
Influence of Temporal Overlap on Time Course of the Simon Effect
32
Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 1 Influence of Temporal Overlap on Time Course of the Simon Effect Giulia Baroni Antonello Pellicano Luisa Lugli Roberto Nicoletti Robert W. Proctor Correspondence to: Giulia Baroni ([email protected] ) Department of Communication Studies, University of Bologna Via A. Gardino 23, 40122 Bologna, Italy
-
Upload
rwth-aachen -
Category
Documents
-
view
1 -
download
0
Transcript of Influence of Temporal Overlap on Time Course of the Simon Effect

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 1
Influence of Temporal Overlap on Time Course of the Simon Effect
Giulia Baroni
Antonello Pellicano
Luisa Lugli
Roberto Nicoletti
Robert W. Proctor
Correspondence to:
Giulia Baroni ([email protected])
Department of Communication Studies, University of Bologna
Via A. Gardino 23, 40122 Bologna, Italy

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 2
Abstract
Two experiments are reported in which we manipulated relevant and irrelevant stimulus dimensions
to assess whether an increase in temporal overlap would influence the time course of a “standard”
Simon effect (obtained when visual stimuli are presented on the left/right of the screen and left/right
responses are performed with uncrossed hands). This procedure is new in two ways: First, the
manipulations were intended to reduce, instead of increase, the distance between conditional and
unconditional response-activation processes. Second, we manipulated the relevant and irrelevant
stimulus dimensions in a manner that did not vary stimulus onset asynchronies (SOAs), precues or
go/no go trials, or alter the stimulus quality. Results were consistent with the hypothesis that when
the two response processes are shifted closer to each other, the Simon effect would be sustained
across time, instead of decreasing as typically found. These findings are discussed in line with the
temporal overlap hypothesis and with an automatic activation account.
Keywords: Simon effect, temporal overlap, time-course analysis, conditional accuracy function,
automatic activation.

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 3
Introduction
In the Simon task, participants are to respond to the colour or shape of a stimulus (i.e., the
relevant dimension) while ignoring its spatial position (i.e., the irrelevant dimension). The Simon
effect refers to the phenomenon wherein the spatial position of the stimulus, despite being
irrelevant, influences performance: Responses are faster and more accurate when the stimulus and
response locations correspond to each other than when they do not (see Proctor & Vu, 2006, and
Simon, 1990, for reviews). Thus, if participants are instructed to press a right key to a red stimulus
and a left key to a green stimulus, their reaction times (RTs) will be shorter and accuracy higher
when the red stimulus is on the right (corresponding trials) than when it is on the left (non-
corresponding trials).
Dual-route models (e.g., De Jong, Liang, & Lauber, 1994) are widely used to account for the
Simon effect. According to these models, two different response processes are involved: On the
one hand, the irrelevant spatial position automatically activates an ipsilateral response through an
unconditional route; on the other hand, the relevant feature of the stimulus is translated, following
the task instructions, into the correct response through a conditional route. Performance on
corresponding trials is faster and more accurate because the same response is activated through the
unconditional and conditional routes. Conversely, non-corresponding trials are slower and less
accurate because a conflict occurs between the automatically activated and the voluntarily translated
response.
The time-courses of these two response processes have been investigated through analysis of
the RT distributions (bin analysis; De Jong et al., 1994; see Proctor, Miles, & Baroni, 2011, for a
review). This technique has revealed that the Simon effect can have two different time courses: A
decreasing effect function (i.e., the magnitude of the Simon effect diminishes as RT increases) is
found when the unconditional response activation occurs soon after the stimulus onset and then
dissipates over time (e.g., Proctor, Yamaguchi, Zhang, & Vu, 2009; Rubichi & Pellicano, 2004).
An increasing or constant effect function is observed, though, when the irrelevant response needs

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 4
more time to reach complete activation and thus to exert the maximum influence on performance
(e.g., Vallesi, Mapelli, Schiff, Amodio, & Umiltà, 2005; Wühr, 2006).
To explain these different effect functions, Wascher, Schatz, Kuder, and Verleger (2001)
proposed that the Simon effect can be generated by two different and dissociable mechanisms: A
visuomotor facilitation of same-side responses and a cognitive interference of codes. The decaying
effect function, yielded by the visuomotor process, would be generated by a “natural” spatial
anatomical mapping, that is, when stimuli and responses are both on the horizontal axis and the
hands are placed in an uncrossed position (see Buhlmann, Wascher, & Umiltà, 2007, Experiments 1
and 2; Wascher et al., 2001, Experiment 1). This task setting has come to be prototypical and is
often referred to as “standard”. Conversely, the stable or increasing effect function, yielded by the
cognitive process, would be due to the lack of a “natural” relation between a visual stimulus and
anatomical effector, namely when (a) the task is performed with the hands crossed (Wascher et al.,
2001, Experiment 1), (b) the stimuli are along the vertical axis and are matched, through a fixed
mapping, with left/right responses (Proctor, Vu, & Nicoletti, 2003; Wiegand & Wascher, 2005;
Wiegand & Wascher, 2007), or (c) the stimuli are presented in an auditory modality (Wascher et al.,
2001, Experiment 2).
The focus of the current study is on the dynamics underlying the decreasing time course of
the “standard” Simon effect. De Jong et al. (1994) firstly attributed this distribution pattern to an
automatic, but short lived, response activation. This interpretation has gathered consistent evidence
in behavioural and neuropsychological literature (see Proctor et al., 2011), and it also matches with
the same-side response activation subsequently illustrated by Wascher et al. (2001) for their so-
called visuomotor Simon effect.
The idea of an automatic activation pattern is also supported by Hommel’s (1993) temporal
overlap hypothesis, which postulates that the Simon effect magnitude is strongly influenced by the
temporal distance between codes from the relevant and the irrelevant stimulus dimension. More
precisely, Hommel claimed, “Every experimental manipulation that markedly increases the
temporal distance between the formation of the relevant stimulus code and the irrelevant spatial

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 5
code leads to a smaller Simon effect – that is, to a smaller effect of irrelevant spatial correspondence
between stimulus and responses” (p. 289). In several of Hommel’s experiments, the Simon effect
tended to disappear whenever the coding of the relevant dimension was postponed relative to the
coding of the irrelevant one. This happened, for example, when the code of the relevant dimension
was slowed both through decreasing the signal quality with a pattern mask or a reduction in contrast
relative to the background, as well as when the stimulus built up gradually (i.e., its colour appeared
at different time gaps, also called stimulus onset asynchronies, SOAs). A similar result was also
obtained in a further study (Hommel, 1994b) in which higher stimulus complexity made the
discrimination of the relevant stimulus value less immediate (see also Hommel, 1994a, and Rubichi,
Iani, Nicoletti, & Umiltà, 1997). Lastly, a Simon effect smaller in magnitude was found even when
the response to lateralised stimuli had to be withheld until a go signal occurred (Simon, Acosta,
Mewaldt, & Speidel, 1976; Vallesi & Umiltà, 2009, Experiment 1) or when a spatial cue preceded
the target stimuli (Vallesi & Umiltà, 2009, Experiment 2).
Hence, consistent with the automatic activation account, all the above mentioned results
demonstrate that when the temporal gap between the relevant and irrelevant dimension is increased,
by delaying the relevant dimension or precuing the irrelevant one, the Simon effect decreased. This
result pattern implies that the spatial code had time to dissipate before response selection occurred.
Recently, Miles and Proctor (2009) provided further support for this automatic activation
account and demonstrated that temporal overlap manipulations can influence the time course of the
Simon effect, as well as its magnitude, when central spatial indicators (“left”/ “right” words) are
presented as stimuli. In their Experiment 1 participants were to press a left or right key according to
the colour of the central word, which could be intact (high discriminability condition) or masked
with a noise overlay (low discriminability condition). The former condition showed an increasing
Simon effect, as is typically obtained for location words (e.g., Pellicano, Lugli, Baroni, & Nicoletti,
2009, Experiment 3). However, for the latter condition, in which the colour processing was
disrupted, the Simon effect was nonsignificant overall and showed a flat effect function across the
RT distribution. The authors interpreted these results as consistent with the temporal overlap

