
Use of forage and plant supplements in organic and free ...
187
Use of forage and plant supplements in organic and free range broiler systems: Implications for production and parasite infections Ph.D. Thesis by GUSTAVO FONSECA DE ALMEIDA DENMARK 2012
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Use of forage and plant supplements in organic and free ...

Use of forage and plant supplements in organic and free range
broiler systems: Implications for production and parasite infections
Ph.D. Thesis by
GUSTAVO FONSECA DE ALMEIDA
DENMARK
2012

Thesis supervisors Principal Supervisor Head of Research Unit John Erik Hermansen Department of Agroecology, Aarhus University Project Supervisor Scientist Klaus Horsted Department of Agroecology, Aarhus University Co-supervisor Professor Stig Milan Thamsborg Danish Centre for Experimental Parasitology, University of Copenhagen Members of the Assessment Committee Senior Scientist Jørgen Eriksen, Aarhus University Dipl. Ing.-Agr. ETH, PhD Veronika Maurer, Res. Inst. of Organic Agriculture (FIBL) Prof. Dr. Dipl. EVPC Arwid Daugschies, University Leipzig

i
Summary
Increased foraging by broilers could improve the use of local resources and support farmers in
conversion to 100% reliance on organic ingredients and thus reduce environmental burdens and
production costs. At same time, however, foraging exposes broilers to parasitic infections.
In this Ph.D. thesis, it was hypothesized that slow-growing broilers restricted in access to
commercial feed were capable of ingesting considerable amounts of herbage and other feed
items available in the range to partially cover their nutritional requirements. In addition, due to a
higher exposure to parasites when raised outdoors, it was hypothesized that the use of plant
extracts under different forms of delivery could be efficient in prevention of coccidiosis and thus
improve broilers performance in production systems where the prophylactic use of chemical
drugs is forbidden.
On this background, the main objective of this study was to investigate the potential contribution
of two different types of vegetation offered to broilers, and to investigate how plants with anti-
parasitic components could reduce parasitic infections and boost performance attributes in free
range and organic production systems.
Two experiments were carried out in Denmark and two in Brazil. In Denmark, a dietary
supplementation of Artemisia annua to free range broilers naturally infected with Eimeria spp.
oocysts was investigated to estimate its effects on performance and infection dynamics. Two
broiler genotypes (White Bresse L40 - a pure slow-growing and Kosmos 8R - a hybrid medium-
growing) were used. In the second experiment, feed intake from two different types of highly
nutritious vegetation (grass/clover vs chicory) and foraging behaviour were investigated to
identify good husbandry practices and to estimate the nutritional contribution of forage in the
finishing growing period of both slow and medium growing broilers.
In Brazil, an on-farm experiment was carried out to investigate the effects of Manihot esculenta,
Artemisia annua and A. vulgaris under two different strategies of delivery (in water and mixed in
feed). In the second experiment conducted in Brazil, three doses of a mixture of A. annua and
Curcuma longa ethanolic extracts in prophylactic treatments were supplemented in water against
coccidiosis. Finally, an exploratory field work was carried out in Brazil on three groups of

ii
farmers producing broilers in alternative systems to investigate developmental potentials with
novel feeding strategies and use of herbal extracts to reduce severity of coccidiosis.
The type of vegetation provided to broilers did not influence the use of the free-range area, and
neither did it affect the performance attributes. Differences in the use of the range area, activity
level and feed intake were observed in relation to genotype, sex, age and also the time of day.
Foraging activity was positively correlated with age. In general, hybrid broilers (medium
growing) spent more time inside and closer to the broiler houses during the day with increased
foraging activity during evenings. In contrast, slow-growing pure breed showed a more uniform
activity during the day. Consumption of forage and beneath ground biomass was able to
substitute 10 to 15% of the daily methionine and 10% of energy requirements for growth and
maintenance in the finishing period.
With respect to the use of herbal extracts to alleviate the problems caused by coccidiosis, the
supplementation of Artemisia annua dried leaves in feed reduced oocyst output. However, the
bitter taste of the leaves inhibited a higher feed intake and thus reduced the capacity for a higher
weight gain. In addition, the manipulation of the raw plant materials was labour-intensive and its
implementation is not practical at farm level. The supplementation with M. esculenta dried
leaves in the feed in the beginning of the rearing period also reduced oocyst output, but
manipulation similarly labour-intensive, which may limit its use by farmers. Based on the
knowledge of an organic farmer, the use of purified extracts in drinking water was investigated
and promising results were found for mixing extracts of A. annua and C. longa in water. Further
research with pure botanicals from these plants and with different plant mixtures to treat avian
coccidiosis is a natural continuation for this project. The study concludes that limitation of
supplemented commercial feed in the finishing period can be used for slow-growing broilers that
have access to highly nutritious vegetation in order to stimulate the appropriate use of the
outdoor area. However, the nutritional contribution is foreseen to be modest. It is further
concluded that the use of ethanolic extracts in drinking water to prevent losses from coccidiosis
seems promising in relation to its effects and practical application.
Research Centre Foulum, October 2012.
Gustavo Fonseca de Almeida

iii
Sammendrag (Danish summary)
Hvis en større andel af økologiske slagtekyllingers ernæringsmæssige behov kan dækkes gennem
fouragering vil det kunne bidrage til at øge anvendelsen af lokale ressourcer og samtidig
reducere miljøbelastningen og produktionsomkostninger. Samtidig betyder øget fouragering dog
også at slagtekyllinger i højere grad udsættes for parasit-infektioner.
I denne ph.d.-afhandling, blev der arbejdet ud fra den hypotese, at langsomt voksende
slagtekyllinger, der kun fik begrænsede mængder kommercielt foder, er i stand til at indtage
betydelige mængder af plantemateriale og andre foderemner fra udearealet til delvis dækning af
deres ernæringsmæssige behov. Samtidig var hypotesen at den højere eksponering for parasitter
(coccidier) i udearealet kunne modvirkes ved brug af planteaktive stoffer der forebygger
coccidiose. Herigennem kunne produktionen forbedres i disse systemer, hvor den profylaktiske
anvendelse af medicin ikke er tilladt. På denne baggrund var det vigtigste formål med projektet
at undersøge det potentielle ernæringsmæssige bidrag fra to forskellige typer af vegetation, der
tilbydes til slagtekyllinger i udearealet, samt at undersøge hvordan planter eller planteekstrakter
med anti-parasitiske komponenter ville reducere coccidieinfektioner og forbedre produktionen i
alternative produktionssystemer.
Der blev gennemført to eksperimenter i Danmark og andre to i Brasilien. I Danmark blev
betydningen af et tilskud af Artemisia annua i foderet til slagtekyllinger, der var inficeret med
Eimeria spp. Oocyster, undersøgt for at vurdere virkninger på tilvækst og infektionsforløb.
Forsøget blev gennemført med to forskellige genotyper af slagtekyllinger – en medium voksende
krydsning (Kosmos 8) fra avlsfirmaet ’Olandia’ og en langsomt voksende ren race (Bresse).
Endvidere blev foderoptagelse og fourageringsadfærd undersøgt fra et udereal med to forskellige
typer vegetation (kløvergræs vs cikorie) for at estimere det ernæringsmæssige bidrag heraf.
I Brasilien blev der hos en økologisk kyllingeproducent gennemført et forsøg for at undersøge
virkningerne af Manihot esculenta, Artemisia annua og A. vulgaris tildelt enten iblandet foder
eller som ekstrakt tildelt i vand. Endvidere blev der på en forsøgsstation undersøgt betydningen
af tre doser af en blanding af A . annua og Curcuma longa ethanolbaserede ekstrakter i vand til
slagtekyllinger. Endelig blev der i Brasilien udført et feltarbejde med tre grupper af familiebrug,
der producerer slagtekyllinger i alternative systemer. Der blev i disse systemer undersøgt

iv
udviklingsmuligheder i forbindelse med nye fodringsstrategier og potentiel anvendelse af
urteekstrakter for at reducere coccidiose.
Typen af vegetation i udearealet i det danske forsøg påvirkede ikke slagtekyllingernes brug af
arealet eller deres produktion. Brugen af udearealet, aktivitetsniveau og foderoptagelse afhang af
genotype, køn, alder samt tidspunktet på dagen. Fourageringsaktiviteten var positivt korreleret
med alderen. Generelt brugte Kosmos 8 kyllingerne (medium voksende) mere tid indenfor og
tættere på kyllingehuset i løbet af dagen og havde en øget fourageringsaktivitet om aftenen. I
modsætning hertil havde den langsomt voksende rene race en mere ensartet aktivitet i løbet af
dagen. I gennemsnit var indtaget af planter og biomasse i jorden i stand til at dække 10 til 15% af
methioninbehovet og 10% af energibehovet til vækst og vedligeholdelse i den sidste del af
produktionsperioden.
Med hensyn til anvendelsen af urter og urteekstrakter til forebyggelse af coccidiose medførte et
tilskud af tørrede Artemisia annua blade i foderet en reduceret oocyst udskillelse, men den bitre
smag af bladene hæmmede foderoptagelsen og dermed tilvæksten. Desuden var bearbejdningen
af de rå plantematerialer arbejdskrævende, og forekommer ikke umiddelbart praktisk at
gennemføre på bedriftsniveau. Tilskud af tørrede M. esculenta blade i foderet i begyndelsen af
opdrætningsperioden reducerede også oocyst udskillelsen, men også her var operationen
arbejdskrævende, hvilket kan begrænse brugen i praksis. Brugen af oprensede planteekstrakter i
drikkevand viste derimod meget lovende resultater – især en kombination af ekstrakter af A.
annua og C. longa i vand. Det konkluderes, at reduceret tildeling af suppleringsfoder i den
afsluttende vokseperiode under forudsætning af adgang til et ernæringsrigt udeareal er en
mulighed for slagtekyllinger til at udnytte disse næringsstoffer, men den samlede betydning for
foderforbruget er kun af mindre betydning. Det konkluderes endvidere, at anvendelse på
ethanolbaserede ekstrakter i drikkevand kan anvendes til forebyggelse af coccidiose i økologisk
slagtekyllingeproduktion i praksis.
Forskningscenter, Foulum, Oktober 2012.
Gustavo Fonseca de Almeida

v
Preface
This thesis is submitted in partial fulfilment of the requirements for the Doctor of Philosophy
(Ph.D.) in Agroecology at the Faculty of Science and Technology, Aarhus University, Denmark.
In this three-year Ph.D. project, three different broiler genotypes, two types of vegetation and
four different herbal supplements with potential anti-coccidial components were investigated.
Supplementation of herbal extracts to chickens in Denmark and in Brazil was undertaken in close
collaboration with scientists from University of Campinas (UNICAMP), University of São Paulo
(USP) in Brazil and the Appalachian Farming Systems Research Center (USDA-ARS) in the U.S.
The research presented in this thesis was compiled from results of two experiments performed in
Denmark (2010) and two performed in Brazil (2011). In addition, an exploratory field study was
carried out on small-scale farms in two different locations in the South of Brazil in a close
collaboration with scientists from EMBRAPA Swine and Poultry Institute and Federal
University of Paraná (UFPR). Interviews and questionnaires were also employed for farmers
linked to a broiler company in the State of São Paulo. In Denmark, a strong inter-departmental
collaboration supported the experiments carried out at Research Centre Foulum, Faculty of
Science and Technology, Aarhus University and I was trained in methods for monitoring
coccidial infections and for identifying endo-parasites in chickens in the Danish Centre for
Experimental Parasitology, Faculty of Health and Medical Sciences, University of Copenhagen.
The head of research unit in the Department of Agroecology, Faculty of Science and Technology,
John Erik Hermansen, was my principal supervisor, while scientist Klaus Horsted from the same
research group acted as my project supervisor. Professor Stig Milan Thamsborg from the Danish
Centre for Experimental Parasitology, Copenhagen University acted as my co-supervisor.
The thesis is supported by two published papers, two submitted manuscripts and one manuscript
in preparation:
I. Almeida, G.F.d.; Horsted, K.; Thamsborg, S.M.; Kyvsgaard N.C.; Ferreira J.F.S.;
Hermansen, J.E. Use of Artemisia annua as a natural coccidiostat in free-range broilers
and its effects on infection dynamics and performance. Veterinary Parasitology, 186
(2012) 178– 187.

vi
II. Almeida, G.F.d.; Hinrichsen, L.K.; Horsted, K.; Thamsborg, S.M. and Hermansen, J.E.
Feed intake and activity level of two broiler genotypes foraging different types of
vegetation in the finishing period. Poultry Science, 91 (2012) 2105– 2113.
III. Almeida, G.F.d.; Thamsborg, S.M.; Magalhães P.M.; Ferreira J.F.S.; Horsted, K.;
Hermansen, J.E. The effects of cassava (Manihot esculenta), annual wormwood
(Artemisia annua) and mugwort (A. vulgaris) on coccidiosis in organically reared pullets
in Brazil. (Manuscript submitted to Asian-Australasian Journal of Animal Sciences).
IV. Almeida, G.F.d.; Thamsborg, S.M.; Madeira, A.M.B.N.; Ferreira J.F.S.; Magalhães, P.M.;
Demattê Filho, L.C.; Horsted, K.; Hermansen, J.E. The effects of combining Artemisia
annua and Curcuma longa ethanolic extracts to broilers challenged with infective oocysts
of Eimeria acervulina and E. maxima. (Manuscript submitted to Parasitology).
V. Almeida, G.F.d.; Figueiredo, E.A.; Molento, M.B.; Demattê Filho, L.C.; Horsted, K.;
Thamsborg, S.M.; Hermansen, J.E. Farmer´s attitudes and perceptions on feeding
strategies and control of coccidiosis in alternative broilers systems in Brazil. (Manuscript
in preparation for submission to Organic Agriculture).
Furthermore, a network project among scientists in Denmark and Brazil and linked to this Ph.D.
project was supported in 2011 by the Danish Ministry of Science Technology and Innovation.
Workshops on novel strategies for livestock production in Denmark and in Brazil were
conducted in both countries (2011) as part of the dissemination activities of this Ph.D. project.
Finally, two oral presentations at an International workshop were given in Denmark in January
2012, while two additional papers were presented at the IFOAM Livestock International
Conference held in Hamburg, Germany, in September 2012.
Research Centre Foulum, October 2012.
Gustavo Fonseca de Almeida

vii
Acknowledgements
xxx

viii
Table of contents
Summary................................................................................................................................... i Sammendrag (Danish Summary)..................................................................................... iii Preface........................................................................................................................................ v Acknowledgements…………................................................................................................ vii List of Figures.......................................................................................................................... ix List of Tables........................................................................................................................... x 1. Introduction....................................................................................................................... 1 1.1 Background........................................................................................................................... 1 1.2 Production systems and its regulations................................................................................. 5 1.3 Nutrient supply by foraging…..…………............................................................................ 14 1.4 Four herbal food supplements and their anti-parasitic properties......................................... 17 1.5 Coccidiosis: relevance in the broiler industry worldwide and the Eimeria spp. life cycle... 20 1.6 Genotypes for different productions systems…………………………………………........ 25 1.7Animal ethics: behaviour as a means of animal welfare assessment in different production systems..........................................................................................................................................
29
1.8 Project justifications, objectives, hypothesis and main research questions.......................... 32 1.9 Outline of the thesis and supporting papers.......................................................................... 35 2. Materials and methods................................................................................................... 36 2.2 Description of methods......................................................................................................... 36 2.2.1 Protocols of experiments performed in Denmark and in Brazil................................... 36 2.2.2 Parasitological methods and animal experimentation……………............................... 38 2.2.3 Analysis of crop content of broilers and sward harvesting ……….............................. 43 2.2.4 The ethanolic extraction of plant components…………………….............................. 46 2.2.5 Investigating farmer’s opinions.................................................................................... 47 3. General results and discussions................................................................................... 48 3.1 Main results from Individual papers..................................................................................... 48 3.2 Turning major results into practical husbandry.................................................................... 53 3.3 Implications for the development of the organic meat sector............................................... 57 4. Conclusions......................................................................................................................... 60 5. Future Outlook.................................................................................................................. 62 6. References........................................................................................................................... 65 7. Supporting papers............................................................................................................ 78 7.1 Use of Artemisia annua as a natural coccidiostat in free-range broilers…………………... 79 7.2 Feed intake and activity level of two broiler genotypes…………………………………... 90 7.3 On-farm study of the effects of cassava, wormwood and mugwort against coccidiosis….. 100 7.4 Combination of Artemisia annua and Curcuma longa ethanolic extracts………………… 126 7.5 Farmer´s attitudes and perceptions on feeding strategies and control of coccidiosis……... 152

ix
List of Figures
Figure 1. Evolution of organic poultry production in Denmark - Layer hens for eggs and broilers for meat with expected market shares for 2015. (Modified after Hermansen, 2011)…..
4
Figure 2. The dilemma cost vs quality – Important factors in decision making………………..
5
Figure 3. Chickens foraging for different ingredients in a Danish free range system………….
15
Figure 4. Starting from the left side: Schematic presentation of Artemisia annua leaves; A. vulgaris leaves and inflorescence; leaves, roots and powder of Curcuma longa and Manihot esculenta foliage………………………………………………………………………………...
18 Figure 5. Eimeria spp. life cycle (Modified after USDA, 2005)……………………………….
22
Figure 6. From the left: A pure slow-growing broiler, a hybrid double-purpose genotype and a fast-growing genotype…………………………………………...…………………………….
28 Figure 7. Oocysts visualised in the McMaster slides used for estimating OPG (100x magnitude) ………………………………………………………………………………............
40
Figure 8. Oocyst visualised at 400x magnitude. ………………………………………............
40
Figure 9. McMaster Chamber and the strategies used for counting oocysts. …………………..
41
Figure 10. Output from the PCR test.………………………………….….................................
42
Figure 11. Content of crops separated into different fractions. From top left: “earthworms, larvae and pupae”; “grass”; “clover”; “chicory”; “insects”; “seeds”; “soil”; “commercial broiler feed”; “grit stones” and “wheat”..………………………………….................................
44 Figure 12. Harvesting biomass for chemical analysis. ………………….................................
45
Figure 13. Price of chicken meat in U$ dollars/Kg from different production systems and the relation price vs system orientation.…………………………………..........................................
58 Figure 14. Slow-growing chickens foraging in plots with fruit trees...........................................
63
Figure 15. Slow-growing chickens managed in movable houses. ..............................................
64

x
List of Tables
Table 1. Production of chicken meat (Million tonnes) and market share by region in 1970, 1990 and 2010 (for six continents and five important production/consumer countries) and per capita consumption 2007, kg……………………………..……….…………………………….
2
Table 2. International regulations on the rearing and marketing of different types of broiler chickens. modified from Nielsen et al. (2003) and according to European regulations Nos. 1538/91, 2092/91, 2891/93 and 436/2001; the French decree from 12.03.1996 for Label Rouge regulation; Coelho et al., (2008) and Brasil (2011) for the Brazilian regulations. Fanatico et al., (2009) for the U.S. regulations on alternative and organic broilers…………….
6
Table 3. Characteristics of sporulation pre-patent period and production of oocysts of Eimeria Acervulina, E. maxima and E. tenella. Modified from Anon (1990)……………………………
23

1
1. Introduction
Meat from chickens is considered a viable source of proteins to humans because it is easy and
fast to produce and has a smaller environmental footprint than other types of meat (de Vries and
de Boer, 2010). However, scaling up the production of chickens has revealed a number of
limitations in animal welfare and quality attributes, especially due to sustainability issues
(Vermeir and Verbeke, 2006). Alternative methods of chicken production have been investigated
to tackle these problems. However, even in alternative production systems there is still a large
dependence on oil seed crops and cereals for feeding the birds, which competes with the use of
the crops for human food resources. This Ph.D. project investigated alternative systems for
feeding chickens and for reducing problems in production related to parasitic infections. This
chapter gives a status of the world market of chicken meat, the differences of the alternative
production systems and the aspects influencing the use of free-range areas, infections caused by
parasites, differences in genetics and animal welfare, all supporting the justifications for this
study and thus leading to the objectives and research questions of this thesis.
1.1 Background
For centuries chickens (Gallus gallus domesticus) have been raised for the production of eggs
and meat for human consumption. From indigenous, and less productive, double-purpose
genotypes, modern breeding programmes have emphasized productivity: either layers for the
production of eggs or broilers for the production of meat (Tixer-Boichard et al., 2012). As a
result of intensive selection, the production of broilers has seen an extraordinary rise in the last
40 years, and in 2010 more than 86 million tonnes of chicken meat was produced worldwide
(FAOstat, 2012a). The United States leads in terms of annual production and consumption, with
a market share of 20% and an annual per capita consumption of more than 50 kg of meat (Table
1). China has doubled its production capacity in the last 20 years and is currently the second-
largest producer at approximately 12 million tonnes with a world market share of 14%. China’s
consumption pattern is different from that in the U.S. with an annual consumption of less than 12
kg per person, and in India the consumption is as low as 0.6 kg chicken meat/person/year. It is

2
expected that the two largest populations in the world (China and India) and the African
continent will experience a huge increase in production and consumption of chicken meat
(Kearney, 2010) in the medium to long term.
Table 1. Production of chicken meat (million tonnes) and market share by region in 1970, 1990 and 2010 (for six continents and five important production/consumer countries) and per capita consumption 2007, kg.
1970 1990 2010 Cons. Per
Production % market Production % market Production % market Capita Africa 543 4 1849 5 4369 5 4.5 N America 4180 32 9265 26 18020 21 49.5 S America 849 6 3782 11 16313 19 27.2 Asia 2247 17 8759 25 28642 33 7.8 Europe 4843 37 10162 29 13766 16 20.3 Oceania 134 1 454 1 1048 1 37.2 U.S. 3840 29 8667 24 16971 20 50.7 China 649 5 2663 7 11841 14 11.8 Brazil 366 3 2356 7 10693 12 31.7 India 81 0.6 531 1.5 2300 2.7 0.6 Denmark 70 0.5 126 0.4 168 0.2 18.2 World 13140 100 35585 100 86205 100 12.6 Source: Production data FAOstat 2012a; Consumption data FAOstat, 2012b.
As presented in Table 1, Brazil was the third largest producer accounting for 12% of global and
65% of South American production in 2010, and together with the U.S. and China produced 46%
of all chicken meat in the world.
The introduction of new technologies for intensive and standardised systems and the
modernisation in crop production has favoured South American and Asian countries due to
cheaper labour and availability of feed ingredients (ABEF, 2012). In Denmark, for example, the
production of broilers has more than doubled in the last 40 years, but its market share has
dropped in a similar proportion to that for Europe combined (Table 1).
In Denmark, the broiler industry is concentrated on a few big farms with flocks being managed
in total confinement with improved cleaning and disinfection procedures. Production systems are
conducted according to the “all in – all out” principle (Permin, 1997). From 1970 to 1990, the

3
production surpluses were exported, especially to Middle East countries (Anon, 2012a).
However, competition from Brazil in the 1990s led Denmark to turn to meet other market
demands by adding value to its products. In 2010, Denmark exported salmonella-free chicken
meat to Sweden, UK, Germany and Latvia (Anon, 2012a).
Many examples of new business orientations exist in developed countries and perhaps it can be
described as a world trend related to urbanisation and increased consumer demands linked to
high education and easy access to information (Castellini et al., 2008). Adding value to products
and thus benefiting from consumers’ willingness to pay premium prices for high quality and safe
food products has been followed by developing countries, but to a lesser extent (Crabone et al.,
2005).
Francisco et al. (2007), studying consumer perceptions in metropolitan areas in the South of
Brazil, reported considerable concerns with food safety (risks associated with bacterial
contamination in the meat). In addition, the same authors verified environmental concerns and
animal welfare as qualitative attributes of chicken meat only when more educated people were
interviewed in accordance with the previous hypothesis by Crabone et al. (2005) and Castellini et
al. (2008).
In recent years, there has been a growing market for antibiotic-free, pathogen-free, free-range or
organically-reared chickens due to changes in consumer behaviour. Motivations are generally
associated with problems related to intensive production systems (Herrmann and Roder, 1995).
Improvements in animal welfare and environmental protection, and health issues - especially the
absence of pesticide and medicine residues - were the most important attributes highlighted by
consumers in two mature organic markets in Europe, Great Britain and Denmark (Wier et al.,
2008) and the same was true for consumers in Canada (Martínez Michel et al., 2011).
Even with a well-known market stratification and growing consumer willingness to pay premium
prices for organic products, high prices for the chicken meat together with supply shortages in

4
supermarkets may inhibit the market growth for organic chicken meat. Torjusen (2004) reported
both factors as the most important obstacles for establishing organic markets in Europe.
Denmark has the highest consumption of organic food per capita in Europe, especially for dairy
products, eggs, vegetables and fruits (Wier et al., 2008). However, consumption of chicken meat
is very low and production of organic broilers in Denmark in 2009 accounted for only 0.15%
(~120.000 animals) of the total broiler production in Denmark (Anon, 2012b).
Figure 1 shows the trend in the organic production of eggs and chicken meat in Denmark since
1996. While the annual production of eggs apparently stabilised in the last decade with an
expected growth in the near future, the production of meat showed a different trend.
Figure 1. Evolution of organic poultry production in Denmark - Laying hens for eggs and broilers for meat with expected production in 2015. The left Y-axis is the production of broilers while the right Y-axis is the production of eggs (Modified after Hermansen, 2011).
According to Krystallis et al. (2012), the main hypothesis for such a market instability in the
Danish sector for organic chicken meat is the difference between high production costs (reflected
in high consumer prices) and poorly experienced (and thus less identifiable) quality attributes of
the organic meat available to consumers.
0
2
4
6
8
10
12
14
0
100
200
300
400
500
1996 1998 2000 2002 2004 2006 2015
# broilers produced (x1000) Million Kg eggs

5
Figure 2. The dilemma cost vs quality – Important factors in decision making.
Figure 2 highlights the differences in size and price between the two types of chicken. It shows a
lack of information for the consumer to make an informed choice. In addition, it points to other
issues that may help understand the current limitations for the development of the market for
organic chicken meat. In practice, while differences in price are easily identifiable, quality
differences in the chicken products on the supermarket shelves are not necessarily directly
apparent. This suggests that the methods used to produce the chickens are not well known and
insufficient information thereof has been given.
1.2 Production systems and its regulations
The modernization that has occurred in the broiler industry in recent years has not only brought
economic advantages to consumers. For example, Adams (2012) reported that the U.S. Federal
Agency for Drug Administration (FDA) has admitted, in June 2012, that chicken meat sold in the
U.S. contains arsenic, a carcinogen, due to the addition of chemicals to the feed of broilers to
avoid disease outbreaks. As a result, some consumers have begun to look for alternatives and
some small-scale farmers were able to benefit from the growing demand for alternative and
“cleaner” food products by adding value to their products (Neufeld, 2002).
In an effort to guarantee the quality attributes of the products, regulations on alternative
production systems were established. Table 2 summarises the most important systems reported in

6
literature for alternatives to the conventional method of producing broilers. Differences in
stocking density, types of feed allowed and the genetic background specified for the production
of the alternative chicken meat are highlighted.
Table 2. International1 regulations on the rearing and marketing of different types of broiler chickens.
Min. growth
period(d)
Min. outdoor
access(d)
Indoor area
(birds/m2)
Outdoor area
(m2/bird)
Organic feed (%)
Genotype Growing
type Natural Antibiotic free - Br Not specified Not specified Not specified Not specified Not specified Any Natural Antibiotic free - US Not specified Not specified Not specified Not specified Not specified Any Free Range - US Not specified ½ life Not specified Not specified Not specified Any Free Range - EU 56 28 13 1 Not specified Any “Caipira” free range – Br 70 45 13 3 Not specified Slow Gr. Traditional Free Range - EU 81 39 12 2 Not specified Slow Gr. Organic U.S. Not specified Not specified Not specified Not specified > 70* Any Organic Brazil Not specified Not specified 10 2.5** 100 Any Organic Denmark 81*** When fully
feathered 10 4 95 Slow Gr.
Organic EU 81 27 10 4 95 Slow Gr. Label Rouge 81 42 11 2 Not specified Slow Gr. Organic Label Rouge 81 42 11 4 95 Slow Gr.
1Taken from Nielsen et al. (2003) and adapted from European regulations Nos. 1538/91, 2092/91, 2891/93, 436/2001 and 834/2007; Anon (2012d) and France (1996) regarding the Label Rouge and organic regulation; Coelho et al. (2008) and Brasil (2011) for the Brazilian regulations; Fanatico et al. (2009) for U.S. regulations for alternative and organic broilers. *Organic label based on three different organic feed ingredients with the minimum requirement of at least 70% organic ingredients in the feed. Up to 95% and 100% are also possible. ** 2.5 m2 per broiler if permanent paddocks. If paddocks are in rotational systems, 0.5 m2 is stipulated for external area per broiler. *** If a slow-growing genotype is used (35g per broiler/day on average estimated in three consecutive flocks), 70 days of age if broilers are from non-organic origin. If the parent stock is raised organically there is no limit on the age for slaughtering.
Rules for alternative broiler productions were created in different locations based on different
cultural aspects and circumstances, with the aim of certifying that products followed specific
high-quality criteria. “Natural”, “Free Range”, “Label Rouge”, “Caipira” and “Organic”

