
Interactions between atmospheric CO2 enrichment and soil fauna
12
Plant and Soil 224: 123–134, 2000. © 2000 Kluwer Academic Publishers. Printed in the Netherlands. 123 Interactions between atmospheric CO 2 enrichment and soil fauna Marie-Madeleine Coûteaux 1, * and Thomas Bolger 2 1 CEFE-CNRS 1919 Route de Mende, F-34293 Montpellier Cedex 5, France and 2 Department of Zoology, University College Dublin, Belfield, Dublin 4, Ireland Key words: global change, soil fauna, decomposition, litter quality, climate Abstract We have reviewed the responses of soil fauna to increased concentrations of atmospheric CO 2 and the consequent climate change. These will affect several attributes of animal populations and communities including their density, biomass, diversity, activity, rates of consumption, life history parameters and migration ability. Changes in the quality and quantity of litter and global warming are the main factors which are expected to modify soil fauna. Although changes have been observed in several attributes of the soil fauna as a consequence of increased con- centrations of atmospheric CO 2 , no general trend which might allow to the prediction of a general pattern of response has been identified. Because of the complexity of the biological mechanisms and the synergetic action of several factors, the few resulting responses reported in the literature are inconclusive. However, some aspects of the situation deserve more attention. These include the consequences of (1) changes in the food resources for soil fauna in the litter layer and in the rhizosphere, (2) the consumption of low quality litter by the macrofauna, (3) the change in life span in response to temperature elevation, (4) the enhancement of earthworm burrowing activity and (5) the changes in community composition arising because of specific differential resistance to adverse conditions. Introduction It is widely accepted that increases in concentrations of the atmospheric CO 2 , and of other greenhouse gases, will greatly modify ecosystem physiology. In terrestrial ecosystems, the effects of this increase on carbon cycling result from the pattern of transloca- tion of C within the plant and its transformation by microorganisms. These are, in turn, controlled by the interactions between soil fauna and this microbial community (Coûteaux and Bottner, 1994). The direct impact of elevated concentrations of CO 2 on the soil fauna are likely to be negligible because these organisms are already adapted to the already high CO 2 concentration in the soil (van Veen et al., 1991). However, indirect effects, such as changes in the quality and quantity of litter, and global warming are expected to affect their density, bio- mass and the composition of the communities. Con- sequently, the impact of soil fauna on soil processes, * Fax: +33 467 41 21 38; E-mail: [email protected] due either to their physical activity such as grinding or burrowing or to their chemical activity (e.g., consump- tion, mineralisation, enzymatic activity), could alter the rate of C transformations. The question is: will soil fauna play a role in the modification of C storage in the soil? In this paper, we review the studies dealing with soil fauna subjected to elevated atmospheric CO 2 , in relation to litter produced under elevated levels of at- mospheric CO 2 and under warmer conditions and we have tried to evaluate the possible effects of global change on the composition and activity of soil fauna and the consequence of these effects on the C soil reservoir as source or sink of CO 2 (Figure 1). Soil fauna under elevated atmospheric CO 2 As indicated above, elevated concentrations of CO 2 are unlikely to have any direct effects on the soil fauna because soil biota are usually exposed to high concentrations in the soil atmosphere. Nevertheless,
Transcript of Interactions between atmospheric CO2 enrichment and soil fauna

Plant and Soil224: 123–134, 2000.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
123
Interactions between atmospheric CO2 enrichment and soil fauna
Marie-Madeleine Coûteaux1,∗ and Thomas Bolger2
1CEFE-CNRS 1919 Route de Mende, F-34293 Montpellier Cedex 5, France and2Department of Zoology,University College Dublin, Belfield, Dublin 4, Ireland
Key words:global change, soil fauna, decomposition, litter quality, climate
Abstract
We have reviewed the responses of soil fauna to increased concentrations of atmospheric CO2 and the consequentclimate change. These will affect several attributes of animal populations and communities including their density,biomass, diversity, activity, rates of consumption, life history parameters and migration ability. Changes in thequality and quantity of litter and global warming are the main factors which are expected to modify soil fauna.Although changes have been observed in several attributes of the soil fauna as a consequence of increased con-centrations of atmospheric CO2, no general trend which might allow to the prediction of a general pattern ofresponse has been identified. Because of the complexity of the biological mechanisms and the synergetic action ofseveral factors, the few resulting responses reported in the literature are inconclusive. However, some aspects ofthe situation deserve more attention. These include the consequences of (1) changes in the food resources for soilfauna in the litter layer and in the rhizosphere, (2) the consumption of low quality litter by the macrofauna, (3) thechange in life span in response to temperature elevation, (4) the enhancement of earthworm burrowing activity and(5) the changes in community composition arising because of specific differential resistance to adverse conditions.
Introduction
It is widely accepted that increases in concentrationsof the atmospheric CO2, and of other greenhousegases, will greatly modify ecosystem physiology. Interrestrial ecosystems, the effects of this increase oncarbon cycling result from the pattern of transloca-tion of C within the plant and its transformation bymicroorganisms. These are, in turn, controlled bythe interactions between soil fauna and this microbialcommunity (Coûteaux and Bottner, 1994).
The direct impact of elevated concentrations ofCO2 on the soil fauna are likely to be negligiblebecause these organisms are already adapted to thealready high CO2 concentration in the soil (van Veenet al., 1991). However, indirect effects, such aschanges in the quality and quantity of litter, and globalwarming are expected to affect their density, bio-mass and the composition of the communities. Con-sequently, the impact of soil fauna on soil processes,
∗ Fax: +33 467 41 21 38;E-mail: [email protected]
due either to their physical activity such as grinding orburrowing or to their chemical activity (e.g., consump-tion, mineralisation, enzymatic activity), could alterthe rate of C transformations. The question is: will soilfauna play a role in the modification of C storage in thesoil?
In this paper, we review the studies dealing withsoil fauna subjected to elevated atmospheric CO2, inrelation to litter produced under elevated levels of at-mospheric CO2 and under warmer conditions and wehave tried to evaluate the possible effects of globalchange on the composition and activity of soil faunaand the consequence of these effects on the C soilreservoir as source or sink of CO2 (Figure 1).
Soil fauna under elevated atmospheric CO2
As indicated above, elevated concentrations of CO2are unlikely to have any direct effects on the soilfauna because soil biota are usually exposed to highconcentrations in the soil atmosphere. Nevertheless,

124
Figure 1. Conceptual schema of the effect of elevated atmosphericCO2 on soil fauna.
changes in microbial biomass and composition havebeen reported (Dhillion et al., 1996; O’Neill, 1994;Zak et al., 1996) which suggest a possible shift infaunal communities in response to modification in thefood supply. In some experiments, where plants weregrown under elevated CO2, responses of some faunalgroups have been examined. No significant changesin the densities of soil protozoa were observed in twoexperiments where a mixed grassland mini-ecosystemwas exposed to double the current concentration ofatmospheric CO2 for 1 year (Coûteaux and Roy, un-published data) and where sweet chestnut trees wereexposed for 2 years in growth chambers (Coûteauxand Mousseau, unpublished data). However, becauseof the high variability in density and the inaccuracyof the counting method, differences between popula-tions of protozoa would have to be very pronounced ifsignificant differences were to be detected.
