
ysis
ofco
mbi
ned
plas
tid
sequ
ence
s.A
,on
eof
the
2000
0m
ost
pars
imon
iou
str
ees
(MP
Ts)
wit
hbr
anch
len
gth
sdr
awn
prop
orti
onal
toth
en
um
ber
ofch
ange
s.B
,st
rict
con
sen
sus
ofth
e20
000
MP
Ts(n
um
bers
abov
ebr
anch
esar
ebo
otst
rap
prop
orti
ons,
not
show
nw
hen
<50
%).
12
3
boj_1200_
PHYLOGENY OF CHLORAEINAE 13
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

remaining species of Chloraea were part of a largepolytomy with the above-mentioned clades (Fig. 5B).
The combined data set of nuclear and plastid DNAsequences comprised 7475 aligned nucleotide posi-tions, of which 1454 (19%) were potentially parsimonyinformative. The heuristic search recovered 19 980MPTs with a length of 6600 steps, CI = 0.46 andRI = 0.70. The strict consensus of the 19 980 trees isshown in Figure 6A. Cranichideae sensu Chase et al.(2003) are strongly supported as monophyletic(BP 100) and they encompass two major clades, Chlo-raeinae (BP 100) and the rest of Cranichideae(BP 70). As in the nrITS analysis, none of the generaof Chloraeinae (excluding monospecific Geoblasta) ismonophyletic. Instead, Chloraea consists of a gradein which C. cylindrostachya and then C. reticulatadiverge first; the latter is sister to the rest of thesubtribe (BP 100). In the strict consensus, theremainder of Chloraeinae (BP 98) form a polytomyconsisting of C. membranacea and four stronglysupported, major clades (marked with numbers1–4 in Fig. 6): (1) [C. grandiflora–(C. gaudichaudii–(C. speciosa–C. magellanica))] (BP 100); (2) paraphyl-etic Bipinnula, including Geoblasta (BP 97); (3)paraphyletic Gavilea, with C. chica nested (BP 99);and (4) ‘core’ Chloraea clade including C. bicallosa toC. volkmanni (BP 94). The Gavilea/Geoblasta andBipinnula/C. chica clades are sister to each otherwith weak support (BP 70).
BAYESIAN ANALYSIS
The Bayesian summary tree is shown in Figure 6B.The overall relationships recovered by the Bayesiananalysis are similar to the strict consensus of theparsimony analysis but slightly more resolved. Anotable exception is the association of C. membrana-cea to the core Chloraea clade, which is stronglysupported (PP 0.98). Likewise, the sister-group rela-tionship between the Bipinnula/Geoblasta clade andthe Gavilea/C. chica clades received strong support inthis analysis (PP 1.00). No instances of strongly sup-ported, contradicting clades between the parsimonyand Bayesian analyses occurred (Fig. 6).
DISCUSSIONPHYLOGENETIC POSITION OF CHLORAEINAE
This study represents the first attempt to reconstructphylogenetic relationships in Chloraeinae by includ-ing all four genera and > 50% of their componentspecies with representatives of all the other subtribescurrently recognized in Cranichideae, plus several ofDiurideae. Monophyly of Chloraeinae sensu Pridgeonet al. (2003) is strongly supported, and our results areconsistent with previous findings by Kores et al.
(1997, 2000, 2001), Clements (1999), Clements et al.(2002) and Salazar et al. (2003) in showing thatChloraeinae could be accommodated in an expandedconcept of Cranichideae. There are a few putativelysynapormophic morphological characters supportingthe inclusion of Chloraeinae in Cranichideae, such asthe possession of fleshy roots either clustered or scat-tered along a rhizome, leaves usually arranged ina basal rosette and a spiranthoid embryo pattern(Clements, 1999).
Cranichideae s.l., including Chloraeinae, is sister toDiurideae, as noted previously in several molecularphylogenetic studies (Cameron et al., 1999; Koreset al., 2000, 2001). The various features of floralmorphology, column structure, exine morphologyand pollen organization shared by Chloraeinae andsome representatives of Diurideae probably repre-sent symplesiomorphies of the whole Diuridae/Cranichideae clade, and thus they do not support aparticularly close relationship between Chloraeinaeand various combinations of Diurideae, as believed bysome taxonomists (e.g. Brieger, 1974–1975; Dressler,1993; Szlachetko & Tukałło, 2008).
PHYLOGENETIC RELATIONSHIPS WITHIN
CHLORAEINAE
The present study shows that none of the genera ofChloraeinae that includes more than one species, i.e.Bipinnula, Chloraea and Gavilea, is monophyletic.Bipinnula and Gavilea are both embedded in Chlo-raea, the monospecific Geoblasta is nested in aparaphyletic Bipinnula and C. chica is embeddedin Gavilea (Fig. 6). This situation would probablyexplain the absence of morphological diagnostic char-acters for Chloraea (see earlier).
