
Defects in mitochondrial clearance predispose human monocytes to interleukin-1β hyper-secretion
Upload
johnshopkinsCategory
view
4download
0
Biochimica et Biophysica Acta 1843 (2014) 923–933
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r .com/ locate /bbamcr
Interleukin-1β and cyclic AMP mediate the invasion of shearedchondrosarcoma cells via a matrixmetalloproteinase-1-dependent mechanism
Pu Wang a,b,1, Pei-Pei Guan a,b,1, Tao Wang a, Xin Yu a, Jian-Jun Guo a,Konstantinos Konstantopoulos b,c,d,e, Zhan-You Wang a,⁎a College of Life and Health Sciences, Northeastern University, Shenyang 110819, PR Chinab Department of Chemical and Biomolecular Engineering, The Johns Hopkins University, Baltimore, MD, USAc Johns Hopkins Institute for NanoBio Technology, The Johns Hopkins University, Baltimore, MD, USAd Johns Hopkins Physical Sciences—Oncology Center, The Johns Hopkins University, Baltimore, MD, USAe Center of Cancer Nanotechonology Excellence, The Johns Hopkins University, Baltimore, MD 21218, USA
Abbreviations: ChIP, chromatin immunoprecipitation; ckin-1β; MMP-1, matrix metalloproteinase-1; OA, osteoar3-kinase⁎ Corresponding author at: College of Life and Health Sc
Shenyang 110004, PR China. Tel./fax: +86 24 22529997.E-mail address: [email protected] (Z.-Y. Wang
1 These authors contributed equally to this work.
0167-4889/$ – see front matter © 2014 Elsevier B.V. All rhttp://dx.doi.org/10.1016/j.bbamcr.2014.01.018
a b s t r a c t
a r t i c l e i n f oArticle history:Received 13 October 2013Received in revised form 15 December 2013Accepted 16 January 2014Available online 25 January 2014
Keywords:Fluid shear stressCyclic AMPInterleukin-1βc-JunMatrix metalloproteinase-1Invasion
Matrix metalloproteinase-1 (MMP-1) is a potential biomarker for chondrosarcoma that is overexpressed at theinvading edges of articular cartilage, and its expression correlates with poor survival rates. However, the molec-ularmechanisms ofMMP-1 regulation and its potential contribution to chondrosarcoma cell invasion have yet tobe elucidated, especially in shear-activated cells. Usingmolecular biology tools and an in vitro fluid shear model,we report that shear stress upregulates cyclic AMP (cAMP) and interleukin-1β (IL-1β) release,which in turn pro-motes the invasion of chondrosarcoma cells via the induction of MMP-1 in a phosphoinositide 3-kinase (PI3-K)-and ERK1/2-dependent manner. Activated PI3-K and ERK1/2 signaling pathways phosphorylate c-Jun, which inturn transactivates MMP-1 in human chondrosarcoma cells. Collectively, fluid shear stress upregulates matrixMMP-1 expression, which is responsible for the enhanced invasion of human chondrosarcoma cells.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Interstitial fluid flow in and around tumor tissue modulates themechanical microenvironment, including the shear stress and pressureforce acting on the cell surface and the tethering force on cell–substrateconnections [1]. Using a mathematical model of interstitial pressure-fluid flow, Jain and colleagues reported that the interstitial fluid velocityin tumors is nearly zero in the center and increases rapidly in the periph-ery (up to 4 μm/s [2]), which corresponds to amaximal shear stress levelof ~10 dyn/cm2 [3]. To date, the effect of fluid shear on tumor cell inva-sion and signaling has not been extensively investigated. Prior work hasshown that application of uniform laminar shear stress (12 dyn/cm2) in-duced G2/M arrest of metastatic tumor cells, including oral squamouscarcinoma, osteosarcoma and chondrosarcoma cells [3]. Such G2/Marrest would facilitate the eradication of tumor cells by the immune
AMP, cyclic AMP; IL-1β, interleu-thritis; PI3-K, phosphoinositide
iences, Northeastern University,
).
ights reserved.
system. However, little is known about the effect of lower levels offluid shear stress on chondrosarcoma cell function. A critical step ofchondrosarcoma cell invasion into normal tissue is the degradation ofthe extracellular matrix (ECM) within and around the tumor. The extra-cellularmatrix of chondrosarcoma cells is composed primarily of fibrillarcollagens, including types I, II, and IV. The degradation of collagens isme-diated by the activity of matrix metalloproteinases (MMPs) which con-stitute a multi-gene family with over 22 members. Specifically, fibrillarcollagens are degraded by interstitial collagenase (MMP-1), neutrophilcollagenase (MMP-8), and collagenase-3 (MMP-13), which maycause a loss of mechanical constraint in invasive cells [4]. Importantly,MMP-1, but not MMP-8 or MMP-13, has been found to be associatedwith an increased rate of chondrosarcoma recurrence [5,6]. Recentwork suggests that the siRNA-induced knockdown of MMP-1 reducesthe invasive potential of human chondrosarcoma cells [7]. However,the role of fluid shear stress in the regulation of MMP-1 in humanchondrosarcoma cells has been largely overlooked. Moreover, it is notclear if fluid shear stress regulates chondrosarcoma cell invasion via aMMP-1-dependent mechanism.
In this study,we delineate the signaling pathway involved inMMP-1induction in shear-activated human chondrosarcoma cells. Specifically,we demonstrate that fluid shear stress augments the release ofcAMP and IL-1β, thereby activating PI3-K/AKT- and ERK1/2 mitogen-

924 P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
activated protein kinase (MAPK)-dependent pathways. PI3-K/AKT andERK1/2 phosphorylate c-Jun,which in turn binds to themmp-1 promoterto mediate MMP-1 mRNA and protein synthesis in shear-stimulatedchondrosarcoma cells. Our findings suggest that fluid shear stress facili-tates the early, local invasion of chondrosarcoma cells via the activationof MMP-1, hence gaining insight into potential therapeutic strategies tocombat the invasion of chondrosarcomas.
2. Materials and methods
2.1. Reagents
IL-1β, forskolin and the inhibitors LY294002, U0126, and SP600125were obtained from Sigma-Aldrich Corp. (St. Louis, MO, USA). Anti-bodies against β-actin, AKT, p-AKT (Ser 473), ERK1/2, p-ERK1/2(Thr202/Tyr204), c-Jun, p-c-Jun (Ser 63), and IL-1β and siRNAstargeting AKT and ERK1/2 were purchased from Cell Signaling Technolo-gy, Inc. (Danvers, MA, USA). Quinazoline (QNZ) was obtained from EnzoLife Sciences (Farmingdale, NY, USA). The IL-1β and cAMPenzyme immu-noassay kits were from Cayman Chemical (Ann Arbor, MI, USA). Recom-binant human MMP-1 was obtained from R&D Systems (Minneapolis,MN, USA). All reagents for the qRT-PCR and SDS-PAGE experimentswere purchased from Bio-Rad Laboratories. All other reagents werefrom Invitrogen (Carlsbad, CA, USA) unless otherwise specified.
2.2. Cell culture and shear stress exposure
SW1353 and HS.819.T cells were grown (37 °C and 5% CO2) on glassslides in DMEM/F12 medium supplemented with 10% FBS as previouslydescribed [8–12]. Before exposure to shear stress, the cells were incubat-ed for 18 h in serum-free medium supplemented with 1% Nutridoma-SP(Roche) [13,14]. Using a streamer goldflowdevice (Flexcell International,Hillsborough, NC, USA), the cells were then subjected to various levels ofshear stress for the prescribed periods of time in medium containing 1%Nutridoma-SP. In select experiments, the indicated concentrations ofthe pharmacological agents were added to the medium immediatelyprior to shearing. In a separate set of experiments, SW1353 cells wereseeded on 6-cm tissue culture dishes (106 cells per dish) in DMEM/F12medium supplemented with 10% FBS [11]. Following incubation for24 h, SW1353 cells were grown in serum-free medium for an additional24 h before incubation with inhibitors in the presence or absence offorskolin or IL-1β.