7
production systems each have their regulations1. These regulations describe the particularities
demanded for each labelling system and differences can be observed.
In Europe, general regulations for the member states for broilers in free-range and organic
systems are described in more detail below, and particularities for national rules in France and
Denmark explored. For the U.S., regulations for natural, free-range and organic systems are
discussed and, finally, the regulations for natural, free-range and organic systems for Brazil are
considered.
Natural systems
Natural systems can be defined as the production of broilers without artificial ingredients or
added colorants in the feed. Natural broilers can be found in the U.S. and in Brazil2 and to some
extent production methods were adapted from the conventional system. Modern infrastructure,
fast-growing genotypes and controlled barn environments are used in the production of Natural
broilers. Some inputs are substituted or removed like antibiotics and growth promoters. Broilers
are produced from diets without antibiotics, coccidiostats or animal-derived ingredients.
Such inputs are typically replaced by probiotic products, vegetal extracts and organic acids.
Stocking density can also be limited and reduced to adhere to market or production requirements,
but no specific rule was found for this aspect. The chicken products labelled as “natural” should
be minimally processed in a way that fundamentally does not alter the raw quality attributes of
the meat. The label of “natural chickens” must explain the use of the term “natural”, i.e. “no
addition of artificial colourings”, “free of animal-derived ingredients”, “antibiotic-free”
“antibiotic, chemotherapeutic and coccidiostat-free” or “minimally processed”.
1 For U.S.: USDA, 2000. Final rule for the NOP - National Organic Program (NOP). Organic labeling and marketing information. & Food Service and inspection service. Fact sheets. Labeling natural and free range chickens. United States Department of Agriculture. - For Brazil: Brasil, 1999. Oficio Regular No 7 from 19 May 1999. Describing rules for production of the "Frango Caipira"; Brasil, 2003. Federal Law n. 10.831. Organic agriculture and sets other provisions and Brasil 2011: Brasil, 2011. Normative Instruction N 64 from 6 October 2011. Describing technical rules for the production of livestock and vegetable in organic systems (All documents for Brazil available in Portuguese). - For Europe: EU, 1991. CR N.1538/91 of 15 June 1991 introducing detailed rules for implementing regulation 1906/90 on certain marketing standards for poultry. Off. J. Eur. Communities (L143), 11-22 & EU, 2007. CR N.834/07 of 28 June 2007 on organic production and labeling of organic products and repealing Regulation (EEC) No 2092/91. Off. J. Eur. Communities (L189), 1-23. - For France: France, 1996. French Decree from 12.03.1996 for the Label Rouge regulation. - For Denmark, Anon (2012c). Vejledning om økologisk Jordbrugsproduktion (in Danish). 2 There is no official/national regulation in Brazil for natural production systems. However, since 2001 AVAL – Alternative Poultry Association - composed by farmers, rural technicians, companies and stakeholders establish rules for production and labeling of chickens in natural systems. For more information in Portuguese: www.aval.org.br

8
Farmers may adopt some or all of those techniques that will determine how they will be able to
label and market their products. Any genotype may be used and in practice fast growing broilers
are raised. In Europe, this type of system is not recognized and labelling and marketing is not
permitted.
Free range systems
Free-range defines production systems where the chickens can, at least in theory, access outdoor
runs for most of their lives. The U.S. regulation differentiates free-range into three main systems:
Pastured, day range and traditional free-range. In the pastured and day-range systems, broilers
get the most benefit from highly nutritional vegetation and will presumably be less challenged
with parasitic infections due to constant movement of the broiler houses. These systems are well
known and extensively used by small-scale farmers in the U.S. and normally slow-growing
broilers are used (Fanatico et al., 2009). In traditional free-range systems in the U.S., houses are
normally fixed, paddocks are permanent and vegetation is less abundant, especially close to the
houses. Parasitic infection is hypothetically one of the major drawbacks in this production.
Fanatico et al. (2007) observed that in the U.S., free-range systems vary widely from large
stationary houses with permanent paddocks to small portable houses that are moved frequently to
new pasture areas. The U.S. definition of free-range system does not include any specification
for how long broilers should remain outdoors and under what conditions they should be raised
and fed. In addition, there is no specification for appropriate genotypes and no minimum age for
slaughter. In order to label broilers as free-range, farmers in the U.S. must demonstrate that birds
have had free access to the outdoors for more than half of their lives. It means that the free-range
label in the U.S. is slightly controversial, since farmers insist that the label applies even if the
birds do not actually go outdoors. Fast-growing genotypes used in these systems may prefer to be
closer to the feeders than foraging in the outdoor areas. For instance, broilers can be outside on a
concrete floor for a short period of time and still be labeled as free-range (Fanatico et al., 2009).
This can also be observed in other systems where fast-growing broilers spend considerably less
time outside than slow-growing broilers (Nielsen et al., 2003).

9
In contrast to the U.S. regulation, the European legislation for free-range systems specifies
maximum stocking rate for indoors and outdoors, minimum age at slaughter and genotype as
well as the feed composition. The European legislation also differentiates free-range into three
main systems: Free-range, traditional free-range and total freedom. In free-range systems poultry
houses are normally fixed and an adequate number of popholes is required based on the size of
the houses and number of animals.
In traditional free-range, as the label rouge systems, stipulates a minimum slaughter age of 81
days and no more than 4,800 broilers can be produced per flock. Mobile houses can be used, but
they are limited to 150 m2 of ground space. If mobile houses are kept open during the night,
indoor stocking rate can be increased to 20 broilers/m2. With movable houses, the total usable
area per house cannot exceed 1,600 m2. Broilers must have continuous access to open-air runs
covered with vegetation from six weeks of age. At least 2 m2 of external area is required per
broiler and only slow-growing genotypes are permitted. In total freedom, also defined in
countries outside France as label rouge with total freedom, the same regulations as for traditional
free-range must be followed, except that the broilers must have continuous access to open-air
runs without fences or a minimum area requirement.
In Brazil, the “caipira” production system specifies regulations for free-range broilers. The
organic regulations in Brazil do not require slow-growing genotypes and in fact the same
genotypes and age of slaughter can be used as in conventional systems. However, the regulations
for “caipira” systems do require slow-growing broilers. “Caipira” systems can also be certified as
organic if the production follows the feed restrictions based on organic regulations.
In practice, broilers in “caipira” systems are raised in fixed houses accessing permanent
paddocks. There is no specification for the vegetation type and, in fact, the presence of
vegetation is not mandatory. There is no limitation in terms of size of the flocks. Caipira systems
are normally practised by small-scale farmers and flocks can vary in size but will not exceed
2,000 broilers (Faria et al., 2012).

10
Feed must be constituted by ingredients exclusively from vegetal origin and the use of growth
promoters of all types is forbidden. The regulation for “caipira” production demands a growing
period of 85 days. However, a recent change has been made and the minimum slaughter age was
reduced to 70 days without limitations for daily growth rates. The change was made to meet a
general demand of farmers, companies and stakeholders due to the high prices of feed
ingredients, especially when the “caipira” broilers are supplemented with organic ingredients to
receive the organic label.
Demattê Filho, 2012 (Personnel communication) disclosed that, additionally, companies offering
slow-growing genotypes for the “caipira” system are in fact supplying hybrids with medium
growth rates. When these animals are fed for 85 days as demanded by the original regulation for
“caipira” systems, broilers grow excessively. As “caipira” broilers are marketed as entire
chickens, consumers may refuse to buy the chickens because they associate the carcass size with
a possible supplementation of hormones and growth enhancers that in fact are not permitted.
The rules for the free-range systems in the U.S., Europe and Brazil supposedly originate from the
French “Label Rouge” system. The term was originally attributed to a quality label (the “red
label”) for poultry produced under a specific guideline thus including small-scale operations, low
stocking rates in housing and slow-growing chickens accessing free-range areas. (More
information on Label Rouge chickens and regulations can be found in the French Rural Code
article L.641-1.)
In short, the Label Rouge designation applies to slow-growing chickens fed low-fat and high-
cereal diets and reared at low stocking densities with outdoor access to a minimum age of 81 d
before being processed (Smith et al., 2012; Lewis et al., 1997). Housing systems in the French
Label Rouge are mostly fixed and permanent as in the traditional U.S. and European free-range
systems and the Brazilian “Caipira” system. The use of a slow-growing genotype is mandatory
and the indoor stocking density allowed is a maximum 10 birds/m2.

11
Organic systems
The French standard for “organic” production is based on the Label Rouge system and thus is
more stringent than the European Regulation (EC) 1804/1999. The total usable surface area for
broilers in any single production unit must not exceed 1,600 m2, and use must not exceed 400 m2
of building per production site. This means that the “organic Label Rouge” is not heavily
concentrated and that houses are well separated (MAFF, 2001).
The French requirements for organic production of broilers demand that only slow-growing birds
of the same age can be grouped in the same building. Furthermore, buildings of more than 200
m2 must be separated into pens by a sealed partition, extended outside by a separation of the
grass runs which broilers cannot cross. This will lead to a maximum flock size per pen of 2,000
broilers (MAFF, 2001). However, no specification on the type and species of vegetation is
available and only “grass” is used to describe the vegetation available to broilers in the organic
Label Rouge regulation. In addition, 95% of the diet must be composed of organic ingredients,
mainly cereals. Allopathic treatments for parasites and infectious diseases are forbidden and the
use of plants and extracts are suggested instead.
In general, regulations for organic production of broilers specify that land should be free of
synthetic fertilizers, pesticides and herbicides for a certain length of time. The dietary
supplementation is subject to some specific requirements. Synthetic ingredients and genetically
modified organisms in feed are customarily forbidden. In some European countries and in the
U.S., some limiting synthetic amino acids are still permitted at low levels if organic sources are
not available to farmers. However, in the EU, 95% of the feed ingredients must be organically
produced, otherwise the meat will not be labelled as organic. From 2015 the requirement will be
for feed to be 100% organic.
In the U.S., organic labelling is available at three different levels based on the amount of organic
feed supplemented to broilers: “100% organic broiler”; “organic broiler”, meaning that at least
95% of the ingredients were organically produced, and “made with organic ingredients”,
meaning that broilers consumed at least 70% of organic ingredients in the feed (USDA, 2012).

12
The U.S. regulation does not specify indoor or outdoor stocking densities, but certification
agencies look for a maximum of eight broilers/m2 indoors (Fanatico et al., 2009). Most organic
production of broilers in the U.S. is performed under intensive, large-scale conditions, similar to
those in the conventional industry (Fanatico et al., 2009). Although organic broilers must have
access to the outdoors, in reality little access is provided in commercial systems using fast-
growing genotypes.
In Denmark, there is a national regulation for the production of organic broilers based on the
European regulation. The maximum size of flocks in Denmark cannot exceed 4,800 broilers and
a maximum stocking rate of 10 birds/m2 is mandatory. If mobile housing is used, the stocking
rate can be increased to 16 broilers/m2, but the maximum size of the flocks cannot be changed
(Anon, 2012c).
All broilers must be able to explore the outdoor chicken runs at stocking rates of 4 m2 per broiler.
The chicken runs used in one production year cannot be used in the following year, to avoid a
high parasitic/microbial burden in permanent paddocks. However, no specification on the type of
vegetation is made and, in fact, the presence of vegetation is in reality not mandatory. As a
peculiarity of the Danish system, broilers should have free access to roughage. Broilers should
have access to the outdoors at the latest when they are fully feathered.
With respect to the minimum age at slaughter, the starting point is the 81 days specified in the
European regulation. However, if a slow-growing genotype is used, the minimum slaughter age
is 70 days if broilers are of non-organic parentage. Additionally, the definition of slow-growing
genotype in the Danish regulation is based on the average daily gain of the broilers. The
definition of slow-growing is broilers that do not grow more than 35 grams per day. The growth
rate is calculated on three successive flocks. However, if the parent stock of the broilers was
raised organically, there is no minimum age for slaughter (Anon, 2012c). In practice, it means
that “less” slow-growing genotypes are used (complying with the 35 g of daily growth).

13
Challenges in the production of broilers in organic and free-range systems
Recently, the European parliament delayed the introduction of the requirement for broiler feed to
be 100% organic to the end of 2014. In addition, the new regulation is expected to support
farmers in that most of the feed ingredients must be produced on-farm or locally. The U.S.
regulatory system adopted the labelling of organic broilers based on the proportion of organic
ingredients in the feed. The Brazilian regulation stipulates a minimum of 80% organic
ingredients in the feed and will change to 100% organic by January 2013. It is important to
mention that in Brazil any genotype can be used and there is no minimum age for slaughtering
organic broilers. Changes and adaptations to the studied regulations highlight the dependence on
oil seed crops and cereals in the feeding strategies for organic broilers, in addition to the use of
(“less”) slow growing genotypes in commercial organic production. It is a challenge for organic
regulation and certification bodies as apparently such systems are not appropriate if the
principles and ideals of IFOAM (IFOAM, 2005) are to be followed during certification
procedures.
Nevertheless, antibiotics, growth promoters, coccidiostats and anthelmintic drugs in prophylactic
treatments are prohibited for all studied regulations. Phytotherapeutics (the use of plants and
plant extracts) and homeopathy are suggested instead. A third party certification company is
responsible for inspecting the production at any time in order to grant certification and thus the
organic label.
With the exception of the organic Label Rouge produced in France, in all other countries “less”
slow-growing genotypes could be used in the production of organic chicken meat. In reality, a
large proportion of the broilers currently marketed as organic are produced in intensive systems,
by the use of fast (or “less” slow) growing genotypes and heavily dependent on organic oil seed
crops and cereals.
European legislation prescribes a minimum age at slaughter for organic production. However,
only systems using slow-growing genotypes are able to keep animals for the minimum period of
81 days before slaughter. Keeping a flock of less slow-growing broilers for the minimum period

14
could create problems for farmers (production costs) and for the welfare of the animals (i.e. leg
problems, low activity, foot-pad dermatitis, etc.).
There is no minimum period specified for outdoor access in the U.S regulations, and in Brazilian
regulations only a small external area of 0.5 m2/broiler is specified for organic systems when
paddocks are rotated. Farmers in Denmark may keep broilers indoors until they are fully
feathered, and this, together with the heavy dependence on oil seed crops and cereals in feeding
strategies, suggests that there are important constraints for the development of the organic sector
when using fast-growing or less slow-growing genotypes. The various regulations commonly
stipulated access to an external range, but the nature and type of vegetation was not specified and,
in practice, most free-range land was just described as grass or vegetation (Walker and Gordon,
2003) and its availability was not even mandatory. The production systems practised revealed
important obstacles in the production of organic chicken meat.
A heavy dependence on oil seed crops and cereals in feeding strategies of fast (or “less” slow)
growing broilers means there is less opportunity for product differentiation of organic meat
products. Even with mandatory access to outdoor areas, herbage has not been mentioned as a
possible source of nutrients, minerals or vitamins in the nutrition of broilers, and with the impact
of grains and cereals on production costs there is good justification for looking for alternatives to
feed broilers and foraging can only be an option if slow-growing genotypes are used (Weeks et
al., 2004).
1.3 Nutrient supply by foraging
In older traditional systems (Prothero, 1936), chickens were normally left to roam more naturally,
seeking essential parts of their nutritional needs on or below the soil surface and consuming not
only oil seed crops and cereals supplied by humans but also vegetation, seeds, fruits, soil
particles, microorganisms, different stages of insects and other arthropods and earthworms
(Lomu et al., 2004; Gordon et al., 2006) (Figure 3). Soil-living organisms have also been

15
reported as an important source of essential amino acids (i.e. Pokarzhevskii et al., 1997; Basser
et al., 1999).
Even though different legislation requires that paddock areas must be mainly covered with
vegetation, the nature and type of the vegetation is not specified (Walker and Gordon, 2003).
Little attention has been given to make outdoor runs attractive and nutritious to broilers as part of
agroecosystem management (van de Weerd et al., 2009).
Figure 3. Chickens foraging for different ingredients in a Danish free range system.
Recently, Horsted and Hermansen (2007) studied laying hens managed on different types of
vegetation and reported that hens were able to meet essential parts of their nutritional
requirements through foraging. Other recent studies have suggested that plots cultivated with
chicory (Cichorium intybus cv. Grassland Puna) were highly attractive to layers based on their
high intake of this plant (Horsted et al., 2006; 2007). In this thesis, it was hypothesised that
chicory could also be attractive to slow-growing broilers.
In strategies for substituting oil seed crops and cereals it is important to identify the amount of
herbage and other living organisms chickens can consume from outdoor areas and the extent it
may contribute to their nutrient supply. Even a modest contribution may be important given that
feed accounts for about 70% of the variable costs of poultry production (Walker and Gordon,
2003) and may be an even higher share in organic broiler systems due to longer periods of

16
growth. It is also important to know the impact of broiler type. Thus, slow-growing broilers
spend considerably more time outside than fast-growing broilers (Kjaer and Mench, 2003) and
this influences foraging behaviour (Nielsen et al., 2003). In practice, it is important for the
farmers to know the value of the different types of ingredients available in the range as a means
of nutrient supplementation for the flocks.
The economic benefit of foraging, however, also depends on how it impacts on the feed
conversion rate (feed used per kg body weight gain). Feed is used partly for maintenance and
partly for tissue growth (Leeson and Summer 1997), where the former is a function of body
weight. Thus, the feed conversion rate is generally impaired with increasing body weight due to
the proportionally higher feed requirement for maintenance compared with growth. This is
particularly important when organic farmers produce in intensive systems.
Similarly, the feed conversion rate is generally impaired in slow-growing broilers due to the
proportionally higher maintenance requirement when foraging compared with growth. Organic
production is typically based on a substantially higher slaughter age and weight – aspects that are
expected to impair feed conversion. If part of this additional feed requirement can be supplied
with herbage and other living organisms, this may improve the profitability of the system
(Walker and Gordon, 2003). The practice of exercise, moving around and searching for food
ingredients in the range combined with age and genetic background of broilers (Castellini et al.,
2008) may give consumers the differentiation in quality (i.e. tenderness) and visual attributes (i.e.
darker colour) that they seek of organic chicken meat (Owens et al., 2006; Delpech et al., 1983).
Supplementing low-protein, low-energy diets, sometimes in a commercial strategy to reduce
costs for industrial broiler systems, may lead birds to consume more of the available feed and
thus increase feed intake (Leeson and Summers, 1997). This suggests that for slow-growing
genotypes in free-range systems, restriction in the commercial diet may stimulate them to
increase feed intake from feed ingredients available in the range.
However, stimulating broilers to explore the free-range land can be a trade-off for the farmers. It
can benefit the nutrition of broilers, but broilers will be more exposed to parasites when

17
accessing permanent paddocks (Permin and Hansen, 1998; Permin et al., 1999; Thamsborg,
2002). And the use of drugs in allopathic treatment against parasites and infection diseases is
forbidden in alternative systems. In all of these alternative systems, the use of substitutes for
medicines to treat microbial and parasitic infections is required, especially in intensive organic
systems.
In the search for new ingredients to substitute chemicals products, the use of plants, roots, crops
and/or its combinations is believed to play an important role because they are supposedly
residue-free and thus well accepted by consumers (Orengo et al., 2012). The original idea behind
this PhD project was to introduce plants with high nutritional content together with plants with
anti-parasitic properties to chicken runs and investigate if broilers were able to forage for
nutrients and at the same time self-medicate. However, due to the time taken for plants to
establish and the high labour input needed for this at farm level, the decision was to supplement
the plant ingredients with what could be practical for the farmers.
1.4 Four herbal food supplements and their anti-parasitic properties
The production of poultry in organic systems promotes animal welfare, health and the quality of
its products. Regulations prohibit the use of drugs for preventive treatment of infectious diseases,
including parasites. As a result, treatment depends on innovative substitutes to the currently-used
drugs. Sometimes vaccination is available, although not always affordable, especially to small-
scale farmers. Vaccination also increases production costs in more intensive systems. Thus, the
use of herbs and their derivatives may represent a viable alternative (Abbas et al., 2012) for
production systems in general.
In organic production systems, broilers are more exposed to parasites. Organic free-range
systems are normally managed in fixed houses with access to permanent paddocks. In intensive
systems using fast-growing genotypes, substitutes for drugs and antimicrobials are even more
necessary. In the alternative systems described previously (section 1.2), control of parasites
through plants, roots or crops offers attractive alternatives.

18
Recently, the herb Artemisia annua L. (Figure 4) has received much attention as an alternative
treatment for drug-resistant Plasmodium falciparum malaria (Willcox et al., 2007) and the
commercial use of artemisinin, the main active ingredient extracted from the plant, is now used
worldwide as a combination therapy (Olliario and Taylor, 2004; Weinberg and Moon, 2009).
Artemisia annua has also been considered a potential candidate for the control of other
apicomplexan parasites, e.g. coccidia infections in chickens (Allen and Fetterer 2002; Arab et al.,
2006).
Figure 4. Starting from the left side: Schematic presentation of Artemisia annua leaves; A. vulgaris leaves and inflorescence; leaves, roots and powder of Curcuma longa and Manihot esculenta foliage.
Allen et al. (1997) reported preventive effects of dried A. annua dried leaves for fast-growing
broilers against individual and dual infections with Eimeria tenella and E. acervulina. They
reported that a prophylactic strategy with a three-week supplementation period prior to the
challenge was required to give a reduction in oocyst output of E. acervulina and increased
performance. Interestingly, the authors reported no significant effects when A. annua dried
leaves, or pure artemisinin, were provided preventively against E. maxima, suggesting that plant
components including artemisinin may have a species-specific mode of action as also suggested
by Arab et al. (2006).

19
A second herb from the genus Artemisia, A. vulgaris has been reported to have anthelmintic and
anti-inflammatory properties in humans (Tigno et al., 2000; Tigno and Gumila, 2000)
supposedly due to high levels of antioxidants (Temraz and El-Tantawy, 2008), since this species
does not contain artemisinin (Mannan, 2010).
Another plant with known anti-protozoan properties is the spice turmeric Curcuma longa,
previously suggested as a potential candidate to treat malaria (Reddy et al., 2005). In Pakistan,
farmers supplement turmeric powder as a feed additive for the control of coccidiosis in intensive
production systems (Abbas et al., 2012). Allen et al. (1998) reported a reduction in intestinal
lesions and suppression in oocyst excretion when 1% dietary inclusion was available to fast-
growing broilers challenged with E. maxima occysts. Apparently, the medicinally active
ingredient of turmeric is curcumin (diferuloylmethane), a phenolic compound with high
antioxidant, anti-inflammatory, and anti-cancerous properties (Conney et al., 1991).
Finally, a relevant feed source for small-scale farmers in Brazil is cassava (Manihot esculenta), a
tropical crop that is traditionally cultivated in developing countries as a source of starch. Apart
from the main product, the root, cassava also produces large amounts of leaf that are left in the
fields as an organic by-product (Sokerya et al., 2007). Fresh cassava forage, including the tender
stems, can be used directly for feeding growing chickens (Ravindran, 1991).
In diets based on maize-soybean grains, the leaf meal was first suggested to be limited to 5 g kg-1
in the diet (Ross and Enriquez, 1969) due to possible anti-nutritional effects of condensed
tannins. Higher levels of inclusion deserve to be investigated as condensed tannins are known to
kill pathogenic bacteria (Barry et al., 1986), to have anthelmintic effects in parasitized dairy
cattle and buffaloes (Netpana et al., 2001), and to have coccidiostatic effects in parasitized goats
(Seng and Rodriguez, 2001) and sheep (Marie-Magdeleine et al., 2010).
Different plant species may have different modes of action when individually supplemented or in
combination. As the commercial production of organic and free-range broilers in general is
performed in short growing periods, some parasites are more relevant during production and thus
deserve more attention than others.

20
Due to its short life cycle and fast dissemination, coccidiosis caused by Eimeria spp. is one of the
major constraints for the production of broilers (Williams, 1999). In addition, in low-density
production or with the use of alternative products, coccidiosis generally remains a subclinical
disease that only affects performance (Fanatico, 2006). The use of plants to alleviate the effects
and severity of coccidiosis may help reduce mortality in low-input systems and also improve
performance for intensive systems by reducing production costs for farmers.
However, there is no clear information in the scientific literature on the mode of action of the
different botanical products. Even when scientists have presented some important evidence, the
modes of action are largely unknown. What has been discussed is the health benefits to animals
of botanical products and the improvement to the immune systems or affect on coccidia parasites
at different stages of its life cycle (Naidoo et al., 2008; Allen et al., 1998).
It is therefore important to gain knowledge on the life cycle of the parasite in order to benefit
from management strategies that may reduce infection severity at farm level. It may also support
the investigation of the mode of action of botanical components and the development of new
strategies to alleviate the effects of infectious diseases like coccidiosis.
1.5 Coccidiosis: relevance in the broiler industry worldwide and the Eimeria
spp. life cycle
Coccidiosis is a parasitic disease caused by the protozoa Eimeria spp. that can lead to severe
losses in poultry production (Williams, 1999). Almost 70% of all economic losses are related to
subclinical diseases thus impacting weight gain and feed conversion rate (De Gussem, 2007).
Velkers (2011) reported that nine species of Eimeria have been described in chickens, of which
at least seven species are relevant for the poultry industry, i.e. (E. acervulina, E. maxima, E.
brunetti, E. necatrix, E. mitis, E. praecox and E. tenella) and some species are more pathogenic
than others. The most frequently diagnosed species in intensively reared poultry include E.
maxima, E. tenella and the most prevalent one, E. acervulina (Williams, 1995).

