
Rapid disassembly of dynamic microtubules upon activation of the capsaicin receptor TRPV1
Upload
independentCategory
view
1download
0
A
(CoiCoim©
Kt
1
tRoDflt
Rls
(
0d
Available online at www.sciencedirect.com
Virus Research 130 (2007) 140–150
Intracellular disassembly of infectious rotavirus particles by depletion ofCa2+ sequestered in the endoplasmic reticulum at the end of virus cycle
Marie Christine Ruiz a,∗, Olga C. Aristimuno a, Yuleima Dıaz a, Franshelle Pena a,Maria Elena Chemello a, Hector Rojas c, Juan Ernesto Ludert b, Fabian Michelangeli a,∗∗
a Laboratorio de Fisiologıa Gastrointestinal, Instituto Venezolano de Investigaciones Cientıficas, Caracas 1020A, Venezuelab Laboratorio de Biologıa de Virus, Instituto Venezolano de Investigaciones Cientıficas, Caracas 1020A, Venezuela
c Laboratorio de Permeabilidad Ionica, Instituto Venezolano de Investigaciones Cientıficas, Caracas 1020A, Venezuela
Received 31 October 2006; received in revised form 31 May 2007; accepted 11 June 2007Available online 23 July 2007
bstract
Rotavirus infection is characterized by a number of Ca2+ dependent virus–cell interactions. The structure of rotavirus triple-layered particlesTLP) is dependent on Ca2+ concentration. Acquisition of the capsid outer layer requires a high Ca2+ concentration inside the ER. Infection modifiesa2+ homeostasis of the cell, increasing ER Ca2+ content, which may be advantageous to virus replication. We studied the role of sequestered Ca2+
n the stabilization of already mature viral particles within the ER. Thapsigargin (TG), a SERCA pump inhibitor, added for 30 min at the end ofnfection depleted ER Ca2+ and reduced the titer of already mature TLP accumulated in the cell. Another inhibitor, cyclopiazonic acid, and twoa2+ ionophores (A23187 and ionomycin) in the presence of EGTA had similar effects. TG eliminated the peak of radiolabeled TLP, increasing that
f DLP in CsCl gradients. Electron microscopy revealed accumulation of clustered particles in the ER, which had lost their integrity. The [Ca2+]n the ER of infected cells is important for virus maturation and for maintaining the integrity of mature TLP. Viral particles in this compartmentay be potentially infectious, already containing VP7 and VP4.2007 Elsevier B.V. All rights reserved.
A11 s
(TtoiPc
op(
eywords: Virus stability; ER calcium; HT29 cells; MA104 cells; OSU strain; Shapsigargin; ER-tracker
. Introduction
Rotavirus is the major etiological agent of viral gastroen-eritis in children and young animals (Kapikian et al., 2001).otavirus infection of the host cell is characterized by a numberf Ca2+ dependent virus–cell interactions (Ruiz et al., 2000).uring the replication cycle, from entry to the release of newly
ormed particles, the forming virions pass through different cel-ular compartments, each one characterized by a distinct [Ca2+]hat is determinant for the replication process.
Rotaviruses are non-enveloped viruses belonging to the
eoviridae family. The infectious virion is an icosahedral triple-ayered particle (TLP) containing a genome formed by 11egments of double-stranded RNA that code for six structural
∗ Corresponding author. Tel.: +58 212 5041164; fax: +58 212 5041093.∗∗ Corresponding author. Tel.: +58 212 5041396; fax: +58 212 5041093.
E-mail addresses: [email protected] (M.C. Ruiz), [email protected]. Michelangeli).
oeibvn
l
168-1702/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.virusres.2007.06.005
train; Ultrastructure; Cyclopiazonic acid; A23187; Ionomycin; BODIPY TR-X
VP) and six non-structural proteins (NSP) (Prasad et al., 1988).he most internal layer of the capsid, formed by VP2, encloses
he genome. The intermediate layer is constituted by VP6. Theuter layer consists of the glycoprotein VP7 and VP4 organizedn dimers or trimers to form 60 spikes (Dormitzer et al., 2004;esavento et al., 2006). The stability of this layer depends onalcium concentration (Shirley et al., 1981).
Rotavirus entry into the cell is accompanied by the lossf VP4 and VP7, thereby converting TLP to double-layeredarticles (DLP). The low Ca2+ concentration in the cytoplasm∼100 nM) is probably responsible for the solubilization of theuter layer and the activation of viral transcription (Chemellot al., 2002; Cohen et al., 1979; Ludert et al., 1987). Rotavirusnfection leads to the formation of perinuclear, non-membrane-ound cytoplasmic inclusions (viroplasms) where synthesized
iral proteins, and dsRNA genome are assembled to form theew DLP (Altenburg et al., 1980).Maturation of new particles proceeds acquiring the outerayer in the endoplasmic reticulum (ER), the main Ca2+ store

esear
oiMEaitbISVaNbotPaC1iaplSts
comadSopsa1
hrtpcCevnilHt(bcm
1aEiIitlHs
Evbtiit
2
2
s(iDg(flc
aLCMccwsg3ci((re
2
M.C. Ruiz et al. / Virus R
f the cell (Estes, 1996). This is a complex process involv-ng numerous steps that have been mainly studied in culture
A104 cells. DLP from the viroplasm bud into the neighboringR through an interaction between the VP6 of DLP and NSP4,glycosylated integral ER membrane protein, which acts as an
ntracellular receptor (Au et al., 1989; Bergmann et al., 1989). Inhis process the particle acquires a transient ER-derived mem-rane (Petrie et al., 1984; Poruchynsky and Atkinson, 1991).solated membrane-enveloped viral intermediates from rotavirusA11-infected MA104 cells comprised the proteins VP1, VP2,P3, VP4, VP6, VP7 and NSP4 (Petrie et al., 1984; Poruchynsky
nd Atkinson, 1991). Subsequent loss of the envelope and itsSP4 component allows condensation of VP7 and the assem-ly of the capsid outer layer. The mechanism of incorporationf VP4 into the layer is as yet not clear. It may occur duringhe budding of the DLP into the ER (Maass and Atkinson, 1990;oruchynsky and Atkinson, 1991). Mature virus is thought to beccumulated in the ER and then released by cell lysis through aa2+ dependent mechanism in MA104 cells (Michelangeli et al.,991; Perez et al., 1998). However, in differentiated and polar-zed Caco2 cells, rotavirus particles seem to be released at thepical pole of the cell without lysis (Jourdan et al., 1997), in arocess dependent on the association of mature viral particles toipid rafts (Cuadras et al., 2006; Cuadras and Greenberg, 2003;apin et al., 2002). On the other hand, it has been proposed that
he acquisition of VP4 takes place post-endoplasmic reticulumite associated to the lipid rafts (Delmas et al., 2004).
Rotavirus morphogenesis is a process dependent on Ca2+
oncentration. During the infection, the experimental disruptionf Ca2+ gradients between the different intracellular compart-ents inhibited the production of infectious particles, observing
n abnormal accumulation of membrane-enveloped interme-iates (Poruchynsky et al., 1991; Shahrabadi et al., 1987;hahrabadi and Lee, 1986). During infection, selective depletionf ER stored calcium by thapsigargin, an inhibitor of the endo-lasmic reticulum calcium pump (SERCA pump), produced theame effect indicating that high Ca2+ concentrations in the ERre required for complete virus maturation (Michelangeli et al.,995).
