
Inhibition of Glycosphingolipid Synthesis Induces a Profound Reduction of Plasma Cholesterol and...
16
ISSN: 1524-4636 Copyright © 2010 American Heart Association. All rights reserved. Print ISSN: 1079-5642. Online 7272 Greenville Avenue, Dallas, TX 72514 Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association. DOI: 10.1161/ATVBAHA.109.201673 2010; 2010;30;931-937; originally published online Feb 18, Arterioscler Thromb Vasc Biol Albert K. Groen Mariska Vos, Patrick C.N. Rensen, Arthur J. Verhoeven, Johannes M. Aerts and Florence Bietrix, Elisa Lombardo, Cindy P.A.A. van Roomen, Roelof Ottenhoff, and Low-Density Lipoprotein Receptor/ Mice Leiden Plasma Cholesterol and Inhibits Atherosclerosis Development in APOE*3 Inhibition of Glycosphingolipid Synthesis Induces a Profound Reduction of http://atvb.ahajournals.org/cgi/content/full/ATVBAHA.109.201673/DC1 Data Supplement (unedited) at: http://atvb.ahajournals.org/cgi/content/full/30/5/931 located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://atvb.ahajournals.org/subscriptions/ Biology is online at Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.org Downloaded from
Transcript of Inhibition of Glycosphingolipid Synthesis Induces a Profound Reduction of Plasma Cholesterol and...

ISSN: 1524-4636 Copyright © 2010 American Heart Association. All rights reserved. Print ISSN: 1079-5642. Online
7272 Greenville Avenue, Dallas, TX 72514Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association.
DOI: 10.1161/ATVBAHA.109.201673 2010;
2010;30;931-937; originally published online Feb 18,Arterioscler Thromb Vasc BiolAlbert K. Groen
Mariska Vos, Patrick C.N. Rensen, Arthur J. Verhoeven, Johannes M. Aerts and Florence Bietrix, Elisa Lombardo, Cindy P.A.A. van Roomen, Roelof Ottenhoff,
and Low-Density Lipoprotein Receptor�/� MiceLeidenPlasma Cholesterol and Inhibits Atherosclerosis Development in APOE*3
Inhibition of Glycosphingolipid Synthesis Induces a Profound Reduction of
http://atvb.ahajournals.org/cgi/content/full/ATVBAHA.109.201673/DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org/cgi/content/full/30/5/931
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://atvb.ahajournals.org/subscriptions/Biology is online at Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

Inhibition of Glycosphingolipid Synthesis Induces aProfound Reduction of Plasma Cholesterol and Inhibits
Atherosclerosis Development in APOE*3 Leiden andLow-Density Lipoprotein Receptor�/� Mice
Florence Bietrix, Elisa Lombardo, Cindy P.A.A. van Roomen, Roelof Ottenhoff, Mariska Vos,Patrick C.N. Rensen, Arthur J. Verhoeven, Johannes M. Aerts, Albert K. Groen
Objective—The iminosugar N-(5�-adamantane-1�-yl-methoxy)-pentyl-1-deoxynoijirimycin (AMP-DNM), an inhibitor ofthe enzyme glucosylceramide synthase catalyzing glycosphingolipid (GSL) biosynthesis, ameliorates diabetes andreduces liver steatosis in ob/ob mice. Because an accumulation of sphingolipids, including sphingomyelin and GSLs,has been reported in atherosclerotic lesions in animal models and in humans, the objective of this study was to determinewhether AMP-DNM also exerts beneficial effects on the development of atherosclerosis.
Methods and Results—APOE*3 Leiden mice, maintained on a high-cholesterol diet, were treated for up to 18 weeks withAMP-DNM. The iminosugar prevented hyperlipidemia, generated a less atherogenic lipid profile, and induced adramatic reduction in the development of atherosclerotic lesions. At the highest dose, no lesions were detectable. Theeffect of AMP-DNM was associated with a decrease in liver cholesterol, an increase in bile secretion, and enhancedexcretion of cholesterol in the feces. Similar effects of AMP-DNM were observed in mice deficient for the low-densitylipoprotein receptor.
Conclusion—By lowering plasma cholesterol, the iminosugar AMP-DNM dramatically reduces the development ofatherosclerosis in APOE*3 Leiden and low-density lipoprotein receptor �/� mice. Thus, targeting GSL synthesis maybe a new treatment modality to prevent cardiovascular disease. (Arterioscler Thromb Vasc Biol. 2010;30:931-937.)
Key Words: atherosclerosis � lipids � lipoproteins
Atherosclerosis is the primary cause of myocardial infarc-tion and stroke and is the underlying cause of �50% of
all deaths in Western countries. The use of statins as primaryand secondary prevention yields a 30% decrease in cardio-vascular risk. To achieve a further substantial drop in cardio-vascular morbidity and mortality, new therapeutic modalitiesneed to be developed. Targeting glycosphingolipid (GSL)synthesis may be an attractive alternative. In humans, theconcentration of plasma GSLs is increased in conditionsassociated with atherosclerosis risk and shows a positivecorrelation with plasma cholesterol.1,2 In addition, it has beenreported that GSLs accumulate in atherosclerotic lesions inhumans and in apolipoprotein E-null (apoE�/�) mice.3
Potential proatherogenic properties have been proposed forGSLs, mainly for lactosylceramide. Lactosylceramide wasfound to promote cholesterol accumulation in macrophagefoam cells, inhibit cholesterol efflux, and induce monocyteadhesion to endothelial cells.4–6 Myriocin, an inhibitor of
serine palmitoyl transferase, which catalyzes the initial step insphingolipid biosynthesis, profoundly reduces the develop-ment of atherosclerosis in apoE�/� mice.7,8 However, itsspecific effect is difficult to interpret, because in addition toGSL synthesis inhibition, myriocin also modulates numerousother sphingolipids, which may have a role in the progressionof the formation of atherosclerotic lesion.