Similarly, Freckman et al. (1991) found that thedensity of nematodes, in soils from stands of westernwheatgrass and blue grama grasses, were unaffectedafter 2 years exposure to elevated concentration ofatmospheric CO2, and Runion et al. (1994) did notfind any significant differences in the densities of mosttypes of nematodes in the rhizosphere of cotton cultiv-ated under 350 and 550µl l−1 in a FACE experiment(free-air CO2 enrichment) after an exposure of 125days. However, in the latter study, the number ofwhat are described as saprophagous nematodes was
significantly higher while the number of parasitic in-dividuals tended to be lower under FACE. The authorssuggested that the additional plant biomass (Prior etal., 1994) and organic carbon (Wood et al., 1994)present under FACE would explain the higher numberof saprophagous nematodes and therefore competitionfor space or resources would have occurred betweenthe two groups. In contrast, Newton et al. (1995) didnot observe differences in the total abundance of nem-atodes and their trophic composition in a pasture soilexposed to 350, 525 and 700µl l−1 atmospheric CO2.
Densities of microarthopods were low in the cot-ton rhizosphere of the FACE experiment describedby Runion et al. (1994). However, there was a trendfor Collembola populations to be higher under FACEin the August sample and higher under wet condi-tions in June. More species of fungi were isolatedfrom Collembola intestines in June. Thus, because ofthe interactions between atmospheric CO2 concentra-tion and seasonal variations, the CO2 concentrationeffect was only evident under favourable moistureconditions.
Enchytraeid abundance was consistently 50–100%higher in a pasture sandy soil under 700µl l−1 CO2and their grazing activity appeared to maintain themicrobial biomass stability despite the fact that be-lowground production was stimulated under high CO2(Newton et al., 1995). Earthworm biomass increasedin this soil (Yeates, 1996, personal communication inTate and Ross, 1997).
There are very few studies of these direct effectsof elevated concentrations of CO2 but they suggestthat the effects arise because of trophic relationships,changes in organic matter quality affecting saprophag-ous groups and changes in the composition of themicrobial community affecting microbivorous anim-als.
Another important indirect stimulatory effect wasobserved by Zaller and Arnone (1997). Earthwormsurface cast production was 35% greater over one yearin open top and open bottom rings under elevated CO2concentrations (610µl l−1). This was particularlyobvious during the wet period. Nevertheless neitherthe total nitrogen nor total organic C content of thecasts were affected. Because of their burrowing activ-ity, earthworm worked soils have higher pore volumes(Hoeksema and Jongrius, 1959), increased infiltrationrates/water-holding capacity (Douglas et al., 1989;Stockdill, 1966), more water-stable aggregates andgreater aeration (Kretzschmar 1978, in Lee 1985) thansoils without earthworms. An important step towards

125
fully understanding the physical effects of earthwormson soils will be to determine the factors that determinetheir movement (Lavelle, 1988). Thus the enhance-ment of the engineering role of earthworms underelevated CO2 could improve the structural quality ofthe soil by burrowing in the soil and mixing organicmatter to mineral particules.
Litter quality
The soil biota represent a sensitive link between plantdetritus and plant-available nutrients. Thus, factorswhich influence detrital quality and quantity are likelyto be highly significant (Swift et al., 1998). Alter-ation of the chemical composition of plant materialproduced under elevated atmospheric CO2 has beenreported widely (Cotrufo et al., 1998a,c; Coûteaux etal., 1995, 1999; Lincoln et al., 1993). Although theresponse of plants to elevated CO2 is species specific,there is a large consensus about the significant reduc-tion of nitrogen concentration in green leaves and alsoin senescent tissue (Cotrufo et al., 1998c; Coûteauxet al., 1999). Variation in lignin concentrations is alsospecies dependent with generally significant increasesfor woody plants and non-significant reductions forherbaceous species (Coûteaux et al., 1999). Some spe-cies have been reported to accumulate starch (Lindrothet al., 1993) but this compound is reallocated duringthe senescence process and does not impact soil fauna.Defensive carbon compounds such as phenols, whichincrease under elevated CO2, can have a negative ef-fect during the early stages of decomposition but theyare leached rapidly.
Effect on populations and communities
Changes in litter quality could affect the abund-ance of some faunal species either directly or indir-ectly through their food supply. A study was carriedout on the decomposition of sweet chestnut litterproduced under elevated (700µl l−1) and ambi-ent concentrations of atmospheric CO2, and incub-ated in the laboratory in the presence of soil faunacombinations of increasing complexity. The unitswere inoculated with four types of faunal combin-ations of increasing complexity: protozoa (P units),P+nematodes (NP units), P+N+Collembola (CNPunits), P+N+C+isopoda (ICNP units). The experi-mental units were leached with 150 ml of distilledwater every two weeks and protozoa, nematodes and
rotifera counted in the leachates on six occasionsbetween the 10th to the 24th week of incubations(Coûteaux et al., 1991). The total numbers of protozoawere depressed in the units containing CO2-enrichedlitter inoculated only with protozoa and in the mostcomplex ones. In these units, flagellates were rare andthe total protozoan density reflected the responses ofthe amoebae and the ciliates which were both lessnumerous in CO2-enriched litter than in the control.Flagellates were almost absent in all the control units.In the NP and CNP units with CO2-enriched litter,flagellates became very abundant after 15 weeks ofincubation. Using litterbags buried in a wheat fieldand in a fallow for 5 months, Frederiksen and Rønn(unpublished data) also found reduced densities ofciliates in straw, grown under 700µl l−1 CO2, whencompared with control straw which had been grownunder ambient air. The number of active ciliates werecorrelated with bacterial biomass.
Rotifera were not detected in the units containingCO2-enriched litter whereas they reached densities ofup to 6000 individuals per unit in the control (Fig-ure 1). At the beginning of the experiment nematodeabundance was lower in the units with CO2-enrichedlitter but after 15 weeks they were far more abundantin those units and decreases were observed in the con-trol units. A decrease in pH was observed during thetime course of the experiment in the CO2-enriched lit-ter, especially in the CNP and the ICNP units. The pHof the leachates from the control increased slowly to6–7, whereas it decreased to 4–5 in the CO2-enrichedlitter (Coûteaux et al., 1991). The low pH may bethe limiting factor in CO2-enriched litter and the highpH the stimulating factor in the control for amoebaeand ciliates. A change in microbial composition, es-pecially an invasion of white-rot fungi after the 12thweek, could explain the late increase of flagellates andnematodes.
Interaction between animals and litter quality andconsequence in terms of mineralisation
The few results which are available indicate differentresponses to different factors associated with globalchange. This is shown clearly in the experiment de-scribed above (Coûteaux et al., 1991). Decompositionrates of CO2-enriched litter, exposed to simple foodwebs were lower than a control litter whereas exposureto more complex food webs induced higher decom-position rates of the CO2-enriched litter comparedto the control. This shift in decomposition poten-

126
tial in response to the food web complexity of theanimal community was explained by different con-comitant processes (i) the dissemination of white-rotfungi propagules by soil fauna, (ii) the lower com-petition within microbial communities in N-poor litterproduced under elevated CO2, and (iii) the recalcit-rance of N-shielded lignin in N-rich litter producedunder ambient air. These results were obtained froma laboratory microcosm experiment but they raise thequestion of the effects of the interaction between thesoil biodiversity and global change on soil organicmatter decomposition. The relatively higher contribu-tion of the fauna to the decomposition of low qualityresidues was also reported by Seastedt (1984) and Tianet al. (1997).
