
Salinity regulates claudin mRNA and protein expression in the teleost gill

Involvement of Cyclic Adenosine Monophosphate–Dependent Protein Kinase A and Pertussis Toxin–SensitiveG Proteins in the Migratory Response of Human CD14�
Mononuclear Cells to Katacalcin
NICOLE C. KANEIDER,1 PETRA EGGER,1 FRANZ J. WIEDERMANN,2 MARKUS RITTER,3
EWALD WOLL,1 and CHRISTIAN J. WIEDERMANN1
ABSTRACT
Katacalcin (KC) belongs to a small family of polypeptides that are encoded by the calc-1 gene and also includecalcitonin (CT) and procalcitonin NH2-terminal cleavage peptide (N-ProCT). Biological roles of KC orN-ProCT are unknown. To determine whether these polypeptides affect leukocyte function, forearm venousblood polymorphonuclear neutrophils and CD14� peripheral blood mononuclear cells (PBMCs) were isolatedfrom healthy human donors. Cell migration was assessed in a blindwell chemotaxis chamber using nitrocel-lulose micropore filters. Cellular levels of cyclic adenosine monophosphate (cAMP) were measured by HPLC;activation of protein kinase A was studied by Western blot. Fluorochrome-labeled peptide binding to cells wasstudied by fluorescence-activated cell sorting (FACS) and intracellular calcium transients were studied byconfocal microscopy with FLUO-3. KC elicited concentration-dependent migration of CD14� PBMC atconcentrations from the atomolar to the micromolar range and deactivated attractant-induced chemotaxis.CT N-terminal flanking peptide had no such effect. Neutrophils did not migrate toward any of those peptidesand their oxygen-free radical release was not affected as measured fluorometrically. Functional responses ofCD14� PBMC to KC correlated to forskolin-sensitive cAMP accumulation in cells and were inhibited byprotein kinase A inhibitor (PKI) and Rp diastereomer of adenosine 3�,5�-cyclic monophosphorothioate.Treatment of CD14� PBMC with KC activated protein kinase AC�. Intracellular calcium was decreased withCT, KC, and procalcitonin (PCT). Binding studies showed that KC might share the binding site with CT andPCT. Data indicate that KC regulates human CD14� PBMC migration via signaling events involving proteinkinase A–dependent cAMP pathways. (J Bone Miner Res 2002;17:1872–1882)
Key words: katacalcin, chemotaxis of CD14� mononuclear cells, RAMPs, inhacellular calcium CT-receptor,cAMP
INTRODUCTION
KATACALCIN (KC) belongs to a small family of polypep-tides that are encoded by the calc-1 gene. Calcitonin
(CT), procalcitonin (PCT) NH2-terminal cleavage peptide
(N-ProCT), and CT gene–related peptide (CGRP) areadditional members of the group. KC (originally termedPDN-21) is the C-terminal flanking peptide of CT in theprecursor polypeptide PCT. On the one hand, KC, CT,and N-ProCT are proteolytically processed from PCT,and, on the other hand, CGRP is derived from calc-1depending on tissue-specific alternative splicing of the
The authors have no conflict of interest.
1Division of General Internal Medicine, Department of Internal Medicine, University of Innsbruck, Innsbruck, Austria.2Department of Anesthesiology and Intensive Care Medicine, University of Innsbruck, Innsbruck, Austria.3Department of Physiology, University of Innsbruck, Innsbruck, Austria.
JOURNAL OF BONE AND MINERAL RESEARCHVolume 17, Number 10, 2002© 2002 American Society for Bone and Mineral Research
1872

gene.(1) The PCT-derived polypeptides come mainlyfrom the thyroid, whereas CGRP is present in both thethyroid and the nervous system.(2,3)
For CT, receptors, gene transcription, signal transduc-tion, and function in the physiology of a variety of organsystems are well described including the relationship tobone mineral density (BMD) and osteoporosis.(4) In1983, Hillyard and coworkers proposed that KC, like CT,may be involved in both plasma calcium regulation andskeletal maintenance(5); however, later the same authorsfailed to confirm a physiological hypocalcemic effect ofsynthetic KC.(6) To our knowledge, except for a proposedcostimulatory function of KC in combination with eitherCT or CGRP on intrathyroidal regulatory mechanisms ofthe follicular cell activity,(7) no physiological roles of KChave been discovered. Similarly, in 1989, N-ProCT wasreported to be a bone cell mitogen(8); also, in this case,however, the original hypothesis was not confirmed.(9)
Therefore, functional roles of KC and N-ProCT are yet tobe discovered.
More recently, intracellular antibody reaction against KCsupported the notion that various types of leukocytes maycontain PCT, suggesting that KC, CT, and N-ProCT may bemore widely distributed than originally thought.(10) Circu-lating levels of PCT are low in healthy subjects, but whenelevated levels were found in patients with bacterial infec-tions, PCT became an important protein in the detection anddifferential diagnosis of inflammatory states.(11,12) C cells inthe thyroid are not believed to be the source of bacterialinfection–induced PCT; probable sites of PCT productionduring inflammation are pulmonary and intestinal neuroen-docrine cells,(11) and because of the association betweenincreased levels of PCT and leukocyte-derived cytokinesduring sepsis, the possible expression of PCT in humanblood leukocytes has been investigated. PCT and CT werefound to be expressed in those cells and their synthesis wasmodulated by bacterial lipopolysaccharide and sepsis-related cytokines.(10,13) Although not uniformly con-firmed,(14) these findings suggested that leukocytes may beamong the sources of elevated PCT levels in patients withsepsis.
Hypothetical roles of PCT such as cytokine networkregulation and anti-inflammatory action are being inves-tigated. PCT inhibits lipopolysaccharide-induced cyto-kine synthesis and chemokine-induced migration inmonocytes,(15,16) which is not surprising because CT as aproteolysis product of PCT has been found to affectcytokine synthesis and migration in monocytes aswell.(17,18) It is not known if other peptide products ofPCT also affect monocytes. Because elevated intracellu-lar KC levels have been found in leukocytes of patientswith infection,(10,13) we were interested to study effectsof KC and N-ProCT on human leukocytes and to comparethe findings with that of CT and PCT. We report that KC,like CT and PCT, but not N-ProCT affects CD14� pe-ripheral blood mononuclear cell (PBMC) migration andinduces cyclic adenosine monophosphate (cAMP) accu-mulation and G protein– and cAMP-dependent activationof protein kinase AC�.
