
Insulin Regulation of Cholesterol 7α-Hydroxylase Expression in Human Hepatocytes ROLES OF FORKHEAD...
10
Insulin Regulation of Cholesterol 7-Hydroxylase Expression in Human Hepatocytes ROLES OF FORKHEAD BOX O1 AND STEROL REGULATORY ELEMENT-BINDING PROTEIN 1c * □ S Received for publication, June 16, 2006, and in revised form, August 1, 2006 Published, JBC Papers in Press, August 2, 2006, DOI 10.1074/jbc.M605815200 Tiangang Li ‡ , Xiaoying Kong ‡ , Erika Owsley ‡ , Ewa Ellis § , Stephen Strom § , and John Y. L. Chiang ‡1 From the ‡ Department of Microbiology, Immunology and Biochemistry, Northeastern Ohio University College of Medicine, Rootstown, Ohio 44272 and § Department of Pathology, University of Pittsburgh Medical Center, Pittsburgh, Pennsylvania 15213 Bile acid synthesis and pool size increases in diabetes, whereas insulin inhibits bile acid synthesis. The objective of this study is to elucidate the mechanism of insulin regulation of cholesterol 7- hydroxylase gene expression in human hepatocytes. Real-time PCR assays showed that physiological concentrations of insulin rapidly stimulated cholesterol 7-hydroxylase (CYP7A1) mRNA expression in primary human hepatocytes but inhibited CYP7A1 expression after extended treatment. The insulin-reg- ulated forkhead box O1 (FoxO1) and steroid regulatory ele- ment-binding protein-1c (SREBP-1c) strongly inhibited hepa- tocyte nuclear factor 4 and peroxisome proliferator-activated receptor coactivator-1 trans-activation of the CYP7A1 gene. FoxO1 binds to an insulin response element in the rat CYP7A1 promoter, which is not present in the human CYP7A1 gene. Insulin rapidly phosphorylates and inactivates FoxO1, whereas insulin induces nuclear SREBP-1c expression in human primary hepatocytes. Chromatin immunoprecipita- tion assay shows that insulin reduced FoxO1 and peroxisome proliferators- activated receptor -coactivator-1 but increased SREBP-1c recruitment to CYP7A1 chromatin. We conclude that insulin has dual effects on human CYP7A1 gene transcription; physiological concentrations of insulin rapidly inhibit FoxO1 activity leading to stimulation of the human CYP7A1 gene, whereas prolonged insulin treatment induces SREBP-1c, which inhibits human CYP7A1 gene transcrip- tion. Insulin may play a major role in the regulation of bile acid synthesis and dyslipidemia in diabetes. The liver plays a central role in lipid metabolism and main- taining whole body lipid homeostasis, which is dysregulated in metabolic syndrome (syndrome X), obesity, and diabetes (1). Bile acid synthesis in the liver is the predominant pathway for cholesterol catabolism and is regulated by cholesterol 7-hy- droxylase (CYP7A1) 2 (2). Bile acids are physiological agents that facilitate biliary cholesterol excretion, intestinal absorp- tion of nutrients, and disposal of toxic metabolites. Bile acids are also signaling molecules that activate bile acid receptors to regulate bile acid synthesis and glucose metabolism (2). In vivo studies show that bile acid pool and excretion increase in dia- betic human patients (3, 4) and in experimental diabetic ani- mals (5, 6). Insulin treatment restores bile acid pool and synthe- sis to the normal levels. It has been reported that physiological concentrations of insulin (1.4 –14 nM) inhibit bile acid synthesis by down-regulation of CYP7A1 (7–9), sterol 12-hydroxylase (CYP8B1) (10), and sterol 27-hydroxylase (CYP27A1) gene transcription (9). We have previously reported that insulin plays a dominant role in the inhibition of CYP7A1 gene tran- scription (7, 8). How insulin regulates human CYP7A1 and what factors mediate the insulin effects remain unknown. Insulin is known to induce more than 100 genes but inhibit only a few hepatic genes involved in glucose metabolism (11). The consensus sequence of negative insulin response element (IRE), T(G/A)TTT(T/G)(G/T), has been identified in promot- ers of phosphoenolpyruvate carboxykinase (PEPCK), glucose- 6-phosphatase, insulin-like growth factor binding protein-1, tyrosine aminotransferase, and apolipoprotein CIII genes (12). These IREs are known to bind hepatocyte nuclear factor 3 (HNF3 or FoxA1), but HNF3 does not mediate insulin action (12). Recently forkhead box O1 (FoxO1), a mammalian homo- log of Caenorhabditis elegans DAF-16 has been identified as an insulin-regulated transcription factor that plays a critical role in mediating insulin inhibition of the PEPCK, glucose-6-phospha- tase, insulin-like growth factor binding protein-1, and apoli- poprotein CIII genes (13–17). Akt/protein kinase B, a down- stream serine kinase of the insulin-signaling pathway, phosphorylates FoxO1 (18). The phosphorylated FoxO1 is excluded from the nucleus and degraded by the ubiquitin pro- teasome pathway, which results in inhibition of FoxO1 target genes (19, 20). FoxO1 exerts a positive effect by binding to its target gene promoters (15). FoxO1 also regulates genes as a * This study was supported by NIDDK, National Institutes of Health Grants DK 58379 and DK44442 (to J. Y. L. C.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental data. 1 To whom correspondence should be addressed: Dept. of Microbiology, Immunology and Biochemistry, Northeastern Ohio University College of Medicine, 4209 State Route 44, Rootstown, OH 44272. Tel.: 330-325-6694; Fax: 330-325-5910; E-mail: [email protected]. 2 The abbreviations used are: CYP7A1, cholesterol 7-hydroxylase; ChIP, chromatin immunoprecipitation; FoxO1, forkhead box O1; HNF4, hepa- tocyte nuclear factor 4; IRE, insulin response element; rIRE, rat IRE; PEPCK, phosphoenolpyruvate carboxykinase; PGC-1, peroxisome proliferators- activated receptor -coactivator-1; SRE, sterol response element; SREBP, SRE-binding protein; LXR, liver orphan receptor ; EMSA, electrophoretic mobility shift assay; HA, hemagglutinin; Luc, luciferase. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 39, pp. 28745–28754, September 29, 2006 © 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. SEPTEMBER 29, 2006 • VOLUME 281 • NUMBER 39 JOURNAL OF BIOLOGICAL CHEMISTRY 28745
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Insulin Regulation of Cholesterol 7α-Hydroxylase Expression in Human Hepatocytes ROLES OF FORKHEAD...