In addition, infection by itself induces changes in calciumomeostasis of the cell that may be advantageous to viruseplication (Michelangeli et al., 1995, 1991). Among these per-urbations we have measured a progressive increase in Ca2+
lasma membrane permeability, which leads to an elevation ofytosolic Ca2+ concentration and enhancement of sequestereda2+ pools in the ER (Michelangeli et al., 1995, 1991). Thisffect is likely due to the activation of SERCA pumps. Theiral product responsible for the change of Ca2+ permeability isot known. However, NSP4 appears as a likely candidate givents many effects on Ca2+ homeostasis. The inducible intracel-ular expression of NSP4-EGFP fusion protein in mammalianEK 293 cells elevates basal intracellular calcium levels more
han two-fold by a phospholipase C independent mechanism
Berkova et al., 2003). On the other hand, expression of recom-inant NSP4 in insect cells (Sf9) induced a change in Ca2+oncentration, with an increase of Ca2+ permeability of the ERembrane but not of the plasma membrane (Tian et al., 1995,
deea
ch 130 (2007) 140–150 141
994). Contrary to our working hypothesis outlined above, theseuthors proposed that a progressive uncompensated change inR permeability would lead to store depletion and a secondary
ncrease in plasma membrane permeability linked to cell death.f this was the case the particles would be, at a point late innfection, in an environment low in Ca2+, not compatible withhe stability of the TLP outer layer. We also showed that infectioneads to a progressive depletion of agonist-releasable ER pools.owever, this effect does not seem to lead to ER Ca2+ depletion
ince the 45Ca2+ uptake is increased (Ruiz et al., 2005).The question remains as to whether Ca2+ sequestered in the
R is required for the stabilization of the mature and infectiousiral particles accumulated in this compartment during the cycleefore their release by cell lysis or another specific transport. Inhis work, we selectively depleted ER calcium at the end of thenfection period, without blocking maturation, to gain deepernsight into its role in the late stages of virus replication and inhe stability of TLP.
. Material and methods
.1. Cell cultures and virus infection
MA104 cells (embryonic rhesus monkey kidney cells, kindlyupplied by J. Cohen) and the colon carcinoma cells HT29from ATCC) were used. MA104 cells were grown in Min-mum Essential Medium (MEM GIBCO) and HT29 cells inulbecco’s modified Eagle Medium (DMEM containing 4.5 g/llucose, from GIBCO) and maintained as previously describedMichelangeli et al., 1995). Cell monolayers were grown to con-uency in 24-well Linbro plates for 3–4 days. At this stage, HT29ells were not differentiated (Ciarlet et al., 2001).
The OSU strain of rotavirus (porcine, serotype P9[7]G5[5]),nd SA11 (Simian Agent-11, clone 3) kindly supplied by F.iprandi (Instituto Venezolano de Investigaciones Cientıficas,aracas, Venezuela) were used. Rotaviruses were replicated inA104 cells in the presence of trypsin (1 �g/ml). Infection was
arried out for 1 h at 37 ◦C. Then, the inoculum was removed, theells were washed with PBS and further incubated with MEMithout fetal calf serum until complete cytopathic effect. Cell
uspensions were frozen–thawed twice and clarified by centrifu-ation. Viral suspensions were treated with 10 �g/ml trypsin for0 min at 37 ◦C to cleave VP4 (Trypsin Type IX; Sigma Chemi-al Co.). Infectivity of the preparations was measured by titrationn microplates in MA104 cells using a monoclonal antibody4B2D2), directed against a common group A antigen of VP6kindly supplied by F. Liprandi (Liprandi et al., 1990) for indi-ect immunofluorescence staining after methanol fixation andxpressed as fluorescent-focus units (FFU) (Ciarlet et al., 1995).
.2. 45Ca2+ uptake
The cell uptake of 45Ca2+ was measured as previously
escribed (Michelangeli et al., 1991). Briefly, cell monolay-rs were grown to confluency in 24-well Linbro plates. At thend of the infection period maintenance medium was removednd replaced by 200 �l of MEM containing 45Ca2+ (1 �Ci per
1 esear
wt3odtvt
2
bAisiBTc(tP
cTonu
2t
oaaiabwltatwcuoo
2c
wi
n3orwp3twRsp1o5psfiC3tm
2
pwpgg(
2
aspoltau
3
3c
c
42 M.C. Ruiz et al. / Virus R
ell). Uptake was stopped after 10 min by washing the cells fourimes by immersion in ice-cold PBS. After drying for 30 min at7 ◦C, cells were dissolved in 250 �l of 0.1% SDS in 0.1N NaOHvernight and then neutralized with 0.5N HCl. Radioactivity wasetermined in 40 �l aliquots by liquid scintillation counting. Ashe number of cells per well was found to be constant, uptakealues for single experiments are normalized and expressed ashe relation to the counts in mock-infected wells.
.3. Confocal microscopy
MA104 cells were grown to 90% confluence on cover slipottom culture plates and infected with OSU strain of rotavirus.t 1 h post-infection the inoculum was removed at 1 h post-
nfection and replaced by MEM (200 �l). At 6 HPI cells weretained according to the protocols that follow. To study the local-zation of SERCA Ca2+ pumps, live cells were incubated withODIPY TR-X thapsigargin (1 �M, 5 min, Molecular Probes).o confirm that thapsigargin binds to the ER compartment, weompared the distribution of fluorescently labeled thapsigarginBODIPY TR-X Tg) in mock and rotavirus-infected cell withhat of ER-tracker, an ER marker (2.5 �M, 5 min, Molecularrobes).
The dual-labeled cells were placed on a laser scanning confo-al microscope stage (LSCM, Nikon C1), mounted in a EclipseE300 Nikon inverted microscope with Nikon 60/1.40 PlanApoil-immersion objective coupled to a C1-LU2 laser unit witheon (543 nm) and argon-cooled air (488 nm) lasers. These lasernits were controlled by a D-eclipse C1 interface.
.4. Quantification of cell-associated viruses after a shortreatment with thapsigargin
Cell monolayers were infected at 37 ◦C, with a multiplicityf infection of approximately 20 FFU/cell. In these conditions,ll cells of the monolayer were infected during the first cycle asscertained by detection of VP6 by immunofluorescence. Thenoculum was removed at 1 h post-infection and infection wasllowed to proceed for different periods of time. Thirty minutesefore the end of a given infection period, the monolayer wasashed with PBS containing 1 mM EGTA to remove extracellu-
ar viruses. Wells were replenished with MEM containing 3 �Mhapsigargin (LC Laboratories). Another set of infected wells,cting as a control, was incubated with MEM until the end ofhe infection period. After 30 min, the monolayers were washedith PBS and frozen to titrate cell-associated infectious parti-
les. In another set of experiments, instead of thapsigargin, wesed cyclopiazonic acid (5 �M) or the Ca2+ ionophores A23187r ionomycin, in the presence of 1.8 mM EGTA, with the aimf depleting Ca2+ from intracellular reservoirs.