MATERIALS AND METHODS
Materials and reagents
Hanks’ balanced salt solution (HBSS) without Ca2� andMg2� was purchased from Gibco BRL, Life Technologies(Vienna, Austria). RPMI 1640 was purchased from Biolog-ical Industries (Kibbutz Beit Haemek, Israel). Lymphoprepwas from Nycomed Pharma (Oslo, Norway), anti-humanCD14 antibodies were from Miltenyi Biotec (BergischGladbach, Germany), and nitrocellulose filters were fromSartorius (Gottingen, Germany). 2�,7�-Dichlorofluorescindiacetate (DCFH-DA; Molecular Probes, Eugene, OR,USA) was dissolved in N,N-dimethylformamide to a stocksolution of 10 mmol/liter. Bovine serum albumin (BSA)was purchased from Dade-Behring (Marburg, Germany);N-formyl-Met-Leu-Phe (fMLP), a general protein kinase Cinhibitor GF 109203X (GFX), wortmannin (WTN),tyrphostin-23, pertussis toxin (PTX), cholera toxin (CTX),and human CT were from Sigma Chemical Co. (St. Louis,MO, USA). Rantes and macrophage chemoattractant pro-tein (MCP) 1 were from PeproTech (London, UK). Proteinkinase A inhibitor (PKI) and Rp diastereomer of adenosine3�,5�-cyclic monophosphorothioate (Rp-cAMPS) were fromSigma Chemical Co.. cAMP was purchased from Calbio-chem (La Jolla, CA, USA). KC and N-ProCT were fromBachem BioSciences, Inc. AG (Bubendorf, Switzerland)and recombinant human PCT was from Brahms Diagnostica(Berlin, Germany). Bombesin was from Neosystems (Strass-bourgh, France). Anti–protein kinase AC� monoclonal anti-body was from Transduction Laboratories (Lexington, KY,USA). Polyvinylidene fluoride (PVDF) membranes werefrom Amersham (Little Chalfont, Buckinghamshire, UK)and NP-40 was from Roche Molecular Biochemicals (Indi-anapolis, IN, USA). Sodium dihydrogen phosphate waspurchased from Sigma Chemical Co.; solvents (water andacetonitrile, HPLC grade) were from Merck & Co. (Darm-stadt, Germany).
Preparation of CD14� PBMC
PBMCs were prepared from peripheral venous blood ofhealthy volunteers (anticoagulated with EDTA). After Lym-phoprep density gradient centrifugation, CD14� PBMCs werecollected and washed three times with normal saline. Positiveselection was performed by adding MACS colloidal superpara-magnetic microbeads (Miltenyi Biotec GmbH, Bergisch Glad-bach, Germany) conjugated with monoclonal anti-humanCD14 antibodies to cooled, freshly prepared PBMC prepara-tions in MACS buffer (PBS with 5 mM of EDTA and 0.5%BSA) according to the manufacturer’s instructions. Cells andmicrobeads were incubated for 15 minutes at 4–6°C. In themeantime the separation column was positioned in the MACSmagnetic field and washed with MACS buffer at room tem-perature. The cells were washed with MACS buffer, resus-pended, and loaded onto the top of the separation column. Theeluent containing CD14� cells was withdrawn and after re-moval of the column from the magnet, trapped CD14� PBMCwere eluted with the sixfold amount of cold MACS buffer,centrifuged, and resuspended in medium containing 0.5%BSA. By immunocytochemistry, preparations yielded a purityof �98%.(19)
1873CELLULAR FUNCTION OF KATACALCIN

Preparation of neutrophils
Neutrophils were obtained from peripheral EDTA anti-coagulated blood of healthy volunteers after discontinuousdensity gradient centrifugation through a layer of Ficoll-Hypaque, followed by hypotonic lysis of contaminatingerythrocytes.(20) Cell preparations yielded �95% neutro-phils (by morphology in GIEMSA stains) and �99% via-bility (by trypan dye exclusion).
Chemotaxis experiments
Leukocyte migration was measured using a modified48-blindwell microchemotaxis chamber equipped with5-�m pore-sized nitrocellulose filters for CD14� PBMCchemotaxis. In some experiments cells were incubated for20 minutes with GFX (500 nmol/liter), staurosporine (10ng/ml), tyrphostin-23 (10 ng/ml), WTN (10 nmol/liter), PKI(from 0.1 to 100 ng/ml) or Rp-cAMPS (from 100 pM to 100�M), or CTX (1 nmol/liter) or PTX (1 nmol/liter). Fordetermination of KC’s potency to deactivate CD14� PBMCchemotaxis toward fMLP, we incubated cells with KC(from 1 amol/liter to 1 �mol/liter) for 20 minutes. Forcontrol, cAMP-independent migration of CD14� PBMCtoward bombesin was tested in some of the experiments.After washing twice, 50 �l of a cell suspension (1 � 106
cells/ml) was put into the upper compartment of the che-motaxis chamber and cells were allowed to migrate for 90minutes toward peptides derived from the calc-1 gene in thelower wells. After these migration periods, the filters weredehydrated, fixed, and stained with hematoxylin and eosin.Migration depth was quantified by microscopy, measuringthe distance from the surface of the filter to the leading frontof three cells. Data are expressed as “chemotaxis index,”which is the ratio between the distance of directed andundirected migration.
Respiratory burst activity of neutrophils
Neutrophil respiratory burst activity was detected by anassay using DCFH-DA, as described previously.(21) Thisassay is based on the oxidation of nonfluorescentDCFH-DA to highly fluorescent 2�,7�-dichlorofluorescein(DCF) both intracellularly and extracellularly.(22) Neutro-phils were primed for 30 minutes at 37°C (5% CO2 atmo-sphere) with various concentrations (from 1 amol/liter to 1�mol/liter) of KC or N-ProCT, washed twice, and resus-pended in HBSS. Then, 100 �l/well (96-well plate) of 2 �105 neutrophils were immersed at 37°C in a 10-�M solutionof DCFH-DA in phenol red–free HBSS containing 1 �Mformyl-Met-Leu-Phe, as a triggering agent, or medium. Theplates were covered with lids and placed in a humidifiedincubator for various time intervals. Fluorescence activitywas determined at 485 � 20 nm excitation and 530 � 25nm emission wavelengths using the CytoFluor 4000 fluo-rescence measurement system (Millipore Corp., Bedford,MA, USA). Because the cells in the plates were not dis-turbed by the measuring procedure, the plates could beincubated for various time periods and reread as desired.Readings were taken every 10 minutes for 30 minutes.
Western blot analysis
Cells were incubated with KC at various concentrations(from 1 amol/liter to 10 nmol/liter) and fMLP (10 nmol/liter) for 30 minutes. Cells were lysed in lysis buffer con-taining NP-40. Proteins were separated on 7.5% SDS poly-acrylamide gels and blotted onto PVDF membranes, whichwere blocked with 5% phosphatase-free milk powder inTris-buffered saline with 0.1% Tween. The antibody thenwas diluted in 0.2% milk to a final concentration 0.05 �g/mland blots were incubated overnight at room tempera-ture. Immunoreactivity was determined using peroxidase-conjugated goat anti-mouse immunoglobulin G (IgG) andthe enhanced chemiluminescence (ECL) reaction (Amer-sham, Pharmacia Biotech, Little Chalfont, Buckingham-shire, England). Intensity of the Western blot bands wasquantified using the Fluor-S MultiImager System and theQuantity One Software (Bio-Rad Laboratories, Hercules,CA, USA).