2.3. Quantitative real-time PCR (qRT-PCR)
qRT-PCR assays were performed with the iCycler iQ detection sys-tem (Bio-Rad) using total RNA and the iScript one-step RT-PCR kitwith SYBR green (Bio-Rad) and the appropriate primers. Using theMMP-1 sequence (GenBank accession number NM_002421), the fol-lowing forward (F) and reverse (R) primers were used: 5′-AAGGCCAGTATGCACAGCTT-3′ and 5′-TGCTTGACCCTCAGAGACCT-3′, respectively.The GenBank accession number and forward and reverse primers forGAPDH, which was used as an internal control, are provided in ourprior publications [11,12]. The reaction mixtures were incubated at50 °C for 15 min, followed by 95 °C for 5 min, and then 35 PCR cycleswere performed with the following temperature profiles: 95 °C for15 s, 58 °C for 30 s, 68 °C for 1 min, and 77 °C for 1 min. Data were col-lected at the final step (77 °C for 1 min) to prevent the inclusion of anyfluorescence from primer dimmers [11,12]. The gene expression valueswere normalized to those of GAPDH.
2.4. Western blot analysis
Static and sheared SW1353 cells were lysed in radioimmune precip-itation assay buffer (25 mM Tris–HCl [pH 7.6], 150mMNaCl, 1% NP-40,1% sodium deoxycholate, and 0.1% SDS) containing protease inhibitor
cocktail (Pierce Chemical Company). The protein content of the cell ly-sates was determined using a bicinchoninic acid (BCA) protein assay re-agent (Pierce Chemical Company). The total cell lysates (4 μg) weresubjected to SDS-PAGE, transferred to a membrane, and probed with apanel of specific antibodies. Each membrane was only probed withone antibody. β-actin was used as a loading control. All western hybrid-izations were performed at least in triplicate using a different cell prep-aration each time.
2.5. Measurement of the cAMP and IL-1β concentration in theculture medium
The cAMP and IL-1β levels in the media of both static and shearedcells were determined using the cAMP and IL-1β enzyme immunoassaykits following the manufacturer's instructions, respectively. The totalprotein in the medium was used as a loading control, and the resultsare expressed as pmol cAMP or pg IL-1β per μg of total protein.
2.6. Transfection
SW1353 chondrosarcoma cells were transfected with 100 nM of anAKT- or ERK1/2-specific siRNA oligonucleotide. In control experiments,the cells were transfected with 100 nM of scrambled siRNA. In promoterassays, SW1353 chondrosarcoma cells were transfected with 1.6 μg/slideof themmp-1 promoter reporter construct together with the pRL-SV40vector. The transfected cells were allowed to recover for at least 12 h ingrowth medium and then incubated overnight in medium containing1% Nutridoma-SP before exposure to shear stress.
2.7. Luciferase promoter constructs
A 4435-base pair (bp) mmp-1 promoter construct, which containsthe 5′-flanking region of the human MMP-1 gene from −4372 to+63 relative to the transcription start site, was generated fromhuman genomic DNA. The construct was created using specifically de-signed forward (5′-CGACGCGTCCTCACATATTTCAAATCCATC-3′) and re-verse (5′-CCGCTCGAGGGCCTTTGTCTTCTTTCTCAGT-3′) primers, whichincluded Mlu1 and Xho1 restriction sites (underlined) at their 5′ endsand 3′ ends, respectively [15]. The amplicon was inserted upstream ofthe luciferase reporter gene in the pGL3-basic vector (Promega). Vari-ous truncations were generated using primers that incorporated therelevant restriction enzyme sites (underlined) to allow for proper orien-tation upstream of the luciferase reporter. The following truncated con-structs were generated using the indicated primers: the −3830 bpconstruct, forward 5′-CGACGCGTTTGCTTAAGCCCAAGAGTTTGA-3′; the−2942 bp construct, forward 5′-CGACGCGTATTCCCCAGCATAAAAAAATAT-3′; the −2872 bp construct, forward 5′-CGACGCGTATATGTTTGAGGCCTTTCACAG-3′; the −2001 bp construct, forward 5′-CGACGCGTTTAGGCAATTTCCTGTCCAATC-3′; the −1551 bp construct, forward5′-CGACGCGTGATATCAGTCTTGACGCAGAAA-3′; the −1194 bp con-struct, forward 5′-CGACGCGTTCTTTTGAATGGAGGAATGCAT-3′; andthe −517 bp construct, forward 5′-CGACGCGTCAGGCAGCTTAACAAAGGCAGA-3′. All constructs were sequenced and confirmed to be identi-cal to the sequence of the mmp-1 promoter [15]. For the NF-κB con-struct, site-directed mutagenesis was used to alter the sequencebetween −2886 and −2878 bp from 5′-ATGGAAAAA-3′ to 5′-ATGGAACCC-3′ in order to block the binding between NF-κB andmmp-1 pro-moter. Similarly, the AP-1 constructwas also altered between at−1949and −1955 bp from 5′-TGAGTTA-3′ to 5′-TGATTTA-3′ [16]. All of theconstructs were confirmed by DNA sequencing.
2.8. Promoter assay
Firefly and Renilla luciferase activities were measured using theDual-Luciferase Report Assay Kit (Promega) [9,12,17–19]. The fireflyluciferase activity was normalized to that of the Renilla luciferase

Fig. 1. MMP-1 expression in human chondrosarcoma tissues and shear-activatedchondrosarcoma cells. (A) MMP-1 expression in human chondrosarcoma tissue speci-mens was determined by immunohistochemical staining using an anti-MMP-1 antibody.Strong immunostaining was detected in ~90% of cancerous tissues, whereas little to noimmunostaining was identified in normal tissues. (B) SW1353 and (C) HS.819.T cellswere subjected to indicated levels of fluid shear stress (0.1–2.0 dyn/cm2) or static condi-tions (0 dyn/cm2) for 48 h.MMP-1mRNA, protein and activity levels were determined byqRT-PCR, western blots and zymography, respectively. The total amounts of GAPDH andMMP-11 served as internal controls in the qRT-PCR and zymography assays, respectively.The data represent the mean ± S.E. of three independent experiments. *, p b 0.05 withrespect to the static control.
925P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
control. The data are expressed as the ratio of the normalized firefly lu-ciferase activity after exposure to shearing to that observed under staticconditions unless otherwise stated.
2.9. Chromatin immunoprecipitation (ChIP)
This assay was performed using the EZ ChIP kit (Upstate Biotechnol-ogy) following the manufacturer's instructions as previously described[11,17,19]. The forward (F) and reverse (R) primers used for the ampli-fication of themmp-1 promoter by qPCRwere 5′-GCAGGCTCTTGCCAGATGGG-3′ and 5′-ACCAAAGAACACTTGCCACT-3′, respectively.
2.10. Casein zymography
The activity of MMP-1 was determined using a previously describedmethod [18]. In brief, the conditioned media from SW1353 cellswas collected andmixed with a non-reducing sample buffer containing0.5M Tris (pH 6.8), 5% SDS, 20% glycerol, and 1% bromophenol blue in a1:1 ratio. The sample was electrophoresed in a 10% SDS-polyacrylamidegel. After electrophoresis, the gel was washed for 1 h at room tempera-ture in a 2.5% (v/v) Triton X-100 solution to remove the SDS, transferredto a zymogram development solution (10 mM CaCl2, 50 mM Tris–HCl[pH 7.4], and 0.02% NaN3), and incubated for 72 h at 37 °C. The gelwas stained for 30 min with 0.1% (w/v) Coomassie brilliant blue (CBB)in a 50% (v/v)methanol and10% (v/v) acetic acid solution anddestainedin a 20% (v/v) methanol and 10% (v/v) acetic acid solution. The areas oflysis were observed as white bands against a black background.