Chloraea cylindrostachya and C. reticulata are suc-cessive sisters to the rest of the subtribe. A similarresult was obtained by Chemisquy & Morrone (2010),whose analysis recovered C. cylindrostachya andC. praecincta Speg. & Kraenzl. (not sampled by us butbelonging to the same geographical group as C. re-ticulata) as successive sisters of all the other Chlo-raeinae. Chloraea cylindrostachya belongs in the‘western group’ and has a wide latitudinal distribu-tion in Chile and Argentina, ranging from 32°S to51°S and occurring at intermediate elevations (1000–2000 m) in the Andean and Coastal Chilean cordille-ras. Therefore, together with C. chica, this is one ofthe species of Chloraea having the largest distribu-tion range. Chloraea cylindrostachya shows some dis-tinctive features, such as a leafy stem, a completelyfleshy labellum and a hood or galea formed by thedorsal sepal and the petals (Correa, 1969; Novoaet al., 2006; Elórtegui & Novoa, 2009). Chloraea cylin-drostachya and C. leptopetala Reiche (not included in
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
boj_1200_
14 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
5 5

Fig
ure
6.P
hyl
ogen
etic
rela
tion
ship
sof
Ch
lora
ein
aein
ferr
edfr
omco
mbi
ned
anal
ysis
ofpl
asti
dan
dn
ucl
ear
ribo
som
alin
tern
altr
ansc
ribe
dsp
acer
(nrI
TS
)se
quen
ces.
A,
stri
ctco
nse
nsu
sof
the
1998
0m
ost
pars
imon
iou
str
ees
(MP
Ts)
fou
nd
byth
epa
rsim
ony
anal
ysis
(nu
mbe
rsab
ove
bran
ches
are
boot
stra
ppr
opor
tion
s,n
otsh
own
wh
en<
50%
).B
,B
ayes
ian
sum
mar
ytr
ee(n
um
bers
abov
ebr
anch
esar
epo
ster
ior
prob
abil
itie
s).
Th
em
ain
clad
esdi
scu
ssed
inth
ete
xtar
ein
dica
ted
byn
um
bers
1–4.
12
3 4
boj_1200_
PHYLOGENY OF CHLORAEINAE 15
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••

our analyses) are the only species of the ‘westerngroup’ that show these features. The presence ofcauline leaves and flowers with reticulate veining isshared with the ‘northern’ Chloraea spp. of north-western Argentina, Bolivia and Peru (Correa, 1969).Chloraea reticulata belongs to this last group, and thepresence of cauline leaves could represent a simple-siomorphy in Chloraeinae. Szlachetko & Tukałło(2008) proposed the new monotypic genus, Correor-chis Szlach., for Chloraea cylindrostachya, whichmight seem justifiable on phylogenetic grounds.However, C. reticulata and C. praecincta also divergein our phylogenetic trees and in those of Chemisquy& Morrone (2010), respectively, prior to the mainradiation of Chloraeinae. If the same criterion appliedin recognizing Correorchis were applied to thesespecies, additional monospecific genera would berequired to reflect their phylogenetic position. Never-theless, we urge taxonomists to refrain from rushingto propose further taxonomic changes until a clearerpicture of the phylogenetic relationships near thebase of the Chloraeinae tree is obtained.
The remaining members of Chloraeinae form astrongly supported clade, which in turn consists offour strongly supported monophyletic groups (1–4in Fig. 6). Clade 1, which includes C. grandiflora,C. gaudichaudii and (C. speciosa–C. magellanica) iseasily identifiable by the reticulate veining of thefloral segments, presence of a hood formed by thedorsal sepal and the petals and entire labellumadorned with clavate calli and with fleshy apex. Allthese species are structurally similar, except in thatthe labellum of C. grandiflora is densely covered byfleshy warts (Correa, 1969). Szlachetko & Tukałło(2008) transferred C. grandiflora to Ulantha Hook.,as U. grandiflora (Poepp.) Szlach., apparently over-looking the fact that the prior combination Ulanthagrandiflora Hook. (type of the genus) implies that hisnew combination created a later homonym. They alsoplaced in Ulantha the species here treated as Bipin-nula apinnula, which, in view of our results, turnsUlantha polyphyletic (see discussion later regardingthe phylogenetic position of B. apinnula).
Clades 2 and 3 were recovered as sisters to eachother and this relationship obtained low bootstrapsupport (BP 70) but a high posterior probability(PP 1.00). Clade 2 is composed of the four Bipinnulaspp. analysed, among which the monotypic genusGeoblasta is nested. Gosewijn (1993) recognized threesections in Bipinnula, all which are represented inour taxonomic sample. Bipinnula fimbriata (Poepp.)I.M.Johnst. (section Multiflorae Gosewijn) is sister toa clade that in turn includes two subclades. The firstof these consists of G. penicillata and B. montanaArechav. (section Bipinnula) and the second includesB. apinnula and B. volkmanni Kraenzl. (section Tri-
lobatae Gosewijn). All these groups obtained strongsupport. Our results fully corroborate the hypothesisput forward by Gosewijn (1993) regarding a closephylogenetic relationship between B. apinnula andB. volkmanni. In contrast, Szlachetko & Tukałło(2008) removed B. apinnula from Bipinnula to placeit in polyphyletic Ulantha (see earlier). Likewise,Szlachetko & Margonska (2001), based on intuitiveassessments of floral characters, speculated thatBipinnula ‘is polyphyletic and consists of two appar-ently unrelated groups.’ They then proposed the newgenus Jouyella Szlach. to accommodate the speciespreviously included in Gosewijn’s (1993) sectionMultiflorae, considering the basal, rosulate leavesthat are present at flowering, the multi-flowered inflo-rescence, arching, shortly pedicellate flowers, the thinlabellum covered by numerous clavate thickening andthe thin sepals as ‘sufficient’ differences to recognizetwo genera. However, their genus Jouyella is identicalin circumscription to Gosewijn’s section Multiflorae,which is the closest relative of the other members ofBipinnula (plus Geoblasta penicillata). Therefore, wedo not see the advantage of inflating nomenclaturewith further genera for which monophyly has notbeen formally tested and we consider Jouyella as asynonym of Bipinnula.