Insulin Regulation of Cholesterol 7�-Hydroxylase Expressionin Human HepatocytesROLES OF FORKHEAD BOX O1 AND STEROL REGULATORY ELEMENT-BINDINGPROTEIN 1c*□S
Received for publication, June 16, 2006, and in revised form, August 1, 2006 Published, JBC Papers in Press, August 2, 2006, DOI 10.1074/jbc.M605815200
Tiangang Li‡, Xiaoying Kong‡, Erika Owsley‡, Ewa Ellis§, Stephen Strom§, and John Y. L. Chiang‡1
From the ‡Department of Microbiology, Immunology and Biochemistry, Northeastern Ohio University College of Medicine,Rootstown, Ohio 44272 and §Department of Pathology, University of Pittsburgh Medical Center, Pittsburgh, Pennsylvania 15213
Bile acid synthesis andpool size increases indiabetes,whereasinsulin inhibits bile acid synthesis. The objective of this study isto elucidate themechanismof insulin regulationof cholesterol 7�-hydroxylase gene expression in human hepatocytes. Real-timePCR assays showed that physiological concentrations of insulinrapidly stimulated cholesterol 7�-hydroxylase (CYP7A1) mRNAexpression in primary human hepatocytes but inhibitedCYP7A1 expression after extended treatment. The insulin-reg-ulated forkhead box O1 (FoxO1) and steroid regulatory ele-ment-binding protein-1c (SREBP-1c) strongly inhibited hepa-tocyte nuclear factor 4� and peroxisome proliferator-activatedreceptor � coactivator-1� trans-activation of the CYP7A1 gene.FoxO1 binds to an insulin response element in the rat CYP7A1promoter, which is not present in the human CYP7A1 gene.Insulin rapidly phosphorylates and inactivates FoxO1,whereas insulin induces nuclear SREBP-1c expression inhuman primary hepatocytes. Chromatin immunoprecipita-tion assay shows that insulin reduced FoxO1 and peroxisomeproliferators- activated receptor �-coactivator-1� butincreased SREBP-1c recruitment to CYP7A1 chromatin. Weconclude that insulin has dual effects on humanCYP7A1 genetranscription; physiological concentrations of insulin rapidlyinhibit FoxO1 activity leading to stimulation of the humanCYP7A1 gene, whereas prolonged insulin treatment inducesSREBP-1c, which inhibits human CYP7A1 gene transcrip-tion. Insulin may play a major role in the regulation of bileacid synthesis and dyslipidemia in diabetes.
The liver plays a central role in lipid metabolism and main-taining whole body lipid homeostasis, which is dysregulated inmetabolic syndrome (syndrome X), obesity, and diabetes (1).Bile acid synthesis in the liver is the predominant pathway forcholesterol catabolism and is regulated by cholesterol 7�-hy-
droxylase (CYP7A1)2 (2). Bile acids are physiological agentsthat facilitate biliary cholesterol excretion, intestinal absorp-tion of nutrients, and disposal of toxic metabolites. Bile acidsare also signaling molecules that activate bile acid receptors toregulate bile acid synthesis and glucose metabolism (2). In vivostudies show that bile acid pool and excretion increase in dia-betic human patients (3, 4) and in experimental diabetic ani-mals (5, 6). Insulin treatment restores bile acid pool and synthe-sis to the normal levels. It has been reported that physiologicalconcentrations of insulin (1.4–14 nM) inhibit bile acid synthesisby down-regulation of CYP7A1 (7–9), sterol 12�-hydroxylase(CYP8B1) (10), and sterol 27-hydroxylase (CYP27A1) genetranscription (9). We have previously reported that insulinplays a dominant role in the inhibition of CYP7A1 gene tran-scription (7, 8). How insulin regulates human CYP7A1 andwhat factors mediate the insulin effects remain unknown.Insulin is known to induce more than 100 genes but inhibit
only a few hepatic genes involved in glucose metabolism (11).The consensus sequence of negative insulin response element(IRE), T(G/A)TTT(T/G)(G/T), has been identified in promot-ers of phosphoenolpyruvate carboxykinase (PEPCK), glucose-6-phosphatase, insulin-like growth factor binding protein-1,tyrosine aminotransferase, and apolipoprotein CIII genes (12).These IREs are known to bind hepatocyte nuclear factor 3�(HNF3� or FoxA1), butHNF3�does notmediate insulin action(12). Recently forkhead box O1 (FoxO1), a mammalian homo-log of Caenorhabditis elegansDAF-16 has been identified as aninsulin-regulated transcription factor that plays a critical role inmediating insulin inhibition of the PEPCK, glucose-6-phospha-tase, insulin-like growth factor binding protein-1, and apoli-poprotein CIII genes (13–17). Akt/protein kinase B, a down-stream serine kinase of the insulin-signaling pathway,phosphorylates FoxO1 (18). The phosphorylated FoxO1 isexcluded from the nucleus and degraded by the ubiquitin pro-teasome pathway, which results in inhibition of FoxO1 targetgenes (19, 20). FoxO1 exerts a positive effect by binding to itstarget gene promoters (15). FoxO1 also regulates genes as a* This study was supported by NIDDK, National Institutes of Health Grants DK
58379 and DK44442 (to J. Y. L. C.). The costs of publication of this articlewere defrayed in part by the payment of page charges. This article musttherefore be hereby marked “advertisement” in accordance with 18 U.S.C.Section 1734 solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental data.
1 To whom correspondence should be addressed: Dept. of Microbiology,Immunology and Biochemistry, Northeastern Ohio University College ofMedicine, 4209 State Route 44, Rootstown, OH 44272. Tel.: 330-325-6694;Fax: 330-325-5910; E-mail: [email protected].
2 The abbreviations used are: CYP7A1, cholesterol 7�-hydroxylase; ChIP,chromatin immunoprecipitation; FoxO1, forkhead box O1; HNF4�, hepa-tocyte nuclear factor 4�; IRE, insulin response element; rIRE, rat IRE; PEPCK,phosphoenolpyruvate carboxykinase; PGC-1�, peroxisome proliferators-activated receptor �-coactivator-1�; SRE, sterol response element; SREBP,SRE-binding protein; LXR�, liver orphan receptor �; EMSA, electrophoreticmobility shift assay; HA, hemagglutinin; Luc, luciferase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 39, pp. 28745–28754, September 29, 2006© 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
SEPTEMBER 29, 2006 • VOLUME 281 • NUMBER 39 JOURNAL OF BIOLOGICAL CHEMISTRY 28745

DNA binding-independent co-activator or co-repressor ofmany transcription factors (21). Dysregulation of FoxO1expression and function leads to hyperglycemia and hypertrig-lyceridemia in the mouse models of type I and type II diabetes(17).Insulin stimulates the activity of sterol regulatory element-
binding proteins (SREBPs), which are key regulators of lipidhomeostasis and insulin actions (22). The predominant SREBPisoform, SREBP-1c in liver and adipose tissues, preferentiallyinduces genes in fatty acid and triglyceride synthesis (23). ASREBPprecursor (125 kDa) forms a complexwith SREBP cleav-age-activating protein in the endoplasmic reticulum. Insulininducing gene-1 and -2a (Insig) anchor the SREBP�SREBPcleavage-activating protein complex to the endoplasmic retic-ulummembrane (24).When cellular sterol levels are low, Insigsrelease the SREBP cleavage-activating protein (SCAP)�SREBPcomplex and allow SCAP to escort SREBPs to the Golgi appa-ratus. Two sterol-regulated proteases act sequentially to releasethe N-terminal basic-helix-loop-helix leucine zipper domain.The mature SREBP (68 kDa) enters the nucleus, binds to thesterol regulatory element (SRE), and induces target gene tran-scription. Insulin stimulates liver orphan receptor � (LXR�)which induces SREBP-1c gene expression (25, 26). Insulin alsopromotes SREBP-1c cleavage and increases insulin-stimulatedlipogenesis by inhibition of Insig-2a expression in livers (11,24). Studies have shown that insulin-dependent proteolyticcleavage is crucial and sufficient for SREBP-1c activation (27).In this study we investigated the insulin regulation of
CYP7A1 gene transcription in human hepatocytes. Resultsindicate that short-term treatment of physiological concentra-tions of insulin rapidly induces, whereas prolonged treatmentof insulin represses human CYP7A1 gene expression. This dualeffect of insulin actionmay bemediated by two insulin sensitivetranscription factors, FoxO1 and SREBP-1c. This study pro-vides new insights into how bile acid synthesis is regulated byinsulin under normal physiological conditions and how dys-regulation of bile acid homeostasis may contribute to glucoseand lipid abnormalities in diabetes.