2.11. Transwell migration and invasion assay
For these assays, 5 × 104 SW1353 cells treated with 10 μM forskolin,100 ng/ml IL-1β, or vehicle control were seeded on 24-well Transwellinserts with an 8-μM pore size (Becton Dickinson, Bedford, MA, USA)in the absence or presence of LY294002 (10 μM), U0126 (10 μM), orSP600125 (10 μM). In select experiments, the SW1353 cells were treatedwith 1 mg/ml rhMMP-1 or vehicle control after seeding. The lowerchamber contained complete medium with 10% FBS. After culture for48 h, the cells on the upper side of the chamber were removed with acotton swab. Next, the membrane was fixed with 3.7% formaldehydeand stained with 0.1% crystal violet in PBS (−). By microscopy underlowmagnification (×100), five fields of vision (up, down, left, right, cen-ter) were analyzed for each membrane. The number of cells that hadpassed through the polycarbonate membrane was determined for eachfield of view to indicate the migratory ability of the tumor cells. In theinvasion experiments, the polycarbonate membrane was pre-coatedwith Matrigel (Becton Dickinson, Bedford, MA, USA) before seeding theSW1353 cells in the upper chamber. Following the same protocol as de-scribed above, the number of cells that had passed through the insertdemonstrated the invasive ability of the tumor cells.
2.12. Immunohistochemistry
Human bone tissue microarrays (US Biomax Inc., Rockville, MD,USA), which were composed of 69 chondrosarcoma and 24 normalbone tissue cores, were fixed on tissue microarray slides. The slideswere first deparaffinized with xylene, rehydrated in a graded series ofethanol, and submerged in 3% hydrogen peroxide to eliminate anyendogenous peroxidase activity. The level of MMP-1 was determinedwith an immunohistochemical staining kit (Invitrogen, Carlsbad, CA,USA) following the manufacturer's instructions.
2.13. Statistical analysis
All data represent the mean ± S.E. of at least three independentexperiments. The statistical significance of the differences between themeans was determined using Student's t-test or one-way ANOVA
where appropriate. If themeanswere found to be significantly different,multiple pairwise comparisons were performed using the Tukey test[14].

926 P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
3. Results
3.1. MMP-1 overexpression in chondrosarcoma tissues and fluid shearstress-stimulated SW1353 chondrosarcoma cells
In view of previous observations suggesting thatMMP-1 contributesto chondrosarcoma cell invasion [5–7], we first evaluated the relativeexpression levels of MMP-1 in chondrosarcoma tissues compared tonormal bone controls. In accordance with prior published data [5,6],MMP-1 was highly immunostained in human chondrosarcomasrelative to normal bone controls (Fig. 1A). Because fluid shear modu-lates tumor metastasis through the regulation of cell adhesion [20],migration, and invasion [21], we next examined the effects of fluidshear on chondrosarcoma cell function. A key step of cell invasion intonormal tissue is the MMP-dependent degradation of the ECM. In lightof MMP-1 overexpression in chondrosarcoma tissues (Fig. 1A), weinvestigated the effect of fluid shear on MMP-1 induction. The humanSW1353 chondrosarcoma cell linewas chosen as amodel systembecauseit is the most commonly used cell line for bone tumor research [22]. Wealso verified our observations using the HS.819.T chondrosarcoma cellline as a second model system. Our data reveal that application of fluidshear stress (2 dyn/cm2 for 48 h) markedly induced the mRNA and pro-tein expression and activity levels of MMP-1 in both SW1353 andHS.819.T cells (Fig. 1B, C). Smaller increases were observed at lowershear stress levels (0.2–1.6 dyn/cm2) (Fig. 1B, C). These data suggestthat fluid shear stress contributes to chondrosarcoma invasion via theinduction of MMP-1.
Fig. 2. Fluid shear stress augments the production of cAMP and IL-1β, which in turn upregulateAKT and ERK1/2 in SW1353 chondrosarcoma cells. SW1353 cells were subjected to fluid shearSW1353 cells were subjected to fluid shear stress (2 dyn/cm2) or static conditions (0 dyn/cm2)anti-IL-1βmAb (1 μg/ml) (C, D). The formation of cAMP (A) and the secretion of IL-1β (B) wermRNA and protein levels were determined by qRT-PCR andwestern blots, respectively. MMP-1as internal controls in the qRT-PCR and zymography assays, respectively (C). Phosphorylated Ademonstrated by the similar intensities of total AKT, ERK1/2, and β-actin. These western blots arepresent the mean ± S.E. of three independent experiments. *, p b 0.05 with respect to the st
3.2. Fluid shear stress induces the synthesis of cAMP and IL-1β, which inturn regulate MMP-1 expression and activity via the PI3-K/AKT andERK1/2 pathways in human SW1353 chondrosarcoma cells
We next sought to elucidate the signaling cascade that regulatesMMP-1 synthesis in shear-activated SW1353 chondrosarcoma cells.Given that exogenously added cAMP and IL-1β induce MMP-1 mRNAsynthesis in human monocytes and chondrosarcoma cells [23,24], weexamined the potential contributions of cAMP and IL-1β to the activationofMMP-1 in sheared SW1353 cells. As a first step, we demonstrated thatthe endogenous levels of cAMP and IL-1βwere elevated in SW1353 cellssubjected to fluid shear (2 dyn/cm2) for 48 h (Fig. 2A, B). This shearexposure markedly upregulated the mRNA and protein expression andenzymatic activity of MMP-1 in SW1353 cells (Fig. 2C). Interestingly, in-cubation of SW1353 cells with the adenylate cyclase inhibitor SQ22536(10 μM) or an anti-IL-1β antibody (1 μg/ml) abrogated the induction ofMMP-1 mRNA and protein synthesis and activity in response to shearstimulation (Fig. 2C). These pharmacological interventions were alsoeffective in suppressing the shear-induced phosphorylation of AKTand ERK1/2 (Fig. 2D). To verify the involvement of the PI3-K/AKTand ERK1/2 signaling pathways in shear-induced MMP-1 synthesis,we treated SW1353 cells with the selective PI3-K inhibitor LY294002(10 μM) or the ERK1/2 inhibitor U0126 (10 μM). These pharmacologicalinhibitors diminished the phosphorylation of AKT and ERK1/2 withoutaffecting their total expression levels (Fig. 3A) and markedly attenuatedshear-induced MMP-1 mRNA and protein expression and activity(Fig. 3B). To confirm the involvement of these signaling pathways in
the mRNA expression and enzymatic activity of MMP-1 as well as the phosphorylation ofstress (2 dyn/cm2) or static conditions (0 dyn/cm2) for 48 h (A, B). In select experiments,for 48 h in the absence or presence of the adenylyl cyclase inhibitor SQ22536 (10 μM)or ane determined using cAMP and IL-1β enzyme immunoassay kits, respectively. The MMP-1activity was quantified by zymography. The total amounts of GAPDH andMMP-11 servedKT and ERK1/2 were detected by immunoblotting using specific Abs. Equal lane loading isre representative of three independent experiments, all revealing similar results. The dataatic control, #, p b 0.05 compared to shear alone.

