
Influence of Frequent Infectious Exposures on General and Varicella-Zoster Virus-Specific Immune...
10
Influence of Frequent Infectious Exposures on General and Varicella- Zoster Virus-Specific Immune Responses in Pediatricians Benson Ogunjimi, a,b,c Evelien Smits, d,e Steven Heynderickx, d,f Johan Van den Bergh, d Joke Bilcke, a Hilde Jansens, g Ronald Malfait, g Jose Ramet, c Holden T. Maecker, h Nathalie Cools, d Philippe Beutels, a,i Pierre Van Damme j Centre for Health Economics Research and Modeling Infectious Diseases (CHERMID), Vaccine and Infectious Disease Institute (VAXINFECTIO), University of Antwerp, Antwerp, Belgium a ; Interuniversity Institute for Biostatistics and Statistical Bioinformatics, Hasselt University, Hasselt, Belgium b ; Department of Pediatrics, Antwerp University Hospital, Edegem, Belgium c ; Laboratory of Experimental Hematology, Vaccine and Infectious Disease Institute (VAXINFECTIO), University of Antwerp, Antwerp, Belgium d ; Center for Oncological Research Antwerp, University of Antwerp, Antwerp, Belgium e ; Center for Cell Therapy and Regenerative Medicine, Antwerp University Hospital, Edegem, Belgium f ; Department of Laboratory Medicine, Antwerp University Hospital, Edegem, Belgium g ; Institute for Immunity, Transplantation, and Infection, Department of Microbiology and Immunology, Stanford University School of Medicine, Stanford, California, USA h ; School of Public Health and Community Medicine, The University of New South Wales, Sydney, Australia i ; Centre for the Evaluation of Vaccination (CEV), Vaccine and Infectious Disease Institute (VAXINFECTIO), University of Antwerp, Antwerp, Belgium j Reexposure to viruses is assumed to strengthen humoral and cellular immunity via the secondary immune response. We studied the effects of frequent exposure to viral infectious challenges on immunity. Furthermore, we assessed whether repetitive expo- sures to varicella-zoster virus (VZV) elicited persistently high immune responses. Blood samples from 11 pediatricians and matched controls were assessed at 3 time points and 1 time point, respectively. Besides the assessment of general immunity by means of measuring T-cell subset percentages, antibody titers and gamma interferon (IFN-)/interleukin 2 (IL-2)-producing T-cell percentages against adenovirus type 5 (AdV-5), cytomegalovirus (CMV), tetanus toxin (TT), and VZV were determined. Pediatricians had lower levels of circulating CD4 -naive T cells and showed boosting of CD8 effector memory T cells. Although no effect on humoral immunity was seen, repetitive exposures to VZV induced persistently higher percentages of IFN--positive T cells against all VZV antigens tested (VZV glycoprotein E [gE], VZV intermediate-early protein 62 [IE62], and VZV IE63) than in controls. T cells directed against latency-associated VZV IE63 benefitted the most from natural exogenous boosting. Although no differences in cellular or humoral immunity were found between the pediatricians and controls for AdV-5 or TT, we did find larger immune responses against CMV antigens in pediatricians. Despite the high infectious burden, we detected a robust and diverse immune system in pediatricians. Repetitive exposures to VZV have been shown to induce a stable increased level of VZV- specific cellular but not humoral immunity. Based on our observations, VZV IE63 can be considered a candidate for a zoster vaccine. T he secondary immune response in humans, elicited after reex- posure to a virus or other pathogens, can reinforce the quan- tity and quality of the immune response against the challenging pathogen. However, the existence of a saturation level or even exhaustion after repetitive natural exposures has not been suffi- ciently studied in humans. Also, some authors have discussed the existence of competition between pathogens in regard to humoral immunity (1, 2). The question thus remains how individuals with frequent exposures to different pathogens and repetitive expo- sures to some pathogens maintain a healthy immune system in balance. In particular, the ubiquitous varicella-zoster virus (VZV) presents a challenging dilemma, as stated by Hope-Simpson (3), who hypothesized that reexposure to VZV might postpone the reactivation of VZV, herpes zoster. As such, simulation programs have predicted temporary increases in herpes zoster incidence af- ter the introduction of a childhood widespread VZV vaccination program (4–6). Although a live attenuated VZV vaccine is cur- rently universally given to children in several countries (e.g., United States, Germany, and Australia), much debate regarding the suitability of such a program remains (5, 7–46), and a recent multidisciplinary systematic review has concluded that so-called exogenous boosting exists, but the true extent of this is yet to be determined (47). In this exploratory study, we have set forth the following goals. First, we aimed to describe the general effects of frequent infec- tious exposures in pediatricians on their humoral and cellular immune responses. Second, we set out to examine virus-specific effects of repeated exposures, particularly for VZV. MATERIALS AND METHODS Study design and subjects. Eleven pediatricians (age range, 33 to 60 years; mean age, 42 years; 7 women) comprising a high-exposure (HE) group donated venous blood samples at three different time points: winter (24 February to 16 March 2012) (HE-WIN), spring (11 May to 8 June 2012) (HE-SPR), and summer (3 to 19 July 2012) (HE-SUM). Information re- garding chickenpox exposure frequencies in pediatricians is shown in Table 1 (as recorded in their study journals). Eleven age (1 year)- and gender-matched normally exposed healthy control individuals (CO) with no known exposure to chickenpox during the past 2 years donated venous blood samples at a single time point (2 to 20 July 2012). Following recently Received 29 December 2013 Accepted 9 January 2014 Published ahead of print 15 January 2014 Editor: R. L. Hodinka Address correspondence to Benson Ogunjimi, [email protected]. Supplemental material for this article may be found at http://dx.doi.org/10.1128 /CVI.00818-13. Copyright © 2014, American Society for Microbiology. All Rights Reserved. doi:10.1128/CVI.00818-13 March 2014 Volume 21 Number 3 Clinical and Vaccine Immunology p. 417– 426 cvi.asm.org 417 on May 16, 2016 by guest http://cvi.asm.org/ Downloaded from
Transcript of Influence of Frequent Infectious Exposures on General and Varicella-Zoster Virus-Specific Immune...

Influence of Frequent Infectious Exposures on General and Varicella-Zoster Virus-Specific Immune Responses in Pediatricians
Benson Ogunjimi,a,b,c Evelien Smits,d,e Steven Heynderickx,d,f Johan Van den Bergh,d Joke Bilcke,a Hilde Jansens,g Ronald Malfait,g
Jose Ramet,c Holden T. Maecker,h Nathalie Cools,d Philippe Beutels,a,i Pierre Van Dammej
Centre for Health Economics Research and Modeling Infectious Diseases (CHERMID), Vaccine and Infectious Disease Institute (VAXINFECTIO), University of Antwerp,Antwerp, Belgiuma; Interuniversity Institute for Biostatistics and Statistical Bioinformatics, Hasselt University, Hasselt, Belgiumb; Department of Pediatrics, AntwerpUniversity Hospital, Edegem, Belgiumc; Laboratory of Experimental Hematology, Vaccine and Infectious Disease Institute (VAXINFECTIO), University of Antwerp, Antwerp,Belgiumd; Center for Oncological Research Antwerp, University of Antwerp, Antwerp, Belgiume; Center for Cell Therapy and Regenerative Medicine, Antwerp UniversityHospital, Edegem, Belgiumf; Department of Laboratory Medicine, Antwerp University Hospital, Edegem, Belgiumg; Institute for Immunity, Transplantation, and Infection,Department of Microbiology and Immunology, Stanford University School of Medicine, Stanford, California, USAh; School of Public Health and Community Medicine, TheUniversity of New South Wales, Sydney, Australiai; Centre for the Evaluation of Vaccination (CEV), Vaccine and Infectious Disease Institute (VAXINFECTIO), University ofAntwerp, Antwerp, Belgiumj
Reexposure to viruses is assumed to strengthen humoral and cellular immunity via the secondary immune response. We studiedthe effects of frequent exposure to viral infectious challenges on immunity. Furthermore, we assessed whether repetitive expo-sures to varicella-zoster virus (VZV) elicited persistently high immune responses. Blood samples from 11 pediatricians andmatched controls were assessed at 3 time points and 1 time point, respectively. Besides the assessment of general immunity bymeans of measuring T-cell subset percentages, antibody titers and gamma interferon (IFN-�)/interleukin 2 (IL-2)-producingT-cell percentages against adenovirus type 5 (AdV-5), cytomegalovirus (CMV), tetanus toxin (TT), and VZV were determined.Pediatricians had lower levels of circulating CD4�-naive T cells and showed boosting of CD8� effector memory T cells. Althoughno effect on humoral immunity was seen, repetitive exposures to VZV induced persistently higher percentages of IFN-�-positiveT cells against all VZV antigens tested (VZV glycoprotein E [gE], VZV intermediate-early protein 62 [IE62], and VZV IE63) thanin controls. T cells directed against latency-associated VZV IE63 benefitted the most from natural exogenous boosting. Althoughno differences in cellular or humoral immunity were found between the pediatricians and controls for AdV-5 or TT, we did findlarger immune responses against CMV antigens in pediatricians. Despite the high infectious burden, we detected a robust anddiverse immune system in pediatricians. Repetitive exposures to VZV have been shown to induce a stable increased level of VZV-specific cellular but not humoral immunity. Based on our observations, VZV IE63 can be considered a candidate for a zostervaccine.