Consumption
Although consumption was not directly measured inthe experiment of Coûteaux et al. (1991), the resultssuggest that the saprophageous soil fauna increasedits consumption when feeding on CO2-enriched litter.In order to satisfy their nitrogen requirements, anim-als consume a higher quantity of N-poor litter. Thisresponse is analogous to that reported for insect herb-ivores which respond to low nitrogen, high starch,CO2-enriched plant by increasing their consumption(Dury et al., 1998; Fajer et al., 1991; Lincoln et al.,1993). Nevertheless, Roth et al. (1998) showed nochange in larval consumption of CO2-enriched foliagebut a reduction of growth and food processing effi-ciency. Cotrufo et al. (1998b) observed that the elev-ated CO2 treatment decreased the rate of consumptionof ash leaf litter byOniscus asellus. Although the res-ults of the experiment stress a possible change of thepalatability of CO2-enriched litter, they are inconclus-ive because of the short duration of the experiment(3 days) and because the animals, which were ableto choose between the two kinds of litter (N-poorand -rich), preferred nutrient-rich litter as expected.However, Rushton and Hassall (1983) showed thatthe isopod,Armadillidium vulgare, can compensatefor low digestibility of some foods by increasing itsconsumption rate. In terms of carbon storage underelevated atmospheric CO2, the question that has tobe raised concerns the possible change in consump-tion rate when animals have no choice. In a laborat-ory experiment, Hättenschwiler et al. (1999) showedthat bothOniscus asellusand Porcellio scabercon-sumed significantly more beech leaf litter and sprucebranchlets which had been grown under elevated CO2
than plant material grown at ambient CO2. They alsoshowed, in a field experiment, that the decomposi-tion of these plant species was significantly higher inlarge mesh units than in small mesh units showing thatthe soil fauna consumption enhanced decomposition.However this effect did not differ between N-rich andN-poor litter because the soil fauna were not forced toconsume any litter.
Litter quantity
Elevated concentrations of atmospheric CO2 are ex-pected to increase litter production through C fertil-isation (e.g. El Kohen et al., 1992; Mousseau andEnoch, 1989; Rouhier et al., 1994). No experiment,with CO2-enriched litter, has been performed to assessthe effect of increased litter production on soil faunaand C storage. Nevertheless an experimental doublingof litter was carried out in a mixed deciduous forestand the dynamics of nematodes, Collembola and mac-rofauna populations were studied over a period of 4years (Arpin et al., 1995; David et al., 1991; Pongeet al., 1993). The density of nematodes was reducedby the increased litter input with an enhanced domin-ance of fungal feeders compared to bacterial feeders.By contrast, no change was observed in the numbersor biomass of Collembola or macrofauna althoughthe decomposition rate of the litter diminished. Theseresults imply that under these conditions there is noevidence that food limitation for the litter-dwellingspecies whatever their diet. A two-fold increase inlitterfall did not modify meso- and macrofauna dens-ity and animals were unable to eat and remove theadditional food supply. This means that meso- andmacrofauna were at their biotic capacity despite thefact that there is no competition for food and space andthat densities are controlled by other intrinsic factorsregulating the stability of the communities. IndeedBengtsson et al. (1998) suggest that the view that suchfood webs are donor controlled should be reconsideredand that the major interactions in some soil are re-ciprocal, i.e. that consumers to some degree controlthe density of their resources. Judas (1990) showeda negative effect of increased litter supply for anecicand endogeic earthworms in a beechwood on lime-stone which confirms that the litter input is not thelimiting factor for these species. However the rangeof predicted increase of litter input is far lower thanthose used in those experimental designs (El Kohen etal., 1992; Mousseau and Enoch, 1989; Rouhier et al.,

127
1994) and negative effects are improbable. Neverthe-less, it is likely that in the most extreme ecosystems,where the production of plant residues is low, changesin plant production could lead to a critical thresholdwhich would have effects on the fauna.
Food resource
For microbivorous animals, the size and compositionof the microbial biomass is a determining factor inthe food resource. Microbial biomass is expected toincrease under elevated CO2 because the annual car-bon input exceeds the energy required for microbialmaintenance. However, this potential increase couldbe counteracted by intensified feeding along the soilfood web (Niklaus and Körner, 1996). Thus micro-bial biomass may not increase and may even decreasedue to grazing but its activity would be enhanced. Soactual biomass size is not an indication of the foodresource for animals but metabolic quotient may bea better indicator. The composition of the microbialbiomass is likely to determine animal response to el-evated atmospheric CO2. Enhancement of mycorrhizalinfection by exposure of seedlings to elevated CO2(O’Neill et al., 1987) could interact with the knowngrazing activity on mycorrhiza (Cromack et al., 1988;Rabatin and Stinner, 1985; Reddel and Spain 1991;Warnock et al., 1982). Resource quality has some con-trol over the biotic factors affecting the relationshipbetween plants and mycorrhizal fungi. For example,the response of fungal respiration to grazing intensityhas been shown to be determined by the nutrient statusof the resource on which the fungus is growing (Han-lon, 1981). Depending on the nitrogen status of a soilthe grazing activity by the soil fauna can cause dif-ferent outcomes (Setälä et al., 1997). In N-poor soilsa complex fauna reduced the amount of pine biomassproduction, presumably through their grazing impacton the ectomycorrhizae, whereas it had no effect inN-rich soils.
When offered a variety of foods, the numbers ofLumbricusspp. associated with each food type wasshown to be negatively correlated with C:N and finalpolyphenol content of the food (Hendriksen, 1990).Earthworms have also been shown to grow morequickly and to produce more cocoons when fed onnitrogen rich diets (Evans and Guild, 1948). Thereis also evidence that food quality can be important indetermining the development and fecundity of Colle-mbola (Booth and Anderson, 1979; Snider, 1971).
Change in bacteria/fungi ratio is expected to induceshift in species composition in microfauna.
In general, the results of dynamic laboratory stud-ies (mainly with microarthropods) have shown that,in the short term (days and weeks), faunal grazingincreases microbial respiration and bacterial popula-tions and that, in some cases, there is a correspondingreduction in fungal populations (Hanlon and Ander-son, 1979, 1980; Huish et al., 1985; Scheu andWolters, 1991; Setälä and Huhta, 1990; Setälä etal., 1988). However, microarthropod grazing pres-sures higher than optimum have been found to havethe reverse effect on respiration (Hanlon and Ander-son 1979, 1980). Setälä and Huhta (1990) found thatafter approximately 1 year, the presence of a complexfauna had a negative effect on decomposition relat-ive to the controls, a similar finding to that of Scheuand Wolters (1991) after 5–6 months. The latter studyconcluded that the influence of fragmentation on res-piration rates was restricted to the initial few weeksafter the event; however, Setälä and Huhta believe thatthe effect was due to change in litter quality as theresource decomposed.
Global warming
In addition to change in litter properties, the effectsof increased greenhouse-gas concentrations have beenpredicted from global atmospheric circulation modelsand there is a general agreement that the primary ef-fect will be a global warming of∼4◦C. Therefore bothaspects cannot be dissociated.