EXPERIMENTAL PROCEDURES
Cell Culture—The human hepatoblastoma cell line, HepG2,was purchased from American Type Culture Collection (Man-assas, VA). The cells were cultured in Dulbecco’s modifiedEagle’s medium and F-12 (Sigma) supplemented with 100units/ml penicillin G/streptomycin sulfate (Mediatech, Hern-don, VA) and 10% (v/v) heat inactivated fetal bovine serum(Irvine Scientific, Santa Ana, CA). Primary human hepato-cytes HH1247 (3 y, M), HH1248 (42 y, F), HH1249 (59 y, M),HH1251 (29 y, M), HH1274 (55 y, F), HH1281 (16 y, M),HH1286 (50 y, M), and HH1308 (64 y, M) were isolated fromhuman donors and were obtained through the Liver TissueProcurement and Distribution System of the National Insti-tutes of Health. For the first 36 h after isolation, cells weremaintained in hepatocyte maintenance medium supple-mented with 10�7 M insulin and dexamethasone (Cambrex,NJ).Reporters and Expression Plasmids—Rat and human
CYP7A1/luciferase (Luc) reporter were constructed as previ-
ously described (Crestani (44); Wang et al. (76)). Expressionplasmid for human peroxisome proliferator-activated receptor� coactivator-1� (PGC-1�) (pcDNA3/HA-PGC-1�) wasobtained from A. Kralli (The Scripps Research Institute, LaJolla, CA). Expression plasmid for pCMV-rHNF3� was kindlyprovided by Robert H. Costa (University of Illinois, Chicago,IL). pCMV5-FoxO1 and pBluescript-FoxO1 were kindly pro-vided by D. Accili (Columbia University, New York). SREBP-1cexpression plasmid (pCMV-SREBP-1c-436) was purchasedfrom the ATCC. pCMX-hLXR� and pCMX-RXR� were pro-vided by D. Mangelsdorf (University of Texas Southwestern)and R. Evans (The Salk Institute for Biological Studies, La Jolla,CA), respectively. The construction of the expression plasmidfor HNF4� (pCMV-HNF4�) was previously described (8). The�-galactosidase expression plasmid (pCMV-�-gal) and themammalian expression vector pcDNA3 were obtained fromClontech (Palo Alto, CA). The reporter vector (pGL3-Basic)was purchased from Promega (Madison, WI). The PEPCKreporter plasmid, p2000Luc, was obtained from R. Hanson(Case Western Reserve University). The synthetic reporter5XUAS-TK-Luc, which contains 5 copies of Gal4 binding siteUAS located upstream of the thymidine kinase promoter andluciferase gene, was provided by A. Takeshita (ToranomonHospital, Tokyo, Japan) (28). The Gal4-HNF4� fusion plasmidpBx-HNF4-LBD was obtained from I. Talianidis (Institute ofMolecular Biology and Biotechnology Foundation for Researchand Technology, Hellas, Herakleion Crete, Greece).Transient Transfection Assay—HepG2 cells were grown to
�80% confluence in 24-well tissue culture plates. Luciferasereporters and expression plasmidswere transfected using Lipo-fectamine 2000 reagent (Invitrogen) following themanufactur-er’s instructions. The pcDNA3 empty vector was added tonormalize the amounts ofDNA transfected in each assay. Lucif-erase and�-galactosidase activities were assayed and expressedin relative luciferase units as described previously (28).RNA Isolation and Quantitative Real-time PCR—Primary
human hepatocytes were cultured in serum-free and insulin-freemedia for 24 h and treatedwith insulin (Sigma) as indicated. RNAisolation, reverse transcription reactions, and real-time PCRwereperformed as described previously (29). All TaqMan Probes,CYP7A1 (HS00167982), FoxO1 (HS00231106), SREBP-1(HS00231674), HNF4� (HS00230853), and UBC (HS00824723),were ordered from Gene Expression Assays (Applied BiosystemsInc., Foster City, CA). PEPCK probe was custommade fromCus-tomGeneExpressionAssays (AppliedBiosystems, Inc.).UbiquitinC was used as an internal control for all PCR amplification reac-tions. Relative mRNA expression was quantified using the com-parativeCt (�Ct)methodandexpressedas2���Ct. Eachassaywasdone in triplicate and expressed as the mean � S.D.Electrophoretic Mobility Shift Assay (EMSA)—FoxO1 and
FoxA1 were synthesized in vitro using the transcription/trans-lation (TNT) system programmed with the FoxO1 and FoxA1expression plasmids according to the manufacturer’s instruc-tion (Promega). [�-32P]dCTP (3000 Ci/mol) was obtained fromPerkinElmer Life Sciences. Synthetic oligonucleotides of com-plimentary strands were labeled with [�-32P]dCTP and incu-bated with in vitro translated proteins (5 �l) as described pre-viously (28). Gels were dried and autoradiographed using a
Insulin Regulation of CYP7A1
28746 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 39 • SEPTEMBER 29, 2006

PhosphorImager 445Si (GE Healthcare). Sequences of theprobes (taggedwithGATCat the 5� end for labeling) used in theexperiments are listed in the supplemental data. Oligonucleo-tides were synthesized by MWG Biotech (High Point, NC).Site-directed Mutagenesis—A PCR-based QuikChange site-
directedmutagenesis kit (Stratagene, La Jolla, CA) was used formutation of the reporter constructs; mutant rIRE2 sequencewas introduced into rat p-344/luc plasmid, and the HNF4�binding site mutation was introduced into ph-1887/Luc plas-mid as described previously (28).Chromatin Immunoprecipitation (ChIP) Assay—Human pri-
mary hepatocytes were plated in T75 tissue culture flasks.HepG2 cells overexpressing HA-PGC-1� were cultured inserum-free and insulin-free media for 24 h. ChIP assays wereperformed as described previously (28). Antibodies againstHNF4� (sc-8987), SREBP-1 (sc-366), and HA tag (sc-805,Lot#K0303) were purchased from Santa Cruz Biotechnology(Santa Cruz, CA), and FoxO1 (#9462) was from Cell Signal-ing Technology, Inc. (Danvers, MA). Antibodies were usedto immunoprecipitate chromatin. Non-immuno IgG (SantaCruz) was used as negative control for immunoprecipitation.A 391-bp fragment containing the HNF4� binding site wasamplified by PCR and analyzed on a 1.5% agarose gel.Isolation of Cell Nuclei—Approximately 107 cells were
washed 3 times with cold 1� phosphate-buffered saline. Cellswere collected by centrifugation at 4 °C and re-suspended in 1ml of buffer A (10 mM Tris-Cl, pH 7.5, 10 mM NaCl, 3 mMMgCl2) containing protease inhibitors (Sigma) and incubatedon ice for 10 min. 100 �l of 10% Nonidet P-40 was then added,and cells were incubated for another 10 min on ice. Cells werelysed by passing through a 22-gauge needle 20 times. Celllysates were centrifuged at 3000 rpm at 4 °C for 10 min to pre-cipitate the nuclei. Nuclei were washed 3 times in ice-coldbuffer A and lysed in SDS lysis buffer for immunoblottinganalysis.Immunoblotting Analysis—Primary human hepatocytes were
treatedwith 10 nM insulin as described above. Total cell lysates ornuclei fractions were analyzed by SDS-polyacrylamide gel elec-trophoresis. An antibody against phospho-FoxO1 (Ser-256)(#9461; Cell Signaling Technology, Inc.) and antibodies againstFoxO1 (sc-9462), HNF4� (sc-8987), and actin (sc-1615) (SantaCruz Biotechnology) were used for Western blotting anddetected by ECL Western blotting detection kit (AmershamBiosciences).
