
Availability Modeling of Computing Systems with Virtual Architectures

INFLUENCE OF DECOMPOSITION OF ROOTS OF
TROPICAL FORAGE SPECIES ON THE AVAILABILITY OF
SOIL NITROGEN
SEGUNDO URQUIAGA,1,2 GEORG CADISCH,2* BRUNO J. R. ALVES,1
ROBERT M. BODDEY1 and KEN E. GILLER2
1EMBRAPA, Centro Nacional de Pesquisa de Agrobiologia (CNPAB), Km 47, Serope dica, Itaguaõ ,23851-970, Rio de Janeiro, Brazil and 2Department of Biological Sciences, Wye College, University of
London, Wye, TN25 5AH, Kent, U.K.
(Accepted 21 May 1998)
SummaryÐImmobilization of mineral N induced by decomposition of roots of four tropical foragespecies (Stylosanthes guianensis, Centrosema sp., Andropogon gayanus and Brachiaria decumbens) in anOxisol was studied under laboratory conditions. Root materials had a high lignin content (12±20%)but total polyphenol content was small (<0.8%). Roots, at 2.5 and 5.0 g kgÿ1, and 10 mg N kgÿ1 of15N-labelled ammonium sulphate (20.3 at.%) were thoroughly mixed with the soil which was main-tained at ®eld capacity for 117 d. Decomposition of the roots (as monitored by CO2 evolution) was in-itially rapid and the legume materials (S. guianensis, Centrosema sp.) with their lower C-to-N ratio andlignin content, decomposed more quickly than the grass roots (A. gayanus, B. decumbens). After 8 d ofincubation the rate of CO2 evolution decreased and was similar for all root materials. CO2 evolutionfrom the decomposing roots in all cases ®tted closely (R2>0.99) a double exponential equation de®ningtwo compartments of root carbon of di�ering susceptibility to decomposition. The equation predictedthat between 43% (Centrosema) and 62% (Brachiaria) of root carbon would not be decomposed evenat in®nite time under incubation conditions. Mineral N in the soil was immobilized rapidly at the startof the incubation, and the immobilization was greatest with the higher rate of application of root ma-terial. Although the C-to-N ratio of legume roots was narrower their higher degradability stimulatedgreater immobilization of soil mineral N than the grass roots. The results are discussed with referenceto N immobilization and carbon sequestration in planted pastures of tropical South America. # 1998Elsevier Science Ltd. All rights reserved
INTRODUCTION
The conservation of organic residues on the soil
surface, or their incorporation into the soil, is aprocedure widely recommended to maintain soil fer-tility. E�ects of residues on the growth and pro-ductivity of crops are often unpredictable, and both
positive and negative e�ects can be observed(Biederbeck et al., 1980; Ladd et al., 1981a;Urquiaga et al., 1982, 1994; Badaruddin and
Meyer, 1990). Reductions in plant growth may becaused directly by liberation of toxins (Halsall etal., 1995; Leigh et al., 1995) or indirectly by the
reduced availability of plant nutrients, especiallynitrogen (Terman and Brown, 1968; Myers andPaul, 1971; Urquiaga et al., 1994).Organic residues are an important source of
nitrogen. Release of N from these materials in soilis determined not only by environmental conditionsand characteristics of the soil, but also by the com-
position or quality of the residues (Cadisch andGiller, 1997). Many studies have shown that the lib-
eration of N is variously related to the C-to-N ratio
of the organic residues (Herman et al., 1977;
Thomas and Asakawa, 1993), their lignin content
(Herman et al., 1977; MuÈ ller et al., 1988; Thomas
and Asakawa, 1993), the content and quality of
polyphenols (Vallis and Jones, 1973; Melillo and
Aber, 1984; Sivapalan et al., 1985; Palm and
Sanchez, 1991), (lignin + polyphenols)-to-N ratio
(Fox et al., 1990) or an active fraction of the poly-
phenols (Handayanto et al., 1994, 1995, 1997).
With the exception of organic residues of high N
content derived from young plant tissues or green
manures which can usually liberate a large pro-
portion of their N, most residues from harvested
crops release only a small proportion of the N in
their tissues immediately, most N being integrated
into soil organic matter (Ladd et al., 1981a,b;
Urquiaga et al., 1982; MuÈ ller et al., 1988). Such
poor quality residues may result in immobilization
of signi®cant quantities of available soil mineral N
during decomposition (Terman and Brown, 1968;
Myers and Paul, 1971; Tomar and Soper, 1981;
Mengel and Schmeer, 1985) thus prejudicing plant
growth.
Soil Biol. Biochem. Vol. 30, No. 14, pp. 2099±2106, 1998# 1998 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0038-0717/98 $19.00+0.00PII: S0038-0717(98)00086-8
*Author for correspondence.E-mail: [email protected]
2099

Few investigators have studied the e�ects of in-corporation of roots or their in situ decomposition.
In some crops, for example sugar cane when subjectto pre-harvest burning (de Oliveira et al., 1994), theroots constitute virtually the only input of organic
carbon to the soil. Fisher et al. (1994) reported thatimproved pastures based on Andropogon gayanus orBrachiaria spp. accumulated the equivalent of 7 t C
haÿ1 yrÿ1 in the soil. Despite such large annual ad-ditions of root residues to the soil in pastures of thetropical savannahs of South America, their pro-
ductivity usually declines after some years. Thispasture decline or ``degradation'' has been attribu-ted to lack of plant-available N in the soil (Robbinset al., 1989; Boddey et al., 1996), but interactions
with root decomposition have not yet been studied.The introduction of legumes into these pastures isthought to alleviate N de®ciencies and while leaf lit-
ters often contribute greatly to net N mineralizationlittle is known about the behaviour of legume rootmaterials. Thus our principal objective was to
compare the e�ect of decomposition of roots fromtropical forage legume and grasses on availability ofsoil N.