21
Infections with E. acervulina, E. maxima and E. tenella are frequently diagnosed in intensively
reared broilers (McDougald et al., 1997) and their control is usually accorded a higher priority
over the other four species due to their pathogenicity (Shirley et al., 2005). In this Ph.D. project,
the focus was on two pathogenic species affecting broilers in Denmark and in Brazil, Eimeria
maxima and E. acervulina, because they cause damage to important parts of the broiler intestines
responsible for nutrient absorption and digestion (Mathis, 2005).
In general, coccidia multiply in epithelial cells in different regions of the intestines, damaging
gut tissues, which, in turn, reduces feed intake and the absorption of nutrients (Morris et al.,
2007). Eimeria acervulina can be found in the upper part of the small intestine – the duodenum -
of chickens, while E. maxima can be found in the middle part of the small intestine (Anon, 1990).
Each species of Eimeria can act alone or in combination. Infections caused by E. acervulina lead
to lesions in the duodenum mucosal surface and in heavier attacks the species may inhibit the
ability of E. maxima to develop in the central part of the intestine (Mathis, 2005).
Depending on infection severity, especially for high stocking density systems, chickens may be
more susceptible to secondary bacterial infections (Cooper and Songer, 2009) and these can
potentially be zoonotic (Al-Sheikhly and Al-Saieg, 1980). In Mexico and Brazil, coccidiosis was
reported as the most important cause of mortality in chickens raised in small-scale systems
(Rodrigues et al., 1997; Luchese et al., 2007).
In intensive conventional systems, the use of ionophorous antibiotics is routine. These drugs
belong to a class of anticoccidial drugs that disrupts the flow of ions across the membrane of
Eimeria parasites, thus causing ion imbalances causing the death of the parasite (Wallach, 2010).
The Eimeria life cycle (Figure 5) is perpetuated when oocysts previously released in faeces from
an infected broiler are ingested by another broiler. Only oocysts undergoing sporulation in the
environment are infective. Due to scratch behaviour, oocysts are involuntarily collected and
ingested. Following ingestion, the grinding action in the chicken gizzard associated with
pancreatic secretions in the upper part of the small intestine induces the rupture of the oocyst

22
wall. It releases four cysts (sporocysts), each containing two infective parasites (sporozoites) that
will invade gut epithelial cells if they do not come into contact with an anticoccidial drug first
(Cervantes, 2008).
Figure 5. Eimeria spp. life cycle (modified from USDA, 2005).
The timing of the cellular invasion will depend on the Eimeria species and may vary by a few
hours from when the oocyst is ingested. Following invasion of a host cell, the sporozoite
becomes a trophozoite which undergoes replication, increases in size and is then termed a
schizont. The schizont ruptures thus destroying the host cell and releasing merozoites. In this
asexual propagation (schizogony), a fast dissemination occurs. Neighbour cells will be invaded
by merozoites and schizogony will repeat. Later male and female gametocytes and gametes are
formed (gametogony) and these will generate new oocysts for completing the cycle in the sexual
stage. Millions of new oocysts produced are released in the environment with faeces (Anon,
1990). The current literature suggests that the release of merozoites from the schizont phase is
repeated two to four times (i.e. Cervantes, 2006) until a reaction from the immune system occurs
23
or it is inhibited by the action of anti-coccidial drugs (Lillehoj and Lillehoj, 2000). Sporozoites,
early developmental stages (trophozoites and first generation schizonts), merozoites and
gametocytes all play a role in induction of protective cellular immunity (Wallach, 2010).
The effects of coccidiosis are associated with the invasion and destruction of epithelial cells, and
associated inflammation of the gut. Rose and Hesketh (1976) suggested that at up to four
generations of merozoites can develop after infection by a single oocyst. It leads to the
hypothesis that if a large number of oocysts is ingested by a single animal in a short period of
time, the health status of the whole flock might be severely compromised in a short period of
time, especially at high stocking densities (Velkers et al., 2010). The number of generations of
Eimeria spp. affecting flocks may depend on the particular species or combination of species
present in the housing (Mathis, 2005). However, after broilers have been exposed to a specific
strain for the first time, when reinfected they apparently develop immunity/resistance (Rose et al.,
1997).
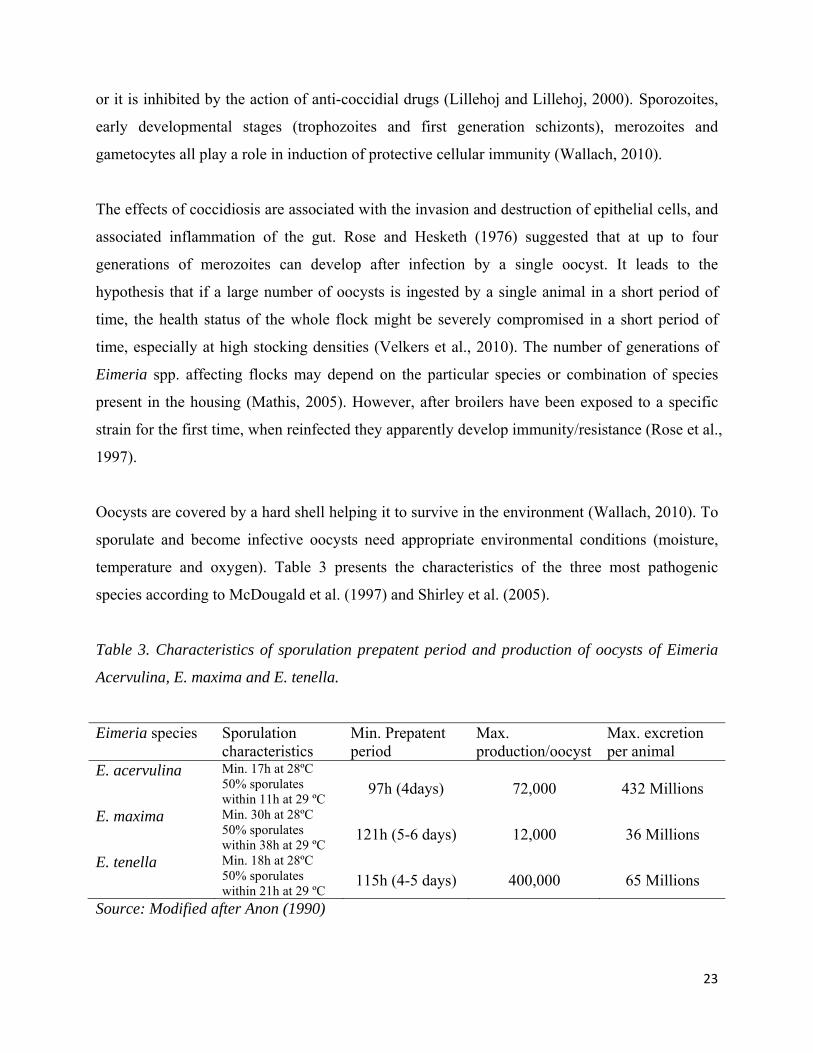
Oocysts are covered by a hard shell helping it to survive in the environment (Wallach, 2010). To
sporulate and become infective oocysts need appropriate environmental conditions (moisture,
temperature and oxygen). Table 3 presents the characteristics of the three most pathogenic
species according to McDougald et al. (1997) and Shirley et al. (2005).
Table 3. Characteristics of sporulation prepatent period and production of oocysts of Eimeria
Acervulina, E. maxima and E. tenella.
Eimeria species Sporulation characteristics
Min. Prepatent period
Max. production/oocyst
Max. excretion per animal
E. acervulina Min. 17h at 28ºC 50% sporulates within 11h at 29 ºC
97h (4days)
72,000
432 Millions
E. maxima Min. 30h at 28ºC 50% sporulates within 38h at 29 ºC
121h (5-6 days)
12,000
36 Millions
E. tenella Min. 18h at 28ºC 50% sporulates within 21h at 29 ºC
115h (4-5 days)
400,000
65 Millions
Source: Modified after Anon (1990)

24
The prepatent period of Eimeria spp. parasites – the period between the intake and release of
new generation of oocysts – varies with species but will not be longer than six days (Anon, 1990).
Depending on the production systems and the genetic background of broilers, severe losses in
production can be observed, especially during the subclinical stage when infections easily remain
unnoticed (De Gussem 2007; Fanatico, 2006).
When coccidiosis reaches a clinical phase, broilers will display a typical “sick bird” attitude with
depression, prostration with huddling under the source of heat as if they feel cold, and producing
watery or bloody droppings (Cervantes, 2006). However, there are only few options for the
diagnostic of subclinical coccidiosis. Oocysts per gram of faeces (OPG) can be counted by using
a practical technique (McMaster) and is a method that can be useful and applicable at farm level
and only demands simple equipment and a trained person. The method can be used for detecting
and measuring infection dynamics of individual broilers (Roepstorff and Nansen, 1997) or a
group of individuals by pooled samples or by litter analysis (Long and Rowell, 1975). However,
OPG data alone gives little indication of severity of infections and, in addition, morphological
examination for speciation is very difficult (Thebo et al., 1998). Analysis of OPG in farmed
chickens may identify the epidemiology of the disease and thus the population dynamics.
However, methods should be used in combination if determination of the role of the subclinical
disease on productivity is the target.
As a complementary method, lesion scoring of intestinal sections can also be applied at farm
level. It is a visual interpretation of the lesions caused by Eimeria spp. at specific locations of the
serosa and mucosal surface in the gut of the broilers. The method uses a scoring system ranging
from 0 to 4 depending on the severity of the infections in the different locations of the intestine
(Johnson and Reid, 1970). Although labour-intensive, this method is currently used in Denmark
to determine the severity of Eimeria spp. infections and also for speciation in commercial flocks
if molecular analysis (PCR) is unavailable or unaffordable (S. Kabell, personal communication,
2011). When affordable to farmers and during scientific experimentation, the use of molecular
analysis is a very important tool for speciation (Haugh et al., 2007; Fernandez et al., 2003).

25
In large-scale conventional systems, supplementation of chemical products to control diseases is
routinely given. In organic intensive systems, the use of vaccination normally integrates a
prophylactic management. However, efficient protection is not always provided and usually
vaccination results in subclinical coccidiosis that affects feed efficiency (Chapman, 2007). It
limits superior utilization of feed and thus may frustrate farmers wishing to reduce production
costs. Also, vaccination is not always affordable, especially in small-scale systems where slow-
growing genotypes are used.
Therefore, in free-range systems it is very important that broilers have sufficient capacity to
withstand parasitic challenges to which they are exposed, especially if permanent paddocks are
used. This may be genetic components, conferring the ability to cope with parasitic infections.
1.6 Genotypes for different productions systems
In the last decades breeding programmes for the production of broilers have focused on genetic
gains to improve growth rate and feed conversion, but this has led to undesirable side-effects
conflicting with the ideals for the production in systems orientated towards sustainability (Olesen
et al., 2000). To change the negative effects imposed by the high selection pressure for
production characteristics, scientists suggest that novel breeding programmes should include
ethical standards by changing from focusing only on increased yields to include traits linked to
the health and welfare of broilers (Decuypere et al., 2010).
As part of the process of searching for a more sustainable production, the concept of
‘‘robustness’’ was proposed (Star et al., 2008). It incorporates ecological resilience into the
production of animals (Holling, 1973). For example, a suitable definition for robust broilers
would be animals with the ability to withstand external pressures without losing production
potential. It suggests that robust broilers might be less susceptible to parasitic infections and will
recover faster and healthier from challenges caused by coccidiosis. From a welfare point of view,
robustness will give broilers better ‘‘coping abilities’’ and flexibility (Decuypere et al., 2010). If
the broilers display this flexibility, welfare is improved (Star et al., 2008).

26
The emergence of drug resistance and increasing pressure from society against the use of
antibiotics is driving the research on coccidiosis towards alternative products to control the
disease like vaccines and herbal extracts and breeding of more resistant types of animals
(Swinkels et al., 2007). Because broilers are bred mainly for economically important traits,
almost no specific selection on disease resistance has taken place (Leshchinsky and Klasing,
2001).
This has led to indirect selection pressure for several unfavourable traits resulting in a decrease
in the overall resistance and poor adaptation capacity. In organic systems, it is hypothetically
beneficial, both for welfare and economic reasons, to have better resistance against infectious
diseases and a better adaptation capacity to less controlled environmental conditions (Swinkels et
al., 2007).
Selection for high performance attributes in fast-growing broilers resulted in alterations to the
immune response when compared to slower-growing genotypes (Cheema et al., 2003). Host
immune reactions to parasites are complex and cell-mediated immunity plays a major role in
disease resistance to coccidial infection (Dalloul and Lillehoj, 2005; Lillehoj, 1998).
Choi et al. (1999) found an increase of a cytokine that is critical for innate and adaptive
immunity against infections caused by E. tenella, but almost none caused by E. acervulina. In
addition, Swinkels et al. (2007) reported an increase of the same cytokine in the duodenum of 7-
day-old fast-growing broilers but not in slow-growing broilers of the same age challenged with E.
acervulina parasites. Swinkels et al. (2006) found that the cytokine response was absent in fast-
growing broilers infected at one day of age, but the same genotype infected at seven days of age
increased its expression. It suggests that vulnerability to Eimeria spp. infections is not only
related to genetics but is also dependent on age and developmental stage of the immune system
of the broilers (Swinkels et al., 2007).
In addition to age affecting the response to parasitic challenges, basic knowledge of genetics
supports the concept of adaptability. The process of genetic resistance and how the environment

27
influences broilers in intensive and free-range systems is far from fully understood (Sørensen,
2010). Genetic resistance can be resistance to coccidiosis through preventing the parasite from
establishing in the gut, which to some extent was observed in the Egyptian Fayoumi genetic line
(Pinard-van der Laan et al., 1998). Resistance to coccidiosis may also be mediated through
immunological characteristics that help to reduce the parasitic effects in the chickens infected
like improvements in “coping abilities” (Sørensen, 2010).
Apparently, a more profitable approach for breeding companies and also for farmers would be a
combination of both factors. Resistance to coccidial infections in organic systems might be
achieved by the genetic capacity of slow-growing broilers to cope with infections associated with
appropriate sanitary measures implemented by the farmer, postponing the age broilers are
challenged. To some extent, this is the basic strategy behind the use of coccidia vaccines. To
stimulate immune response, a low level of infection associated with less pathogenic strains is the
key for a successful vaccination programme against coccidisosis (Williams, 2002).
However, vaccination at a very early stage may be compromised if chicks are then repeatedly
challenged by infective oocysts available in the ground. To some extent, farmers may be able
through management practices to influence environmental factors and exposure of broilers to
challenges, allowing the development of the immune systems and thus promoting resistance to
the disease. In a complementary approach, if the use of herbal extracts reduces the parasite
infectivity (Del Cacho et al., 2010), it may likewise reduce infections, thus allowing time for the
immune system to respond to the first challenge and for the broilers to acquire resistance to the
Eimeria spp. parasite.
In intensive systems with repeated turnover of flocks where vaccination is not always efficient
(Min et al., 2005) or reduction in the production costs is a target (Abbas et al., 2012), fast-
growing genotypes (Figure 6) may benefit from protection provided by botanical food
supplements through improvements of the intestinal health and in this way reduce the effects of
coccidial infections.

28
In free-range systems, the robustness of slow-growing genotypes aided by herbal
supplementation complies with organic principles and ideals (IFOAM, 2005). For example,
Thorp and Luiting (2000) selected fast growing broilers for high or low antibody response to an
externally induced disease challenge. Broilers with a high antibody response had a lower body
weight gain. In other words, when fast-growing broilers selected for high production parameters
are exposed to such challenges, they may be more severely affected than slow-growing
genotypes. Stearns (1976) defines this cause-effect relation by a theory of resource allocation.
While positive correlations can be observed when selecting for higher growing rates, negative
genetic correlations among fast-growing parameters and resistance to diseases may also exist as
it does for reproduction traits (Rauw et al. 1998). Broilers selected for high yields may allocate
more energy to tissue growth and as a consequence they will have less energy available to fight
coccidial challenges.
Figure 6. From the left: A pure slow-growing broiler, a hybrid double-purpose genotype and a fast-growing genotype.
Broilers managed under intensive organic systems are normally challenged with coccidiosis at an
early stage of life due to the opportunism and fast dissemination of the parasites. Depending on
infection severity, opportunist bacteria may benefit from the lesions in the gut mucosa and from
the weakness of the immune system of chickens (Al-Sheikhly and Al-Saieg, 1980). When
infection caused by parasites and bacteria achieves a level of economic damage, mortality rates
may increase and efficiency in feed utilization will decrease as a consequence. This may cause

29
pain and compromise animal welfare. In addition, it will lead to higher production costs, which
will be reflected in higher prices for consumers.
More than for any other domesticated species, breeding programmes for broilers have narrowed
their selection objectives driven by economic priorities (Sørensen, 2010). In contrast, “caipira”,
free-range, Label Rouge and organic free-range systems normally practised on small-scale farms
are likely to support the use of slow-growing genotypes. Systems without the use of prophylactic
treatment of parasites may provide safer products for consumers due to the absence of drug
residues but broilers may suffer more depending on the genetic background, stocking density and
the severity of the coccidial challenges. This gives a dilemma with considerations for animal
welfare on the one side and production on the other, which will be explored in the following
section.
1.7 Animal ethics: behaviour as a means of animal welfare assessment in different production systems
International regulations for chickens managed in organic and free-range systems emphasize
animal welfare (EU, 1999a; USDA, 2000; Brazil, 2003). However, the meaning and
interpretation of the concept “animal welfare” varies based on the values of who is using the
concept (Sandøe and Christiansen, 2008). Vanhonacker et al. (2012) suggested that different
approaches for assessing animal welfare could be conceptualized in that animal welfare
“comprises the state of the animal’s body and mind, and the extent to which its nature is
satisfied”. However, even with a definition conceptualized, different aspects of welfare (body,
mind, and nature) could be in conflict, resulting in practical and ethical challenges (Duncan,
2005; Hewson, 2003). The concept of integrity was then supported by eliminating such conflicts.
Animal integrity provides an integrated view, thus supporting the above definition of health and
welfare (Decuypere et al., 2010). For example, restriction of food (caused by humans) and high
temperature (caused by climate change) can influence the welfare of broilers. Animal integrity
can only be affected by the human action. It means that integrity demands human respect,

30
suggesting that animal integrity is more a philosophical than an empirical issue (Christiansen and
Sandøe, 2000).
Consideration for the welfare of chickens has increased in recent years and has been prompted
mainly by the negative effects of intensification of production systems, as by the concerns of
society with sustainability issues (Vermeir and Verbeke, 2006).
The foundation of civil associations or organizations pushing governments to change legislation
and encouraging farmers to produce animals in more sustainable systems with minimum
standards for animal welfare has been one of the results of public engagement. For example,
IFOAM, the International Federation of Organic Agriculture Movement, was founded in the
1970s due to concerns on sustainability issues and animal welfare. Nowadays, IFOAM
influences legislation for organic production worldwide based on its principles and ideals
(IFOAM, 2005). Such principles encourage resilient production systems based on ecological
processes where genetic variability and local conditions should be explored. Animal welfare, in
the IFOAM concepts, are very much in accordance with what is expected for animal integrity, in
which farmed animals should be respected and access opportunities and conditions should
encourage their natural behaviour.
In intensive organic systems, drugs and antibiotics are substituted by alternative products and
broilers are raised for longer periods with a minimum age for slaughter. In addition, broilers are
allowed access to outdoor areas even though in practice little or no access may be observed
(Fanatico et al., 2007).
Substitution of the production inputs is part of the transition process to sustainable methods of
food production (Gliesman, 2000). However, few examples of processes in holistic systems go
beyond the input substitution. In this respect, a strong debate in the organic movement refers to
this approach as the “conventionalisation” of the organic agriculture. Oelofse et al. (2011),
Darnhofer et al. (2010) and Guthman (2004) provide interesting insights into this debate, but
with focus on vegetables fruits and crops.

31
In the case of organic broilers, it is fair to refer to “conventionalised” intensive systems because
these are often heavily dependent on organic oil seed crops and cereals and other input
substitutes. Such dependence leads to high production costs, reflected in higher prices for
consumers. In addition, conventionalised systems may fail to meet the principles and ideals of
organic farming. Organic grains and cereals are the main ingredients for feed and to a high extent
compete with food for humans. Higher mortality rates may also be observed than in conventional
systems where supplementation of chemotherapeutics is routine. In the conventionalised organic
broilers systems, the absence of alternative products to prevent or alleviate the effects of
coccidiosis is a very important limitation as it affects animal welfare.
In the production of chickens, it has been difficult to find examples of holistic systems following
the principles and ideals suggested by IFOAM (2005). An example of a more holistic approach is
the organic Label Rouge production in France that uses slow-growing genotypes, limits stocking
density and size of the flocks. However, the production is normally managed on farms where
permanent plots are used, increasing the exposure to parasites (Permin and Hansen, 1998).
Whether intensive organic production or not, in both systems coccidiosis is one of the most
important obstacles for the production of organic chicken meat. Animal welfare may be
compromised depending on the severity of the infections. However, the degree of such
compromise may vary according to the production system because slow-growing broilers are
expected to cope better with infections (Star et al., 2008).
Humans have different values, ethical backgrounds and thus perceive animal welfare differently
(Sandøe and Christiansen, 2008) which will be reflected in their decision on whether to buy
organic chicken meat. In addition, the different ethical backgrounds of farmers influence their
choice of system for organic meat production. If a more economic orientation is observed, the
input substitution would be sufficient to produce organic broilers and thus benefit from premium
prices. In another perspective, farmers may try to diversify their production systems and thus
benefit from synergies and complementarities of the inclusion of free-range broilers in their
production systems. In addition, the availability of vegetation would be an important advantage
in the eyes of the consumer (Heckendorn et al., 2009).

32
In both perspectives, however, more information is necessary to support the production of
broilers in organic systems, especially in search for alternative nutrient sources and to control
coccidiosis.
1.8 Project justification, objectives, hypothesis and main research questions
In organic systems broilers must be fed diets composed almost entirely of organic ingredients.
These ingredients are currently mainly oil seed crops and cereals. Organically produced crops
and cereals are expensive for feeding broilers due to the competition with human nutrition. In the
past, chickens were commonly left to roam, seeking essential parts of their nutritional needs on
or below the soil surface (Prothero, 1936). For example, Robinson (1961) estimated feed savings
of 10% when fresh grass was available to hens in free-range systems. More recently, Horsted and
Hermansen (2007) reported that a modern genotype of laying hens restricted in commercial feed
and foraging different types of vegetation was able to meet essential parts of its nutritional
requirements by foraging and it was hypothesized that it would also be possible for broilers.
Even though the genetic background of chickens influences foraging behaviour (Nielsen et al.,
2003), animal husbandry and management practices are very important. In commercial free-
range systems, broilers are normally raised in fixed houses with access to permanent outside
areas. On the one hand, very little attention has been given to making outdoor runs attractive and
nutritious to broilers (Van de Weerd et al., 2009). On the other hand, chickens are more exposed
to parasites when raised with access to permanent paddocks (Permin et al., 1999). In addition,
access to pasture plays an important role in the development of the immune system of chickens
due to a higher exposure to different feed resources and forms of life (Combes et al., 2003) with
effects on product quality and on animal health (Lopez-Bote et al., 1998; Sossidou et al., 2010).
More specifically, it improves the oxidative stability of the meat (Castellini et al., 2006).
Slow-growing broilers are apparently immunologically different in terms of resistance to
pathogens when compared with fast-growing genotypes (Pinard-Van Der Laan et al., 1998). This

33
was proposed after comparing different lines for resistance to coccidiosis and observing an
Egyptian genotype with a strong protective response against Eimeria tenella infections.
Interestingly, different lines from the same breed (Fayoumi) showed differences in susceptibility
to avian coccidiosis (Kim et al., 2008). Ayssiwede et al. (2011) also reported higher resistance of
Senegal indigenous genotypes to multiple infections caused by Eimeria spp. compared to fast-
growing broilers.
On this background, the overall hypothesis of this project was that natural foraging could
contribute to the nutrient supply of slow-growing broilers, and that production systems thus
could be developed where access to outdoor areas could meet animal health concerns related to
endo-parasitic infections. Herbal products and extracts could herein be used in the management
strategies to reduce severity of avian coccidiosis, especially in systems highly dependent on
substitution of inputs.
The overall objective was to measure the nutritional contribution of two different types of
vegetation to slow-growing broilers naturally exposed to Eimeria spp. parasites and to identify
the extent to which herbal extracts supplemented under different strategies of delivery would
alleviate problems caused by coccidiosis.
Thus, the specific objectives of this project were:
- Investigate the effect of artemisinin, a sesquiterpene lactone mostly available in the
leaves of Artemisia annua, supplemented prophylactically in the diet for two different
slow-growing broiler genotypes on coccidiosis; (addressed in paper I);
- Investigate foraging behaviour of two different broiler genotypes with access to two
different types of vegetation in the finishing period and estimate the nutritional
contribution of forage; (addressed in paper II);

34
- Investigate strategies for on-farm use of herb-based products as a means of “natural”
prevention in input substitution strategies to control avian coccidiosis (addressed in
paper III and IV);
- Investigate farmers’ perceptions and ideas for strategies to substitute the use of oil seed
crops and cereals and delivery strategies of herbal products to be implemented at farm
level (addressed in paper V).
The expected outcome of this project was the identification of opportunities for improved
alternative systems for broilers in Denmark and in Brazil. A reduced reliance on oil seed crops in
feeding strategies and botanical compounds to prevent coccidiosis would improve the health
status of the broilers, improve feed efficiency and thus be reflected in lower costs to consumers.
On this background, it was assumed that this study would contribute with new scientific
knowledge and at the same time support farmers engaged in the production of broilers in
alternative systems. The Ph.D. thesis was then focused on the following research questions:
1. Are forage crops suitable to supply nutrients to slow growing broilers? If yes, which crops, to
what extent and under which management strategy?
2. Can plant extracts be used to improve the health status of broilers and thus substitute the use
of drugs in coccidia management? If yes, which plants and strategies of supplementation would
be reasonable at farm level? At which level of inclusion or dosage?
To answer these questions, this Ph.D. project benefited from four experiments and field work
with small-scale farmers in Brazil. In the next section, the outline of this thesis is presented
including the research papers supporting this work.

35
1.9 Outline of the thesis and supporting papers
This three-year project investigated strategies for the supply of nutrients to slow-growing broiler
genotypes using two different types of vegetation and four different botanical food supplements
to alleviate problems caused by coccidiosis in alternative production systems. In addition to the
four experiments, the project benefited from an exploratory field study with three smallholder
case studies producing broilers in Brazil.
This Ph.D. thesis is thus supported by the following research papers:
I. Use of Artemisia annua as a natural coccidiostat in free-range broilers and its effects on
infection dynamics and performance. Veterinary Parasitology, 186 (2012) 178– 187.
II. Feed intake and activity level of two broiler genotypes foraging different types of
vegetation in the finishing period. Poultry Science, 91 (2012) 2105– 2113.
III. The effects of cassava (Manihot esculenta), annual wormwood (Artemisia annua) and
mugwort (A. vulgaris) on coccidiosis in organically reared pullets in Brazil. (Manuscript
submitted to Asian-Australasian Journal of Animal Sciences).
IV. The effects of combining Artemisia annua and Curcuma longa ethanolic extracts to
broilers challenged with infective oocysts of Eimeria acervulina and E. maxima.
(Manuscript submitted to Parasitology).
V. Farmer´s attitudes and perceptions on feeding strategies and control of coccidiosis in
alternative broilers systems in Brazil. (Manuscript in preparation for submission to
Organic Agriculture).
In the next section, a more detailed description of the methods used in this thesis is presented.

36
2. Materials and methods
The aim of this section is to present the overall methodology used in this Ph.D. project. Initially,
the outline of the thesis illustrates the organisation of this work based on the five supporting
papers. The following items describe the experimental protocols and the methods used in animal
experimentation to evaluate the effects of herbal extracts on coccidiosis and the strategies used to
measure the contribution of forage in the nutrient supply of the slow-growing broilers.
2.2 Description of methods
2.2.1 Protocols of experiments performed in Denmark and in Brazil
Two experiments were performed at Research Centre Foulum, Faculty of Science and
Technology at Aarhus University, Denmark (56° 48′ N, 9° 58′ E).
The first experiment is presented in Paper I. A 2 x 2 factorial design was used – with two
genotypes (White Bresse L40 and Kosmos 8R) and two dietary treatments (with or without
supplementation with A. annua dried leaves) as main factors – and with three replicates for each
treatment combination. Twelve small flocks each with 35 broilers of mixed sex were raised for
70 days. The dissemination of coccidia in the flocks by seeder and contact broilers and
parasitological assays to follow infections were used and are described in this chapter. Dried
leaves of A. annua produced in Denmark were mixed in the diet to the level of 3% DM and fed
to broilers to evaluate the effects of the raw material on infection dynamics and performance.
The second experiment performed in Denmark, presented in Paper II, was a continuation of the
first experiment and also used a 2 x 2 factorial design. The rationale behind the second
experiment was that the feed conversion rate would be impaired in the finishing period and the
access to highly nutritious vegetation would reduce the use of oil seed crops and cereals in the
broilers’ nutrition. Twenty-five broilers of mixed sexes were randomly distributed to 12 flocks.
In this study, the main factors were the access to two different types of mixed vegetation: grass
(Lolium perenne) and clover (Trifolium repens) versus a mix of Chicory (Cichorium intybus cv.

37
Grassland Puna) and spontaneous weeds (Lolium perenne, Trifolium repens, Artemisia vulgaris,
Senecio sp. and Tripleurospermum sp.). Weight gain, feed intake and feed conversion rate were
estimated. In addition, feed selection habits and forage intake were evaluated by measuring crop
content while foraging activity was estimated by behavioural studies. The aim of this study was
to estimate the contribution of forage to the overall nutrition of two different slow-growing
genotypes.
The third experiment is described in Paper III and was performed at an organic commercial
farm located in the estate of São Paulo in Brazil (22°38′ S, 47°00′ W). The experiment was
linked to the farm routine, following its production schedule. One-day-old Isa Brow pullets were
randomly allocated to one of 14 pens (165 pullets per pen) thus allowing two replicates for each
of seven different treatments. In this study, the main objective was to identify the effects of the
ingredients cassava (Manihot esculenta), annual wormwood (Artemisia annua) and mugwort (A.
vulgaris) on coccidiosis under different strategies of delivery − in water and supplementation in
feed to the level of 3% DM weight.
The fourth experiment, described in Paper IV, compared different products to control
coccidiosis in the production of fast-growing broilers, investigating strategies for input
substitution. The study was performed in an experimental facility of a Brazilian company (Korin
Agropecuária Ltd.) engaged in the production of organic and natural (antibiotic-free) broilers in
vertically integrated intensive systems, located in the estate of Sao Paulo (22° 24′ S, 47° 41′ W).
Fast-growing male broilers (Cobb S500) were randomly distributed to 48 pens with 180 broilers
per treatment. A combination of Artemisia annua and Curcuma longa ethanolic extracts in three
different dosages were supplemented via drinking water to broilers infected with E. acervulina
and E. maxima oocysts. The combination effects against dual infections were compared to
options available in the market for organic and conventional production systems. Weight gain,
feed intake, feed conversion rate, lesion score, OPG (oocyst per gram of faeces) and mortality
were assessed to compare different treatments.
In Paper V, three groups of small-scale farmers producing “caipira”, natural (antibiotic-free) and
organic broilers situated in four different regions in Brazil were investigated. An assessment of

38
the most relevant constraints based on different production systems and animal welfare was
conducted. The aim was to examine the main limitations from the farmers’ point of view in
terms of novel strategies for the supply of nutrients to broilers and for the control of avian
coccidiosis. The field work was conducted through the complementarities provided by a
combination of sociological and agronomic research methods based on qualitative interviews,
observational studies and questionnaires with semi-structured questions.
2.2.2 Parasitological methods and animal experimentation
Two different procedures were used for infecting experimental broilers. In the first experiment
(Paper I) 60 broilers (half from each genotype) were raised indoors for 10 days and their
bedding material was contaminated with manure (20 kg scattered homogeneously on the ground)
obtained from a backyard farm and tested positive for Eimeria spp. oocysts (mean ~ 48.000
oocysts/g of bedding material from three replicated samples). After this period, these seeder
broilers began to shed oocysts, ranging from 40 to 560 oocysts/g of faeces (OPG), and five
randomly selected seeders were introduced to each plot with contemporary 30 broilers to
transmit the disease by contact as suggested by Velkers et al (2010).
In the third experiment (Paper III), pullets were naturally infected with the seven species of
Eimeria that were available in the chicken house from previous flocks.
In the fourth experiment (Paper IV) three fast-growing broilers per pen (n=126) (representing 10%
of broilers in each pen) were artificially inoculated by gavage with a 1 mL distilled water
suspension containing 5 x 104 E. maxima + 2 x 105 E. acervulina sporulated oocysts. These
broilers were nominated as seeders and were raised for nine days with contemporary broilers
with the aim of transmitting the disease by contact (Velkers et al., 2010).
For monitoring subclinical infections, OPG (Papers I, III and IV) and lesion score (Papers I
and IV) were used. A standardized McMaster technique adapted from Roepstorff and Nansen
(1997) for monitoring the dynamics of Eimeria spp. was used.