The secondary immune response in humans, elicited after reex-posure to a virus or other pathogens, can reinforce the quan-
tity and quality of the immune response against the challengingpathogen. However, the existence of a saturation level or evenexhaustion after repetitive natural exposures has not been suffi-ciently studied in humans. Also, some authors have discussed theexistence of competition between pathogens in regard to humoralimmunity (1, 2). The question thus remains how individuals withfrequent exposures to different pathogens and repetitive expo-sures to some pathogens maintain a healthy immune system inbalance. In particular, the ubiquitous varicella-zoster virus (VZV)presents a challenging dilemma, as stated by Hope-Simpson (3),who hypothesized that reexposure to VZV might postpone thereactivation of VZV, herpes zoster. As such, simulation programshave predicted temporary increases in herpes zoster incidence af-ter the introduction of a childhood widespread VZV vaccinationprogram (4–6). Although a live attenuated VZV vaccine is cur-rently universally given to children in several countries (e.g.,United States, Germany, and Australia), much debate regardingthe suitability of such a program remains (5, 7–46), and a recentmultidisciplinary systematic review has concluded that so-calledexogenous boosting exists, but the true extent of this is yet to bedetermined (47).
In this exploratory study, we have set forth the following goals.First, we aimed to describe the general effects of frequent infec-
tious exposures in pediatricians on their humoral and cellularimmune responses. Second, we set out to examine virus-specificeffects of repeated exposures, particularly for VZV.
MATERIALS AND METHODSStudy design and subjects. Eleven pediatricians (age range, 33 to 60 years;mean age, 42 years; 7 women) comprising a high-exposure (HE) groupdonated venous blood samples at three different time points: winter (24February to 16 March 2012) (HE-WIN), spring (11 May to 8 June 2012)(HE-SPR), and summer (3 to 19 July 2012) (HE-SUM). Information re-garding chickenpox exposure frequencies in pediatricians is shown inTable 1 (as recorded in their study journals). Eleven age (�1 year)- andgender-matched normally exposed healthy control individuals (CO) withno known exposure to chickenpox during the past 2 years donated venousblood samples at a single time point (2 to 20 July 2012). Following recently
Received 29 December 2013 Accepted 9 January 2014
Published ahead of print 15 January 2014
Editor: R. L. Hodinka
Address correspondence to Benson Ogunjimi, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/CVI.00818-13.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/CVI.00818-13
March 2014 Volume 21 Number 3 Clinical and Vaccine Immunology p. 417– 426 cvi.asm.org 417
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

proposed guidelines, venipuncture was performed at fixed sampling sites(48). This study was approved by the ethics board of the University Hos-pital Antwerp, Antwerp, Belgium. Written informed consent was ob-tained from all study participants.
White blood cell count determination. One fresh EDTA tube persample was analyzed using an automated hemocytometer (Sysmex XS-1000i) in order to obtain the total white blood cell and differential counts.
Blood processing and flow cytometric analyses. Peripheral bloodmononuclear cells (PBMC) were isolated by Ficoll-Paque Plus gradientseparation (Amersham Biosciences, Uppsala, Sweden) from freshly ob-tained heparinized blood. The freezing of PBMC was performed in 90%fetal bovine serum (FBS) (Gibco Invitrogen, Ghent, Belgium) supple-mented with 10% dimethyl sulfoxide (DMSO) (Sigma-Aldrich, Stein-heim, Germany). Filled cryovials were put into Nalgene cryoboxes beforefreezing at �80°C, and afterwards, the cells were stored in liquid nitrogen.Thawing of the cells for batch analysis was done in a water bath at 37°C.After washing with preheated (37°C) medium supplemented with 10%FBS, flow cytometric analysis was performed. For this, PBMC were leftunstimulated or stimulated overnight in the presence of GolgiPlug (BDBiosciences) with (i) phytohemagglutinin (PHA) plus ionomycin, (ii)VZV intermediate-early protein 62 (IE62) PepMix (JPT, Berlin, Ger-many), (iii) VZV IE63 PepMix (JPT), (iv) VZV glycoprotein E (gE) Pep-Mix (JPT), (v) tetanus toxin (TT) mix (PANATecs, Heilbronn, Ger-many), (vi) cytomegalovirus (CMV) pp65 peptide mix (NIH, Bethesda,MD), or (vii) human adenovirus type 5 (AdV-5) penton PepMix (JPT).Per condition, 1 � 10E6 PBMC per ml Iscove’s modified Dulbecco’s me-dium (Gibco Life Technologies, Ghent, Belgium) plus 10% FBS werestimulated using 1 �g peptide/ml. Afterwards, the cells were washed andstained using the antibody panel as described in Table 2. In doing so, thecell subsets were determined based on membrane expression levels ofCD3, CD4, CD8, CD45RA, and CCR7 (see Fig. S1 in the supplementalmaterial for the gating strategy). Next, the cells were fixed and permeab-ilized using fluorescence-activated cell sorting (FACS) lysing solution andFACS permeabilizing solution 2 (BD Biosciences), according to the man-ufacturer’s protocol, before staining to determine intracellular expressionlevels of gamma interferon (IFN-�) and interleukin 2 (IL-2). Nonreactiveisotype-matched antibodies were used as controls for IL-2, IFN-�, andCCR7. Flow cytometric analysis was performed on a BD FACSAria II flowcytometer (BD Biosciences). If possible, 1,000,000 events were measured.The FlowJo software version 9.6.2 (Tree Star, Inc., Ashland, OR) was usedfor cytometric data analysis. As suggested recently, absolute cell countswere obtained by multiplying the ratio of marker-positive cells to thawed,viable, and single cells with the number of lymphocytes plus monocytesper volume unit (49). However, we noted no major differences betweenthe analyses based on the relative and absolute data.
Determination of antibody responses. Blood serum samples werefrozen at �80°C until batch analysis. All serological analyses were per-formed for both IgM and IgG. For the analysis of VZV-specific antibodies,two methods were used, a VZV IgM/G glycoprotein-based enzyme-linkedimmunosorbent assay (gpELISA) (Serion, Germany) and VZV-infectedcell lysate on XL Liaison using a DiaSorin kit (USA). IgG directed againstCMV pp150, pp28, p38, and p52 and IgM against CMV pp150, p52(RoCO, Basel, Switzerland), AdV-5 hexon-specific IgM/G (Demeditec,Germany), and tetanus toxin-specific IgM/G (Demeditec) titers were de-
termined as well. The positivity thresholds used were those given by themanufacturers.
Data management and statistics. Hematocytological cell counts andIgG titers against the different pathogens were compared between thedifferent sampling points (HE-WIN, HE-SPR, HE-SUM, and CO). Forthe flow cytometric analysis, only the time points HE-SPR and HE-SUMwere retained for further analysis due to a low viability in the PBMC fromHE-WIN.
First, T-cell subset percentages of CD3� T cells were compared be-tween the different sampling groups (HE-SPR, HE-SUM, and CO). TheCD3� CD4� and CD3� CD8� subsets were further subtyped as CCR7�
CD45RA� (naive T cells [Tna]), CCR7� CD45RA� (central memory Tcells [Tcm]), CCR7� CD45RA� (effector memory T cells [Tem]), andCCR7� CD45RA� (effector T cells [Tef]). Next, the percentages ofIFN-�- or/and IL-2-producing T-cell subtypes were calculated perstimulus by subtracting the percentage from the unstimulated sam-ples. Fisher’s exact test for contingency tables, comparing stimulatedwith unstimulated samples, was used to retain positive differences, witha P value of �0.05 (one-sided). Comparisons with a P value of 0.05 butwith �2,000 cells in the parent group were left blank due to the low cellcount (465/4,752 blanks with 144 due to one unusable PBMC sample inthe HE-SUM group). The proportions of antigen-responsive individualsper sampling group and per antigen were compared with the seropreva-lence per sampling group and per pathogen. The comparison was notformally made between AdV-5 hexon antibody and penton cytokine re-sponses due to the broader range of responses against AdV-5 hexon IgGby different adenovirus types compared to those against the AdV-5 pen-ton peptide mix. Individuals were considered immune to VZV or CMV ifthey had a positive serological or cytokine response against VZV or CMV,respectively. All individuals were considered immune against TT (due tovaccination), and for the cellular analyses against AdV-5 penton, onlyindividuals with positive cytokine responses were considered immune toAdV-5. The proportions of antigen-responsive individuals were com-pared per antigen between the three sampling groups. Next, antigen-stim-ulus-induced IFN-�-producing total T-cell and T-cell subset percentageswere calculated per pathogen for the pathogen-immune individuals.Comparisons were made between antigens and between the samplinggroups. Spearman’s correlations were calculated between the VZV IgGtiters and VZV IE62, VZV IE63, and VZV gE responses. Although ourmain focus was directed toward the antiviral cytokine IFN-�, a sensitivityanalysis included the combination of IFN-�- and IL-2-producing cells aswell.