MATERIALS AND METHODS
Plant material and soil
Roots were obtained from four tropical foragespecies: two legumes; Stylosanthes guianensis cv.Bandeirante and Centrosema sp. (hybrid C.pubescens�C. acutifolium cv. ItaguaõÂ ), and two
grasses; Andropogon gayanus cv. Planaltina andBrachiaria decumbens cv. Basilisk. These plants hadbeen established for some years on an acid soil,
ItaguaõÂ series (Typic Hapludult; Ramos et al.,1973), of low nutrient availability, at the ®eldstation of the National Centre for Agrobiology
Research (EMBRAPA-CNPAB), ItaguaõÂ , Rio deJaneiro. Roots were excavated to 30 cm depth, sep-arated by sieving, carefully washed and dried at608C for 72 h. Root material used for the incu-
bation experiment was of diameter 0.5±3 mm andground to pass a 1 mm mesh. Some chemicalcharacteristics of these root tissues are shown in
Table 1.
The soil used for the incubation experiment wasa dark-red latosol (Oxisol) from the central savan-
nah region of Brazil (EMBRAPA-CPAC,Planaltina, Federal District of BrasõÂ lia). The soil ofclay-loam texture, 1.0% organic C, 0.085% total N,
pH 5.5, cation exchange capacity 1.5 cmol(+) kgÿ1,
Ca2++ Mg2+, Al3+ and K+, 1.0, 0.4 and0.1 cmol(+) kg
ÿ1, respectively, and low P availability
(<2 mg P g soilÿ1, Mehlich).
Experimental procedures
At 14 d before the addition of root material thesoil was amended with single superphosphate, pot-
assium sulphate, magnesium sulphate and zinc sul-phate at the following rates kgÿ1 dry soil: 150 mg P,100 mg K, 30 mg Mg and 4.5 mg Zn, respectively,
and lime at 1 cmol Ca kgÿ1 to avoid nutrient de-®ciencies other than N. Aliquots of either 300 or30 g of the fertilized soil were distributed in 500 and250 ml glass bottles for the evaluation of CO2 evol-
ution and soil mineral N dynamics, respectively.Roots were mixed into the soil samples and fol-lowed by 10 mg N kgÿ1 of 15N-labelled ammonium
sulphate (20.3 at.% 15N) added in 5 ml of solutionfor each 30 g of soil and thoroughly mixed in. Thesoil in the ¯asks was then moistened to 100% of
water holding capacity (23.8% w/w). For the evalu-ation of CO2 evolution root materials were incor-porated at 5 g kg soilÿ1, while for the evaluation of
soil mineral N two rates of addition of root ma-terials, 2.5 and 5.0 g kg soilÿ1, were used.The treatments consisted of additions of root ma-
terial of the four di�erent forage species, plus a
control treatment with no added roots. Sequentialevaluations of CO2 evolution, by trapping CO2 in25 ml of 0.1±0.5 M KOH, were performed from the
four replicate ¯asks of each treatment according tothe technique of Anderson et al. (1983) during theintervals between sampling times of 0, 0.5, 1, 2, 3,
3.5, 4, 8, 15, 29, 51, 71 and 117 d. Microbial bio-mass N was evaluated in 30 g of soil taken from theCO2 ¯asks at the end of the experiment (117 d)using the fumigation±extraction technique followed
by analysis of the total N in the extract (Amatoand Ladd, 1988).For the evaluations of mineral N concentration,
separate bottles were prepared for sampling every
Table 1. Chemical characteristics of plant roots used in the experiment
Plant root of: Na (%) Total Ca (%) C-to-N ratio Ligninb (%)Lignin-to-N
ratio
Solublepolyphenolsc
(%) Soluble Cd (%)
Stylosanthes 1.01 43.9 44 12 11.9 0.80 naCentrosema 1.58 43.3 27 16 10.1 0.41 0.6Andropogon 0.61 42.4 70 18 29.5 0.68 0.7Brachiaria 0.33 42.7 129 20 60.6 0.55 0.6
aTotal N and C were determined with a CHN autoanalyser.bLignin was determined by the acid detergent ®bre method (Van Soest and Wine, 1968).cSoluble polyphenols, expressed as tannic acid, determined by the Folin±Denis method (Anderson and Ingram, 1989).dHot water extractable (408C for 20 min).
S. Urquiaga et al.2100
7 d until 28 d then subsequently at 56, 77 and
117 d. The soil (30 g) samples were extracted with
90 ml of 2 M KCl and subsequently steam distilledin the presence of 200 mg of Devarda's alloy and
200 mg of MgO into dilute hydrochloric acid
according to the procedure of Keeney and Nelson
(1982) but modi®ed, replacing boric acid with hy-drochloric acid, as the boric acid led to problems in
the automated CN analyzer. After titration with
NaOH solution to determine the total mineral N
content (NH4++NO3
ÿ), the solutions were acidi®edand evaporated to dryness. The resulting residues
were placed in tin capsules for analysis of 15N
enrichment using a Roboprep automatic CN analy-
ser (Europa Scienti®c, Crewe, U.K.) coupled to a
VG 622 mass spectrometer (VG Isogas,Middlewich, U.K.).
Net mineralization±immobilization of soil mineral
N in each treatment was expressed as a percentage
of the mineral N available in the control treatment
at time zero (60 mg N g soilÿ1).