Determination of cAMP
Cells were incubated with KC at various concentrations(from 1 amol/liter to 1 �mol/liter), washed twice in ice-coldPBS, and lysed with a buffer containing 0.01 M of Tris-HCl,0.5% NP-40 and 1 mmol/liter phenylmethylsulfonyl fluo-ride (PMSF). Thereafter, the lysate was centrifuged at15,000g for 5 minutes to pellet the insoluble material. Fiftymicroliters of each sample was subjected to HPLC analysis.Detection of cAMP was performed by ion-pair chromatog-raphy with an Inertsil ODS-2 (250 mm � 4 mm; 5 �m;Wagner-Munz, Vienna, Austria). The signal was monitoredwith a variable-wavelength UV-visible detector at a wave-length of 260 nm. The chromatographic conditions were asfollows: isocratic elution at ambient temperature with 97%NaH2PO4 (0.1 M, pH 6.0) and 3% acetonitrile; flow rate, 1.0ml/minute; injection volume, 50 �l; retention time, 7.7minutes; elution time, 16 minutes; posttime, 0 minutes.
Measurement of intracellular calcium concentration
Measurements of intracellular calcium concentrationchanges were carried out using the calcium-sensitive dyeFLUO 3. This dye does not allow for ratiometric measure-ment and thus for absolute quantification of intracellularcalcium concentrations, but does exhibit a superb sensi-tivity to subtle changes in intracellular calcium concen-tration. Suspended CD14� PBMCs were incubated for 20minutes in RPMI 1640 medium with 1 �M of FLUO3-acetoxymethylester (Molecular Probes) and allowed toadhere to uncoated glass coverslips. Thereafter, the glasscoverslip was mounted into a perfusion chamber allowingfor fluid exchange and the cells were superfused continu-ously with control or experimental solution. The controlsolution was composed of NaCl, 130 mmol/liter; KCl, 5mmol/liter; MgCl2, 1 mmol/liter; CaCl2, 2 mmol/liter; glu-cose, 5 mmol/liter; HEPES, and 10 mmol/liter; pH 7.4. Forcalcium-free solutions, CaCl2 and MgCl2 were omitted and10 mM of EGTA was added. Where indicated KC, CT, orPCT was added to the control solution at a final concentra-tion of 1 pmol/liter each or 1 �M of ionomycin was added
1874 KANEIDER ET AL.

to the control and calcium-free solution. Adherent cells wereallowed to adapt for at least 5 minutes before the experiments.Measurements were performed with an inverted microscope(IX 70, magnification �40; Olympus, Vienna, Austria; objec-tive Plan NeoFluar UApo/340; Olympus). Light from a mono-chromator (Polychrome IV, excitation wavelength of 505 nmand illumination pulses of 25–30 ms duration at 1-s intervals;TILL Photonics, Graefelfing, Germany) was deflected by adichroic mirror (Q515LP, AHF Analysentechnik, Tuebingen,Germany) to the object and the emitted light was passedthrough the dichroic mirror and a bandpass filter (HQ535/30,AHF Analysentechnik) to a cooled CCD camera (IMAGOCCD Super VGA; TILL Photonics). The monochromator andCCD camera were controlled by the TILL VISION software(version 4.0; TILL Photonics). Data analysis was performedusing the same software. Fluorescence values were obtained asmean gray values from selected regions of interest coveringindividual CD14� PBMCs. These values were corrected forcellular autofluorescence and background subtracted and ex-pressed as relative fluorescence values with respect to themeasured mean gray values under control conditions 120 sbefore addition of the respective hormone. Maximum andminimum concentrations of intracellular calcium were ob-tained by application of 1 �M of ionomycin (Sigma ChemicalCo.) in the presence and absence of calcium, respectively.
Receptor binding assays
Labeling of peptides was performed with theFluorescein-EX Protein Labeling kit (Molecular Probes).Labeling studies were performed at 4°C. CD14� PBMCswere first incubated with human IgG (150 �g/ml) to
avoid unspecific binding of substances. After 20 minutes,cells were washed and fluorescein isothiocyanate(FITC)–labeled KC or CT was added to the cells for 20minutes. After another washing step, cells were incubatedwith unlabeled KC, PC, CT, or N-ProCT at variousconcentrations (from 1 amol/liter to 1 �mol/liter) foranother 20 minutes. After washing, samples were sub-jected to FACS analyses on a Becton Dickinson (SanJose, CA, USA) FACScan. Data were analyzed byCellQuest (San Jose, CA, USA) software.
Statistical analysis
Data are expressed as mean and standard error of themean (SEM). Means were compared by Mann-Whitney Utest or paired t-test after Kruskal-Wallis analysis of variance(ANOVA). A p-value � 0.05 was considered significant.Analyses were performed using the StatView softwarepackage (Abacus Concepts Inc. Berkeley, CA, USA).
RESULTS
Effects of PCT-derived polypeptides on CD14�
PBMCs and neutrophil migration
To determine whether PCT-derived polypeptides induceCD14� PBMCs and neutrophil chemotaxis, we tested invitro migration of these cells toward KC, N-ProCT, CT, andPCT at a wide range of concentrations (from 1 amol/liter to1 �mol/liter). KC, CT, and PCT induced CD14� PBMCchemotaxis in a concentration-dependent manner, with CTand PCT being effective in the range of 100 amol/liter–
FIG. 1. Influence of PCT-derived polypeptides on CD14� PBMC migration. (A) Directed migration of CD14� PBMCs was measured using amodified 48-blindwell microchemotaxis chamber equipped with 5-�m pore-sized nitrocellulose filters for CD14� PBMC chemotaxis. Cells wereallowed to migrate toward various concentrations of PCT-derived peptides (1 aM to 1 �M) for 90 minutes. Migration depth was quantified bymicroscopy, measuring the distance from the surface of the filter to the leading front of three cells. Data are expressed as chemotaxis index, whichis the ratio between the distance of directed and undirected migration. Mean distance of undirected migration was 57 � 4.5 �m. (B) Influenceof PCT-derived peptides on CD14� PBMC migration toward fMLP (10 nM) was tested after pretreatment of cells with the peptides (1 aM to 1�M). After washing, cells were allowed to migrate toward fMLP for 90 minutes. Data are expressed as chemotaxis index, the ratio betweendirected and random migration. Mean distance of random migration was 48 � 5.8 �m. Statistical analysis: Mann-Whitney U test afterKruskal-Wallis (p � 0.01; *p � 0.05; **p � 0.01; n 6 for each experiment).
1875CELLULAR FUNCTION OF KATACALCIN

1�mol/liter and KC at concentrations between 10 fmol/literand 1 �mol/liter; N-ProCT did not affect migration ofCD14� PBMCs. Maximum activities were seen at concen-trations between 1 pmol/liter and 10 nmol/liter (Fig. 1A).All of the polypeptides failed to induce chemotactic activityin neutrophils (data not shown). When compared with mean(n 6) chemotaxis indices of 1.8 � 0.11 and 2.2 � 0.22 ofRantes (20 ng/ml) and fMLP (10 nmol/liter) in CD14�
PBMCs and neutrophils, respectively, the PCT-derived pep-tides appeared to be of similar potencies in CD14� PBMCsbut not neutrophils.
To test whether KC-induced migration of CD14� PBMCis dependent on concentration gradients (chemotaxis) orrepresents enhanced speed or frequency of spontaneousmigration only (chemokinesis), checkerboard analysis ofCD14� PBMC migration to KC was performed. Resultsshow that the type of response of CD14� PBMC toward KCreflects true chemotaxis (Table 1).