All comparative statistical analyses were performed using either a two-sided t test (paired or unpaired) when the variable was deemed normallydistributed, or a nonparametric test (Wilcoxon signed-rank or Mann-Whitney rank-sum test) if not. A nonparametric test was used if nonnor-mality was concluded by the Shapiro-Wilk statistic for at least one of thedistributions, and otherwise both a t test and a nonparametric test wereused. However, if the nonparametric test proved to have more power todetect differences than did the t test (an indication of nonnormality), theresults from the nonparametric test were used. In addition, all statisticalanalyses based on cytokine-producing cell counts were performed using
TABLE 2 Antibody panel for PBMC staining
Target Fluorochromea Clone Company
IFN-� FITC B27 BDCCR7 PE 150503 BDCD3 PE-Texas Red S4.1 Life TechnologiesCD45RA PE-Cy7 HI100 BDLIVE/DEAD Violet Life TechnologiesIL-2 APC 5344.111 BDCD8 Pacific orange 3B5 Life TechnologiesCD4 APC-H7 RPA-T4 BDa FITC, fluorescein isothiocyanate; PE, phycoerythrin; APC, allophycocyanin.
TABLE 1 Monthly chickenpox exposure frequencies in pediatriciansa
Data
No. of exposures in:
February March April May June
Range 1–5 0–10 2–15 2–23 1–12Median 2 5 5 9 5No. of respondents 4 7 7 7 9a All pediatricians had an exposure to chickenpox within, at most, 11 weeks prior to thefirst sampling point, but mostly within a few weeks.
Ogunjimi et al.
418 cvi.asm.org Clinical and Vaccine Immunology
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

nonparametric tests due to the skewed distributions of these counts. Mat-lab 2012b and SPSS 20 were used for the analyses. The results were con-sidered significant if there was a P value of �0.05 or tended to be signifi-cant if the P value was 0.05 and �0.10.
RESULTSComparison of white blood cell counts. The total white blood cellcount tended to be higher in the HE-SUM (summer sampling ofpediatricians) group than in the HE-SPR (spring sampling of pe-diatricians), HE-WIN (winter sampling of pediatricians), and CO(control group) groups (factor 1.1, P 0.066; factor 1.1, P 0.079; factor 1.2, P 0.034, respectively). The neutrophil countwas higher in the HE-SUM group than in the HE-SPR group (fac-tor 1.2, P 0.028). The monocyte count tended to be higher in theHE-SUM group than in the HE-WIN group (factor 1.2, P 0.053).
Comparison of T-cell subset percentages (Fig. 1). The CD4�
CCR7� CD45RA� (CD4� Tna) percentage was lower in the HE-SUM group than in the CO group (factor 1.6, P 0.041). TheCD4� Tna percentage also tended to be lower in the HE-SPR
group than in the CO group (factor 1.4, P 0.076). In addition,we found that the CD8� Tna percentage tended to be lower in theHE-SUM group versus the HE-SPR group (factor 1.1, P 0.092).The CD8� CCR7� CD45RA� (CD8� Tem) percentage tended tobe higher in the HE-SPR and HE-SUM groups than in the COgroup (factor 1.3, P 0.076; factor 1.4, P 0.099, respectively).No differences were found for the total CD4�, total CD8�,CCR7� CD45RA� (Tcm), or CCR7� CD45RA� (Tef) percent-ages.
Antibody detection against common pathogens. Pathogenseropositivity remained constant at all time points in the HEgroups. All study participants were VZV IgG seropositive (forboth the VZV gpELISA and VZV cell lysate). All HE samples wereAdV-5 hexon seropositive, whereas the IgG titer was negative forone CO participant and unequivocal for another. For CMV only,a tendency (P 0.085) toward more IgG positive individuals wasshown in the HE samples (9/11) than in the CO samples (4/11)(Fig. 2 and Table 3).
None of the pathogens’ IgG titers indicated statistically mean-
FIG 1 Comparison of T-cell subset percentages (of CD3� PBMC). Distribution of T-cell subsets is presented per sampling as a box plot (with medians ashorizontal lines, interquartile range as boxes, ranges as vertical lines, and outliers as dots). Data for CD4� Tna, CO versus HE-SPR (P 0.076); for CD4� Tna,CO versus HE-SUM (P 0.041); for CD8� Tna, HE-SPR versus HE-SUM (P 0.092); for CD8� Tem, CO versus HE-SPR (P 0.076); and for CD8� Tem, COversus HE-SUM (P 0.099). All other P values are 0.10.
Effect of Frequent Natural Viral Exposures on Immunity
March 2014 Volume 21 Number 3 cvi.asm.org 419
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

ingful differences between the sampling groups, although theVZV cell lysate titer had a tendency to be higher in the HE-WINgroup than in the HE-SUM group (factor 1.1, P 0.090), and theCMV IgG titer showed a trend toward higher values in the HEgroups (Fig. 2). We note that several CMV IgG titers in the HEgroups reached the experimental upper limit.
Pathogen-specific T cells. Although CMV-specific IgG titerswere detected in only 4/11 CO samples, we detected CMV pp65-specific T cells in 10/11 CO samples when using IFN-� or/and IL-2intracellular cytokine staining upon stimulation (P 0.024). Inaddition, we observed that CMV positivity in the HE samplesreached 100% while using cellular immunology instead of serol-ogy as a measurement tool. We also noted only 6/11 CO sampleshaving VZV IE63-specific T cells (P 0.036), whereas all CO andHE samples had responses to VZV IE62. The percentage of sam-ples with cellular responses to VZV gE T cells ranged between 73and 90%. The percentage of individuals with TT-specific T cellswas significantly lower in CO (6/11) samples than in the HE-SPR(11/11, P 0.036) and HE-SUM (10/10, P 0.036) samples.CD4� and CD8� responses were observed against all antigenstested (Table 3).
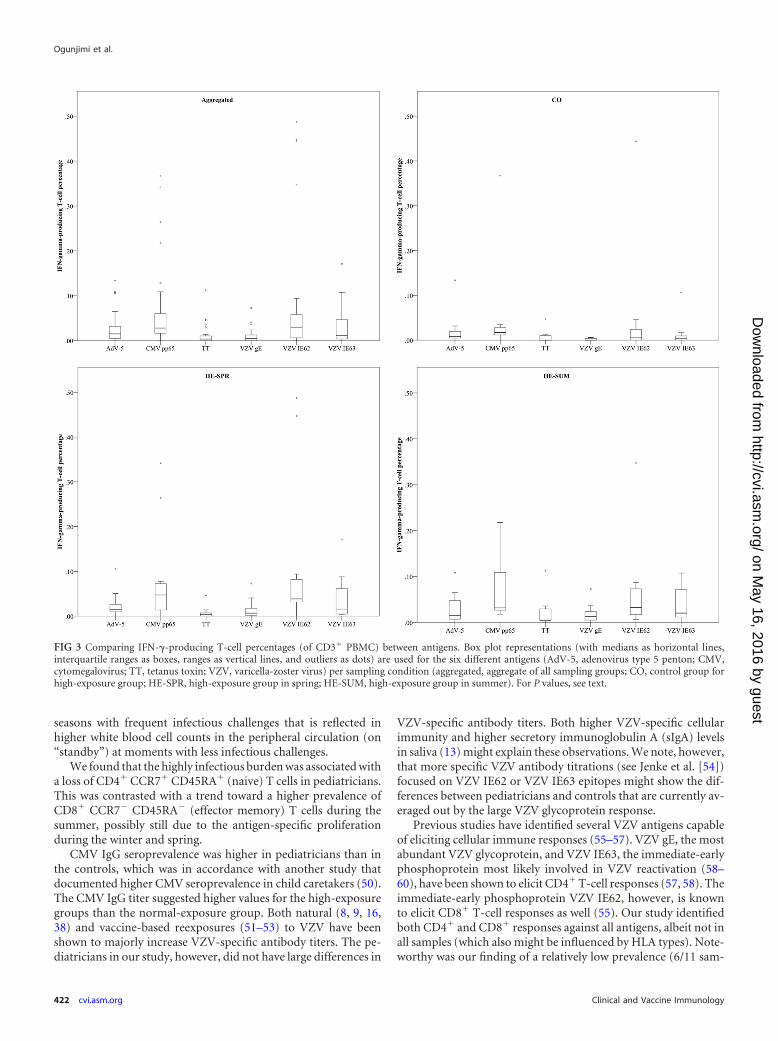
Comparing IFN-� T-cell percentages between antigens andbetween groups. Altogether, we noted that CMV pp65- and VZV
IE62-specific T-cell responses were the most abundant, althoughsome differences existed between the CO and HE samples (Fig. 3and 4).