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 6
hypothesis: The delay of the conditional process allowed the automatic activation to decay prior to
the conflict arising. This interpretation was extended to the results of Experiment 2, in which the
discriminability of both the relevant and the irrelevant dimensions was reduced in order to delay
both the conditional and unconditional response processes. The Simon effect in this low
discriminability condition was significant and increased across bins, as for the non-disrupted
condition of Experiment 1. The only difference between these manipulations was the position of
the Simon effect distribution function along the abscissa (RT): The function for Experiment 2 was
shifted rightward by more than 100 ms because of the overall higher task complexity.
The aim of the current study was to investigate more deeply how temporal overlap
manipulations can shape the time course of the Simon effect. However, we followed an opposite
rationale with respect to most of the above discussed studies. Our experiments aimed at decreasing,
instead of increasing, the temporal distance between stimulus codes relative to a baseline condition,
thus yielding greater temporal overlap between the resulting response processes. As in Miles and
Proctor’s (2009) study, an automatic activation account was used as the theoretical framework. But
unlike that study, all of our tasks shared a spatial-anatomical mapping between stimuli and
responses. Participants were in fact always faced with lateralised, instead of centrally presented,
stimuli. This difference is not trivial since, according to Wascher et al. (2001), such a “natural” task
set should yield a decreasing time course.
We hypothesized, though, that greater temporal overlap would turn this typical decreasing
effect found for the “standard” Simon task into a sustained one. Indeed, reducing the distance
between the relevant and irrelevant stimulus dimensions would make participants respond during
the time window in which the automatic activation had not yet dissipated. As a consequence,
performance would be influenced by the irrelevant information for a longer time span, thus yielding
a significant Simon effect across the whole RT distribution. To assess this hypothesis, we
conducted two experiments in which the temporal overlap was enhanced through discrete
manipulations to reduce the time to process the relevant stimulus dimension (i.e., Experiment 1,
wide coloured stimuli condition) or increase the time to process the irrelevant stimulus dimension

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 7
(i.e., Experiment 2, arrow stimuli condition). Note that, unlike most of the previously discussed
studies, the temporal proximity was increased without dissociating the two stimulus codes by means
of SOAs, precues, or go/no go signals. Moreover, we also avoided altering the stimulus
discriminability through masking patterns or variations of the stimulus contrast.
Experiment 1 was composed of three conditions, baseline, wide coloured stimuli and wide
unfilled stimuli. The baseline condition aimed at replicating the “standard” Simon settings:
Lateralized coloured squares were used as stimuli and matched with left/right keypress responses
performed by the left and the right hand, respectively. In the wide coloured stimuli condition
participants were faced with the same coloured squares, but larger in size. This manipulation aimed
at facilitating and speeding up the colour discrimination (i.e., the relevant distinction), thus shifting
the conditional component back in time, closer to the automatic one. Finally, in the wide unfilled
stimuli condition, participants were also faced with wide squares, but coloured only along their
borders, instead of their whole area. This condition served as a control for the wide coloured stimuli
condition, in order to disambiguate the possible effects of increased colour information from that of
increased size. Indeed, the stimulus width was kept equal to the wide coloured stimuli condition but
the amount of colour information was set similar to the baseline condition.
Two different results were thus expected for these three conditions: In line with the above
mentioned literature, a decreasing time course was expected for the baseline and also for the wide
unfilled stimuli condition, since the temporal proximity between the relevant and irrelevant stimulus
codes is assumed to be similar. Conversely, in the wide coloured stimuli condition, the increased
temporal overlap was expected to yield a constant or increasing Simon effect.
A sustained time course was also hypothesized for the arrow stimuli condition of
Experiment 2, where the temporal overlap was enhanced, though, by a manipulation on the
irrelevant stimulus dimension. Lateralised arrow signals, similar in width to the baseline stimuli,
were used in fact in place of squares. The spatial code given by the direction of such indicators was
expected to sum up to the one provided by their location in space, leading to a stronger and longer
lasting stimulus spatial code with respect to that of the baseline condition of Experiment 1.

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 8
To summarize, the goal of the wide coloured and arrow stimuli conditions of Experiments 1
and 2, respectively, was to increase the temporal overlap between the two stimulus codes. As a
consequence, a sustained Simon effect function was expected to be found for both conditions,
despite their “standard” task set would typically yield a decaying time course. Results that match
our predictions would provide reliable further evidence that temporal overlap influences the time
course of the Simon effect.
Experiment 1
The experiment included baseline, wide coloured stimuli, and wide unfilled stimuli
conditions, all implemented with a “standard” Simon task: A square was presented to the left or to
the right on the computer screen, and the responses to its colour was performed with uncrossed
hands by pressing one of two buttons placed on the left and right of the body midline. The
difference between the three conditions was in the saliency of the colours (see Figure 1).
The aim of the colour saliency manipulation was as follows. First of all, presenting wide
stimuli filled with colour (i.e. wide coloured stimuli condition) was meant to facilitate and speed up
the colour discrimination task, relative to that of the baseline condition, in order to shift the
selection of the correct response (conditional process) closer to the (fast) activation of the
unconditional one. This goal of a higher degree of temporal overlap had a converse rationale with
respect to Hommel’s (1994a, 1994b) studies (see also Rubichi et al., 1997), in which the stimulus
complexity was enhanced to slow down identification of the relevant dimension. Presenting wide
squares, but only coloured along their borders (i.e., wide unfilled stimuli), should not facilitate the
task-instructed colour discrimination, thus yielding a similar degree of temporal overlap between
stimulus codes to that of baseline.
Our predictions were as follows. For the baseline condition, we expected the Simon effect to
be largest in the shortest RT bins and then to decrease across the longer ones. This result would
replicate the findings shown in the current literature (e.g., Proctor et al., 2009; Rubichi & Pellicano,
2004) and would be consistent with the evidence that such a declining effect function is bound to a
spatial-anatomical mapping between stimuli and responses (Wascher et al., 2001). For the wide