Effects of PCT-derived polypeptides onchemoattractant-stimulated migrationof CD14� PBMCs
To test whether PCT-derived polypeptides have the po-tency to deactivate CD14� PBMC migration toward knownattractants, cells were incubated with KC, N-ProCT, CT, orPCT in concentrations between 10 fmol/liter and 1 �mol/liter for 20 minutes and then washed twice and allowed tomigrate toward 10 nmol/liter of fMLP for 90 minutes.PCT-pretreated cells showed a significantly reduced chemo-tactic response at concentrations from 1 pmol/liter to 1�mol/liter; CT-pretreated CD14� PBMC showed a signif-icant decrease in chemotaxis at concentrations of 100 pmol/liter and 10 nmol/liter; KC pretreatment led to aconcentration-dependent decrease at concentrations be-tween 1 amol/liter and 10 fmol/liter and was a more potentinhibitor of fMLP-induced chemotaxis than CT or PCT(Fig. 1B). N-ProCT did not affect the chemotactic responseof CD14� PBMCs to fMLP.
KC deactivated CD14� PBMC chemotaxis not only to-ward fMLP but also toward other attractants of the chemo-kine family (heterologous deactivation) as well as towardPCT and CT. Pretreatment of CD14� PBMCs with KC alsodeactivated subsequent chemotaxis toward KC itself (ho-mologous deactivation; Table 2).
Effect of PCT-derived polypeptides on spontaneousand triggered respiratory burst activity of neutrophils
No effects of KC, CT, N-ProCT, or PCT were seen whenneutrophil basal respiratory burst was tested, and fMLP-triggered (1 �mol/liter) release of oxygen-free radicals wasnot significantly altered by the presence of the peptideswhen tested at concentrations from 1 amol/liter to 1 �mol/liter (data not shown).
Effects of PTX, CTX, and signaling enzyme inhibitorson CD14� PBMC migration
To investigate which signaling mechanisms might be in-volved in KC-induced chemotaxis of CD14� PBMCs, cellswere incubated with several enzyme blockers. The protein
TABLE 2. HOMOLOGEOUS AND HETEROLOGEOUS
DEACTIVATION OF CD14� PBMC CHEMOTAXIS BY KC
Attractant
Treatment
MediumKC
(100 nmol/liter)
Mean SEM Mean SEM
Medium 1.00 — 1.03 0.070fMLP (10 nmol/liter) 2.21* 0.217 1.14† 0.083MCP-1 (0.1 �mol/liter) 1.87* 0.149 1.12† 0.105Rantes (20 ng/mliter) 1.82* 0.114 1.13† 0.089KC (100 nmol/liter) 1.73* 0.127 1.08† 0.089PCT (100 nmol/liter) 1.92* 0.147 1.19† 0.140CT (100 nmol/liter) 1.88* 0.173 1.18† 0.127
CD14� PBMCs were incubated with medium or KC (CT) for 20minutes, washed twice, and allowed to migrate toward solubleattractants in the lower chamber as described in Table 1. Statisticalanalysis: Mann-Whitney U-test after Kruskal-Wallis ( p � 0.01;*p � 0.05 vs. medium after pretreatment with medium; †p �0.05 vs. attractant after pretreatment with medium; n � 3).
TABLE 1. CHECKERBOARD ANALYSIS OF EFFECTS OF CONCENTRATION GRADIENTS OF KC IN CD14� PBMC MIGRATION
KC (mol/liter), upper chamber
Medium 10�10 10�8 10�6
KC (mol/liter), Medium 1.000 � 0.000* 1.183 � 0.125 0.857 � 0.008 0.773 � 0.157Lower chamber 10�10 1.788 � 0.016* 1.196 � 0.108 0.998 � 0.087 0.954 � 0.165
10�8 2.116 � 0.080*0.080.080* 1.816 � 0.152* 1.062 � 0.052 0.884 � 0.17710�6 2.274 � 0.158* 1.662 � 0.067* 1.576 � 0.153* 0.780 � 0.046
Directed migration of CD14� PBMCs to KC was tested by checkerboard analysis. Cells were allowed to migrate to variousconcentration gradients of KC for 90 minutes. Migration depth was quantified by microscopy. Data are expressed as chemotaxis index,which is the ratio between directed and undirected migration of cells. Mean distance of migration to medium in the upper and lowerchamber was 52 � 4.2 �m. Statistical analysis: Mann-Whitney U-test after Kruskal-Wallis analysis of variance ( p � 0.001; n � 3;*p � 0.05 vs. medium in the upper and lower chamber; only those groups within the matrix being sufficient to distinguish betweenchemokinesis and chemotaxis were compared statistically by Mann-Whitney U-test).
1876 KANEIDER ET AL.

kinase A and C inhibitor staurosporine, the protein kinase Cinhibitor GFX, the tyrosine kinase inhibitor tyrphostin-23, andthe phospholipase 3 inhibitor WTN were first used for block-ing signaling enzymes; involvement of G proteins was testedwith PTX, which is known to induce Gi proteins, and CTX,which induces Gs proteins. Migration of CD14� PBMCs to-ward all, PCT, CT, KC, and CGRP was inhibited by stauro-sporine but not by GFX or any of the other enzyme blockers;for comparison, migration of CD14� PBMCs toward fMLP
was inhibited by staurosporine and tyrphostin-23; bombesin-induced chemotaxis was inhibited by tyrphostin-23 (Table 3).
In the analysis of G protein involvement in calc-1 gene-derived polypeptide chemotaxis by using PTX or CTX, allPCT, CT, KC, and CGRP were significantly affected in theiractivation of migration by PTX, which was not so afterpretreatment with CTX (Table 3).
Effect of PCT-derived polypeptides on cAMPaccumulation in CD14� PBMCs
Effects of KC, N-ProCT, and CT on signaling events inCD14� PBMCs were tested further by measuring cellularlevels of cAMP after preincubating cells with the peptidesfollowed by washing. KC, like CT, stimulated cAMP accu-
FIG. 2. Accumulation of intracellular cAMP after KC exposure ofCD14� PBMCs. CD14� PBMCs (106 cells/ml) were incubated withKC at various concentrations (1 amol/liter to 1 �mol/liter), CT (1amol/liter to 1 �mol/liter) or N-ProCT (pmol/liter). After washing,cells were lysed and lysates were subjected to HPLC analysis. cAMPcontent was determined using an UV-visible wavelength detector.Statistical analysis: Mann-Whitney U test after Kruskal-Wallis (p �0.01; *p � 0.05, versus CD14� PBMC incubation with medium; n 3 for KC; n 1 for N-ProCT and CT).
FIG. 3. Activation of protein kinase AC� by KC. For 20 minutes,CD14� PBMCs were treated with KC (1 aM to 10 nM) or fMLP (10nmol/liter). Cells then were washed twice and lysed. Equal proteinconcentrations of lysates were loaded onto lanes. Activated proteinkinase A was visualized with a monoclonal antibody against subunitC� after 7.5% SDS-PAGE.