The control individuals had more CMV pp65-specific T cellsthan T cells against VZV gE (P � 0.001), VZV IE63 (P 0.020), orTT (P � 0.001). Also, specific T cells against AdV-5 penton andVZV IE62 were more prevalent than those against VZV gE (P 0.0039 and P 0.079, respectively). Similarly, pediatricians hadhigher CMV pp65-specific T-cell responses both during springand summer than against VZV gE (P 0.042 and P 0.0052,respectively) or TT (P 0.0024 and P 0.015, respectively).Comparable to the controls, VZV IE62 responses were more fre-quent than VZV gE responses in both the HE-SPR (P 0.020) andHE-SUM (P 0.044) groups. However, VZV IE62 responses werealso more frequent than TT responses in both the HE-SPR (P 0.0031) and HE-SUM (P 0.024) groups and more frequent thanAdV-5 penton responses during the spring (P 0.049). We alsonoted that during the spring, TT-specific T-cell responses wereless frequent than those against AdV-5 penton (P 0.011) andVZV IE63 (P 0.0063).
Overall, we found positive correlations between the percent-ages of T cells against VZV IE62 and VZV IE63 (data not shown).Aggregating the different groups also showed a minor correlation
FIG 2 Antibody detection against common pathogens. IgG titers per pathogen (AdV-5 hexon [IU/ml], CMV [IU/ml], TT [IU/100 ml], VZV cell lysate[mIU/ml], VZV gp [mIU/ml]) and per sampling group are presented using a box plot (with medians as horizontal lines, interquartile ranges as boxes, ranges asvertical lines, and outliers as dots). Data for VZV cell lysate, HE-WIN versus HE-SUM (P 0.090). All other P values are 0.10.
Ogunjimi et al.
420 cvi.asm.org Clinical and Vaccine Immunology
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

between the percentages of T cells against VZV gE, VZV IE62, andVZV IE63 (data not shown).
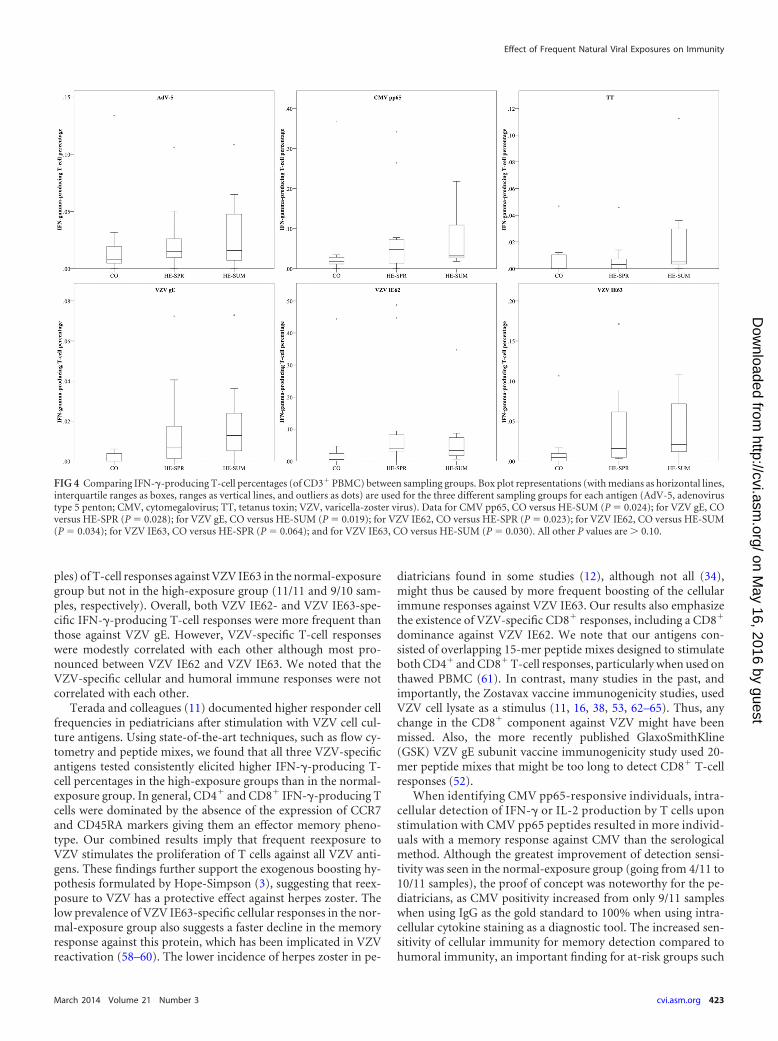
The antigen-specific analysis between the sampling groups(Fig. 4) found the percentage of CMV-specific T cells to be higherin the HE-SUM group than in the CO (P 0.024) group. AllVZV-specific T-cell percentages were lower in controls than inhighly exposed individuals (both spring and summer): VZV IE62was lower in the CO group than in the HE-SPR (P 0.023) andHE-SUM (P 0.034) groups, VZV IE63 was lower in the COgroup than in the HE-SPR (P 0.064) and HE-SUM (P 0.030)groups, and VZV gE was lower in the CO group than in the HE-SPR (P 0.028) and HE-SUM (P 0.0019) groups. No differ-ences between the sampling groups were found for AdV-5 pentonor TT.
Analysis of within-individual distribution of antigen-spe-cific T-cell subsets. The CO, HE-SPR, and HE-SUM antigen-spe-cific T-cell subset proportions showed for all antigens tested thatIFN-�-producing T cells mainly had a CD4� Tem, CD8� Tem, orcombined Tem subset. Noteworthy are the CD4� Tem domi-nance for VZV gE and the CD8� Tem dominance for VZV IE62(Fig. 5; see also Table S1 in the supplemental material).
Combining IFN-�- and IL-2-specific T-cell responses. Ingeneral, we noted that the inclusion of IL-2 did not have a major
effect on the results obtained with IFN-� alone. However, theproportion of CD4� Tcm responses was increased with the inclu-sion of IL-2 for all antigens tested (see Fig. S2 and Table S1 in thesupplemental material).
DISCUSSION
This exploratory study was designed to analyze the cellular andhumoral immune responses at different time points (winter,spring, and summer) in a group with high exposure to infections(represented by pediatricians) and to compare these to single timepoint responses (summer) in a group with what we considered anormal exposure load (non-MDs, nonteachers, etc.). The aim wasto analyze both the general indices of immune and antigen-spe-cific responses, particularly against VZV but also against CMV,adenovirus type 5, and tetanus toxin.