.5. Separation of virus particles by isopycnicentrifugation
MA104 cells grown in Falcon flask (175 cm2) were infectedith OSU rotavirus with 20 FFU/cell (3 ml). At 1 HPI, the
noculum was removed and replaced by MEM without methio-
aieo
ch 130 (2007) 140–150
ine. At 3 HPI the medium was replaced by MEM containing5S-methionine (50 �Ci/ml). Thirty minutes before the endf the experiment (6 HPI), the medium was removed andeplaced by one with or without 3 �M thapsigargin. Cellsere then washed three times with PBS, scraped and resus-ended in PBS (150 mM NaCl, 1 mM MgSO4, 1 mM CaCl2,M K2HPO4, 0.6 M NaH2PO4). Cells were subjected to
hree cycles of freeze–thawing. Nuclei and larger debrisere removed from the lysates by low-speed centrifugation.adiolabeled viral particles were extracted with Freon 113
eparately from the supernatant and the pellet fraction resus-ended in a buffered medium (10 mM Tris–HCl [pH 7.8],00 mM NaCl, 10 mM CaCl2, 2 mM phenylmethylsulfonyl flu-ride, and 1% Nonidet P-40). After centrifugation (2 min at000 × g) the respective supernatants were pooled and virusarticles were concentrated by ultracentrifugation on a 45%ucrose cushion at 30,000 × g in a Beckman SW40Ti rotoror 4 h. The pellet was resuspended in a medium contain-ng 50 mM Tris–HCl [pH 7.4], 87 mM NaCl and 10 mMaCl2, and banded in a CsCl gradient (SW40Ti rotor, 48 h,5,000 × g). Fractions (400 �l) were collected and their radioac-ivity (by liquid scintillation counting) and density were
easured.
.6. Analysis of labeled proteins by PAGE
Fractions corresponding to the radioactivity peaks wereooled and analyzed by SDS-PAGE. Fractions were dilutedith PBS, centrifuged (30,000 × g, 90 min) and the pellet resus-ended in 50 �l of PBS. Sodium dodecyl sulfate polyacrylamideel electrophoresis (SDS-PAGE) on 12.5% polyacrylamideels and autoradiography were performed as described beforeGorziglia et al., 1985).
.7. Electron microscopy of infected cells
MA104 cells were grown in 25 cm2 Falcon flask and infecteds described above. Infected cells were treated or not with thap-igargin from 1 h or at 30 min before the end of the infectioneriod. At 6 HPI, the cells were scraped, washed with PBS, fixedvernight at 4 ◦C with 2.5% glutaraldehyde in 0.1 M caccody-ate buffer containing 10 mM CaCl2 and postfixed with osmiumetraoxyde (1%). Fixed cells were resin embedded, sectionednd viewed under the electron microscope after staining withranyl acetate and lead citrate.
. Results
.1. Release of accumulated Ca2+ in the ER of infectedells by thapsigargin treatment
In the experiments presented in Fig. 1, we measured the timeourse of the increase of total calcium content during infection
nd the effect of thapsigargin added 30 min before the end of thenfection period. As we have previously shown (Michelangelit al., 1991), rotavirus infection induced a progressive increasef 45Ca2+ uptake in MA104 cells, which reached almost eight-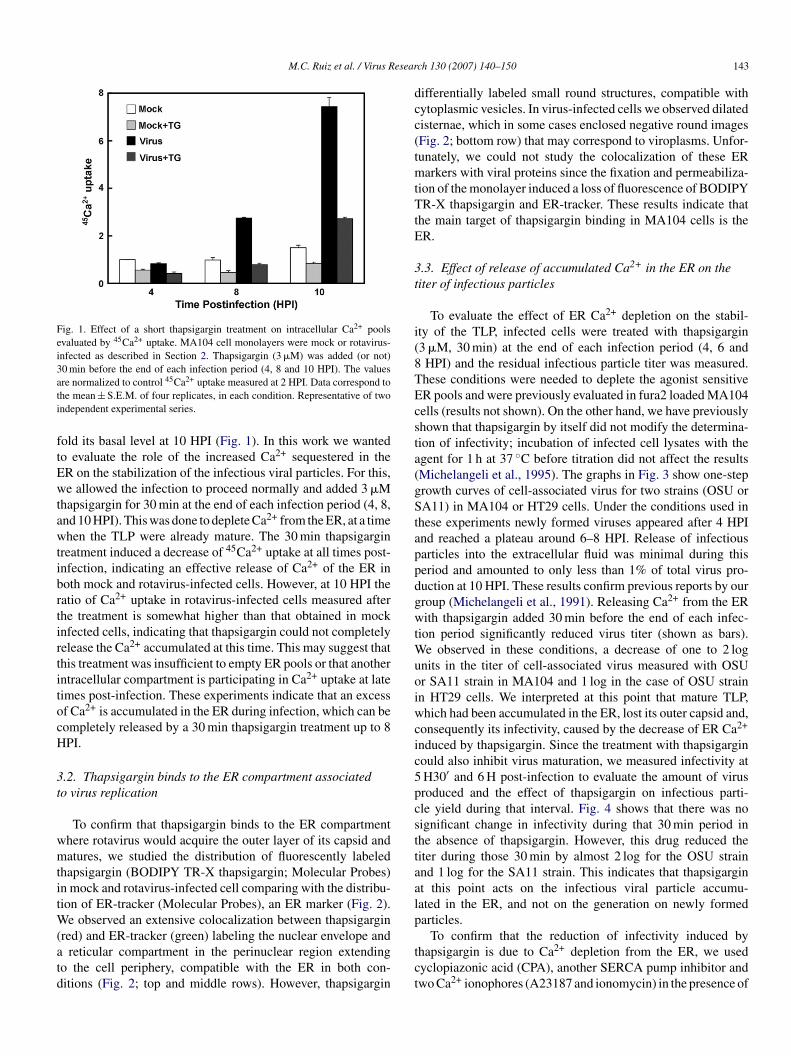
M.C. Ruiz et al. / Virus Resear
Fig. 1. Effect of a short thapsigargin treatment on intracellular Ca2+ poolsevaluated by 45Ca2+ uptake. MA104 cell monolayers were mock or rotavirus-infected as described in Section 2. Thapsigargin (3 �M) was added (or not)30 min before the end of each infection period (4, 8 and 10 HPI). The valuesati
ftEwtawtibrtirtitocH
3t
wmtitW(atd
dcc(tmtTtE
3t
i(8TEcsta(gStappdgwtWuoiwcic5pcsttaalp
re normalized to control 45Ca2+ uptake measured at 2 HPI. Data correspond tohe mean ± S.E.M. of four replicates, in each condition. Representative of twondependent experimental series.
old its basal level at 10 HPI (Fig. 1). In this work we wantedo evaluate the role of the increased Ca2+ sequestered in theR on the stabilization of the infectious viral particles. For this,e allowed the infection to proceed normally and added 3 �M
hapsigargin for 30 min at the end of each infection period (4, 8,nd 10 HPI). This was done to deplete Ca2+ from the ER, at a timehen the TLP were already mature. The 30 min thapsigargin
reatment induced a decrease of 45Ca2+ uptake at all times post-nfection, indicating an effective release of Ca2+ of the ER inoth mock and rotavirus-infected cells. However, at 10 HPI theatio of Ca2+ uptake in rotavirus-infected cells measured afterhe treatment is somewhat higher than that obtained in mocknfected cells, indicating that thapsigargin could not completelyelease the Ca2+ accumulated at this time. This may suggest thathis treatment was insufficient to empty ER pools or that anotherntracellular compartment is participating in Ca2+ uptake at lateimes post-infection. These experiments indicate that an excessf Ca2+ is accumulated in the ER during infection, which can beompletely released by a 30 min thapsigargin treatment up to 8PI.