TABLE 3. EFFECTS OF PTX, CTX, AND SIGNALING ENZYME INHIBITORS ON CD14� PBMCS MIGRATION TOWARD calc-1GENE–DERIVED POLYPEPTIDES
Treatment
Chemotaxis index
KC mean(SEM)
PCT mean(SEM)
CT mean(SEM)
CGRP mean(SEM)
fMLP mean(SEM)
Bombesinmean (SEM)
Medium 1.6 (0.12) 1.9 (0.17) 1.8 (0.15) 1.7 (0.08) 1.8 (0.05) 1.9 (0.05)GFX (500 nmol/liter) 1.5 (0.07) 1.7 (0.10) 1.6 (0.12) 1.6 (0.12) 1.6 (0.14) 2.0 (0.06)Staurosporine (10 ng/mliter) 0.8 (0.09)† 0.9 (0.06)† 0.9 (0.08)† 0.8 (0.11)† 1.2 (0.01)* 1.6 (0.3)Tyrphostin-23 (10 ng/mliter) 1.6 (0.06) 1.8 (0.09) 1.7 (0.09) 1.5 (0.15) 1.7 (0.03)* 1.3 (0.132)*WTN (10 nmol/liter) 1.5 (0.12) 1.6 (0.20) 1.9 (0.11) 1.6 (0.17) 1.9 (0.07) 1.5 (0.19)CTX (1 nmol/liter) 1.6 (0.07) 1.6 (0.19) 1.6 (0.12) 1.4 (0.15) 1.9 (0.06) 1.4 (0.26)PTX (1 nmol/liter) 1.2 (0.13)* 0.8 (0.03)† 1.0 (0.05)† 1.0 (0.14)* 1.3 (0.04)* 1.0 (0.04)*
CD14� PBMCs were incubated with various signaling enzyme blockers for 20 minutes. After washing, chemotaxis experiments towardKC (1 pmol/liter), PCT (1 pmol/liter), CT (1 pmol/liter), CGRP (1 nmol/liter), fMLP (10 nmol/liter), and bombesin (1 nmol/liter) wereperformed as described in Table 1. Data are expressed as chemotaxis index, which is the ratio between directed and random migration.Mean distance of random migration was 54 � 3.2 �m. Statistical analysis: Mann-Whitney U-test after Kruskal-Wallis ( p � 0.01;*p � 0.05, †p � 0.01 vs. pretreatment with medium; n � 3).
1877CELLULAR FUNCTION OF KATACALCIN

mulation in CD14� PBMC concentration dependently withmaximum effects seen in the picomolar range, whereasN-ProCT did not (Fig. 2). KC-stimulated cAMP accumula-
tion was increased further by the presence of forskolin (datanot shown).
Protein kinase A activation in CD14� PBMC by KC
The observation of cAMP accumulation in CD14� PB-MCs by KC and the inhibition of KC-induced migration ofCD14� PBMCs by staurosporine but not GFX suggestedthat protein kinase A activity might be involved in theobserved function of KC. To confirm these functional find-ings, Western blot analysis was performed. Using an anti–protein kinase AC� antibody, concentration-dependent sub-unit dissociation of activated protein kinase A was seen inresponse to KC (Fig. 3).
Effects of cAMP/protein kinase A inhibitors on PCT-derived polypeptide-induced migration
The cAMP/protein kinase A inhibitor Rp-cAMPS and theprotein kinase A inhibitor PKI were used to confirm the roleof cAMP-dependent protein kinase A activation in CD14�
PBMC migration by KC. Chemotaxis of CD14� PBMCtoward KC (1 pmol/liter) was inhibited by pretreatment withPKI (Fig. 4). PCT- and fMLP-induced chemotaxis also wasinhibited by pretreatment with Rp-cAMPS, whereas chemo-taxis toward bombesin that was used as control attractantwas not (Fig. 5).
Effect of KC, CT, and PCT on calcium transient inCD14� PBMC
Addition of 1 pmol/liter of KC, CT, or PCT led to withina 220-s decrease of the relative fluorescence from 98.8 �
FIG. 4. The protein kinase A inhibitor PKI inhibits KC-inducedCD14� PBMC chemotaxis. After pretreatment with PKI (0.1–100ng/ml) for 20 minutes, cells were washed and chemotaxis toward KC(1 pM) was tested as described for Fig. 1. Data are expressed aschemotaxis index, which is the ratio between directed and undirectedmigration of cells. Statistical analysis: Mann-Whitney U test afterKruskal-Wallis (p � 0.01; *p � 0.05; n 3).
FIG. 5. Rp-cAMPS inhibits CD14� PBMCmigration toward KC. Cells were incubated withvarious concentrations of Rp-cAMPS (100 pMto 100 �M) for 20 minutes. After washing, che-motaxis toward KC, PCT, fMLP, and bombesinwas tested in modified Boyden chemotaxischambers as described for Fig. 1. Data are ex-pressed as chemotaxis index, which is the ratiobetween directed and undirected migration. Sta-tistical analysis: Mann-Whitney U test afterKruskal-Wallis (p � 0.01; *p � 0.05; n 3).
1878 KANEIDER ET AL.
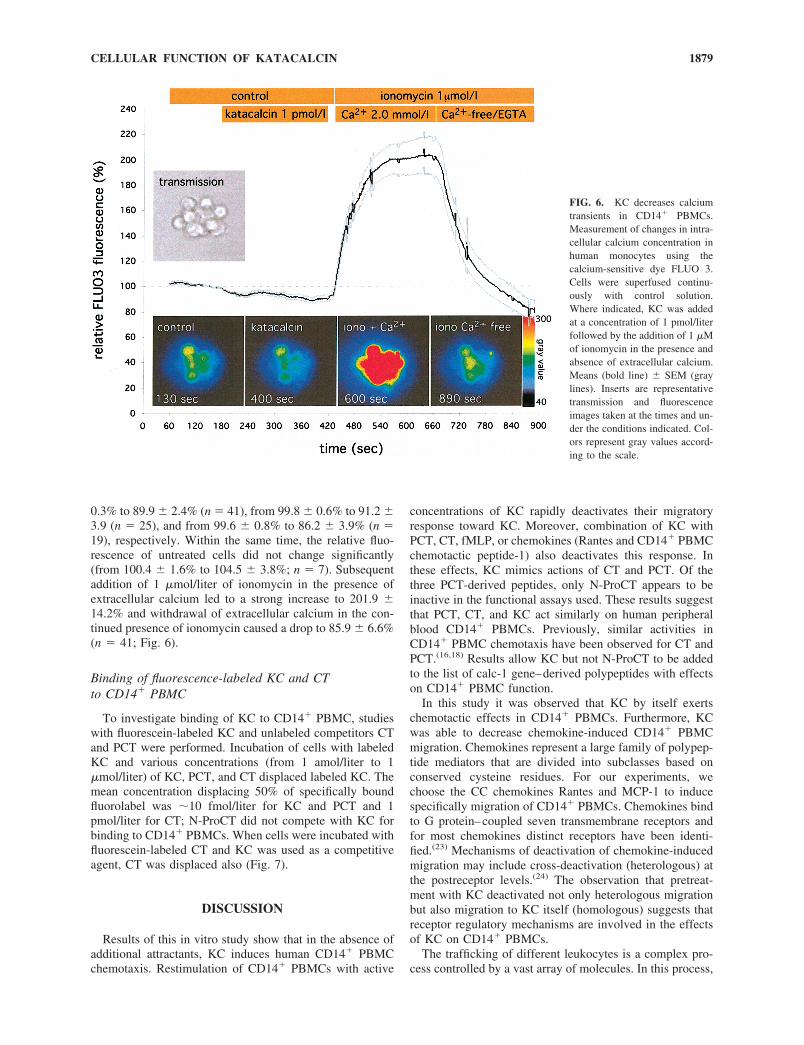
0.3% to 89.9 � 2.4% (n 41), from 99.8 � 0.6% to 91.2 �3.9 (n 25), and from 99.6 � 0.8% to 86.2 � 3.9% (n 19), respectively. Within the same time, the relative fluo-rescence of untreated cells did not change significantly(from 100.4 � 1.6% to 104.5 � 3.8%; n 7). Subsequentaddition of 1 �mol/liter of ionomycin in the presence ofextracellular calcium led to a strong increase to 201.9 �14.2% and withdrawal of extracellular calcium in the con-tinued presence of ionomycin caused a drop to 85.9 � 6.6%(n 41; Fig. 6).