The iminosugar N-(5�-adamantane-1�-yl-methoxy)-pentyl-1-deoxynoijirimycin (AMP-DNM) is a specific inhibitor ofglucosylceramide (GlcCer) synthase, the enzyme catalyzingthe initial step in GSL biosynthesis.9 By decreasing theavailability of GlcCer, AMP-DNM reduces the synthesis ofmore complex GSLs, such as gangliosides, which play acrucial role in cell membrane properties.10 Previous work hasshown that the use of the iminosugar improves glucosetolerance, reduces hepatic steatosis, and enhances insulinresponsiveness in rodent models for type 2 diabetes.11,12 Inlean mice, AMP-DNM treatment led to a decrease of plasma
Received on: December 9, 2009; final version accepted on: January 27, 2010.From Department of Medical Biochemistry, Academic Medical Center, University of Amsterdam, Amsterdam, the Netherlands (F.B., E.L.,
C.P.A.A.v.R., R.O., M.V., A.J.V., J.M.A., A.K.G.); Department of General Internal Medicine, Endocrinology and Metabolic Diseases, Leiden UniversityMedical Center, Leiden, the Netherlands (P.C.N.R.). A.K.G. is currently affiliated with the Department of Pediatrics, University Medical CenterGroningen, Groningen, the Netherlands.
Correspondence to Johannes M. Aerts, Department of Medical Biochemistry, Academic Medical Center, University of Amsterdam, Meibergdreef 15,1105 AZ, Amsterdam, the Netherlands. E-mail [email protected]
© 2010 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.109.201673
931 at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

lipids and to a stimulation of reverse cholesterol transport thatwas illustrated by an increase of biliary lipid secretion and anincrease of fecal sterol excretion.13 Furthermore, this com-pound was shown to convey a potent antiinflammatoryeffect.14,15 On the basis of all these observations, we postu-lated that AMP-DNM might also show beneficial effects oncardiovascular disease. Therefore, the aim of this study wasto evaluate the effect of AMP-DNM on the progression ofatherosclerosis.
Materials and MethodsMaterialsAMP-DNM was synthesized as previously described.16,17 All sol-vents and reagents used were of analytic grade.
Mice and DietsAPOE*3 Leiden transgenic mice18 were maintained on a C57Bl/6homogeneous background and housed at the Institute Animal CoreFacility in a temperature- and humidity-controlled room with a12-hour light/dark cycle. Studies were performed using femaleAPOE*3 Leiden mice (8 to 12 weeks old) fed for 18 weeks with ahigh-cholesterol diet (1% cholesterol, 15% fatty acids; Arie Blok,Woerden, the Netherlands) supplemented with 0, 0.3, or 0.6 g ofAMP-DNM � kg diet�1 to obtain a calculated dose of 50 or 100 mgof AMP-DNM � kg body weight (bw)�1 � day�1, respectively. Whenfemale low-density lipoprotein receptor (LDLR�/�) mice wereused, the diet contained 0.25% cholesterol for a period of 12 weeks.Experiments were performed with the approval of the EthicalCommittee for Animal Experiments.
Plasma and Tissue SamplingBlood samples of nonfasting animals were collected via the tail veinat the time points indicated in the figures. At the end of theexperiments, large blood samples were collected by abdominal aortapuncture, and plasma samples were stored at �20°C. Hearts wereflushed with PBS to remove the excess of blood before fixation informaldehyde 1% (Formal-Fixx, Thermo Electron Corporation,Pittsburgh, Pa.) for 24 hours and storage at �80°C embedded intissue medium (Tissue-Tek O.C.T., Sakura, Zoeterwoude, the Neth-erlands). Other tissues were snap frozen in liquid N2 and stored at�80°C.
Characterization of Atherosclerotic LesionsFrozen sections from the aortic sinus were prepared according toPaigen et al.19 Surface lesion area was measured after Oil Red Ostaining by computer-assisted image quantification with Leica QWinsoftware (Leica Microsystems, Wetzlar, Germany). Images werecaptured with a Leica DFC 420 video camera. At least 5 sections permouse were examined for each staining.
Bile and Feces SamplingTwenty-four hours of feces were collected from cages during thelast day of the treatment. Mice were kept in their original cages toavoid stressful procedures. Feces were freeze-dried, weighed, andground before storage at �20°C. Cannulation of the gall bladderand bile collection were performed as described previously.20
After cannulation, bile was collected for 15 minutes, and bilesamples were immediately frozen and stored at �20°C. Bile flowwas determined gravimetrically, assuming a density of 1 g � mL�1
for bile.
Measurement of Intestinal Cholesterol AbsorptionIntestinal cholesterol absorption was determined by the fecaldual-isotope ratio method21 on mice fed for 6 weeks with thehigh-cholesterol diet (see above) supplemented without or withAMP-DNM. Mice (n�5 per group) were given an intragastricbolus of 150 �L of olive oil containing 1 �Ci of [14C]cholesterol(Amersham Biosciences) and 2 �Ci of [3H]sitostanol (AmericanRadiolabeled Chemicals, St. Louis, Mo.) by gavage. Mice werehoused with 2 to 3 animals per cage. Total feces per cage werecollected for 3 days, dried, and ground before extraction asdescribed below. The ratios of the 2 radiolabels in the fecalextracts and in the dosing mixture were used to calculate thepercentage of cholesterol absorption.
Analytic ProceduresCeramide (Cer) and GlcCer were determined in liver and plasmasamples after lipids extraction according to Folch et al22 by high-performance liquid chromatography analysis of orthophtaldehyde-conjugated lipids according to a procedure described previously.11
Fecal neutral sterols were extracted from the feces and measured bygas chromatography.23 Biliary cholesterol, bile salt, and phospholip-ids from bile were measured by fluorescent methods as describedpreviously.24 Cholesterol and triglycerides in plasma were deter-mined using colorimetric enzymatic kits from Biolabo (Maizy,France). Plasma cholesterol concentrations in the main lipoproteinclasses were determined in a pool of plasma of each group (5 mice)separated by high-performance gel filtration chromatography asdescribed before.25
Statistical AnalysisValues presented in the figures represent mean�SEM. Statisticalsignificance between control and treated groups was determined byANOVA (*P�0.05; **P�0.01).