The total white blood cell count was about 10% to 20% higherfor the high-exposure group during the summer than in the win-ter and spring and than in the normal-exposure group. The neu-trophil and monocyte counts in the high-exposure group werehigher during the summer than during the spring and winter,respectively. Importantly, we did not find any differences for thelymphocyte counts. These results might imply that highly exposedindividuals have an upregulated innate immune system during
TABLE 3 Humoral and cellular immunity against common pathogensa
Pathogen Cellular antigen Group
No. ofindividualswith positiveresponses/totalno. ofindividuals for:
P for IgGvs CMI
CMI
P for:
No. ofindividuals withresponsive cells/total no. ofindividuals for:
P for CD4�
vs CD8�IgG CMICO vsHE-SPR
CO vsHE-SUM
HE-SPR vsHE-SUM
CD4�
cellsCD8�
cells
AdV-5 AdV-5 penton CO 9/10 9/11 NSb NS NS 6/11 6/10 NSHE-SPR 11/11 11/11 7/11 9/11 NSHE-SUM 11/11 10/10 9/10 6/10 NS
CMV CMV pp65 CO 4/11 10/11 0.024 NS NS NS 8/10 6/10 NSHE-SPR 9/11 11/11 NS 10/11 9/11 NSHE-SUM 9/11 10/10 NS 9/10 9/10 NS
Tetanus TT CO 10/11 6/11 NS 0.036 0.036 NS 6/11 3/11 NSHE-SPR 11/11 11/11 NS 8/11 5/11 NSHE-SUM 11/11 10/10 NS 10/10 5/10 0.033
VZV VZV gE CO 11/11 8/11 NS NS NS NS 6/11 3/10 NSHE-SPR 11/11 9/11 NS 9/11 3/11 0.016HE-SUM 11/11 9/10 NS 9/10 4/10 0.058
VZV VZV IE62 CO 11/11 11/11 NS NS NS NS 10/11 7/11 NSHE-SPR 11/11 11/11 NS 9/11 9/11 NSHE-SUM 11/11 10/10 NS 10/10 9/10 NS
VZV VZV IE63 CO 11/11 6/11 0.036 0.036 NS NS 5/11 5/11 NSHE-SPR 11/11 11/11 NS 8/11 9/11 NSHE-SUM 11/11 9/10 NS 8/10 6/10 NS
a Shown is memory against pathogens using either humoral (IgG) or cell-mediated immunity (CMI) (IFN-�- or/and IL-2-producing T cells) as proxy. Fisher’s exact test was usedto compare immunity as assessed by IgG or CMI (against an antigen) for three sampling groups (P values of �0.10 are shown). CMI is compared among the three groups (P valuesof �0.10 are shown). For each antigen, the presence of CD4� and CD8� IFN-� or/and IL-2 responses are assessed and compared with each other per antigen and per group (Pvalues of �0.10 are shown).b NS, nonsignificant.
Effect of Frequent Natural Viral Exposures on Immunity
March 2014 Volume 21 Number 3 cvi.asm.org 421
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from
seasons with frequent infectious challenges that is reflected inhigher white blood cell counts in the peripheral circulation (on“standby”) at moments with less infectious challenges.
We found that the highly infectious burden was associated witha loss of CD4� CCR7� CD45RA� (naive) T cells in pediatricians.This was contrasted with a trend toward a higher prevalence ofCD8� CCR7� CD45RA� (effector memory) T cells during thesummer, possibly still due to the antigen-specific proliferationduring the winter and spring.
CMV IgG seroprevalence was higher in pediatricians than inthe controls, which was in accordance with another study thatdocumented higher CMV seroprevalence in child caretakers (50).The CMV IgG titer suggested higher values for the high-exposuregroups than the normal-exposure group. Both natural (8, 9, 16,38) and vaccine-based reexposures (51–53) to VZV have beenshown to majorly increase VZV-specific antibody titers. The pe-diatricians in our study, however, did not have large differences in
VZV-specific antibody titers. Both higher VZV-specific cellularimmunity and higher secretory immunoglobulin A (sIgA) levelsin saliva (13) might explain these observations. We note, however,that more specific VZV antibody titrations (see Jenke et al. [54])focused on VZV IE62 or VZV IE63 epitopes might show the dif-ferences between pediatricians and controls that are currently av-eraged out by the large VZV glycoprotein response.
Previous studies have identified several VZV antigens capableof eliciting cellular immune responses (55–57). VZV gE, the mostabundant VZV glycoprotein, and VZV IE63, the immediate-earlyphosphoprotein most likely involved in VZV reactivation (58–60), have been shown to elicit CD4� T-cell responses (57, 58). Theimmediate-early phosphoprotein VZV IE62, however, is knownto elicit CD8� T-cell responses as well (55). Our study identifiedboth CD4� and CD8� responses against all antigens, albeit not inall samples (which also might be influenced by HLA types). Note-worthy was our finding of a relatively low prevalence (6/11 sam-
FIG 3 Comparing IFN-�-producing T-cell percentages (of CD3� PBMC) between antigens. Box plot representations (with medians as horizontal lines,interquartile ranges as boxes, ranges as vertical lines, and outliers as dots) are used for the six different antigens (AdV-5, adenovirus type 5 penton; CMV,cytomegalovirus; TT, tetanus toxin; VZV, varicella-zoster virus) per sampling condition (aggregated, aggregate of all sampling groups; CO, control group forhigh-exposure group; HE-SPR, high-exposure group in spring; HE-SUM, high-exposure group in summer). For P values, see text.
Ogunjimi et al.
422 cvi.asm.org Clinical and Vaccine Immunology
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from
ples) of T-cell responses against VZV IE63 in the normal-exposuregroup but not in the high-exposure group (11/11 and 9/10 sam-ples, respectively). Overall, both VZV IE62- and VZV IE63-spe-cific IFN-�-producing T-cell responses were more frequent thanthose against VZV gE. However, VZV-specific T-cell responseswere modestly correlated with each other although most pro-nounced between VZV IE62 and VZV IE63. We noted that theVZV-specific cellular and humoral immune responses were notcorrelated with each other.
Terada and colleagues (11) documented higher responder cellfrequencies in pediatricians after stimulation with VZV cell cul-ture antigens. Using state-of-the-art techniques, such as flow cy-tometry and peptide mixes, we found that all three VZV-specificantigens tested consistently elicited higher IFN-�-producing T-cell percentages in the high-exposure groups than in the normal-exposure group. In general, CD4� and CD8� IFN-�-producing Tcells were dominated by the absence of the expression of CCR7and CD45RA markers giving them an effector memory pheno-type. Our combined results imply that frequent reexposure toVZV stimulates the proliferation of T cells against all VZV anti-gens. These findings further support the exogenous boosting hy-pothesis formulated by Hope-Simpson (3), suggesting that reex-posure to VZV has a protective effect against herpes zoster. Thelow prevalence of VZV IE63-specific cellular responses in the nor-mal-exposure group also suggests a faster decline in the memoryresponse against this protein, which has been implicated in VZVreactivation (58–60). The lower incidence of herpes zoster in pe-
diatricians found in some studies (12), although not all (34),might thus be caused by more frequent boosting of the cellularimmune responses against VZV IE63. Our results also emphasizethe existence of VZV-specific CD8� responses, including a CD8�
dominance against VZV IE62. We note that our antigens con-sisted of overlapping 15-mer peptide mixes designed to stimulateboth CD4� and CD8� T-cell responses, particularly when used onthawed PBMC (61). In contrast, many studies in the past, andimportantly, the Zostavax vaccine immunogenicity studies, usedVZV cell lysate as a stimulus (11, 16, 38, 53, 62–65). Thus, anychange in the CD8� component against VZV might have beenmissed. Also, the more recently published GlaxoSmithKline(GSK) VZV gE subunit vaccine immunogenicity study used 20-mer peptide mixes that might be too long to detect CD8� T-cellresponses (52).
When identifying CMV pp65-responsive individuals, intra-cellular detection of IFN-� or IL-2 production by T cells uponstimulation with CMV pp65 peptides resulted in more individ-uals with a memory response against CMV than the serologicalmethod. Although the greatest improvement of detection sensi-tivity was seen in the normal-exposure group (going from 4/11 to10/11 samples), the proof of concept was noteworthy for the pe-diatricians, as CMV positivity increased from only 9/11 sampleswhen using IgG as the gold standard to 100% when using intra-cellular cytokine staining as a diagnostic tool. The increased sen-sitivity of cellular immunity for memory detection compared tohumoral immunity, an important finding for at-risk groups such
FIG 4 Comparing IFN-�-producing T-cell percentages (of CD3� PBMC) between sampling groups. Box plot representations (with medians as horizontal lines,interquartile ranges as boxes, ranges as vertical lines, and outliers as dots) are used for the three different sampling groups for each antigen (AdV-5, adenovirustype 5 penton; CMV, cytomegalovirus; TT, tetanus toxin; VZV, varicella-zoster virus). Data for CMV pp65, CO versus HE-SUM (P 0.024); for VZV gE, COversus HE-SPR (P 0.028); for VZV gE, CO versus HE-SUM (P 0.019); for VZV IE62, CO versus HE-SPR (P 0.023); for VZV IE62, CO versus HE-SUM(P 0.034); for VZV IE63, CO versus HE-SPR (P 0.064); and for VZV IE63, CO versus HE-SUM (P 0.030). All other P values are 0.10.
Effect of Frequent Natural Viral Exposures on Immunity
March 2014 Volume 21 Number 3 cvi.asm.org 423
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

as transplant patients and pregnant women, was recently alreadyshown for CMV (66) and had already been shown for variouspathogens, such as hepatitis C virus (67). CMV pp65-specificIFN-�-producing T cells were more frequent than those againstVZV IE63, VZV gE, TT, and AdV penton. The relatively highprevalence of CMV-specific T cells is consistent with the resultsfrom other studies (68, 69). The CMV pp65-specific T-cell fre-quencies were also higher in samples from pediatricians than inthose from the controls. We found no systematic differences in theT-cell frequencies against TT or AdV-5 penton between the pedi-atricians and controls, although TT-specific IFN-�- or IL-2-pro-ducing CD4� CCR7� CD45RA� (effector memory) cells weremore frequent at both sampling points for pediatricians than forcontrols.