927P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
MMP-1 regulation, SW1353 cells were transfectedwith siRNAs targetingAKT or ERK1/2, which effectively knocked down the protein expressionof AKT or ERK1/2, respectively, relative to cells transfected with thescrambled siRNA control (Fig. 3C). Depletion of AKT or ERK1/2 alsoinhibited MMP-1 mRNA and protein expression and enzymatic activity(Fig. 3D).
To further verify the involvement of cAMP and IL-1β in the regula-tion of MMP-1 induction in shear-stimulated SW1353 cells, static ex-periments were conducted by incubating the cells with forskolin(10 μM), which activates cAMP formation, or exogenous IL-1β(100 ng/ml). Treatment of SW1353 cells with forskolin (10 μM) orIL-1β (100 ng/ml) markedly increased the phosphorylation of AKTand ERK1/2 (Fig. 4A, B), which in turn resulted in the enhanced mRNAand protein expression of MMP-1 (Fig. 4C, D). In addition, treatmentwith LY294002 (10 μM) or U0126 (10 μM) blocked forskolin- orIL-1β-induced mRNA and protein expression of MMP-1 (Fig. 4C, D).The observed reduction in MMP-1 can be attributed to the abilityof LY294002 and U0126 to inhibit the phosphorylation of AKT andERK1/2, respectively, in human chondrosarcoma cells (Fig. 4A, B).Taken together, our data reveal that fluid shear stress upregulates theproduction of both cAMP and IL-1β, which in turn induce MMP-1 ex-pression and enzymatic activity via a PI3-K/AKT and ERK1/2 signalingpathways.
Fig. 3. Fluid shear stress induces MMP-1 synthesis in human chondrosarcoma cells via a PI3-K(2 dyn/cm2) or static conditions (0 dyn/cm2) for 48 h (A–D). In select experiments, SW1353U0126 (10 μM) in the presence of fluid shear stress (A, B). In another set of experiments, SWERK1/2 before being exposed to fluid shear stress for 48 h (C, D). (A, C) Phosphorylated AKT anonstrated by the similar intensities of total AKT, ERK1/2, andβ-actin. These gels are representativprotein and activity levels were determined by qRT-PCR, western blots and zymography, respeassays, respectively. The data represent the mean ± S.E. of three independent experiments. *, ptreatment.
3.3. c-Jun, but not NF-κB, is involved in shear-induced MMP-1mRNA synthesis
Wenext aimed to identify the key transcription factor(s) responsiblefor MMP-1 mRNA induction in shear-activated chondrosarcoma cells.The promoter of themmp-1 gene contains several consensus sequences,including those for AP-1 and NF-κB [15,16]. Given the increased phos-phorylation of AKT and ERK1/2 in shear-activated or forskolin- orIL-1β-treated chondrosarcoma cells, we examined the potential role ofAKT and ERK1/2 in modulating the activities of c-Jun and NF-κB. Asshown in Fig. 5, the treatment of SW1353 cells with LY294002 orU0126 diminished the phosphorylation of c-Jun in shear-activated orforskolin- or IL-1β-treated cells (Fig. 5A–C). These data suggest thatc-Jun is involved in transmitting fluid shear stress signals to regulatethe synthesis of MMP-1 in human chondrosarcoma cells. To validatethis hypothesis, SW1353 cells were treated with the JNK inhibitorSP600125 (10 μM) and stimulated with fluid shear stress, forskolin, orIL-1β. Our data reveal that SP600125 markedly suppressed the phos-phorylation of c-Jun in shear stress-, forskolin-, or IL-1β-stimulatedchondrosarcoma cells (Fig. 5D–F). Similarly, MMP-1 mRNA and proteinexpression and enzymatic activity were also inhibited upon the treat-ment of shear stress-, forskolin-, or IL-1β-activated chondrosarcomacells with SP600125 (Fig. 5G–I). In contrast, the treatment of SW1353
/AKT- and ERK1/2-dependent pathway. SW1353 cells were subjected to fluid shear stresscells were incubated with the PI3-K inhibitor LY294002 (10 μM) or the ERK1/2 inhibitor1353 cells were transfected with a siRNA oligonucleotide sequence specific for AKT or
d ERK1/2 were detected by immunoblotting using specific Abs. Equal lane loading is dem-e of three independent experiments, all revealing similar results. (B, D) TheMMP-1mRNA,ctively. GAPDH and MMP-11 served as internal controls in the qRT-PCR and zymographyb 0.05 with respect to the static control. §, p b 0.05 compared with the fluid shear stress

Fig. 4. cAMPand IL-1β upregulateMMP-1 expression and activity in SW1353 chondrosarcoma cells. SW1353 cells were treatedwith 10 μM forskolin (A, C) or 100 ng/ml IL-1β (B, D) in theabsence or presence of the PI3-K inhibitor LY294002 (10 μM) or the ERK1/2 inhibitor U0126 (10 μM). The levels of phospho-AKT and phospho-ERK1/2were determined in the cell lysatesbywestern blot analysis using specific Abs. Equal lane loading is demonstrated by the similar intensities of total AKT, ERK1/2, andβ-actin (A, B). The gels are representative of at least threeindependent experiments, all revealing similar results. The protein expression and proteolytic activity ofMMP-1 in the SW1353 culture media was evaluated bywestern blots and caseinzymography (C, D, upper panel), inwhichMMP-11 served as a loading control. TheMMP-1mRNA levelswere determined by qRT-PCR, inwhichGAPDH served as an internal control (C, D,lower panel). The data represent themean±S.E. of three independent experiments. *, p b 0.05with respect to the static control. #, p b 0.05 comparedwith the forskolin or IL-1β treatment.
928 P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
cells with the NF-κB inhibitor quinazoline (QNZ) (2 μM) failed to signif-icantly alter the expression or enzymatic activity of MMP-1 upon stim-ulation with shear stress, forskolin, or IL-1β (Fig. S1).
To verify the involvement of c-Jun in the regulation of shear-inducedMMP-1 mRNA expression, a series of mmp-1 promoter constructs wasgenerated using the luciferase reporter plasmid pGL3-basic (Fig. 6A,left panel). First, SW1353 cells were transiently transfected with a con-struct encompassing the 5′-flanking region of the human MMP-1 genefrom −4372 to +63 bp prior to exposure to fluid shear stress. The ap-plication of fluid shear stress (2 dyn/cm2) for 48 h markedly increased(~10-fold)mmp-1 promoter activity in SW1353 cells (Fig. 6A). A similar10-fold upregulation was observed upon the transfection of SW1353cells with plasmids containing deletions from −4372 to −3830 bp(−3830/+63), −3830 to −2942 bp (−2942/+63), −2942 to−2872 bp (−2872/+63), or −2872 to −2001 bp (−2001/+63). Al-together, these results suggest that the DNA region between −4372and −2001 bp upstream of the transcriptional start site is not criticalto the induction of shear-induced mmp-1 promoter activity (Fig. 6A).However, a further deletion from −2001 to −1551 bp (−1551/+63)significantly suppressed the luciferase activity of the reporter(Fig. 6A). Bioinformatics analysis of the consensus sequence in the−2001/−1551 region revealed the presence of an AP-1 site situatedat −1949/−1955 bp, which may be responsible for shear-inducedMMP-1 synthesis. Indeed, introduction of a point mutation into theaforementioned AP-1 site, but not the NF-κB binding site, resulted in amarked decrease in shear-activated luciferase activity relative to thereported wild-type value (Fig. 6B).