ResultsAMP-DNM Treatment Inhibits Atherosclerosisand Ameliorates Hyperlipidemia in APOE*3Leiden MiceTo investigate the effect of AMP-DNM on lesion forma-tion, APOE*3 Leiden mice were treated for 18 weeks witha high-cholesterol diet (1% weight/weight) supplementedwith AMP-DNM to obtain doses of 0, 50, and 100 mg ofAMP-DNM � kg bw�1 � day�1. A repeated dose of 100mg � kg bw�1 � day�1 of AMP-DNM is well tolerated anddoes not exert liver toxicity in mice.12 In the APOE*3Leiden mice, the drug did decrease food consumption from(2.4�0.3 g of diet � mouse�1 � day�1 at 0 mg of AMP-DNM versus 2.2�0.3 g of diet at 50 mg to 2.0�0.3 g ofdiet at 100 mg) (Supplemental Figure IA, available onlineat http://atvb.ahajournals.org). At the end of the treatment,the weight of the 2 treated groups was significantly lowerthan that of the control group (24.1�0.6 g at 0 mg ofAMP-DNM versus 21.3�0.3 g at 50 mg versus 18.9�0.2 g at 100 mg) (Supplemental Figure IB). Quantificationof the lesions was performed after staining of the lipiddeposits in the aortic sinus with Oil Red O (Figure 1A and1B). AMP-DNM treatment had a spectacular inhibitoryeffect on the development of atherosclerosis in this model.Mice receiving the dose of 50 mg of AMP-DNM showeda reduction of 86% in the amount of lesions compared to
932 Arterioscler Thromb Vasc Biol May 2010
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

control animals. At 100 mg of AMP-DNM, no lesions weredetectable in the aortic sinus of the mice.
To investigate the mechanism by which AMP-DNMlimited atherogenesis, we first studied the effects ofthe iminosugar on plasma lipids. AMP-DNM dose-dependently lowered plasma cholesterol (Figure 1C) aswell as triglyceride concentrations (Figure 1D). This effectof AMP-DNM was observed from 6 weeks of administra-tion and was stable over time, and was reflected in animproved lipoprotein profile, that is, a reduction of cho-lesterol in the very low density lipoprotein/low-densitylipoprotein fraction (Figure 1E).
The Table depicts the effect of AMP-DNM treatment onplasma GSL content. The high-cholesterol diet withoutAMP-DNM induced a 10-fold increase of plasma GlcCerand a 3-fold increase of Cer. At the end of the experimentalperiod, the mice treated with AMP-DNM showed a dose-dependent reduction of the amount of GlcCer and a
concomitant reduction of the amount of Cer (Table). Theamount of sphingomyelin determined in a pool of plasmaalso decreased at the end of the treatment: 68% and 83% ofreduction with 50 and 100 mg of AMP-DNM, respectively(data not shown).
AMP-DNM Treatment Lowers Lipids in the Liverof APOE*3 Leiden MiceNext, we focused on the effect of AMP-DNM on hepatic lipidmetabolism. As expected, by inhibiting GlcCer synthase,AMP-DNM dose-dependently lowered the amount of hepaticGlcCer without a change in the amount of Cer (Table). Thecholesterol and triglyceride contents were also decreasedsignificantly in the liver of treated animals. Cholesteroldecreased in a dose-dependent manner (Figure 2A), whereasthe reduction in triglycerides was similar for the 2 doses ofAMP-DNM (Figure 2B). No effect on hepatic choline-basedphospholipids was observed (Figure 2C).
As a consequence of lowered hepatic cholesterol levels,26
expression of 3-hydroxy-3-methylglutaryl coenzyme A re-ductase (5.1-fold) and one of its main regulators, SREBP-2(2-fold), were increased. In accordance, the expression ofLXR, ABCA1, ABCG5, and ABCG8 were decreased,whereas the expression of the very low density lipoproteinreceptor and the LDLR were upregulated (SupplementalFigure IIA).
AMP-DNM Treatment Increases CholesterolOutput in APOE*3 Leiden MiceBecause AMP-DNM treatment strongly decreased choles-terol in plasma and in livers of the treated animals, wehypothesized that AMP-DNM may have increased biliarylipid secretion and total sterol output. At the end of thetreatment period, we therefore collected hepatic bile for 15minutes. AMP-DNM treatment induced an increase of cho-lesterol secretion from 5.0�0.3 nmol � min�1 � 100 g bw�1 at0 mg of AMP-DNM to 8.8�0.6 at 100 mg of AMP-DNM(Figure 3A), and bile salt (Figure 3B), phospholipid secretion(Figure 3C), and bile flow (Figure 3D) increased in a similarfashion. Interestingly, despite the high load of cholesterol inthe diet, the secretion of fecal neutral sterol increased from182 �mol � 24 hour�1 � 100 g bw�1 in the control group to232 �mol � 24 hour�1 � 100 g bw�1 with AMP-DNM treat-
Table. Plasma and Liver GlcCer and Cer Concentrations inAPOE*3 Leiden Mice Fed With a High-Cholesterol Diet Withoutor With AMP-DNM at the Indicated Doses for 18 Weeks
Plasma (nmol � mL�1) Liver (nmol � g�1 liver)
GlcCer Cer GlcCer Cer
Pretreatment 4.1�0.6 8.0�1.8
Control 41.6�4.7 22.1�2.6 35.1�1.9 97.8�7.3
50 mg 13.1�1.1* 8.3�2.5* 26.1�2.4* 120.9�18.2
100 mg 3.0�0.5* 3.3�0.4* 19.7�2.0* 116.1�13.0
Data are expressed as mean�SEM of 5 mice. Pretreatment values weredetermined in 3 different pools of plasma collected before the start of the diet.
*P�0.01.
Figure 1. AMP-DNM treatment inhibits the development of ath-erosclerosis and ameliorates hyperlipidemia in APOE*3 Leidenmice. APOE*3 Leiden mice were treated as described in Materi-als and Methods. A, Representative photomicrographs of OilRed O–stained fatty streaks in the aortic root of the indicatedmice (magnification �5). B, Quantitative analysis of atheroscle-rosis lesion size. C and D, Plasma concentration of cholesterol(C) and triglycerides (D) before the treatment (pre) and at theindicated time points. Data are expressed as mean�SEM;n�10. **P�0.01. E, Distribution of cholesterol over the differentlipoprotein fractions determined in a pool of plasma (5 mice) foreach group. CTRL indicates control.
Bietrix et al Lowering GSL Synthesis Reduces Atherosclerosis 933
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

ment (Figure 4A). This increase was not accompanied by adecrease of cholesterol uptake in the intestine, as determinedwith the fecal dual-isotope ratio method (Figure 4B).