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 9
coloured stimuli condition, the easier colour discrimination was expected to yield overall shorter
RT with respect to the baseline condition and, more importantly, a stable or increasing Simon effect
function. The perceptual manipulation on the relevant stimulus dimension was intended to allow
participants to respond during the time window in which the automatic activation had its maximum
strength and it had not yet dissipated. Finally, in the wide unfilled stimuli condition we
hypothesized that reducing the amount of colour information within the stimulus compared to the
wide coloured stimuli would result in the conditional response process not being shifted closer to
the unconditional one. Consequently, we expected that condition to yield an RT range and a Simon
effect function similar to those of the baseline condition.
To summarize, we hypothesized that, within a “standard” Simon task, a higher degree in
temporal overlap would shift the typically found decreasing time course, into a sustained or
increasing one.
Method
Participants. Thirty-six students from the University of Bologna (24 females; mean age =
20.4 years; SD = 1.8 years) volunteered to participate in the experiment. Twelve took part in the
baseline condition, 12 in the wide coloured stimuli condition and the remaining 12 in the wide
unfilled stimuli one. The participants were all right-handed, had normal or corrected-to-normal
vision, and were naïve as to the purpose of the experiment. They received extra credit for their
participation in the study.
Apparatus and stimuli. The experiment was carried out in a dimly lit and noiseless room.
The participant was seated facing a 17 in. cathode-ray tube screen driven by a 700 MHz processor
computer. His/her head was positioned in an adjustable head-and-chin rest. The screen was
positioned so that stimuli appeared at eye level at a distance of 47 cm. Stimulus selection, response
timing and data collection were controlled by the E-Prime software (www.pstnet.com; Psychology
Software Tools, Inc.)
A fixation cross 1-cm wide and 1-cm high (1.2° x 1.2° of visual angle) was presented at the
centre of the screen against a black background. For the baseline and wide coloured stimuli

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 10
conditions, the stimuli were filled green and red squares presented at the left or right of the fixation
cross. For the wide unfilled stimuli condition, though, the stimuli were black squares with green/red
borders also presented at the left/right screen edges. The square width and height was 0.8 cm (1° x
1°), for the baseline condition, and 4.6 cm (5.6° x 5.6°) for the wide coloured and wide unfilled
stimuli conditions (see Figure 1). The width of the border for the wide unfilled stimuli condition
was 0.4 cm. The stimulus total coloured area for the baseline, wide coloured and wide unfilled
stimuli conditions was: 0.6; 21.2 and 13 cm2, respectively. Stimuli of all three conditions were
centred at a distance from the fixation, being separated by 9.4 cm (11.5° centre to centre).
Responses were made by pressing either the ‘‘Z’’ or the ‘‘M’’ key (left and right keys) on the
computer keyboard with the left or the right index finger, respectively.
Insert Figure 1 about here
Procedure. At the beginning of the experiment, instructions were presented on the screen,
and each participant was required to place the tip of his/her left and right index finger on the left
and right key, respectively. When the participant’s fingers were appropriately placed on the keys,
the experimenter started the experiment by pressing the space bar. The central fixation cross was
presented and then remained visible across the whole experiment. After a 1000-ms intertrial
interval, the stimulus appeared on the left or on the right of the screen and remained visible for 250
ms followed by a 1000-ms blank interval for response collection. Stimulus presentations occurred
according to a random sequence. RT was recorded as the time between the stimulus onset and the
pressing of the response key. The instructions stressed the speed and accuracy of performance.
One-third of the participants was randomly assigned to the baseline condition, another third
to the wide coloured stimuli condition and the remaining third to the wide unfilled stimuli one
(between-subjects design). Within each condition, the instructions were balanced since half of the
participants were instructed to respond to the red colour by pressing the right key and to the green
colour by pressing the left key, whereas the other half were instructed with the reverse assignment.
An error message was given as feedback for omitted (> 1250 ms), anticipated (< 150 ms), and
incorrect responses. The feedback for incorrect responses was also enhanced by a low pitch tone of

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 11
600 ms in duration. Each participant completed a block of 40 practice trials, which was followed by
400 experimental trials divided into 4 blocks of 100 trials each (25 for each stimulus type).
Participants were allowed to take a short break between blocks.
Results
We excluded from the analysis trials for which the RT was more than two standard
deviations smaller (0.1 %) or greater (3.9 %) than the participant’s overall mean RT. Incorrect
responses (4.3 %) were also discarded. Mean correct RTs and arcsin-transformed accuracy rates
(ARs), that is, percentage of correct responses, were analyzed separately.
To measure the Simon effect, corresponding (i.e., the position of the response key
corresponded to the position of the square) and non-corresponding (i.e., the response key did not
correspond to the position of the square) responses for each condition were compared. The time
course of the Simon effect for RT was investigated by applying the Vincentizing procedure
(Ratcliff, 1979). The RT distribution for each participant and correspondence condition was divided
into deciles (bins), and the mean of RT for each decile was calculated. Because the variance was
higher for the last bin than for the others, it was eliminated from the analysis (see, e.g., Buhlmann,
et al., 2007; Wascher et al., 2001). We calculated the size of the Simon effect for each remaining
bin, subtracting the mean RT for the corresponding condition from the mean RT for the non-
corresponding condition.
Mean reaction times. RTs were submitted to a three-factor ANOVA with Bin (1–9) and
Correspondence (corresponding vs. non-corresponding S-R pairings) as within-subjects factors and
Condition (Baseline vs. Wide Coloured Stimuli vs. Wide Unfilled Stimuli) as a between-subjects
factor. It should be noted that, considering the way that the data were grouped, the Bin main effect
necessarily turned out to be significant in all analyses. Therefore, it is not reported and discussed
here or in Experiment 2. When sphericity was violated, the Huynh-Feldt correction was applied,
although the original degrees of freedom are reported.
The Correspondence factor was significant, F(1,33) = 113.43, p < .001, indicating that
corresponding responses were faster than non-corresponding ones (381 vs. 404 ms). The Condition