.2. Thapsigargin binds to the ER compartment associatedo virus replication
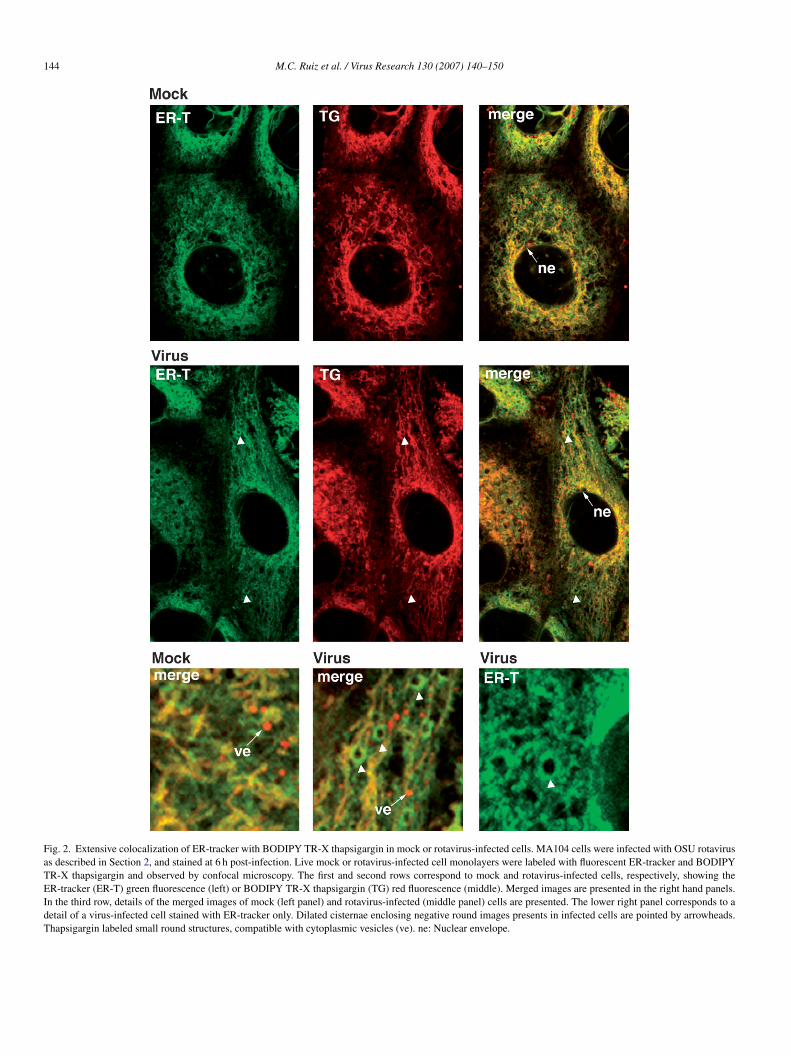
To confirm that thapsigargin binds to the ER compartmenthere rotavirus would acquire the outer layer of its capsid andatures, we studied the distribution of fluorescently labeled
hapsigargin (BODIPY TR-X thapsigargin; Molecular Probes)n mock and rotavirus-infected cell comparing with the distribu-ion of ER-tracker (Molecular Probes), an ER marker (Fig. 2).
e observed an extensive colocalization between thapsigargin
red) and ER-tracker (green) labeling the nuclear envelope andreticular compartment in the perinuclear region extendingo the cell periphery, compatible with the ER in both con-itions (Fig. 2; top and middle rows). However, thapsigargin
tct
ch 130 (2007) 140–150 143
ifferentially labeled small round structures, compatible withytoplasmic vesicles. In virus-infected cells we observed dilatedisternae, which in some cases enclosed negative round imagesFig. 2; bottom row) that may correspond to viroplasms. Unfor-unately, we could not study the colocalization of these ER
arkers with viral proteins since the fixation and permeabiliza-ion of the monolayer induced a loss of fluorescence of BODIPYR-X thapsigargin and ER-tracker. These results indicate that
he main target of thapsigargin binding in MA104 cells is theR.
.3. Effect of release of accumulated Ca2+ in the ER on theiter of infectious particles
To evaluate the effect of ER Ca2+ depletion on the stabil-ty of the TLP, infected cells were treated with thapsigargin3 �M, 30 min) at the end of each infection period (4, 6 andHPI) and the residual infectious particle titer was measured.hese conditions were needed to deplete the agonist sensitiveR pools and were previously evaluated in fura2 loaded MA104ells (results not shown). On the other hand, we have previouslyhown that thapsigargin by itself did not modify the determina-ion of infectivity; incubation of infected cell lysates with thegent for 1 h at 37 ◦C before titration did not affect the resultsMichelangeli et al., 1995). The graphs in Fig. 3 show one-steprowth curves of cell-associated virus for two strains (OSU orA11) in MA104 or HT29 cells. Under the conditions used in
hese experiments newly formed viruses appeared after 4 HPInd reached a plateau around 6–8 HPI. Release of infectiousarticles into the extracellular fluid was minimal during thiseriod and amounted to only less than 1% of total virus pro-uction at 10 HPI. These results confirm previous reports by ourroup (Michelangeli et al., 1991). Releasing Ca2+ from the ERith thapsigargin added 30 min before the end of each infec-
ion period significantly reduced virus titer (shown as bars).e observed in these conditions, a decrease of one to 2 log
nits in the titer of cell-associated virus measured with OSUr SA11 strain in MA104 and 1 log in the case of OSU strainn HT29 cells. We interpreted at this point that mature TLP,hich had been accumulated in the ER, lost its outer capsid and,
onsequently its infectivity, caused by the decrease of ER Ca2+
nduced by thapsigargin. Since the treatment with thapsigarginould also inhibit virus maturation, we measured infectivity atH30′ and 6 H post-infection to evaluate the amount of virusroduced and the effect of thapsigargin on infectious parti-le yield during that interval. Fig. 4 shows that there was noignificant change in infectivity during that 30 min period inhe absence of thapsigargin. However, this drug reduced theiter during those 30 min by almost 2 log for the OSU strainnd 1 log for the SA11 strain. This indicates that thapsigargint this point acts on the infectious viral particle accumu-ated in the ER, and not on the generation on newly formedarticles.
To confirm that the reduction of infectivity induced byhapsigargin is due to Ca2+ depletion from the ER, we usedyclopiazonic acid (CPA), another SERCA pump inhibitor andwo Ca2+ ionophores (A23187 and ionomycin) in the presence of
144 M.C. Ruiz et al. / Virus Research 130 (2007) 140–150
Fig. 2. Extensive colocalization of ER-tracker with BODIPY TR-X thapsigargin in mock or rotavirus-infected cells. MA104 cells were infected with OSU rotavirusas described in Section 2, and stained at 6 h post-infection. Live mock or rotavirus-infected cell monolayers were labeled with fluorescent ER-tracker and BODIPYTR-X thapsigargin and observed by confocal microscopy. The first and second rows correspond to mock and rotavirus-infected cells, respectively, showing theER-tracker (ER-T) green fluorescence (left) or BODIPY TR-X thapsigargin (TG) red fluorescence (middle). Merged images are presented in the right hand panels.In the third row, details of the merged images of mock (left panel) and rotavirus-infected (middle panel) cells are presented. The lower right panel corresponds to adetail of a virus-infected cell stained with ER-tracker only. Dilated cisternae enclosing negative round images presents in infected cells are pointed by arrowheads.Thapsigargin labeled small round structures, compatible with cytoplasmic vesicles (ve). ne: Nuclear envelope.