From a geographical standpoint, Bipinnula com-prises three disjunct groups, which are correlatedwith both the sections proposed by Gosewijn (1993)and our molecular results. The first group corre-sponds to section Multiflorae (see above for morpho-logical details). It is composed of species endemic toChile and mainly restricted to coastal areas andlowland valleys in northern and central Chile. Thesecond group matches section Trilobatae and includestwo species endemic to the Andes of south-centralChile between 35°S and 37°S and is restricted tointermediate elevations (1500–2000 m; Novoa et al.,2006). Both species share a few-flowered inflorescence(character intermediate between sections Bipinnulaand Multiflorae) and a trilobate labellum fullycovered by colourful appendages and warts. The lastgroup, section Bipinnula, consists of one-floweredspecies (see later for details) from the Río de la Platacoastal region in eastern Argentina, Uruguay andsouthern Brazil (Izaguirre, 1973).
As already mentioned, our analyses recover Bipin-nula as paraphyletic, but monophyly would beachieved by transferring Geoblasta penicillata toBipinnula. In our analysis, this species is sister toB. montana from Uruguay, which occurs in the samepart of South America, although it not nearly aswidespread, as G. penicillata. Correa (1968) rein-stated the monospecific genus Geoblasta, distinguish-ing it from Chloraea based on characters of thelabellum and column and suggested that Geoblasta is
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
boj_1200_
16 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
6 6
7 7
88

more closely related to Bipinnula than to Chloraea,which is in agreement with our results. The species ofBipinnula section Bipinnula share several featureswith G. penicillata, including the absence of nectarif-erous channels, wingless column, one-flowered inflo-rescence, insect-like labellum (in most of the species)and similar geographical distribution. The phyloge-netic position of G. penicillata has to be reassessedwhen more single-flowered Bipinnula spp. becomeavailable for molecular study.
Clade 3 encompasses all the sampled species ofGavilea, with C. chica nested among them and thepair C. chica/G. australis obtained strong supportin our combined parsimony and Bayesian ana-lyses (BP 99, PP 1.00) (Fig. 6). Thus, contrary toChemisquy & Morrone (2010), our results do notsupport the monophyly of Gavilea, although this dis-crepancy might have resulted from differences in thetaxonomic sampling between the two studies. In ouranalyses, C. chica is consistently placed as the sisterof G. australis, a species not included in the analysisof Chemisquy & Morrone (2010). Although the place-ment of C. chica in the Gavilea clade might seemunexpected at a first glance, C. chica is able of propa-gating itself vegetatively by producing new plantsfrom creeping rhizomes (Fig. 1A). This feature is alsofound at least in G. araucana (Phil.) M.N.Correa(Fig. 1B) and is otherwise unknown in Chloraeinae.The species pair C. chica/G. australis is in turn sisterto the rest of Gavilea.
All other Chloraea spp. are grouped in clade 4 and,in our Bayesian analysis, C. membranacea is thesister of all the others (Fig. 6). Chloraea membrana-cea is a member of the ‘eastern’ group, together withC. bella Hauman (not sampled for this study). Thisgroup occurs in southern Brazil and adjacent Argen-tina, differing from other Chloraea in the straightcolumn wings broader near the column apex and thestigma longer than two-thirds of the column length.The remaining species includes the type species ofChloraea, C. virescens (Willd.) Lindl., and therefore itmight be referred to as ‘core Chloraea’. This group ischaracterized by the membranaceous flowers, longi-tudinal veining (except C. prodigiosa Rchb.f.) insepals and petals and entire to three-lobed labellumusually adorned with several keels or longitudinalrows of laminar or thickened excrescences. Thespecies of this clade display a high degree of morpho-logical variation, and species delimitation within thislineage is particularly problematic (Correa, 1969).Two major subclades were recovered within thisgroup. The first subclade includes the Chileanendemic species C. bicallosa, C. cuneata and C. nudi-labia (Novoa et al., 2006). These species have basalleaves that do not form a rosette and are narrow withacute apex and sometimes spathulate. The species of
this subclade are frequently found living in forests ofAraucaria araucana (Molina) C.Koch and speciesof Nothofagus Blume, which are restricted to inter-mediate elevations (1000–2000 m) in the southernAndean and Coastal Chilean cordilleras. Further-more, C. nudilabia is characterized by the presence ofa type C peloria (Mondragón-Palomino & Theißen,2009), i.e. having a labellum similar in shape, sizeand colouration to rest of the floral segments, givingthe flowers a similar appearance to that of the Aus-tralasian diurid genus Thelymitra T.Forst. & G.Forst.Mondragón-Palomino & Theißen (2009) pointed outthe possibility of independent occurrences of rareactinomorphic-like species within zygomorphic groupsof subfamily Orchidoideae. It is likely that autogamyand cleistogamy, frequent in groups with these flowercharacteristics, have contributed to the developmentof stable ‘prospecies’ (Rudall & Bateman, 2003).However, pollination data are available for only a fewChloraea spp. and the species studied so far are allself-compatible and allogamous (Humaña, Cisternas& Valdivia, 2008). According to our results, none ofthose allogamous species (C. bletioides Lindl., C. chry-santha Poepp., C. crispa Lindl. and C. galeata Lindl.)is closely related to C. nudilabia.