Binding of fluorescence-labeled KC and CTto CD14� PBMC
To investigate binding of KC to CD14� PBMC, studieswith fluorescein-labeled KC and unlabeled competitors CTand PCT were performed. Incubation of cells with labeledKC and various concentrations (from 1 amol/liter to 1�mol/liter) of KC, PCT, and CT displaced labeled KC. Themean concentration displacing 50% of specifically boundfluorolabel was �10 fmol/liter for KC and PCT and 1pmol/liter for CT; N-ProCT did not compete with KC forbinding to CD14� PBMCs. When cells were incubated withfluorescein-labeled CT and KC was used as a competitiveagent, CT was displaced also (Fig. 7).
DISCUSSION
Results of this in vitro study show that in the absence ofadditional attractants, KC induces human CD14� PBMCchemotaxis. Restimulation of CD14� PBMCs with active
concentrations of KC rapidly deactivates their migratoryresponse toward KC. Moreover, combination of KC withPCT, CT, fMLP, or chemokines (Rantes and CD14� PBMCchemotactic peptide-1) also deactivates this response. Inthese effects, KC mimics actions of CT and PCT. Of thethree PCT-derived peptides, only N-ProCT appears to beinactive in the functional assays used. These results suggestthat PCT, CT, and KC act similarly on human peripheralblood CD14� PBMCs. Previously, similar activities inCD14� PBMC chemotaxis have been observed for CT andPCT.(16,18) Results allow KC but not N-ProCT to be addedto the list of calc-1 gene–derived polypeptides with effectson CD14� PBMC function.
In this study it was observed that KC by itself exertschemotactic effects in CD14� PBMCs. Furthermore, KCwas able to decrease chemokine-induced CD14� PBMCmigration. Chemokines represent a large family of polypep-tide mediators that are divided into subclasses based onconserved cysteine residues. For our experiments, wechoose the CC chemokines Rantes and MCP-1 to inducespecifically migration of CD14� PBMCs. Chemokines bindto G protein–coupled seven transmembrane receptors andfor most chemokines distinct receptors have been identi-fied.(23) Mechanisms of deactivation of chemokine-inducedmigration may include cross-deactivation (heterologous) atthe postreceptor levels.(24) The observation that pretreat-ment with KC deactivated not only heterologous migrationbut also migration to KC itself (homologous) suggests thatreceptor regulatory mechanisms are involved in the effectsof KC on CD14� PBMCs.
The trafficking of different leukocytes is a complex pro-cess controlled by a vast array of molecules. In this process,
FIG. 6. KC decreases calciumtransients in CD14� PBMCs.Measurement of changes in intra-cellular calcium concentration inhuman monocytes using thecalcium-sensitive dye FLUO 3.Cells were superfused continu-ously with control solution.Where indicated, KC was addedat a concentration of 1 pmol/literfollowed by the addition of 1 �Mof ionomycin in the presence andabsence of extracellular calcium.Means (bold line) � SEM (graylines). Inserts are representativetransmission and fluorescenceimages taken at the times and un-der the conditions indicated. Col-ors represent gray values accord-ing to the scale.
1879CELLULAR FUNCTION OF KATACALCIN

cells must be able to sense continuously small changes inchemoattractant gradients. Migration through a chemotacticgradient probably uses an on-off mechanism in which che-moattractant receptor desensitization may be an importantstep. For a large number of chemoattractant receptors, rapiddesensitization appears to involve agonist-promoted receptorphosphorylation by G protein–coupled receptor kinases.(25–
27) G protein–coupled receptor kinase–mediated phosphor-ylation of serine/threonine residues in the carboxyl tail andintracellular loops of G protein–coupled receptors increasesthe affinity for arrestin-type proteins, the binding of whichprevents any further coupling between the receptor and Gproteins.(28,29) The uncoupled receptors are subsequentlyremoved from the plasma membrane in a process termedinternalization. Recent evidence suggests that both Gprotein–coupled receptor kinases and arrestins play a keyrole in the sequestration process.(30,31) Receptor tyrosinekinases, G protein–coupled receptors, phosphoinositide-3-OH kinase, and the Rho family of GTPases receive andtransduce the signals that lead to actin nucleation that is re-quired for cell migration.(32) It is known that neitherphosphatidylinositol-3-kinase, extracellular signal-regulatedkinase activity, nor protein kinase C are required for CD14�
PBMC migration toward CD14� PBMC chemoattractant pro-tein 1 or fMLP.(33) Therefore, to further define the signalingpathways of KC in CD14� PBMC chemotaxis, effects ofsignaling enzyme blockers were tested.
Staurosporine but not the specific protein kinase C inhib-itor GFX inhibited chemotaxis, suggesting that protein ki-nase A may be one of the signaling pathways of KC inCD14� PBMCs. This hypothesis was confirmed by show-ing that the cAMP-dependent protein kinase A inhibitorRp-cAMPS and the protein kinase A inhibitor PKI alsoabolished effects of CD14� PBMC migration.
Pretreatment of CD14� PBMCs with increasing concen-trations of KC was associated with a progressive increase inthe cytosolic concentration of cAMP. The accumulation ofcAMP in CD14� PBMCs correlated closely with the abilityof KC to affect CD14� PBMC chemotaxis. The ability ofKC to increase the cellular levels of cAMP was synergisti-cally enhanced by pretreating the cells with forskolin, anactivator adenylate cyclase. Several hormones, includingcatecholamines, histamine, and prostaglandin E1, areknown to regulate CD14� PBMC functions by stimulatingthe accumulation of cAMP,(34) and inhibition of chemokine-induced migration by cellular cAMP has been reported.(35)
Thus, results observed on KC theoretically fit well intocurrent hypotheses on the mechanisms of regulation ofCD14� PBMC migration.