M.C. Ruiz et al. / Virus Research 130 (2007) 140–150 145
Fig. 3. Effect of ER Ca2+ released by thapsigargin at the end of infection period on infectious particle yield. MA104 or HT29 cell monolayers were infected byOSU or SA11 rotavirus strain. Thirty minutes before the end of each infection period, the medium was replaced by one containing (or not) 3 �M thapsigargin. Then,the monolayers were washed twice and frozen for infectious particle titration in MA104 cell monolayers. In control conditions without thapsigargin treatment ao circlr ponda
Eelemttco
FibIttFdi
3treatment with thapsigargin
To determine whether the outer layer of TLP had been solubi-lized during the 30-min thapsigargin treatment, we analyzed the
ne-step growth curve was performed by titration of cell-associated virus (closedepresented by hatched bars. The results are expressed as FFU/ml. Values corresnd MA104/SA11 and, n = 7 for five independent experiments in HT29/SA11).
GTA (1.8 mM) to empty the cell reservoirs (Fig. 5). In parallelxperiments measuring cytoplasmic Ca2+ concentration in fura2oaded MA104 cells we could confirm that these treatmentsffectively released ER Ca2+ (not shown). These experimentalaneuvers at the end of the infection period induced a reduc-
ion of infectious virus associated to the cells. The fact that these
reatments deplete Ca2+ reservoirs by different mechanisms indi-ates that Ca2+ depletion of the ER is responsible for the effectf thapsigargin on virus titer.5
6
7
8
9
OSU SA11
Control 5:30 HPI
Control 6:00 HPI
TG 5:30-6:00 HPI
Infe
ctiv
ity
(lo
gF
FU
/ml)
ig. 4. Evaluation of the amount of virus produced between 5 H30′ and 6 H post-nfection and the effect of thapsigargin. MA104 cell monolayers were infectedy OSU or SA11 rotavirus strains. Infection was carried out for 5 H30′ or 6 H.n one condition, the medium was replaced at 5 H30′ by one containing 3 �Mhapsigargin. Then, the monolayers were washed twice and frozen for infec-ious particle titration in MA104 cell monolayers. The results are expressed asFU/ml. The values correspond to the mean ± S.E.M. (n = 9 for three indepen-ent experiments in MA104/OSU and, n = 6 for two independent experimentsn MA104/SA11).
FtMbiEwlpBaf
es, continuous line). The residual infectivity titer after thapsigargin treatment isto the mean ± S.E.M. (n = 10 for five independent experiments in MA104/OSU
.4. Analysis of viral particle types obtained after short
ig. 5. Effect of ER Ca2+ released by cyclopiazonic acid and Ca2+ ionophores inhe presence of EGTA at the end of infection period on infectious particle yield.
A104 cell monolayers were infected by OSU rotavirus strains. Thirty minutesefore the end of each infection period, the medium was replaced by one contain-ng (or not) either cyclopiazonic acid (CPA, 10 �M), A23187 (5 �M) + 1.8 mMGTA or ionomycin (5 �M) + 1.8 mM EGTA. Then, the monolayers wereashed twice and frozen for infectious particle titration in MA104 cell mono-
ayers. In control conditions without treatments a one-step growth curve waserformed by titration of cell-associated virus (closed circles, continuous line).ars represent the residual infectivity titer after different treatments. The resultsre expressed as FFU/ml. The values correspond to the mean ± S.E.M. (n = 4or two independent experiments).

146 M.C. Ruiz et al. / Virus Resear
Fig. 6. Analysis of viral particles isolated from infected cells treated with thap-sigargin. MA104 cells were infected with OSU rotavirus; at 3 HPI the mediumwas replaced by one containing 35S-methionine. Thirty minutes before the endof the infection period (6 HPI) 3 �M thapsigargin was added. Virus particlesextracted from cell lysates were separated by CsCl centrifugation as describedin Section 2. Gradient fractions were analyzed for density and radioactivity (Aand B). The positions of TLP (1.36 g/ml), DLP (1.38 g/ml) and empty particlesin gradients are indicated. The fractions with a radioactivity peak correspondingto the density of different types of particles were pooled and proteins analyzedby SDS-PAGE (C). Lanes 1 and 2 correspond to fractions 10–13 and 14–17 fromvae
vcwdTistoip
F(puwVsctiwedVtw(
3b
tptcsspiTvcwiclttmIoc(cbtd
4
C
irus-infected cells, respectively. Lanes 3 and 4 correspond to fractions 10–13nd 26–28 from thapsigargin treated infected cells, respectively. Representativexperiment of two.
iral particle types after separation in CsCl gradients. MA104ells were infected for 6 h and the newly synthesized proteinsere radiolabeled with 35S-methionine. Fractions from the gra-ients were then analyzed for radioactivity and density (Fig. 6A).he radioactivity measurements in the gradient fractions from
nfected cells show a main peak at a density of 1.36 g/ml and amall one at 1.38 g/ml corresponding to TLP and DLP, respec-
ively. By comparison, the TLP peak disappeared as an effectf the short thapsigargin treatment, with a concomitant increasen the peak corresponding to DLP. We could also see a neweak that corresponded to empty particles (around 1.31 g/ml;irtt
ch 130 (2007) 140–150
ig. 6B). The PAGE analysis of fractions around these peaks10–13, 14–17, and 26–28) confirmed the identity of particlesresent in the different peaks (Fig. 6C). In fractions 14–17 ofntreated infected cells, aside from VP1, VP2, VP3 and VP6,e could identify the outer layer proteins, VP4 (and VP5*) andP7, thus corresponding to TLP. In the 10–13 fractions, corre-
ponding to DLP density, a small amount of VP7 but not VP4ould be observed. The small amount of VP7 is probably dueo overlap of the DLP and TLP bands in the fraction harvest-ng (Fig. 6C, lane 1). In the 10–13 fractions from cells treatedith thapsigargin, neither VP4 nor VP7 were detected. Inter-
stingly, treatment with thapsigargin produced empty particlesetected in the fractions 26–28, in which VP1, VP2, VP3 andP6 could be identified. Furthermore, the identity of viral par-
icles in the different bands in the fractions of CsCl gradientsas confirmed by electron microscopy after negative staining
results not shown).