The second subclade of core Chloraea includes, forthe most part, species endemic to Chile, and they arerestricted to coastal or lowland valleys (Novoa et al.,2006). Genetic differences between the species in thisgroup are extremely low, this resulting in a lack ofsupported resolution for the relationships amongspecies of core Chloraea, and is suggestive of a rela-tively recent diversification of this lineage of Chloraeain western South America. The greatest concentrationand diversity of Chloraea and Gavilea occur in theAndean cordillera (Correa & Sánchez, 2003), andAndean orogeny may have played a role in promotingvicariant speciation events that resulted in the dis-parity in species diversity of this group between theeastern and western sides of the Andes.
Our analyses clearly show that, as currently delim-ited, Chloraea is polyphyletic and the generic limits inthe whole subtribe Chloraeinae are in urgent need ofrevision. Recently, several changes in the circumscrip-tion of the genera have been proposed, notably bySzlachetko and co-workers (Szlachetko & Margonska,2001; Szlachetko & Tukałło, 2008). These authorsresurrected Bieneria Rchb.f and Ulantha, and createdthe new genera Jouyella, Chileorchis Szlach. andCorreorchis, in addition to making various transfersof species between genera. However, as noted earlier,Ulantha is polyphyletic, Jouyella is arguably super-fluous, as its content is identical to section Multifloraeof Bipinnula and what remains of both Bipinnulaand Chloraea if those genera are accepted are non-monophyletic assemblages of species. We have not
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
boj_1200_
PHYLOGENY OF CHLORAEINAE 17
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
99
1010

been able to test the monophyly and phylogeneticposition of Bieneria because no species assigned tothat genus by Szlachetko & Tukałło (2008) have beenavailable for molecular study. Several of their genericconcepts have been based on unreliable characters,such as the degree of lobulation and ornamentationof the labellum (e.g. Ulantha sensu Szlachetko &Tukałło, 2008), and it is worth noting that the delimi-tation of ‘Chloraeinae’ of Szlachetko and co-workers(Szlachetko, 1995; Szlachetko & Rutkowski, 2000;Szlachetko & Tukałło, 2008) represents a grossly poly-phyletic mixture of genera that, according to severalphylogenetic studies, are dispersed among at leastthree distinct tribes, namely Codonorchideae, Cran-ichideae and Diurideae (e.g. Kores et al., 2001; Clem-ents et al., 2002).
Our study does not support the circumscriptions ofChloraea proposed either by Correa (1969), Correa &Sánchez (2003) or Szlachetko & Tukałło (2008; seeearlier). Various sections have been proposed withinChloraea based on combinations of characters, suchas number of flowers per inflorescence, presence/absence and shape of calli, crests and warts on thelabellum and type of veining of the floral segments(e.g. Kraenzlin, 1904; Reiche, 1910), but none of thosesections turns out to be monophyletic. Regarding thegeographical groups proposed by Hauman (1922) andCorrea (1969), it is clear that the ‘western’ group doesnot correspond to clades recovered in our analyses.For instance, C. cylindrostachya, a member of the‘western’ group, is sister to the rest of Chloraeinae,and thus it is not associated with other members ofthat group, such as C. disoides Lindl. and C. philippiiRchb.f. Only one species of the ‘eastern’ and ‘northern’groups was analysed here (C. membranacea andC. reticulata, respectively), so we are unable at thistime to draw conclusions concerning whether thespecies included in these groups by previous authorsare closely related to one another or not, but overallour results indicate that there is no clear correlationbetween the clades and the geographical groups.
The present study has increased considerably thesample of both species and characters analysed pre-viously (Chemisquy & Morrone, 2010). However,inclusion in future analyses of further Chloraea spp.,especially from Peru and northern Argentina, and ofBipinnula spp. from Uruguay and Argentina, willhelp to improve our understanding of the phylo-genetic relationships and the taxonomic limits inChloraeinae.
ACKNOWLEDGEMENTS
The authors thank Laura Márquez Valdelamar (Labo-ratorio de Biología Molecular, Instituto de Biología,Universidad Nacional Autónoma de México) for
assistance with DNA sequencing; Santiago Benitez-Vieyra andAndrea Cocucci (Universidad Nacional deCórdoba, Argentina) and Orfeo Crosa (Universidad dela República de Uruguay) for plant material and fieldassistance; and the PUCV-UCONC and FONDEFD06I1079 projects for providing financial support.
REFERENCES
Ackerman JD, Williams NH. 1981. Pollen morphology ofthe Chloraeinae (Orchidaceae: Diurideae) and related sub-tribes. American Journal of Botany 68: 1392–1402.
Barriel V, Tassy P. 1998. Rooting with multiple outgroups:consensus versus parsimony. Cladistics 14: 193–200.
van den Berg C, Goldman DH, Freudenstein JV, Prid-geon AM, Cameron KM, Chase MW. 2005. An overviewof the phylogenetic relationships within Epidendroideaeinferred from multiple DNA regions and recircumscriptionof Epidendreae and Arethuseae (Orchidaceae). AmericanJournal of Botany 92: 613–624.
van den Berg C, Higgins WE, Dressler RL, Whitten WM,Soto MA, Culham A, Chase MW. 2000. A phylogeneticanalysis of Laeliinae (Orchidaceae) based on sequence datafrom nuclear transcribed spacers (ITS) of nuclear ribosomalDNA. Lindleyana 15: 96–114.
Brieger FG. 1974–1975. Unterfamilie: Neottioideae. In:Brieger FG, Maatsch R, Senghas K, Schlechter R, eds. DieOrchideen. Berlin: Paul Parey, ••–••.