Effect on populations and communities
Soil warming experiments in natural ecosystems havebeen used to address some important questions in-cluding the effects of soil warming on biochemicalprocesses (e.g. decomposition, rock weathering, nitro-gen cycle, and trace gas emission), and the feedbackthat changes in these processes may have on atmo-sphere and climate change. Buried heating cable hasbeen used to elevate soil temperature by 5◦C (Peter-john et al., 1993, 1994); infrared radiators suspendedabove the surface have been used to simulate an ac-tual mechanism by which anthropogenic greenhousegases warms the ground (Harte et al., 1995). Thissoil warming influenced the soil moisture dependingon the depth, the year, the period of the year andthe microclimate. Soil mesofauna responded to these

128
variations in various ways: artificial soil heating in-creased mesofaunal biomass and diversity, except inthe dry zone during a dry year suggesting a pos-sible soil moisture and temperature optimum (Harteet al., 1996). Nevertheless these authors concludedthat within the natural range of interannual and spa-tial variation, no unique set of microclimate conditionexists, the biomass response to a change in micro-climate by either the warming treatment or by theinterannual variation is opposite to that in response toa change in microclimate across a spatial gradient. In-stead of an optimum, this response to soil heating canbe interpreted in conjunction with a moisture thresholdconcept: soil heating increases soil mesofauna bio-mass and diversity provided that soil moisture does notdecrease under a defined threshold which was reachedby Harte et al. (1996) when the spatial effect (dryzone) is combined with the interannual effect (dryyear). Coulson et al. (1996) conducted an experimentto measure the effect of summer warming on the totalpopulation densities of soil-dwelling microarthropods.Small polythene tents were used in order to increasethe heat budget available to the soil animals by about10% over 3 years in a polar semi-desert and a tundraheath, in Norway. Abundance of oribatid mites didnot respond to the temperature increase, but numbersof Collembola were significantly reduced at the polarsemi-desert but not at the tundra heath. It was alsopointed out that the abundance of Collembola declinedat both sites during a dry midsummer.
Translocating undisturbed soil cores into areaswith warmer climates is another method used to sim-ulate global warming. The latitudinal translocationtechnique has been used to expose soil to differentchemical and physical climates with a minimum ofdisturbance and in a natural environment (Berg etal., 1997; Bottner et al., 1999). Enchytraeids, dip-tera larvae and tardigrades were studied in a similarexperiment using altitudinal translocation (Briones etal., 1997). Soil cores were transplanted from a highaltitude site to a lower site, resulting in a mean annualtemperature increase of 2.5◦C. The experiment lastedfor about 3 months over a summer period. The re-sponse of the enchytraeids was species dependent. Theabundance ofCognettia sphagnetorumincreased withtemperature but its vertical distribution was linkedto moisture suggesting a migratory ability. This wasnot the case forCernosvitoviella atratawhich diedunder the dry conditions induced by the increase intemperature.Achaeta eiseniwas more tolerant andits density increased with temperature. Diptera lar-
vae were sensitive to lower moisture content inducedby higher temperature whereas tardigrades using theirability to become anhydrobiose were non-affected bydry conditions. In a mesocosm experiment, Sulkava etal. (1996) showed that, at low soil moisture content,microarthropods reduced the abundance of nematodesand enchytraeids and that these reductions were re-flected in reduced nitrogen mineralisation. However,under conditions of high soil moisture, the enchytraeidpopulation, in particular, was not controlled by themicroarthropods and nitrogen mobilization was notaffected by the presence of the arthropods.
Life cycle
Living organisms differ in terms of time to first repro-duction, fecundity, number of times they breed, lifespan and other life history features. Changes in tem-perature are generally reported to directly influencethe lengths of life cycles or parts thereof (David etal., 1999; Tsukamoto and Watanabe, 1977). Temper-ature controls cocoon production, cocoon incubationand hatchling growth of enchytraeids and earthworms(Briones et al., 1997; Butt et al., 1992). Tsukamotoand Watanabe (1977) showed that the incubation timeof the cocoons became shorter as temperature rose,but the hatchability and the number of young wormsemerging from a single cocoon decreased with the risein temperature. Nevertheless, these life history traitshave an optimum value beyond which the response isreversed.
Climate change is expected to modify seasonlength which is one of the major determinant factorsfor species such asPolydesmus inconstans(Snider,1981a). In northern areas, if the timing of snowmelt isadvanced by warming and the freezing period appearslater, a longer summer period may allow more stadiato be achieved within a year which could thus reducethe longevity of the individuals which might be com-pensated by an increase in fecundity (Snider, 1981b;Sutton et al., 1984). Sutton et al. (1984), comparingpopulations of the terrestrial isopod,Armadillidiumvulgare, observed that populations from an area ofMediterranean climate in California had a shorter timeto maturation than English ones. On the other hand ifthe summer temperature sum becomes lower, the lifecycle from egg to juvenile to egg-laying adults maynot be completed (Swift et al., 1998). Low temperatureduring the summer was shown to reduce the supercool-ing ability of microarthropods in the Arctic (Coulsonet al., 1995).
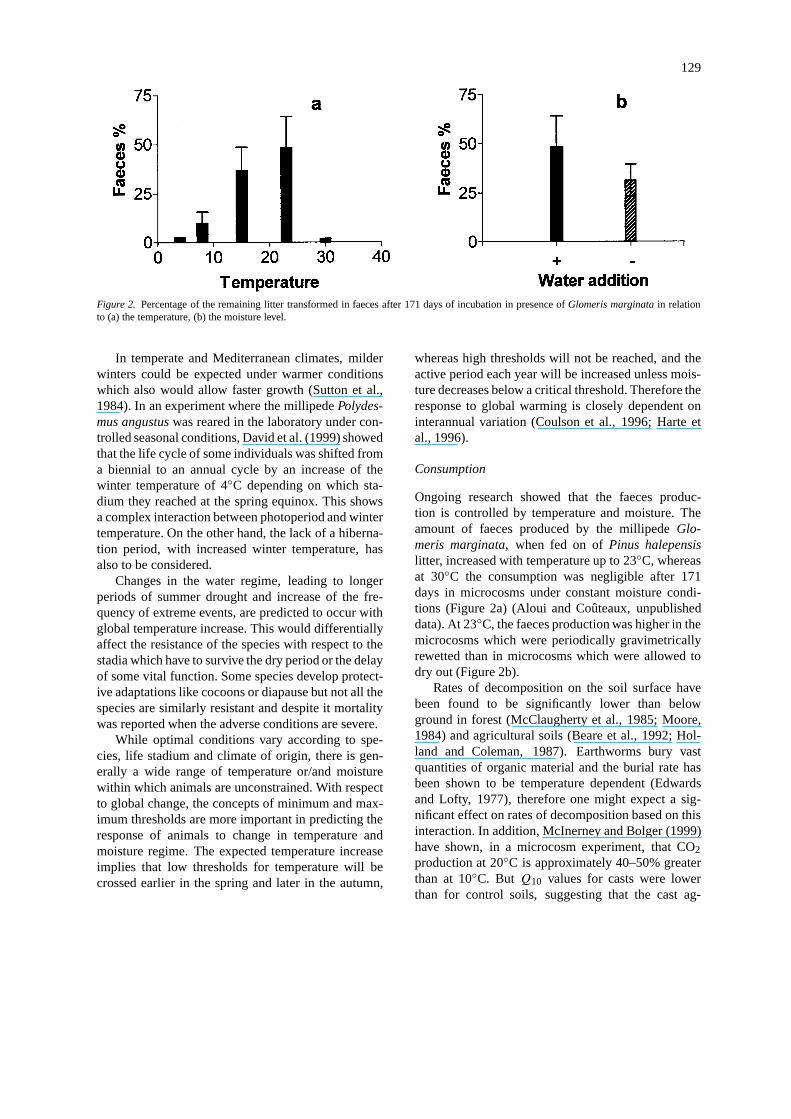
129
Figure 2. Percentage of the remaining litter transformed in faeces after 171 days of incubation in presence ofGlomeris marginatain relationto (a) the temperature, (b) the moisture level.