AMP-DNM Treatment Strongly AffectsAtherosclerosis Development in LDLR�/� MiceTo confirm the effect of AMP-DNM on atherosclerotic lesionformation, we performed a similar series of experiments inLDLR�/� mice. Female LDLR�/� mice were fed aWestern-type diet (0.25% cholesterol) supplemented with thesame amount of AMP-DNM to obtain the doses of 0, 50, and100 mg of AMP-DNM � kg bw�1 � day�1. Although both thediet-induced lesions and the diet-induced changes in plasma
lipids were more significant in the LDLR�/� mice than inthe APOE*3 Leiden mice, also in this model AMP-DNMinhibited lesion formation (Figure 5A and 5B). Drug treat-ment resulted in a reduction of plasma cholesterol (Figure5C) and triglycerides (Figure 5D) and an amelioration in thelipoprotein profile (Figure 5E).
DiscussionThe present study reveals that AMP-DNM, an inhibitor ofGSL synthesis, dramatically reduces plasma lipid concentra-
Figure 2. AMP-DNM treatment lowersplasma triglycerides and cholesterol in theliver. The content of cholesterol (A), tri-glycerides (B), and phospholipids (C) wasdetermined in the liver of the animals atthe end of the experiment. Data areexpressed as mean�SEM; n�10.**P�0.01. CTRL indicates control.
Figure 3. AMP-DNM treatment stimulates bile lipids secretion.Biliary secretion of cholesterol (A), bile salts (B), and phospholip-ids (C) was determined after bile collection for 15 minutes asdescribed in Materials and Methods. Bile flow (D) was deter-mined assuming a density of 1 g � mL�1. Data are expressed asmean�SEM; n�10. **P�0.01. CTRL indicates control.
Figure 4. AMP-DNM treatment stimulates fecal neutral steroloutput without affecting cholesterol uptake. A, Analysis of thesterol content in 24 hours of feces. Mean values of 2 cages (5mice per cage) are indicated by the horizontal bars. The deter-mination of neutral sterols had a coefficient of variation of lessthan 5%. B, Percentage of cholesterol absorption calculatedwith the fecal dual-isotope ratio method21 after 6 weeks ofAMP-DNM treatment. Mean values of 2 cages (2 or 3 mice percage) are indicated by the horizontal bars. CTRL indicatescontrol.
934 Arterioscler Thromb Vasc Biol May 2010
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

tion and atherosclerosis in APOE*3 Leiden mice. Thisbeneficial effect of AMP-DNM was observed in APOE*3Leiden mice and in LDLR-deficient mice. In both models, weobserved a strong dose-dependent reduction of plasma cho-lesterol and triglycerides in combination with a dramaticreduction in the development of atherosclerotic lesions.Despite the addition of large amounts of cholesterol to thediet, in APOE*3 Leiden mice, plasma cholesterol remainedclose to wild type values with the dose of 100 mg ofAMP-DNM, and plasma cholesterol did not rise above thethreshold necessary to induce the formation of lesions.27 Itshould be noted that AMP-DNM treatment of APOE*3Leiden mice reduced food consumption and body weightparticularly, at the highest dose. This effect was not observedin all mouse models. In fact, with the dose of 50 mg ofAMP-DNM, despite a strong reduction of plasma cholesterol,the food consumption of the LDLR�/� mice remainedsimilar to what was observed for the control group during thedosing period (Supplemental Figure IIIA). It therefore seems
very unlikely that the beneficial effects of AMP-DNM can befully attributed to reduced food intake.
Although the ability of AMP-DNM to lower plasmacholesterol levels seems pivotal in the reduction of lesiondevelopment, other effects of AMP-DNM have to be consid-ered. The decrease of Cer and sphingomyelin in the plasma oftreated animals may play an additional role in the inhibitionof atherosclerosis. Leventhal et al have shown that cholesterolefflux is impaired in acid sphingomyelinase-deficient macro-phages.28 Levels of sphingomyelin in plasma correlate withthe occurrence of coronary heart disease in clinical studies,29
and Cer is involved in lipoprotein aggregation and foam cellformation.30,31 Reduction in both Cer and sphingomyelin isnot expected to occur as the result of inhibition of GlcCersynthase. In fact, the ratio of cholesterol to Cer and the ratioof cholesterol to sphingomyelin in the plasma of the differentgroups did not change; they remained stable despite theextreme modulation of plasma cholesterol. This suggests thata combined elimination of excessive cholesterol, Cer, andsphingomyelin occurs with AMP-DNM treatment. The mech-anism underlying this intriguing phenomenon has not yetbeen elucidated.
It is generally accepted that inflammatory processes playan important role in atherogenesis. AMP-DNM has beenshown to convey a potent antiinflammatory effect in animalmodels of inflammatory bowel disease induced by chronicmacrophage activation,14 and our group also reported abeneficial effect of AMP-DNM in mouse models of themetabolic syndrome, a disease characterized by chroniclow-grade inflammation.15,32 In the APOE*3 Leiden micestudied here, AMP-DNM treatment also decreased tumornecrosis factor-� and monocyte-chemoattactant protein-1(MCP-1/CCL2) expression (Supplemental Figure II), andtherefore its antiinflammatory action may have contributed tothe strong antiatherogenic effect of AMP-DNM. Similareffects were noted in LDLR�/� mice; we observed areduction of 40% and 70% of plasma CCL2 levels in animalstreated with 50 and 100 mg of AMP-DNM, respectively (datanot shown).
An important observation in our study was the increase ofbile salt and cholesterol secretion in bile of AMP-DNMtreated animals. A concomitant increase in the excretion ofneutral sterol in the feces was observed. It should be notedthat when extrapolated to 24 hours, the stimulation of biliarycholesterol secretion could explain at most 30% of theincrease in neutral fecal sterol excretion. The phenotype ofanimals treated with AMP-DNM resembles that of micetreated with ezetimibe, a potent inhibitor of intestinal choles-terol uptake.33 In lean animals, ezetimibe induces a 6-foldincrease of neutral sterol in the feces.34 In apoE�/� mice,ezetimibe induces a massive reduction in plasma cholesteroland a strong inhibition of the development of atherosclero-sis.35 Thus, to determine whether AMP-DNM exerted anezetimibe-like effect, we used the fecal dual-isotope ratiomethod to determine the rate of cholesterol absorption. Noeffect on intestinal cholesterol uptake after treatment withAMP-DNM was found. In fact, the absorption seemed to beslightly increased with the dose of 100 mg of AMP-DNM. Inaddition the expression of NPC1L1, the main protein respon-
Figure 5. AMP-DNM treatment inhibits the development of ath-erosclerosis and ameliorates hyperlipidemia in LDLR�/� mice.LDLR�/� mice were treated as described in Materials andMethods. A, Representative photomicrographs of Oil RedO–stained fatty streaks in the aortic root of the indicated mice(magnification �5). B, Quantitative analysis of atheroscleroticlesion size. Data represent mean�SEM for at least 5 mice ineach group. **P�0.01. The concentrations of cholesterol (C) andtriglyceride (D) were determined in the plasma of the indicatedmice at the end of the treatment. Data are expressed asmean�SEM of 10 mice in each group. E, Distribution of thecholesterol over the different lipoprotein fractions determined ina pool of plasma (5 mice) for each group. CTRL indicatescontrol.