39
In summary, faeces samples were examined for coccidia oocysts by the modified McMaster-
technique using saturated sodium chloride solution with 50% glucose monohydrate as flotation
fluid. Faeces were diluted and well homogenized in tapwater depending on sample weight,
proportional to 4 g of faeces in 56-mL of tapwater. After 30 minutes standing and re-suspension,
the solution was filtrated through gauze (aperture 250 μm) to remove debris. Graded centrifuge
tubes were identified and filled to the limit of 10-mL and centrifuged (1200 rpm) for seven
minutes.
The supernatant was removed and the pellet containing oocysts was stored in the fridge. Next
day, tubes were left for 30 minutes at room temperature and the flotation fluid was added to 4-
mL. A disposable pipette was used to mix flotation solution with the pellet and quickly transfer
the homogenized mixture to the McMaster slide. After five minutes standing to allow oocysts to
float, oocysts were counted at 100x amplification using a microscope. Each oocyst visually
identified in each of the 12 rows scanned on the McMaster slide corresponded to 20 oocysts per
gram of faeces (OPG).
Including the samples analysed during my learning activities and during a practice run to master
the technique before experiments began, more than 2000 samples were processed in this PhD
project. During the experiments, the McMaster method was applied to identify infection in
individual broilers (Paper I), results pooled representing 10% of broiler flocks (Paper IV) and
in flocks (Paper III).
In Paper I, droppings from ten individuals in each plot (males and females) were collected twice
weekly during four consecutive weeks. In Paper III, droppings were collected from the ground
according to Long and Rowell (1975) on a weekly basis, thus representing flocks of up to 200
young chickens. In total, faeces from 14 pens were collected over a period of 14 weeks.
In Paper IV, pooled samples from “tracers” representing 10% of the flocks were collected twice
weekly during four consecutive weeks, representing infection dynamics of each of the 48 pens.

40
Droppings were collected from individual cages (Paper I), collective cages (Paper IV) and from
the ground (Paper III). Faeces were commonly stored in disposable containers, kept in
refrigerated boxes and transported to laboratories within 24 hours of sampling.
In the laboratory, a reading procedure was developed as part of the McMaster method. After
adjusting the focus of the microscope (using 40 times focus lens) and thus identifying the rows of
the McMaster chambers, the 100x magnitude lens (10x10) was used for counting the oocysts.
Figure 7 shows oocysts at this amplification.
Figure 7: Oocysts visualised in the McMaster slides used for estimating OPG (100x magnitude).
If morphological evaluation is intended, 400x amplification may be used as shown in Figure 8.
However, morphological identification is difficult as oocysts are very similar in shape and size
(Thebo et al., 1998). Even a trained person would require molecular evaluation to a 100% precise
diagnostic for species involved in infection.
Figure 8. Oocyst visualised at 400x magnitude.

41
During the adaptation of the McMaster method, for each reading method described below, a
check was performed to ensure repeatability of the counting procedure. Based on the number of
oocysts observed in a first visualisation on the McMaster slide, five different counting strategies
were used as follows.
The McMaster slide (Figure 9) contains two chambers. Each chamber contains six rows. In total,
there are 12 rows in the slide. All OPG counting used in this PhD study followed a first rule:
Start counting from the first row on the left side of the left chamber in the McMaster slide. From
this first visual evaluation:
Figure 9. McMaster Chamber and the strategies used for counting oocysts
1. If no oocysts were identified in the first row scanned, all rows were thoroughly
scanned and the total number of oocysts available in the 12 rows summed. The total
number of oocysts was equalised to N and then multiplied by 20 (sensitivity of the
test) to estimate the number of oocysts available per gram of faeces N x 20 = OPG ;
2. If many oocysts were identified in the first visual scan in the McMaster slide, the
number in the first row was counted and summed. When the number (N) of oocysts in
the first row was in the range of 200 and 300, oocysts in other two randomly chosen
rows were scanned and counted. The numbers counted in the three rows were them
summed, divided by three and multiplied by 12 - number of rows in the McMaster
slide and then multiplied once again by 20 (sensitivity of the test).
(N row 1 + N row 2 + N row 3) / 3 x 12 x 20 = OPG ;

42
3. If the number (N) of oocysts in the first row was in the range of 300 and 500,
counting only one row was enough and then N was multiplied by 12 (rows).
N row 1 x 12 x 20 = OPG ;
4. Where a large number of oocysts was observed in a first visual scan, a specific area
on the McMaster chamber was chosen and the number of oocysts available in that
specific eye field was counted. Counting too many oocysts is a challenge and a useful
strategy was to choose a specific area for setting up the eye field close to air bubbles
that represent a mechanical clock. In this strategy, counting could be performed in
“slices of pizza” and the air bubbles were used as references. By estimating the area
for the eye field, the total N number was then multiplied by 1060 to apply the same
sensitivity of the test. N x 1060 = OPG.
5. If the number (N) of oocysts counted in the first chamber summed to more than 500,
only one chamber was counted and N multiplied by two and thereafter by 20.
N chamber 1 x 2 x 20 = OPG
Molecular evaluation (Figure 10) is necessary for a 100% confident diagnosis of species of
Eimeria causing infections.
In Paper I, one polled faecal sample from all ten broilers
(males and females) per pen (n = 12) was stored as pellets
(after dilution of faeces as presented before) at −18°C for
four months before being examined by PCR for seven
species of Eimeria (Haug et al., 2007) at the Laboratory of
Parasitology, Swedish Veterinary Institute. The protocol
(GeneRelease) is expected to detect from 0.4 to 10 oocysts
per PCR for the seven species of Eimeria obtained from
field samples (oocysts in the presence of debris).
Figure 10. Output from the PCR test

43
In Paper III, oocysts were purified from pooled faecal samples and analysed by multiplex PCR
(Fernandez et al., 2003) in the Laboratory of Parasitology, Institute of Biological Sciences -
University of São Paulo, Brazil.
2.2.3 Analysis of crop content of broilers and sward harvesting
A practical method to determine feed selection habits of chickens raised in free-range systems is
to evaluate the content of the crop at different times of the day (Horsted, 2006). Only a few
studies have evaluated the food habits of highly productive hens in free-range systems (Horsted
et al., 2007; Horsted, 2006; Antell and Ciszuk, 2006) and in scavenging indigenous genotypes in
developing countries (Mwalusanya et al., 2002; Sonaiya, 2004). No literature on broilers bred for
the production of meat was found.
Mongin (1976) reported that the crop can be expected to be full in the evening since the amount
of dry matter (DM) in the crop follows a cyclic variation influenced by the photoperiod, which
agrees with the findings of Horsted (2006). A similar hypothesis could be expected for foraging
broilers, but variation could also be expected based on the broilers’ genetic background and sex.
In practical terms, broilers (males and females) were harvested (cervical dislocation) at two
different times of the day (morning and evening) over three different periods in the growing
period (beginning, midway and final part of the experiment) and the entire crop was removed
immediately after harvesting (Paper II). Crops were stored in plastic bags, identified with the
bird ID and plot number and stored in the freezer for future analysis.
Following Jensen and Korschgen (1947) and more recently applied by Horsted et al. (2007),
individual crops were thawed in the laboratory and the content separated by forceps into different
fractions as schematically presented on Figure 11: “earthworms, larvae and pupae”; “grass”;
“clover”; “chicory”; “insects”; “seeds”; “soil”; “commercial broiler feed”; “grit stones” and
“wheat”.

44
All fractions were dried in a forced air-drying oven at 60ºC for approximately 24 hours and
calculation of crop content was based on weight of air-dried fractions (Antell and Ciszuk, 2006;
Horsted et al., 2007).
Figure 11. content of crops separated into different fractions. From top left: “earthworms,
larvae and pupae”; “grass”; “clover”; “chicory”; “insects”; “seeds”; “soil”; “commercial
broiler feed”; “grit stones” and “wheat”.
The total plant intake was estimated by summing the total herbage material found in the crops to
evaluate the vegetative food intake and consequently the food habits during the experiment and
from different periods of the day (morning and evening) (Paper II).
Before broilers accessed the paddocks with vegetation, samples were harvested from two 0.25-
m² patches from each of the 12 plots investigated. The location of the patches for sampling was
decided beforehand and the same locations were used for all plots investigated. The vegetation
was cut approximately 2 cm above ground (Figure 12) and the harvested biomass from the two
patches was pooled for each plot and stored in the freezer for further chemical analysis.

45
Figure 12. harvesting biomass for chemical analysis
With data on daily feed intake and nutritional value for the restricted diet (commercial broiler
feed and whole wheat) and for the vegetation found in the crop of the broilers, an estimation of
the daily contribution of forage (including genotypes and sex) on the nutrient supply of broilers
could be calculated.
Supplement of herbage to broilers as a strategy of nutrient supply requires good husbandry
practices. To identify degradation of vegetation with time, visual assessments were performed.
Vegetation degradation was estimated by four different visual measures in a 1-m² area in each
plot with the same location used for all plots.
The percentages of the main components of grass, clover, chicory, weeds, bare soil and
decomposed vegetation were estimated for each plot to a sum of 100%. The change in relative
vegetative cover of the different components was compared to represent the degradation of
herbage with time and results were averaged according to treatment combinations.
Based on the contribution of forage to the nutrient supply of slow-growing broilers in addition to
the degree of degradation in the vegetation areas, strategies for paddock rotation could be
suggested for Danish conditions.

46
2.2.4 The ethanolic extraction of plant components
Previously (section 1.4), four different species of plants were described and were used in this
PhD study as a supplement to chickens as a means of coccidial management. Whole A. annua
plants produced in Denmark were dried in a ventilated room at approximate 20°C. Leaves were
ground by a hammer mill and sifted through a 3-mm mesh and the powder was added to the diet
based on 3% w/w of slow-growing broilers (Paper I). Leaves of M. esculenta (cassava) were
dried in the shade at room temperature (22-23°C) on the farm and supplemented at 3% w/w in
the diet to pullets (Paper III). In the Brazilian experiment, A. annua whole plants were dried in
the sun for one week and leaves were separated from secondary stems by hand. Leaves were
ground by a hammer mill and sifted through a 5-mm mesh and were also included in the diet of
pullets to the same level of inclusion - 3% w/w (Paper III).
The dosage used for the ethanolic extracts was initiated following suggestions presented by a
visionary organic farmer who was using the ethanolic extract of A. vulgaris in the control of
clinical signals of coccidiosis in the production of organic pullets (Paper III). The farmer’s
Ethanolic herbal extracts were prepared by using dried leaves of
A. annua (Paper III and IV) and A. vulgaris (Paper III) and
powder from the root of C. longa (Paper IV). In summary, in 1-L
Erlenmeyer flasks wrapped with aluminium foil to prevent
oxidation and degradation by light, 100 g dried leaves of A.
annua and A. vulgaris (without small branches and crushed by
hand) or 100 g gridded powder of C. longa were mixed with 1 L
ethanol 70% (700 mL ethanol 97° + 300 mL distilled water).
The mixtures were kept in a dark room at a controlled temperature
(22°C) and homogenised with circular movements twice per day
for 20 days. The content was filtered through paper filters and
transferred to 50mL flasks with dropping glass dispensers. Flasks
were identified and stored in the fridge until needed.

47
approach was to supplement 15 drops in approximately 25 L of water on a daily basis to groups
of 160 pullets after the appearance of the clinical signals of coccidiosis.
Allen et al. (1997) reported preventive effects of dried A. annua leaves in caged fast-growing
male broilers against individual and dual infection with Eimeria tenella and E. acervulina. In
their study, the leaves were added to the diet, at maximum dosages of 4-5 mg artemisinin per kg
body weight per day, and the birds were not subjected to re-infection. These authors reported that
a prophylactic strategy by a three-week supplementation period prior to the challenge was
required to give a reduction in oocyst output of E. acervulina and increased performance.
To this end, supplementation of A. annua dried leaves was provided before any infection was
expected (Paper I and paper III) and ethanolic extracts were supplemented before expected
infections based on the farm circumstances (Paper III) and for the total growing period when
fast-growing broilers were investigated (Paper IV).
2.2.5 Investigating farmer’s opinions
In Paper V, three farm level case studies of alternative production of broilers were performed
with two main objectives: Firstly, to investigate the extent coccidosis affect the production of
broilers in systems not relying on chemical medicines, and, secondly, exploring farmers’ ideas
on alternative ingredients to substitute the use of maize and soybeans in nutrient supplementation
to broilers, to tackle the future challenges of the production of broilers in free-range systems in
Brazil. The research was developed through the complementarities provided by a combination of
sociological and agronomic research methods, based on qualitative interviews (Kvale and
Brinkmann, 2009) and a questionnaire with semi-structured questions (Almeida and Abreu,
2009).
Each case study consisted of a sample of collaborative farmers. Case areas were selected
following discussions with local researchers and stakeholders based upon the following criteria:
(1) that smallholders were producing broilers without chemical remedies and medicines in
alternative production systems in the regions where most of the production of chickens for meat

48
takes place in Brazil, (2) that farmers should be small-scale producers or family farmers, and (3)
they should be linked to an alternative supply markets.
Data collection acquired household information, detailed production data as well as qualitative
information of farmers' perceptions of aspects related to farming systems. The focus of the
interviews was to gain knowledge of the beliefs of the farmers and the constraints to strategies to
reduce the use of oil seed crops and cereals in broiler nutrition by suggesting local alternative
feed sources and forage crops for foraging broilers.
3. General results and discussion
This section is structured according to the scientific application of the results obtained in this
Ph.D. thesis. Thus, three main topics discussing the results are presented in respect to
management strategies for feeding and controlling coccidiosis in the production of broilers. The
first topic discusses the main results obtained in the four experiments described in Papers I to
IV and reported by farmers, described in Paper V. The second topic discusses how the major
results obtained in this Ph.D. study may find applications in husbandry practices for the
production of broilers, thus supporting further development for the sector. The third topic deals
with some implications of the different production systems investigated for the development of
the organic sector.
3.1 Main results from the individual papers
In Paper I, the prophylactic supplementation of A. annua in feed reduced the number of excreted
oocysts (60-70%) with no interaction with genotype (Pure bred slow-growing White Bresse L40
and hybrid medium-growing Kosmos 8R). In general, females excreted more oocysts than males,
supposedly due to differences in foraging behaviour. The overall body weight gain and the daily
weight gain when infections were subdued showed a three-way interaction between genotype,
sex and treatment - mainly because hybrid females responded positively to the A. annua

49
treatment, while the hybrid males responded negatively, and the only minor differences were
found between sexes for the pure slow-growing genotype.
Females were more active in pecking for feed ingredients in the range and this behaviour
presumably contributed to an involuntarily higher intake of oocysts from the ground compared to
males. In this respect, the effect of treatment was more pronounced in females of the hybrid
genotype Kosmos 8R. Untreated Kosmos females excreted 75% more oocysts than treated
females of the same genotype and gained around 100 g less weight in the experimental period as
a consequence of the higher coccidial challenge (Paper I).
Oocysts are excreted with faeces and faeces accumulated closer to the broiler houses and to the
feeders. Due to less time spent foraging, this was more pronounced in plots with Kosmos broilers
as also found by Dawkins et al. (2003), who reported differences in foraging behaviour and
attributing these to the genetic background of broilers. However, hybrid Kosmos males treated
with A. annua dried leaves gained less weight than untreated broilers without differences in
oocyst excretion. A similar result in another livestock has previously been attributed to the
unpalatability of components of the A. annua dried leaves restricting feed intake (Caygill and
Mueller-Harvey, 1999). In general, the supplementation of A. annua dried leaves reduced the
oocyst output. However, the bitter taste of the leaves inhibited feed intake and thus reduced the
capacity for a higher weight gain for the flock as a whole (Paper I). These results highlight the
need of identifying alternatives for the supply of the herbal components with anti-coccidial
activity without compromising performance attributes in production.
A three-week supplementation period of 4 mg artemisinin/kg/day prior to oocyst exposure was
necessary to provide protection against E. acervulina infections. In addition to the unpalatable
components and in accordance with Allen et al. (1997), the supplementation of A. annua dried
leaves was not efficient in controlling E. maxima infections. Arab et al. (2006) also reported no
effects of pure artemisinin against E. maxima, suggesting that coccidial ingredients available in A.
annua have a species-specific mode of action.

50
In Paper II, the same broilers from the previous experiment were restricted in supplementary
feed (50 g commercial diet/broiler/day), while whole wheat was supplied ad libitum in the
finishing growing period. Broilers of both genotypes were allowed to forage two different mixed
vegetations (grass/clover or chicory/weeds) in order to identify the benefits of herbage on
nutrition during this period (80 to 113 days of age).
The type of vegetation available to the broilers did not influence the use of the free-range area,
nor performance attributes. By estimating forage degradation with time, it was observed that the
grass/clover vegetation was more resistant and persistent under grazing compared to the chicory
plots. Differences in the use of the range area, activity level and feed content in the crops were
observed in relation to genotype, sex, age and also for the time of the day. Foraging activity was
positively correlated with age of broilers. In general, the hybrid broilers spent more time inside
and closer to the houses during the day with increased foraging activity during evenings, in
contrast to the pure slow-growing broilers that showed a more uniform level of activity during
the day (Paper II).
Based on the measurement of crop content, it was estimated that the pure-bred slow-growing
broilers had a daily intake of 5-8 g of forage DM per day, while the hybrid broilers had an intake
of 9 g for females and 20 g for males. This was a surprise and it was hypothesized that due to a
relative higher restriction in supplementary feed (a fixed amount of supplemental feed, but a
higher growth rate), hybrid male broilers compensated by increasing the intake of vegetation.
The limitation of supplemented commercial feed in the finishing period is practical for slow-
growing broilers that have access to highly nutritious vegetation. However, the consumption of
forage was able to substitute only 10 to 15% of the methionine and 10% of energy requirements
for growth and maintenance of the broilers (Paper II). Perhaps a tighter restriction would have
increased the intake of forage as was reported by Horsted and Hermansen (2007) for layers
restricted in commercial feed and foraging in highly nutritious vegetation.
In Paper III, the results of an on-farm experiment to investigate potential natural products and
strategies of delivery of natural coccidiostats is presented. In summary, one-day-old pullets were
randomly allocated to 14 pens (165 pullets per pen) thus allowing two replicates for each of

51
seven different treatments. One group of pullets was supplemented with 3% of DM cassava
(Manihot esculenta) dried leaves during the first eight days of life, while two groups were
supplemented with 3% of DM Artemisia annua either for the first eight weeks or from week 4 to
8. One group was supplemented with concentrated ethanolic extract of A. vulgaris in water
before expected appearance of clinical symptoms, while another group was supplemented with A.
annua extract in water after the expected appearance of clinical symptoms. The response of the
above five treatments was compared to a vaccinated group and to the farmer’s routine of
supplementing an ethanolic extract of A. vulgaris in water after appearance of clinical symptoms.
Weight gain, feed intake and coccidia (Eimeria spp.) oocyst excretion (OE) were monitored
weekly from 1 to 14 weeks of age.
In this study, it was found that the supplementation of A. annua at 3% of DM reduced the growth
rates of pullets without affecting oocyst excretion (OE), the lower growth rate was supposedly
because the bitter taste imposed by sesquiterpene lactones available in the A. annua leaves that
give a low palatability restricting feed intake. In addition, in this experiment, as opposed to the
first experiment reported in Paper I, a prophylactic supplementation before the infection took
place was not possible due to availability of oocysts in the farm environment at an early stage.
This supports the hypothesis previously provided by Allen et al. (1997) that a three-week
supplementation period prior to the challenge was required to reduce oocyst output of E.
acervulina and increase performance.
In the present work, supplementation of cassava (M. esculenta) dried leaves markedly suppressed
OE (50-60%) without affecting the growth rate compared to the vaccinated group. While the
effects of ethanolic plant extracts with or without artemisinin were limited in terms of OE,
probably because the time of supplementation was insufficient to provide protection, a daily 3%
of DM supplementation of cassava dried leaves in the beginning of the rearing period was found
to effectively reduce Eimeria spp. OE (Paper III). The effect of cassava dried leaves against
Eimeria spp. was attributed to the presence of condensed tannins (CT) and the daily dose given
over eight consecutive days was estimated at 2 mg CT/pullet.

52
Based on results from papers I and III with respect to the strategy of delivery (in diet or in water)
of A. annua extracts, in addition to its failure in treating Eimeria maxima parasites − another
economically important species affecting the production of broilers (De Gussen, 2007) − a
combination partner to A. annua was sought. Curcuma longa was reported to be used in the
control of avian coccidiosis by farmers and, more specifically, it has been reported to be efficient
against Eimeria maxima (Allen et al., 1998). Three different dosages of a combination of herbal
extracts supplemented in drinking water were therefore compared to chemical coccidiostats and
to products available for organic systems - vaccination and a commercial product based on
oregano oil extract against dual infections caused by E. acervulina and E. maxima (Paper IV).
In Paper IV fast-growing broilers (Cobb S500) supplemented with chemical coccidiostats gave
superior performance attributes compared to all other groups investigated. However, a dose-
dependent reduction of lesion scores was observed for the mixture of herbal extracts (Artemsia
annua + Curcuma longa). On average, broilers supplemented with the highest dosage of herbal
extracts achieved intermediate scores, which was higher than broilers supplemented with
chemical coccidiostats but lower than broilers supplemented with the products available for
organic systems (vaccine and a product based on oregano oil extract) and negative controls. A
trend for lower mortality (p=0.08) in the last part of the growing period (23-43d) was observed in
broilers supplemented with the highest dosage of the herbal mixture Artemisia annua + Curcuma
longa ethanolic extracts as compared to broilers supplemented with chemical coccidiostats. This
suggests that the delivery strategy is practical for implementation at farm level and that this
herbal combination has an interesting future in alternative strategies for coccidia treatment. The
highest dose (27 ppb Artemisinin + 107 ppb Curcumin) was supplemented prophylactically in
drinking water from day 1 of the broiler’s life, two to three weeks before infections were
introduced by contact broilers (Velkers et al., 2010) to imitate the disease transmission that
normally takes place at farm level (Papers I and IV).
In Paper V, two groups of farmers raising free-range slow-growing broilers and organized in
cooperatives in the South of Brazil and a third group of farmers producing antibiotic-free (AF)
fast-growing broilers linked to a private company in the State of São Paulo were interviewed.
Individual assessments of production systems were performed for 21 farms in the three case

53
studies. The main limitations, advantages, and challenges in production were investigated.
Groups producing slow-growing broilers in free-range systems linked to the two cooperatives
revealed high costs of feed ingredients and the organization of the production chains was the
most important limiting factor. The third group ranked coccidiosis as the most important
limitation due to the absence of prophylactic treatments in the production of the AF broilers.
Free-range broilers were exposed to permanent paddocks with very little vegetation and plots
were not managed with the aim of supplying nutrients to the broilers.
Fast-growing genotypes used in the production of the AF broilers were challenged with
coccidiosis and necrotic enteritis based on the farmers’ opinions. Farmers producing free-range
broilers were demanding alternative sources of feed ingredients to reduce production costs,
while farmers producing the AF broilers were demanding better strategies to control coccidiosis
and thus improve yield and welfare of broilers. Moreover, based on farmers’ opinions, a list of
ingredients to substitute maize and soybeans in feeding strategies and a list of forage species
with potential use in paddock cultivation for foraging broilers are presented for the Brazilian
conditions. Finally, most of the farmers engaged in the production of alternative broilers
suggested drinking water as the most efficient delivery strategy to supplement herbal products in
strategies to control avian coccidiosis.
3.2 Turning major results into practical husbandry
With the aim of turning the results into practical improvements in production systems, results are
discussed in relation to three main phases in broiler production as suggested by Pedersen et al.
(2003).
After observing pioneer farmers producing organic broilers in Denmark, Pedersen et al (2003)
suggested that the starting period is from when chicks are one day old and until broilers are
moved to other farms or until they are moved to a house with restricted access to an outside area
at the age of 20 to 24 days. In a second period, defined as the transition period, broilers are raised
with access to free-range areas until 50 days of age. The (third) finishing period was suggested to

54
comprehend the period from 50 days to the end of the production cycle when broilers are
slaughtered.
Even though this division was originally proposed to describe the production of free-range
broilers in Denmark by pioneer organic farmers, the three main periods suggested would also be
appropriate for describing the production of natural and organic broilers raised in intensive
systems in Brazil. In these systems, differences in husbandry are associated with changes in
dietary supplementation. Feed programmes are normally divided into five main periods,
depending on the dietary content of protein: initial (1-8 days), growth (9-22 days), fattening I
(23-29 days), fattening II (30-36 days) and final (37-43 days). In organic intensive productions,
the finishing period remains until 48-56 days and the same diet is supplemented in the final
period. Due to differences in the nutritional level of the five types of diet (data not presented)
they could easily be grouped into three main periods: starter (including initial + growth),
fattening and final.
Initial phase
It is suggested that a successful sanitary management before allocating the broilers indoors in the
first period may determine the timing and level of exposure to Eimeria spp. parasites and thus
influence the future productivity of the flocks. By postponing the coccidial challenge, broilers
would develop their immune systems and providing better resistance to the disease at a later age
(Swinkels et al., 2006; 2007). Strong sanitary measures include simple practices. For example,
cleaning and using brushfire to kill surviving oocysts from previous flocks is practical, also by
small-scale farmers. Giving time for the immune systems of broilers to develop before being
infected would reduce the chances for the primary pathogen to contaminate and also reduce
possible infections by secondary opportunist bacteria. The use of herbal supplements as part of
the farm routine in this period may improve the health status of the flocks and thus alleviate
problems caused by coccidiosis and also by secondary opportunist bacteria (Papers III and IV).

55
Transition phase
In the transition period, broilers may explore outdoor areas in free-range systems or receive a
different type of diet with less protein content. As a consequence of stress caused by different
external environmental influences, broilers may be more exposed to microbial challenges either
in the chicken houses or in the chicken runs, depending on the production system. In this
transition period, it was observed that coccidial infections transmitted by contact broilers
(Velkers et al., 2010) and the severity of the infections were not influenced by the genetic
background of broilers for the free-range system (Paper I), which disagrees with the hypothesis
of pure slow-growing breeds being better able to cope with the coccidial infections (Pinard-van
der Laan et al., 1998). However, it remains speculative how much the reduction in oocyst output
observed in this experiment (60-70%) may impact the epidemiology of the disease based on the
genotype of the broilers studied. In addition, how different broiler genotypes respond to the
treatment with natural products like A. annua at higher infection levels remains a challenge for
scientists.
While the complete elimination of the parasite seems unattainable with a natural treatment using
herbal products, a smaller infection level will lead to a fast build-up of immunity in infected
birds, supporting the overall flock resistance (Velkers et al., 2011). This is a desirable outcome
for free-range and organic systems and may still reduce the risk of infections by secondary
opportunistic organisms like bacteria.
High exposure to parasites is this period may occur, especially if housing is fixed and paddocks
are permanent in the case of free-range systems. In natural or organic intensive systems, the
bigger challenge could be the absence of ionophore drugs to treat coccidiosis. In this situation,
we found a higher infection peak at 23 days of age, coincidentally when broilers received a diet
containing less protein (Paper IV). Cases of necrotic enteritis (caused by the opportunist bacteria
Clostridium perfrigens) reported by farmers in Paper V were the consequence of primary
pathogens (coccidia) that usually pave the way for an invasion of opportunist bacteria (Colier et
al., 2008).