In temperate and Mediterranean climates, milderwinters could be expected under warmer conditionswhich also would allow faster growth (Sutton et al.,1984). In an experiment where the millipedePolydes-mus angustuswas reared in the laboratory under con-trolled seasonal conditions, David et al. (1999) showedthat the life cycle of some individuals was shifted froma biennial to an annual cycle by an increase of thewinter temperature of 4◦C depending on which sta-dium they reached at the spring equinox. This showsa complex interaction between photoperiod and wintertemperature. On the other hand, the lack of a hiberna-tion period, with increased winter temperature, hasalso to be considered.
Changes in the water regime, leading to longerperiods of summer drought and increase of the fre-quency of extreme events, are predicted to occur withglobal temperature increase. This would differentiallyaffect the resistance of the species with respect to thestadia which have to survive the dry period or the delayof some vital function. Some species develop protect-ive adaptations like cocoons or diapause but not all thespecies are similarly resistant and despite it mortalitywas reported when the adverse conditions are severe.
While optimal conditions vary according to spe-cies, life stadium and climate of origin, there is gen-erally a wide range of temperature or/and moisturewithin which animals are unconstrained. With respectto global change, the concepts of minimum and max-imum thresholds are more important in predicting theresponse of animals to change in temperature andmoisture regime. The expected temperature increaseimplies that low thresholds for temperature will becrossed earlier in the spring and later in the autumn,
whereas high thresholds will not be reached, and theactive period each year will be increased unless mois-ture decreases below a critical threshold. Therefore theresponse to global warming is closely dependent oninterannual variation (Coulson et al., 1996; Harte etal., 1996).
Consumption
Ongoing research showed that the faeces produc-tion is controlled by temperature and moisture. Theamount of faeces produced by the millipedeGlo-meris marginata, when fed on ofPinus halepensislitter, increased with temperature up to 23◦C, whereasat 30◦C the consumption was negligible after 171days in microcosms under constant moisture condi-tions (Figure 2a) (Aloui and Coûteaux, unpublisheddata). At 23◦C, the faeces production was higher in themicrocosms which were periodically gravimetricallyrewetted than in microcosms which were allowed todry out (Figure 2b).
Rates of decomposition on the soil surface havebeen found to be significantly lower than belowground in forest (McClaugherty et al., 1985; Moore,1984) and agricultural soils (Beare et al., 1992; Hol-land and Coleman, 1987). Earthworms bury vastquantities of organic material and the burial rate hasbeen shown to be temperature dependent (Edwardsand Lofty, 1977), therefore one might expect a sig-nificant effect on rates of decomposition based on thisinteraction. In addition, McInerney and Bolger (1999)have shown, in a microcosm experiment, that CO2production at 20◦C is approximately 40–50% greaterthan at 10◦C. But Q10 values for casts were lowerthan for control soils, suggesting that the cast ag-

130
gregate structure limited decomposition rates at hightemperatures.
Daniel (1991) found that food consumption in-creased with temperature up to 20◦C but declinedabove 22◦C. The consumption also declined whenthe soil water potential was below−7.0 kPa. Danielalso showed that high temperatures, low food avail-ability and low water potential, led to an increasein assimilation efficiency. At−7 kPa and 15◦C L.terrestris juveniles assimilated 55 and 43% of inges-ted dandelion leaves when 0.25 and 1.00 g of food(d.w.) were available per gram earthworm (d.w.) perweek, respectively. He speculated that coprophagy orprolonged gut passage time increased assimilation.
Spatial response to global change
For soil animals which have the ability to move, mi-gration could be an response to climate change. Thedynamics of the vertical distribution of animals in thesoil profile reflect this ability to migrate. For some spe-cies dryer conditions, induced by soil heating, leads toa vertical migration whereas others died in situ (Bri-ones et al., 1997). The downward migration ability ofsome species in response to soil drying or temperaturedropping was reported by Warburg et al. (1984) in areview study. Lateral migration is more rarely repor-ted. Dispersion of the isopod,Hemilepistus reaumuri,was shown to be as great as 500 m after 4–5 days.Buddelundiasp. andPorcellio sp. swarm. In suchswarms most animals are juveniles, with males com-prising only one-fifth of the population. The behaviourin these swarms was abnormal because the animalswere active during both night and day. Normally theseanimals are not active in the light (Warburg et al.,1984). This behaviour represents one of the criteriaof migration: movement as a priority activity. Thisability, even if it is not frequent, indicates that theseanimals could invade new space and settle in if the lifeconditions are favourable.
Diversity
Biodiversity is likely to be modified but the directionof this change will vary between the climatic zones. Inregions where temperature increase will create morefavourable conditions, such as longer summers withno moisture deficit, it might be expected that diversitywould increase by migration as suggested above and
through improved survivorship under milder condi-tions. However, in many areas, the increase of summertemperature will be associated with longer summerdrought. Species which have not evolved droughttolerances are likely to disappear with a consequen-tial decrease in biodiversity. The decline of diversitywould be more critical in epigeic communities thanin endogeic ones. Nevertheless, milder winter in areaswhere there is no frost or where frost will disappearmay compensate for the adverse summer conditions.
Biodiversity can affect average levels of ecosystemperformance, but the extent to which different speciesmake unique or redundant contributions to ecosys-tem processes remains controversial (Andrén et al.,1999; Ehrlich and Ehrlich, 1981; Johnson et al., 1996;Naeem et al., 1994, 1995; Schulze and Mooney, 1993).The question of how important species diversity isto ecosystem function boils down to consideration ofhow species in the system interact (Walker, 1992).Grime (1997) concluded, from a review of many stud-ies of the relationships between species richness andcommunity/ecosystem function, that the most imme-diate problem was to identify irreplaceable speciesand functional types and to discover whether there aresituations in which ecosystem variability depends onunusually high diversity. A certain number of rivetor driver species are clearly needed, even to main-tain landscape diversity, and ecologists have begun tolook at ways to identify those that are critical to eco-system functioning. It has been contended that mostecosystems exhibit levels of species richness higherthan that required for efficient biogeochemical andtrophic function (di Castri and Younès, 1990; Freck-man et al., 1997). This implies that within at leastsome functional types there are multiple species thatperform the same function and within any system therewill be: (i) functional diversity (the number of differ-ent functional pathways or types in a community); (ii)functional analogues (taxa belonging to the same func-tional type); and (iii) functional redundancy whichoccurs when there is more than one analogue in acommunity (Cowling et al., 1994).
The kernel of the functional redundancy hypo-thesis is the proposition that the contribution of aspecies must be very different from the contributionsof other species before its deletion or diminution willhave consequences for the habitual functioning of theecosystem. Despite the widespread appreciation of aninvolvement by the less conspicuous, though vastlynumerous, members of the biota in nutrient cyclingand litter decomposition within the soil (Verhoef and

131
Brussard, 1990), it is far from clear that the greatprofusion of species is entirely necessary for the main-tenance of such services. Although the functionaldiversity of soil organisms can be quite high, it isapparently exceeded to a large degree by the richnessof soil species. It is therefore sometimes inferred thatthe loss of species will not necessarily be reflected bya change in rates of biologically mediated processesand biochemical transformations. However, though re-dundancy in a single function may be common amongsoil biota, the suite of functions attributable to anyone species is unlikely to be redundant. Furthermore,functionally similar organisms often have differentenvironmental tolerances, physiological requirementsand microhabitat preferences. As such they are likelyto play quite different roles in the soil system (Beareet al., 1995).