RESULTS
Effect of Insulin on CYP7A1 and Transcription Factor mRNAExpression in Human Primary Hepatocytes—We first studiedthe effect of insulin on CYP7A1 mRNA expression in humanprimary hepatocytes using quantitative real-time PCR. Fig. 1Ashows that increasing doses of insulin (0.1 nM to 100 nM) rapidlyinduced CYP7A1 mRNA levels in 2 h, with a maximum induc-tion of �22-fold at the physiological concentration of 10 nM inthis donor liver. Higher concentrations of insulin had less stim-ulatory effect on CYP7A1mRNA expression. In contrast, insu-lin treatment for 6 h inhibited CYP7A1mRNA by �40% at 100nM (Fig. 1B). As a positive control, insulin strongly reducedPEPCKmRNAexpression in primary hepatocytes at both 2 and
6 h (Fig. 1, A and B). To further study the insulin effect onCYP7A1 mRNA expression, we treated primary hepatocyteswith insulin (10 nM) for a period of time from 2 to 24 h. Quan-titative real-time PCR analysis of primary hepatocytes from fivedonors showed that insulin induced CYP7A1 mRNA expres-sion levels at 2 h by an average of �9-fold and inhibitedCYP7A1 mRNA at 6 and 24 h by �36 and �39%, respectively(supplemental Table 1). As a positive control, insulin stronglyinhibited PEPCK mRNA in a time-dependent manner.We also measured the mRNA expression levels of HNF4�,
the key regulator of human CYP7A1 gene transcription, andSREBP-1c and FoxO1, two transcription factors that are knownto mediate the effects of insulin (Table 1, supplemental data).Insulin significantly induced HNF4� mRNA levels at 2 h by�30% but inhibited it at 6 and 24 h. On the other hand, insulininduced SREBP-1c by about 2-fold, consistent with a previousreport (30). Insulin had no effect on FoxO1 mRNA expression,consistent with previous studies that insulin regulates FoxO1mainly at post-translational levels (15, 18, 19).Identification of the IREs in the CYP7A1 Gene—FoxO1 is
known to bind to the consensus IRE, T(G/A)TTT(T/G)(G/T),in the promoters of glucose-6-phosphatase, PEPCK, insulin-like growth factor binding protein-1, and tyrosine aminotrans-ferase and mediates the inhibitory effect of insulin on these
FIGURE 1. Real-time PCR assays of the effect of insulin on CYP7A1 andPEPCK mRNA expression in human primary hepatocytes. Primary humanhepatocytes (HH 1251) were treated with increasing concentrations of insulinfor 2 h (A) or 6 h (B). CYP7A1 or PEPCK mRNA expression was assayed intriplicate and expressed as the mean � S.D.
Insulin Regulation of CYP7A1
SEPTEMBER 29, 2006 • VOLUME 281 • NUMBER 39 JOURNAL OF BIOLOGICAL CHEMISTRY 28747

FIGURE 2. EMSA of FoxO1 and FoxA1 interaction with the putative rat and human IREs. A, putative IREs in rat (r) and human (h) CYP7A1 promoter. B, EMSAof in vitro synthesized FoxO1 and FoxA1 binding to rat and human IRE probes. A glucose-6-phosphatase IRE probe was used as a positive control. Unpro-grammed TNT lysates (T3 and T7) were used as negative controls for nonspecific bindings. Sequences of IRE probes used are described in the supplementaldata. C, EMSA of in vitro synthesized FoxO1 and FoxA1 with 32P-labeled rat CYP7A1 IRE2 and mutant IRE2 probes. Competition assays were done with a 100-foldexcess of unlabeled probes. Mutations in rIRE2 probes is shown. G6Pase, glucose 6-phosphatase. D, effects of FoxO1 on rat CYP7A1 reporter activities. WT, wildtype rat CYP7A1 reporter (p-344/luc). RLU, relative light units. Mutant, rat CYP7A1 reporter (p-344/luc) containing single nucleotide mutation in IRE2 sequence.Reporter (0.2 �g) and the indicated expression plasmids (0.1 �g) were transfected into HepG2 cells. Transient transfection and luciferase assays were per-formed as described “Experimental Procedures.” A mutation in rat IRE2 is shown. Statistical analysis was performed by Student’s t test; *, significant, p � 0.05.
Insulin Regulation of CYP7A1
28748 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 39 • SEPTEMBER 29, 2006

genes (31). Analysis of the nucleotide sequences in the CYP7A1promoter identified four putative IREs in the rat gene and twoputative IREs in the human gene (Fig. 2A). Only the IRE locatedin the bile acid response element-I (�81 to�75) is conserved inrat and humanCYP7A1 genes.We then performed an EMSA totest if FoxO1 bound to these IREs in the CYP7A1 gene. Fig. 2Bshows that FoxO1 bound strongly to the rat IRE2 (rIRE2) butdid not bind to rIRE1, rIRE3, rIRE4, human (h) IRE1 and hIRE2despite the similarity in nucleotide sequences (Fig. 2A). FoxO1binding to rIRE2 was competed out by a large excess of unla-beled rIRE2 probe and a glucose-6-phosphatase probe that con-tained three FoxO1 binding sites (Fig. 2C) (16). This rIRE2 alsoboundFoxA1 (HNF3�) as expected.A singlemutation ofG toTin the rIRE2 abolished Foxo1 binding (Fig. 2C). To test theeffect of FoxO1 on rat CYP7A1 gene transcription, reporterassays were performed in HepG2 cells with a rat CYP7A1 pro-moter/luciferase construct. Co-transfection of a FoxO1 expres-sion plasmid stimulated rat CYP7A1 reporter activity by�4-fold (Fig. 2D, left panel). When a single mutation that abol-ished FoxO1 binding was introduced into the rat CYP7A1reporter, the basal reporter activity was reduced by more than80%. Co-transfection of FoxO1 failed to stimulate the mutantreporter activity (Fig. 2D, left panel). This mutant reporter didrespond to HNF4� and LXR� because it retained the HNF4�and LXR� binding sites as in the wild type plasmid (Fig. 2D,right panel).FoxO1 Inhibited Human CYP7A1 Reporter Activity by Block-