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 12
factor was close to the .05 level, F(2,33) = 3.13, p = .058, showing a trend for responses to be faster
in the wide coloured stimuli condition (363 ms) than in the baseline and the wide unfilled stimuli
conditions (411 vs. 403 ms, respectively).
The Correspondence × Condition interaction was not significant, F(1,22) < 1, indicating that
the Simon effect was of similar magnitude for the baseline (400 vs. 423 ms), wide coloured stimuli
(351 vs. 374 ms), and wide unfilled stimuli (392 ms vs. 415 ms) conditions (see Table 1).
Insert Table 1 about here
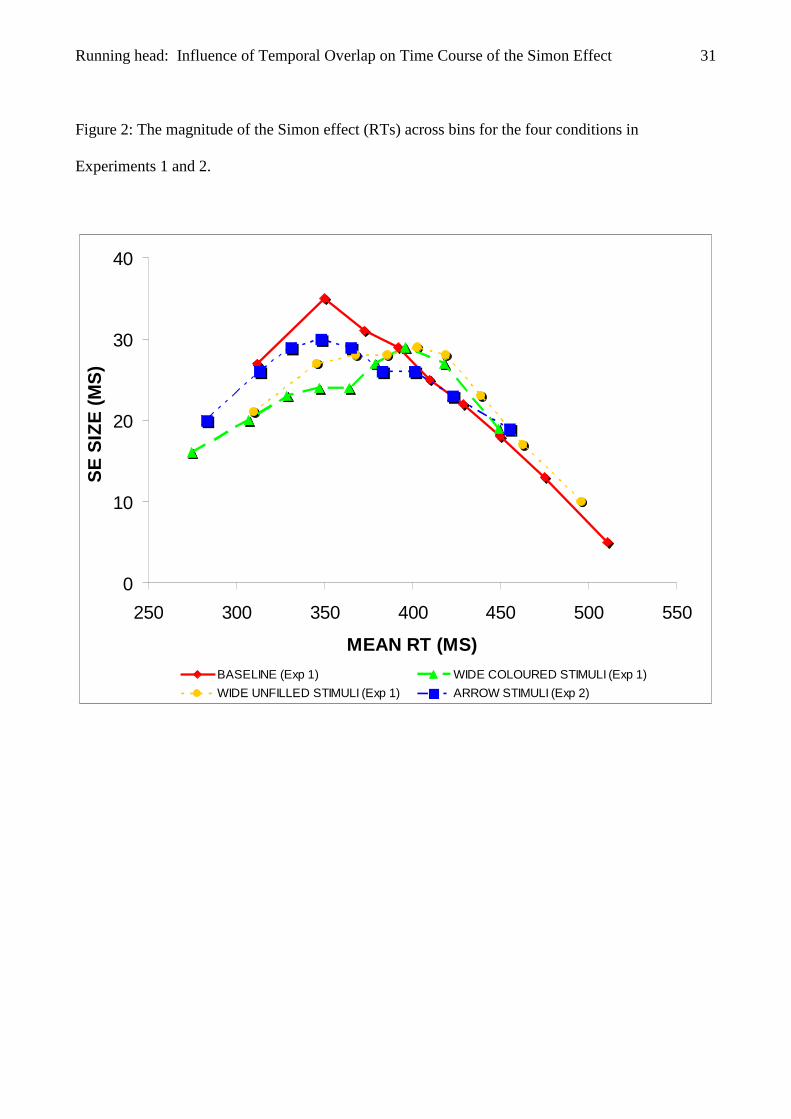
Crucially, the Bin × Correspondence x Condition interaction was significant, though,
F(16,264) = 3.71, p = .005 (see Figure 2). Consequently, we performed two post-hoc ANOVAs to
test the predictions that (a) the baseline condition would yield a similar (i.e., decaying) time course
to that of the wide unfilled stimuli condition and (b) the time course of both these conditions would
differ from that of the wide coloured stimuli condition (i.e., sustained).
First, a three-factor ANOVA with Bin × Correspondence × Condition (Baseline vs. Wide
Unfilled Stimuli) was run. As hypothesized, the baseline and wide unfilled stimuli conditions
yielded similar overall RTs (411 and 403, respectively), as shown by the lack of significance of the
Condition main effect, F(1,22) < 1.0. Moreover, they had similar time courses as shown by the
nonsignificant Condition × Correspondence × Bin interaction, F(8,176) = 1.65, p = .196. In the
baseline condition, paired sample t-tests showed that the Simon effect was significant at all bins
except for the last one, ts(11) > 3.70, ps < .003, and t(11) = 1.1, p = .295, for bins 1 to 8 and for bin
9, respectively. Furthermore, Helmert contrasts showed that the size of the Simon effect did not
differ significantly from bin 1 to bin 2 (27 and 35 ms), F(1,11) = .957, p = .349, but then decreased
significantly from bin 2 to bin 9 (35, 31, 29, 25, 22, 18, 13 and 5 ms , respectively), Fs(1,11) > 8.05,
ps < .02. In the wide unfilled stimuli condition, paired sample t-tests showed that the Simon effect
was also significant at all bins except for the last one, ts(11) > 4.00, ps = .002, and t(11) = 1.80, p =
.099, for bins 1 to 8 and for bin 9, respectively. Helmert contrasts showed that the size of the Simon
effect did not differ significantly from bin 1 to bin 5 (21, 27, 28, 28 and 29 ms), Fs(1,11) > .425; ps
> 0.68, but then decreased significantly from bin 5 to bin 9 (29, 28, 23, 17 and 10 ms, respectively),

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 13
Fs(1,11) > 5.85, ps = .034 (see Figure 2). Thus, significant decreasing effect functions were found
for both baseline and wide unfilled stimuli conditions.
A second three-factor ANOVA compared the time course of the wide coloured stimuli
condition to those of the baseline and wide unfilled stimuli conditions averaged together. As
hypothesized, the Condition × Correspondence × Bin interaction was significant, F(8,272)= 5.74, p
= .003, indicating that while the Simon effect in the baseline and wide unfilled stimuli conditions
had a decreasing time course, the wide coloured stimuli condition yielded a sustained one. This
sustained effect function was confirmed by a Bin × Correspondence ANOVA on the single wide
coloured stimuli condition which displayed a nonsignificant interaction F(8,88) = 1.71, p = .192,
indicating that the size of the Simon effect did not change reliably across bins: It was 16, 20, 23, 24,
24, 27, 29, 27 and 19 ms from bin 1 to bin 9, respectively (see Figure 2).
Insert Figure 2 about here
Accuracy rates (ARs). Arcsine-transformed ARs of each condition were submitted to a
repeated-measures ANOVA with Correspondence and Condition as the within- and between-
subjects factors, respectively. For clarity, the original accuracy rates are reported. When sphericity
was violated, the Huynh-Feldt correction was applied, although the original degrees of freedom are
reported.
The main effect of Correspondence was significant, F(1,33) = 11.60, p = .002, indicating
that corresponding responses were more accurate than non-corresponding ones (96.9 % vs. 94.8%).
Paired sample t tests on the single conditions, indicated that the Simon effect for ARs was
significant for both wide coloured and wide unfilled stimuli conditions (2.7 % vs. 3.3 %), ts(12) >
2.52, p < .03, respectively, but failed to reach significance for the baseline condition, t(12) > 1 (see
Table 1). The Condition factor was not significant, F(2,33) < 1, but the Condition ×
Correspondence interaction approached the .05 level, F(2,33) = 3.17, p = .055. Independent sample
t tests indicated that the Simon effect for ARs was larger for the wide unfilled stimuli than for the
baseline condition, t(22) = 2.45, p = .023, and the wide coloured stimuli condition also tended to
have a larger Simon effect with respect to that of the baseline condition, t(22) = 1.90, p = 0.71.

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 14
Discussion
The results of the time-course investigations confirmed our hypothesis: A decreasing time
course was found for both the baseline and the wide unfilled stimuli conditions and, crucially, a
sustained time course was shown by the wide coloured stimuli condition (see Figure 2). The
baseline result was consistent with current literature (e.g., Proctor et al., 2009) and confirmed the
fast and transient nature of the automatically activated response (De Jong et al., 1994). This
transient activation pattern was later attributed by Wascher et al. (2001) to visuomotor facilitation of
same side responses, depending on an S-R spatial-anatomical mapping. It is worth mentioning that,
to make this condition an effective baseline for the following ones, we used stimuli similar in
dimensions to those of previously conducted studies that used lateralised coloured shapes (e.g., De
Jong et al., 1994; Proctor, Pick, Vu & Anderson, 2005; Proctor et al., 2009; Vu & Proctor, 2008).
Findings from the wide coloured stimuli condition provided evidence that temporal overlap
was an important factor in shaping the effect function of a “standard” Simon task. In that condition,
the Simon effect did not differ reliably across the distribution function, showing a tendency for the
largest Simon effect to occur later in the RT distribution, when the temporal proximity between the
two stimulus codes, and thus between the resulting response processes, was increased. This shift in
the effect function was independent of the S-R task settings, which remained spatial-anatomical as
it was for the baseline condition. Moreover, since our stimuli were intact, this change in the time
course could not be attributed to temporal dissociation between the two stimulus dimensions nor to
imposition of masking patterns or overlays (see Hommel, 1993, 1994a, 1994b; Miles & Proctor,
2009; Vallesi & Umiltà, 2009).
Someone might object to these conclusions, though, by saying that wide coloured squares
may not only have facilitated the task-relevant colour discrimination but also altered some aspects
related to processing of the irrelevant stimulus spatial value and, thus, linked to spatial attention
mechanisms. For instance, increasing stimulus size may have, on one hand, decreased the perceived
distance between fixation and the stimulus and, on the other hand, affected location processing by
altering the probability and/or size of attention shifts toward the stimulus. To avoid this possible