Burns-Balogh P, Funk V. 1986. A phylogenetic analysis ofthe Orchidaceae. Smithsonian Contributions to Botany 61.Washington, DC: Smithsonian Institution.
Cameron KM, Whitten WM, Kores PJ, Jarrell DC, AlbertVA, Yukawa T, Hills HG, Goldman DH. 1999. A phyloge-netics analysis of the Orchidaceae: evidence from rbcL nucle-otide sequences. American Journal of Botany 86: 208–224.
Chase MW. 2003. Phylogenetics of Chloraeeae. In: PridgeonAM, Cribb PJ, Chase MW, Rasmussen FN, eds. Generaorchidacearum, vol. 3. Orchidoideae (part two). Oxford:Oxford University Press, ••–••.
Chase MW, Cameron KM, Barret RL, Freudenstein JV.2003. DNA data and Orchidaceae systematics: a new phy-logenetic classifications. In: Dixon KW, Kell SP, Barret RL,Cribb PJ, eds. Orchid conservation. Kota Kinabalu: NaturalHistory Publications (Borneo), ••–••.
Chemisquy MA, Morrone O. 2010. Phylogenetic analysis ofthe subtribe Chloraeinae (Orchidaceae): a preliminaryapproach based on three chloroplast markers. AustralianJournal of Botany 23: 38–46.
Chiron G, Oliveira RP, Santos TM, Bellvert F, BertrandC, Van den Berg C. 2009. Phylogeny and evolution ofBaptistonia (Orchidaceae, Oncidiinae) based on molecularanalyses, morphology and floral oil evidences. Plant System-atics and Evolution 281: 35–49.
Ciotek L, Giorgis P, Benitez-Vieyra S, Cocucci AA. 2006.First confirmed case of pseudocopulation in terrestrialorchids of South America: pollination of Geoblastapennicillata (Orchidaceae) by Campsomeris bistrimacula(Hymenoptera, Scoliidae). Flora 201: 365–369.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
646566676869707172737475767778798081828384858687888990919293949596979899
100101102103104105106107108109110111112
boj_1200_
18 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
1111 12
1313
1414
1515

Clements MA. 1999. Embryology. In: Pridgeon AM, Cribb PJ,Chase MW, Rasmussen FN, eds. Genera orchidacearum,vol. 1. General introduction, Apostasioideae, Cypripedio-ideae. Oxford: Oxford University Press, ••–••.
Clements MA, Jones DL, Sharma IK, Nightingale ME,Garratt MJ, Fitzgerald KJ, Mackenzie AM, Molloy BP.2002. Phylogenetics of Diurideae (Orchidaceae) based onthe internal transcribed spacer (ITS) regions of nuclearribosomal DNA. Lindleyana 17: 135–171.
Correa MN. 1956. Las especies argentinas del géneroGavilea. Boletín de la Sociedad Argentina de Botánica 6:73–86.
Correa MN. 1968. Rehabilitación del género Geoblasta Barb.Rodr. Revista del Museo de la Plata, Sección Botánica 11:69–74.
Correa MN. 1969. Chloraea, género sudamericano de Orchi-daceae. Darwiniana 15: 374–500.
Correa MN. 2003. Chloraeeae (key to the genera). In: Prid-geon AM, Cribb PJ, Chase MW, Rasmussen FN, eds. Gen-eraorchidacearum, vol. 3. Orchidoideae (part two). Oxford:Oxford University Press, ••–••.
Correa MN, Sánchez M. 2003. Chloraeeae. In: Pridgeon AM,Cribb PJ, Chase MW, Rasmussen FN, eds. Genera orchi-dacearum, vol. 3. Orchidoideae (part two). Oxford: OxfordUniversity Press, ••–••.
Cribb PJ, Kores PJ. 2000. The systematic position of Codo-norchis (Orchidaceae: Orchidoideae). Lindleyana 15: 169–170.
Douzery E, Pridgeon AM, Kores PJ, Kurzweil H, LinderP, Chase MW. 1999. Molecular phylogenetics of Diseae(Orchidaceae): a contribution from nuclear ribosomal ITSsequences. American Journal of Botany 86: 887–899.
Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedurefor small quantities of fresh leaf tissue. PhytochemistryBulletin, Botanical Society of America 19: 11–15.
Dressler RL. 1981. The orchids: natural history and classi-fication. Cambridge: Harvard University Press.
Dressler RL. 1993. Phylogeny and classification of the orchidfamily. Portland, OR: Dioscorides Press.
Dressler RL, Dodson CH. 1960. Classification and phylog-eny in the Orchidaceae. Annals of the Missouri BotanicalGarden 47: 25–68.
Elórtegui S, Novoa P. 2009. Orquídeas de la región deValparaíso. Viña del Mar: Taller La Era.
Felsenstein J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39: 783–791.
Figueroa C, Salazar GA, Zavaleta HA, Engleman EM.2008. Root character evolution and systematics in Cran-ichideae, Prescottiinae and Spiranthinae (Orchidaceae,Cranichideae). Annals of Botany 101: 509–520.
Fitch WM. 1971. Toward defining the course of evolution:minimum change for a specific tree topology. SystematicZoology 20: 406–416.
Gosewijn WJN. 1993. Descubrimiento de una sorprendenteorquídea en Talca. Gayana Botanica 50: 11–16.
Gravendeel B, Chase MW, de Vogel EF, Roos MC, MesTHM, Bachmann K. 2001. Molecular phylogeny of Coel-ogyne (Epidendroideae; Orchidaceae) based on plastid
RFLPs, matK, and nuclear ribosomal ITS sequences: evi-dence for polyphyly. American Journal of Botany 88: 1915–1927.