56
Finishing phase
In the finishing period, a positive correlation between age and foraging activity was observed,
suggesting that broilers do learn how to explore range areas with age, so the sooner the broilers
access the chicken runs, the sooner they may become proficient in foraging (Paper II). Another
important aspect is that broilers have preferred times of the day for exploring the range, which is
strongly correlated with the genetic background of the broilers (Paper II). It is possible to
increase the capacity of broilers to explore the forage areas by restricting the commercial diet.
This can be done based on the timings of broilers’ preferences for the outdoors (Dawkins et al.,
2003). Perhaps supplementing commercial feed to broilers before mid-day would stimulate a
higher activity in the outdoor runs during mornings and evenings. In this Ph.D. study, the daily
supplementation of 50g/head at around 10 am was reflected in higher foraging activity in the
evenings. This was more pronounced in hybrid broilers (Kosmos 8R) and can be verified by the
larger amount of vegetation found in the crop of males. Kosmos males ingested more than
double the vegetation of the Kosmos females and the males of the slow-growing pure breed
(Paper II). However, the dietary restriction imposed on the pure slow-growing broilers did not
influence their foraging behaviour as was suggested for the hybrid broilers, especially males. The
insignificant amounts of soil particles found in broiler crops suggest that the dietary restriction
was not sufficient to stimulate animals to seek their amino acids from elsewhere as reported by
Horsted et al. (2007), who found larger amounts of soil in the crops of layers with no access to a
commercial diet compared to layers with access.
However, for the sake of animal welfare, restricted access should be given to encourage broilers
to explore the range areas, although the supply of nutrients from forage to free-range broilers is
more uncertain if large flocks are targeted, which limits the prospects of upscaling our findings.
For the Danish farmer intent on supplementing 100% organic feed to broilers, it should be borne
in mind that broilers can consume considerable amounts of forage and other feed ingredients as
part of their nutritional needs and their intake is mainly influenced by genotype, age, time of the
day and level of restriction of commercial diet.

57
3.3 Implications for the development of the organic meat sector
Rosset and Altieri (1995) and also Gliessman (2000) suggested as a first step for converting
conventional into organic systems the reduction in the use of chemical products and their
substitution by other non-polluting resources. Afterwards, by combining the use of such inputs
with other practices like crop rotation, reuse of crop residues and animal manures, legumes,
green manures for fertilization and the control of pests and disease, new production systems and
agro-ecological landscape would jointly evolve and be achieved after some time of transition.
Ideally, this is the process to convert to more sustainable food systems.
However, due to an increased number of consumers willing to pay premium prices for the
organic products, organic farming becomes attractive and a large number of farmers who use
modern machinery and infrastructure adopted monocultures attracted by the economic
advantages of selling organic products at higher prices. This process brought to academia a
debate called “the conventionalization debate” (Guthman, 2004; Darnhofer et al., 2010).
Farmers following this regime, described as conventionalised farmers, are trapped in an input
substitution process that keeps them dependent on suppliers (many of a corporate nature) of a
variety of organic inputs (Altieri and Nicholls, 2005). Today, the majority of the organic and
natural broilers marketed worldwide are produced in strategies dependent on input substitution.
Conventionalised farmers producing organic broilers can justify, based on a range of reasons and
motivations, their decisions to use fast-growing broilers, be they economic reasons (high yield
and better feed conversion rate) or environmental benefits from avoiding the use of chemical
fertilizers and pesticides in the production of grains and cereals used to feed the broilers. The
problem appears when consumers are not willing to pay for very expensive meat products.
By substituting inputs to produce organic broilers for chicken meat, farmers will not be
competitive in the long term if they rely on fast-growing genotypes and on oil seed crops and
cereals to feed the broilers (Figure 13). The Danish case is illustrative. The dependence on

58
imports for composing the diet based on organic oil seed crops and cereals results in highly
expensive meat on the supermarket shelves with consumers reluctant to buy.
The idea behind Figure 13 was to present not only the price of chicken originating from different
production systems. Additionally, it was intended to place the price of the different chickens in a
context of systems orientation: from conventional production based on fast-growing broilers and
oil seed crops, over the substitution of inputs in production and to a more holistic system
supported by the IFOAM organic principles and ideas and also by the concepts originally
proposed by Rosset and Altieri (1995) and also by Gliessman (2000).
The closer the point representing the production system is to the right hand side of the figure, the
closer it is to the ideals and principles of the organic movement and the more advanced the
systems are in the conversion to more sustainable forms of production.
Figure 13. Price of chicken meat related to different production systems organized on a scale from conventional over different degrees of input substitution to redesigned systems. On the Y-axis: Price of chicken meat in U$ dollar/Kg (in the store). On the X-axis: System orientation based on conversion to sustainable food systems after Gliessman (2000). (Source: Prices were obtained in local currency in August 2012 - after interviewing stakeholders in Brazil, France, United States and Denmark. Prices were adjusted for U$ dollar/Kg using the exchange rate from 12 August 2012).

59
By interpreting the costs based on the production systems starting from the left side of Figure 13,
it is possible to observe a slightly higher price of the natural broilers compared to the
conventional broilers in U.S. and in Brazil. Natural broilers are produced without coccidiostats,
growth enhancers, and animal origin residues and it could be interpreted as a first step (reduction
and substitution of chemical inputs in the production of foods) in the conversion process to
sustainable food systems (Rosset and Altieri, 1995; Gliessman, 2000).
Conventionalized organic broilers are more expensive to produce than natural broilers mainly
because of the use of organic crops and cereals in the feeding strategies and this is reflected in
the price of the meat. The differences in price were more pronounced in the Danish case,
apparently because there is a higher cost associated with the import of organic ingredients
compared to organic broilers in Brazil and in the U.S. where organic grains and cereals are
comparatively cheap.
Less differentiation may be observed in the organic meat from these systems because fast-
growing broilers are used fed diets based on oil seed crops and cereals. Housing infrastructure is
similar and the inputs used in production are substituted by organic ingredients in
conventionalized systems as suggested by Darnhofer et al. (2010) and by Guthman (2004). Due
to controversies related to the real access to outdoor areas and also in respect to the genetic
background of the broilers used in the production of free-range in the U.S. (Fanatico et al., 2009),
the free-range U.S chicken meat was placed closer to conventionalized systems (Figure 13).
By interpreting the Rosset and Altieri (1995) and Gliessman (2000) approaches for converting to
sustainable food systems, the use of slow-growing broilers managed in free-range systems is
more resilient than conventionalized systems, and is closer to the principles and ideals of
IFOAM (2005). Conversion to 100% organic feed is expected to be stipulated in new European
regulations and this will supports the development of free-range broilers in more holistic systems,
especially if these ingredients originate from local resources. However, this will demand a re-
think and re-design of production systems.

60
A range of strategies may help improve free-range systems that could be financially competitive.
For example, the use of hybrid genotypes with appropriate husbandry strategies that integrate the
welfare of the broilers (i.e. guarantee of animal integrity by assuming the responsibility in
restricting commercial feed) and allow access to highly nutritious vegetation would reduce the
use of oil seed crops and cereals in the production of broilers. However, the amount of nutrients
that could be supplied to broilers in a forage area (herbage, worms, insects, etc.) may differ
widely among farms and will also depend on the time of the year, stocking rate and the quality
and utilization of the range (Bassler et al., 1999).
Free-range systems in Brazil, U.S. and in France were apparently more resilient than
conventionalized organic systems (Figure 13) because of the use of slow-growing and more
robust genotypes and access to free-range areas.
Even though these systems are characterized as more resilient, the nutritional dependence on oil
seed crops and cereals is still limiting the development of the organic meat sector. In addition,
the search for new housing systems to avoid the use of permanent paddocks and the search for
strategies to reduce parasite contamination in permanent paddocks is also necessary. In
conventionalised systems, alternative feed ingredients to substitute oil seed crops and cereals are
needed.
4. Conclusions
This Ph.D. project aimed to investigate if nutrient-rich vegetation would be an option for the part
substitution of oil seed crops and cereals in the nutrition of broilers and if herbal products could
be used in strategies to alleviate problems caused by coccidiosis. It was found that the
grass/clover combination in the Danish free-range system was more resistant and persistent
during grazing by broilers than the chicory/weeds combination. In addition, a positive correlation
was observed between age of the broilers and activity level, suggesting that the sooner broilers
are given access to the range areas, the better use can be achieved. It was estimated that the pure
slow-growing broilers had a daily intake of 5-8 g of forage DM per day, while the hybrid broilers

61
had an intake of 9 g for females and 20 g for males. In addition, consumption of forage was
estimated to substitute 10 to 15% of the methionine and 10% of energy requirements for growth
and maintenance of the broilers. Restriction of commercial feed in the finishing period increased
foraging activity, especially for the hybrid broilers, and is practical to implement by farmers.
The preventive use of A. annua as a botanical coccidiostat, at a daily intake above 4 mg
artemisinin/kg body weight, can markedly reduce oocyst output (60-70%) in broilers reared
under common organic free-range conditions and exposed to natural coccidia infections. The
oocyst output was much higher for non-supplemented females, suggesting an influence of sex,
related to either a more active foraging behaviour (resulting in higher oocyst intake), or a
physiological/immunological difference in how males and females cope with infections.
Supplementation of A. annua in feed restricted feed intake and the strategy of supplementation
did not bring major advantages that can be recommended for implementation at farm level.
Supplementation of Manihot esculenta dried leaves in feed, on the other hand, at a daily intake of
2 mg of condensed tannins during the first eight days of life did not impair feed intake and
markedly suppressed oocyst output (50-60%). However, for all supplementations, the
manipulation of raw materials to be mixed in the diet are labour-intensive and may compromise
its application by farmers.
According to farmers, the use of ethanolic extracts in water would be practical to implement at
farm level and the combination of ethanolic extracts of A. annua and Curcuma longa in
prophylactic supplementation at a daily intake level of 27 ppb artemisinin + 107 ppb curcumin
reduced lesions scores caused by a dual infections of E. maxima and E. acervulina parasites with
apparent benefits in the health status of broilers. The delivery strategy is perceived as affordable
and practical to implement at farm level. However, more investigation in the dose and
composition of the mixtures is necessary for a further recommendation for its practical use by
farmers.

62
5. Future outlook
Synthetic coccidiostats are expected to be phased out as feed additives before 1 January 2013
(COM, 2008) and developmental strategies for different production systems can be supported
based on the results of this study.
In conventional systems, perhaps the use of herbal ingredients in alternate mixtures managed
together with vaccination and other husbandry practices would support the production of food
for humans with less use of chemical contaminants and thus be more attractive to consumers.
In conventionalised systems, the main challenge remains of adapting the production system – by
searching for inputs to be substituted – in terms of feed resources, products to treat parasites and
hybrid broilers to substitute fast-growing genotypes. In such systems, herbal extracts in different
mixtures could possibly be implemented together with other allowed strategies such as
vaccination. However, these systems are still very dependent on oil seed crops and cereals.
Forage will not be feasible if fast-growing broilers are used and other feed ingredients that could
be industrialised and adapted to the vertical production chains deserve more attention.
A list of vegetal species suggested by farmers as feed resources and species that could be used in
forage areas to part supplement nutritional needs and protection of broilers was presented in this
thesis and deserves further investigation.
If the objective is to improve system resilience, the main challenge would be to adjust the
production system – by using nutrient-rich pasture rotational systems and other environmental
enrichment strategies like integration with fruit/crop production - and then introducing slow-
growing broilers in these systems (Figure 14). The herbage, fruit residues, and living organisms
available in the range would be potential feed sources for the broilers.

63
Figure 14. Slow-growing chickens foraging in plots with fruit trees.
In a complementary perspective, integration with other livestock species would also be a
beneficial strategy. Farmers suggested small ruminants as the most appropriate species to be
integrated with broilers, because they do not compete for feed resources. For example, broilers
could be managed side by side or after small ruminants (sheep or goats) in a rotational system.
If broilers are managed afterwards, ruminants could be fed by forage and moved to an adjacent
plot. Controlled foraging could provide a short sward that, on the one hand, would not
compromise the recovery of the vegetation and, on the other hand, would not hide predators, thus
providing broilers with an area to explore more closely to the ground. In such a management
system, broilers could benefit from eating larvae and insects (attracted by and) available in the
faeces from ruminants and in the vegetation. Broilers could also spread the manure when
practising their scratching behaviour, distributing nutrients in the plots that could also affect the
life cycle of internal and external parasites that could alleviate further problems for the ruminants.

64
The production of slow-growing broilers in Denmark would benefit from a large amount of
roughage during the winter season and more enrichment in the houses with alternative feed
resources. During late spring, summer and autumn, the use of movable houses (Figure 15), also
beneficial for smallholders in Brazil, would be useful for farmers producing broilers in free-
range systems, making them less dependent on oil seed crops and cereals and thus more resilient
than was observed in this study. If more holistic systems are the objective, integration with fruits
and ruminants could be an option and this would accord with the ideals and principles of organic
farming.
Figure 15. Slow-growing chickens managed in movable houses.

65
6. References
Abbas, R.Z., Colwell, D.D. and Gilleard, J., 2012. Botanicals: an alternative approach for the control of avian coccidiosis. World Poultry Science Journal 68, 203-215.
ABEF – Associação Brasileira dos Exportadores de Frango do Brasil. Estatísticas. Disponível em: http://www.abef.com.br/Estatisticas/MercadoMundial/MercadoMundial.asp. Acessed June 2012.
Adams, M., 2012. FDA finally admits chicken meat contains cancer-causing arsenic (but keep eating it, yo!). Article from Naturalnews.com from 6 June 2012. Available at: http://www.naturalnews.com/032659_arsenic_chicken.html. Accessed June 2012.
Allen, P.C. and Fetterer, R.H., 2002. Recent advances in biology and immunobiology of Eimeria species and in diagnosis and control of infection with these coccidian parasites of poultry. Clinical Microbiology Reviews 15, 58-65.
Allen, P.C., Danforth, H.D. and Augustine, P.C., 1998. Dietary modulation of avian coccidiosis. International Journal for Parasitology 8, 1131-1140.
Allen, P.C., Lydon, J. and Danforth, H.D., 1997. Effects of components of Artemisia annua on coccidia infections in chickens. Poultry Science 76, 1156-1163.
Almeida, G.F.d. and Abreu. L.S.de., 2009. Estratégias produtivas e aplicação de princípios da agroecologia: o caso dos agricultores familiares da Cooperativa dos agropecuaristas solidários de Itápolis - COAGROSOL. ISSN 1981-4771. Revista de Economia Agrícola 56, 37-53.
Al-Sheikhly, F. and Al-Saieg, A., 1980. Role of Coccidia in the Occurrence of Necrotic Enteritis of Chickens. Avian Diseases 24 (2), 324-333.
Altieri, M.A. and Nicholls, C.I., 2005. Agroecology and the Search for a Truly Sustainable Agriculture. First Edition. In: P.N.U.M.A. Basic Textbooks for Environmental Training. Mexico. Available at: http://www.agroeco.org/doc/agroecology-engl-PNUMA.pdf. Access July 2012.
Anonymous, 1990. Diagnosis of coccidiosis in chickens. Janssen Pharmaceutica, Beerse, Belgium.
Anonymous, 2012a. Danish Poultry Meat Association. Available in Danish at: http://www.danskslagtefjerkrae.dk/Aktiviteter/Dansk_fjerkare_ude_og_hjemme.aspx. Accessed June 2012.
Anonymous. 2012b. Report from the Danish Poultry Council 2010. Copenhagen, Danish Poultry Council. Anonymous, 2012c. Denmark. The Ministry of Food, Agriculture and Fishery. Vejledning om økologisk Jordbrugsproduktion 178 pp (In Danish).
Anonymous, 2012d. Basic rules for the production of the label rouge chickens. Available at: http://www.poultrylabelrouge.com/014_differentes_volailles_poulet.php. Accessed June 2012.

66
Antell, S. and Ciszuk P., 2006. Forage consumption of laying hens - The crop content as an indicator of feed intake and AME content of ingested forage. Archive fur Geflugelkunde 70, 154-160.
Arab, H.A., Rahbari, S., Rassouli, A., Moslemi, M.H. and Khosravirad, F.D.A., 2006. Determination of artemisinin in Artemisia sieberi and anticoccidial effects of the plant extract in broiler chickens. Tropical Animal Health and Production 38, 497-503.
Ayssiwede, S.B., N'Dri, K.M., Gbati, O., and Missohou, A., 2011. Comparative study of different breeds of chickens' sensibility to avian coccidiosis. Reveu de Medecine Veterinaire 162 (3), 138-142.
Barry, T.N., Manley, T.R. and Duncan, S.J., 1986. The role of condensed tannin in the nutritional value of Lotus pedunculatus for sheep: Site of carbohydrate and protein digestion as influenced by dietary reactive tannin concentration. British Journal of Nutrition 55, 123-137.
Basser, A., Ciszuk, P. and Sjelin, K., 1999. Management of Laying hens in móbile houses – A review of experiences. In Ecological animal husbandry in the Nordic countries. Hermansen, J.E., Lund, V. and Thuen, E. (Eds). Proceedings of the NJF – seminar No.303. Horsens, Denmark 16-17 September, p. 45-50.
BRASIL, 2003. Federal Law n. 10.831. Organic agriculture and sets other provisions. Available in English at: http://www.prefiraorganicos.com.br/media/45792/lei_10.831_ingles.pdf. Accessed January 2012.
BRASIL, 2011. Federal Normative Instruction n. 46. Regulates the vegetable and animal production in organic systems. Available in Portuguese at: http://www.in.gov.br/visualiza/index.jsp?data=07/10/2011&jornal=1&pagina=4&totalArquivos=248 Accessed January 2012.
Castellini, C., Berri, C., Le Bihan-Duval, E. and Martino, G., 2008. Qualitative attributes and consumer perception of organic and fee-range poultry meat. World's Poultry Science Journal 64, 500 – 512.
Castellini, C., Dal Bosco, A., Mugnai, C. and Pedrazzoli, M., 2006. Comparison of two chicken genotypes organically reared: oxidative stability and other qualitative traits. Italian Journal of Animal Science 5, 29.
Caygill, J. C. and Mueller-Harvey, I. (Eds.), 1999. Secondary Plant Products - Antinutritional and beneficial actions in animal feeding. Nottingham University Press, Nottingham, UK, 129 pp.
Cervantes, H., 2006. Incidence of subclinical diseases and pathological conditions in clinically normal broilers from 3 production complexes sorted by sex and age. 143rd Annual Convention of the American Veterinary Medical Association and 50th Annual Meeting of the American Association of Avian Pathologists, July 15-19, Honolulu, Hawaii.

67
Cervantes, H., 2008. Coccidiosis Control. Available at: http://www.ces.ncsu.edu/depts/poulsci/conference_proceedings/supervisors_shortcourse/2008/cervantes_2008.pdf. Access July 2012.
Chapman, H.D., 2007. Rotation programmes for coccidiosis control. International Poultry production 15, 7-9.
Cheema, M.A., Qureshi, M.A. and Havenstein, G.B. 2003. A Comparison of the Immune Response of a 2001 Commercial Broiler with a 1957 Random bred Broiler Strain When Fed Representative 1957 and 2001 Broiler Diets. Poultry Science 82, 1519–1529.
Choi, K.D., H.S. Lillehoj, and D.S. Zarlenga., 1999. Changes in local IFN-gamma and TGF-beta4 mRNA expression and intraepithelial lymphocytes following Eimeria acervulina infection. Veterinary Immunology and Immunopathology 71, 263-275.
Christiansen, S.B., and Sandøe, P., 2000. Bioethics: limits to the interference with life. Animal Reproduction Science 60, 15–29
Coelho, A.A.D., Sanino, J.M. and Rosário, M.F. do, 2008. Frango Feliz: Caminhos para a avicultura alternativa. Piracicaba: FEALQ. 88p.
Collier, C.T., Hofacre, C.L., Payne, A.M., Anderson, D.B., Kaiser, P., Mackie, R.I. and Gaskins, H.R., 2008. Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth. Veterinary Immunology and Immunopathology 122, 104–115.
COM (2008). COM/2008/0233. Report from the Commission to the Council and the European Parliament on the use of coccidiostats and histomonostats as feed additives, submitted pursuant to article 11 of regulation (EC) no. 1831/2003 of the European Parliament and of the Council of 22 September, 2003 on additives for use in animal nutrition, http://www.ipex.eu/ipex/cms/home/Documents/dossier_COM20080233.
Combes, S., Lebas, F., Lebreton, L., Martin, T., Jehl, N., Cauquil, L., Darche, B. and Corboeuf, M.A., 2003. Comparison lapin « Bio »/ lapin standard: Caractéristique des carcasses et composition chimique de 6 muscles de la cuisse. Proc. 10èmes Journées de la Recherche Cunicole, Paris, 133–136.
Conney, A.H, Lysz, T., Ferraro, T., Abidi, T.F., Manchand, P.S., Laskin, J.D. and Huang, M.T., 1991. Inhibitory effect of curcumin and some related dietary compounds on tumor promotion and arachidonic metabolism in mouse skin. Advances in Enzyme Regulation 31, 385-396.
Cooper, K.K. and Songer, J.G., 2009. Necrotic enteritis in chickens: A paradigm of enteric infection by Clostridium perfringens type A. Anaerobe 15, 55-60.
Crabone, G.T., Moori, R.G. and Sato, G.S., 2005. The key factors for organic chicken purchase decision and the impact in the production chain. Organizacão Rural & Agroindustria 7 (3), 312-323.

68
Dalloul, R.A., and H.S. Lillehoj, 2005. Recent advances in immunomodulation and vaccination strategies against coccidiosis. Avian Diseases 49, 1-8.
Darnhofer, I., Lindenthal,T., Bartel-Kratochvil, R., Zollitsch, W., 2010. Conventionalisation of organic farming practices: from structural criteria towards an assessment based on organic principles. A review. Agronomy for Sustainable Development 30, 67–81.
Dawkins, M. S., P. A. Cook, M. J. Whittingham, K. A. Mansell, and A. E. Harper. 2003. What makes free-range broiler chickens range? In situ measurement of habitat preference. Animal Behaviour 66, 151-160.
De Gussem, M., 2007. Coccidiosis in poultry: Review on diagnosis, control, prevention and interaction with overall gut health. In: Proceedings of the 16th European Symposium on Poultry Nutrition. pg 253–261. World’s Poultry Science Association, Beekbergen, the Netherlands.
de Vries, M. and de Boer, I.J.M., 2010. Comparing environmental impacts for livestock products: A review of life cycle assessments Livestock Science 128 (1), 1-11.
Decuypere, E., Bruggeman, V., Everaert, N., Li, Y., Boonen, R., Tavernie, J. De, Janssens, S. and Buys, N., 2010. The Broiler Breeder Paradox: ethical, genetic and physiological perspectives, and suggestions for solutions. British Poultry Science 51 (5), 569-579.
Del Cacho, E., Gallego, M., Francesch, M., Quílez, J. and Sánchez-acedo, C., 2010. Effect of artemisinin on oocyst wall formation and sporulation during Eimeria tenella infection. Parasitology International 4, 506-511.
Delpech, P., Dumont, B. L. and Nefzaoui, A., 1983. Influence du rationnement et du patrimoine génétique de poulets sur les caractéristiques physico-chimiques et sensorielles de la viande à différents âges. Proceedings of the 6th European Symposium on the Quality of Poultry Meat, pp. 21-27.
Duncan, I.J.H., 2005. Science-based assessment of animal welfare: Farm animals. Revue Scientifique et Technique-Office International des Epizooties 24, 483–492.
Duncan, I.J.H., and Fraser, D., 1997. Understanding animal welfare. In M. Appleby & B. O. Hughes (Eds.). Animal welfare (pp. 19–31). Wallingford: CABI Publisher.
EU, 1991b. Commission regulation N.1538/91 of 15 June 1991 introducing detailed rules for implementing regulation 1906/90 on certain marketing standards for poultry. Official Journal of the European Communities (L143), 11-22.
EU, 1993. Commission Regulation (EEC) No 2891/93 of 21 October 1993 amending Regulation (EEC) No 1538/91 introducing detailed rules for implementing Regulation (EEC) No 1906/90 on certain marketing standards for poultry meat. Official Journal of the European Communities (L 263), 12-30.
EU, 2001. Commission Regulation (EC) No 436/2001 of 2 March 2001 amending Annex II to Council Regulation (EEC) No 2092/91 on organic production of agricultural products and

69
modifications referring thereto on agricultural products and foodstuffs. Official Journal of the European Communities (L 63), 16-18.
EU, 2007. Commission regulation N.834/07 of 28 June 2007 on organic production and labelling of organic products and repealing Regulation (EEC) No 2092/91. Official Journal of the European Communities (L189), 1-23.
EU. 1999a. Council Regulation 1804/1999 supplementing Regulation 2092/91 on organic production. Official Journal of the European Communities 42(L222), 1-28.
Fanatico, A., 2006. Organic poultry production in the United States. ATTRA - National Sustainable Agriculture Information Service. Available at: http://www.attra.org/attra-pub/PDF/coccidiosis.pdf. Accessed January 2012.
Fanatico, A.C., Pillai, P.B., Emmert, J.L. and Owens, C.M., 2007. Meat Quality of Slow- and Fast-Growing Chicken Genotypes Fed Low-Nutrient or Standard Diets and Raised Indoors or with Outdoor Access. Poultry Science 86, 2245–2255.
Fanatico, A.C., Owens, C.M. and Emmert J.L., 2009. Organic poultry production in the United States: Broilers. The Journal of Applied Poultry Research 18, 355–366.
FAOstat 2012a. Food and Agriculture Organization of the United Nations. Data on production; Livestock Primary (from 1970, 1990 and 2010). Available at: http://faostat.fao.org/site/569/DesktopDefault.aspx?PageID=569#ancor. Accessed June 2012.
FAOstat 2012b. Food and Agriculture Organization of the United Nations. Data on food supply; Livestock and Fish Primary Equivalent (from 2007). Available at: http://faostat.fao.org/site/610/default.aspx#ancor. Accessed June 2012.
Faria, P.B., Vieira, J.O., Souza, X.R., Rocha, M.F.M. and Pereira, A.A., 2012. Quality of broiler meat of the free-range type submitted to diets containing alternative feedstuffs/ Qualidade da carne de frangos de corte do tipo caipira submetidos a dietas contendo alimentos alternativos. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 64 (2), 389-396.
Fernandez, S., Pagotto, A.H., Furtado, M.M., Katsuyama, A.M., Madeira, A.M.B.N and Gruber, A., 2003. A multiplex PCR assay for the simultaneous detection and discrimination of the seven Eimeria species that infect domestic fowl. Parasitology 127, 317-325.
FRANCE, 1996. Relatif à la partie législative du livre VI (nouveau) du code rural. Art. L. 641-2. - Les produits agricoles ou alimentaires, bruts ou transformés, peuvent se voir reconnaître exclusivement une appellation d'origine contrôlée. Available at: http://www.senat.fr/leg/96-TA108.html. Accessed August 2012.
Francisco, D.C., Nascimento, V.P., Loguercio, A.P. and Camargo L., 2007. Caracterização do consumidor de carne de frango da cidade de Porto Alegre Ciência Rural 37 (1) Santa Maria jan./fev. 2007.

70
Gliessman, S.R., 2000. Agroecologia: processos ecológicos em agricultura sustentável. Porto Alegre: UFRGS. 170 p.
Gordon, S.H.B., Cottrill, M., Omiczek, T. Verhoeven, R. Weightman, R. Safford, F. Nicholson,
Guthman, J., 2004. The trouble with ‘organic lite’ in California: a rejoinder to the ‘conventionalisation’ debate. Sociologia Ruralis 44, 301–315.
Haug, A., Thebo, P., and Mattsson, J.G., 2007. A simplified protocol for molecular identification of Eimeria species in field samples. Veterinary Parasitology 1, 35-45.
Heckendorn, F., Haring, D.A., Amsler, Z. and Maurer, V., 2009. Do stocking rate and a simple run management practice influence the infection of laying hens with gastrointestinal helminths? Veterinary Parasitology 159, 60-68.
Hermansen, J.E., 2011. Livestock Production in Organic Systems in Denmark. In: International Workshop in Organic Agriculture. Curitiba, Brazil. August 17.th 2011.
Herrmann, R. and Roder, C., 1995. Does food consumption converge internationally?: measurement, empirical tests and determinants. European Review of Agricultural Economics 22, 400 - 414.
Hewson, C.J., 2003. What is animal welfare? Common definitions and their practical consequences. Canadian Veterinary Journal 44(6), 496–499.
Holling, C.S., 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4, 1-23.
Horsted, K. and Hermansen, J.E., 2007. Whole wheat versus mixed layer diet as supplementary feed to layers foraging a sequence of different forage crops. Animal 1, 575-585.
Horsted, K., 2006. PhD thesis Increased foraging in organic layers. The Royal Veterinary and Agricultural University & Department of Agroecology, Aarhus University. 141p.
Horsted, K., Hammershøj, M., and Hermansen, J.E., 2006. Short-term effects on productivity and egg quality in nutrient-restricted versus non-restricted organic layers with access to different forage crops. Acta Agriculturae Scandinavica 56, 42-54.
Horsted, K., Hermansen, J.E., and Ranvig, H., 2007. Crop content in nutrient-restricted versus non-restricted organic laying hens with access to different forage vegetations. British Poultry Science 48 (2), 177-184.
IFOAM, 2005. The principle aims of organic production and processing. Basic standards. http://www.ifoam.org/about_ifoam/standards/norms/norm_documents_library/Norms_ENG_V4_20090113.pdf . Accessed September 2010.
Jensen, G.H. and L.J. Korschgen, 1947. Contents of crops, gizzards, and droppings of bobwhite quail force-fed known kinds and quantities of seeds. Journal of Wildlife Management 11, 37-43.