.5. Effect of thapsigargin on particle structure as assessedy electron microscopy
The impact of Ca2+ depletion of ER by a 30-min incuba-ion with thapsigargin (3 �M) before the end of the infectioneriod (6 HPI) on particle structure was evaluated by elec-ron microscopy of infected cells (Fig. 7). Untreated infectedells present electron-dense structures in the cytoplasm, corre-ponding to viroplasms, in close apposition to dilated ribosometudded cisternae of rough ER (RER) (Fig. 7A). Double-layeredarticles emerging from the viroplasm were observed buddingnto the RER, and acquiring a membrane during this process.hese membrane-enveloped particles (MEP) were seen near theiroplasms or in proximity to the RER membrane. Towards theenter of dilated cisternae, TLP devoid of enveloping membrane,ith a mean diameter of 75 nm were present (Fig. 7A and D). As
t has been reported before, thapsigargin treatment of infectedells from 1 to 6 HPI (Fig. 7B) induced an abnormal accumu-ation of membrane-enveloped particles within the cisternae ofhe RER (Michelangeli et al., 1995). Therefore, the addition ofhapsigargin early in the infection (1 HPI) seems to arrest the
orphogenesis of rotavirus at the membrane-enveloped stage.n the case of cells treated with thapsigargin for 30 min at the endf infection (6 HPI), membrane-enveloped particles were seenlose to the RER membrane similar to the untreated conditionsFig. 7C and E). However, we can observe an accumulation oflustered particles, with a defined electron-dense core separatedy an unidentifiable amorphous material. These images suggesthat mature TLP had lost their capsid integrity as a result of Ca2+
epletion (Fig. 7E).
. Discussion
The structure of rotavirus infectious particles is dependent ona2+ concentration. Ca2+ has a stabilizing effect on the physical
ntegrity and infectivity of rotavirus (Shirley et al., 1981). In aotavirus suspension, Ca2+ chelation results in the removal ofhe outer layer of the capsid with a concomitant loss of infec-ivity (Cohen et al., 1979; McCrae and Faulkner-Valle, 1981).

M.C. Ruiz et al. / Virus Research 130 (2007) 140–150 147
Fig. 7. Effect of thapsigargin on the ultrastructure of viral particles in rotavirus-infected MA104 cells. Monolayers were infected by OSU rotavirus as described inSection 2. (A) In untreated infected cells double-layered particles can be observed emerging from the viroplasm (V), budding into the RER, acquiring ER membrane.These membrane-enveloped particles (MEP) can be seen near the viroplasm or in proximity to the RER membrane. Towards the center of dilated cisternae, triple-layerparticles (TLP) devoid of enveloping membrane can be observed. (B) In infected cells treated with 1 �M thapsigargin from 1 HPI, viroplasms and images of particlebudding into the RER are also observed; only membrane-enveloped particles are accumulated within the cisternae of the RER. (C) In infected cells treated with3 velop( of pa( ).
Ftidla
cD
�M thapsigargin for 30 min before the end of infection (6 HPI), membrane-encl) separated by an amorphous material can be observed. In (D and E) details30 min, in E). Bars correspond to 200 nm in (A–C), and to 100 nm in (D and E
urthermore, Ca2+ was shown to be an integral constituent ofhe outer layer of TLP (Shahrabadi et al., 1987). In experiments
n vitro, a low Ca2+ concentration in the range of 20–600 nM,epending on the strain, induces the solubilization of the outerayer of the capsid (Ruiz et al., 1996). This phenotype is associ-ted to VP7 where two proline residues are found in a putativetcf2
ed particles can be observed. Instead of TLP, a cluster of electron-dense coresrticles in the RER of untreated infected cells (D) or treated with thapsigargin
alcium-binding site (Gajardo et al., 1997; Ruiz et al., 1996).uring its entry into the cell, rotavirus may take advantage of
he low Ca2+ concentration in the cytoplasm or in an endocyticompartment to loose its capsid outer layer. TLP would trans-orm to DLP, active for the transcription process (Ruiz et al.,000).

1 esear
tvfwT(tCta
pclSptptaepmcvo(Ctp
rtvawbpi
3itfimaflctspNioe
(opr
itRtctvoC
eap2eotbwm(Co
dbanctdcmmto(a
iitMtpiUg
48 M.C. Ruiz et al. / Virus R
Viral transcription, translation and RNA replication occur inhe cytoplasm, where assembly of new particles takes place iniroplasms up to the DLP stage. The maturation of the particlesollows by budding of the DLP through the RER membrane,here the two viral glycoproteins, VP7 and NSP4 are localized.he third layer of the capsid is acquired during this process
Altenburg et al., 1980; Bergmann et al., 1989). Since the struc-ure of the TLP is not compatible with physiological cytoplasmica2+ concentrations (50–100 nM), it seems logical to think that
he acquisition of the outer layer of the capsid must take place incompartment with a high Ca2+ concentration, such as the ER.
The role of Ca2+ in rotavirus morphogenesis has been studiedreviously. Reduction of extracellular Ca2+ below a critical con-entration (0.17 mM) drastically reduced the virus titer and VP7evels due to degradation of this protein (Shahrabadi et al., 1987;hahrabadi and Lee, 1986). Calcium depletion of all cell com-artments using a Ca2+ ionophore in calcium-free medium alonghe infection cycle (from 1 HPI) also inhibited the infectiousarticle yield (Poruchynsky et al., 1991). This treatment alteredhe formation of the hetero-oligomeric complex of NSP4, VP7,nd VP4 in the ER and induced an accumulation of membrane-nveloped intermediate particles (Poruchynsky et al., 1991). In arevious work, we evaluated the role of ER Ca2+ pools in virusaturation. We showed that selective depletion of ER stored
alcium by the addition of thapsigargin at the beginning of theiral cycle (1 HPI), arrested rotavirus maturation at the stagef membrane-enveloped particles that accumulated in the ERMichelangeli et al., 1995). These results suggested that the higha2+ concentration of the ER in infected cells was required for
he final assembly of the outer layer of TLP containing VP7 and,erhaps VP4.
The present study represents a different approach, withespect to our previous work (Michelangeli et al., 1995), aimingo elucidate the importance of ER Ca2+ in the late stages of theiral cycle and rotavirus morphogenesis when TLP were alreadyssembled and accumulated in this compartment. The strategyas to deplete Ca2+ from the sites where mature virus wereeing accumulated. For this we used thapsigargin, a sesquiter-ene lactone isolated from the plant Thapsia garganica, whichs a specific SERCA pump inhibitor (Thastrup et al., 1990).
The treatment of infected cells with 3 �M thapsigargin for0 min induced a decrease of 45Ca2+ uptake down to mocknfected values. We used this high drug concentration to be surehat the ER Ca2+ pools were effectively depleted within the timerame selected for this experiment. This treatment must havenduced a Ca2+ depletion of the ER and any other compart-ent where SERCA pumps may be localized, such as the Golgi
pparatus (Van Baelen et al., 2004). Extensive colocalization ofuorescent thapsigargin and the ER marker (ER-tracker) indi-ates that most of the SERCA pumps in MA104 cells reside inhe ER. However, differential labeling of vesicle-like structuresuggest the presence of SERCA pumps in another compartment,erhaps Golgi-associated vesicles (Van Baelen et al., 2004).
evertheless, these structures did not seem modified by virusnfection. On the other hand, in the ER of infected cell we couldbserve dilated cisternae enclosing structures not stained withither fluorescent marker, which may correspond to viroplasms
Emso
ch 130 (2007) 140–150
Fig. 2). These observations are in accordance with the imagesbtained by electron microscopy which show that rotavirusarticles are budding from viroplasms and accumulated intoibosome studded dilated ER cisternae (Fig. 7A).