Hauman L. 1922. La distribución geográfica del género Chlo-raea. Physis 4: 293–295.
Huelsenbeck JP, Ronquist F. 2001. MrBAYES: Bayesianinference of phylogenetic trees. Bioinformatics 17: 754–755.
Humaña AM, Cisternas MA, Valdivia CE. 2008. Breedingsystem and pollination of selected orchids of Chloraea genusfrom central Chile. Flora 203: 469–473.
Izaguirre P. 1973. Las especies uruguayas de Bipinnula.Boletín Sociedad Argentina de Botánica 15: 261–276.
Jones DL, Clements MA, Sharma IK, Mackenzie AM,Molloy BPJ. 2002. Nomenclatural notes arising fromstudies into the tribe Diurideae (Orchidaceae). Orchadian13: 437–468.
Katoh K, Asimenos G, Toh H. 2009. Multiple alignment ofDNA sequences with MAFFT. In: Posada D, ed. Bioinfor-matics for DNA sequence analysis. Methods in MolecularBiology 537. New York: Humana Press, ••–••.
Katoh K, Kuma K, Toh H, Miyata T. 2005. MAFFT ver-sion 5: improvement in accuracy of multiple sequence align-ment. Nucleic Acids Research 33: 511–518.
Kores PJ, Cameron KM, Molvray M, Chase MW. 1997.The phylogenetic relationships of Orchidoideae and Spiran-thoideae (Orchidaceae) as inferred from rbcL plastidsequences. Lindleyana 12: 1–11.
Kores PJ, Molvray M, Weston PH, Hopper SD, BrownAP, Cameron KM, Chase MW. 2001. A phylogeneticanalysis of Diurideae (Orchidaceae) based on plastid DNAsequence data. American Journal of Botany 88: 1903–1914.
Kores PJ, Weston PH, Molvray M, Chase MW. 2000.Phylogenetic relationships within the Diurideae (Orchi-daceae): inferences from plastid matK sequences. In: WilsonKL, Morrison DA, eds. Monocots: systematics and evolution.Melbourne: CSIRO, ••–••.
Kraenzlin F. 1904. Orchidacearum genera et species, vol. 2,part 1. Berlin: Bayer & Müller.
Lavarack P. 1976. The taxonomic affinities of the AustralianNeottioideae. Taxon 25: 289–296.
Molvray M, Kores PJ, Chase MW. 2000. Polyphyly ofmycoheterotrophic orchids and functional influences onfloral and molecular characters. In: Wilson KL, MorrisonDA, eds. Monocots: systematics and evolution. Melbourne:CSIRO, ••–••.
Mondragón-Palomino M, Theißen G. 2009. Why are orchidflowers so diverse? Reduction of evolutionary constraints byparalogues of class B floral homeotic genes. Annals ofBotany 104: 583–594.
Monteiro SHN, Selbach-Schnadelbach A, de OliveriraRP, van den Berg C. 2010. Molecular phylogeneticsof Galeandra (Orchidaceae: Catasetinae) based on plastidand nuclear DNA sequences. Systematic Botany 35: 476–486.
Muasya AM, Simpson DA, Culham A, Chase MW. 1998.An assessment of suprageneric phylogeny in Cyperaceaeusing rbcL sequences. Plant Systematics and Evolution 211:257–271.
123456789
10111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455565758
5960616263646566676869707172737475767778798081828384858687888990919293949596979899
100101102103104105106107108109110111112113114115116
boj_1200_
PHYLOGENY OF CHLORAEINAE 19
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
16 16
17 17
18 18
19 19
20 20
21 21
2222
2323
2424
2525
2626

Novoa P, Espejo J, Cisternas M, Domínguez E. 2006.Guía de campo de las orquídeas chilenas. Concepción:CORMA.
Posada D, Crandall KA. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14: 817–818.
Pridgeon AM, Bateman RM, Cox AV, Hapeman JR,Chase MW. 1997. Phylogenetics of subtribe Orchidinae(Orchidoideae, Orchidaceae) based on nuclear ITS sequ-ences. 1. Intergeneric relationships and polyphyly of Orchissensu lato. Lindleyana 12: 89–109.
Pridgeon AM, Chase MW. 1995. Subterranean axes in tribeDiurideae (Orchidaceae): morphology, anatomy, and system-atic significance. American Journal of Botany 82: 1473–1495.
Pridgeon AM, Cribb PJ, Chase MW, Rasmussen FN.2001. Genera orchidacearum vol. 2: Orchidoideae part 1.Oxford: Oxford University Press.
Pridgeon AM, Cribb PJ, Chase MW, Rasmussen FN.2003. Genera orchidacearum vol. 3: Orchidoideae part 2,Vanilloideae. Oxford: Oxford University Press.
Rasmussen FN. 1982. The gynostemium of the neottioidorchids. Opera Botanica 65: 1–96.
Rasmussen FN. 1985. Orchids. In: Dahlgren RMT, CliffordHT, Yeo PF, eds. The families of the monocotyledons: struc-ture, evolution and taxonomy. Berlin: Springer, ••–••.
Reiche K. 1910. Orchidaceae chilenses: ensayo de unamonografía de las orquídeas de Chile. Anales del MuseoNacional de Chile, Segunda Sección, Botánica 18: 1–88.