A decrease of intracellular calcium was observed byaddition of CT, KC, and PCT. There is some evidence thatCT might inhibit a Na�/Ca2� exchanger in the rabbitnephron.(36) On rat cortical collecting ducts, there also iscontradictory literature suggesting enhanced calcium influxin response to CT because of an opening of low-affinityCa2� channels.(37) However, CGRP decreases intracellularcalcium by inhibiting Ca2� influx through membrane hy-perpolarization mediated partly by activation of the largeconductance Ca2�-activated potassium channels and by adecrease in Ca2�, presumably by sequestrating cytosolicCa2� into thapsigargin-sensitive Ca2� storage sites as was
FIG. 7. Displacement of fluoro-labeled KC and CT by calc-1 gene–derived peptides. (A) Incubation of cells with fluorochrome-labeled KC(1 �mol/liter) and various concentrations (1 amol/liter to 1 �mol/liter)of KC, PCT, N-ProCT, and CT. Mean � SEM fluorescence activityshown on the y axis and serial dilution of competitor shown on the xaxis (n 3). (B) Cells incubated with fluorescein-labeled CT and KCused as competitive agent (n 3).
1880 KANEIDER ET AL.

shown in rat intramural coronary arteries.(38) No data onintracellular calcium regulation in CD14� PBMCs by calc-1–derived peptides have been described so far. The mech-anism of the decrease in free intracellular calcium inducedby CT, PCT, and KC needs to be clarified in additionalstudies. Because CGRP acts on CT receptors depending onreceptor activity–modifying protein (RAMP),(39) it is notsurprising to find similar calcium responses induced by CT,KC, and PCT.
A PTX-sensitive, Gi protein–coupled receptor appears tobe involved in the response of CD14� PBMCs to KC,whereas CTX, which induces Gs proteins, did not abrogatethe effect of KC. Such a response pattern has been describedpreviously as being typical of a variety of chemoattractantreceptors including chemokine and formyl peptide recep-tors.(40)
KC specifically binds to CD14� PBMCs, which can bedisplaced by PCT or CT but not N-ProCT, suggesting thatthese calc-1–derived peptides use the same binding site onthe cells. Effects of CT are known to be mediated by the CTreceptor, a G protein–linked receptor that couples to Gs,Gi/o, and Gq and whose stimulation leads to the activationof multiple signaling effectors, including adenylyl cyclase,protein kinase A, phospholipases C, D, and A2, and proteinkinase C, as well as extracellular signal–regulated kinases(Erk1/2).(41–44) Because several of these signaling pathwayshave been reported as being involved in regulating CD14�
PBMC migration and were confirmed here as mediatingeffects of CT and CGRP in CD14� PBMCs, it might be thatthe CT receptor directly regulates cell migration. Involve-ment of cAMP-dependent protein kinase A and PTX-sensitive G proteins has been reported for CGRP-inducedreceptor signaling in other types of cells(45) that has beendescribed also as regulating leukocyte migration.(46)
RAMPs are a family of single transmembrane domainproteins shown to be important for the transport and ligandspecificity of the CT receptor.(39,47) Because the CT gene–derived polypeptides, KC, CT, PCT, and CGRP seem to beable to affect CD14� PBMC migration via similar signaltransduction pathways identified as being typical of CTreceptors, that is, cAMP-dependent protein kinase A andPTX-sensitive G proteins, it may well be that depending onRAMPs, KC also acts via CT receptors of CD14� PBMCs.
REFERENCES
1. Horn DA, Latchman DS 1993 A distal region of the CALC-1gene is necessary for regulated alternative splicing. FEBS Lett324:123–126.
2. Maruna P, Nedelnikova K, Gurlich R 2000 Physiology andgenetics of procalcitonin. Physiol Res 49(Suppl 1):S57–S61.
3. Seitz PK, Cooper CW 1989 Cosecretion of calcitonin andcalcitonin gene-related peptide from cultured rat medullarythyroid C cells. J Bone Miner Res 4:129–134.
4. Pondel M 2000 Calcitonin and calcitonin receptors: Bone andbeyond. Int J Exp Pathol 81:405–422.
5. Hillyard CJ, Myers C, Abeyasekera G, Stevvensvenson JC,Craig RK, MacIntyre I 1983 Katacalcin: A new plasmacalcium-lowering hormone. Lancet 1:846–848.
6. MacIntyre I, Hillyard CJ, Reynolds JJ, Gaines Das RE, CraigRK 1984 A second plasma calcium-lowering peptide from thehuman calcitonin precursor—a re-evaluation. Nature 308:84.
7. Ahren B 1989 Effects of calcitonin, katacalcin, and calcitoningene-related peptide on basal and TSH-stimulated thyroid hor-mone secretion in the mouse. Acta Physiol Scand 135:133–137.
8. Burns DM, Forstrom JM, Friday KE, Howard GA, Roos BA1989 Procalcitonin’s amino-terminal cleavage peptide is abone-cell mitogen. Proc Natl Acad Sci USA 86:9519–9523.
9. Hassager C, Bonde SK, Anderson MA, Rink H, Spelsberg TC,Riggs BL 1991 Procalcitonin NH2-terminal cleavage peptidehas no mitogenic effect on normal human osteoblast-like cells.J Bone Miner Res 6:489–493.
10. Oberhoffer M, Vogelsang H, Jager L, Reinhart K 1999 Kata-calcin and calcitonin immunoreactivity in different types ofleukocytes indicate intracellular procalcitonin content. J CritCare 14:29–33.
11. Wiedermann FJ, Innerhofer P, Margreiter J, Fuchs D, Schober-sberger W 1999 Procalcitonin and neopterin in infectiousdiseases. Pteridines 10:125–132.
12. Assicot M, Gendrel D, Carsin H, Raymond J, Guilbaud J,Bohuon C 1993 High serum procalcitonin concentrations inpatients with sepsis and infection. Lancet 341:515–518.
13. Oberhoffer M, Stonans I, Russwurm S, Stonane E, VogelsangH, Junker U, Jager L, Reinhart K 1999 Procalcitonin expres-sion in human peripheral blood mononuclear cells and itsmodulation by lipopolysaccharides and sepsis-related cyto-kines in vitro. J Lab Clin Med 134:49–55.
14. Monneret G, Laroche B, Bienvenu J 1999 Procalcitonin is notproduced by circulating blood cells. Infection 27:34–35.
15. Monneret G, Pachot A, Laroche B, Picollet J, Bienvenu J 2000Procalcitonin and calcitonin gene-related peptide decreaseLPS-induced TNF production by human circulating bloodcells. Cytokine 12:762–764.
16. Wiedermann FJ, Kaneider NC, Egger P, Tiefenthaler W,Wiedermann CJ, Lindner KH, Schobersberger W 2002 Migra-tion of human monocytes in response to procalcitonin. CritCare Med 30:1112–1117.
17. Aida S, Okawa-Takatsuji M, Aotsuka S, Yokohari R 1993Down-regulation by eel calcitonin of interleukin-1 release andproduction by peripheral blood monocytes from patients withrheumatoid arthritis. Clin Exp Rheumatol 11:503–507.
18. Sacerdote P, Bianchi M, Panerai AE 1990 Human monocytechemotactic activity of calcitonin and somatostatin relatedpeptides: Modulation by chronic peptide treatment. J ClinEndocrinol Metab 70:141–148.