Bietrix et al Lowering GSL Synthesis Reduces Atherosclerosis 935
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

sible for cholesterol uptake in the intestine,36 was not mod-ulated by the inhibitor (Supplemental Figure IIB).
Because neither inhibition of cholesterol uptake in theintestine and increase in biliary secretion of cholesterol couldaccount for the excess in fecal sterol excretion observed withAMP-DNM, it may have been caused by an increase intransintestinal cholesterol excretion.37 As yet, individual pro-teins involved in transintestinal cholesterol excretion have notbeen identified. The 2 half transporters ABCG5 and ABCG8are known to be involved in cholesterol efflux from thehepatocyte to the biliary lumen, as well as in cholesterolefflux from the enterocyte to the intestinal lumen. AMP-DNM treatment did not increase expression of these trans-porters in the intestine or liver. It remains uncertain at presentwhether ABCG5 and ABCG8 are involved in the AMP-DNMinduced cholesterol excretion.
The results of our study may suggest the use of GlcCersynthase inhibitors for prevention or treatment of atheroscle-rosis. Pharmacological lowering of GSLs by inhibition of theGlcCer synthase is a valid therapeutic approach. A compoundstructurally related to AMP-DNM and also an inhibitor of theGlcCer synthase, Zavesca (1,5-(butylimino)-1,5-dideoxy-D-glucitol), is already registered for treatment of Gaucherdisease.38 In addition, another well-tolerated inhibitor ofGSLs biosynthesis (Genz-112638) is presently being devel-oped as oral therapy for Gaucher disease.39 Our investigationindicates that additional studies are warranted regardingthe use of GSL biosynthesis inhibitors for cardioprotectivetherapy.
In conclusion, our study points to the close association ofthe metabolism of GSLs, lipoprotein metabolism, and athero-genesis. We clearly demonstrate that AMP-DNM, by inhib-iting the biosynthesis of GSLs, profoundly lowers plasmalipids by promoting reverse cholesterol transport and bilecholesterol secretion, and it most likely stimulates transintes-tinal cholesterol excretion. AMP-DNM thereby protectsagainst atherosclerosis development in mouse models closelyrelated to human pathology.
AcknowledgmentsWe are grateful to Han Levels for helping with the lipoprotein profileand to Renze Boverhof and Frans Stellaard for helping with themeasurement of neutral sterol.
Sources of FundingThis work was supported by the Netherlands Heart Foundation, grant2007B030. F.B. was supported by the Fondation pour la RechercheMedicale (SPE20061108543).
DisclosuresNone.
References1. Chatterjee SB, Dey S, Shi WY, Thomas K, Hutchins GM. Accumulation
of glycosphingolipids in human atherosclerotic plaque and unaffectedaorta tissues. Glycobiology. 1997;7:57–65.
2. Dawson G, Kruski AW, Scanu AM. Distribution of glycosphingolipids inthe serum lipoproteins of normal human subjects and patients with hypo-and hyperlipidemias. J Lipid Res. 1976;17:125–131.
3. Garner B, Priestman DA, Stocker R, Harvey DJ, Butters TD, Platt FM.Increased glycosphingolipid levels in serum and aortae of apolipoproteinE gene knockout mice. J Lipid Res. 2002;43:205–214.
4. Garner B, Mellor HR, Butters TD, Dwek RA, Platt FM. Modulation ofTHP-1 macrophage and cholesterol-loaded foam cell apolipoprotein Elevels by glycosphingolipids. Biochem Biophys Res Commun. 2002;290:1361–1367.
5. Glaros EN, Kim WS, Quinn CM, Wong J, Gelissen I, Jessup W, GarnerB. Glycosphingolipid accumulation inhibits cholesterol efflux via theABCA1/apolipoprotein A-I pathway: 1-phenyl-2-decanoylamino-3-morpholino-1-propanol is a novel cholesterol efflux accelerator. J BiolChem. 2005;280:24515–24523.
6. Gong N, Wei H, Chowdhury SH, Chatterjee S. Lactosylceramide recruitsPKCalpha/epsilon and phospholipase A2 to stimulate PECAM-1expression in human monocytes and adhesion to endothelial cells. ProcNatl Acad Sci U S A. 2004;101:6490–6495.
7. Park TS, Panek RL, Rekhter MD, Mueller SB, Rosebury WS, RobertsonA, Hanselman JC, Kindt E, Homan R, Karathanasis SK. Modulation oflipoprotein metabolism by inhibition of sphingomyelin synthesis in ApoEknockout mice. Atherosclerosis. 2006;189:264–272.
8. Glaros EN, Kim WS, Quinn CM, Jessup W, Rye KA, Garner B. Myriocinslows the progression of established atherosclerotic lesions in apoli-poprotein E gene knockout mice. J Lipid Res. 2008;49:324–331.
9. Aerts JM, Hollak C, Boot R, Groener A. Biochemistry of glycosphin-golipid storage disorders: implications for therapeutic intervention. PhilosTrans R Soc Lond B Biol Sci. 2003;358:905–914.
10. Allende ML, Proia RL. Lubricating cell signaling pathways with gan-gliosides. Curr Opin Struct Biol. 2002;12:587–592.
11. Aerts JM, Ottenhoff R, Powlson AS, Grefhorst A, van EM, DubbelhuisPF, Aten J, Kuipers F, Serlie MJ, Wennekes T, Sethi JK, O’Rahilly S,Overkleeft HS. Pharmacological inhibition of glucosylceramide synthaseenhances insulin sensitivity. Diabetes. 2007;56:1341–1349.