Ronquist F, Huelsenbeck JP, Van der Mark P. 2005.MrBayes 3.1 manual, draft 5/17/2005. Available at: http://morphbank.ebc.uu.se/mrbayes/
Rudall P, Bateman R. 2003. Evolutionary change in flowersand inflorescences: evidence from naturally occurringterata. Trends in Plant Science 2: 76–82.
Salazar GA, Cabrera LI, Madriñán S, Chase MW. 2009.Phylogenetic relationships of Cranichidinae and Prescotti-nae (Orchidaceae, Cranichideae) inferred from plastid andnuclear DNA sequences. Annals of Botany 104: 403–416.
Salazar GA, Chase MW, Soto MA, Ingrouille M. 2003.Phylogenetics of Cranichideae with emphasis on
Spiranthinae (Orchidaceae, Orchidoideae): evidence fromplastid and nuclear DNA sequences. American Journal ofBotany 90: 777–795.
Shaw J, Lickey EB, Beck JT, Farmer SB, Liu W, MillerJ, Siripun KC, Winder CT, Schilling EE, Small RL.2005. The tortoise and the hare II: relative utility of 21noncoding chloroplast DNA sequences for phylogeneticanalysis. American Journal of Botany 92: 142–166.
Steele KP, Vilgalys R. 1994. Phylogenetic analyses of thePolemoniaceae using nucleotide sequences of the plastidgene matK. Systematic Botany 19: 126–142.
Swofford DL. 2002. PAUP*. Phylogenetic analysis using par-simony (*and other methods). Version 4. Sunderland, MA:Sinauer.
Szlachetko DL. 1991. Thelymitroideae, a new subfamilywithin Orchidaceae. Fragmenta Floristica et Geobotanica36: 33–49.
Szlachetko DL. 1995. Systema orchidalium. Fragmenta Flo-ristica et Geobotanica 3 (Suppl): 1–152.
Szlachetko DL, Margonska HB. 2001. Genera et speciesorchidalium 4. Polish Botanical Journal 46: 123–125.
Szlachetko DL, Rutkowski P. 2000. Gynostemia orchida-lium I. Apostasiaceae, Cypripediaceae, Orchidaceae (The-lymitroideae, Orchidoideae, Tropidioideae, Spiranthoideae,Neottioideae, Vanilloideae). Acta Botanica Fennica 169:1–379.
Szlachetko DL, Tukałło P. 2008. Notes on the subtribeChloraeinae (Orchidaceae). Acta Societatis BotanicorumPoloniae 77: 111–116.
Taberlet P, Gielly L, Pautou G, Bouvet J. 1991. Universalprimers for amplification of three non-coding regions ofchloroplast DNA. Plant Molecular Biology 17: 1105–1109.
White T, Bruns JT, Lee S, Taylor J. 1990. Amplificationand direct sequencing of fungal ribosomal RNA genes forphylogenetics. In: Innis M, Gelfand D, Sninsky J, White TJ,eds. PCR protocols: a guide to methods and applications.San Diego, CA: Academic Press, ••–••.
123456789
101112131415161718192021222324252627282930313233343536373839
404142434445464748495051525354555657585960616263646566676869707172737475767778
boj_1200_
20 M. A. CISTERNAS ET AL.
© 2011 The Linnean Society of London, Botanical Journal of the Linnean Society, 2011, ••, ••–••
27 27
28 28
29 29
30 30
3131

AUTHOR QUERY FORM
Dear Author,During the preparation of your manuscript for publication, the questions listed below have arisen. Please
attend to these matters and return this form with your proof.Many thanks for your assistance.
QueryReferences
Query Remark
q1 AUTHOR: Dressler, 1974 has not been included in the Reference List, pleasesupply full publication details.
q2 AUTHOR: On the one hand – this phrase has been inserted here to balancethe use of On the other hand later in the paragraph. Please check that this isthe correct place to insert this phrase and amend if necessary
q3 AUTHOR: a closing bracket was missing and has been inserted here. Is this thecorrect place?
q4 AUTHOR: Álvarez-Molina & Cameron, 2009 has not been included in theReference List, please supply full publication details.
q5 AUTHOR: the brackets have been changed to curved brackets enclosed bysquare brackets – is that OK? Also, are the en-rules correct or should they behyphens?
q6 AUTHOR: Chemisquy & Morrone (2009 has been changed to Chemisquy &Morrone (2010 so that this citation matches the Reference List. Please confirmthat this is correct.
q7 AUTHOR: the square brackets have been changed to curved – is that OK?Also, is this en-rule OK or should this be a hyphen?
q8 AUTHOR: Szlachetko & Margonska (2001b has been changed to Szlachetko &Margonska (2001 so that this citation matches the Reference List. Pleaseconfirm that this is correct.
q9 AUTHOR: Mondragón-Palomino & Theißen, 2008 has been changed toMondragón-Palomino & Theißen, 2009 so that this citation matches theReference List. Please confirm that this is correct.
q10 AUTHOR: Mondragón-Palomino & Theißen (2008) has been changed toMondragón-Palomino & Theißen (2009) so that this citation matches theReference List. Please confirm that this is correct.
q11 AUTHOR: Please confirm the editor Schlechter R is correct for Brieger;1974–1975.
q12 AUTHOR: Please supply page range for Reference Brieger 1974–1975.
q13 AUTHOR: Chase, 2003 has not been cited in the text. Please indicate where itshould be cited; or delete from the Reference List.
q14 AUTHOR: Please supply page range for Reference Chase 2003.
q15 AUTHOR: Please supply page range for Reference Chase, Cameron, Barret,Freudenstein 2003.