ChIP assays were performed to demonstrate the binding ofphosphorylated c-Jun to its putative site on the mmp-1 promoter. Asshown in Fig. 6C, the binding of c-Jun to themmp-1 promoterwasmark-edly increased upon the exposure of SW1353 cells to fluid shear stressfor 48 h. It is noteworthy that SW1353 cells treated with exogenousSQ22536 (10 μM) or an anti-IL-1β antibody (1 μg/ml) displayed an ap-parent reduction in the amount of c-Jun bound to themmp-1 promoterafter exposure to shear stress for 48 h (Fig. 6C). Similar observationsweremade upon the treatment of SW1353 cells with the PI3-K inhibitorLY294002 (10 μM), the ERK1/2 inhibitor U0126 (10 μM), or the JNKinhibitor SP600125 (10 μM) (Fig. 6C). Similar data were also obtainedusing forskolin or IL-1β-stimulated human chondrosarcoma cells(Fig. 6D, E). Collectively, these data support the notion that c-Jun, butnot NF-κB, functions as a pivotal transcriptional factor that mediatesshear-regulated MMP-1 expression.
3.4. Fluid shear stress activates MMP-1, which in turn induces humanchondrosarcoma cell migration and invasion in vitro
We next wished to decipher the molecular mechanism by whichfluid shear stress promotes chondrosarcoma cell migration and inva-sion. Previous studies have reported that MMP-1 activation is involvedin chondrosarcoma cell invasion andmetastasis [4–7]. Fluid shear stressstimulation increased the production of cAMP and IL-1β, suggestingthat cAMP and/or IL-1β transmit the shear stress signals to regulatehuman chondrosarcoma cell invasion. In line with this hypothesis,both forskolin (10 μM) and IL-1β (100 ng/ml) enhanced the migration

Fig. 5. The key role of c-Jun in fluid shear stress-, forskolin or IL-1β-inducedMMP-1 synthesis. SW1353 cells were subjected tofluid shear stress (2 dyn/cm2) or static conditions (0 dyn/cm2)for 48 h (A, D, G) in the absence or presence of the PI3-K inhibitor LY294002 (10 μM), the ERK1/2 inhibitor U0126 (10 μM) (A), or the JNK inhibitor SP600125 (10 μM) (D, G). In select ex-periments, SW1353 cells were treatedwith 10 μM forskolin (B, E, H) in the absence or presence of the PI3-K inhibitor LY294002 (10 μM), the ERK1/2 inhibitor U0126 (10 μM) (B), or the JNKinhibitor SP600125 (10 μM) (L, H). In another set of experiments, SW1353 cellswere treatedwith 100ng/ml IL-1β (C, F, I) in the absence or presence of the PI3-K inhibitor LY294002 (10 μM),theERK1/2 inhibitor U0126 (10 μM) (C), or the JNK inhibitor SP600125 (10 μM) (F, I). The levels of phosphorylated and total c-Jun in the cell lysateswere determined bywestern blot analysisusing specific Abs. Equal lane loading is demonstrated by the similar intensities of total β-actin (A–F). The protein expression and proteolytic activity of MMP-1 in the SW1353 culturemediawas evaluated by western blots and casein zymography (G–I, upper panel), in whichMMP-11 served as a loading control. The MMP-1mRNA levels were determined by qRT-PCR, in whichGAPDH served as an internal control (G–I, lower panel). The gels are representative of at least three independent experiments, all revealing similar results. The data represent themean±S.E.of three independent experiments. *, p b 0.05 with respect to the static control. #, p b 0.05 compared with fluid shear stress, forskolin, or IL-1β treatment alone.
929P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
and invasion of SW1353 cells (Fig. 7A–D). Interestingly, treatment ofSW1353 cells with the PI3-K inhibitor LY294002 (10 μM), the ERK1/2inhibitor U0126 (10 μM)or the JNK inhibitor SP600125 (10 μM)blockedthe ability of exogenously added cAMP and IL-1β to enhance SW1353cell migration and invasion (Fig. 7A–D). Moreover, incubation ofSW1353 cells with rhMMP-1 significantly increased their migrationand invasion, as shown in Transwell assays (Fig. 7E, F). In addition, treat-ment of SW1353 cells with anti-MMP-1 (1 μg/ml) antibody suppressedcell migration and invasion (Fig. 7G, H). Collectively, our data demon-strate that fluid shear stress induces the production of both endogenouscAMP and IL-1β, which in turn increase MMP-1 mRNA expression andenzymatic activity via a PI3-K-, ERK1/2-, and c-Jun-dependent path-ways, thereby promoting the invasive potential of chondrosarcomacells (Fig. 8).
4. Discussion
MMP-1 upregulation has been reported in chondrosarcoma cells[5–7]. Yet, the underlying mechanisms of MMP-1 regulation have yetto be fully elucidated. Given the elevated expression levels of MMP-1in chondrosarcoma tissues relative to normal controls [5–7] and therole of fluid shear stress in regulating tumor metastasis [1,25], wesought to elucidate the signaling pathway governingMMP-1 regulationin fluid shear stress-stimulated human chondrosarcoma cells. Specifi-cally, we demonstrate that fluid shear stress augments the release ofcAMP and IL-1β, which in turn regulate MMP-1 expression via PI3-K-,
ERK1/2-, and JNK-dependent c-Jun activating pathways in SW1353cells (Fig. 8).
MMP-1 is tightly regulated and maintained at basal levels underphysiological conditions. Previous studies detected MMP-1 immuno-staining in more than 90% of human chondrosarcomas, and showedthat the positive score of MMP-1 staining is significantly higher inhuman chondrosarcomas than in benign chondroid lesions [26]. In addi-tion, the ratio ofMMP-1 to TIMP-1 gene expression serves as a biomarkerof chondrosarcomas and is indicative of the rate of chondrosarcoma re-currence [6]. In accordancewith these prior observations, our data revealthat MMP-1 immunostainingwas increased in human chondrosarcomascompared to normal bone controls (Fig. 1A). We also found thatfluid shear stress upregulates MMP-1 expression and activity in humanchondrosarcoma cells. In linewith this finding, other studies have report-ed that fluid shear stress induces MMP-1 synthesis in other cell types,including human periodontal ligament cells [27], rat vascular musclesmooth cells [28], and human chondrocytes [29]. Collectively, our datasuggest that fluid shear stress potentially contributes to chondrosarcomacell migration and invasion via the induction of MMP-1 expression.
Because fluid shear is a physiologically relevant signal that regulatesintracellular signaling pathways, we delineated the molecular mecha-nisms regulating MMP-1 synthesis in sheared SW1353 cells. We foundthat the endogenous production of cAMP and IL-1β by sheared chon-drosarcoma cells induced MMP-1 expression. This finding is in accordwith previous observations showing that cAMP agonists and IL-1β arecapable of inducing MMP-1 mRNA expression in human monocytes

Fig. 6. c-Jun is the critical transcription factor for MMP-1 synthesis in human chondrosarcoma cells. SW1353 cells were transfected withmmp-1 promoter expression plasmids (A) or de-letion constructs (B) before exposure to fluid shear stress (2 dyn/cm2) for 48 h. The resultant luciferase activities,whichwere normalized to the Renilla luciferase activities, weremeasuredusing the Dual-Luciferase Reporter Assay Kit. The data represent the mean ± S.E. of three independent experiments. *, p b 0.05 with respect to pMMP-1 (−4372/+63). In select exper-iments, SW1353 cells were subjected to fluid shear stress (2 dyn/cm2) or static conditions (0 dyn/cm2) for 48 h in the absence or presence of the adenylyl cyclase inhibitor SQ22536(10 μM), an anti-IL-1β mAb (1 μg/ml), the PI3-K inhibitor LY294002 (10 μM), the ERK1/2 inhibitor U0126 (10 μM), or the JNK inhibitor SP600125 (10 μM) (C). In distinct experiments,SW1353 cells were treated with 10 μM forskolin in the absence or presence of the PI3-K inhibitor LY294002 (10 μM), the ERK1/2 inhibitor U0126 (10 μM), or the JNK inhibitorSP600125 (10 μM) (D). In another set of experiments, SW1353 cells were treated with 100 ng/ml IL-1β in the absence or presence of the PI3-K inhibitor LY294002 (10 μM), the ERK1/2 inhibitor U0126 (10 μM), or the JNK inhibitor SP600125 (10 μM) (E). Cross-linked chromatin was immunoprecipitated using an anti-c-Jun antibody. In the ChIP assays, the anti-RNApolymerase II antibody was used as a positive control. DNA purified from immunoprecipitated (IP) and preimmune (Input) specimens was subjected to qPCR amplification using primersfor themmp-1 promoter. All experiments are representative of three independent experiments, all revealing similar results. The data represent the mean ± S.E. of three independentexperiments. *, p b 0.05 with respect to the static or vehicle treatment control. #, p b 0.05 compared with fluid shear stress, forskolin, or IL-1β treatment alone.