Conclusions
General trends in soil fauna responses to changes inclimate are difficult to predict because of the paucityof data and because those that do exist are based onshort-term experiments in restricted areas and artificialconditions. Because of the limited quantity of materialthat can be produced under elevated CO2, the resultsrelated to litter quality are generally provided by ex-periments in the laboratory. Extrapolations to largertemporal and spatial scales must therefore be donewith caution because the soil fauna comprises a largerange of species in terms of size, life cycle, scale of ac-tion, adaptation or migration potential that is currentlynot very well known and because due to their intimateinteractions with vegetation and soil microflora, it de-pends to a large extent on the responses of these otherbiota to global change.
The main patterns of responses of soil fauna tomodifications of litter quantity and quality, and cli-mate change are most likely to be interactive ratherthan direct or unidirectional (Swift et al., 1998) andthe resulting effect would lead either to positive ornegative feedback depending on the dominant effecton the combined response. A positive feedback i.e. anincreased release of CO2 to the atmosphere could beexpected to arse from an increase of activity (espe-cially in the winter) and fecundity as a consequenceof temperature increase, and an increase in consump-tion due to both temperature increase and change oflitter quality. A negative feedback, i.e. a sink of at-mospheric CO2 can be expected due to, for example,
summer mortality or a reduction in life expectancy.Other expected responses like downward and hori-zontal migration, use of protective features such asencystment, diapause, cocoon formation, favouringadapted species would change the species compositionand the decomposer community might shift to organ-isms with different climatic sensitivity, but because ofthe apparently high functional redundancy in speciesrichness, resilience in functional capacity might not beaffected unless keystone species are in danger. How-ever the keystone soil animals are not clearly identifiedbecause the role of animals at the ecosystem level isindirect, controlling microbial functions, rather thandirect and disappearance of a soil species would affectthe intensity of a function but not the function itself.
Most of the aspects of the consequence of globalchange on soil fauna are poorly explored under ex-perimental conditions and there is a large range ofpriorities to develop in order to overcome the currentwide range of speculation. There is an urgent need tofocus on interactions between soil fauna and the othersoil biota which have a key role in the maintenanceof the ecosystems and are expected to be modifiedby global changes, e.g., faunal grazing on roots andmycorrhizae. Moreover, a better knowledge of plantresidue consumption and physiological responses ofsoil fauna to abiotic factors, with respect to criticalstadia and life history trait, would throw light on theirspecific responses to global change.
References
Andrén O, Brussaard L and Clarholm M 1999 Soil organism in-fluence on ecosystem-level processes – bypassing the ecologicalhierarchy. Appl. Soil Ecol. 11, 177–188.
Arpin P, Ponge JF and Vannier G 1995 Experimental modificationsof litter supplies in a forest mull and reaction of the nematodefauna. Fundam. Appl. Nematol. 18, 371–389.
Beare HM, Parmelee WR, Hendrix FP, Cheng W, Coleman DCand Crossley DA Jr 1992 Microbial and faunal interactions andeffects on litter nitrogen and decomposition in agroecosystems.Ecol. Monogr. 62, 569–591.
Beare MH, Coleman DC, Crossley DA Jr, Hendrix PF and OdumEP 1995 A hierarchical approach to evaluating the significanceof soil biodiversity to biogeochemical cycling. Plant Soil 170,5–22.
Bengtsson J, Lundqvist H, Saetre P, Sohlenius B and SolbreckB 1998 Effects of organic matter removal on soil food web:Forestry practices meet ecological theory. Appl. Soil Ecol. 9,137–143.
Berg MP, Verhoef HA, Anderson JM, Beese F, Bolger T, CoûteauxMM, Ineson P, McCarthy F, Palka L, Raubuch M, Splatt P andWillison T 1997 Effects of air pollutant–temperature interactionson mineral-N dynamics and cation leaching in replicate forestsoil transplantation experiment. Biogeochemistry. 39, 295–326.

132
Booth RG and Anderson JM 1979 The influence of fungal foodquality on the growth and fecundity ofFolsomia candida(Colle-mbola: Isotomidae). Oecologia 38, 317–323.
Bottner P, Coûteaux MM, Anderson JA, Berg B, Billès G, BolgerT, Casabianca H, Romanyà J and Rovira P 1999 Decompositionof 13C and15N labelled plant material in a 65–40◦ latitudinaltransect of West European coniferous forest soils: simulation ofclimate change by translocation of soils. Soil Biol. Biochem. 31(in press).
Briones MJI, Ineson P and Piearce TG 1997 Effects of climatechange on soil fauna; responses of enchytraeids, Diptera larvaeand tardigrades in a transplant experiment. Appl. Soil Ecol. 6,117–134.
Butt KR, Frederickson J and Morris RM 1992 The intensive produc-tion of Lumbricus terrestrisL. for soil amelioration. Soil Biol.Biochem. 24, 1321–1325.
Cotrufo MF, Berg B and Kratz W 1998a Increased atmospheric CO2and litter quality. Environ. Rev. 6, 1–12.
Cotrufo MF, Briones MJI and Ineson P 1998b Elevated CO2 affectsfield decomposition rate and palatability of tree leaf litter: im-portance of changes in substrate quality. Soil Biol. Biochem. 30,1565–1571.
Cotrufo MF, Ineson P and Scott A 1998c Elevated CO2 reduces thenitrogen concentration of plant tissues. Glob. Change Biol. 4, 43–54.
Coulson SJ, Hoskinson ID, Block W, Webb NR and Worland MR1995 Low summer temperature: a potential mortality factor forhigh arctic soil microarthropods. J. Insect Physiol. 9, 783–792.
Coulson SJ, Hodkinson ID, Webb NR, Block W, Bale JS, Strath-dee AT, Worland MR and Wooley C 1996 Effects of experi-mental temperature elevation on high-arctic soil microarthopodspopulations. Polar Biol. 16, 147–153.
Coûteaux MM and Bottner P 1994 Biological interactions betweenfauna and the microbial community.In Beyond the Biomass:Compositional and Functional Analysis of Soil Microbial Com-munities. Eds K Ritz, J Dighton and K E Giller. pp 159–172.John Wiley, Chichester.
Coûteaux MM, Mousseau M, Célérier M L and Bottner P 1991Increased atmospheric CO2 and litter quality: decompositionof sweet chestnut leaf litter with animal food web of differentcomplexity. Oikos 61, 54–64.
Coûteaux MM, Bottner P and Berg B 1995 Litter decomposition,climate and litter quality. Trends Ecol. Evol. 10, 63–66.
Coûteaux MM, Kurz C, Bottner P and Raschi A 1999 Influence ofthe increased atmospheric CO2 concentration on quality of plantmaterial and litter decomposition. Tree Physiol. 19, 301–311.
Cowling RM, Mustart PJ, Laurie H and Richards MB 1994 Spe-cies diversity; functional diversity and functional redundancy infynbos communities. S. Afr. J. Sci. 90, 333–337.