RESULTS
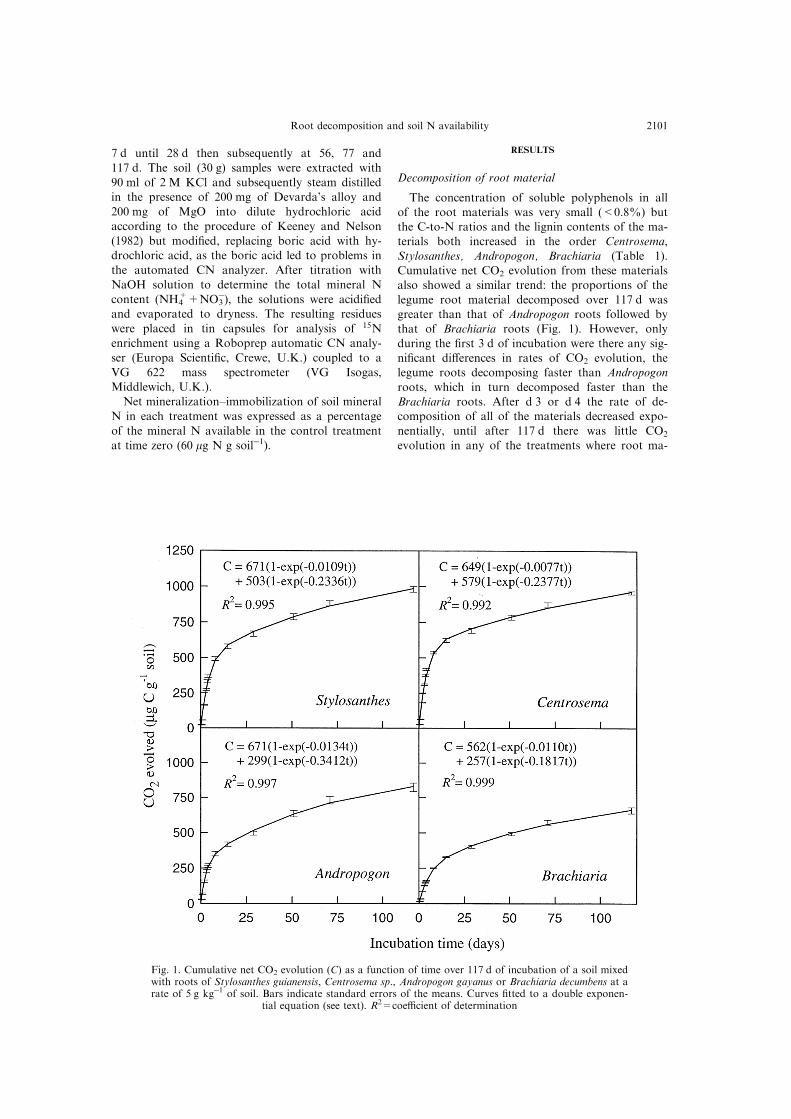
Decomposition of root material
The concentration of soluble polyphenols in allof the root materials was very small (<0.8%) butthe C-to-N ratios and the lignin contents of the ma-terials both increased in the order Centrosema,
Stylosanthes, Andropogon, Brachiaria (Table 1).Cumulative net CO2 evolution from these materialsalso showed a similar trend: the proportions of the
legume root material decomposed over 117 d wasgreater than that of Andropogon roots followed bythat of Brachiaria roots (Fig. 1). However, only
during the ®rst 3 d of incubation were there any sig-ni®cant di�erences in rates of CO2 evolution, thelegume roots decomposing faster than Andropogon
roots, which in turn decomposed faster than theBrachiaria roots. After d 3 or d 4 the rate of de-composition of all of the materials decreased expo-nentially, until after 117 d there was little CO2
evolution in any of the treatments where root ma-
Fig. 1. Cumulative net CO2 evolution (C) as a function of time over 117 d of incubation of a soil mixedwith roots of Stylosanthes guianensis, Centrosema sp., Andropogon gayanus or Brachiaria decumbens at arate of 5 g kgÿ1 of soil. Bars indicate standard errors of the means. Curves ®tted to a double exponen-
tial equation (see text). R2=coe�cient of determination
Root decomposition and soil N availability 2101
terial was added (approx. 1 mg CO2-C g soilÿ1 dÿ1)although it was still greater than that of the control
(0.34 mg CO2-C g soilÿ1 dÿ1). High coe�cients of re-gression were obtained (R2 between 0.992 and0.999) when these data were ®tted to the equation:
C � a1�1ÿ exp�ÿk1t�� � a2�1ÿ exp�ÿk2t��,where C is the total CO2-C evolved as a function oftime (t), a1 and a2 represent the size of two distinct
carbon compartments in the roots material ofdi�erent rates of decomposition, and k1 and k2 rep-resent the decomposition rate constants of these
compartments, respectively.