930 P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
and chondrosarcoma cells [23,24]. In contrast, Zayed et al. [30] reportedthat treatment of human osteoarthritic chondrocytes with cAMP-elevating agents, such as 8-Br-cAMP and forskolin, suppresses IL-1β-
induced MMP-1 expression. This discrepancy may reflect the divergentresponses of distinct cell types (e.g., chondrosarcoma versus osteoar-thritic chondrocytes).
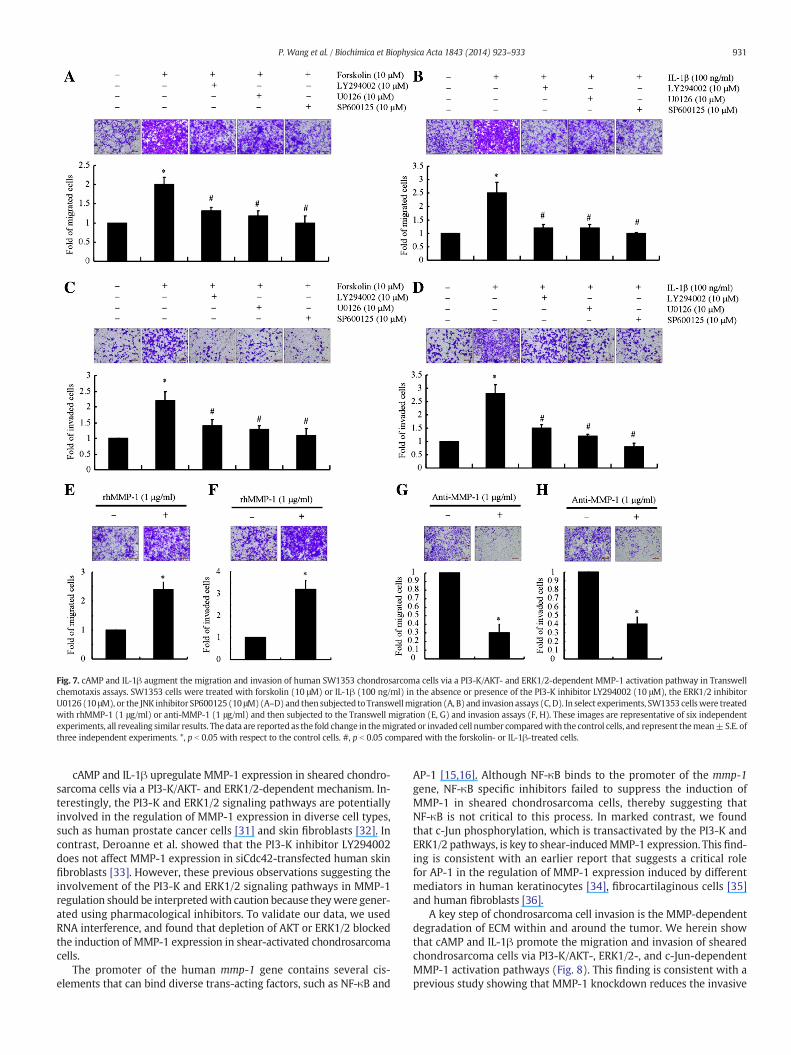
Fig. 7. cAMP and IL-1β augment the migration and invasion of human SW1353 chondrosarcoma cells via a PI3-K/AKT- and ERK1/2-dependent MMP-1 activation pathway in Transwellchemotaxis assays. SW1353 cells were treated with forskolin (10 μM) or IL-1β (100 ng/ml) in the absence or presence of the PI3-K inhibitor LY294002 (10 μM), the ERK1/2 inhibitorU0126 (10 μM), or the JNK inhibitor SP600125 (10 μM)(A–D)and then subjected to Transwellmigration (A, B) and invasion assays (C, D). In select experiments, SW1353 cellswere treatedwith rhMMP-1 (1 μg/ml) or anti-MMP-1 (1 μg/ml) and then subjected to the Transwell migration (E, G) and invasion assays (F, H). These images are representative of six independentexperiments, all revealing similar results. Thedata are reported as the fold change in themigrated or invaded cell number comparedwith the control cells, and represent themean±S.E. ofthree independent experiments. *, p b 0.05 with respect to the control cells. #, p b 0.05 compared with the forskolin- or IL-1β-treated cells.
931P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
cAMP and IL-1β upregulate MMP-1 expression in sheared chondro-sarcoma cells via a PI3-K/AKT- and ERK1/2-dependent mechanism. In-terestingly, the PI3-K and ERK1/2 signaling pathways are potentiallyinvolved in the regulation of MMP-1 expression in diverse cell types,such as human prostate cancer cells [31] and skin fibroblasts [32]. Incontrast, Deroanne et al. showed that the PI3-K inhibitor LY294002does not affect MMP-1 expression in siCdc42-transfected human skinfibroblasts [33]. However, these previous observations suggesting theinvolvement of the PI3-K and ERK1/2 signaling pathways in MMP-1regulation should be interpretedwith caution because theywere gener-ated using pharmacological inhibitors. To validate our data, we usedRNA interference, and found that depletion of AKT or ERK1/2 blockedthe induction of MMP-1 expression in shear-activated chondrosarcomacells.
The promoter of the human mmp-1 gene contains several cis-elements that can bind diverse trans-acting factors, such as NF-κB and
AP-1 [15,16]. Although NF-κB binds to the promoter of the mmp-1gene, NF-κB specific inhibitors failed to suppress the induction ofMMP-1 in sheared chondrosarcoma cells, thereby suggesting thatNF-κB is not critical to this process. In marked contrast, we foundthat c-Jun phosphorylation, which is transactivated by the PI3-K andERK1/2 pathways, is key to shear-inducedMMP-1 expression. This find-ing is consistent with an earlier report that suggests a critical rolefor AP-1 in the regulation of MMP-1 expression induced by differentmediators in human keratinocytes [34], fibrocartilaginous cells [35]and human fibroblasts [36].
A key step of chondrosarcoma cell invasion is the MMP-dependentdegradation of ECM within and around the tumor. We herein showthat cAMP and IL-1β promote the migration and invasion of shearedchondrosarcoma cells via PI3-K/AKT-, ERK1/2-, and c-Jun-dependentMMP-1 activation pathways (Fig. 8). This finding is consistent with aprevious study showing that MMP-1 knockdown reduces the invasive

Fig. 8. Proposed cascade of the signaling events regulatingMMP-1 synthesis in shear-activated human SW1353 chondrosarcoma cells. Fluid shear stress (2 dyn/cm2) induces the produc-tion and release of cAMPand IL-1β by human SW1353 chondrosarcoma cells. The elevated levels of cAMP and IL-1β stimulate the activity of the PI3-K/AKT and ERK1/2 pathways, which inturn induce MMP-1 synthesis via a c-Jun-dependent manner. The activation of MMP-1 augments chondrosarcoma cell migration and invasion.