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 15
critique, the wide unfilled stimuli condition was run, where stimulus width was the same as that of
the wide coloured stimuli condition but with colour presented only along the squares’ borders
instead of within their whole area. A decreasing time course, similar to that of baseline, would
corroborate the hypothesis that the sustained time course found for the wide coloured stimuli
condition was due to the facilitation of colour discrimination and thus to an increase in temporal
overlap. Results confirmed our expectations. First, the time course of the baseline condition did not
differ from that of the wide unfilled stimuli condition. Namely, the Simon effect tended to dissipate
across time for both these conditions. Second, the time course of the wide stimuli condition differed
from that of the baseline and wide unfilled stimuli conditions, being sustained rather than
decreasing. Third, the difference in RT ranges supported the hypothesis that the colour
discrimination was easier for the wide coloured stimuli condition with respect to the remaining
conditions. Indeed, there was a trend for the wide stimuli condition to have the fastest RTs and,
conversely, for the baseline and wide unfilled stimuli conditions to have similar mean RTs.
Hence, taken together, these findings broaden the evidence in favour of the automatic
activation account, which has previously been investigated mainly by way of manipulations which
decreased, instead of increasing, the temporal overlap. To corroborate our conclusions, we
conducted a second experiment in which an increased temporal overlap was also obtained by
manipulating the irrelevant, instead of the relevant, stimulus dimension.
Experiment 2
In line with the wide coloured stimuli condition of Experiment 1, the current experiment was
conducted to assess the time-course of the “standard” Simon effect when the temporal overlap
between the conditional and unconditional response mechanisms was increased. However, as
opposed to Experiment 1, we achieved this goal by manipulating the task irrelevant spatial
dimension of the stimulus instead of the relevant dimension. Lateralised arrows were employed as
visual stimuli in place of the squares used in Experiment 1. Converging evidence has been provided
about the strength and effectiveness of such spatial indicators: Even when presented centrally,
arrows have been found to quickly activate a spatial code and also to trigger a reflexive shift of

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 16
attention similar to the one caused by the abrupt onset of a peripheral non-predictive visual stimulus
(e.g., Bonato, Priftis, Marenzi, & Zorzi, 2008; Hommel, Pratt, Colzato, & Godijn, 2001; Ristic &
Kingstone, 2006). This finding was also confirmed by a recent study conducted by Pellicano et al.
(2009) that focused on the time course of such indicators. The authors demonstrated that the spatial
value of centrally presented arrows affected performance from the shortest RTs, as typically found
for lateralised stimuli. Furthermore, they influenced performance for a longer time than lateralised
stimuli, yielding a large Simon effect along the whole RT distribution (see also Proctor et al., 2009;
Ricciardelli, Bonfiglioli, Iani, Rubichi, & Nicoletti, 2007).
Considering the above mentioned findings, in the current experiment we decided to use
arrows presented at corresponding lateralised locations, instead of centrally presented ones. These
stimuli should produce two spatial codes, one given by the arrow direction and the other by its
location on the screen. Because the two always corresponded, the resulting spatial value, and also
the following unconditional response activation, would be stronger and longer lasting with respect
to that yielded by the square stimuli in the baseline condition of Experiment 1.
Hence, we hypothesized that a sustained Simon effect function, instead of a decreasing one
(i.e. baseline condition of Experiment 1), would be observed. Moreover, we also hypothesized that
the analysis of the conditional accuracy function (CAF), that is, the time course analysis of ARs,
would provide further evidence of such stronger unconditional activation. Many studies by
Ridderinkhof and colleagues (see Ridderinkhof, 2002a, 2002b; Wylie et al., 2009; Wylie,
Ridderinkhof, Bashore, & van den Wildenberg, 2010) suggest that the greater the strength of the
automatic response activation, the lower the ARs for non-corresponding trials mostly during shorter
RTs, since participants are more affected by the stimulus spatial value. A belated reaching of a near
perfect level of accuracy, thus, was expected for the current experiment compared to the baseline
condition of Experiment 1.
To summarize, the goal and the expected results of the current experiment were in line with
those of the wide coloured stimuli condition of Experiment 1: Increased temporal overlap was
supposed to yield a sustained time course for the Simon effect. However, in place of facilitating the

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 17
relevant colour task, we decided to enhance the irrelevant stimulus code. Hence, lateralised arrow
signals replaced the previously utilised squares stimuli. Their stronger spatial code was supposed to
influence performance for a longer time gap, thus yielding a constant or increasing Simon effect
across the RTs distribution and, also, a delayed increase of the ARs level in the CAF.
Method
Twelve new right-handed students from the same pool (6 females; mean age = 23.1 years,
SD = 2.1 years) were selected as before. The apparatus, stimuli and procedure were the same as in
Experiment 1 except that the stimuli were coloured (green/red) arrows, pointing to the left or to the
right, presented on the left/right of the screen (see Figure 1). Arrows were 1.5 cm wide and 1 cm
high (1.8° x 1.2°). The total coloured area for the arrow stimuli was: 0.65 cm2. As for the previous
Experiment, the distance of each stimulus from fixation was 9.4 cm (11.5° centre to centre). The
arrow direction was always congruent with its position: Arrows pointing to the left were presented
on the left side of the screen and the opposite was true for arrows pointing to the right.
Results
As for the previous experiment, we excluded from the analysis responses that were more
than two standard deviations smaller (0.1 %) or greater (4.2 %) than the participant’s overall mean
RT. Incorrect responses were also discarded (4.6 %). Mean correct RTs and ARs were analyzed
separately.
Mean reaction times. To test whether the shape of the effect function for RTs changed
significantly from the baseline condition of Experiment 1 to the arrow stimuli condition of
Experiment 2, we performed a repeated-measures ANOVA with Condition (Baseline vs. Arrow
Stimuli) as the between-subjects factor and Correspondence and Bin factors as the within-subjects
ones. When sphericity was violated, the Huynh-Feldt correction was applied, although the original
degrees of freedom are reported.
The Correspondence factor was significant, F(1,22)= 70.06, p < 001, indicating that
corresponding responses were faster than non-corresponding ones (377 vs. 401 ms). The Condition
factor was significant, F(1,22) = 5.15, p = .033, indicating that participants had faster performance