This exploratory study was limited by its sample size, and conse-quently, its lower power for finding significant differences that mightexist (e.g., in regard to antibody responses). Despite this, several sys-tematic and significant results were found. The limited sample sizemade it impossible to assess immunity dysregulation caused byCMV, as was recently proposed for VZV (70). Nonetheless, someimportant conclusions can be made based on this study.
Future work should focus on a reanalysis of the VZV vaccine im-munogenicity studies by making use of 15-mer peptide mixes. Inaddition, our (albeit indirect) support for the importance of VZVIE63 in VZV reactivation might stimulate the development of a her-
pes zoster vaccine directed at VZV IE63. Future work should analyzethe long-term effects of frequent exposures on the specific and non-specific immune system, keeping in mind that the population in thecurrent study consisted mainly of young individuals.
In conclusion, while comparing immune responses betweenhighly exposed and normally exposed individuals, we observed atransient depletion of naive CD4� T cells and boosting of CD8�
effector memory T cells. Although no effect on humoral immu-nity was seen, we found that all VZV-specific antigens tested hadpersistently higher IFN-�-producing T-cell percentages afterstimulation in the highly exposed than in the normally exposedindividuals, thereby illustrating the stimulatory effects of the sec-ondary immune response.
ACKNOWLEDGMENTS
We thank all the participants for their participation and M. Claeys for hisconstructive comments.
The human CMV pp65 peptide pool was obtained through the NIHAIDS Research and Reference Reagent Program (Division of AIDS,NIAID, NIH). This work was supported by grants of the Research Foun-dation Flanders (project grant, predoctoral fellowship to B.O., postdoc-toral fellowships to J.B. and N.C.), the Hercules Foundation-Belgium, andthe University of Antwerp (predoctoral fellowship to J.V.D.B.). Thefunders had no role in study design, data collection or analysis, the deci-sion to publish, or preparation of the manuscript.
FIG 5 Comparing differences in IFN-�-producing T-cell subsets between sampling groups. Box plot representations (with medians as horizontal lines,interquartile ranges as boxes, ranges as vertical lines, and outliers as dots) are used to compare T-cell subset percentages (of CD3� PBMC) for the three differentsampling groups. See Table S1 in the supplemental material for P values.
Ogunjimi et al.
424 cvi.asm.org Clinical and Vaccine Immunology
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

REFERENCES1. Amanna IJ, Slifka MK. 2010. Mechanisms that determine plasma cell
lifespan and the duration of humoral immunity. Immunol. Rev. 236:125–138. http://dx.doi.org/10.1111/j.1600-065X.2010.00912.x.
2. Radbruch A, Muehlinghaus G, Luger EO, Inamine A, Smith KG,Dörner T, Hiepe F. 2006. Competence and competition: the challenge ofbecoming a long-lived plasma cell. Nat. Rev. Immunol. 6:741–750. http://dx.doi.org/10.1038/nri1886.
3. Hope-Simpson RE. 1965. The nature of herpes zoster: a long-term studyand a new hypothesis. Proc. R. Soc. Med. 58:9 –20.
4. Bilcke J, van Hoek AJ, Beutels P. 2013. Childhood varicella-zoster virusvaccination in Belgium: cost-effective only in the long run or withoutexogenous boosting? Hum. Vaccin. Immunother. 9:812– 822. http://dx.doi.org/10.4161/jv.23334.
5. Brisson M, Edmunds WJ, Gay NJ, Law B, De Serres G. 2000.Modelling the impact of immunization on the epidemiology of vari-cella zoster virus. Epidemiol. Infect. 125:651– 669. http://dx.doi.org/10.1017/S0950268800004714.
6. Schuette MC, Hethcote HW. 1999. Modeling the effects of varicellavaccination programs on the incidence of chickenpox and shingles. Bull.Math. Biol. 61:1031–1064. http://dx.doi.org/10.1006/bulm.1999.0126.
7. Gershon AA, Steinberg SP, Borkowsky W, Lennette D, Lennette E.1982. IgM to varicella-zoster virus: demonstration in patients with andwithout clinical zoster. Pediatr. Infect. Dis. 1:164 –167. http://dx.doi.org/10.1097/00006454-198205000-00007.
8. Arvin AM, Koropchak CM, Wittek AE. 1983. Immunologic evidence ofreinfection with varicella-zoster virus. J. Infect. Dis. 148:200 –205. http://dx.doi.org/10.1093/infdis/148.2.200.
9. Gershon AA, Steinberg SP. 1990. Live attenuated varicella vaccine: pro-tection in healthy adults compared with leukemic children. National In-stitute of Allergy and Infectious Diseases Varicella Vaccine CollaborativeStudy Group. J. Infect. Dis. 161:661– 666.
10. Garnett GP, Grenfell BT. 1992. The epidemiology of varicella-zostervirus infections: the influence of varicella on the prevalence of herpeszoster. Epidemiol. Infect. 108:513–528. http://dx.doi.org/10.1017/S0950268800050019.
11. Terada K, Kawano S, Yoshihiro K, Morita T. 1993. Proliferative re-sponse to varicella-zoster virus is inverse related to development of highlevels of varicella-zoster virus specific IgG antibodies. Scand. J. Infect. Dis.25:775–778. http://dx.doi.org/10.3109/00365549309008578.
12. Solomon BA, Kaporis AG, Glass AT, Simon SI, Baldwin HE. 1998. Lastingimmunity to varicella in doctors study (L.I.V.I.D. study). J. Am. Acad. Der-matol. 38:763–765. http://dx.doi.org/10.1016/S0190-9622(98)70207-5.
13. Terada K, Niizuma T, Yagi Y, Miyashima H, Kataoka N, Sadahiro T.2000. Low induction of varicella-zoster virus-specific secretory IgA anti-body after vaccination. J. Med. Virol. 62:46 –51. http://dx.doi.org/10.1002/1096-9071(200009)62:1�46::AID-JMV83.0.CO;2-Y.
14. Brisson M, Gay NJ, Edmunds WJ, Andrews NJ. 2002. Exposure tovaricella boosts immunity to herpes-zoster: implications for mass vacci-nation against chickenpox. Vaccine 20:2500 –2507. http://dx.doi.org/10.1016/S0264-410X(02)00180-9.
15. Thomas SL, Wheeler JG, Hall AJ. 2002. Contacts with varicella or withchildren and protection against herpes zoster in adults: a case-control study.Lancet 360:678–682. http://dx.doi.org/10.1016/S0140-6736(02)09837-9.
16. Vossen MT, Gent MR, Weel JF, de Jong MD, van Lier RA, KuijpersTW. 2004. Development of virus-specific CD4� T cells on reexposure tovaricella-zoster virus. J. Infect. Dis. 190:72– 82. http://dx.doi.org/10.1086/421277.
17. Jumaan AO, Yu O, Jackson LA, Bohlke K, Galil K, Seward JF. 2005.Incidence of herpes zoster, before and after varicella-vaccination-associated decreases in the incidence of varicella, 1992–2002. J. Infect. Dis.191:2002–2007. http://dx.doi.org/10.1086/430325.
18. Mullooly JP, Riedlinger K, Chun C, Weinmann S, Houston H. 2005.Incidence of herpes zoster, 1997–2002. Epidemiol. Infect. 133:245–253.http://dx.doi.org/10.1017/S095026880400281X.
19. Yavuz T, Ozdemir I, Sencan I, Arbak P, Behçet M, Sert E. 2005.Seroprevalence of varicella, measles and hepatitis B among female healthcare workers of childbearing age. Jpn. J. Infect. Dis. 58:383–386.
20. Yih WK, Brooks DR, Lett SM, Jumaan AO, Zhang Z, Clements KM,Seward JF. 2005. The incidence of varicella and herpes zoster in Massachu-setts as measured by the Behavioral Risk Factor Surveillance System (BRFSS)
during a period of increasing varicella vaccine coverage, 1998–2003. BMCPublic Health 5:68. http://dx.doi.org/10.1186/1471-2458-5-68.
21. Chaves SS, Santibanez TA, Gargiullo P, Guris D. 2007. Chickenpoxexposure and herpes zoster disease incidence in older adults in the U.S.Public Health Rep. 122:155–159.