71
Johnson J. and Reid W.M., 1970. Anticoccidial drugs lesion scoring techniques in battery and floor-pen experiments with chickens. Experimental Parasitology 28, 30-6.
Kearney, J. 2010. Food consumption trends and drivers. Philosophical Transactions of the Royal Society B 365, 2793–2807.
Kim, D.K., H.S. Lillehoj, Y.H. Hong, D.W. Park, S.J. Lamont, J.Y. Han, and E.P. Lillehoj, 2008. Immune-related gene expression in two B-complex disparate genetically inbred Fayoumi chicken lines following Eimeria maxima infection. Poultry Science 87, 433-443.
Kjaer, J.B. and Mench, J.A., 2003. Behaviour problems associated with selection for increased Production. Pages 67-82 in Poultry Genetics, Breeding and Biotechnology. Muir, W.M. and Aggrey, S.E. (Eds). Wallingford, CAB International.
Krystallis, A. de Barcellos, M.D. Grunert, K.G. Perrea, T. and Verbeke, W., 2012. Consumer Attitudes towards Sustainability Aspects of Food Production: Insights from Three Continents. Journal of Marketing Management 28 (3-4), 334-372.
Kvale, S. and Brinkmann, S., 2009. InterViews, Learning the Craft of Qualitative Research Interviewing. SAGE Publications.
Leeson, S., and Summers, J.D., 1997. Pages 207-254 in Commercial Poultry Nutrition. 2nd ed. University Books, Guelph, Ontario, Canada.
Leshchinsky, T.V. and Klasing, K.C., 2001. Relationship between the levels of dietary vitamin E and the immune response of broiler chickens. Poultry Science 80, 1590-1599.
Lewis, P.D., Perry, G.C., Farmer, L.J. and Patterson, R.L.S., 1997. Responses of two genotypes of chicken to the diets and stocking densities typical of UK and ‘Label Rouge’ production systems: I. Performance, behavior, and carcass composition. Meat Science 45, 501–516.
Lillehoj, H.S. and Lillehoj, E.P., 2000. Avian coccidiosis: a review of acquired intestinal immunity and vaccination strategies. Avian Diseases 44, 408–425.
Lillehoj, H.S., 1998. Role of T lymphocytes and cytokines in coccidiosis. International Journal for Parasitology 28, 1071-1081.
Lomu, M.A., Glatz, P.C. and Ru., Y.J., 2004. Metabolizable energy of crop contents in free-range hens. International Journal of Poultry Science 3, 728-732.
Long P.L. and Rowell, J.G., 1975. Sampling broiler house litter for coccidial oocysts. British Poultry Science 16, 583-592.
Lopez-Bote, C.J., Gray, J.I., Gomaa, E.A. and Flegal, C.J., 1998. Effect of dietary administration of oil extracts from rosemary and sage on lipid oxidation in broiler meat. British Poultry Science 39, 235-240.

72
Luchese, F.C., Perin, M., Aita, R.S., Mottin, V.D., Molento, M.B., and Monteiro, S.G., 2007. Prevalência de espécies de Eimeria em frangos de criação industrial e alternativa. Brazilian Journal of Veterinary Research and Animal Science 44, 81-86.
MAFF, 2001. A study tour of organic poultry production in France – examining the current and future implementation of standards and EU legislation. Maff project code OF 0303. Available at: http://orgprints.org/6612/1/OF0303_2551_FRP.pdf. Accessed July 2012.
Mannan, A., Ahmed, I., Arshad, W., Asim, M.F., Qureshi, R.A., Hussain, I. and Mirza, B., 2010. Survey of artemisinin production by diverse Artemisia species in northern Pakistan. Malaria Journal 9, 310. http://dx.doi.org/10.1186/1475-2875-9-310.
Marie-Magdeleine, C., Mahieu, M., Philibert, L., Despois, P. and Archimède, H., 2010. Effect of cassava (Manihot esculenta) foliage on nutrition, parasite infection and growth of lambs. Small Ruminant Research 93, 10-18.
Martínez Michel L., Anders, S. and Wismer W.V., 2011. Consumer Preferences and Willingness to Pay for Value-Added Chicken Product Attributes. Journal of Food Science 76 (8) S469 –S477.
Mathis, G.F., 2005. Reasons for field problems with E. maxima: E. acervulina vs E. maxima. Proceeding of the IXth International coccidiosis conference. Foz do Iguacú, Brazil. 19-23 Sep 2005.
McDougald, L.R., Fuller, L. and Mattiello, R. 1997. A Survey of Coccidia on 43 Poultry Farms in Argentina. Avian Diseases 41, 923-929.
Min, W., H.S. Lillehoj, C.M. Ashwell, C.P. van Tassell, R.A. Dalloul, L.K. Matukumalli, J.Y. Han, and E.P. Lillehoj. 2005. Expressed sequence tag analysis of Eimeria-stimulated intestinal intraepithelial lymphocytes in chickens. Molecular Biotechnology 30, 143–150.
Mongin, P., 1976. Composition of crop and gizzard contents in the laying hen. British Poultry Science 17, 499-507.
Morris, G.M., Woods, W.G., Richards, D.G. and Gasser, R.B., 2007. Investigating a persistent coccidiosis problem on a commercial broiler–breeder farm utilizing PCR-coupled capillary electrophoresis. Parasitology Research 101, 583-589.
Mwalusanya, N.A., Katule, A.M., Mutayoba, S.K., Minga, U.M., Mtambo, M.M.A. and Olsen, J.E., 2002. Nutrient status of crop contents of rural scavenging local chickens in Tanzania. British Poultry Science 43, 64-69.
Naidoo, V., McGaw, L.J., Bisschop, S.P.R., Duncan, N. and Eloff, J.N., 2008. The value of plant extracts with antioxidant activity in attenuating coccidiosis in broiler chickens. Veterinary Parasitology 153, 214-219.
Netpana, N., Wanapat, M., Poungchompu, O. and Toburan, W., 2001. Effect of condensed tannins in cassava hay on fecal parasitic egg counts in swamp buffaloes and cattle. In: Preston, T.R., Ogle, B., Wanapat, M. (Ed.), Proceedings of the International Workshop on Current

73
Research and Development on Use of Cassava as Animal Feed, Khon Kaen University, Thailand. Available at: http://www.mekarn.org/procKK/wana1.htm. Accessed November 2011.
Neufeld, L., 2002. Consumer preferences for organic/free range chicken. Agriculture Marketing Service, Kansas State University. Available at: http://www.agmrc.org/media/cms/ksufreerangech_588C29A8ED362.pdf. Accessed June 2012.
Nielsen, B.L., Thomsen, M.G., Sorensen, P. and Young, J.F., 2003. Feed and strain effects on the use of outdoor areas by broilers. British Poultry Science 44, 161-169.
Oelofse, M., Høgh-Jensen, H., Abreu, L.S. de, Almeida, G.F. d, El-Araby, A., Hui, Q.Y., Sultan, T. Neergaard, A., 2011. Organic farm conventionalization and farmer practices in China, Brazil and Egypt. Agronomy for Sustainable Development 31, 689-698.
Olesen, I., Groen, A.F. and Gjerde B., 2000. Definition of animal breeding goals for sustainable production systems. Journal of Animal Science 78, 570-582.
Olliaro, P.L. and Taylor, W.R.J., 2004. Developing artemisinin-based drug combinations for the treatment of drug-resistant falciparum malaria: a review. Journal of Postgraduate Medicine 50, 40-44.
Orengo, J., Buendía, A.J., Ruiz-Ibáñez, M.R., Madrid, J., Del Río, L., Catalá-Gregori, P., García, V. and Hernández, F., 2012. Evaluating the efficacy of cinnamaldehyde and Echinacea purpurea plant extract in broilers against Eimeria acervulina. Veterinary Parasitology 185, 158-163.
Owens, C. M., Fanatico, A. C., Pillai, P. B., Meullenet, J. F. and Emmert, J. L., 2006. Evaluation of alternative genotypes and production systems for natural and organic poultry markets in the U.S. In: Proceedings XII European Poultry Conference, Verona.
Pedersen, M.A., Thamsborg, S.M., Fisker, C., Ranvig, H. and Christensen, J.P., 2003. New Production Systems: Evaluation of Organic Broiler Production in Denmark. Journal of Applied Poultry Research 12, 493-508.
Permin, A. and Hansen J.W., 1998. The epidemiology, diagnosis and control of poultry parasites. A FAO Handbook, FAO, Rome, Italy.
Permin, A., 1997. Helminths and helminthosis in poultry with special emphasis on Ascaridia galli in chickens. PhD thesis. The Royal Veterinary and Agricultural University, Copenhagen, Denmark.119 p.
Permin, A., Bisgaard, M., Frandsen, F., Pearman, M., Kold, J. and Nansen, P., 1999. The prevalence of gastrointestinal helminths in different poultry production systems. British Poultry Science 40, 439-443.
Pinard-van der Laan, M.H., Monvoisin, J.L., Pery, P., Hamet, N. and Thomas, M., 1998. Comparison of Outbred Lines of Chickens for Resistance to Experimental Infection with Coccidiosis (Eimeria tenella). Poultry Science 77, 185–191.

74
Pokarzhevskii, A.D., Zaboyev, D.P., Ganin, G.N. and Gordienko, S.A., 1997. Amino acids in earthworms: are earthworms ecosystemivorous? Soil Biology and Biochemistry 29, 559-567.
Prothero, R.E. 1936. English farming past and present. Heinemann and Frank Cass and Co. London, 1961 ed. First published in 1912.
Rauw, W.M., Kanis, E., Noordhuizen-Stassen, E.N. and Grommers, F.J. 1998. Undesirable side effects of selection for high production efficiency in farm animals: a review. Livestock Production Science 56, 15-33.
Ravindran, V., 1991. Preparation of cassava leaf products and their use as animal feed. In: Roots, tubers, plantains and bananas in animal feeding. APHP Paper 95: 81-98, FAO: Rome. Available at:http://www.fao.org/WAICENT/FAOINFO/AGRICULT/AGA/AGAP/FRG/AHPP95/95111.pdf. Accessed February 2012.
Reddy, R.C., Vathsala, P.G., Keshamouni, V.G., Padmanaban, G. and Rangarajan, P.N., 2005. Curcumin for malaria therapy. Biochemical and Biophysical Research Communications 326, 472–474.
Robinson, L. 1961. Modern Poultry Husbandry. Crosby Lockwood. London.
Rodriguez, J.C., Segura, J.C., Alzina, A. and Guterrez, M.A., 1997. Factors affecting mortality of crossbred and exotic chicken kept under backyard systems in Yucatan, Mexico. Tropical Animal Health and Production 29, 151-157.
Roepstorff, A. and Nansen, P., 1997. The epidemiology, diagnosis and control of helminth parasites of swine. A FAO Handbook, FAO, Rome, Italy.
Rose, M.E. and Hesketh, P., 1976. Immunity to coccidiosis: stages of the life-cycle of Eimeria maxima which induce, and are affected by, the response of the host. Parasitology 73, 25–57.
Rose, M.E., Hesketh, P. and Wakelin, D., 1997. Oral vaccination against coccidiosis: responses in strains of mice that differ in susceptibility to infection with Eimeria vermiformis. Infection and Immunity 65, 1808–1813.
Ross, E. and Enriquez, F.Q., 1969. The nutritive value of cassava leaf meal. Poultry Science 48, 846-853.
Rosset, P. and Altieri, M.A., 1995. Agroecology and the conversion of large-scale conventional systems to sustainable management. International Journal of environmental studies 50 (3/4), 165-185.
S. Holmes, T. Turner, and H.A. van de Weerd., 2006. Organic egg production – a desk study on sustainable and innovative methods for meeting the hen's protein requirements. ADAS report on Defra project OF0357. Web Defra http://randd.defra.gov.uk/Default.aspx?Menu=Menu&Module=More&Location=None&ProjectID=13671&FromSearch=Y&Publisher=1&SearchText=OF0357&SortString=ProjectCode&SortOrder=Asc&Paging=10#Description. Accessed February 2011.

75
Sandøe, P. and Christiansen, S. B., 2008. Ethics of Animal Use, Blackwell ed.
Seng, S. and Rodriguez, L., 2001. Foliage from cassava, Flemingia macrophylla and bananas compared with grasses as forage sources for goats: effects on growth rate and intestinal nematodes. Livestock Research for Rural Development 13, 1-6.
Shirley, M.W., Smith, A.L. and Tomley, F.M., 2005. The biology of avian Eimeria with an emphasis on their control by vaccination. Advanced Parasitology 60, 285–330.
Smith, D.P., Northcutt, J.K. and Steinberg, E.L., 2012. Meat quality and sensory attributes of a conventional and a Label Rouge-type broiler strain obtained at retail. Poultry Science 91 (6), 1489-1495.
Sokerya, S., Waller, P.J., Ledin, I. and Höglund, J., 2007. The effects of short-term feeding of fresh cassava (Manihot esculenta) foliage on gastrointestinal nematode parasite infections in goats in Cambodia. Tropical Biomedicine 24, 47-54.
Sonaiya, E.B., 2004. Direct assessment of nutrient resources in free-range and scavenging systems. World’s Poultry Science Journal 60, 523-535.
Sørensen, P., 2010. Chicken genetic resources used in smallholder production systems and opportunities for their development, by FAO. Smallholder Poultry Production Paper No. 5. Rome.
Sossidou, E.N., Dal Bosco, A., Elson, H.A. and Fontes, C., 2010. Pasture-based systems for poultry production: Implications and perspectives. World’s Poultry Science Journal 67, 47–58.
Star, L., Ellen, D., Uitdehaag, K. and Brom, F.W.A., 2008. A plea to implement robustness into a breeding goal: poultry as an example. Journal of Agricultural and Environmental Ethics 21, 109–125.
Stearns, S.C., 1976. Life-history tactics: A review of the ideas. The Quarterly Review of Biology 51, 3-47.
Swinkels, W.J., Post, J., Cornelissen, J.B., Engel, B., Boersma, W.J. and Rebel, J.M., 2006. Immune responses in Eimeria acervulina infected one-day-old broilers compared to amount of Eimeria in the duodenum, measured by real-time PCR. Veterinary Parasitology 138, 223–233.
Swinkels, W.J., Post, J., Cornelissen, J.B., Engel, B., Boersma, W.J. and Rebel, J.M., 2007. Immune responses to an Eimeria acervulina infection in different broilers lines. Veterinary Immunology and Immunopathology 117, 26–34.
Temraz, A. and El-Tantawy, W.H., 2008. Characterization of antioxidant activity of extract from Artemisia vulgaris. Pakistan journal of pharmaceutical sciences 21, 321-326.
Thamsborg, S.M., 2002. Organic Farming in the Nordic Countries - Animal Health and Production. Acta veterinaria Scandinavica. Supplementum 43, 7-15.

76
Thebo, P., Lunden, A., Uggla, A. and Hooshmand-Rad, P., 1998. Identification of seven Eimeria species in Swedish domestic fowl. Avian Pathology 6, 613-617.
Thorp, B.H. and Luiting, E., 2000. Breeding for resistance to production disease in poultry. In R.F.E. Axford, S.C. Bishop, F.W. Nicholas & J.B. Owen, eds. Breeding for disease resistance in farm animals, pp. 357–378. Wallingford UK, CABI Publishing.
Tigno, X.T. and Gumila, E., 2000. In vivo microvascular actions of Artemisia vulgaris L. in a model of ischemia-reperfusion injury in the rat intestinal mesentery. Clinical hemorheology and microcirculation 23, 159-165.
Tigno, X.T., De Guzman, F. and Flora, A.M., 2000. Phytochemical analysis and hemodynamic actions of Artemisia vulgaris L. Clinical hemorheology and microcirculation 23, 167-175.
Tixer-Boichard, M., Leenstra, F., Flock, D.K., Hocking P.M. and Weigend, S., 2012. A century of poultry genetics. World's Poultry Science Journal 68, 307-321.
Torjusen, H., Sangstad, L., O’Doherty Jensen, K. and Kjærnes, U., 2004. European consumers’ conceptions of organic food: a review of available research. Professional Report No. 4, National Institute for Consumer Research, Oslo.
USDA 2000. United States Department of Agriculture – NOP - National Organic Program (NOP). Agricultural Marketing Service (AMS), U.S. Department of Agriculture. Available at: http://www.ams.usda.gov/AMSv1.0/ams.fetchTemplateData.do?template=TemplateA&navID=NationalOrganicProgram&leftNav=NationalOrganicProgram&page=NOPNationalOrganicProgramHome&acct=nop. Accessed June 2012.
USDA, 2012. United States Department of Agriculture. Food Service and inspection service. Fact sheets. Food labeling. Available at: http://www.fsis.usda.gov/Factsheets/Meat_&_Poultry_Labeling_Terms/index.asp. Accessed June 2012.
USDA-ARS, 2005. United States Department of Agriculture. Agricultural Research Service. In: Animal parasitic diseases: coccidiosis. Available at: http://www.ars.usda.gov/Main/docs.htm?docid=11018#part2. Accessed November 2009.
Van de Weerd, H.A., Keatinge R. and Roderick S., 2009. A review of key health-related welfare issues in organic poultry production. World's Poultry Science Journal 65, 649-684.
Vanhonacker, F., Verbeke, W., Van Poucke, E, Pieniak, Z., Nijs, G. and Tuyttens, F., 2012. The Concept of Farm Animal Welfare: Citizen Perceptions and Stakeholder Opinion in Flanders, Belgium. Journal of Agricultural and Environmental Ethics 25, 79–101.
Velkers, F.C., Swinkels, W.J.C., Rebel, J.M.J., Bouma, A., Daemen, A.J.J.M., Klinkenberg, D., Boersma, W.J.A., Stegeman, J.A., de Jong, M.C.M. and Heesterbeek, J.A.P., 2010. Effect of Eimeria acervulina infection history on the immune response and transmission in broilers. Veterinary Parasitology 173, 184-192.

77
Velkers, F. C., 2011. Transmission dynamics of Eimeria acervulina in broilers. PhD Thesis. Wageningen University, Wageningen, the Netherlands. 156 p.
Vermeir, I., and Verbeke, W., 2006. Sustainable food consumption: Exploring the consumer ‘‘Attitude— behavioral intention’’ gap. Journal of Agricultural and Environmental Ethics 19, 169–194.
Walker, A and Gordon, S., 2003. Intake of nutrients from pasture by poultry. Proceedings of the Nutrition Society 62, 253–256.
Wallach, M., 2010. Role of antibody in immunity and control of chicken coccidiosis. Trends in Parasitology 26 (8), 382–387.
Weeks, C.A., Nicol, C.J., Sherwin, C.M. and Kestin, S.C., 1994. Comparison of the behaviour of broiler chicken in indoor and free-range environments. Animal Welfare 3, 179-192.
Weinberg, E.D. and Moon, J., 2009. Malaria and iron: history and review. Drug Metabolism Reviews 41, 644-662.
Wier, M., Jensen, K.O., Andersen, L.M. and Millock, K., 2008. The character of demand in mature organic food markets: Great Britain and Denmark compared. Food Policy 33, 406–421.
Willcox, M., Falquet, J., Ferreira, J.F.S., Gilbert, B., HSU, E., Magalhães, P.M. de, Plaizer-Vercammen, J. and Wright, C.W., 2007. Artemisia annua as a herbal tea for malaria. African Journal of Traditional, Complementary and Alternative Medicines 4, 121-123.
Williams, R.B., 1995. Epidemiological studies of coccidiosis in the domesticated fowl (Gallus gallus). II: Physical condition and survival of Eimeria acervulina oocysts in poultry‐house litter. Applied Parasitology 36, 90-96.
Williams, R.B., 1999. A compartmentalized model for the estimation of the cost of coccidiosis to the world’s chicken production industry. International Journal for Parasitology 8, 1209-1229.
Williams, R.B., 2002. Fifty years of anticoccidial vaccines for poultry, 1952-2002. Avian Diseases 46, 755-802.

78
SUPPORTING PAPERS

79
Paper I
Use of Artemisia annua as a natural coccidiostat in free-range broilers and its
effects on infection dynamics and performance. Veterinary Parasitology, 186
(2012) 178– 187.
Almeida, G.F.d.; Horsted, K.; Thamsborg, S.M.; Kyvsgaard N.C.; Ferreira J.F.S.; Hermansen, J.E.
Reprinted with permission of Elsevier

Ua
GJa
Db
c
d
a
ARR1A
KANFBSC
g
0d
Veterinary Parasitology 186 (2012) 178– 187
Contents lists available at SciVerse ScienceDirect
Veterinary Parasitology
jou rn al h om epa ge: www.elsev ier .com/ locate /vetpar
se of Artemisia annua as a natural coccidiostat in free-range broilersnd its effects on infection dynamics and performance
ustavo F. de Almeidaa,∗, Klaus Horsteda, Stig M. Thamsborgb, Niels C. Kyvsgaardc,orge F.S. Ferreirad, John E. Hermansena
Department of Agroecology, Faculty of Sciences and Technology, Aarhus University, Research Centre Foulum, Blichers Allé, 20, P.O. Box 50, DK-8830 Tjele,enmarkDanish Centre for Experimental Parasitology, Faculty of Life Sciences, University of Copenhagen, Dyrlægevej 100, DK-1870 Frederiksberg C, DenmarkDepartment of Veterinary Disease Biology, Faculty of Life Sciences, University of Copenhagen, Stigbojlen 4, DK-1870 Frederiksberg CDenmarkUSDA-ARS, Appalachian Farming Systems Research Center, 1224 Airport Rd., Beaver, WV 25813, USA
r t i c l e i n f o
rticle history:eceived 24 May 2011eceived in revised form4 November 2011ccepted 16 November 2011
eywords:rtemisia annua L.atural anti-protozoa drugree-range systemsroiler Genotypesex effectsoccidiosis
a b s t r a c t
This work investigated the preventive effect of Artemisia annua L. dried leaves supplied asa botanical coccidiostat to two broiler genotypes reared in a Danish free-range system in afactorial experiment (two genotypes and ± supplement of dried A. annua leaves). The geno-types White Bresse L40, a pure slow-growing line, and Kosmos 8 Ross, a hybrid genotypewith medium growing characteristics, were used. Broilers were raised indoor until 29-days-old and kept free of parasites. Twelve groups of 30 randomly selected broilers were placedin the range forming three replicates for each treatment combination. The paddocks werecultivated with a mix of grass and clover. A separate group of broilers was naturally infectedwith Eimeria spp. oocysts and five animals nominated as “seeders” were introduced to theabove mentioned 12 groups, 10 days after its formation, with each group consisting of 35animals per plot. This infection strategy was meant to imitate the transmission pathwayobserved at farm level. Ten individual birds from each of the 12 groups, in total 120 animalsof mixed sex, were monitored twice weekly for 30 days for oocysts excretion. PCR of pooledfaecal samples, oocyst morphology and localization upon necropsy were used to identifythe Eimeria species involved in the infection. In general, broilers from both genotypes inthe range coped well with a coccidia infection caused by Eimeria acervulina and Eimeriamaxima as no clinical symptoms, or deaths, were reported during the experiment. In gen-eral, broilers supplemented with A. annua dried leaves showed a significantly (p < 0.05)reduced number of excreted oocysts during the infection with no interaction to genotype.Females generally had a significantly higher shedding of oocysts than males (p < 0.05). Theoverall body weight gain and the daily weight gain when infection was subdued showed athree-way interaction among genotype, sex and treatment – accounted mainly for the fact
that Kosmos females responded positively to the Artemisia treatment while Kosmos malesresponded negatively, and only minor differences were found between sexes for the WhiteBresse genotype. In conclusion, supply of A. annua dried leaves as a botanical coccidiostatsignificantly reduced oocyst output in free ranged broilers and thus may form part of a strategy to prevent comm∗ Corresponding author. Tel.: +45 8715 4757; fax: +45 8715 4798.E-mail addresses: [email protected],
[email protected] (G.F.d. Almeida).
304-4017/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.vetpar.2011.11.058
ercial losses.© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Organic principles (IFOAM, 2005) encourage farmers tointegrate livestock in agro-ecosystems to improve nutri-tional balance, reduce farmers’ dependence on market and