Immobilization of soil mineral nitrogen
Before addition of fertilizers and root material
the dry soil had a total mineral N content of40.6 mg N gÿ1, of which 32.5 mg N gÿ1 was NO3
ÿ
and 8.1 mg N gÿ1 NH4+. After the addition of fertili-
zer, root material and 10 mg N gÿ1 of 15N-labelled
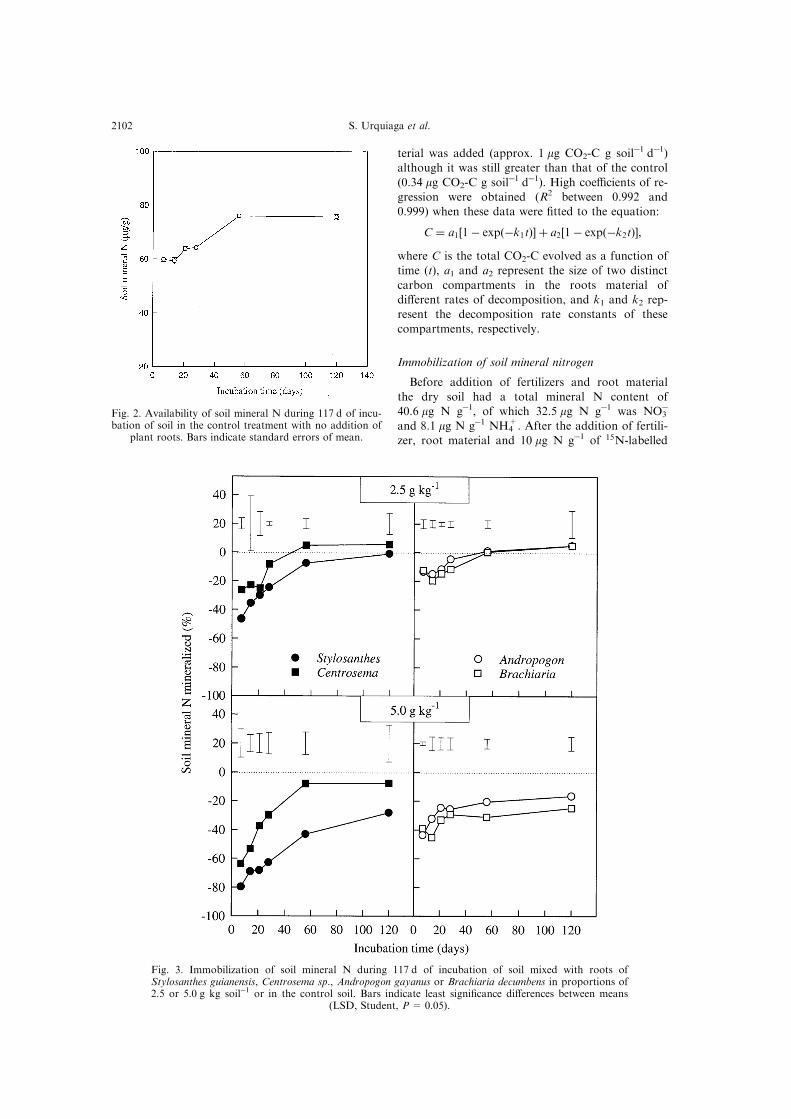
Fig. 2. Availability of soil mineral N during 117 d of incu-bation of soil in the control treatment with no addition of
plant roots. Bars indicate standard errors of mean.
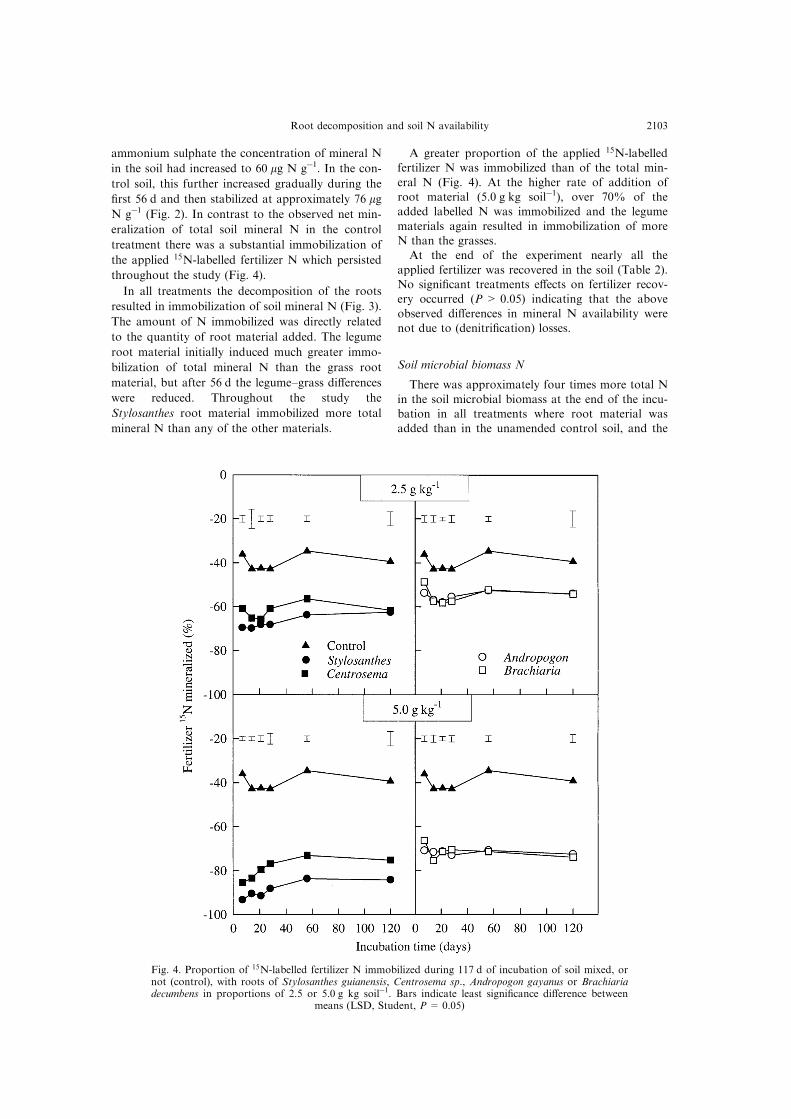
Fig. 3. Immobilization of soil mineral N during 117 d of incubation of soil mixed with roots ofStylosanthes guianensis, Centrosema sp., Andropogon gayanus or Brachiaria decumbens in proportions of2.5 or 5.0 g kg soilÿ1 or in the control soil. Bars indicate least signi®cance di�erences between means
(LSD, Student, P = 0.05).
S. Urquiaga et al.2102
ammonium sulphate the concentration of mineral N
in the soil had increased to 60 mg N gÿ1. In the con-
trol soil, this further increased gradually during the
®rst 56 d and then stabilized at approximately 76 mgN gÿ1 (Fig. 2). In contrast to the observed net min-
eralization of total soil mineral N in the control
treatment there was a substantial immobilization of
the applied 15N-labelled fertilizer N which persisted
throughout the study (Fig. 4).