19. Schratzberger P, Dunzendorfer S, Reinisch N, Kahler CM,Herold M, Wiedermann CJ 1998 Release of chemoattractantsfor human monocytes from endothelial cells by interactionwith neutrophils. Cardiovasc Res 38:516–521.
20. Wiedermann CJ, Niedermuhlbichler M, Braunsteiner H 1992Priming of polymorphonuclear neutrophils by atrial natriureticpeptide in vitro. J Clin Invest 89:1580–1586.
21. Dunzendorfer S, Wiedermann CJ 2000 Modulation of neutro-phil migration and superoxide anion release by metoprolol. JMol Cell Cardiol 32:915–924.
22. Rosenkranz AR, Schmaldienst S, Stuhlmeier KM, Chen W,Knapp W, Zlabinger GJ 1992 A microplate assay for thedetection of oxidative products using 2�,7�-dichlorofluorescin-diacetate. J Immunol Methods 156:39–45.
23. Wells TN, Power CA, Proudfoot AE 1998 Definition, functionand pathophysiological significance of chemokine receptors.Trends Pharmacol Sci 19:376–380.
24. Richardson RM, Pridgen BC, Haribabu B, Snyderman R 2000Regulation of the human chemokine receptor CCR1. Cross-regulation by CXCR1 and CXCR2. J Biol Chem 275:9201–9208.
25. Lefkowitz RJ 1993 G protein-coupled receptor kinases. Cell74:409–412.
26. Premont RT, Inglese J, Lefkowitz RJ 1995 Protein kinases thatphosphorylate activated G protein-coupled receptors. FASEB J9:175–182.
1881CELLULAR FUNCTION OF KATACALCIN

27. Lohse MJ, Krasel C, Winstel R, Mayor F 1996 G-protein-coupled receptor kinases. Kidney Int 49:1047–1052.
28. Bohm SK, Grady EF, Bunnett NW 1997 Regulatory mecha-nisms that modulate signalling by G-protein-coupled recep-tors. Biochem J 322:1–18.
29. Sterne-Marr R, Benovic JL 1995 Regulation of G protein-coupled receptors by receptor kinases and arrestins. VitamHorm 51:193–234.
30. Ferguson SS, Zhang J, Barak LS, Caron MG 1998 Role ofbeta-arrestins in the intracellular trafficking of G-protein-coupled receptors. Adv Pharmacol 42:420–424.
31. Ruiz-Gomez A, Mayor F 1997 Beta-adrenergic receptor kinase(GRK2) colocalizes with beta-adrenergic receptors duringagonist-induced receptor internalization. J Biol Chem 272:9601–9604.
32. Jones GE 2000 Cellular signaling in macrophage migrationand chemotaxis. Leukoc Biol 68:593–602.
33. Fine JS, Byrnes HD, Zavodny PJ, Hipkin RW 2001 Evaluationof signal transduction pathways in chemoattractant-inducedhuman monocyte chemotaxis. Inflammation 25:61–67.
34. Motulsky HJ, Smith D, Terman BI, Feldman R 1986 Regula-tion of hormone-stimulated cyclic AMP accumulation in intacthuman mononuclear leukocytes by blood plasma. J CyclicNucleotide Protein Phosphor Res 11:329–343.
35. Tanaka H, Minoshima T, Endo N 1995 The effect of a syn-thetic 7-thiaprostaglandin E1 derivative, TEI-6122, on mono-cyte chemoattractant protein-1 induced chemotaxis in THP-1cells. Br J Pharmacol 116:2298–2302.
36. Zuo Q, Claveau D, Hilal G, Leclerc M, Brunette MG 1997Effect of calcitonin on calcium transport by the luminal andbasolateral membranes of the rabbit nephron. Kidney Int 51:1991–1999.
37. Chakraborty M, Chatterjee D, Gorelick FS, Baron R 1994 Cellcycle-dependent and kinase-specific regulation of the apicalNa/H exchanger and the Na,K-ATPase in the kidney cell lineLLC-PK1 by calcitonin. Proc Natl Acad Sci USA 91:2115–2119.
38. Sheykhzade M, Berg Nyborg NC 2001 Mechanisms of CGRP-induced relaxation in rat intramural coronary atreries. Br JPharmacol 132:1235–1246.
39. Armour SL, Foord S, Kenakin T, Chen WJ 1999 Pharmaco-logical characterization of receptor-activity-modifying pro-teins (RAMPs) and the human calcitonin receptor. J PharmacolToxicol Methods 42:217–224.
40. Sozzani S, Locati M, Zhou D, Rieppi M, Luini W, Lamorte G,Bianchi G, Polentarutti N, Allavena P, Mantovani A 1995Receptors, signal transduction, and spectrum of action ofmonocyte chemotactic protein-1 and related chemokines.J Leukoc Biol 57:788–794.
41. Chen Y, Shyu JF, Santhanagopal A, Inoue D, David JP, DixonSJ, Horne WC, Baron R 1998 The calcitonin receptor stimu-lates Shc tyrosine phosphorylation and Erk1/2 activation. In-volvement of Gi, protein kinase C, and calcium. J Biol Chem273:19809–19816.
42. Shyu JF, Zhang Z, Hernandez-Lagunas L, Camerino C, ChenY, Inoue D, Baron R, Horne WC 1999 Protein kinase Cantagonizes pertussis-toxin-sensitive coupling of the calcitoninreceptor to adenylyl cyclase. Eur J Biochem 262:95–101.
43. Naro F, Perez M, Migliaccio S, Galson DL, Orcel P, Teti A,Goldring SR 1998 Phospholipase D- and protein kinase Cisoenzyme-dependent signal transduction pathways activatedby the calcitonin receptor. Endocrinology 139:3241–3248.
44. Chabre O, Conklin BR, Lin HY, Lodish HF, Wilson E, IvesHE, Catanzariti L, Hemmings BA, Bourne HR 1992 A recom-binant calcitonin receptor independently stimulates 3�,5�-cyclic adenosine monophosphate and Ca2�/inositol phosphatesignaling pathways. Mol Endocrinol 6:551–556.
45. Disa J, Parameswaran N, Nambi P, Aiyar N 2000 Involvementof cAMP-dependent protein kinase and pertussis toxin-sensitive G-proteins in CGRP mediated JNK activation inhuman neuroblastoma cell line. Neuropeptides 34:229–233.
46. Dunzendorfer S, Kaser A, Meierhofer C, Tilg H, WiedermannCJ 2001 Cutting edge: Peripheral neuropeptides attract imma-ture and arrest mature blood-derived dendritic cells. J Immunol166:2167–72.
47. Sexton PM, Albiston A, Morfis M, Tilakaratne N 2001 Re-ceptor activity modifying proteins. Cell Signal 13:73–83.
Address reprint requests to:Christian J. Wiedermann, M.D.
Department Internal MedicineUniversity of Innsbruck
Anichstrasse 35A-6020 Innsbruck, Austria
Received in original form August 15, 2001; in revised form March6, 2002; accepted April 17, 2002.
1882 KANEIDER ET AL.
Copyright © 2022 FDOKUMEN




