ry Para
G.F.d. Almeida et al. / Veterinastimulate agro-ecological biodiversity. Organic farming isregulated by legislative rules (EU, 2007) and organic poul-try production overemphasizes animal welfare (EU, 1991).Animals with appropriate genetic background must be ableto explore their natural behaviour by having access tooutdoor areas and be able to perform the physiologicalactivities related to their species during most part of theirlife (Jensen and Toates, 1993; Vestergaard, 1996; Duncan,2002). As a consequence of access to permanent paddocks,animals will be more exposed to parasites (Permin andHansen, 1998; Permin et al., 1999; Thamsborg, 2002).
Coccidiosis is a parasitic disease caused by the proto-zoa Eimeria spp. that can cause severe losses in poultryproduction (Williams, 1999). Parasites multiply in theintestines damaging gut tissues which, in turn, reducesfeed intake and the absorption of nutrients (Morris et al.,2007). Animals are also more susceptible to secondary bac-terial infections (Cooper and Songer, 2009) and these maypotentially be zoonotic (Al-Sheikhly and Al-Saieg, 1980).Rodriguez et al. (1997) in Mexico, and Luchese et al. (2007)in Brazil, referred to coccidiosis as the most importantcause of chicken mortality also in smallholder systems.However, in low-density production or with the use ofpreventive medication, coccidiosis generally remains asubclinical disease that only affects performance (Fanatico,2006).
In organic farming and other low-input systems, wheremedical prevention of coccidiosis is either prohibited or notaffordable, control through locally available plants, rootsor crops offers an attractive alternative. Recently, the herbArtemisia annua L. has received much attention as an alter-native treatment for drug-resistant Plasmodium falciparummalaria (Willcox et al., 2007) and the commercial use ofartemisinin, the main active ingredient extracted from theplant, is now widespread worldwide as a combination ther-apy (Olliaro and Taylor, 2004; Weinberg and Moon, 2009).
Artemisia annua has also been considered as a poten-tial candidate to control other apicomplexan parasites, e.g.coccidia infections in chicken (Allen and Fetterer, 2002;Arab et al., 2006). Allen et al. (1997) reported preventiveeffects of dried A. annua leaves, in caged fast-growing malebroilers, against individual and dual infection with Eime-ria tenella and E. acervulina. The leaves were added to thediet, at maximum dosages of 4–5 mg artemisinin per kgbody weight per day, and the birds were not subjectedto re-infection. These authors reported that a prophylacticstrategy by a three-week supplementation period prior tothe challenge was required to cause a reduction in oocystoutput of E. acervulina and increased performance. Inter-estingly, the authors reported no significant effects whenA. annua dried leaves, or pure artemisinin, were providedpreventively against E. maxima, suggesting that plant com-ponents including artemisinin may have a species-specificmode of action.
Arab and co-workers (2006) in another confinementtrial reported a positive effect of pure artemisinin, suppliedorally, as a curative strategy, one day after the inocula-
tion of the infective oocysts. Reduction on E. acervulina andE. tenella oocyst output as well as increased animal per-formance at doses of 1 and 2.5 mg/kg body weight werereported when pure artemisinin was supplied three times asitology 186 (2012) 178– 187 179
day, during five days, to fast-growth broilers not subjectedto re-infection. However, the same authors reported nosignificant effects of pure artemisinin, supplied curatively,against E. maxima. Brisibe et al. (2008) found coccidioci-dal activity and improved animal performance, similar to acommercial coccidiostat, when dried A.annua leaves wereadded at 10 and 20% of the daily diet of caged fast-growingmale broilers continuously challenged with E. tenella.
Rodriguez et al. (1997) and Luchese et al. (2007) bothreported that indigenous genotypes, commonly reared insmall-scale farms, could cope with Eimeria spp. infectionsbetter than hybrid/fast-growing genotypes, as measuredby lower mortality rates. Others have also documented therole of genetics in relation to responses to Eimeria spp.infections (Bumstead and Millard, 1992; Lillehoj and Trout,1996).
On this background, we hypothesized that the slowgrowing genotype chosen for our trial would cope betterwith Eimeria spp. infections in a natural organic environ-ment and that dried leaves of A. annua could also providepositive effects in prevention of coccidiosis. Therefore, theaim of the present work was to investigate the effect ofartemisinin-rich A. annua supplementation in two differ-ent genotypes (slow- and moderate-growing) in relationto infection rates, oocyst excretion dynamics and subse-quent impact on live weight gain after a challenge withcoccidia species under conditions closely resembling com-mon farming practices.
2. Materials and methods
2.1. Experimental design and genotypes
The experiment was conducted at the Research Cen-tre Foulum, Aarhus University, Denmark (56◦48′N, 9◦58′E).The study used a 2 × 2 factorial design – with two geno-types and two dietary treatments (supplemented with A.annua dried leaves or not) as main factors – and with threereplicates for each treatment combination. In total, theexperiment thus included 12 flocks of 35 broilers of mixedsex. Each group of 35 birds was allocated to a specific plot of105 m2 (12 m × 8.75 m), yielding a maximum stocking rateof 3 m2 of outside area per animal, and a mobile chickenhouse (4.6 m2) with cover and perches in compliance withthe European regulation for organic systems. Each plotincluded two feed silos (commercial diet and whole wheat)located inside the houses and a 25 L water tank locatedone meter outside the houses which all were refilled twiceweekly. The two genotypes in question were White BresseL40 from the Research Center Foulum, a pure slow growingline, and Kosmos 8 Ross, a hybrid genotype with dual pur-pose and medium growing characteristics purchased froman Italian breeding company (http://www.olandia.it).
During the study, animals were fed a typical organicconcentrate for broilers in a restriction scheme, whilewhole wheat and water was supplied ad libitum. Chickswere hatched on July 6th 2010 and transferred to a cleaned
and disinfected stable. The experiment consisted of 420chicks, half from each genotype and it began when birdswere 29-days-old. Broilers were allocated in paddocks pre-viously cultivated with a mixture of grass (Lolium perenne)
1 ry Para
atiD
2o
2sisv1wataneftmRbp
2i
dectTsaimoEmats(
2a
sweai“
80 G.F.d. Almeida et al. / Veterina
nd clover (Trifolium repens) not used for poultry produc-ion in the previous two years. The experimental protocolsnvolving broilers in our study were in accordance with theanish legislation regarding animal care and health.
.2. Indoor pre-experimental rearing period (1–29 daysf age)
All the one-day-old chicks were raised until they were9-days-old on concrete floor with wood shavings in fiveeparate brooder pens (4.5 m × 2.1 m). All bird groups dur-ng the indoor period were maintained on an organictarter diet for broilers on tray feeders, and were pro-ided with water ad libitum. Four groups indoor (with00 animals each, half from each genotype) were treatedith toltrazuril (Baycox®, Bayer Animal Health Ltd., DK)
t recommended dose preventively to avoid early infec-ion with Eimeria spp. The fifth pen, allocated with 50nimals of each genotype (designated as “seeders”) didot received toltrazuril in the drinking water and it wasxpected that these birds would acquire Eimeria infectionsrom the environment due to lack of resistance and oppor-unism of the parasite (Sharman et al., 2010). A weekly
onitoring system of oocyst excretion adapted fromoepstorff and Nansen (1997) was used to detect infection,ut the “seeders” excreted no oocysts during the indooreriod.
.3. Outdoor adaptation rearing period and naturalnfection of the “seeders” (29–39 days of age)
Broilers were randomly allocated to the 12 plots out-oor when they were 29-days-old. Six groups, three ofach genotype, were fed Artemisia mixed with a starteroncentrate as a dough type, made available in movableray feeders to allow broilers to adapt to the new diet.he non-treated groups received a dough type with theame amounts of starter concentrate but without A. annuadded. In the same 10 days period, the “seeders” remainedndoors but their bedding material was contaminated with
anure (20 kg scattered homogeneously on the ground)btained from a backyard farm and found positive forimeria spp. oocysts (mean ∼48.000 oocysts/g of beddingaterial from three replicated samples). After the 10 days
daptation period, broilers in the range were consuminghe diet with Artemisia added and the “seeders” began tohed oocysts, ranging from 40 to 560 oocysts/g of faecesOPG).
.4. Outdoor experimental rearing period (39–74 days ofge)
Five “seeders” of each genotype were randomlyelected, caged for counting of oocysts, and then mixedith the outdoor groups. The dough diet was adjusted to
nsure the required amount of artemisinin recommendeds a prophylactic treatment (Allen et al., 1997). The exper-ment was terminated four weeks after introduction ofseeders” to the paddocks.
sitology 186 (2012) 178– 187
2.5. Feeding strategy during the outdoor rearing period(29–74 days of age)
The dough with Artemisia mixed was supplied to thetreated groups every morning around 8:00 am from thefirst day outdoor to the end of the experiment. The dailyamount of concentrate was restricted from 6:00 pm untilthe following morning after the supply of the dough. Thedough was prepared daily by mixing a portion of pow-dered A. annua dried leaves with a specific amount of starterdiet (10 g/head) in the presence of water. When the doughwas introduced to the birds during the adaptation period,A. annua powder was added to the mixture at 5% of theexpected daily food intake (calculated twice per week afterre-filling the feed silos). However, 5% was deemed too highto ensure a rapid intake of the dough. Therefore, during the10 days adaptation period, the percent of Artemisia driedleaves in the dough diet was gradually reduced to 3% of theestimated daily food intake. This percentage guaranteeda minimum consumption of artemisinin above 4 mg/kgbroiler.
Daily, after feed was available to the broilers for threehours, all remaining dough from each plot was collectedat 11:00 am, transported indoors and dried in the shadein a ventilated area at room temperature (20 ◦C) duringthe following three days. From 11:00 am to 6:00 pm allbirds had access to growth concentrate ad libitum. Whenthe remaining dough was dried, it was carefully collected,stored in plastic bags, weighed and the amount of A. annuaconsumed daily was estimated.
2.6. Plant material and determination of artemisinincontent
The A. annua L. (var. Artemis, Mediplant) used in thefeed was an improved variety for high artemisinin content,kindly supplied by the Department of Horticulture, Facultyof Science and Technology, Aarhus University. Plants weregrown on a sandy loam soil at Aarslev Research Centre inDenmark (55◦18′N, 10◦27′E) and harvested in the autumnof 2009 just before flowering. The whole plant was driedin a ventilated room at approximate 20 ◦C and stored inplastic bags in a controlled environment for one year. Twoweeks before the trial, leaves were separated by hand frommain and secondary stems. Leaves were ground by a ham-mer mill and sifted through a 3-mm mesh and the powderstored in 10-L buckets. Artemisinin, dihydroartemisinicacid, and artemisinic acid were extracted from three sep-arate 500-mg samples of powdered leaves, mixed withdiatomaceous earth using a mortar and pestle, transferredto 10-mL stainless steel cells, and extracted with an accel-erated solvent extraction system (ASE 350, Dionex, USA)using petroleum ether as the solvent at a temperature of100 ◦C, pressurized to 1500 psi with nitrogen, for a statictime of 5 min. The petroleum ether extract was transferred
to a fume hood, evaporated to dryness, and reconstitutedin 20 mL of acetonitrile (two washes of 10 mL each). Afterfiltration through a 0.45 �m nylon filter, artemisinin, dihy-droartemisinic acid, and artemisinic acid were quantified
ry Para
G.F.d. Almeida et al. / Veterinaby HPLC–UV, based on standard curves generated with purestandards, according to Ferreira and Gonzales (2009).
2.7. Monitoring the infection dynamics
A total of 120 broilers, 10 from each group, were ran-domly selected and tagged in the same day when broilerswere introduced to the outdoor paddocks. The ten birds perplot were caged individually twice weekly (Monday andThursday), and after 20 min individual droppings were col-lected and stored. When broilers achieved 55 days of age,sex differentiation was performed visually (64 males and56 females of tagged animals). In the laboratory, faecessamples were examined for coccidia oocysts by a modi-fied McMaster-technique using saturated sodium chloridesolution with 50% glucose monohydrate, as flotation fluid(Roepstorff and Nansen, 1997). Faeces were diluted andwell homogenized in tap water depending on sampleweight, proportional to 4 g of faeces in 56-mL of tap water.After 30 min standing and re suspension, the solutionwas filtrated through gauze (aperture 250 �m) to removedebris.
Graded centrifuge tubes were identified and filled to thelimit of 10-mL and centrifuged (1200 rpm) for 7 min. Thesupernatant was removed and the pellet containing oocystswas stored in the fridge. Next day, tubes were left for 30 minat room temperature and the flotation fluid was added to 4-mL. A disposable pipette was used to mix flotation solutionwith the pellet and quickly transfer the homogenized mix-ture to the McMaster slide. After 5 min standing to allowoocysts to float, oocysts were counted at 100× amplifica-tion using a microscope. For each oocyst visually identifiedin each of the twelve rows scanned on the McMaster slidecorresponded to 20 oocysts per gram of faeces (OPG).
The species of Eimeria present in the study were deter-mined by different means. Five “seeders” and five “nonseeders” from each genotype (total n = 20) were examinedpost mortem the day after the introduction of “seeders” tothe paddocks, when animals were 40-days-old. In addition,at the end of study, when animals were 74-days-old, fivebirds from each group were necropsied (n = 60) and scoredfor lesions. The location and number of intestinal lesionson the serosal surface were noted for three species of Eime-ria commonly found in Danish poultry systems: E. maxima,E. acervulina and E. tenella. Lesion counts are presented asgroup means. Then, the intestines were opened and lesionsin the mucosa were scored (0–4) according to Johnsonand Reid (1970) as modified by Anonymous (1990). Thismethod is currently used in Denmark to determine theseverity of Eimeria spp. infections and also for speciationin commercial flocks if molecular analysis (PCR) is unavail-able or unaffordable (S. Kabell, personal communication,2011). One pooled faecal sample from all tagged animalsper group, collected when animals were 59-days-old (totaln = 12) and stored as pellets under −18 ◦C for four monthswas examined by PCR for seven species of Eimeria (Hauget al., 2007) at the Laboratory of Parasitology, Swedish Vet-
erinary Institute. For field samples (oocysts in the presenceof debris), the protocol (GeneRelease) described by Hauget al. (2007) is expected to detect from 0.4 to 10 oocystsper PCR for seven species of Eimeria. Lastly, morphologicalsitology 186 (2012) 178– 187 181
examinations of oocysts were performed during the rou-tine of OPG counting and on the pooled samples accordingto Thebo et al. (1998).
2.8. Performance measurements
Consumption of concentrate and whole wheat wererecorded for each group twice a week during the out-door period and a mean individual feed consumption wascalculated for both genotypes. All broilers under studywere weighed individually four times during the study:at the beginning, when broilers were 29-days-old; when42-days-old (before expected infection build-up); when56-days-old (during expected infection), and at the end ofthe experiment when they were 74-days-old (after infec-tion declined). The total weight gain and the daily weightgain in those three specific periods were calculated indi-vidually, and the feed conversion rate was calculated perplot.
2.9. Statistical analysis
The infection dynamics was estimated by eight consec-utive measures from the same ten individual birds fromeach of the 12 groups. The mean OPG of males and femalesfrom each of the treatment groups over the period werepresented in curves to visually compare relevant effects.For the statistical analysis, we summed the eight consec-utive mean OPGs from each individual bird and log 10transformed this sum in order to obtain normality andhomogeneity of variance (Quinn and Keough, 2002). Anal-ysis of variance of log 10 transformed data was performedusing the MIXED procedure in SAS software (version 9.2)(SAS Institute, 2000; Littell et al., 1996) by the followingmodel:
Yijklmn = � + ˛i + ˇj + �k + (˛ˇ)ij + (˛ˇ)ik + (ˇ�)jk
+ (˛ˇ�)ijk + Am(ij) + εijklmn
where Yijklmn was the logarithm of the summed numberof oocysts per gram of faeces (OPG) of the individ-ual broiler; � = the mean value; ˛i = the genotype, i = 1,2(White Bresse L40, Kosmos 8 Ross); ˇj = the treatment,j = 1,2 (A. annua, control); �k = the sex, k = 1,2 (male,female); (˛ˇ)ij = the interaction genotype × treatment;(˛�)ik = the interaction genotype × sex; (ˇ�)jk = the inter-action treatment × sex; (˛ˇ�)ijk = the interaction geno-type × treatment × sex; Am(ij) = the random effect of plotnumber (genotype × treatment) and εijklmn = the error. Thep-value of the non-significant two and three-way inter-actions (higher than 0.20) were removed and a reducedmodel was used for the analysis of differences betweengroups.
For the statistical analysis of total weight gain, dailyweight gain and total food consumed during the exper-imental period, the above full model was used. Data onweight gain were not transformed as they were found to
be normally distributed. Data on food consumption werepresented at flock basis, and thus not available by sex.Interim food conversion rate (FCR) was estimated by thetotal amount of food consumed (mean) divided by the total
182 G.F.d. Almeida et al. / Veterinary Para
Fig. 1. HPLC–UV chromatogram of powdered A. annua leaves* used asa botanical coccidiostat added to the daily diet of broilers during thestudy. * Leaves contained an average of 0.93% artemisinin (Art), 0.19%dpb
gdV
3
3c
(4spcbdd(F
Fq“
ihydroartemisinic acid (DHAA), and 0.03% artemisinic acid (AA). The twineak with Art was identified as deoxiartemisinin (DOArt), which has noiological activity but is also produced by the leaves.
ain weight (mean). Tukey’s test was applied in order toetermine statistical differences between group means. p-alues < 0.05 were considered statically significant.
. Results
.1. Artemisinin content in feed and estimatedonsumption
The three different samples of powdered A. annuacv. Artemis) leaves, collected just after grinding, 20, and0 days after storage at room temperature showed veryimilar concentration of the three main bioactive sesquiter-ene lactones usually found in A. annua leaves. The averageompound concentration identified in the powder leavesy HPLC–UV, based on pure standards, all with a stan-
ard deviation of 0.01 were: Artemisinin (0.93 g/100 g),ihydroartemisinic acid (0.19 g/100 g) and artemisinic acid0.03 g/100 g). The HPLC–UV chromatogram is presented inig. 1. Based on an average artemisinin value of 0.93%, theig. 2. Average daily consumption of artemisinin* (mg/kg body weight) per genotyuantification of 0.93 g of artemisinin per 100 g of dry leaf powder added to dailySeeders” were introduced in the plots at day 10 when broilers were 39-days-old
sitology 186 (2012) 178– 187
estimated mean consumption of artemisinin provided inthe feed during the experiment is shown in Fig. 2 in orderto present that the prophylactic strategy succeeded.
3.2. Composition of supplementary feed and estimationof feed consumed
The composition of the food ingredients supplied tothe broilers during the experiment and the estimated feedconsumption during the trial are both available as supple-mental material to this article (link to Vet. Parasitol.). Asexpected, the total feed intake of the hybrid Kosmos washigher (+23.9%) than that of the pure White Bresse line(p < 0.001). No significant differences in feed consumptionwere found between groups, treated or not with A. annua.
3.3. Infection dynamics
The mean number of oocysts excreted by the “seeders”when introduced to the 12 plots outdoor was not different(p < 0.05). In addition, no clinical symptoms of disease werereported during the trial.
To identify Eimeria species, a first scoring of lesionswas performed at necropsy the day after the “seeders”were introduced. No pathological lesions were noted inbroilers previously treated with toltrazuril (n = 10) whilein the “seeders” group (n = 10) the identification of scat-tered white lesions in the duodenum mucosal surface –the development site of E. acervulina – in the intestine oftwo broilers (score 1 for both broilers, one of each geno-type) were noted. Small red petechiae on the serosal sideof the mid-intestine (site of E. maxima) were also noted infour of the “seeders” (two animals from each genotype). Nopathological lesions were noted in the caecal walls (site ofE. tenella).
At the end of the trial, 5 male broiler per plot (totaln = 60) were examined post mortem. No lesions were iden-tified in the intestine of 25 broilers (12 from the hybrid
Kosmos and 13 from the White Bresse line). In 35 birds,lesions were detected on the serosal surface with mean(± s.d.) numbers of lesions of 0.6 ± 0.2 vs. 0.6 ± 0.2 for theWhite Bresse genotype, treated and control respectively,pe. * Artemisinin mg equivalents/kg was calculated according to HPLC–UV diet. Adaptation period to the diet (from day 1 to day 10 of treatment)..

G.F.d. Almeida et al. / Veterinary Parasitology 186 (2012) 178– 187 183
Table 1Levels of significance of factors influencing coccidia infection and performance (averaged cumulative body gain weight between 29 and 74 days of age andmean daily weight gain over three specific periods of the broilers development: initial – from 29 to 42 days of age; mid – from 42 to 56 days of age and lateperiod – from 56 to 74 days of age).
Mean oocystexcretiona
Averaged bodyweight gain (g)
Mean daily weight gain (g/day)
Initial period 29–42d Mid period 42–56d Late period 56–74d
Treatment <0.05 NS <0.05 NS <0.05Genotype NS <0.001 <0.001 <0.001 <0.001Genotype × treatment NS NS NS NS NSSex <0.05 <0.001 <0.001 <0.001 <0.001Treatment × sex – <0.05 NS NS <0.05
NN
Genotype × sex – <0.05
Treatment × genotype × sex – <0.05
a log 10-Transformed for each experimental animal.
and 0.7 ± 0.1 vs. 0.5 ± 0.1 for the hybrid Kosmos, treatedand control respectively (no significant differences). 34(94%) lesions on the serosal surface were found in the mid-intestine and supposed to be in a healing phase, perhapsrelated to consecutive bacterial infections. Only two lesionswere found in the serosal surface of duodenum in two ani-mals, one from each genotype. A maximum of two lesionswere noticed for an individual bird (one of each genotype)in the serosal surface. No pathological lesions were notedin any part of the mucosa in any animal after opening ofthe intestines.
From PCR of pooled samples at 59-days of age, only E.acervulina could be identified. Exams from pooled sam-ples found occysts of the same size and morphology of E.acervulina. From morphological exams during routine OPGcounting, E. maxima stood out by detached size and shape.
Within the first two weeks after the introduction of
“seeders”, the prevalence of coccidia infection increasedfrom nil to 100% in all groups with the level of oocystdetection varying according to sex and treatment, and fol-lowing a similar trend in relation to the group replications.Table 2Effect of Artemisia annua on mean oocyst excretion (back transformed values frobody gain weight between 29 and 74 days of age and mean daily weight gain ove42 days of age; mid – from 42 to 56 days of age and late period – from 56 to 74 da
Variables Mean oocystexcretion (k OPGa)
Averagweight
Genotype Sex Treatment
W. Bresse Male + 40.0 1470
W. Bresse Male − 61.3 1474
W. Bresse Female + 56.3 1169
W. Bresse Female − 91.3 1183
Kosmos Male + 68.8 1957
Kosmos Male − 72.5 2060
Kosmos Female + 56.3 1665
Kosmos Female − 145.0 1571
SEMb 27.5 31.4
Average + 55.0 1565
− 92.5 1572
SEMb 17.5 15.7
Average Males 60.0 1740
Females 87.5 1397
SEMb 13.8 15.7
a k OPG = 1000 OPG.b SEM, standard error of the mean.
S NS <0.05S NS <0.05
During the experimental period, the measured oocyst out-put was significantly influenced by treatment (p < 0.05) andsex (p < 0.05) (Table 1). Thus, non-treated broilers showed67% higher oocyst output than broilers with access to A.annua dried leaves and on average, females excreted 45%more oocysts than males (Table 2). No effect of geno-type was found and no interaction between genotype andtreatment was observed. However, the difference in oocystoutput was consistently high in infected, untreated groupsparticularly in Kosmos females, which excreted twice asmuch oocysts compared to Kosmos males.
The course of oocyst excretion according to sex andtreatment is shown in Fig. 3. The mean oocyst countsrose to a maximum three weeks after the introduction ofthe “seeders” and significant reduced number of oocysts(p < 0.05) was observed during the identified infection onbirds of treated groups. A sharp decline in mean OPG
counts was observed in all groups in week 4, when animalswere 70-days-old. According to the infection dynamicspresented on the plotted curves (Fig. 3), apparently thenon-treated broilers from both sexes showed a tendency,m least square means of log 10) and performance (averaged cumulativer three specific periods of the broilers development: initial – from 29 toys of age).
ed body gain (g)
Mean daily weight gain (g/day)
Initial period29–42d
Mid period42–56d
Late period56–74d
26.6 30.7 38.628.8 30.2 37.622.3 25.3 29.123.8 25.6 28.534.4 43.6 50.740.9 44.2 50.529.9 38.2 41.634.2 35.4 35.0
1.3 1.1 1.0
28.3 34.5 40.031.9 33.9 37.9
0.8 0.6 0.5
32.7 37.2 44.427.5 31.1 33.6
0.6 0.5 0.5

184 G.F.d. Almeida et al. / Veterinary Parasitology 186 (2012) 178– 187
F * Sampla t period
eaab
3
itsgfmoda(
bmw(dwst
ig. 3. Mean oocyst excretion over time* in relation to sex and treatment.fter the introduction of the “seeders” to the plots in respect to pre-paten
ven debatable, to a small initial peak when 56-days of agend a later, consistent and larger peak when 63-days of ages compared to a single peak at 59-days of age from treatedroilers.
.4. Body weight gain
The overall body weight gain and the weight gain whennfection was subdued (i.e. after decline in oocyst excre-ion) showed a three-way interaction among genotype,ex and treatment (Table 1). A higher average weightain was reported in benefit to the supplemented Kosmosemales while the opposite was found in relation to supple-
ented Kosmos males. Associated to the minor differencesbserved between sexes for the White Bresse genotype, noifferences for the overall performance of flocks treatednd untreated were reported at the end of the trialTable 2).
Before the coccidial infection build-up (29–42 days),roilers from both genotypes and sex fed the diet supple-ented with A. annua had significantly lower average bodyeight gains (p < 0.05) than broilers in the control groups
Table 2). There were no significant differences for mean
aily weight gain in the intermediate period (42–56 days)hile in the late period (56–74 days), treated groups gainedignificant more weight (p < 0.05) than control birds afterhe subclinical coccidial challenge (Table 2).
ings for OPG counts begun when broilers were 45-days-old, the sixth days of Eimeria spp.
4. Discussion
Our study showed that adding A. annua to the feedsignificantly reduced the oocyst output per gram of fae-ces (OPG) in subclinical coccidia infections and increasedweight gain when infections were subdued. Our resultsagree with those of Allen et al. (1997) who reported 80%suppression in OPG, and that A. annua protected broilersagainst E. acervulina, the predominant species in our trial.Arab and colleagues (2006) reported OPG reductions of E.acervulina ranging from 90 to 95% resulting from supple-menting two different doses of pure artemisinin curativelyand it suggests that artemisinin can lead to a continuousand significant reduction in OPG that may reduce sever-ity of the disease, even under heavy challenge. However,it remains speculative to what degree the reduction inoocysts output observed in our study (60–70%) will pos-itively impact the epidemiology of the disease at higherinfection levels. While the complete elimination of the par-asite seems unattainable with a natural treatment suchas A. annua, a smaller infection level will lead to a build-up of immunity in infected birds, supporting the overallflock resistance. This is a desirable outcome for free rangeand organic systems and may reduce the risk of infections
by secondary opportunistic organisms like bacteria. Onthis background, our results indicate that supplementationwith dried A. annua leaves to broilers raised in a free-rangeorganic system may be used as a botanical coccidiostat. In
ry Para
G.F.d. Almeida et al. / Veterinaaddition, we observed that the effect of treatment was notdependent on genotype.
Assuming the positive effect of the preventive treat-ment, and based on reduced infection levels for treatedgroups (Table 2 and Fig. 3), we hypothesize that broilersfrom the non-supplemented groups were more suscepti-ble to infections than the treated broilers supplementedwith A. annua dried leaves. Eimeria acervulina is capableof excreting oocysts in faeces 12-fold higher than E. max-ima (Anonymous, 1990; Eckert et al., 1995). According tothe PCR results from pooled samples in our trial, only E.acervulina oocysts were found when broilers were 59-daysof age reconfirming its predominance in our trial. Assum-ing that broilers were first infected when they were around49-days of age, after the introduction of the “seeders”, thepre-patent period of the parasites suggests that the highestoocyst excretion is expected two weeks after the first inges-tion of oocysts (Velkers et al., 2010), in accordance to thepeaks identified in the non-treated groups, when broilerswere approximately 63-days-old (Fig. 3).
Although our HPLC analyses revealed that this cul-tivar contained 0.93% artemisinin and related bioactivesesquiterpene compounds (dihydroartemisinic acid andartemisinic acid), it is also known that the plant is a richsource of antioxidants, including flavonoids and pheno-lic acids that are believed to potentiate the bioactivity ofartemisinin against malaria (Ferreira et al., 2010). Otherantioxidant flavonoids unrelated to A. annua have docu-mented effects against coccidiosis in broilers (Naidoo et al.,2008; Wang et al., 2008). Hence, we cannot attribute ourfindings solely to the presence of artemisinin, although itis probably the major anti-coccidian compound producedby A. annua.
Interestingly, we found a sex effect on oocyst excre-tion. Females in the control group shed 75% more oocyststhan males (Table 2 and Fig. 3). As males and females werereared together, this finding suggests a true sex-related dif-ference. Zhu et al. (2000) estimating correlation parametersfor breeding programs for resistance to coccidiosis reportedno differences in oocyst excretion related to sex, but founddifferences between sexes for immunological responsesafter high dose inoculations. Danforth (1998) reported nodifferences between sexes for development of immunityafter vaccination for fast growing genotypes. Other thanthis report, we found no supportive evidence in the litera-ture for sex-related differences in avian coccidiosis.
Meanwhile, it was also observed that females from bothgenotypes in our study spent more time moving around,interacting socially and pecking for food when comparedto males (unpublished data), which agrees with Masic et al.(1974) and Savory (1975) who also reported females withhigher activity in the range compared to males in foragingbehaviour of chicken.
Infective oocysts are mainly available in faeces whichwere more abundant close to the broiler-house and to thesources of food. Thus, we consider that females were moreexposed to infection than males and this may explain the
sex differences in oocyst excretion in the untreated groups.Artemisinin is believed to interfere with parasite repro-duction against at least two species of Eimeria (Allenet al., 1997; Arab et al., 2006) and also to reduce the
sitology 186 (2012) 178– 187 185
capacity of oocysts to sporulate or survive in the environ-ment (del Cacho et al., 2010). However, the mechanism ofaction is largely unknown. Artemisinin may act directlyon parasitic stages or indirectly through interaction withthe intestinal micro-flora. It may cause suppression ofpro-inflammatory factors triggered by the parasite andimmunological responses to Eimeria spp. depending on thegenetic background of the birds. It was apparent, althoughnot significant, that artemisinin, and possibly other natu-ral compounds from A. annua, influenced the parasite-hostrelationship according to sex. In addition, the response totreatment in terms of weight gain was influenced by sex.Several explanations may be plausible in this context.
In different domesticated animal species, drugmetabolism (Sanvordeker and Lambert, 1974; Tam, 1983)and immunological responses to xenobiotics (Takahashiet al., 1993; Redmond et al., 2010) are known to beinfluenced by age, sex and genetic background. Anothercomplementary explanation might be that sexual differ-ences in behaviour among broilers could have influencedeven potentiated, other relevant factors as well as theresponse to A. annua compounds. Thus, to our knowledge,this is the first time a sex effect is reported when supplyinga botanical coccidiostat to broilers in a free-range system.
Even though A. annua was recently recommended asa rich source of protein and amino acids (Brisibe et al.,2009), its bitter taste imposed by sesquiterpenes leads tolow palatability to be supplied as a feed ingredient in free-range systems where other sources of food are availableto the birds. Allen et al. (1997) found that addition of 5%A. annua dried leaves affected negatively body weight gainover a three-week period compared to diets with lowerlevels of inclusion. In association with previous reports ofunpalatable components of plants used in livestock feedingstrategies (e.g., Caygill and Mueller-Harvey, 1999), we sug-gest that such bitter taste imposed by high doses suppliedin a short period of time (3 h) have interfered negativelywith the broilers’ performance before the build-up of coc-cidia infections, in particular regarding the hybrid Kosmosmales from our study. Maybe the addition of molassesor other natural sweetener (e.g., Stevia rebaudiana leaves)could mask the bitter taste of Artemisia and improve feedconsumption.
The interpretation of our results regarding the influ-ence of A. annua on performance is limited by the fact thatwe considered it logistically impossible to manage treatedand untreated non-infected groups with appropriate repli-cates. Birds are highly exposed to parasites when runningin free range systems (Permin and Hansen, 1998; Perminet al., 1999) and we found it unrealistic to avoid Eimeria spp.infections due to their opportunism and fast dissemination(Sharman et al., 2010). However, in view of the findingsby Brisibe et al. (2008) that uninfected layers consumingA. annua had increased feed intake and egg laying ratescompared to uninfected hens consuming a commercial dietwithout A. annua, the inclusion of an uninfected-treatedgroup would increase our knowledge on the benefits of this
plant to broiler chickens. Toghyani et al. (2010) recentlyshowed increased weight gains in caged uninfected broilersafter addition of thyme (Thymus vulgaris). It is also impor-tant to consider that different sources of food are available
1 ry Para
tchapfsb
cbtaadpmci
vpdtptfes
5
sbecTseoeetsFbcbe
C
orta
86 G.F.d. Almeida et al. / Veterina
o poultry in free range systems compared to a uniqueommercial diet fed to test birds in caged systems. Theigher nutritional variability and diversity of free-rangegroecosystems may inhibit (or increase) intake of anti-arasitic ingredients available to animals. This deservesurther investigation before new strategies for nutrientupply to poultry in organic and free range systems cane devised.
According to Velkers et al. (2010), the introduction ofoccidia with “seeders” provided rapid dissemination toroilers in all groups. It is a useful technique for experimen-ation with animals reared in organic systems as it avoidsrtificial inoculations, an undesirable practice regardingnimal welfare. Conversely, the contamination of the bed-ing material, as used in our experiment, without therevious knowledge of the Eimeria species present in theanure, may create experimental barriers and perhaps
ompromise supplementation studies of botanical coccid-ostats like A. annua due to species-specific mode of action.
The present method for provision of A. annua as a pre-entive measure was labour intensive and deemed notractical under normal farm conditions. Furthermore, itid not show major advantages in performance whenhe whole flock was considered. Nevertheless, due to theotential coccidiostatic effect of A. annua, alternative wayso provide natural bioactive plant components must beurther investigated to transfer this technology to farm-rs engaged in organic and free-range broiler productionystems.
. Conclusion
The preventive use of A. annua as a botanical coccidio-tat, at a daily intake level superior to 4 mg artemisinin/kgody weight, can markedly reduce oocyst output in broil-rs reared under common organic free-ranging farmingonditions and exposed to natural coccidia infections.he significant increase in oocysts output found in non-upplemented females suggests a sex influence related toither a more active foraging behaviour (resulting in higherocysts intake), or to a physiological/immunological differ-nce in how males and females cope with infections. A sexffect was also noted in relation to weight gain in responseo treatment with A. annua as an increased productivity waseen mainly in females from the hybrid Kosmos genotype.uture studies may indicate whether positive effects ofioactive compounds of A. annua can be enhanced withoutompromising feed intake of broilers in free range systemsy investigating different strategies for its extraction, deliv-ry and supplementation.
onflict of interest statement
The authors declare no conflicts of interest. The mention
f a commercial product is solely for the convenience of theeader and does not imply endorsement from the authorso any commercial products or brands over similar onesvailable in the market.sitology 186 (2012) 178– 187
Acknowledgements
Aarhus University and SOAR – Research School forOrganic Agriculture and Food Systems – are thanked forfinancing our study. The authors wish to thank Orla Nielsenfor his great technical support during the trial. Senior scien-tist Kai Grevsen is thanked for supplying Artemisia material.Also we want to thank Dr. Susanne Kabell from the DanishKnowledge Center for Agriculture for lesion score as wellas Drs. Per Thebo and Karin Troell from Swedish Veteri-nary Institute (SVA) for Eimeria speciation by PCR. Thanksare also due to Mr. Barry Harter for his valuable helpon HPLC–UV analysis. Finally, we wish to thank the twoanonymous reviewers which constructively contributed toincrease the quality of the paper.
Appendix A. Supplementary data
Supplementary data associated with this arti-cle can be found, in the online version, atdoi:10.1016/j.vetpar.2011.11.058.
References
Allen, P.C., Lydon, J., Danforth, H.D., 1997. Effects of components ofArtemisia annua on coccidia infections in chickens. Poult. Sci. 76,1156–1163.
Allen, P.C., Fetterer, R.H., 2002. Recent advances in biology and immunobi-ology of Eimeria species and in diagnosis and control of infection withthese coccidian parasites of poultry. Clin. Microbiol. Rev. 15, 58–65.
Al-Sheikhly, F., Al-Saieg, A., 1980. Role of coccidia in the occurrence ofnecrotic enteritis of chickens. Avian Dis. 24, 324–333.
Anonymous, 1990. Diagnosis of Coccidiosis in Chickens. Janssen Pharma-ceutica, Beerse, Belgium.
Arab, H.A., Rahbari, S., Rassouli, A., Moslemi, M.H., Khosravirad, F.D.A.,2006. Determination of artemisinin in Artemisia sieberi and anticoc-cidial effects of the plant extract in broiler chickens. Trop. Anim. HealthProd. 38, 497–503.
Bumstead, N., Millard, B., 1992. Variation in susceptibility of inbred linesof chickens to seven species of Eimeria. Parasitology 104, 407–413.
Brisibe, E.A., Umoren, U.E., Owai, P.U., Brisibe, F., 2008. Dietary inclusionof dried Artemisia annua leaves for management of coccidio-sis and growth enhancement in chickens. Afr. J. Biotechnol. 7,4083–4092.
Brisibe, E.A., Umoren, U.E., Briside, F., Magalhães, P.M., Ferreira, J.F.S.,Luthria, D., Prior, R.L., 2009. Nutritional characterisation and antiox-idant capacity of different tissues of Artemisia annua L. Food Chem.115, 1240–1246.
Caygill, J.C., Mueller-Harvey, I. (Eds.), 1999. Secondary Plant Products –Antinutritional and Beneficial Actions in Animal Feeding. NottinghamUniversity Press, Nottingham, UK, p. 129.
Cooper, K.K., Songer, J.G., 2009. Necrotic enteritis in chickens: A paradigmof enteric infection by Clostridium perfringens type A. Anaerobe 15,55–60.
Danforth, H.D., 1998. Use of live oocyst vaccines in the control of aviancoccidiosis: experimental studies and field trials. Int. J. Parasitol. 28,1099–1109.
Del Cacho, E., Gallego, M., Francesch, M., Quílez, J., Sánchez-acedo, C., 2010.Effect of artemisinin on oocyst wall formation and sporulation duringEimeria tenella infection. Parasitol. Int. 4, 506–511.
Duncan, I.J.H., 2002. Poultry welfare: science or subjectivity? Brit. Poult.Sci. 5, 643–652.
Eckert, J., Taylor, M., Catchpole, J., Licois, D., Coudert, P., Bucklar, H.,1995. Morphological characteristics of oocysts. In: Eckert, J., Braun, R.,Shirley, M., Coudert, P. (Eds.), COST 89/820. Biotechnology. Guidelines
of Techniques in Coccidiosis Research. Office for Official Publicationsof the European Communities, Luxembourg, pp. 103–119.EU, 1991. Commission regulation N.1538/91 of 15 June 1991 introduc-ing detailed rules for implementing regulation 1906/90 on certainmarketing standards for poultry. Off. J. Eur. Communities L143, 11–22.