Cromack K Jr, Fichter BL, Moldenke AM, Entry J A and InghamER 1988 Interactions between soil animals and ectomycorrhizalfungal mats.In Biological interactions in soil. Eds CA Ed-wards, BR Stinner, D Stinner and S Rabatin. pp 161–168. Agric.Ecosystems Environ. 24, Elsevier, Amsterdam.
Daniel O 1991 Leaf-litter consumption and assimilation by ju-veniles ofLumbricus terrestrisL. (Oligochaeta, Lumbricidae)under different environmental conditions. Biol. Fertil. Soils 12,202–208.
David JF, Ponge JF, Arpin P and Vannier G 1991 Reactions of themacrofauna of a forest mull to experimental perturbations oflitter supply. Oikos 61, 316–326.
David JF, Célérier ML and Geoffroy JJ 1999 Periods of dormancyand cohort-splitting in the millipedePolydesmus angustus(Dip-lopoda, Polydesmidae). Eur. J. Entomol. 96, (in press).
Dhillion SS, Roy J and Abrams M 1996 Assessing the impact ofelevated CO2 on soil microbial activity in a mediterranean modelecosystem. Plant Soil 187, 333–342.
di Castri F and Younès T 1990 Ecosystem function of biologicaldiversity. Biol. Int. 22, 20.
Douglas JT, Goss MJ and Hill D 1989 Measurements of pore char-acteristics in a clay soil under ploughed and drilling. Soil Till.Res. 1, 11–18.
Dury SJ, Good JEG, Perrins CM, Buse A and Kaye T 1998 The ef-fects of increasing CO2 and temperature on oak leaf palatabilityand the implications for herbivorous insects. Glob. Change Biol.4, 55–61.
Edwards CA and Lofty JR 1977 Biology of Earthworms, 2nd ed.Chapman and Hall, London.
Ehrlich PR and Ehrlich AH 1981 Extinction. The Causes and Con-sequences of the Disappearance of Species. pp xiv, 306. RandomHouse, New York.
El Kohen A, Rouhier H and Mousseau M 1992 Changes in dryweight and nitrogen partitioning induced by elevated CO2 de-pend on soil nutrient availability in sweet chestnut (Castaneasativa Mill). Ann. Sci. For. 49, 83–90.
Evans AC and Guild WJ Mc L 1948 Studies of the relationshipsbetween earthworms and soil fertility. IV On the life cycles ofsome British Lunbricidae. Ann. Appl. Biol. 35, 471–484.
Fajer ER, Bowers ED and Bazzaz FA 1991 The effects of enrichedCO2 atmospheres on the buckeye butterfly,Junonia coenia.Ecology 72, 751–754.
Freckman DW, Moore JC, Hunt HW and Elliott ET 1991 Theeffects of elevated CO2 and climate change on soil nematodecommunity structure of prairie sod.. Bull. Ecol. Soc. Am. 72-2,119.
Freckman DW, Blackburn TW, Brussaard L, Hutchings P, PalmerMA and Snelgrove PVR 1997 Linking biodiversity and ecosys-tem functioning of soils and sediments. Ambio 26, 556–562.
Grime JP 1997 Biodiversity and ecosystem function: the debatedeepens. Science 277, 1260–1261.
Hanlon RDG 1981 Influence of grazing by Collembola on the activ-ity of senescent fungal colonies grown on media of differentnutrient concentration. Oikos 36, 362–367.
Hanlon RDG and Anderson JM 1979 The effects of collembolagrazing on microbial activity in decomposing leaf litter. Oeco-logia 38, 93–99.
Hanlon RDG and Anderson JM 1980 Influence of macroarthopodfeeding activities on microflora in decomposing oak leaves. SoilBiol. Biochem. 12, 255–261.
Harte J, Rawa A and Price V 1996 Effects of manipulated soilmicroclimate on mesofaunal biomass and diversity. Soil Biol.Biochem. 28, 313–322.
Harte J, Torn MS, Chang F, Feifarek B, Kinzig A P, Shaw R andShen K 1995 Global warming and soil microclimate: results froma meadow-warming experiment. Ecol. Appl. 5, 132–150.
Hättenschwiler S, Bühler S and Körner C 1999 Quality, decom-position and isopod consumption of tree litter produced underelevated CO2. Oikos 85, 271–281.
Hendriksen, NB 1990 Leaf litter selection by detritivore and geo-phagous earthworms. Biol. Fertil. Soils 10, 17–21.
Hoeksema KJ and Jongrius A 1959 On the influence of earthwormson the soil structure in mulched orchards. Proc. Int. Symp. SoilStruct. 188–194.
Holland EA and Coleman DC 1987 Litter placement effects onmicrobial and organic matter dynamics in an agroecosystem.Ecology 8, 425–433.
Huish S, Leonard MA and Anderson JM 1985 Wetting and dry-ing effects on animal/microbial mediated nitrogen mineralization

133
and mineral element losses from deciduous forest litter and rawhumus. Pedobiologia 28, 177–183.
Johnson KH, Vogt KA, Clarke HJ, Schmitz OJ and Vogt D J 1996Biodiversity and the production and stability of ecosystems.Trends Ecol. Evol. 11, 372–377.
Judas M 1990 The development of earthworm populations follow-ing manipulation of the canopy leaf litter in a beechwood onlimestone. Pedobiologia 34, 247–255.
Kretzschmar A 1978 Quantification écologique des galeries deLombriciens. Techniques et premières estimations. Pedobiologia18, 31–38.
Lavelle P 1988 Earthworm activities and the soil system. Biol.Fertil. Soils 6, 237–251.
Lee KE 1985 Earthworms; their Ecology and Relationships withSoils and Land Use. Academic Press, London.
Lincoln DE, Fajer ER and Johnson RH 1993 Plant-insect herbivoreinteractions in elevated CO2 environments. Trends Ecol. Evol. 8,64–68.
Lindroth RL, Kinney KK and Platz CL 1993 Responses of decidu-ous trees to elevated atmospheric CO2: productivity, and insectperformance. Ecology 74, 763–777.
McClaugherty CA, Pastor J, Aber JD and Melillo JM 1985 Forestlitter decomposition in relation to soil nitrogen dynamics andlitter quality. Ecology 66, 266–275.
McInerney M and Bolger T 1999 Effects of temperature, wet-ting/drying cycles, and soil texture on carbon and nitrogendynamics in stabilized earthworm casts derived from two forestsoils. Soil Biol. Biochem. 31 (in press).
Moore TR 1984 Litter decomposition in a subarctic spruce-lichenwoodland, Eastern Canada. Ecology 65, 299–308.
Mousseau M and Enoch HZ 1989 Effect of doubling atmosphericCO2 concentration on growth, dry matter distribution and CO2exchange of 2 yr old sweet chestnut trees (Castanea sativaMill.).Ann. Sci. For. 46, 506s–508s.
Naeem S, Thompson LJ, Lawler SP, Lawton JH and WoodfinRM 1994 Declining biodiversity can alter the performance ofecosystems. Nature 368, 734–737.
Naeem S, Thompson LJ, Lawler SP, Lawton JH and Woodfin RM1995 Empirical evidence that declining species diversity may al-ter the performance of terrestrial ecosystems. Phil. Trans. R. Soc.London 347, 249–262.
Newton PCD, Clark H, Bell CC, Glasgow EM, Tate KR, Ross DJ,Yeates GW and Saggar S 1995 Plant growth and soil processesin temperate grassland communities at elevated CO2. J. Biogeog.22, 235–240.