ing HNF4� Trans-activation Activity—The effect of FoxO1 onhuman CYP7A1 reporter activity was studied using transfec-tion assays. In contrast to the stimulatory effect on the ratCYP7A1 reporter, co-transfection of FoxO1 inhibited humanCYP7A1 reporter activities of ph-1887/Luc, ph-371/Luc, andph-150/Luc by �70% (Fig. 3A). Further deletion of thesequence downstream of �150 abolished the inhibitory effectof FoxO1. Thus, the inhibitory effect of FoxO1 required theregion from �150 to �135, which was previously mapped asthe bile acid response element-II that contained an HNF4�binding site (32). It has been reported that FoxO1 interacts withtheDNAbinding domain ofHNF4� and inhibits HNF4� trans-activation activity and its DNA binding (21). To test if theHNF4� binding site was involved in FoxO1 regulation of thehuman CYP7A1 gene, the HNF4� binding site in the ph-1887/Luc reporter was mutated. Fig. 3B shows that the HNF4� bind-ing site mutant reporter (mHNF4�-ph-1887/luc) had a muchlower basal activity, and FoxO1 did not inhibit the mutantreporter activity. These results suggest that FoxO1 mightinhibit human CYP7A1 gene by inhibiting HNF4� trans-acti-vation activity. A mammalian one-hybrid assay was used tostudy the effect of FoxO1 on HNF4� trans-activation of a Gal4(5�UAS)/TK/Luc reporter. Fig. 3C shows that Gal4-HNF4�activation of the GAL4 reporter activity was drasticallyincreased by PGC-1�, and FoxO1 dose-dependently inhibitedreporter activity (Fig. 3C). These results suggest that in additionto interfering with HNF4� binding to DNA, FoxO1 might alsoblock HNF4� and PGC-1� interaction. The effect of FoxO1 onHNF4� and PGC-1� co-activation of human CYP7A1 reporteractivity (Fig. 3D) was also studied. HNF4� or PGC-1� stimu-lated human CYP7A1 reporter activity by less than 100%, likely
the result of abundant HNF4� expression in HepG2 cells.FoxO1 strongly inhibited the human CYP7A1 reporter activitystimulated by HNF4� and/or PGC-1�. Taken together, theseresults suggest that FoxO1 could act as a DNA binding inde-pendent co-repressor of HNF4� and interfere withHNF4� andPGC-1� co-activation of CYP7A1 gene transcription.SREBP-1c Inhibited Human CYP7A1 Gene Transcription by
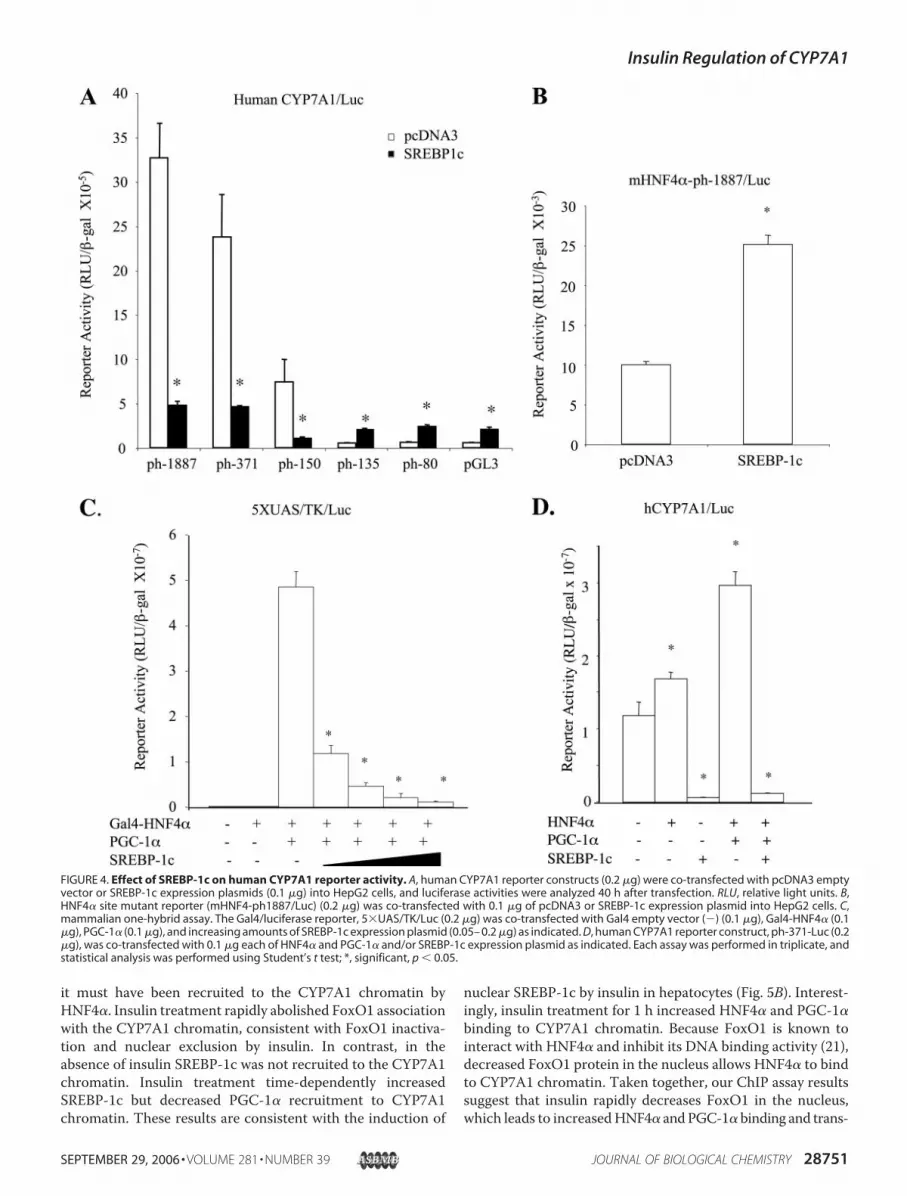
Blocking HNF4� Recruitment of PGC-1�—To test if SREBP-1cwas involved inmediating insulin effect onhumanCYP7A1, theeffect of SREBP-1c on humanCYP7A1 reporterswas studied bytransfection assays in HepG2 cells. Co-transfection of thenuclear form of SREBP-1c strongly inhibited the humanCYP7A1 reporter (ph-1887/Luc) by more than 80% (Fig. 4A).Deletion analyses of the human CYP7A1 promoter localized aregion from �150 to �135 that mediated the inhibitory effectof SREBP-1c (Fig. 4A). We then tested the effect of SREBP-1con the HNF4� binding site mutant reporter (mHNF4�-ph-1887/luc). As shown in Fig. 4B, mutation of theHNF4� bindingsitemarkedly reduced the basal reporter activity, and SREBP-1cfailed to inhibit the mutant reporter activity. These results sug-gest that SREBP-1cmight inhibit humanCYP7A1 by inhibitingHNF4� trans-activation ofCYP7A1. SREBP-1c has been shownto interact with HNF4�-LBD and block its recruitment ofPGC-1� to the PEPCK gene (33). Thus, a mammalian one-hybrid assay was used to study the effect of SREBP-1c onPGC-1� and Gal4-HNF4� co-activation of a GAL4 5�UAS/TK/Luc reporter. Fig. 4C shows that SREBP-1c inhibited Gal4-HNF4�-LBD andPGC-1� co-activation of reporter activity in adose-dependent manner. Furthermore, SREBP-1c stronglyinhibited the basal activity of a human CYP7A1 reporter andthe activity stimulated by HNF4� and PGC-1� (Fig. 4D). Thus,SREBP-1c could act as a DNA binding independent co-repres-sor of HNF4� and interfere with HNF4� and PGC-1� co-acti-vation of CYP7A1 gene transcription.Western Blot Analysis of Effects of Insulin on FoxO1, SREBP-
1c, and HNF4� Protein Expressions in Primary HumanHepatocytes—To study effect of insulin on phosphorylation ofFoxO1 and expression of SREBP-1c protein in primary humanhepatocytes, immunoblot analysis of FoxO1 and SREBPproteinexpression was conducted. Fig. 5A shows that without insulintreatment, FoxO1 was not phosphorylated and was mainlylocated in the nucleus. Insulin treatment for 30min led to phos-phorylation of FoxO1 and its rapid nuclear exclusion and deg-radation. Immunoblot analysis did not detect SREBP-1c pre-cursor (125 kDa) expression in hepatocytes (data not shown).Insulin treatment increased the amount of mature SREBP-1c(68 kDa) in cell lysates in a time-dependent manner (Fig. 5B).Insulin treatment did not significantly affect HNF4� proteinlevels in hepatocytes (Fig. 5C). The time courses of insulin inac-tivation of FoxO1 and induction of SREBP-1c in hepatocytescorrelate well with quantitative real-time PCR results onCYP7A1 expression and support our hypothesis that insulininactivates FoxO1 to rapidly induce CYP7A1 gene expressionand that prolonged insulin treatment induces SREBP-1c toinhibit CYP7A1 gene expression.ChIP Assay of HNF4� Binding and Recruitment of SREBP-1c,
FoxO1, and PGC-1� to the Human CYP7A1 Chromatin—Tofurther confirm the roles of FoxO1 and SREBP-1c in mediating
Insulin Regulation of CYP7A1
SEPTEMBER 29, 2006 • VOLUME 281 • NUMBER 39 JOURNAL OF BIOLOGICAL CHEMISTRY 28749

insulin regulation of human CYP7A1 gene transcription, ChIPassays were performed using primary human hepatocytestreatedwith 10 nM insulin for a period of time from1 to 24 h.Anantibody against FoxO1, SREBP-1c, HNF4�, or PGC-1� wasused to immunoprecipitate chromatin complexes for ChIP