In all treatments the decomposition of the roots
resulted in immobilization of soil mineral N (Fig. 3).
The amount of N immobilized was directly related
to the quantity of root material added. The legume
root material initially induced much greater immo-
bilization of total mineral N than the grass root
material, but after 56 d the legume±grass di�erences
were reduced. Throughout the study the
Stylosanthes root material immobilized more total
mineral N than any of the other materials.
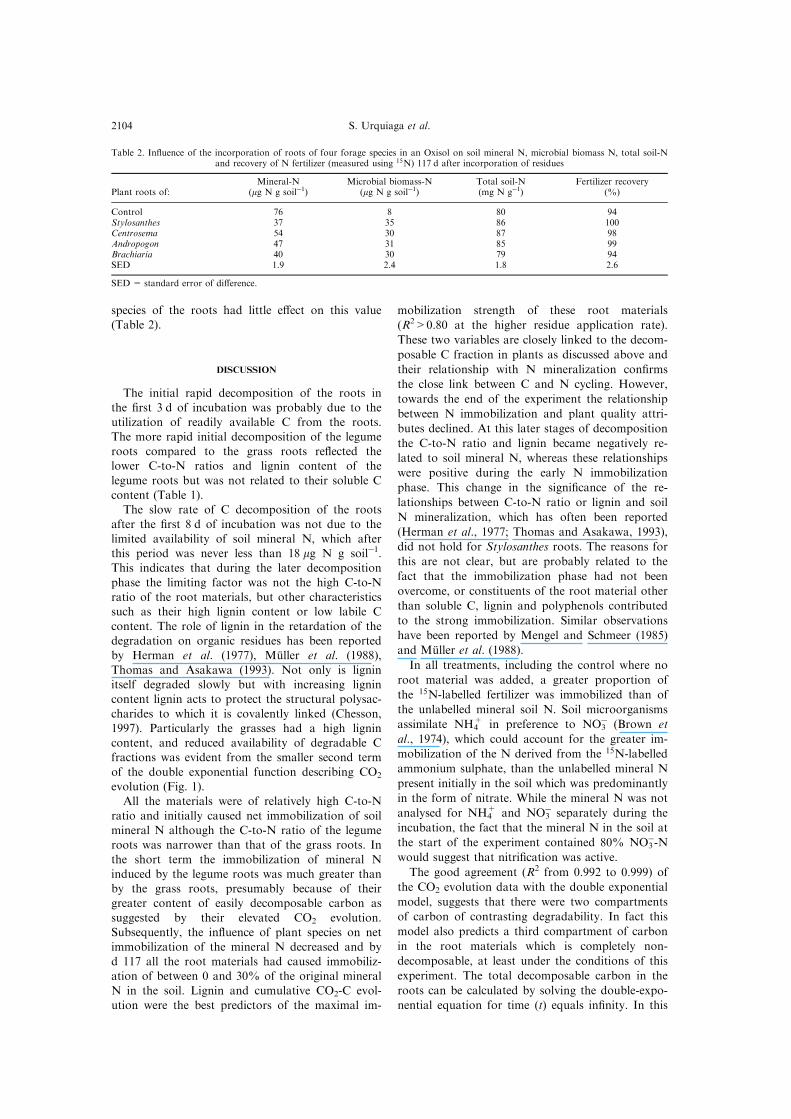
A greater proportion of the applied 15N-labelledfertilizer N was immobilized than of the total min-
eral N (Fig. 4). At the higher rate of addition ofroot material (5.0 g kg soilÿ1), over 70% of theadded labelled N was immobilized and the legume
materials again resulted in immobilization of moreN than the grasses.At the end of the experiment nearly all the
applied fertilizer was recovered in the soil (Table 2).No signi®cant treatments e�ects on fertilizer recov-ery occurred (P > 0.05) indicating that the above
observed di�erences in mineral N availability werenot due to (denitri®cation) losses.
Soil microbial biomass N
There was approximately four times more total N
in the soil microbial biomass at the end of the incu-bation in all treatments where root material wasadded than in the unamended control soil, and the
Fig. 4. Proportion of 15N-labelled fertilizer N immobilized during 117 d of incubation of soil mixed, ornot (control), with roots of Stylosanthes guianensis, Centrosema sp., Andropogon gayanus or Brachiariadecumbens in proportions of 2.5 or 5.0 g kg soilÿ1. Bars indicate least signi®cance di�erence between
means (LSD, Student, P = 0.05)
Root decomposition and soil N availability 2103
species of the roots had little e�ect on this value(Table 2).
DISCUSSION
The initial rapid decomposition of the roots inthe ®rst 3 d of incubation was probably due to theutilization of readily available C from the roots.
The more rapid initial decomposition of the legumeroots compared to the grass roots re¯ected thelower C-to-N ratios and lignin content of the
legume roots but was not related to their soluble Ccontent (Table 1).The slow rate of C decomposition of the roots
after the ®rst 8 d of incubation was not due to thelimited availability of soil mineral N, which afterthis period was never less than 18 mg N g soilÿ1.This indicates that during the later decomposition
phase the limiting factor was not the high C-to-Nratio of the root materials, but other characteristicssuch as their high lignin content or low labile C
content. The role of lignin in the retardation of thedegradation on organic residues has been reportedby Herman et al. (1977), MuÈ ller et al. (1988),
Thomas and Asakawa (1993). Not only is ligninitself degraded slowly but with increasing lignincontent lignin acts to protect the structural polysac-
charides to which it is covalently linked (Chesson,1997). Particularly the grasses had a high lignincontent, and reduced availability of degradable Cfractions was evident from the smaller second term
of the double exponential function describing CO2
evolution (Fig. 1).All the materials were of relatively high C-to-N
ratio and initially caused net immobilization of soilmineral N although the C-to-N ratio of the legumeroots was narrower than that of the grass roots. In
the short term the immobilization of mineral Ninduced by the legume roots was much greater thanby the grass roots, presumably because of theirgreater content of easily decomposable carbon as
suggested by their elevated CO2 evolution.Subsequently, the in¯uence of plant species on netimmobilization of the mineral N decreased and by
d 117 all the root materials had caused immobiliz-ation of between 0 and 30% of the original mineralN in the soil. Lignin and cumulative CO2-C evol-
ution were the best predictors of the maximal im-
mobilization strength of these root materials
(R2>0.80 at the higher residue application rate).