12. Bijl N, Sokolovic M, Vrins C, Langeveld M, Moerland PD, Ottenhoff R,van Roomen CP, Claessen N, Boot RG, Aten J, Groen AK, Aerts JM, vanEijk M. Modulation of glycosphingolipid metabolism significantlyimproves hepatic insulin sensitivity and reverses hepatic steatosis in mice.Hepatology. 2009;50:1431–1441.
13. Bijl N, van Roomen CP, Triantis V, Sokolovic M, Ottenhoff R, ScheijS, van Eijk M, Boot RG, Aerts JM, Groen AK. Reduction of glyco-sphingolipid biosynthesis stimulates biliary lipid secretion in mice.Hepatology. 2009;49:637– 645.
14. Shen C, Bullens D, Kasran A, Maerten P, Boon L, Aerts JM, Van AG,Geboes K, Rutgeerts P, Ceuppens JL. Inhibition of glycolipid biosyn-thesis by N-(5-adamantane-1-yl-methoxy-pentyl)-deoxynojirimycinprotects against the inflammatory response in hapten-induced colitis. IntImmunopharmacol. 2004;4:939–951.
15. van Eijk M, Aten J, Bijl N, Ottenhoff R, van Roomen CP, Dubbelhuis PF,Seeman I, Ghauharali-van der Vlugt K, Overkleeft HS, Arbeeny C, GroenAK, Aerts JM. Reducing glycosphingolipid content in adipose tissue ofobese mice restores insulin sensitivity, adipogenesis and reduces inflam-mation. PLoS One. 2009;4:e4723.
16. Overkleeft HS, Renkema GH, Neele J, Vianello P, Hung IO, Strijland A,van der Burg AM, Koomen GJ, Pandit UK, Aerts JM. Generation ofspecific deoxynojirimycin-type inhibitors of the non-lysosomal glucosyl-ceramidase. J Biol Chem. 1998;273:26522–26527.
17. Wennekes T, van den Berg RJ, Donker W, van der Marel GA, StrijlandA, Aerts JM, Overkleeft HS. Development of adamantan-1-yl-methoxy-functionalized 1-deoxynojirimycin derivatives as selective inhibitors ofglucosylceramide metabolism in man. J Org Chem. 2007;72:1088–1097.
18. van den Maagdenberg AM, Hofker MH, Krimpenfort PJ, de Bruijn I, vanVlijmen B, van der Boom H, Havekes LM, Frants RR. Transgenic micecarrying the apolipoprotein E3-Leiden gene exhibit hyperlipoprotein-emia. J Biol Chem. 1993;268:10540–10545.
19. Paigen B, Morrow A, Holmes PA, Mitchell D, Williams RA. Quantitativeassessment of atherosclerotic lesions in mice. Atherosclerosis. 1987;68:231–240.
20. Oude Elferink RP, Ottenhoff R, van Wijland M, Smit JJ, Schinkel AH,Groen AK. Regulation of biliary lipid secretion by mdr2 P-glycoproteinin the mouse. J Clin Invest. 1995;95:31–38.
21. Wang DQ, Carey MC. Measurement of intestinal cholesterol absorptionby plasma and fecal dual-isotope ratio, mass balance, and lymph fistula
936 Arterioscler Thromb Vasc Biol May 2010
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

methods in the mouse: an analysis of direct versus indirect method-ologies. J Lipid Res. 2003;44:1042–1059.
22. Folch J, Lees M, Sloane Stanley GH. A simple method for the isolationand purification of total lipides from animal tissues. J Biol Chem. 1957;226:497–509.
23. Gerhardt KO, Gehrke CW, Rogers IT, Flynn MA, Hentges DJ. Gas-liquidchromatography of fecal neutral steroids. J Chromatogr. 1977;135:341–349.
24. Frijters CM, Ottenhoff R, Van Wijland MJ, Van Nieuwkerk C, GroenAK, Oude Elferink RP. Influence of bile salts on hepatic mdr2 P-glyco-protein expression. Adv Enzyme Regul. 1996;36:351–363.
25. Levels JH, Lemaire LC, van den Ende AE, van Deventer SJ, vanLanschot JJ. Lipid composition and lipopolysaccharide binding capacityof lipoproteins in plasma and lymph of patients with systemic inflam-matory response syndrome and multiple organ failure. Crit Care Med.2003;31:1647–1653.
26. Goldstein JL, Bose-Boyd RA, Brown MS. Protein sensors for membranesterols. Cell. 2006;124:35–46.
27. Groot PH, Van Vlijmen BJ, Benson GM, Hofker MH, Schiffelers R,Vidgeon-Hart M, Havekes LM. Quantitative assessment of aortic athero-sclerosis in APOE*3 Leiden transgenic mice and its relationship to serumcholesterol exposure. Arterioscler Thromb Vasc Biol. 1996;16:926–933.
28. Leventhal AR, Chen W, Tall AR, Tabas I. Acid sphingomyelinase-deficient macrophages have defective cholesterol trafficking and efflux.J Biol Chem. 2001;276:44976–44983.
29. Jiang XC, Paultre F, Pearson TA, Reed RG, Francis CK, Lin M, BerglundL, Tall AR. Plasma sphingomyelin level as a risk factor for coronaryartery disease. Arterioscler Thromb Vasc Biol. 2000;20:2614–2618.
30. Williams KJ, Tabas I. The response-to-retention hypothesis of earlyatherogenesis. Arterioscler Thromb Vasc Biol. 1995;15:551–561.
31. Kinnunen PK, Holopainen JM. Sphingomyelinase activity of LDL: a linkbetween atherosclerosis, ceramide, and apoptosis? Trends CardiovascMed. 2002;12:37–42.
32. Wellen KE, Hotamisligil GS. Inflammation, stress, and diabetes. J ClinInvest. 2005;115:1111–1119.
33. Clarenbach JJ, Reber M, Lutjohann D, von BK, Sudhop T. The lipid-lowering effect of ezetimibe in pure vegetarians. J Lipid Res. 2006;47:2820–2824.
34. Vrins CL, Van der Velde AE, Van den Oever K, Levels JH, Huet S,Oude Elferink RP, Kuipers F, Groen AK. PPARd activation leads toincreased trans intestinal cholesterol efflux. J Lipid Res. 2009;50:2046 –2054.
35. Davis HR Jr, Compton DS, Hoos L, Tetzloff G. Ezetimibe, a potentcholesterol absorption inhibitor, inhibits the development of atheroscle-rosis in ApoE knockout mice. Arterioscler Thromb Vasc Biol. 2001;21:2032–2038.