assays using primers designed to amplify a 391-bp fragment ofhuman CYP7A1 promoter region that included an HNF4�binding site. In the absence of insulin, ChIP assays detectedFoxO1 and HNF4� in the CYP7A1 chromatin (Fig. 6A).Because FoxO1 does not bind to the humanCYP7A1 promoter,
FIGURE 3. Effect of FoxO1 on human CYP7A1/Luc reporter activity. A, CYP7A1 reporter constructs (0.2 �g) were co-transfected with expression plasmids(0.1 �g) into HepG2 cells, and luciferase activity (relative light units (RLU)) was analyzed 40 h after transfection. RLU, relative light units. B, HNF4� site mutantreporter (mHNF4-ph1887-Luc) was transfected in HepG2 cells. pcDNA3 empty vector or FoxO1 expression plasmid was co-transfected as indicated. C, mam-malian one-hybrid assay. The Gal4/luciferase reporter 5�UAS/TK/Luc (0.2 �g) was co-transfected with Gal4 empty vector (�) (0.1 �g), Gal4-HNF4� (0.1 �g),PGC-1� (0.1 �g), and increasing amounts of FoxO1 expression plasmid (0.05– 0.2 �g) as indicated. D, a human CYP7A1 reporter, ph-371-Luc (0.2 �g), wasco-transfected with FoxO1, HNF4�, and/or PGC-1� (0.1 �g) as indicated into HepG2 cells. Each assay was performed in triplicate, and statistical analysis wasperformed using Student’s t test; *, significant, p � 0.05.
Insulin Regulation of CYP7A1
28750 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 39 • SEPTEMBER 29, 2006
it must have been recruited to the CYP7A1 chromatin byHNF4�. Insulin treatment rapidly abolished FoxO1 associationwith the CYP7A1 chromatin, consistent with FoxO1 inactiva-tion and nuclear exclusion by insulin. In contrast, in theabsence of insulin SREBP-1c was not recruited to the CYP7A1chromatin. Insulin treatment time-dependently increasedSREBP-1c but decreased PGC-1� recruitment to CYP7A1chromatin. These results are consistent with the induction of
nuclear SREBP-1c by insulin in hepatocytes (Fig. 5B). Interest-ingly, insulin treatment for 1 h increased HNF4� and PGC-1�binding to CYP7A1 chromatin. Because FoxO1 is known tointeract with HNF4� and inhibit its DNA binding activity (21),decreased FoxO1 protein in the nucleus allows HNF4� to bindto CYP7A1 chromatin. Taken together, our ChIP assay resultssuggest that insulin rapidly decreases FoxO1 in the nucleus,which leads to increasedHNF4� andPGC-1�binding and trans-
FIGURE 4. Effect of SREBP-1c on human CYP7A1 reporter activity. A, human CYP7A1 reporter constructs (0.2 �g) were co-transfected with pcDNA3 emptyvector or SREBP-1c expression plasmids (0.1 �g) into HepG2 cells, and luciferase activities were analyzed 40 h after transfection. RLU, relative light units. B,HNF4� site mutant reporter (mHNF4-ph1887/Luc) (0.2 �g) was co-transfected with 0.1 �g of pcDNA3 or SREBP-1c expression plasmid into HepG2 cells. C,mammalian one-hybrid assay. The Gal4/luciferase reporter, 5�UAS/TK/Luc (0.2 �g) was co-transfected with Gal4 empty vector (�) (0.1 �g), Gal4-HNF4� (0.1�g), PGC-1� (0.1 �g), and increasing amounts of SREBP-1c expression plasmid (0.05– 0.2 �g) as indicated. D, human CYP7A1 reporter construct, ph-371-Luc (0.2�g), was co-transfected with 0.1 �g each of HNF4� and PGC-1� and/or SREBP-1c expression plasmid as indicated. Each assay was performed in triplicate, andstatistical analysis was performed using Student’s t test; *, significant, p � 0.05.
Insulin Regulation of CYP7A1
SEPTEMBER 29, 2006 • VOLUME 281 • NUMBER 39 JOURNAL OF BIOLOGICAL CHEMISTRY 28751

activation of the CYP7A1 gene. Insulin induces SREBP-1c, whichis recruited toCYP7A1chromatin toblockHNF4� recruitmentofPGC-1�, and results in inhibitingCYP7A1 gene transcription.ChIP assays were also performed using HepG2 cells overex-
pressing HA-PGC-1� to study the effect of insulin on HNF4�recruitment of PGC-1� to CYP7A1 chromatin. Fig. 6B showsthat insulin (100 nM) treatment for 20 h reduced PGC-1�recruitment to CYP7A1 chromatin. T0901317, a potent LXR�agonist, also strongly reduced PGC-1� recruitment. These dataare consistent with a previous report that insulin and LXR�induce SREBP-1c, which blocks PGC-1� interaction withHNF4� (33). Furthermore, we overexpressed SREBP-1c orFoxO1 in HepG2 cells to test their effect on PGC-1� recruit-ment to CYP7A1 chromatin. Fig. 6C shows that overexpressionof SREBP-1c or FoxO1 abolished PGC-1� recruitment toCYP7A1 chromatin.
DISCUSSION
In the current study we investigated the insulin regulation ofhuman CYP7A1 gene transcription in human hepatocytes.Quantitative real-time PCR analyses have revealed that physi-ological concentrations of insulin strongly and rapidly induce
CYP7A1 mRNA expression. Interestingly, prolonged insulintreatment results in inhibition of human CYP7A1 mRNAexpression. Results indicate that the dual effect of insulin actionon human CYP7A1 is likely mediated through two insulin-reg-ulated transcription factors, FoxO1 and SREBP-1c. FoxO1binds to the IRE in the rat CYP7A1 gene promoter and stimu-lates rat CYP7A1 reporter activity. However, this IRE is notconserved in the human CYP7A1 promoter. Our results sug-
FIGURE 5. Western blot analysis of FoxO1, SREBP-1c, and HNF4� in pri-mary human hepatocytes treated with insulin. Primary human hepato-cytes were treated with 10 nM insulin for the times indicated. Total cell lysatesor nuclei fractions from donor HH1286, HH1281, and HH1249 were used fordetection of FoxO1 (A), SREBP-1c (B), and HNF4� (C ) by antibodies, respec-tively. P-FoxO1, phosphorylated FoxO1.