These two variables are closely linked to the decom-
posable C fraction in plants as discussed above and
their relationship with N mineralization con®rms
the close link between C and N cycling. However,
towards the end of the experiment the relationship
between N immobilization and plant quality attri-
butes declined. At this later stages of decomposition
the C-to-N ratio and lignin became negatively re-
lated to soil mineral N, whereas these relationships
were positive during the early N immobilization
phase. This change in the signi®cance of the re-
lationships between C-to-N ratio or lignin and soil
N mineralization, which has often been reported
(Herman et al., 1977; Thomas and Asakawa, 1993),
did not hold for Stylosanthes roots. The reasons for
this are not clear, but are probably related to the
fact that the immobilization phase had not been
overcome, or constituents of the root material other
than soluble C, lignin and polyphenols contributed
to the strong immobilization. Similar observations
have been reported by Mengel and Schmeer (1985)
and MuÈ ller et al. (1988).
In all treatments, including the control where no
root material was added, a greater proportion of
the 15N-labelled fertilizer was immobilized than of
the unlabelled mineral soil N. Soil microorganisms
assimilate NH4+ in preference to NO3
ÿ (Brown et
al., 1974), which could account for the greater im-
mobilization of the N derived from the 15N-labelled
ammonium sulphate, than the unlabelled mineral N
present initially in the soil which was predominantly
in the form of nitrate. While the mineral N was not
analysed for NH4+ and NO3
ÿ separately during the
incubation, the fact that the mineral N in the soil at
the start of the experiment contained 80% NO3ÿ-N
would suggest that nitri®cation was active.
The good agreement (R2 from 0.992 to 0.999) of
the CO2 evolution data with the double exponential
model, suggests that there were two compartments
of carbon of contrasting degradability. In fact this
model also predicts a third compartment of carbon
in the root materials which is completely non-
decomposable, at least under the conditions of this
experiment. The total decomposable carbon in the
roots can be calculated by solving the double-expo-
nential equation for time (t) equals in®nity. In this
Table 2. In¯uence of the incorporation of roots of four forage species in an Oxisol on soil mineral N, microbial biomass N, total soil-Nand recovery of N fertilizer (measured using 15N) 117 d after incorporation of residues
Plant roots of:Mineral-N
(mg N g soilÿ1)Microbial biomass-N
(mg N g soilÿ1)Total soil-N(mg N gÿ1)
Fertilizer recovery(%)
Control 76 8 80 94Stylosanthes 37 35 86 100Centrosema 54 30 87 98Andropogon 47 31 85 99Brachiaria 40 30 79 94SED 1.9 2.4 1.8 2.6
SED = standard error of di�erence.
S. Urquiaga et al.2104

case the fractions of non-decomposable carbon
become 43, 47, 54 and 62% in the Centrosema,
Stylosanthes, Andropogon and Brachiaria root ma-
terials, respectively. These large proportions of non-
decomposable root fraction occurred despite the
grinding of the root material which may have
increased the surface area for microbial attack.
These data suggest that a large proportion of root
carbon of these tropical forage species may persist
in the soil which could partly account for the large
annual accumulation of carbon observed in pastures
of Brachiaria and Andropogon spp. in Colombia by
Fisher et al. (1994). Assuming that 50% of root car-
bon is virtually undecomposable, between 4 and 28
t haÿ1 of root carbon in an improved grass±legume
pasture must be deposited additionally each year to
account for the accumulation of the 2 to 14
t haÿ1 yrÿ1 of carbon, most of it in the subsoil,
reported by Fisher et al. (1994). This is a very large
amount of root C even though a total aerial dry
matter production of approximately 22 t haÿ1 yrÿ1
by Brachiaria humidicola (not fertilized with N) has
been reported for an area in the Atlantic forest
region of Southern Bahia, Brazil (Boddey et al.,
1995). Few data on in situ root production and
turnover are available but current estimates of root
biomass in tropical pastures are around 2 to 6
t haÿ1 (Cadisch et al., 1994; Rao et al., 1996) which
implies that root turnover must be extremely rapid
to account for the annual C deposition rates
reported by Fisher et al. (1994).
While the very slow turnover of a large fraction
of the organic matter of the root material leads to
the conservation or accumulation of carbon in the
soil, the immobilization of available soil mineral N
caused by the decomposing roots, as well as the
actual N sequestered in the non-degradable fraction
of this organic matter, can result in extremely small
concentrations of plant-available N in the long
term. Studies in Australia on Panicum maximum
pastures on the Vertisols of Brigalow regions of
Queensland, suggest that the immobilization of N
caused by the deposition of litter and root materials
of high C-to-N ratio are the principal causes of pas-
ture decline in this region (Robbins et al., 1989;
Robertson et al., 1993a,b). Our results suggest that
this may also be an important factor in the
observed decline, or degradation, of Brachiaria pas-
tures in Brazil and other areas of tropical South
America (Toledo, 1985; Boddey et al., 1996). As the
roots were ground, our ®ndings give estimates of
the N immobilization potentials of the root ma-
terials which may be moderated under ®eld con-
ditions. Nevertheless the contribution which
decomposing legume roots can make to alleviate N
de®ciencies are negligible (and probably negative) at
least in the short-term. Thus the positive contri-
bution of legumes to N cycling is largely through
recycling of above-ground residues.