36. Altmann SW, Davis HR Jr, Zhu LJ, Yao X, Hoos LM, Tetzloff G, IyerSP, Maguire M, Golovko A, Zeng M, Wang L, Murgolo N, Graziano MP.Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterolabsorption. Science. 2004;303:1201–1204.
37. van der Velde AE, Vrins CL, van den Oever K, Kunne C, Oude ElferinkRP, Kuipers F, Groen AK. Direct intestinal cholesterol secretion con-tributes significantly to total fecal neutral sterol excretion in mice. Gas-troenterology. 2007;133:967–975.
38. McEachern KA, Fung J, Komarnitsky S, Siegel CS, Chuang WL, HuttoE, Shayman JA, Grabowski GA, Aerts JM, Cheng SH, Copeland DP,Marshall J. A specific and potent inhibitor of glucosylceramide synthasefor substrate inhibition therapy of Gaucher disease. Mol Genet Metab.2007;91:259–267.
39. Elstein D, Dweck A, Attias D, Hadas-Halpern I, Zevin S, Altarescu G,Aerts JF, van WS, Zimran A. Oral maintenance clinical trial withmiglustat for type I Gaucher disease: switch from or combination withintravenous enzyme replacement. Blood. 2007;110:2296 –2301.
Bietrix et al Lowering GSL Synthesis Reduces Atherosclerosis 937
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
1
SUPPLEMENTAL MATERIALS
Supplemental methods
Determination of mRNA levels
Total RNA was isolated from approximately 100 mg of liver tissue or from the first 10 cm of
the proximal intestine of APOE*3 Leiden mice using Trizol reagent according to the
manufacturer's protocol (Invitrogen, Breda, The Netherlands). Purified RNA was treated with
RQ1 Rnase-free Dnase (1 units/2 µg of total RNA, Promega, Leiden, The Netherlands) and
reverse transcribed with SuperScript II Reverse Transcriptase and random hexamers
(Invitrogen, Breda, The Netherlands) according to protocols supplied by the manufacturer.
Gene expression analysis was performed on a Bio-Rad MyIQ Single-color Real-Time PCR
Detection System by using the Bio-Rad IQ SYBR Green Supermix (Bio-Rad Laboratories
Inc., Hercules, CA). Acidic ribosomal phosphoprotein (36B4) was used as standard
housekeeping genes.
List of primers used for real time PCR.
forward reverse
PPARα TGAACAAAGACGGGATG TCAAACTTGGGTTCCATGAT
LXR TCAGCATCTTCTCTGCAGACCGG TCATTAGCATCCGTGGGAACA
FXR ACGAAGATCAGATTGCTTTGCTC CTCCGCCGAACGAAGAAAC
SREBP-2 TGAAGCTGGCCAATCAGAAAA ACATCACTGTCCACCAGACTGC
HMGCoAr TCTGGCAGTCAGTGGGAACTATT CCTCGTCCTTCGATCCAATTT
SREBP-1 GACCTGGTGGTGGGCACTGA AAGCGGATGTAGTCGATGGC
SCD1 TACTACAAGCCCGGCCTCC CAGCAGTACCAGGGCACCA
FAS CGAACCTGGCTGCCTACTAC GAGCCGTCAAACAGGAAGAG
MTP TCAAGAGAGGCTTGGCTAGCTT GGCCTGGTAGGTCACTTTACAATC
Cyp7A1 TGAACAAAGACGGGATG TCAAACTTGGGTTCCATGAT
BSEP TGGAAAGGAATGGTGATGGG CAGAAGGCCAGTGCATAACAGA
MDR-2 AGGCAGCGAGAAACGGAAC TGGTTGCTGATGCTGCCTAG
ABCA1 GGTTTGGAGATGGTTATACAATAGTTGT TTCCCGGAAACGCAAGTC
ABCG5 TGGCCCTGCTCAGCATCT ATTTTTAAAGGAATGGGCATCTCTT
ABCG8 CCGTCGTCAGATTTCCAATGA GGCTTCCGACCCATGAATG
SR-BI GGCTGCTGTTTGCTGCG GCTGCTTGATGAGGGAGGG
LDL-R CTGTGGGCTCCATAGGCTATCT GCGGTCCAGGGTCATCTTC
LRP TGGGTCTCCCGAAATCTGTT ACCACCGCATTCTTGAAGGA
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
2
VLDL-R GGCTCTGGCGAGTGCATT GCCGTCCTTGCAGTCAGG
F4/80 CTTTGGCTATGGGCTTCCAGTC GCAAGGAGGACAGAGTTTATCGTG
CCL-2 AGGTCCCTGTCATGCTTCTGG CTGCTGCTGGTGATCCTCTTG
TNFα ACGGCATGGATCTCAAAGAC AGATAGCAAATCGGCTGACG
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from
SUPPLEMENTAL MATERIAL
3
SUPPLEMENTAL FIGURES
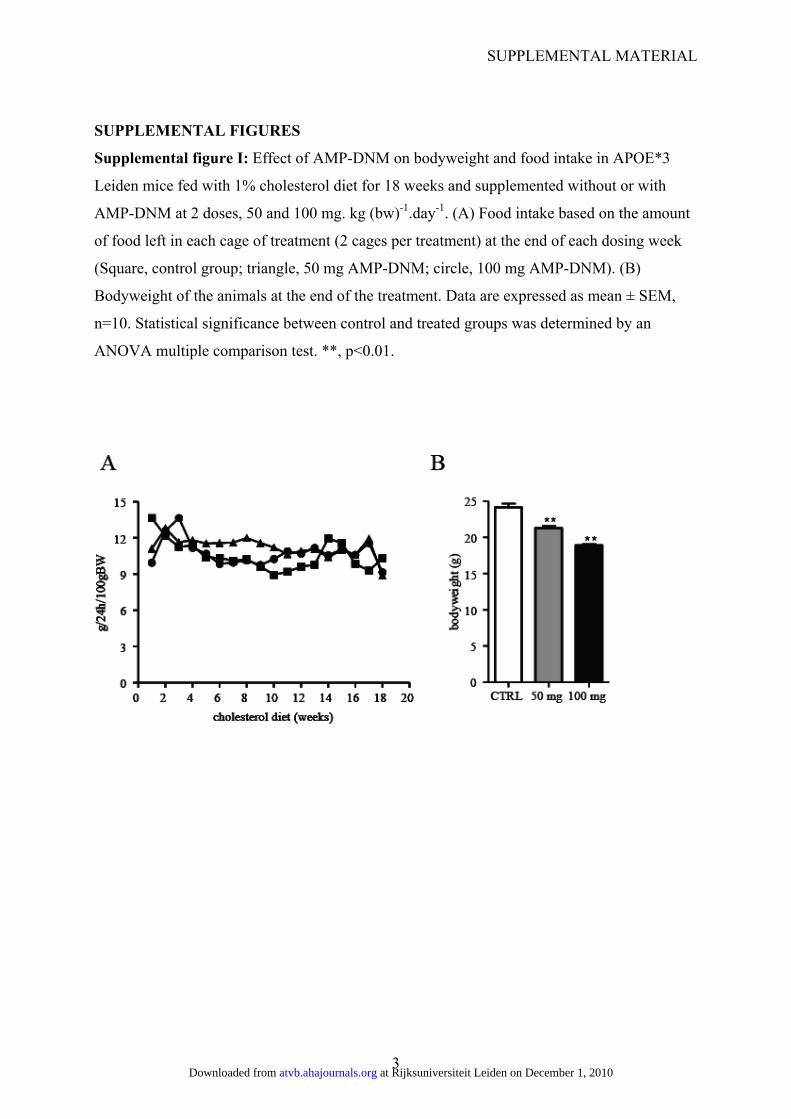
Supplemental figure I: Effect of AMP-DNM on bodyweight and food intake in APOE*3
Leiden mice fed with 1% cholesterol diet for 18 weeks and supplemented without or with
AMP-DNM at 2 doses, 50 and 100 mg. kg (bw)-1.day-1. (A) Food intake based on the amount
of food left in each cage of treatment (2 cages per treatment) at the end of each dosing week
(Square, control group; triangle, 50 mg AMP-DNM; circle, 100 mg AMP-DNM). (B)