Thapsigargin treatment induced a drop of the number ofnfectious particles that had been already produced at any timeested. This effect seems to be due to Ca2+ depletion of theER where virus particles are localized. Thapsigargin appears
o be acting by a specific inhibition of the SERCA pump sinceyclopiazonic acid (CPA), another SERCA pump inhibitor hadhe same effect. Furthermore, the reduction of cell-associatedirus in MA104 cells obtained with Ca2+ ionophores (A23187r ionomycin) in the presence of EGTA (1.8 mM) confirms thata2+ depletion is responsible for the loss of infectivity.
Depletion of ER Ca2+ may affect viral progeny through differ-nt mechanisms: decapsidation of TLP, arresting of maturation,nd inhibition of protein synthesis by activation of the unfoldedrotein response (UPR) of the cell (Brostrom and Brostrom,003; Harding et al., 2002). If the effect of thapsigargin at thend of viral cycle on the infectious titer were due to the arrestingf virus maturation or inhibition of protein synthesis, the virusiter measured at 5 H30′ post-infection without treatment shoulde the same as that obtained at 6 HPI after the 30 min treatmentith the drug. This was not the case; instead, thapsigargin treat-ent decreased the number of infectious particles by over 90%
almost 2 log). This supports the contention that the effect of ERa2+ depletion on virus titer is the solubilization of the capsiduter layer of accumulated TLP.
The purification and separation by CsCl gradient of theifferent types of viral particles obtained after the short incu-ation with thapsigargin confirmed that this treatment produced
solubilization of the outer layer, transforming TLP intoon-infectious DLP. The electron micrographs show, in theseonditions, the presence of membrane-enveloped particles inhe RER, and an accumulation of particles with a morphologyifferent to those present in untreated conditions. These parti-les were clustered and separated by an amorphous material thatay correspond to precipitated proteins from the outer layer. Theembrane-enveloped particles appeared to be stable in condi-
ions where the RER has been depleted of Ca2+. This had beenbserved previously using a long treatment with thapsigarginfrom 1 HPI) (Michelangeli et al., 1995) or a medium containingCa2+ ionophore with EGTA (Poruchynsky et al., 1991).
It is interesting to note that the loss of infectious capacitynduced by RER Ca2+ depletion suggests that the viral particlesn this compartment are mature and infectious, already con-aining VP7 and VP4. This may suggest that in non-polarized
A104 and HT29 cells, VP4 is acquired in the ER. This con-ention is in accordance with the results of Cuadras et al. whoroposed that assembly of VP4 into mature virions takes placen the late stages of transit through the ER (Cuadras et al., 2006).nfortunately, we could not study the colocalization of thapsi-argin with viral proteins since BODIPY TR-X thapsigargin and
R-Tracker fluorescence disappeared upon cell fixation and per-eabilization. On the other hand, the involvement of an atypicalub-compartment of the ER containing raft, could not be ruledut (Chwetzoff and Trugnan, 2006; Delmas et al., 2004).

esear
tib
A
maJsygET
R
A
A
B
B
B
C
C
C
C
C
C
C
D
D
E
G
G
H
J
K
L
L
M
M
M
M
P
P
P
P
P
P
R
R
R
S
M.C. Ruiz et al. / Virus R
In conclusion, the evidence presented in this paper supportshe role of the high Ca2+ concentration stored in the RER ofnfected cells not only in the maturation of infectious particlesut also in their stability before release from the cell.
cknowledgments
The authors wish to thank Dr. C. Sapin for constructive com-ents on the manuscript, Y. Higuerey, M. Romano. F. Sanchez
nd M. Granados for technical assistance and M. Diaz and. Rivas of Servicio de Fotografıa Cientifica (IVIC). Projectupported by Consejo Nacional de Investigaciones Cientıficas
Tecnologicas (FONACIT), Venezuela through FONACITrants (S1-2001000329, G2001000637) and the FONACIT-COSNORD international program (PI 2002000905) and byOTAL Venezuela S.A.
eferences
ltenburg, B.C., Graham, D.Y., Estes, M.K., 1980. Ultrastructural study ofrotavirus replication in cultured cells. J. Gen. Virol. 46, 75–85.
u, K.S., Chan, W.K., Burns, J.W., Estes, M.K., 1989. Receptor activity ofrotavirus nonstructural glycoprotein NS28. J. Virol. 63, 4553–4562.
ergmann, C.C., Maass, D., Poruchynsky, M.S., Atkinson, P.H., Bellamy, A.R.,1989. Topology of the non-structural rotavirus receptor glycoprotein NS28in the rough endoplasmic reticulum. Embo. J. 8 (6), 1695–1703.
erkova, Z., Morris, A.P., Estes, M.K., 2003. Cytoplasmic calcium measurementin rotavirus enterotoxin-enhanced green fluorescent protein (NSP4-EGFP)expressing cells loaded with Fura-2. Cell Calcium 34 (1), 55–68.
rostrom, M.A., Brostrom, C.O., 2003. Calcium dynamics and endoplasmicreticular function in the regulation of protein synthesis: implications for cellgrowth and adaptability. Cell Calcium 34 (4–5), 345–363.
hemello, M.E., Aristimuno, O.C., Michelangeli, F., Ruiz, M.C., 2002. Require-ment for vacuolar H+ -ATPase activity and Ca2+ gradient during entry ofrotavirus into MA104 cells. J. Virol. 76 (24), 13083–13087.
hwetzoff, S., Trugnan, G., 2006. Rotavirus assembly: an alternative model thatutilizes an atypical trafficking pathway. Curr. Top. Microbiol. Immunol. 309,245–261.
iarlet, M., Crawford, S.E., Estes, M.K., 2001. Differential infection of polarizedepithelial cell lines by sialic acid-dependent and sialic acid-independentrotavirus strains. J. Virol. 75 (23), 11834–11850.
iarlet, M., Ludert, J.E., Liprandi, F., 1995. Comparative amino acid sequenceanalysis of the major outer capsid protein (VP7) of porcine rotaviruses withG3 and G5 serotype specificities isolated in Venezuela and Argentina. Arch.Virol. 140 (3), 437–451.
ohen, J., Laporte, J., Charpilienne, A., Scherrer, R., 1979. Activation ofrotavirus RNA polymerase by calcium chelation. Arch. Virol. 60, 177–186.
uadras, M.A., Bordier, B.B., Zambrano, J.L., Ludert, J.E., Greenberg, H.B.,2006. Dissecting rotavirus particle-raft interaction with small interferingRNAs: insights into rotavirus transit through the secretory pathway. J. Virol.80 (8), 3935–3946.
uadras, M.A., Greenberg, H.B., 2003. Rotavirus infectious particles use lipidrafts during replication for transport to the cell surface in vitro and in vivo.Virology 313 (1), 308–321.
elmas, O., Gardet, A., Chwetzoff, S., Breton, M., Cohen, J., Colard, O., Sapin,C., Trugnan, G., 2004. Different ways to reach the top of a cell. Analysis ofrotavirus assembly and targeting in human intestinal cells reveals an originalraft-dependent, Golgi-independent apical targeting pathway. Virology 327(2), 157–161.
ormitzer, P.R., Nason, E.B., Prasad, B.V., Harrison, S.C., 2004. Structuralrearrangements in the membrane penetration protein of a non-envelopedvirus. Nature 430 (7003), 1053–1058.
stes, M.K., 1996. Rotaviruses and their replication. In: Fields, B.N., Knipe,P.M., Howley, P.M., Chanock, R.M., Melnick, J.L., Monath, T.P., Roizman,
S
ch 130 (2007) 140–150 149
B., Straus, S.E. (Eds.), Fields Virology, vol. 2.2, third ed. Lippincott-RavenPublishers, Philadelphia, pp. 1625–1655.