FIGURE 6. Chromatin immunoprecipitation assays of the effect of insulinon FoxO1, SREBP-1c, and HNF4� recruitment to CYP7A1 chromatin. A,human primary hepatocytes (HH1308) were treated with 10 nM insulin for thetime indicated. Cells were harvested for ChIP assay as described “Experimen-tal Procedures.” An antibody against HNF4�, SREBP-1, or FoxO1 was used toimmunoprecipitate chromatin for PCR amplification and analysis. 10% of celllysate was used as input. Non-immune IgG was used as negative controls. B,ChIP assays of the effect of insulin and T0901317 on HNF4� recruitment ofPGC-1� to CYP7A1 chromatin. HepG2 cells in 100 mm dishes were trans-fected with HA-tagged PGC-1� (10 �g). Cells were then treated with insulin(100 nM) or T0901317 (1 �M) for 24 h. Chromatin was precipitated (IP) with anantibody against HA tag or HNF4� for ChIP assays. 10% of cell lysate was usedas input. Non-immune IgG was used as negative controls. C, ChIP assays ofHepG2 cells overexpressing SREBP-1c or FoxO1. HepG2 cells in a 100-mmdishes were transfected with 10 �g of HA-tagged PGC-1�, pcDNA3 emptyvector, and FoxO1, or SREBP-1c expression plasmid was overexpressed asindicated. Cells were harvested for ChIP assay as described under “Experi-mental Procedures.” An antibody against HA tag or HNF4� was used to immu-noprecipitate chromatin for PCR amplification and analysis. 10% of cell lysatewas used as input. Non-immune IgG was used as a negative control.
Insulin Regulation of CYP7A1
28752 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 39 • SEPTEMBER 29, 2006

gest that FoxO1 functions as a co-repressor that inhibits thehuman CYP7A1 gene by interfering with the HNF4� andPGC-1� co-activation of the gene. The ChIP assay reveals thatFoxO1 is present in CYP7A1 chromatin without insulin treat-ment, and FoxO1may interact withHNF4� to block its recruit-ment of PGC-1� to CYP7A1 chromatin (Fig. 6A). Upon insulintreatment, FoxO1 is released, and PGC-1� is recruited byHNF4� to transiently stimulate CYP7A1 mRNA expression inhuman hepatocytes. Prolonged insulin treatment induces themature form of SREBP-1c, which is recruited to CYP7A1 chro-matin to inhibit HNF4� and PGC-1� trans-activation of thehuman CYP7A1 gene. This study suggests that CYP7A1 is atarget gene of both FoxO1 and SREBP-1c, which play criticalroles in regulating bile acid synthesis in response to insulinlevels in hepatocytes.Bile acids are known to activate a nuclear receptor farnesoid X
receptor, which induces a negative nuclear receptor small het-erodimer partner to inhibit nuclear receptor-regulated genetranscription (34). Several recent reports have shown that bileacids play critical roles in the regulation of glucose and lipidmetabolism (35–37). During the postprandial state, seruminsulin levels increase transiently to stimulate glucose metabo-lism. According to our study, insulin may rapidly induceCYP7A1 and bile acid synthesis to facilitate nutrient absorptionin the intestine during the postprandial state. Bile acids inhibitthe PEPCK gene and prevent hyperglycemia (38). During thepost-absorptive state and starvation, glucagon levels increase toinhibit CYP7A1 and bile acid synthesis while stimulating glu-coneogenesis to prevent hypoglycemia (39).In type II diabetes, FoxO1 nuclear exclusion is impaired due
to insulin resistance (17). Hyperinsulinemia may inhibitCYP7A1 and bile acid synthesis by inducing SREBP-1c, whichstimulates lipogenesis and contributes to hypertriglyceridemia.Inhibition of bile acid synthesis stimulates gluconeogenesis andlipogenesis. In type I diabetes, lack of insulin activates FoxO1,which stimulates gluconeogenesis, but inhibits bile acid synthe-sis and also causes hyperglycemia and dyslipidemia. All thesemetabolic changes are caused by insulin and may contribute tothe development of diabetic dyslipidemia and cardiovasculardiseases.It should be noted that we have demonstrated distinct spe-
cies differences in FoxO1 and SREBP-1c regulation of rat andhuman CYP7A1 gene transcription. FoxO1 induces ratCYP7A1 by binding to an IRE (TGTTTAC) in the rat genepromoter, but FoxO1 does not bind and inhibit the humanCYP7A1 gene. Analysis of homologous CYP7A1 genes revealsthat this rat IRE sequence is also conserved in mouse and ham-ster genes. In the mouseCYP7A1 gene this IRE is located in thesame region and is likely a FoxO1 binding site. A recent reportshows a 2-fold higher expression of CYP7A1 mRNA in FoxO1transgenic mice, suggesting induction of mouse CYP7A1 byFoxO1 (40). In the hamster CYP7A1 gene this IRE is locatedupstream of a putative IRE that binds HNF3� and HNF4� (41).SREBP-1c also differentially regulates the rat and humanCYP7A1 genes. SREBP-1c binds to the E-box (CANNTG) andSRE motifs and activates gene transcription (42). However,SREBP-1c also can function as a DNA binding-independentrepressor by interfering with HNF4� interaction with PGC-1�
and inhibiting PEPCK gene transcription (33). This studyshows that the same mechanism also inhibits the humanCYP7A1 gene. In contrast, we found that SREBP-1c stronglystimulated rat CYP7A1 reporter activity (data not shown).Analysis of the rat CYP7A1 gene promoter sequence revealsthree E-boxes and several GC-rich sequences located down-stream of �344 of the rat CYP7A1 promoter. These E-boxmotifs are not conserved in the human CYP7A1 gene. Furtherstudy is needed to identify the SREBP-1c binding site in the ratCYP7A1 gene and the mechanism by which FoxO1 andSREBP-1c regulates the rat CYP7A1 gene. Previously we havereported that in contrast to the induction of CYP7A1 mRNAexpression in mouse liver, LXR� does not stimulate humanCYP7A1 gene transcription because the human CYP7A1 genelacks a LXR� response element (43). Species differences in reg-ulation of the CYP7A1 gene and lipid metabolism underscorethe importance of studying human CYP7A1 gene regulation inhuman hepatocyte models.In summary, we studied the mechanisms of insulin regula-
tion of CYP7A1 gene transcription mediated by FoxO1 andSREBP-1c. Our results suggest that insulin has a dual effect onhumanCYP7A1 gene transcription; that is, a rapid induction atphysiological concentrations to control glucose and lipidmetabolism, and later, inhibition to maintain glucose and lipidhomeostasis. Understanding the complex mechanism of insu-lin regulation of bile acid synthesis and lipid homeostasis willlead to developing novel drug therapies for treating diabeticdyslipidemia.
Acknowledgments—The technical assistance of Bethany Spalding-Yoder and Sandra Del Signore is gratefully acknowledged. Primaryhuman hepatocytes were supplied by the Liver Procurement and Dis-tribution System funded by Contract DK92310 from NIDDK.
REFERENCES1. Reaven, G., Abbasi, F., andMcLaughlin, T. (2004) Recent Prog. Horm. Res.
59, 207–2232. Chiang, J. Y. L. (2003) Am. J. Physiol. Gastrointest. Liver Physiol. 284,
349–3563. Bennion, L. J., and Grundy, S. M. (1977) N. Engl. J. Med. 296, 1365–13714. Andersen, E., Karlaganis, G., and Sjovall, J. (1988) Eur. J. Clin. Investig. 18,
166–1725. Hassan, A. S., Ravi Subbiah, M. T., and Thiebert, P. (1980) Proc. Soc. Exp.