932 P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
potential of human chondrosarcoma cells [7]; however, the authors didnot elucidate the underlying signaling mechanism [7]. cAMP and IL-1βhave been reported to be involved in the invasion of human tropho-blasts [37] and ovarian carcinoma cells [38] via an MMP-dependentpathway. The activation of PI3-K, which is downstream of cAMP andIL-1β (Fig. 8), induces the invasion of human prostate cancer cells inan MMP-1-dependent manner [31]. In addition, ERK1/2 is involved inmelanomaMMP-1-dependent invasion [39]. As PI3-K and ERK1/2 phos-phorylate c-Jun, which in turn regulates MMP-1 expression, our datasuggest that c-Jun is involved in chondrosarcoma cell invasion. Indeed,we found that treatment of SW1353 chondrosarcoma cells with thec-Jun inhibitor SP600125 suppressed cell invasion induced by exoge-nous forskolin or IL-1β. Ourfinding is in linewith previous data showingthat c-Jun inducesMMP-1 expression and regulates the invasive activityof 143B osteosarcoma cells [40].
In summary, we have delineated the signaling pathway by whichfluid shear stress promotes chondrosarcoma cell invasion via an MMP-1-dependent mechanism. Fluid shear stimulates the production ofcAMP and IL-1β, which induces the phosphorylation of AKT andERK1/2. Activated PI3-K/AKT and ERK1/2 signalingpathways phosphor-ylate c-Jun, which in turn transactivates MMP-1 that is responsible forthe potentiation of the invasion of human chondrosarcoma cells.
5. Conclusions
In this study, we found that MMP-1 is markedly induced in humanchondrosarcoma tissues and shear-activated chondrosarcoma cells.We also deciphered the signaling pathway by which MMP-1 promotesthe invasion of shear-activated chondrosarcoma cells. By reconstructingand deciphering the signaling pathways involved in MMP-1 induction,potential therapeutic strategies that interfere with chondrosarcomacell invasion may be identified.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.bbamcr.2014.01.018.
Competing financial interests
The authors declare no competing financial interests.
Acknowledgements
This work was supported in part or whole by National NaturalScience Foundation of China (31300777), the Fundamental ResearchFunds of China (N120520001 and N120320001), the Liaoning ProvinceHigh School Talent Support Program of China (LJQ2013029) and NIH/NCI U54 CA143868.
References
[1] J.M. Rutkowski, M.A. Swartz, A driving force for change: interstitial flow as amorphoregulator, Trends Cell Biol. 17 (2007) 44–50.
[2] S.R. Chary, R.K. Jain, Direct measurement of interstitial convection and diffusion ofalbumin in normal and neoplastic tissues by fluorescence photobleaching, Proc.Natl. Acad. Sci. U. S. A. 86 (1989) 5385–5389.
[3] S.F. Chang, C.A. Chang, D.Y. Lee, P.L. Lee, Y.M. Yeh, C.R. Yeh, C.K. Cheng, S. Chien, J.J.Chiu, Tumor cell cycle arrest induced by shear stress: roles of integrins and Smad,Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 3927–3932.
[4] N. Koshikawa, G. Giannelli, V. Cirulli, K. Miyazaki, V. Quaranta, Role of cell surfacemetalloprotease MT1-MMP in epithelial cell migration over laminin-5, J. Cell Biol.148 (2000) 615–624.
[5] K.R. Berend, A.P. Toth, J.M. Harrelson, L.J. Layfield, L.A. Hey, S.P. Scully, Association be-tween ratio of matrix metalloproteinase-1 to tissue inhibitor of metalloproteinase-1and local recurrence, metastasis, and survival in human chondrosarcoma, J. BoneJoint Surg. Am. 80 (1998) 11–17.
[6] S.P. Scully, K.R. Berend, W.N. Qi, J.M. Harrelson, Collagenase specificity inchondrosarcoma metastasis, Braz. J. Med. Biol. Res. 32 (1999) 885–889.
[7] X. Jiang, C.M. Dutton, W.N. Qi, J.A. Block, N. Garamszegi, S.P. Scully, siRNA mediatedinhibition of MMP-1 reduces invasive potential of a human chondrosarcoma cellline, J. Cell. Physiol. 202 (2005) 723–730.
[8] J.P. Abulencia, R. Gaspard, Z.R. Healy,W.A. Gaarde, J. Quackenbush, K. Konstantopoulos,Shear-induced cyclooxygenase-2 via a JNK2/c-Jun-dependent pathway regulatesprostaglandin receptor expression in chondrocytic cells, J. Biol. Chem. 278 (2003)28388–28394.
[9] Z.R. Healy, N.H. Lee, X. Gao, M.B. Goldring, P. Talalay, T.W. Kensler, K. Konstantopoulos,Divergent responses of chondrocytes and endothelial cells to shear stress: cross-talk

933P. Wang et al. / Biochimica et Biophysica Acta 1843 (2014) 923–933
among COX-2, the phase 2 response, and apoptosis, Proc. Natl. Acad. Sci. U. S. A. 102(2005) 14010–14015.
[10] Z.R. Healy, F. Zhu, J.D. Stull, K. Konstantopoulos, Elucidation of the signalingnetwork of COX-2 induction in sheared chondrocytes: COX-2 is induced via aRac/MEKK1/MKK7/JNK2/c-Jun-C/EBPbeta-dependent pathway, Am. J. Physiol. CellPhysiol. 294 (2008) C1146–C1157.
[11] P. Wang, F. Zhu, K. Konstantopoulos, Prostaglandin E2 induces interleukin-6 expres-sion in human chondrocytes via cAMP/protein kinase A- and phosphatidylinositol3-kinase-dependent NF-kappaB activation, Am. J. Physiol. Cell Physiol. 298 (2010)C1445–C1456.
[12] P. Wang, F. Zhu, N.H. Lee, K. Konstantopoulos, Shear-induced interleukin-6 synthesisin chondrocytes: roles of E prostanoid (EP) 2 and EP3 in cAMP/protein kinase A- andPI3-K/Akt-dependent NF-kappaB activation, J. Biol. Chem. 285 (2010) 24793–24804.
[13] M.B. Goldring, Culture of immortalized chondrocytes and their use as models ofchondrocyte function, Methods Mol. Med. 100 (2004) 37–52.
[14] M.B. Goldring, J.R. Birkhead, L.F. Suen, R. Yamin, S. Mizuno, J. Glowacki, J.L. Arbiser,J.F. Apperley, Interleukin-1 beta-modulated gene expression in immortalizedhuman chondrocytes, J. Clin. Invest. 94 (1994) 2307–2316.
[15] J.L. Rutter, U. Benbow, C.I. Coon, C.E. Brinckerhoff, Cell-type specific regulation of humaninterstitial collagenase-1 gene expression by interleukin-1 beta (IL-1 beta) in humanfibroblasts and BC-8701 breast cancer cells, J. Cell. Biochem. 66 (1997) 322–336.
[16] J.A. Green, P.T. Elkington, C.J. Pennington, F. Roncaroli, S. Dholakia, R.C. Moores, A.Bullen, J.C. Porter, D. Agranoff, D.R. Edwards, J.S. Friedland, Mycobacterium tubercu-losis upregulates microglial matrix metalloproteinase-1 and -3 expression andsecretion via NF-kappaB- and activator protein-1-dependent monocyte networks,J. Immunol. 184 (2013) 6492–6503.