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 18
in the arrow stimuli condition compared to baseline (367 vs. 411 ms). The lack of significance of
the Correspondence × Condition interaction, F(1,22) < 1, indicates that the arrow stimuli condition
did not differ from the baseline condition as regard to the size of the Simon effect (354 vs. 380 ms
and 400 vs. 423 ms, for corresponding and non-corresponding trials of the arrow stimuli and
baseline condition, respectively, see Table 1). As hypothesized, the Bin × Correspondence ×
Condition interaction was significant, F(8,176) = 4.40 p = .005, showing that the arrow stimuli and
the baseline conditions yielded different time courses being stable the former and decreasing the
latter (see Figure 2). Indeed, the nonsignificant Bin × Correspondence interaction, F(8,88) = 2.21, p
= .121, of an ANOVA conducted on the single arrow stimuli condition, indicated that the size of the
Simon effect did not change reliably across bins: It was 20, 26, 29, 30, 29, 26, 26, 23 and 19 ms
from bin 1 to bin 9, respectively (see Figure 2).
Accuracy rates. To assess the CAF, that is, the time course of accuracy rates, ARs were
calculated for each of the nine RT bins and both corresponding and non-corresponding trials. Data
were then submitted to a repeated-measures ANOVA with the same factors as those of the RTs
analysis. When sphericity was violated, the Huynh-Feldt correction was applied, although the
original degrees of freedom are reported.
The Condition factor was significant, F(1,22) = 5.06, p = .035, showing that participants had
a less accurate performance for the arrow stimuli condition with respect to the baseline condition.
Taken together with the RT results, these finding indicate that participants adopted a faster but less
accurate criterion in responding in the arrow stimuli condition compared to baseline (367 vs. 411
ms and 94.5% vs. 96.4 %, for RTs and ARs, respectively). Thus, these results denote that a less
conservative speed-accuracy criterion was adopted on average in Experiment 2. The
Correspondence factor and the Correspondence × Condition interaction both failed to reach
significance, Fs(1,22) = 2.57, p =.123 and = 3.10, p = .092, respectively. The latter result indicates
that that two conditions did not differ as regard to the size of the Simon effect, even if the arrow
stimuli condition showed a clear trend to have a larger amplitude compared to the baseline one (0.1
% vs. 3.6 %, see Table 1).

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 19
Crucially, the Bin × Correspondence × Condition interaction was significant, F (8,176) =
3.06, p = . 004, indicating, in line with our hypothesis, that the ARs function for the arrow stimuli
condition differed to that of baseline. To investigate deeper this result, a repeated-measure ANOVA
on non-corresponding trials only was run where Bin and Condition were the within and between
subjects factors, respectively. In line with our prediction, the Bin × Condition interaction was
significant, F(8,176) = 3.02 p = .006, indicating that the arrow stimuli curve took a longer time to
reach an almost perfect level of accuracy compared to that of baseline (see Figure 3). Indeed, in an
ANOVA conducted on the non-corresponding trials of the baseline condition only, the Bin factor
resulted significant, F(8,88) = 9.05, p < .001, and Helmert contrasts showed that the ARs for non-
corresponding trials steeply increased from bin 1 to bin 3 (83 %, 93% and 97%, respectively),
Fs(1,11) > .9.36, ps < .02, for then remaining constant around a very high accuracy rate until bin 9
(97 %, 98%, 99%, 97% 99%, 98% and 99%, respectively), Fs(1,11) > 1. Crucially, when the same
ANOVA was run for the arrow stimuli condition, the Bin factor also turned out to be significant,
F(8,88) = 18.40, p < .001 and Helmert contrasts showed that, in line with our prediction,
participants took longer to reach a close-to-perfection level. Indeed, ARs for non-corresponding
trials increased significantly from bin 1 to bin 4 (78%, 87%, 90% and 93%, respectively), Fs(1,11)
> 17.17, ps < .003, slightly decreased from bin 4 to bin 5 (93 % and 91%), F(1,11) =15.90, p =
.002, increased again from bin 5 to bin 6 (91% and 97%, respectively), F(1,11) =12.00, p = .005,
and then remained constant until the last ninth bin (97%, 99%, 98% and 99%, respectively),
Fs(1,11) > 1 (See Figure 3). An ANOVA on corresponding trials only, with Bin and Condition as
the within- and between-subjects factors, respectively, was also run. As hypothesized, the ARs
functions of the two conditions did not differed between each others, as indicated by the
nonsignificant Bin × Condition interaction F(8,176) < 1. Indeed, for both the baseline and arrow
stimuli conditions participants had an almost perfect level of accuracy for corresponding trials along
all bins.
Discussion

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 20
The results of both the RTs and ARs time course investigations confirmed our predictions:
Lateralised arrow stimuli produced a stronger and longer lasting interference effect on performance.
The time course analysis revealed that the Simon effect was sustained across the whole RT
distribution, a result in line with that obtained for the wide coloured stimuli condition of Experiment
1, but different with respect to the decaying time course yielded by the baseline condition.
Moreover, the time course analysis of accuracy rates (see Ridderinkhof, 2002a, 2002b; Wylie et al.,
2008, Wylie et al., 2010) supported the idea of a stronger interference effect on performance along
bins: A near perfect level of accuracy for non-corresponding trials was reached later in time in the
current experiment compared to the baseline condition of Experiment 1. Hence, this finding
indicated that arrow stimuli had a stronger spatial code, compared to the squares one, which caused
participants to be more error prone for a longer time gap.
To conclude, these results supported the previously reported evidence showing that an
increased temporal overlap can affect the time course of a “standard” Simon effect, turning it from
decaying to stable. Moreover, these findings were further corroborated by the CAF analysis results,
which showed a stronger activation of the automatic response process.
General Discussion
The current study aimed to investigate the role of temporal overlap in shaping the time
course of a “standard” Simon effect. Two main conclusions can be drawn from our results. First,
enhancing the temporal proximity between stimulus codes, and thus between the resulting response
processes, yielded a sustained Simon effect across the whole RT distribution. This finding is new
with respect to literature where a decreasing time course has typically been found when a
“standard” setting is implemented, that is, when lateralised visual stimuli are matched to left/right
responses performed with uncrossed hands (e.g., Proctor et al., 2009). However, an automatic
activation account can encompass and explain both of these outcomes.
De Jong et al. (1994) were the first to attribute the decreasing time course of the “standard”
Simon effect to a fast, but stimulus locked, spatially correspondent response activation. Despite
being questioned (Roswarski & Proctor, 2003; Zhang & Kornblum, 1997), this hypothesis has

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 21
gathered converging evidence from both neuropsychological and behavioral studies. Among the
first ones are studies employing lateralized readiness potentials (LRPs), a technique developed in
the late 1980s (De Jong, Wierda, Mulder, & Mulder, 1988; Gratton, Coles, Sirevaag, Eriksen, &
Donchin, 1988) and which has been widely used within the Simon effect paradigm to investigate
early response preparation (see Leuthold, 2011, for a review). More precisely, the LRP: “exhibits
greater negativity in the motor cortex contralateral then ipsilateral to the responding hands a few
milliseconds before overt response onset” (Leuthold, 2011, p. 205). Several studies by Wascher and
colleagues provided data showing that this cortical asymmetry tended to disappear, or be drastically
reduced, when a non “standard” Simon task was implemented, that is, when stimuli were presented
in a vertical position or in the auditory modality, or when responses were made with crossed hands
(Buhlmann et al., 2007; Wascher et al., 2001; Wiegand & Wascher, 2005; Wiegand & Wascher,
2005). Such findings led Wascher et al. (2001) to claim that automatic activation was only yielded
by a good visuomotor connection, that is, when stimuli and responses share a spatial-anatomical
mapping.
Of particular relevance to our research are the results from behavioural studies which
showed a reduction, or even inversion, of the Simon effect for cases in which the temporal distance
between the conditional and unconditional response processes was increased. In these cases, the
automatically activated response had enough time to dissipate before response selection occurred
(Hommel, 1993, 1994a, 1994b; Simon et al, 1976; Vallesi & Umiltà, 2009). In a recent study, Miles
and Proctor (2009) showed that, besides its magnitude, such a decrease in temporal overlap had an
impact on the Simon effect function, when centrally presented spatial words (“left”/ “rights”) were
used.
The temporal overlap hypothesis was embraced by our experiments as well. However, we
assessed it by mean of a converse rationale with respect to the above mentioned studies: The
relevant and irrelevant stimulus dimensions were manipulated in such a way as to reduce, instead of
increase, the temporal distance between the conditional and unconditional responses. More
precisely, we first facilitated the discrimination of the relevant stimulus dimension (i.e., wide