Toppan Best-set Premedia LimitedJournal Code: BOJ Proofreader: ElsieArticle No: 1200 Page Extent: 20

q16 AUTHOR: Please supply page range for Reference Clements 1999.
q17 AUTHOR: Please supply page range for Reference Correa 2003.
q18 AUTHOR: Please supply page range for Reference Correa, Sánchez 2003.
q19 AUTHOR: Douzery, Pridgeon, Kores, Kurzweil, Linder, Chase, 1999 has notbeen cited in the text. Please indicate where it should be cited; or delete fromthe Reference List.
q20 AUTHOR: Dressler, Dodson, 1960 has not been cited in the text. Pleaseindicate where it should be cited; or delete from the Reference List.
q21 AUTHOR: Figueroa, Salazar, Zavaleta, Engleman, 2008 has not been cited inthe text. Please indicate where it should be cited; or delete from the ReferenceList.
q22 AUTHOR: Huelsenbeck, Ronquist, 2001 has not been cited in the text. Pleaseindicate where it should be cited; or delete from the Reference List.
q23 AUTHOR: Please supply page range for Reference Katoh, Asimenos, Toh 2009.
q24 AUTHOR: Please supply page range for Reference Kores, Weston, Molvray,Chase 2000.
q25 AUTHOR: Lavarack, 1976 has not been cited in the text. Please indicate whereit should be cited; or delete from the Reference List.
q26 AUTHOR: Please supply page range for Reference Molvray, Kores, Chase2000.
q27 AUTHOR: Pridgeon, Bateman, Cox, Hapeman, Chase, 1997 has not been citedin the text. Please indicate where it should be cited; or delete from theReference List.
q28 AUTHOR: Pridgeon, Cribb, Chase, Rasmussen, 2001 has not been cited in thetext. Please indicate where it should be cited; or delete from the Reference List.
q29 AUTHOR: Please supply page range for Reference Rasmussen 1985.
q30 AUTHOR: Please check this website address and confirm that it is correct forRonquist, Huelsenbeck, Van der Mark; 2005. (Please note that it is theresponsibility of the author(s) to ensure that all URLs given in this article arecorrect and useable.)
q31 AUTHOR: Please supply page range for Reference White, Bruns, Lee, Taylor1990.

USING e-ANNOTATION TOOLS FOR ELECTRONIC PROOF CORRECTION
Required software to e-Annotate PDFs: Adobe Acrobat Professional or Adobe Reader (version 8.0 or
above). (Note that this document uses screenshots from Adobe Reader X)
The latest version of Acrobat Reader can be downloaded for free at: http://get.adobe.com/reader/
Once you have Acrobat Reader open on your computer, click on the Comment tab at the right of the toolbar:
1. Replace (Ins) Tool – for replacing text.
Strikes a line through text and opens up a text
box where replacement text can be entered.
How to use it
Highlight a word or sentence.
Click on the Replace (Ins) icon in the Annotations
section.
Type the replacement text into the blue box that
appears.
This will open up a panel down the right side of the document. The majority of
tools you will use for annotating your proof will be in the Annotations section,
pictured opposite. We’ve picked out some of these tools below:
2. Strikethrough (Del) Tool – for deleting text.
Strikes a red line through text that is to be
deleted.
How to use it
Highlight a word or sentence.
Click on the Strikethrough (Del) icon in the
Annotations section.
3. Add note to text Tool – for highlighting a section
to be changed to bold or italic.
Highlights text in yellow and opens up a text
box where comments can be entered.
How to use it
Highlight the relevant section of text.
Click on the Add note to text icon in the
Annotations section.
Type instruction on what should be changed
regarding the text into the yellow box that
appears.
4. Add sticky note Tool – for making notes at
specific points in the text.
Marks a point in the proof where a comment
needs to be highlighted.
How to use it
Click on the Add sticky note icon in the
Annotations section.
Click at the point in the proof where the comment
should be inserted.
Type the comment into the yellow box that
appears.

USING e-ANNOTATION TOOLS FOR ELECTRONIC PROOF CORRECTION
For further information on how to annotate proofs, click on the Help menu to reveal a list of further options:
5. Attach File Tool – for inserting large amounts of
text or replacement figures.
Inserts an icon linking to the attached file in the
appropriate pace in the text.
How to use it
Click on the Attach File icon in the Annotations
section.
Click on the proof to where you’d like the attached
file to be linked.
Select the file to be attached from your computer
or network.
Select the colour and type of icon that will appear
in the proof. Click OK.
6. Add stamp Tool – for approving a proof if no
corrections are required.
Inserts a selected stamp onto an appropriate
place in the proof.
How to use it
Click on the Add stamp icon in the Annotations
section.
Select the stamp you want to use. (The Approved
stamp is usually available directly in the menu that
appears).
Click on the proof where you’d like the stamp to
appear. (Where a proof is to be approved as it is,
this would normally be on the first page).
7. Drawing Markups Tools – for drawing shapes, lines and freeform
annotations on proofs and commenting on these marks.
Allows shapes, lines and freeform annotations to be drawn on proofs and for
comment to be made on these marks..
How to use it
Click on one of the shapes in the Drawing
Markups section.
Click on the proof at the relevant point and
draw the selected shape with the cursor.
To add a comment to the drawn shape,
move the cursor over the shape until an
arrowhead appears.
Double click on the shape and type any
text in the red box that appears.
Copyright © 2022 FDOKUMEN




