Biol. Med. 164, 449–4526. Villanueva, G. R., Herreros,M., Perez-Barriocanal, F., Bolanos, J. P., Bravo,
P., and Marin, J. J. (1990) J. Lab. Clin. Med. 115, 441–4487. Wang, D. P., Stroup, D., Marrapodi, M., Crestani, M., Galli, G., and
Chiang, J. Y. (1996) J. Lipid Res. 37, 1831–18418. Crestani,M., Sadeghpour, A., Stroup, D., Galli, G., and Chiang, J. Y. (1998)
J. Lipid Res. 39, 2192–22009. Twisk, J., Hoekman, M. F., Lehmann, E. M., Meijer, P., Mager, W. H., and
Princen, H. M. (1995) Hepatology 21, 501–51010. Ishida, H., Yamashita, C., Kuruta, Y., Yoshida, Y., and Noshiro, M. (2000)
J. Biochem. (Tokyo) 127, 57–6411. Foufelle, F., and Ferre, P. (2002) Biochem. J. 366, 377–39112. O’Brien, R. M., and Granner, D. K. (1996) Physiol. Rev. 76, 1109–116113. Hall, R. K., Yamasaki, T., Kucera, T., Waltner-Law, M., O’Brien, R., and
Granner, D. K. (2000) J. Biol. Chem. 275, 30169–3017514. Schmoll, D.,Walker, K. S., Alessi, D. R., Grempler, R., Burchell, A., Guo, S.,
Walther, R., and Unterman, T. G. (2000) J. Biol. Chem. 275, 36324–3633315. Puigserver, P., Rhee, J., Donovan, J., Walkey, C. J., Yoon, J. C., Oriente, F.,
Insulin Regulation of CYP7A1
SEPTEMBER 29, 2006 • VOLUME 281 • NUMBER 39 JOURNAL OF BIOLOGICAL CHEMISTRY 28753

Kitamura, Y., Altomonte, J., Dong, H., Accili, D., and Spiegelman, B. M.(2003) Nature 423, 550–555
16. Nakae, J., Kitamura, T., Silver, D. L., and Accili, D. (2001) J. Clin. Investig.108, 1359–1367
17. Altomonte, J., Cong, L., Harbaran, S., Richter, A., Xu, J., Meseck, M., andDong, H. H. (2004) J. Clin. Investig. 114, 1493–1503
18. Nakae, J., Park, B. C., andAccili, D. (1999) J. Biol. Chem. 274, 15982–1598519. Zhang, X., Gan, L., Pan, H., Guo, S., He, X., Olson, S. T., Mesecar, A.,
Adam, S., and Unterman, T. G. (2002) J. Biol. Chem. 277, 45276–4528420. Matsuzaki, H., Daitoku, H., Hatta, M., Tanaka, K., and Fukamizu, A.
(2003) Proc. Natl. Acad. Sci. U. S. A. 100, 11285–1129021. Hirota, K., Daitoku, H., Matsuzaki, H., Araya, N., Yamagata, K., Asada, S.,
Sugaya, T., and Fukamizu, A. (2003) J. Biol. Chem. 278, 13056–1306022. Foretz, M., Pacot, C., Dugail, I., Lemarchand, P., Guichard, C., Le Liepvre,
X., Berthelier-Lubrano, C., Spiegelman, B., Kim, J. B., Ferre, P., andFoufelle, F. (1999)Mol. Cell. Biol. 19, 3760–3768
23. Horton, J. D., Goldstein, J. L., and Brown,M. S. (2002) J. Clin. Investig. 109,1125–1131
24. Engelking, L. J., Kuriyama, H., Hammer, R. E., Horton, J. D., Brown, M. S.,Goldstein, J. L., and Liang, G. (2004) J. Clin. Investig. 113, 1168–1175
25. Tobin, K. A., Ulven, S. M., Schuster, G. U., Steineger, H. H., Andresen,S. M., Gustafsson, J. A., and Nebb, H. I. (2002) J. Biol. Chem. 277,10691–10697
26. Repa, J. J., Liang, G., Ou, J., Bashmakov, Y., Lobaccaro, J. M., Shimomura,I., Shan, B., Brown, M. S., Goldstein, J. L., and Mangelsdorf, D. J. (2000)Genes Dev. 14, 2819–2830
27. Hegarty, B. D., Bobard, A., Hainault, I., Ferre, P., Bossard, P., and Foufelle,F. (2005) Proc. Natl. Acad. Sci. U. S. A. 102, 791–796
28. Li, T., and Chiang, J. Y. (2004) Am. J. Physiol. Gastrointest. Liver Physiol.288, 74–84
29. Song, K.H., Li, T., andChiang, J. Y. (2006) J. Biol. Chem.281, 10081–10088
30. Cagen, L. M., Deng, X., Wilcox, H. G., Park, E. A., Raghow, R., and Elam,M. B. (2004) Biochem. J. 385, 207–216
31. O’Brien, R. M., Noisin, E. L., Suwanichkul, A., Yamasaki, T., Lucas, P. C.,Wang, J. C., Powell, D. R., and Granner, D. K. (1995) Mol. Cell. Biol. 15,1747–1758
32. Stroup, D., and Chiang, J. Y. (2000) J. Lipid Res. 41, 1–1133. Yamamoto, T., Shimano, H., Nakagawa, Y., Ide, T., Yahagi, N.,Matsuzaka,
T., Nakakuki,M., Takahashi, A., Suzuki, H., Sone,H., Toyoshima,H., Sato,R., and Yamada, N. (2004) J. Biol. Chem. 279, 12027–12035
34. Chiang, J. Y. (2002) Endocr. Rev. 23, 443–46335. Stayrook, K. R., Bramlett, K. S., Savkur, R. S., Ficorilli, J., Cook, T., Christe,
M. E., Michael, L. F., and Burris, T. P. (2005) Endocrinology 146, 984–99136. Watanabe,M., Houten, S.M.,Wang, L.,Moschetta, A.,Mangelsdorf, D. J.,
Heyman, R. A., Moore, D. D., and Auwerx, J. (2004) J. Clin. Investig. 113,1408–1418
37. Cariou, B., Duran-Sandoval, D., Kuipers, F., and Staels, B. (2005) Endocri-nology 146, 981–983
38. Yamagata, K., Daitoku, H., Shimamoto, Y., Matsuzaki, H., Hirota, K.,Ishida, J., and Fukamizu, A. (2004) J. Biol. Chem. 279, 23158–23165
39. Song, K. H., and Chiang, J. Y. (2006) Hepatology 43, 117–12540. Zhang, W., Patil, S., Chauhan, B., Guo, S., Powell, D. R., Le, J., Klotsas, A.,
Matika, R., Xiao, X., Franks, R., Heidenreich, K. A., Sajan, M. P., Farese,R. V., Stolz, D. B., Tso, P., Koo, S. H., Montminy, M., and Unterman, T. G.(2006) J. Biol. Chem. 281, 10105–10117
41. De Fabiani, E., Crestani, M., Marrapodi, M., Pinelli, A., Golfieri, V., andGalli, G. (2000) Biochem. J. 347, 147–154
42. Osborne, T. F. (2000) J. Biol. Chem. 275, 32379–3238243. Chiang, J. Y., Kimmel, R., and Stroup, D. (2001) Gene (Amst.) 262,
257–26544. Crestani, M., Stroup, D., and Chiang, J. Y. L. (1995) J. Lipid Res. 36,
2419–2432
Insulin Regulation of CYP7A1
28754 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 39 • SEPTEMBER 29, 2006




