Niklaus PA and Körner C 1996 Responses of soil microbiota of alate successional alpine grassland to long term CO2 enrichment.Plant Soil 184, 219–229.
O’Neill EG 1994 Responses of soil biota to elevated atmosphericcarbon dioxide. Plant Soil 165, 55–65.
O’Neill EG, Luxmoore RJ and Norby R J 1987 Increases in mycor-rhizal colonization and seedling inPinus echinataandQuercusalba in an enriched CO2 atmosphere. Can. J. For. Res. 17,878–883.
Peterjohn WT, Melillo JM, Bowles FP and Steudler PA 1993Soil warming and trace gas fluxes: experimental design andpreliminary flux results. Oecologia 93, 18–24.
Peterjohn WT, Mellilo JM, Steudler PA and Newrick KM 1994 Re-sponses of trace gas fluxes and N availability to experimentallyelevated soil temperature. Ecol. Appl. 4, 617–625.
Ponge JF, Arpin P and Vannier G 1993 Collembolan response toexperimental perturbations of litter supply in a temperate forestecosystem. Eur. J. Soil Biol. 29, 141–153.
Prior SA, Rogers HH, Runion GB and Mauney JR 1994 Effectsof free-air CO2 enrichment on cotton root growth. Agr. For.Meteorol. 70, 69–86.
Rabatin SC and Stinner BR 1985 Arthropods as consumers ofvesicular-arbuscular mycorrhizal fungi. Mycologia 77, 320–322.
Reddel P and Spain AV 1991 Earthworms as vectors of viable pro-pagules of mycorrhizal fungi. Soil Biol. Biochem. 23, 767–774.
Roth S, Lindroth RL, Volin JC and Kruger EL 1998 Enriched atmo-spheric CO2 and defoliation: effects on tree chemistry and insectperformance. Glob. Change Biol. 4, 419–430.
Rouhier H, Billès G, El Kohen A, Mousseau M and Bottner P1994 Effect of elevated CO2 on carbon and nitrogen distributionwithin a tree-soil system; 14C studies on sweet chestnut trees(Castanea sativaMill.). Plant Soil 162, 281–292.
Runion GB, Curl EA, Rogers HH, Backman PA, Rodriguez-KabanaR and Helms BE 1994 Effects of free-air CO2 enrichment onmicrobial populations in the rhizosphere and phyllosphere ofcotton. Agric. For. Meteor. 70, 117–130.
Rushton SP and Hassall M 1983 Food and feeding rates of the ter-restrial isopodArmadillidium vulgare(Latreille). Oecologia 57,415–419.
Scheu S and Wolters V 1991 Influence of fragmentation and bioturb-ation on the decomposition of14C-labelled beech leaf litter. SoilBiol. Biochem. 23, 1029–1034.
Schulze E-D and Mooney HA 1993 Biodiversity and EcosystemFunction. Springer-Verlag, Berlin.
Seastedt T R 1984 The role of microarthropods in decompositionand mineralization processes. Annu. Rev. Entomol. 29, 25–46.
Setälä H and Huhta V 1990 Evaluation of the soil fauna impact ondecomposition in a simulated coniferous forest soil. Biol. Fertil.Soils 10, 163–169.
Setälä H, Haimi J and Huhta V 1988 A microcosm study on therespiration and weight loss in birch litter and raw humus asinfluenced by soil fauna. Biol. Fertil. Soils 5, 282–287.
Setälä H, Rissanen J and Markkola AM 1997 Conditional outcomesin the relationship between pine and ectomycorrhizal fungi inrelation to biotic and abiotic environments. Oikos 80, 112–122.
Snider RJ 1971 Dietary influence on the growth and fecundity ofOnychiurus justii (Denis), (Onychiuridae, Collembola).In Or-ganismes du Sol et production primaire. Proc. 4th Coll. SoilZool., pp 225-234. I.N.R.A. Dijon.
Snider RM 1981a Growth and survival ofPolydesmus inconstans(Diplopoda: Polydesmidae) at constant temperautre. Pedobiolo-gia 22, 345–353.
Snider RM 1981b The reproductive biology ofPolydesmus in-constans(Diplopoda: Polydesmidae) at constant temperature.Pedobiologia 22, 354–365.
Stockdill SMJ 1966 The effect of earthworms on pastures. Proc.N.Z. Ecol. Soc. 13, 68–75.
Sulkava P, Huhta V and Laakso J 1996 Impact of soil faunalstructure on decomposition and N-mineralisation in relationto temperature and moisture in forests soil. Pedobiologia 40,505–513.
Sutton SL, Hassall M, Willows R, Davis RC, Grundy A and Sun-derland KD 1984 Life histories of terrestrial Isopods: A study ofintra- and interspecific variations. Symp. Zool. Soc. London. 53,269–294.
Swift MJ, Andrén O, Brussaard L, Briones M, Coûteaux MM, Ek-schmitt K, Kjøller A, Loiseau P and Smith P 1998 Global change,soil biodiversity, and nitrogen cycling in terrestrial ecosystems:three case studies. Glob. Change Biol. 4, 729–743.
Tate KR and Ross DJ 1997 Elevated CO2 and moisture effects onsoil carbon storage and cycling in temperate grasslands. Glob.Change Biol. 3, 225–235.

134
Tian G, Brussaard L, Kang BT and Swift MJ 1997 Soil fauna-mediated decomposition of plant residues under constrained en-vironmental and residue quality conditions.In Driven by Nature,Plant litter quality and decomposition, Eds G Cadish, KE Giller.pp 125–144. Cab International, Wallingford, Oxon.
Tsukamoto J and Watanabe H 1977 Influence of temperature onhatching and growth ofEisenia foetida(Oligochaeta, Lumbri-cidae). Pedobiologia 17, 338–342.
van Veen JA, Liljeroth E and Lekkerkerk LJA 1991 Carbon fluxesin plant-soil systems at elevated atmospheric CO2 levels. Ecol.Appl. 1, 175–181.
Verhoef HA and Brussaard L 1990 Decomposition and nitrogenmineralization in natural and agroecosystems: the contributionof soil animals. Biogeochemistry 11, 175–211.
Walker B 1992 Biodiversity and ecological redundancy. Conserv.Biol. 6, 18–23.
Warburg MR, Linsenmair KE and Bercovitz K 1984 The effect of
climate on the distribution and abundance of isopods. Symp.Zool. Soc. London 53, 339–367.
Warnock AJ, Fitter AH and Usher MB 1982 The influence ofthe springtailFolsomia candidaon the mycorrhizal associationof leek Allium porumand the vesicular arbuscular mycorrhizalendophyteGlomus fasciculatus. New Phytol. 90, 285–292.
Wood CW, Torbert HA, Rogers HH, Runion GB and Prior SA 1994Free-air CO2 enrichment effects on soil carbon and nitrogen.Agric. For. Meteor. 70, 103–116.
Zak DR, Ringelberg DB, Pregitzer KS, Randlett DL, White DCand Curtis PS 1996 Soil microbial communities beneathPopu-lus grandidentatagrown under elevated atmospheric CO2. Ecol.Appl. 6, 257–262.
Zaller JG and Arnone JA 1997 Activity of surface-casting earth-worms in a calcareous grassland under elevated atmosphericCO2. Oecologia 111, 249–254.




