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 22
coloured stimuli condition, Experiment 1), presenting stimuli with a wider coloured area compared
to those of baseline. In line with our prediction, the magnitude of the Simon effect did not vary with
time, indicating that the facilitation of the color discrimination shifted the conditional process closer
to the unconditional one. To make sure that this result was due to temporal overlap manipulations
and not to spatial attention matters related to coding of the irrelevant stimulus dimension, we ran a
control condition where the same stimuli were presented but with color presented only along their
borders, instead of within their whole area (i.e., wide unfilled stimuli condition). As hypothesized,
this condition yielded a decreasing Simon effect function similar to that of the baseline condition.
In Experiment 2, we then manipulated the irrelevant stimulus dimension using arrow stimuli in
place of squares (i.e., arrow stimuli condition) and found, as hypothesized, a sustained time course
as it was for the wide coloured stimuli condition. Hence, we interpreted our results as being in line
with an automatic activation account: Increasing the temporal overlap made participants respond
during the time span wherein the automatic activation had not decayed yet, thus yielding a stable
Simon effect function even for “standard” Simon tasks.
Although the manipulations intended to increase temporal overlap yielded stable, instead of
decreasing, effect functions, the overall mean Simon effect was no larger in those conditions than in
the baseline condition. The main reason why the mean effects did not differ seems to be that more
responses occurred prior to the build up of peak activation in the conditions with increased temporal
overlap (the wide coloured stimuli and arrow stimuli conditions) than in the baseline condition (see
Fig. 2). Thus, in addition to moving slow responses into the range prior to dissipation of the
corresponding-response activation, the conditions with increased temporal overlap apparently
moved more fast responses into the range during which the automatic activation was increasing.
An alternative possibility is that the mean Simon effect is determined by factors other than those
influencing the distribution functions, but given that mean data often show differences in Simon-
effect size expected on the basis of temporal overlap, this possibility seems less likely (see Proctor
et al., 2011, for a review).

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 23
It is worth noting that we manipulated the two stimulus dimensions, neither dissociating
them by mean of SOAs, pre- or post-cues, nor by altering the quality of the stimulus signal or
contrast. That is, we avoided adopting the most predominant paradigms that have been used in the
literature so far (Hommel, 1993, 1994a, 1994b; Miles & Proctor, 2009; Rubichi et al., 1997; Simon
et al., 1976; Vallesi & Umiltà, 2009).
The second conclusion concerns the CAF analysis. We showed in fact that this tool was
further informative on the effects of the strengthened stimulus spatial code and the consequent
automatic response activation. The spatial code of arrow indicators used for Experiment 2 was
demonstrated to be stronger with respect to the one of the square stimuli presented in the baseline
condition of Experiment 1, because participants tended to be error prone for a longer period in the
former condition than in the latter condition, thus showing greater difficulty in containing the
influence that arrows had on their performance. These results integrate well with those from RTs,
where arrows were found to exert a prolonged influence on performance, as shown by their
sustained effect function.
The CAF analysis was first implemented by Ridderinkhof and colleagues (Ridderinkhof
2002a, 2002b; Wylie et al., 2009, 2010) with the aim of investigating individual and group
differences in the activation and subsequent containment of the automatic response process. We
think that the CAF analysis could be a valid and informative tool to place side by side to the time
course investigation of RTs, even when the subject pool is composed of healthy adults and it is not
focused on individual or group differences.
To summarize, the current study aimed at increasing, instead of reducing, the temporal
overlap between the conditional and unconditional responses processes. As hypothesized, this
manipulation yielded a positive Simon effect with a sustained time course even for a “standard”
Simon task, a result which is new with respect to the literature. However, our interpretation was in
line with the temporal overlap hypothesis and with an automatic activation account.
More research is needed to further investigate the role of temporal overlap in shaping the
time course of the Simon effect. In particular, the current study only focused on the “standard”

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 24
Simon task, without investigating the other non “standard” tasks (e.g., see Wühr ,2006), for which
Wascher et al. (2001) argued that the Simon effect has a different, “cognitive” basis. Moreover, our
results did not disentangle the issue of whether the dissipation of the automatically activated
response is due to a passive decay or to an active inhibition. Evidence on both sides has been
gathered, but, to our knowledge, a definitive answer has yet to be given. What our results do show
is that by increasing temporal overlap, all responses can be made prior to much dissipation being
evident.
Acknowledgements
This work was supported by the EU FP7 project “Emergence of communication in Robots
through Sensorimotor and Social Interaction” (ROSSI), Grant Agreement 216125
(www.rossiproject.eu). The authors would also thank Christian Frings, Peter Wühr and Edmund
Wascher for helpful comments during the revision process.

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 25
References
Buhlmann, I., Umiltà, C., & Wascher, E. (2007). Response coding and visuomotor transformation
in the Simon task: The role of action goals. Journal of Experimental Psychology: Human
Perception and Performance, 33, 1269-1282.
Bonato, M., Priftis, K., Marenzi, R., & Zorzi, M. (2009). Normal and impaired reflexive orienting
of attention following central non-predictive cues. Journal of Cognitive Neuroscience, 21,
745-759.
De Jong, R., Liang, C-C., & Lauber, E. (1994). Conditional and unconditional automaticity: A
dual-process model of effects of spatial stimulus-response correspondence. Journal of
Experimental Psychology: Human Perception and Performance, 20, 731-750.
De Jong, R., Wierda, M., Mulder, G., & Mulder, L. J. M. (1988). The use of partial information in
response processing. Journal of Experimental Psychology: Human Perception and
Performance, 14, 682-692.
Gratton, G., Coles, M. G. H., Sirevaag, E. J., Eriksen, C. W., & Donchin, E. (1988). Pre- and
poststimulus activation of response channels: A psychophysiological analysis. Journal of
Experimental Psychology: Human Perception and Performance, 14, 331-344
Hommel (1993). The relationship between stimulus processing and response selection in the Simon
task: Evidence for a temporal overlap. Psychological Research, 55, 280-290.
Hommel, B. (1994a). Spontaneous decay of response-code activation. Psychological Research, 56,
261–268.
Hommel, B. (1994b). Effects of irrelevant spatial S’R compatibility depend on stimulus complexity.
Psychological Research, 56, 179–184.
Hommel, B., Pratt, J., Colzato, L., & Godijn, R. (2001). Symbolic control of visual attention.
Psychological Science, 12, 360-365.
Leuthold, H. (2011). The Simon effect in cognitive electrophysiology: A short review. Acta
Psychologica, 136, 203–211.
Miles, J., & Proctor, R. W. (2009). Reducing and restoring stimulus-response compatibility effects