22. Saadatian-Elahi M, Mekki Y, Del Signore C, Lina B, Derrough T, CaulinE, Thierry J, Vanhems P. 2007. Seroprevalence of varicella antibodiesamong pregnant women in Lyon-France. Eur. J. Epidemiol. 22:405– 409.http://dx.doi.org/10.1007/s10654-007-9136-z.
23. Bonmarin I, Santa-Olalla P, Lévy-Bruhl D. 2008. Modelling the impactof vaccination on the epidemiology of varicella zoster virus. Rev. Epide-miol. Sante Publique 56:323–331. (In French.) http://dx.doi.org/10.1016/j.respe.2008.07.087.
24. Patel MS, Gebremariam A, Davis MM. 2008. Herpes zoster-relatedhospitalizations and expenditures before and after introduction of thevaricella vaccine in the United States. Infect. Control Hosp. Epidemiol.29:1157–1163. http://dx.doi.org/10.1086/591975.
25. Valdarchi C, Farchi F, Dorrucci M, De Michetti F, Paparella C, Babud-ieri S, Spano A, Starnini G, Rezza G. 2008. Epidemiological investigationof a varicella outbreak in an Italian prison. Scand. J. Infect. Dis. 40:943–945. http://dx.doi.org/10.1080/00365540802308449.
26. Toyama N, Shiraki K, Society of the Miyazaki Prefecture Dermatol-ogists. 2009. Epidemiology of herpes zoster and its relationship tovaricella in Japan: a 10-year survey of 48,388 herpes zoster cases inMiyazaki prefecture. J. Med. Virol. 81:2053–2058. http://dx.doi.org/10.1002/jmv.21599.
27. Brisson M, Melkonyan G, Drolet M, De Serres G, Thibeault R, De WalsP. 2010. Modeling the impact of one- and two-dose varicella vaccinationon the epidemiology of varicella and zoster. Vaccine 28:3385–3397. http://dx.doi.org/10.1016/j.vaccine.2010.02.079.
28. Carville KS, Riddell MA, Kelly HA. 2010. A decline in varicella but anuncertain impact on zoster following varicella vaccination in Victoria,Australia. Vaccine 28:2532–2538. http://dx.doi.org/10.1016/j.vaccine.2010.01.036.
29. Donahue JG, Kieke BA, Gargiullo PM, Jumaan AO, Berger NR, Mc-Cauley JS, Belongia EA. 2010. Herpes zoster and exposure to the varicellazoster virus in an era of varicella vaccination. Am. J. Public Health 100:1116 –1122. http://dx.doi.org/10.2105/AJPH.2009.160002.
30. Grant KA, Carville KS, Kelly HA. 2010. Evidence of increasing frequencyof herpes zoster management in Australian general practice since the in-troduction of a varicella vaccine. Med. J. Aust. 193:483.
31. Karhunen M, Leino T, Salo H, Davidkin I, Kilpi T, Auranen K. 2010. Mod-ellingtheimpactofvaricellavaccinationonvaricellaandzoster.Epidemiol. Infect.138:469–481. http://dx.doi.org/10.1017/S0950268809990768.
32. Nelson MR, Britt HC, Harrison CM. 2010. Evidence of increasing fre-quency of herpes zoster management in Australian general practice sincethe introduction of a varicella vaccine. Med. J. Aust. 193:110 –113.
33. Rimland D, Moanna A. 2010. Increasing incidence of herpes zosteramong veterans. Clin. Infect. Dis. 50:1000 –1005. http://dx.doi.org/10.1086/651078.
34. Wu CY, Hu HY, Huang N, Pu CY, Shen HC, Chou YJ. 2010. Do thehealth-care workers gain protection against herpes zoster infection? A6-year population-based study in Taiwan. J. Dermatol. 37:463– 470. http://dx.doi.org/10.1111/j.1346-8138.2010.00804.x.
35. Gaillat J, Gajdos V, Launay O, Malvy D, Demoures B, Lewden L,Pinchinat S, Derrough T, Sana C, Caulin E, Soubeyrand B. 2011. Doesmonastic life predispose to the risk of Saint Anthony’s fire (herpes zoster)?Clin. Infect. Dis. 53:405– 410. http://dx.doi.org/10.1093/cid/cir436.
36. Jardine A, Conaty SJ, Vally H. 2011. Herpes zoster in Australia:evidence of increase in incidence in adults attributable to varicellaimmunization? Epidemiol. Infect. 139:658 – 665. http://dx.doi.org/10.1017/S0950268810001949.
37. Leung J, Harpaz R, Molinari NA, Jumaan A, Zhou F. 2011. Herpeszoster incidence among insured persons in the United States, 1993–2006:evaluation of impact of varicella vaccination. Clin. Infect. Dis. 52:332–340.http://dx.doi.org/10.1093/cid/ciq077.
38. Ogunjimi B, Smits E, Hens N, Hens A, Lenders K, Ieven M, VanTendeloo V, Van Damme P, Beutels P. 2011. Exploring the impact ofexposure to primary varicella in children on varicella-zoster virus immu-nity of parents. Viral Immunol. 24:151–157. http://dx.doi.org/10.1089/vim.2010.0031.
39. Salleras M, Domínguez A, Soldevila N, Prat A, Garrido P, Torner N,Borrás E, Salleras L. 2011. Contacts with children and young people and
Effect of Frequent Natural Viral Exposures on Immunity
March 2014 Volume 21 Number 3 cvi.asm.org 425
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from

adult risk of suffering herpes zoster. Vaccine 29:7602–7605. http://dx.doi.org/10.1016/j.vaccine.2011.08.023.
40. Tanuseputro P, Zagorski B, Chan KJ, Kwong JC. 2011. Population-based incidence of herpes zoster after introduction of a publicly fundedvaricella vaccination program. Vaccine 29:8580 – 8584. http://dx.doi.org/10.1016/j.vaccine.2011.09.024.
41. van Hoek AJ, Melegaro A, Zagheni E, Edmunds WJ, Gay N. 2011.Modelling the impact of a combined varicella and zoster vaccination pro-gramme on the epidemiology of varicella zoster virus in England. Vaccine29:2411–2420. http://dx.doi.org/10.1016/j.vaccine.2011.01.037.
42. Carville KS, Grant KA, Kelly HA. 2012. Herpes zoster in Australia.Epidemiol. Infect. 140:599 – 600 (Author reply, 600 – 601). http://dx.doi.org/10.1017/S095026881100149X.
43. Chao DY, Chien YZ, Yeh YP, Hsu PS, Lian IB. 2012. The incidence ofvaricella and herpes zoster in Taiwan during a period of increasing vari-cella vaccine coverage, 2000 –2008. Epidemiol. Infect. 140:1131–1140.http://dx.doi.org/10.1017/S0950268811001786.
44. Lasserre A, Blaizeau F, Gorwood P, Bloch K, Chauvin P, Liard F,Blanchon T, Hanslik T. 2012. Herpes zoster: family history and psycho-logical stress-case-control study. J. Clin. Virol. 55:153–157. http://dx.doi.org/10.1016/j.jcv.2012.06.020.
45. Ogunjimi B, Van Damme P, Beutels P. 2012. Zoster in monasteries:some clarification needed. Clin. Infect. Dis. 54:305–306 (Author reply,306 –307). http://dx.doi.org/10.1093/cid/cir841.
46. Hales CM, Harpaz R, Joesoef MR, Bialek SR. 2013. Examination of linksbetween herpes zoster incidence and childhood varicella vaccination.Ann. Intern. Med. 159:739 –745. http://dx.doi.org/10.7326/0003-4819-159-11-201312030-00006.
47. Ogunjimi B, Van Damme P, Beutels P. 2013. Herpes zoster risk reduc-tion through exposure to chickenpox patients: a systematic multidisci-plinary review. PLoS One 8:e66485. http://dx.doi.org/10.1371/journal.pone.0066485.
48. Ogunjimi B, Peeters D, Hens N, Malfait R, Van Tendeloo V, VanDamme P, Beutels P, Smits E. 2012. Sampling site matters when count-ing lymphocyte subpopulations. PLoS One 7:e41405. http://dx.doi.org/10.1371/journal.pone.0041405.
49. Ogunjimi B, Hens N, Malfait R, Van Tendeloo V, Smits E. 2013.Creating a robust framework for the analysis of cryopreserved samples inquantitative immunological experiments. J. Immunol. Methods 392:63–67. http://dx.doi.org/10.1016/j.jim.2013.03.006.
50. de Villemeur AB, Gratacap-Cavallier B, Casey R, Baccard-Longère M,Goirand L, Seigneurin JM, Morand P. 2011. Occupational risk for cyto-megalovirus, but not for parvovirus B19 in child-care personnel in France.J. Infect. 63:457– 467. http://dx.doi.org/10.1016/j.jinf.2011.06.012.