[17] P. Wang, F. Zhu, K. Konstantopoulos, Interleukin-6 synthesis in human chondrocytesis regulated via the antagonistic actions of prostaglandin PGE2 and 15d-PGJ2, PLoSOne 6 (2011) e27630.
[18] P. Wang, F. Zhu, K. Konstantopoulos, The antagonistic actions of endogenousinterleukin-1beta and 15-deoxy-Delta12,14-prostaglandin J2 regulate the temporalsynthesis of matrix metalloproteinase-9 in sheared chondrocytes, J. Biol. Chem. 287(2012) 31877–31893.
[19] P.Wang, F. Zhu, Z. Tong, K. Konstantopoulos, Response of chondrocytes to shear stress:antagonistic effects of the binding partners Toll-like receptor 4 and caveolin-1, FASEB J.25 (2011) 3401–3415.
[20] E. Bastida, L. Almirall, M.C. Bertomeu, A. Ordinas, Influence of shear stress ontumor-cell adhesion to endothelial-cell extracellular matrix and its modulation byfibronectin, Int. J. Cancer 43 (1989) 1174–1178.
[21] H. Qazi, Z.D. Shi, J.M. Tarbell, Fluid shear stress regulates the invasive potential ofglioma cells via modulation of migratory activity and matrix metalloproteinaseexpression, PLoS One 6 (2011) e20348.
[22] L. Ottaviano, K.L. Schaefer, M. Gajewski, W. Huckenbeck, S. Baldus, U. Rogel, C.Mackintosh, E. de Alava, O. Myklebost, S.H. Kresse, L.A. Meza-Zepeda, M. Serra, A.M.Cleton-Jansen, P.C. Hogendoorn, H. Buerger, T. Aigner, H.E. Gabbert, C. Poremba,Molecular characterization of commonly used cell lines for bone tumor research: atrans-European EuroBoNet effort, Genes Chromosomes Cancer 49 (2010) 40–51.
[23] T.L. Chen, M.T. Sheu, Y.C. Liang, Y.J. Lin, M.S. Hsieh, C.H. Chen, Disease-modifying ef-fects of glucosamine on Interleukin-1b-treated chondrosarcoma cells (SW1353)under normoxic and hypoxic conditions, J. Exp. Clin. Med. 2 (2010) 17–28.
[24] B. Reel, G.B. Sala-Newby,W.C.Huang,A.C.Newby, Diverse patternsof cyclooxygenase-independent metalloproteinase gene regulation in human monocytes, Br. J.Pharmacol. 163 (2011) 1679–1690.
[25] R.K. Jain, Normalization of tumor vasculature: an emerging concept in antiangiogenictherapy, Science 307 (2005) 58–62.
[26] A. Kawashima, Y. Okada, I. Nakanishi, Y. Ueda, K. Iwata, A. Roessner, Immunolocal-ization of matrix metalloproteinases and tissue inhibitors of metalloproteinases inhuman chondrosarcomas, Gen. Diagn. Pathol. 142 (1997) 129–137.
[27] L. Zheng, Y. Huang, W. Song, X. Gong, M. Liu, X. Jia, G. Zhou, L. Chen, A. Li, Y. Fan,Fluid shear stress regulates metalloproteinase-1 and 2 in human periodontalligament cells: involvement of extracellular signal-regulated kinase (ERK) and P38signaling pathways, J. Biomech. 45 (2012) 2368–2375.
[28] Z.D. Shi, X.Y. Ji, D.E. Berardi, H. Qazi, J.M. Tarbell, Interstitial flow induces MMP-1expression and vascular SMC migration in collagen I gels via an ERK1/2-dependentand c-Jun-mediated mechanism, Am. J. Physiol. Heart Circ. Physiol. 298 (2010)H127–H135.
[29] H. Yokota, M.B. Goldring, H.B. Sun, CITED2-mediated regulation of MMP-1 andMMP-13 in human chondrocytes under flow shear, J. Biol. Chem. 278 (2003)47275–47280.
[30] N. Zayed, H. Afif, N. Chabane, L. Mfuna-Endam, M. Benderdour, J. Martel-Pelletier, J.P.Pelletier, R.K. Motiani, M. Trebak, N. Duval, H. Fahmi, Inhibition of interleukin-1beta-inducedmatrixmetalloproteinases 1 and 13 production in human osteoarthriticchondrocytes by prostaglandin D2, Arthritis Rheum. 58 (2008) 3530–3540.
[31] Z.Z. Zeng, Y. Jia, N.J. Hahn, S.M. Markwart, K.F. Rockwood, D.L. Livant, Role of focaladhesion kinase and phosphatidylinositol 3′-kinase in integrin fibronectinreceptor-mediated, matrix metalloproteinase-1-dependent invasion by metastaticprostate cancer cells, Cancer Res. 66 (2006) 8091–8099.
[32] C.-H. Park, J.H. Chung, Epidermal growth factor-inducedmatrix metalloproteinase-1expression is negatively regulated by p38 MAPK in human skin fibroblasts,J. Dermatol. Sci. 64 (2011) 134–141.
[33] C.F. Deroanne, D. Hamelryckx, T.T. Ho, C.A. Lambert, P. Catroux, C.M. Lapiere, B.V.Nusgens, Cdc42 downregulates MMP-1 expression by inhibiting the ERK1/2 path-way, J. Cell Sci. 118 (2005) 1173–1183.
[34] K.A. Kang, R. Zhang, M.J. Piao, D.O. Ko, Z.H. Wang, K. Lee, B.J. Kim, T. Shin, J.W. Park,N.H. Lee, B.S. Yoo, J.W. Hyun, Inhibitory effects of triphlorethol-A onMMP-1 inducedby oxidative stress in human keratinocytes via ERK and AP-1 inhibition, J. Toxicol.Environ. Health A 71 (2008) 992–999.
[35] S. Kapila, Y. Xie, W. Wang, Induction of MMP-1 (collagenase-1) by relaxin infibrocartilaginous cells requires both the AP-1 and PEA-3 promoter sites, Orthod.Craniofac. Res. 12 (2009) 178–186.
[36] X.Wang, Z. Bi, W. Chu, Y. Wan, IL-1 receptor antagonist attenuates MAP kinase/AP-1activation and MMP1 expression in UVA-irradiated human fibroblasts induced byculture medium from UVB-irradiated human skin keratinocytes, Int. J. Mol. Med.16 (2005) 1117–1124.
[37] E. Staun-Ram, S. Goldman, E. Shalev, Ets-2 and p53 mediate cAMP-induced MMP-2expression, activity and trophoblast invasion, Reprod. Biol. Endocrinol. 7 (2009)135.
[38] C. Denkert, I. Koch, S. Berger, M. Kobel, A. Siegert, S. Hauptmann, Cytokine-suppressive anti-inflammatory drugs (CSAIDs) inhibit invasion andMMP-1 produc-tion of ovarian carcinoma cells, Cancer Lett. 195 (2003) 101–109.
[39] M.B. Weiss, E.V. Abel, M.M. Mayberry, K.J. Basile, A.C. Berger, A.E. Aplin, TWIST1 is anERK1/2 effector that promotes invasion and regulates MMP-1 expression in humanmelanoma cells, Cancer Res. 72 (2012) 6382–6392.
[40] R. Kimura, C. Ishikawa, T. Rokkaku, R. Janknecht, N. Mori, Phosphorylated c-Jun andFra-1 induce matrix metalloproteinase-1 and thereby regulate invasion activity of143B osteosarcoma cells, Biochim. Biophys. Acta 1813 (2011) 1543–1553.
Copyright © 2022 FDOKUMEN




