Bodyweight of the animals at the end of the treatment. Data are expressed as mean ± SEM,
n=10. Statistical significance between control and treated groups was determined by an
ANOVA multiple comparison test. **, p<0.01.
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
4
Supplemental figure II: Effect of AMP-DNM on expression of key genes involves in lipid
homeostasis and inflammation in the liver (A) and the proximal intestine (B) of APOE3*
Leiden mice. mRNA was prepared from individual mice (n = 5 per group) and data are
presented as means of 5 animals performed in duplicate ± SEM. Expression values are
normalized to 36B4 and expression values in the control group were set as 1.00. Statistical
significance between control and treated groups was determined by an ANOVA multiple
comparison test. *, p<0.05; **, p<0.01.
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
5
Supplemental figure III: Effect of AMP-DNM on plasma lipids in LDLR -/- mice fed with
0.25 % cholesterol diet for 12 weeks and supplemented without or with AMP-DNM at 2
doses, 50 and 100 mg. kg (bw)-1.day-1. (A) Food intake based on the amount of food left in
each cage of treatment (2 cages per treatment) at the end of each dosing week (Square, control
group; triangle, 50 mg AMP-DNM; circle, 100 mg AMP-DNM). (B) Bodyweight of the
animals at the end of the treatment. Data are expressed as mean ± SEM, n=10. Statistical
significance between control and treated groups was determined by an ANOVA multiple
comparison test. **, p<0.01.
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
6
SUPPLEMENTAL TABLES
Supplemental table I: Concentration of plasma glucosylceramide (GlcCer) and ceramide
(Cer) in LDLR -/- mice fed with 0.25 % cholesterol diet for 12 weeks and supplemented
without or with AMP-DNM at 2 doses, 50 and 100 mg. kg (bw)-1.day-1. Data are expressed as
mean ± SEM, n=5. Statistical significance between control and treated groups was determined
by an ANOVA multiple comparison test. *, p<0.05; **, p<0.01.
Control 50 mg 100 mg
GlcCer
(nmol.ml-1) 117.8±9.0 27.1±5.0** 7.2±0.7**
Cer
(nmol.ml-1) 47.7±5.5 28.0±5.1* 10.7±2.4**
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
7
Supplemental table II: Concentration of lipids in liver of LDLR -/- mice fed with 0.25 %
cholesterol diet for 12 weeks and supplemented without or with AMP-DNM at 2 doses, 50
and 100 mg. kg (bw)-1.day-1. Data are expressed per gram of tissue as mean ± SEM (n=5) for
glucosylceramide (GlcCer) and ceramide (Cer) levels and as mean ± SEM (n=10) for
cholesterol and triglyceride levels. Statistical significance between control and treated groups
was determined by an ANOVA multiple comparison test. *, p<0.05; **, p<0.01.
Control 50 mg 100 mg
GlcCer
(nmol.g-1 liver) 74.6±9.2 27.8±1.8** 28.1±4.4**
Cer
(nmol.g-1 liver) 149.6±20.6 225.2±30.9 262.8±23.6*
cholesterol
(μmol.g-1 liver) 11.1±0.5 9.5±0.7 7.9±1.0*
triglycerides
(μmol.g-1 liver) 5.6±0.5 4.0±0.3* 3.6±0.4**
phospholipids
(μmol.g-1 liver) 9.2±0.4 8.8±0.1 8.6±0.6
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from

SUPPLEMENTAL MATERIAL
8
Supplemental table III: Bile secretion in LDLR -/- mice fed with 0.25 % cholesterol diet for
12 weeks and supplemented without or with AMP-DNM at 2 doses, 50 and 100 mg. kg (bw)-
1.day-1. Bile was collected for 15 min and biliary lipids were determined as described in the
materials and methods. Data are expressed as mean ± SEM, n=5. **, p<0.01.
Bile
control 50 mg 100 mg
bile flow
(μl.min-1.100g bw-1) 3.6±0.8 9.2±1.2** 5.6±0.3
cholesterol
(nmol.min-1.100g bw-1) 1.2±0.3 3.7±0.4** 2.5±0.4
bile salt
(nmol.min-1.100g bw-1) 253.0±57.6 620.6±87.7** 376.1±47.6
phospholipids
(nmol.min-1.100g bw-1) 36.2±8.1 93.3±11.3** 60.2±5.0
at Rijksuniversiteit Leiden on December 1, 2010 atvb.ahajournals.orgDownloaded from




