In the ®eld, roots would be exposed to more bio-tic and abiotic factors than in this study. Amongst
these may be the cycles of wetting and drying,physical disturbance and their consumption of theroot material by macrofauna, especially termites,
which are all important factors in soil organic mat-ter turnover in tropical soils (Lavelle et al., 1992).The in¯uences of these factors on decomposition of
roots and litter of tropical forage species constituteimportant priorities for further research.
AcknowledgementsÐWe thank the Brazilian NationalResearch Council (CNPq) for Research Fellowships for S.Urquiaga and R. M. Boddey and Carlos AragaÄ o and JoseÂCarlos C. dos Santos for their technical assistance inchemical analysis of plant roots. Part of this work wasfunded by the International Scienti®c Cooperation pro-gramme of the EEC.
REFERENCES
Amato M. and Ladd J. N. (1988) Assay for microbial bio-mass based on ninhydrin-reactive nitrogen in extracts offumigated soils. Soil Biology and Biochemistry 20, 107±114.
Anderson J. M., Proctor J. and Vallack H. W. (1983)Ecological studies in four contrasting lowland rain for-est in Gunung Mulu national park, Sarawak. Journal ofEcology 71, 503±527.
Anderson J. M. and Ingram J. S. I. (1989) Tropical SoilBiology and Fertility: a Handbook of Methods. CABInternational, Wallingford.
Badaruddin M. and Meyer D. W. (1990) Green-manurelegume e�ects on soil nitrogen, grain yield, and nitrogennutrition of wheat. Crop Science 30, 819±825.
Biederbeck V. O., Campbell C. A., Bowren K. E. andMcIver R. N. (1980) E�ect of burning cereal straw onsoil properties and grain yields in Saskatchewan. SoilScience Society of America Journal 44, 103±111.
Boddey R. M., Rao I. M. and Thomas R. J. (1996)Nutrient cycling and environmental impact ofBrachiaria pastures. In The Biology Agronomy andImprovement of Brachiaria, eds. J. W. Miles, B. L.Maass and C. B. do Valle, pp. 72±86. CIAT, Cali,Colombia and EMBRAPA, Campo Grande, Brazil.
Boddey R. M., de Resende C. P., Pereira J. M.,Cantarutti R. B., Alves B. J. R., Ferreira E., RichterM., Cadisch G. and Urquiaga S. (1995) The nitrogencycle in pure grass and grass/legume pastures: evalu-ation of pasture sustainability. In Nuclear Techniques inSoil±Plant Studies for Sustainable Agriculture andEnvironmental Preservation, eds. FAO/IAEA, pp 307±319. FAO/IAEA, Vienna.
Brown C. M., MacDonald-Brown D. S. and Meers J.L. (1974) Physiological aspects of microbial inorganicmetabolism. Advances in Microbial Physiology 11, 1±52.
Cadisch G., Giller K. E., Urquiaga S., Miranda C. H. B.,Boddey R. M. and Schunke R. M. (1994) Does phos-phorus supply enhance soil-N mineralization inBrazilian pastures? European Journal of Agronomy 3,339±345.
Cadisch G. and Giller K. E. (1997) Driven by Nature:Plant Litter Quality and Decomposition. CABInternational, Wallingford.
Chesson A. (1997) Plant degradation by ruminants: paral-lels with litter decomposition in soils. In Driven byNature: Plant Litter Quality and Decomposition, eds. G.Cadisch and K. E. Giller, pp. 47±66. CABInternational, Wallingford.
Root decomposition and soil N availability 2105

Fisher M. J., Rao I. M., Ayarza M. A., Lascano C. E.,Sanz J. I., Thomas R. J. and Vera R. R. (1994) Carbonstorage by introduced deep-rooted grasses in the SouthAmerican savannas. Nature 371, 15.
Fox R. H., Myers R. J. K. and Vallis I. (1990) The nitro-gen mineralization rate of legume residues in soils asin¯uenced by their polyphenol, lignin and nitrogen con-tents. Plant and Soil 129, 251±259.
Halsall D. M., Leigh J. H., Gollasch S. E. and HolgateM. D. (1995) The role of allelopathy in legume declinein pastures. II. Comparative e�ects of pasture, crop andweed residues on germination, nodulation and rootgrowth. Australian Journal of Agriculture Research 46,189±207.
Handayanto E., Cadisch G. and Giller K. E. (1994)Nitrogen release from prunings of legume hedgerowtrees in relation to quality of the prunings and incu-bation method. Plant and Soil 160, 237±248.
Handayanto E., Cadisch G. and Giller K. E. (1995)Manipulation of quality and mineralization of tropicallegume tree prunings by varying nitrogen supply. Plantand Soil 176, 149±160.
Handayanto E., Giller K. E. and Cadisch G. (1997)Regulating N release from legume tree prunings by mix-ing residues of di�erent quality. Soil Biology andBiochemistry 29, 1417±1426.
Herman W. A., McGill W. B. and Dormaar J. F. (1977)E�ects of initial chemical composition on decompositionof roots of three grass species. Canadian Journal of SoilScience 57, 205±215.
Keeney D. R. and Nelson D. W. (1982) Nitrogen ± inor-ganic forms. In Methods of Soil Analysis, Part 2.Chemical and Microbiological Properties, eds. A. L.Page, R. H. Miller and D. R. Keeney, pp. 643±698.American Society of Agronomy, Madison, WI.
Ladd J. N., Oades J. M. and Amato M. (1981a)Distribution and recovery of nitrogen from legume resi-dues decomposing in soils sown to wheat in the ®eld.Soil Biology and Biochemistry 13, 252±256.