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 26
by decreasing the discriminability of location words. Acta Psychologica, 130, 95-102.
Pellicano, A., Lugli, L., Baroni, G., & Nicoletti, R. (2009). The Simon effect with conventional
signals. A time-course analysis. Experimental Psychology, 56, 219-227.
Proctor, R. W., Miles, J. D., Baroni, G. (2011). Reaction-time distribution analysis of spatial
correspondence effects. Psychonomic Bullettin & Review, 18, 242–266.
Proctor, R. W., Pick, D. F., Vu, K.-P. L., & Anderson, R. E. (2005). The enhanced Simon effect for
older adults is reduced when the irrelevant location information is conveyed by an accessory
stimulus. Acta Psychologica, 119, 21-40.
Proctor, R. W., & Vu, K-P. L. (2006). Stimulus-response compatibility principles: Data, theory,
and application. Boca Raton, FL: Taylor & Francis.
Proctor, R. W., Vu, K.-P. L., & Nicoletti, R. (2003). Does right-left prevalence occur for the Simon
effect? Perception & Psychophysics, 65, 1318-1329.
Proctor, R. W., Yamaguchi, M., Zhang, Y., & Vu, K.-P. L. (2009). Influence of visual stimulus
mode on transfer of acquired spatial associations. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 35, 434-445.
Ratcliff, R. (1979). Group reaction time distributions and an analysis of distribution statistics.
Psychological Bulletin, 86, 446-461.
Ricciardelli, P., Bonfiglioli, C., Iani, C., Rubichi, S., & Nicoletti, R. (2007). Spatial coding and
central patterns: Is there something special about the eyes? Canadian Journal of
Experimental Psychology, 61, 79–90.
Ridderinkhof, R.K. (2002a). Micro- and macro-adjustments of task set: activation and suppression
in conflict tasks. Psychological Research, 66, 312-323.
Ridderinkhof, R.K. (2002b). Activation and suppression in conflict tasks: empirical clarification
through distributional analyses. In W. Prinz & B. Hommel (Eds.), Common Mechanisms in
Perception and Action: Attention & Performance, Vol. XIX (pp. 494–519). Oxford: Oxford
University Press.
Ristic, J., & Kingstone, A. (2006). Attention to arrows: Pointing to a new direction. The Quarterly

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 27
Journal of Experimental Psychology, 59, 1921-1930.
Roswarski, T. E., & Proctor, R. W. (2003). Intrahemispherical activation, visuomotor transmission,
and the Simon effect: Comment on Wascher et al. (2001). Journal of Experimental
Psychology: Human Perception and Performance, 29, 152-158.
Rubichi, S., Nicoletti, R., Iani, C., & Umiltà, C. (1997). The Simon effect occurs relative to the
direction of an attention shift. Journal of Experimental Psychology: Human Perception and
Performance, 23, 1353-1364.
Rubichi, S., & Pellicano A. (2004). Does the Simon effect affect movement execution? European
Journal of Cognitive Psychology, 16, 825-840.
Simon, J.R. (1990). The effects of an irrelevant directional cue on human information processing. In
R.W. Proctor & T.G. Reeve (Eds.), Stimulus–response compatibility: An integrated
perspective. Amsterdam: North-Holland.
Simon, J. R., Acosta, E., Mewaldt, S. P., & Speidel, C. R. (1976). The effect of an irrelevant
directional cue on choice reaction time: Duration of the phenomenon and its relation to
stages of processing. Perception & Psychophysics, 19, 16-22.
Vallesi, A., & Umiltà, C. A. (2009). Decay of stimulus spatial code in horizontal and vertical Simon
tasks. Journal of General Psychology, 136, 350–373.
Vallesi, A., Mapelli, D., Schiff, S., Amodio, P., & Umiltà, C. (2005). Horizontal and Vertical
Simon effect: different underlying mechanisms? Cognition, 96, 33-43.
Vu, K.-P. L., & Proctor, R. W. (2008). Age differences in response selection for pure and mixed
stimulus-response mappings and tasks. Acta Psychologica, 129, 49-60.
Wascher, E., Schatz, U., Kuder, T., & Verleger, R. (2001). Validity and boundary conditions of
automatic response activation in the Simon task. Journal of Experimental Psychology:
Human Perception and Performance, 27, 731-751.
Wiegand, K., & Wascher, E. (2005). Dynamic aspects of stimulus-response correspondence.
evidence for two mechanisms involved in the Simon effect. Journal of Experimental
Psychology: Human Perception and Performance, 31, 453–464.

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 28
Wiegand, K., & Wascher, E. (2007). The Simon effect for vertical S–R relations: changing the
mechanism by randomly varying the S–R mapping rule? Psychological Research, 71, 219-
233.
Wühr, P. (2006). The Simon effect in vocal responses. Acta Psychologica, 121, 210-226.
Wylie, S.A., Ridderinkhof, K.R., Bashore, T.R., van der Wildemberg, W.P. (2010). The effect of
Parkinson’s disease on the dynamics of on-line proactive cognitive control during action
selection. Journal of Cognitive Neuroscience, 22, 2058-2073.
Wylie, S.A., van den Wildenberg, W.P., Ridderinkhof, K.R., Bashore, T.R., Powell, V.D.,
Manning, C.A., Wooten, G.F. (2009). The effect of speed-accuracy strategy on response
interference control in Parkinson's disease. Neuropsychologia, 47(8-9), 1844-53.
Zhang, J., & Kornblum, S. (1997). Distributional analysis and De Jong, Liang, and Lauber’s (1994)
dual-process model of the Simon effect. Journal of Experimental Psychology: Human
Perception and Performance, 23, 1543–1551.

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 29
Table 1: Corresponding (C) and Non corresponding (NC) values and the resulting Simon effect
(SE; * = significant) for RT (ms) and AR (%) measures.
EXPERIMENT C NC SE
Experiment 1 - Baseline
Mean RTs (ms) 400 423 23*
ARs (%) 96.5 % 96.4 % 0.1 %
Experiment 1 – Wide Coloured Stimuli
Mean RTs (ms) 351 374 23*
ARs (%) 96.9 % 94.2 % 2.7* %
Experiment 1 – Wide Unfilled Stimuli
Mean RTs (ms) 392 415 23*
ARs (%) 97.2 % 93.9 % 3.3* %
Experiment 2 – Arrow Stimuli
Mean RTs (ms) 354 380 26*
ARs (%) 96.3 % 92.7 % 3.6 %

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 30
Figure 1: The stimuli used for Experiment 1 and for Experiment 2. The stimuli used for the baseline
condition (on the left side of the rightmost and leftmost panels) are matched with the stimuli used
for the wide coloured stimuli condition of Experiment 1 (right side of the leftmost panel) and with
those presented for arrow stimuli condition of Experiment 2 (right side of the rightmost panel). The
central panel displays the stimuli used for the wide coloured stimuli condition of Experiment 1
(right side) and those of the wide unfilled stimuli (left side).
Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 31
Figure 2: The magnitude of the Simon effect (RTs) across bins for the four conditions in
Experiments 1 and 2.
0
10
20
30
40
250 300 350 400 450 500 550
MEAN RT (MS)
SE
SIZ
E (
MS
)
BASELINE (Exp 1) WIDE COLOURED STIMULI (Exp 1)
WIDE UNFILLED STIMULI (Exp 1) ARROW STIMULI (Exp 2)

Running head: Influence of Temporal Overlap on Time Course of the Simon Effect 32
Figure 3: Accuracy rates for non-corresponding trials across bins for the baseline condition of
Experiments 1 and the arrow stimuli condition of Experiment 2. Accuracy for corresponding
condition was close to 100%.
70%
75%
80%
85%
90%
95%
100%
200 250 300 350 400 450 500 550
BASELINE (Exp 1) ARROW STMULI (Exp 2)




