ajardo, R., Vende, P., Poncet, D., Cohen, J., 1997. Two proline residues areessential in the calcium-binding activity of rotavirus VP7 outer capsid pro-tein. J. Virol. 71 (3), 2211–2216.
orziglia, M., Larrea, C., Liprandi, F., Esparza, J., 1985. Biochemical evidencefor the oligomeric (possibly trimeric) structure of the major inner capsidpolypeptide (45 K) of rotaviruses. J. Gen. Virol. 66 (Pt. 9), 1889–1900.
arding, H.P., Calfon, M., Urano, F., Novoa, I., Ron, D., 2002. Transcriptionaland translational control in the Mammalian unfolded protein response. Annu.Rev. Cell. Dev. Biol. 18, 575–599.
ourdan, N., Maurice, M., Delautier, D., Quero, A.M., Servin, A.L., Trugnan,G., 1997. Rotavirus is released from the apical surface of cultured humanintestinal cells through nonconventional vesicular transport that bypasses theGolgi apparatus. J. Virol. 71 (11), 8268–8278.
apikian, A.Z., Hoshino, Y., Chanock, R.M., 2001. Rotaviruses. In: Knipe,D.M., Howley, P.M. (Eds.), Fields Virology, vol. 2.2, fourth ed. Lippin-cott/The Williams & Wilkins Co., Philadelphia, pp. 1787–1833.
iprandi, F., Lopez, G., Rodriguez, I., Hidalgo, M., Ludert, J.E., Mattion, N.,1990. Monoclonal antibodies to the VP6 of porcine subgroup I rotavirusesreactive with subgroup I and non-subgroup I non-subgroup II strains. J. Gen.Virol. 71 (Pt. 6), 1395–1398.
udert, J.E., Michelangeli, F., Gil, F., Liprandi, F., Esparza, J., 1987. Penetrationand uncoating of rotaviruses in cultured cells. Intervirology 27, 95–101.
aass, D.R., Atkinson, P.H., 1990. Rotavirus proteins VP7, NS28 and VP4 formoligomeric structures. J. Virol. 64, 2632–2641.
cCrae, M.A., Faulkner-Valle, G.P., 1981. Molecular biology of rotaviruses. I.Characterization of basic growth parameters and pattern of macromolecularsynthesis. J. Virol. 39 (2), 490–496.
ichelangeli, F., Liprandi, F., Chemello, M.E., Ciarlet, M., Ruiz, M.C., 1995.Selective depletion of stored calcium by thapsigargin blocks rotavirus mat-uration but not the cytopathic effect. J. Virol. 69 (6), 3838–3847.
ichelangeli, F., Ruiz, M.C., del Castillo, J.R., Ludert, J.E., Liprandi, F., 1991.Effect of rotavirus infection on intracellular calcium homeostasis in culturedcells. Virology 181, 520–527.
esavento, J.B., Crawford, S.E., Estes, M.K., Prasad, B.V., 2006. Rotavirusproteins: structure and assembly. Curr. Top. Microbiol. Immunol. 309, 189–219.
etrie, B.L., Greenberg, H.B., Graham, D.Y., Estes, M.K., 1984. Ultrastructurallocalization of rotavirus antigens using colloidal gold. Virus Res. 1, 133–152.
erez, J.F., Chemello, M.E., Liprandi, F., Ruiz, M.C., Michelangeli, F., 1998.Oncosis in MA104 cells is induced by rotavirus infection through an increasein intracellular Ca2+ concentration. Virology 252 (1), 17–27.
oruchynsky, M.S., Atkinson, P.H., 1991. Rotavirus protein rearrangementsin purified membrane-enveloped intermediate particles. J. Virol. 65,4720–4727.
oruchynsky, M.S., Maass, D.R., Atkinson, P.H., 1991. Calcium depletionblocks the maturation of rotavirus by altering the oligomerization of virus-encoded proteins in the ER. J. Cell Biol. 114, 651–656.
rasad, B.V.V., Wang, G.J., Clerx, J.P., Chiu, W., 1988. Three-dimensionalstructure of rotavirus. J. Mol. Biol. 199 (2), 269–275.
uiz, M.C., Charpilienne, A., Liprandi, F., Gajardo, R., Michelangeli, F., Cohen,J., 1996. The concentration of Ca2+ that solubilizes outer capsid pro-teins from rotavirus particles is dependent on the strain. J. Virol. 70 (8),4877–4883.
uiz, M.C., Cohen, J., Michelangeli, F., 2000. Role of Ca(2+)in the replicationand pathogenesis of rotavirus and other viral infections. Cell Calcium 28(3), 137–149.
uiz, M.C., Diaz, Y., Pena, F., Aristimuno, O.C., Chemello, M.E., Michelangeli,F., 2005. Ca2+ permeability of the plasma membrane induced by rotavirusinfection in cultured cells is inhibited by tunicamycin and brefeldin A.Virology 333 (1), 54–65.
apin, C., Colard, O., Delmas, O., Tessier, C., Breton, M., Enouf, V., Chwet-
zoff, S., Ouanich, J., Cohen, J., Wolf, C., Trugnan, G., 2002. Rafts promoteassembly and atypical targeting of a nonenveloped virus, rotavirus, in Caco-2cells. J. Virol. 76 (9), 4591–4602.hahrabadi, M.S., Babiuk, L.A., Lee, P.W.K., 1987. Further analysis of the roleof calcium in rotavirus morphogenesis. Virology 158, 103–111.

1 esear
S
S
T
T
T
50 M.C. Ruiz et al. / Virus R
hahrabadi, M.S., Lee, P.W.K., 1986. Bovine rotavirus maturation is a calcium-dependent process. Virology 152, 298–307.
hirley, J.A., Beards, G.M., Thouless, M.E., Flewett, T.H., 1981. The influenceof divalent cations on the stability of human rotavirus. Arch. Virol. 67 (1),
1–9.hastrup, O., Cullen, P.J., Drobak, B.K., Hanley, M.R., Dawson, A.P., 1990.Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by spe-cific inhibition of the endoplasmic reticulum Ca2(+)-ATPase. Proc. Natl.Acad. Sci. U.S.A. 87 (7), 2466–2470.
V
ch 130 (2007) 140–150
ian, P., Estes, M.K., Hu, Y., Ball, J.M., Zeng, C.Q., Schilling, W.P., 1995.The rotavirus nonstructural glycoprotein NSP4 mobilizes Ca2+ from theendoplasmic reticulum. J. Virol. 69 (9), 5763–5772.
ian, P., Hu, Y., Schilling, W.P., Lindsay, D.A., Eiden, J., Estes, M.K., 1994. The
nonstructural glycoprotein of rotavirus affects intracellular calcium levels.J. Virol. 68, 251–257.an Baelen, K., Dode, L., Vanoevelen, J., Callewaert, G., De Smedt, H., Mis-siaen, L., Parys, J.B., Raeymaekers, L., Wuytack, F., 2004. The Ca2+/Mn2+
pumps in the Golgi apparatus. Biochim. Biophys. Acta 1742 (1–3), 103–112.
Copyright © 2022 FDOKUMEN




