Ladd J. N., Oades J. M. and Amato M. (1981b) Microbialbiomass formed from 14C, 15N-labelled plant materialdecomposing in soils in the ®eld. Soil Biology andBiochemistry 13, 119±126.
Lavelle P., Spain A., Blanchart E., Martin A., Martin S.and Schaefer R. (1992) Impact of soil fauna on theproperties of soils in the humid tropics. In Myths andScience of Soils of the Tropics, eds. R. Lal and P.Sanchez, pp. 157±185. Soil Science Society of America,Madison.
Leigh J. H., Halsall D. M. and Holgate M. D. (1995) Therole of allelopathy in legume decline in pastures. I.E�ects of pasture and crop residues on germination andsurvival of subterranean clover in the ®eld and nursery.Australian Journal of Agriculture Research 46, 179±188.
Melillo J. M. and Aber J. D. (1984) Nutrient immobiliz-ation in decaying litter: an example of carbon nutrientinteractions. In Trends in Ecological Research for the1980s, eds. J. H. Cooley and F. B. Golley, pp. 193±215.Plenum, New York.
Mengel K. and Schmeer H. (1985) E�ect of straw, cellu-lose and lignin on the turnover and availability oflabelled ammonium nitrate. Biology and Fertility of Soils1, 175±181.
MuÈ ller M. M., Sundman V., Soininvaara O. andMerilainen A. (1988) E�ect of chemical composition onthe release of nitrogen from agricultural plant materialsdecomposing in soil under ®eld conditions. Biology andFertility of Soils 6, 78±83.
Myers R. J. K. and Paul E. A. (1971) Plant uptake andimmobilization of 15N-labelled ammonium nitrate in a®eld experiment with wheat. In Nitrogen-15 in Soil Plant
Studies, ed. IAEA, pp. 55±64. International AtomicEnergy Agency, Vienna.
de Oliveira O. C., Urquiaga S. and Boddey R. M. (1994)Burning cane: the long-term e�ects. International SugarJournal 96, 272±274.
Palm C. A. and Sanchez P. A. (1991) Nitrogen releasefrom the leaves of some tropical legumes as a�ected bytheir lignin and polyphenolic contents. Soil Biology andBiochemistry 23, 83±88.
Ramos D. C., Castro A. F. and de Camargo M. N. (1973)Levantamento detalhado de solos da a rea daUniversidade Federal Rural de Rio de Janeiro. PesquisaAgropecuaria Brasileira 8, 1±27.
Rao I. M., Ayarza M. A., and Herrera P. (1996) Root dis-tribution and production in native and introduced pas-tures in the South American savannas. In Proceedings ofthe 5th Symposium of the International Society of RootResearch, ed. J. E. Box. Kluwer, Dordrecht.
Robbins G. B., Bushell J. J. and McKeon G. M. (1989)Nitrogen immobilization in decomposing litter contrib-utes to productivity decline in ageing pastures of greenpanic (Panicum maximum var. trichoglume). Journal ofAgricultural Science, Cambridge 113, 401±406.
Robertson F. A., Myers R. J. K. and Sa�gna P.G. (1993a) Distribution of carbon and nitrogen in along-term cropping system and permanent pasture sys-tem. Australian Journal of Agricultural Research 44,1323±1336.
Robertson F. A., Myers R. J. K. and Sa�gna P.G. (1993b) Carbon and nitrogen mineralization in culti-vated and grassland soils in subtropical Queensland.Australian Journal of Soil Research 31, 611±619.
Sivapalan K., Fernando V. and Thenabadu M. W. (1985)N-mineralization in polyphenol-rich plant residues andtheir e�ect on nitri®cation of applied ammonium sul-phate. Soil Biology and Biochemistry 4, 547±551.
Terman G. L. and Brown M. A. (1968) Uptake of fertili-zer and soil nitrogen by ryegrass, as a�ected by carbon-aceous residues. Soil Science Society of AmericaProceedings 32, 86±90.
Thomas R. J. and Asakawa N. M. (1993) Decompositionof leaf litter from tropical forage grasses and legumes.Soil Biology and Biochemistry 25, 1351±1361.
Toledo J. M. (1985) Pasture development for cattle pro-duction in the major ecosystems of the tropicalAmerican lowlands. In XI International GrasslandCongress, pp. 74±81. Japanese Grassland Society,Kyoto, Japan.
Tomar J. S. and Soper R. J. (1981) Fate of tagged urea Nin the ®eld with di�erent methods of N and organicmatter placement. Agronomy Journal 73, 991±995.
Urquiaga S., Libardi P. L., Reichard K., Padovese P.,Moraes S. and Victoria L. R. (1982) Estudo da minera-lizac° aÄ o do 15N orgaà nico num Oxisol (EL) e sua absor-c° aÄ o por uma graminea (Melinis minuti¯ora Beauv). InColoquio Regional Sobre Materia Organica do Solo, eds.C. C. Cerri, D. Athie and D. Sodrzeieski, pp. 197±207.Promocet, Sao Paulo.
Urquiaga S., Giller K. E. and Cadisch G. (1994) Tracingmechanisms of nitrogen transfer from legume to grassin tropical pastures. In Soil Management in SustainableAgriculture, eds. H. Lee and H. Cook, pp. 104±112.Wye College, Wye.
Vallis I. and Jones R. J. (1973) Net mineralization ofnitrogen in leaves and leaf litter of Desmodium intortumand Phaseolus atropurpureus mixed with soil. SoilBiology and Biochemistry 5, 391±398.
Van Soest P. J. and Wine R. H. (1968) Determination oflignin and cellulose in acid-detergent ®ber with per-manganate. Journal of the Association of O�cialAnalytical Chemists 51, 780±785.
S. Urquiaga et al.2106
Copyright © 2022 FDOKUMEN




