51. Levin MJ, Schmader KE, Gnann JW, McNeil SA, Vesikari T, Betts RF,Keay S, Stek JE, Bundick ND, Su SC, Zhao Y, Li X, Chan IS, Annun-ziato PW, Parrino J. 2013. Varicella-zoster virus specific antibody re-sponses in 50-59-year-old recipients of zoster vaccine J. Infect. Dis. 208:1386 –1390. http://dx.doi.org/10.1093/infdis/jit342.
52. Leroux-Roels I, Leroux-Roels G, Clement F, Vandepapelière P, VassilevV, Ledent E, Heineman TC. 2012. A phase 1/2 clinical trial evaluatingsafety and immunogenicity of a varicella zoster glycoprotein e subunitvaccine candidate in young and older adults. J. Infect. Dis. 206:1280 –1290.http://dx.doi.org/10.1093/infdis/jis497.
53. Levin MJ, Oxman MN, Zhang JH, Johnson GR, Stanley H, HaywardAR, Caulfield MJ, Irwin MR, Smith JG, Clair J, Chan IS, Williams H,Harbecke R, Marchese R, Straus SE, Gershon A, Weinberg A, VeteransAffairs Cooperative Studies Program Shingles Prevention Study Inves-tigators. 2008. Varicella-zoster virus-specific immune responses in elderlyrecipients of a herpes zoster vaccine. J. Infect. Dis. 197:825– 835. http://dx.doi.org/10.1086/528696.
54. Jenke AC, Klein S, Baiker A, Wirth S, Sander M, Noelting C, BoecherO, Vizoso-Pinto MG. 2012. Serologic analysis of the IgG antibody re-sponse in children with varicella zoster virus wild-type infection and vac-cination. Pediatr. Infect. Dis. J. 31:1148 –1152. http://dx.doi.org/10.1097/INF.0b013e31826bac27.
55. Frey CR, Sharp MA, Min AS, Schmid DS, Loparev V, Arvin AM. 2003.Identification of CD8� T cell epitopes in the immediate early 62 protein(IE62) of varicella-zoster virus, and evaluation of frequency of CD8� T cellresponse to IE62, by use of IE62 peptides after varicella vaccination. J.Infect. Dis. 188:40 –52. http://dx.doi.org/10.1086/375828.
56. Jones L, Black AP, Malavige GN, Ogg GS. 2007. Phenotypic analysis ofhuman CD4� T cells specific for immediate-early 63 protein of varicella-
zoster virus. Eur. J. Immunol. 37:3393–3403. http://dx.doi.org/10.1002/eji.200737648.
57. Malavige GN, Jones L, Black AP, Ogg GS. 2008. Varicella zoster virusglycoprotein E-specific CD4� T cells show evidence of recent activationand effector differentiation, consistent with frequent exposure to replica-tive cycle antigens in healthy immune donors. Clin. Exp. Immunol. 152:522–531. http://dx.doi.org/10.1111/j.1365-2249.2008.03633.x.
58. Malavige GN, Rohanachandra LT, Jones L, Crack L, Perera M, Fer-nando N, Guruge D, Ogg GS. 2010. IE63-specific T-cell responses asso-ciate with control of subclinical varicella zoster virus reactivation in indi-viduals with malignancies. Br. J. Cancer 102:727–730. http://dx.doi.org/10.1038/sj.bjc.6605542.
59. Zerboni L, Sobel RA, Ramachandran V, Rajamani J, Ruyechan W,Abendroth A, Arvin A. 2010. Expression of varicella-zoster virus imme-diate-early regulatory protein IE63 in neurons of latently infected humansensory ganglia. J. Virol. 84:3421–3430. http://dx.doi.org/10.1128/JVI.02416-09.
60. Nagel MA, Choe A, Traktinskiy I, Cordery-Cotter R, Gilden D, CohrsRJ. 2011. Varicella-zoster virus transcriptome in latently infected humanganglia. J. Virol. 85:2276 –2287. http://dx.doi.org/10.1128/JVI.01862-10.
61. Maecker HT, Dunn HS, Suni MA, Khatamzas E, Pitcher CJ, Bunde T,Persaud N, Trigona W, Fu TM, Sinclair E, Bredt BM, McCune JM,Maino VC, Kern F, Picker LJ. 2001. Use of overlapping peptide mixturesas antigens for cytokine flow cytometry. J. Immunol. Methods 255:27– 40.http://dx.doi.org/10.1016/S0022-1759(01)00416-1.
62. Weinberg A, Zhang JH, Oxman MN, Johnson GR, Hayward AR,Caulfield MJ, Irwin MR, Clair J, Smith JG, Stanley H, Marchese RD,Harbecke R, Williams HM, Chan IS, Arbeit RD, Gershon AA, SchodelF, Morrison VA, Kauffman CA, Straus SE, Schmader KE, Davis LE,Levin MJ, U.S. Department of Veterans Affairs (VA) Cooperative StudiesProgram Shingles Prevention Study Investigators. 2009. Varicella-zostervirus-specific immune responses to herpes zoster in elderly participants ina trial of a clinically effective zoster vaccine. J. Infect. Dis. 200:1068 –1077.http://dx.doi.org/10.1086/605611.
63. Weinberg A, Lazar AA, Zerbe GO, Hayward AR, Chan IS, Vessey R,Silber JL, MacGregor RR, Chan K, Gershon AA, Levin MJ. 2010.Influence of age and nature of primary infection on varicella-zoster virus-specific cell-mediated immune responses. J. Infect. Dis. 201:1024 –1030.http://dx.doi.org/10.1086/651199.
64. Irwin MR, Levin MJ, Laudenslager ML, Olmstead R, Lucko A, Lang N,Carrillo C, Stanley HA, Caulfield MJ, Weinberg A, Chan IS, Clair J,Smith JG, Marchese RD, Williams HM, Beck DJ, McCook PT, ZhangJH, Johnson G, Oxman MN. 2013. Varicella zoster virus-specific im-mune responses to a herpes zoster vaccine in elderly recipients with majordepression and the impact of antidepressant medications. Clin. Infect.Dis. 56:1085–1093. http://dx.doi.org/10.1093/cid/cis1208.
65. Weinberg A, Huang S, Song LY, Fenton T, Williams P, Patterson J,Tovar-Salazar A, Levin MJ. 2012. Immune correlates of herpes zoster inHIV-infected children and youth. J. Virol. 86:2878 –2881. http://dx.doi.org/10.1128/JVI.06623-11.
66. Loeth N, Assing K, Madsen HO, Vindeløv L, Buus S, Stryhn A. 2012.Humoral and cellular CMV responses in healthy donors; identificationof a frequent population of CMV-specific, CD4� T cells in seronegativedonors. PLoS One 7:e31420. http://dx.doi.org/10.1371/journal.pone.0031420.
67. Heller T, Werner JM, Rahman F, Mizukoshi E, Sobao Y, Gordon AM,Sheets A, Sherker AH, Kessler E, Bean KS, Herrine SK, Stevens M,Schmitt J, Rehermann B. 2013. Occupational exposure to hepatitis Cvirus: early T-cell responses in the absence of seroconversion in a longitu-dinal cohort study. J. Infect. Dis. 208:1020 –1025. http://dx.doi.org/10.1093/infdis/jit270.
68. Gillespie GM, Wills MR, Appay V, O’Callaghan C, Murphy M, SmithN, Sissons P, Rowland-Jones S, Bell JI, Moss PA. 2000. Functionalheterogeneity and high frequencies of cytomegalovirus-specific CD8(�)T lymphocytes in healthy seropositive donors. J. Virol. 74:8140 – 8150.http://dx.doi.org/10.1128/JVI.74.17.8140-8150.2000.
69. Khan N, Shariff N, Cobbold M, Bruton R, Ainsworth JA, Sinclair AJ,Nayak L, Moss PA. 2002. Cytomegalovirus seropositivity drives the CD8T cell repertoire toward greater clonality in healthy elderly individuals. J.Immunol. 169:1984 –1992.
70. Ogunjimi B, Theeten H, Hens N, Beutels P. Serology indicates cytomeg-alovirus infection is associated with varicella-zoster virus reactivation. J.Med. Virol., in press. http://dx.doi.org/10.1002/jmv.23749.
Ogunjimi et al.
426 cvi.asm.org Clinical and Vaccine Immunology
on May 16, 2016 by guest
http://cvi.asm.org/
Dow
nloaded from








![[Herpes Zoster and its prevention in Italy. Scientific consensus statement]](https://static.fdokumen.com/doc/165x107/6332d5755f7e75f94e094855/herpes-zoster-and-its-prevention-in-italy-scientific-consensus-statement.jpg)