ry Para
G.F.d. Almeida et al. / VeterinaEU, 2007. Commission regulation N.834/07 of 28 June 2007 on organicproduction and labeling of organic products and repealing Regulation(EEC) No. 2092/91. Off. J. Eur. Communities L189, 1–23.
Fanatico, A., 2006. Organic poultry production in the United States. ATTRA– National Sustainable Agriculture Information Service. Availableat: http://www.attra.org/attra-pub/PDF/coccidiosis.pdf (accessedJanuary 2011).
Ferreira, J.F.S., Gonzales, J.M., 2009. Analysis of underivatized artemisininand related sesquiterpene lactones by high-performance liquidchromatography with ultraviolet detection. Phytochem. Anal. 2,91–97.
Ferreira, J.F.S., Luthria, D.L., Sasaki, T., Heyerick, A., 2010. Flavonoids fromArtemisia annua L. as antioxidants and their potential synergism withartemisinin against malaria and cancer. Molecules 15, 3135–3170.
Haug, A., Thebo, P., Mattsson, J.G., 2007. A simplified protocol for molecu-lar identification of Eimeria species in field samples. Vet. Parasitol. 1,35–45.
IFOAM, 2005. The principle aims of organic production and processing.Basic standards. http://www.ifoam.org/about ifoam/standards/norms/norm documents library/Norms ENG V4 20090113.pdf(accessed September 2010).
Jensen, P., Toates, F.M., 1993. Who needs behavioral needs – motivationalaspects of the needs of animals. Appl. Anim. Behav. Sci. 37, 154–164.
Johnson, J., Reid, W.M., 1970. Anticoccidial drugs lesion scoring techniquesin battery and floor-pen experiments with chickens. Exp. Parasitol. 28,30–36.
Lillehoj, H.S., Trout, J.M., 1996. Avian gut-associated lymphoid tissues andintestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev.9, 349–360.
Littell, R.C., Milliken, G.A., Stroup, W.W., Wolfinger, R.D., 1996. SAS Systemfor Mixed Models. SAS Institute, Cary, NC.
Luchese, F.C., Perin, M., Aita, R.S., Mottin, V.D., Molento, M.B., Monteiro,S.G., 2007. Prevalência de espécies de Eimeria em frangos de criac ãoindustrial e alternativa. Braz. J. Vet. Res. Anim. Sci. 44, 81–86.
Masic, B., Wood-gush, D.G.M., Duncan, I.J.H., McCorquodale, C., Savory,C.J., 1974. A comparison of the feeding behaviour of young broiler andlayer males. Brit. Poult. Sci. 15, 499–505.
Morris, G.M., Woods, W.G., Richards, D.G., Gasser, R.B., 2007. Investigat-ing a persistent coccidiosis problem on a commercial broiler–breederfarm utilizing PCR-coupled capillary electrophoresis. Parasitol. Res.101, 583–589.
Naidoo, V., McGaw, L.J., Bisschop, S.P.R., Duncan, N., Eloff, J.N., 2008. Thevalue of plant extracts with antioxidant activity in attenuating coc-cidiosis in broiler chickens. Vet. Parasitol. 153, 214–219.
Olliaro, P.L., Taylor, W.R.J., 2004. Developing artemisinin-based drug com-binations for the treatment of drug-resistant falciparum malaria: areview. J. Postgrad. Med. 50, 40–44.
Permin, A., Hansen, J.W., 1998. The Epidemiology, Diagnosis and Controlof Poultry Parasites. A FAO Handbook. FAO, Rome, Italy.
Permin, A., Bisgaard, M., Frandsen, F., Pearman, M., Kold, J., Nansen, P.,1999. Prevalence of gastrointestinal Helminths in different poultry
production systems. Brit. Poult. Sci. 40, 439–443.Quinn, G.P., Keough, M.J., 2002. Experimental Design and Data Analysisfor Biologists, 6th printing, 2007. Cambridge University Press.
Redmond, S.B., Tell, R.M., Coble, D., Mueller, C., Palic, D., Andreasen,C.B., Lamont, S.J., 2010. Differential splenic cytokine responses to
sitology 186 (2012) 178– 187 187
dietary immune modulation by diverse chicken lines. Poult. Sci. 89,1635–1641.
Rodriguez, J.C., Segura, J.C., Alzina, A., Guterrez, M.A., 1997. Factors affect-ing mortality of crossbred and exotic chicken kept under backyardsystems in Yucatan, Mexico. Trop. Anim. Health Prod. 29, 151–157.
Roepstorff, A., Nansen, P., 1997. The Epidemiology, Diagnosis and Controlof Helminth Parasites of Swine. A FAO Handbook. FAO, Rome, Italy.
SAS Institute, 2000. SAS Online Doc® . Version 9.2. SAS Institute, Cary, NC.Sanvordeker, D.R., Lambert, H.J., 1974. Environmental modification of
mammalian drug metabolism and biological response. Drug Metab.Rev. 3, 201–254.
Savory, C.J., 1975. A growth study of broiler and layer chicks reared insingle-strain and mixed-strain groups. Brit. Poult. Sci. 16, 315–318.
Sharman, P.A., Smith, N.C., Wallach, M.G., Katrib, M., 2010. Chasingthe golden egg: vaccination against poultry coccidiosis. Parasitol.Immunol. 32, 590–598.
Tam, Y., 1983. Individual variation in 1st-pass metabolism. Clin. Phar-makon. 25, 300–328.
Thamsborg, S.M., 2002. Organic farming in the nordic countries – animalhealth and production. Acta Vet. Scand. Suppl. 43, 7–15.
Takahashi, K., Akiba, Y., Horiguchi, M., 1993. Sex-related differences ofhepatic microsomal mixed function oxidase system and antibody pro-duction in broilers implanted with corticosterone and/or fed ascorbicacid. Comp. Biochem. Physiol. 15, 159–164.
Thebo, P., Lunden, A., Uggla, A., Hooshmand-Rad, P., 1998. Identificationof seven Eimeria species in Swedish domestic fowl. Avian Pathol. 6,613–617.
Toghyani, M., Tohidi, M., Gheisari, A.A., Tabeidian, S.A., 2010. Perfor-mance, immunity, serum biochemical and hematological parametersin broiler chicks fed dietary thyme as alternative for an antibioticgrowth promoter. Afr. J. Biotechnol. 9, 6819–7682.
Velkers, F.C., Swinkels, W.J.C., Rebel, J.M.J., Bouma, A., Daemen, A.J.J.M.,Klinkenberg, D., Boersma, W.J.A., Stegeman, J.A., de Jong, M.C.M.,Heesterbeek, J.A.P., 2010. Effect of Eimeria acervulina infection historyon the immune response and transmission in broilers. Vet. Parasitol.173, 184–192.
Vestergaard, K.S., 1996. Assessing animal welfare: the significance ofcausal studies of behavior at the motivational level. Acta Agric. Scand.Sect. A: Anim. Sci. 27, 61–63.
Wang, M.L., Suo, X., Gu, J.H., Zhang, W.W., Fang, Q., Wang, X., 2008.Influence of grape seed proanthocyanidin extract in broiler chick-ens: effect on chicken coccidiosis and antioxidant status. Poult. Sci.87, 2273–2280.
Weinberg, E.D., Moon, J., 2009. Malaria and iron: history and review. DrugMetab. Rev. 41, 644–662.
Willcox, M., Falquet, J., Ferreira, J.F.S., Gilbert, B.H.S.U., Magalhães,E., de, P.M., Plaizer-Vercammen, J., Wright, C.W., 2007. Artemisiaannua as a herbal tea for malaria. Afr. J. Trad. Comp. Alt. Med. 4,121–123.
Williams, R.B., 1999. A compartmentalized model for the estimation ofthe cost of coccidiosis to the world’s chicken production industry. Int.
J. Parasitol. 8, 1209–1229.Zhu, J.J., Lillehoj, H.S., Allen, P.C., Yun, C.-H., Pollock, D., Sadjadi, M.,Emara, M.G., 2000. Analysis of disease resistance-associated param-eters in broiler chickens challenged with Eimeria maxima. Poult. Sci.79, 619–625.

90
Paper II
Feed intake and activity level of two broiler genotypes foraging different types
of vegetation in the finishing period. Poultry Science, 91 (2012) 2105– 2113.
Almeida, G.F.d.; Hinrichsen, L.K.; Horsted, K.; Thamsborg, S.M. and Hermansen, J.E.
Reprinted with permission of Poultry Science

INTRODUCTION In organic production systems, broilers must have ac-
cess to an outside area during part of the rearing period (EU, 2007). This is in addition to the high standards of animal welfare, avoidance of water and soil pollution, and increased agroecological biodiversity/resilience set out in the organic ideals (IFOAM, 2005) that guide worldwide certification procedures for livestock pro-duction in organic systems. In Europe, legislation for organic systems (EU, 1991) specifies that the outside areas available to broilers must be mainly covered with vegetation. However, the nature and type of the vegeta-tion is not specified, and in practice most free-range land is just identified as grass or plants (Walker and Gordon, 2003). Little attention has been given to make outdoor runs attractive and nutritious to broilers as
part of agroecosystem management (Van de Weerd et al., 2009).
In older traditional systems (Prothero, 1936), chick-ens were normally left to roam more naturally, seeking essential parts of their nutritional needs on or below the soil surface and consuming not only oil seed crops and cereals supplied by humans but also vegetation, seeds, fruits, soil particles, microorganisms, different stages of insects, and other arthropods and earthworms (Lomu et al., 2004; Gordon et al., 2006). Thus, Robinson (1961) estimated that feed savings could be up to 10% when fresh grass was available to hens in free-range systems. In a more recent investigation by Horsted and Her-mansen (2007), laying hens managed on different types of vegetation were able to meet a considerable part of their nutritional requirements through foraging. Other recent studies have suggested that plots cultivated with chicory (Cichorium intybus cv. Grassland Puna) were attractive to laying hens and reported a high intake of this plant (Horsted et al., 2006, 2007). A natural exten-sion of this hypothesis is that similar benefits might be achieved for broilers.
Feed intake and activity level of two broiler genotypes foraging different types of vegetation in the finishing period
G. F. d. Almeida ,*1 L. K. Hinrichsen ,† K. Horsted ,* S. M. Thamsborg ,‡ and J. E. Hermansen *
* Department of Agroecology, and † Department of Animal Science, Faculty of Science and Technology, Aarhus University, Blichers Allé 20, Research Centre Foulum, PO Box 50, DK-8830 Tjele, Denmark; and ‡ Danish Centre
for Experimental Parasitology, University of Copenhagen, Dyrlaegevej 100, DK-1870 Frederiksberg C, Denmark
ABSTRACT A study was performed with 2 broiler genotypes (slow and medium growth) restricted in supplementary feed and foraging 2 different mixed veg-etations (grass/clover or chicory) to identify possible benefits of herbage on nutrition during the finishing period (80 to 113 d of age). Three hundred birds were included in a 2 × 2 factorial design with groups of 25 birds replicated 3 times. The use of outdoor areas, performance, and forage intake were investigated. To identify possible differences in foraging activity, the use of the range was monitored one day per week at 4 dif-ferent times of the day. Feed intake from foraging was estimated by killing 4 birds per plot (2 males and 2 fe-males) in the morning and in the evening on 3 d during the experiment and measuring crop content. Vegetation type did not influence broiler use of the free-range area, feed intake, or performance. Differences in the use of
the range area, activity level, and feed content in the crops were observed in relation to genotype, sex, age of broilers, and also the time of day. Foraging activity was positively correlated with age. Medium-growth broilers spent more time inside and closer to the broiler houses during the day with increased foraging activity during evenings, in contrast to the slow-growing broilers that showed a more uniform activity during the day. Based on the measurement of crop content it was estimated that the slow-growing genotype had a daily intake of 5 to 8 g of forage per day, whereas the medium-growing genotype had an intake of 9 g for females and 20 g for males. In conclusion, limitation of supplemented pro-tein feed in the finishing period may be acceptable for broilers that have access to highly nutritious vegeta-tion.
Key words: broiler , crop content , forage , free-range , organic system
2012 Poultry Science 91 :2105–2113http://dx.doi.org/ 10.3382/ps.2012-02187
2105
Received January 25, 2012. Accepted May 2, 2012. 1 Corresponding author: [email protected] or gfaru-
© 2012 Poultry Science Association Inc.

More information on the feeding habits of broilers managed in free-range systems is needed to reassure farmers of the value of different types of vegetation as a means of nutrient supplementation. In a previous investigation on the foraging habits of different broiler genotypes, slow-growing broilers were found to spend considerably more time outside than fast-growing broil-ers, and this might influence foraging behavior (Nielsen et al., 2003).
The economic benefit of foraging, however, also de-pends on how it affects the feed conversion rate (FCR; feed used per kg of BW gain). Feed is used partly for maintenance and partly for tissue growth (Leeson and Summer 1997), where the former is a function of BW. Thus, FCR is generally impaired with increasing BW because of the proportionally higher feed requirement for maintenance compared with growth. Similarly, FCR is generally impaired in slow-growing birds because of proportionally higher demands for maintenance com-pared with growth. Organic production is typically based on a substantially higher slaughter weight and use of moderate- to slow-growing genotypes—aspects that are expected to impair feed conversion. If a part of this additional feed requirement can be supplied with herbage, this may improve the profitability of the pro-duction and reduce the load of nutrients at the outdoor area. Also, stimulating to foraging may be a means to counteract the higher cost of supplementary feed used for slow-growing broilers.
In addition, supplementing low-protein, low-energy diets, sometimes in a commercial strategy to reduce costs for industrial broiler systems, may lead birds to consume more of the available feed and thus increase feed intake (Leeson and Summers, 1997). This suggests that for free-range systems, restriction in commercial diet may stimulate broilers to increase feed intake of herbage available in the range.
On this background we investigated foraging activ-ity, performance attributes (weight gain, feed intake, and FCR) of 2 broiler genotypes—a slow-growing (pure breed) and a moderate-growing (hybrid) genotype—raised with access to different types of vegetation in the finishing phase of their growing period in a system closely resembling common farming practices to explore the contribution of foraging to the nutritional needs of the broilers.
MATERIALS AND METHODS
Study Site and Study DesignThe experiment was performed at Research Centre
Foulum, Faculty of Sciences and Technology, Aarhus University, Denmark (56°48′ N, 9°58′ E) in the sum-mer and autumn of 2010. The experiment used a 2 × 2 factorial design of 2 genotypes of broilers with access to 2 different types of mixed vegetation: grass (Lolium perenne) and clover (Trifolium repens) versus a mix of chicory (Cichorium intybus cv. Grassland Puna) and
spontaneous weeds (Lolium perenne, Trifolium repens, Artemisia vulgaris, Senecio sp., and Tripleurospermum sp.). The experimental treatments were replicated 3 times, thus including 12 flocks, each with 25 broilers of mixed sex, for a total of 300 broilers. Each group of 74-d-old birds was allocated a specific plot of 105 m2 (12 × 8.75 m) separated from each other by nylon net fences, yielding a stocking rate of 4.2 m2 of outdoor area per animal and a mobile hut (4.6 m2) with cover and perches in compliance with the European regula-tion for organic broilers (EU, 1999). The experiment began when the broilers were aged 80 d and terminated when they were 113 d old.
Experimental Birds and HousingThe 2 genotypes in question were White Bresse line
L40 (L40) from the Research Centre Foulum, a pure breed with slow-growing characteristics, and Kosmos 8 Red (K8R), a hybrid genotype with medium-grow-ing characteristics purchased from an Italian breeding company (http://www.olandia.it). The study was sub-divided into 3 different periods as suggested by Peder-sen et al. (2003) for the production of organic broilers in Denmark: inside (1 to 29 d of age), transition to the range areas during which coccidial infections in broilers of both genotypes were monitored (29 to 74 d of age), and the finishing period (from 74 to 113 d of age). In the latter period, the use of range area, activity level, feed intake, and performance attributes of broilers were all investigated.
Day-old chicks were grown in a clean and disinfected chicken house until the age of 29 d. Figure 1 illustrates the timeline of our study. In the transition period, the chicks were randomly selected and allocated to plots (n = 12) cultivated with grass/clover so they could adapt to the outdoor range and where subclinical coccidial in-fections were monitored until they were subdued when broilers were aged 74 d. Between d 45 and 74, 10 birds per group were monitored individually for coccidia oo-cyst excretion twice weekly as described in a previous study (Almeida et al., 2012). Prior to the experimental period in the present study, the broilers were randomly redistributed in the 12 flocks within genotype. In addi-tion, to avoid any effects or bias from intestinal nema-tode infections, all birds were treated (from 74 to 80 d of age) with flubendazole (Flubenol, Janssen Animal Health Ltd., Beerse, Belgium) via drinking water at recommended doses.
During the experimental period (from 80 to 113 d of age), animals of both genotypes were fed a typical organic feed for broilers in a restriction scheme (50 g/bird) to stimulate birds to forage especially for pro-tein and whole wheat and water supplied ad libitum. Two feeders with supplementary feed (wheat and con-centrate) were located inside the broiler houses, and a 25-L water tank was located outside. The experimental protocols involving broilers conformed to Danish legis-lation regarding animal care and health.
2106 ALMEIDA ET AL.

The consumption of whole wheat was recorded twice a week before refilling the feeders. Fifty grams per bird of commercial diet was filled into the feeders daily at 1100 h. All broilers under study were weighed individu-ally when 84 and 113 d old. From this strategy, it was possible to estimate the daily and accumulated gain, total weight gain, and FCR for both genotypes.
Assessment of the Use of Outdoor Areas and Broiler Activity Level
The number of broilers found outside and their ac-tivity were assessed by the same observer twice a week during the experiment. In the transition, preexperimen-tal period (29 to 74 d), the observer was trained in us-ing the instrument of data collection and at the same time the broilers became used to the presence of the observer.
Observations were performed when broilers were 80, 87, 94, 101, and 108 d of age from 0830 to 1000 h and again 1130 to 1300 h (Wednesdays) and from 1530 to 1700 h and 1830 to 2000 h (Thursdays). Weather condi-tions were found to be approximately similar for the 2 consecutive observation days in each week, so they were combined to give only 1 day of observation per week.
To evaluate broiler behavior, the paddocks were visu-ally subdivided into 5 main areas by inserting marking sticks in each corner of the subplots (Figure 2). The decision on which plot to start observations for each pe-riod of the day was made randomly by using a 12-sided die, each side representing one of the plots. After the initial plot had been determined, the remaining plots followed in numerical order. Observation of broiler be-havior started after the observer had stood for 1 min at a specified point outside the plot (marked 1 in Figure 2) to get the chickens used to the presence and snap-shots were then taken as suggested previously by Mar-tin and Bateson (2007). From the snapshots, the num-ber of broilers found outdoors in each subdivision was noted. The broilers categorized as being outdoors were those found in subplots 2 to 5, whereas broilers found either in subplot 1, drinking water, not visually found, or close to the houses were considered to be inside. The type of outdoor activity broilers were engaged in was
also recorded in 2 main categories: either resting with the body in contact with the soil/dust bathing or mov-ing around/searching for food (foraging) in the range.
Data were systematized and grouped for each plot according to treatment (type of vegetation) and geno-type. Results are presented as the mean percentage of broilers found outdoors for each period of the day and also as the percentage of broilers for the specific activi-ties based on the number of broilers found outdoors.
Crop Content, Sward Harvesting, and Estimation of Forage Degradation
Three times during the experiment, 4 broilers from each plot were necropsied to estimate the food habits at ages 84, 100, and 112 d. Two broilers (one male and one female), randomly selected from each plot, were killed (cervical dislocation) in the morning (between 0800 and 0900 h) and 2 broilers (one male and one female) in the evening (between 1800 and 1945 h). The entire crop was removed immediately afterward as suggested by Horsted et al. (2007), stored in plastic bags, identified with the bird ID and plot number, and stored in the freezer for future analysis.
As originally suggested by Jensen and Korschgen (1947), individual crops were thawed in the labora-tory and the content separated by forceps into differ-ent fractions: commercial broiler feed; wheat; grass; clover; chicory; other plant material; seeds; insects; earthworms, larvae, and pupae; grit stones; and soil. All fractions were dried in a forced air-drying oven at 60°C for approximately 24 h, and calculation of crop content was based on air-dried fractions (Antell and Ciszuk, 2006; Horsted et al., 2007). Because of the very small amount of seeds, insects, earthworms, larvae, pu-pae, grit stones, and soil found in the crops, these were
Figure 1. Study timeline and description of activities in relation to age of broilers. Broilers were randomized and allocated to the experi-mental plots at 74 d of age.
Figure 2. Plot spatial subdivision (areas 1 to 5) during behavioral studies, and point 1 outside the plots is the position of the observer when taking measurements and recordings.
2107TWO BROILER GENOTYPES FORAGING DIFFERENT VEGETATION

omitted from the analysis and we based our analysis on the contents of supplementary feed (commercial broiler feed and wheat) and herbage found in the crops. In our results we categorized the vegetative ingredients as grass, clover, chicory, and other plants. We estimated the total plant intake by summing the total herbage material found in the crops to evaluate the vegetative food intake and consequently the food habits during the experiment and from different periods of the day (morning and evening).
Before the experiment began, samples of vegetation were harvested from two 0.25-m2 patches from each plot. The location of the patches for sampling was de-cided beforehand, and the same locations were used for all 12 plots. The vegetation was cut approximately 2 cm above ground, and the harvested biomass from the 2 patches was pooled for each plot and stored in the freezer for further analysis.
To identify degradation of vegetation with time, 6 visual assessments (one per week) were performed from when the broilers were first introduced to the experi-mental plots. Vegetation degradation was estimated by 4 different visual measures in a 1-m2 area in each plot with the same location used for the 12 plots. The percentages of the main components of grass, clover, chicory, weeds, bare soil, and decomposed vegetation were estimated for each plot to a sum of 100%. The change in relative cover of the different components was compared to represent the degradation of herbage with time, and results were averaged according to treatment combinations.
Statistical MethodsEffects of treatments on the use of the range, crop
content, and performance attributes were subjected to ANOVA by linear models in SAS (SAS Institute, 2000). Data on the use of the range were based on
plot observations (plot as the experimental unit) and were found to be not normally distributed. Logarithmic transformation was performed to obtain normality and homogeneity of variance (Quinn and Keough, 2002). All main factors and their possible interactions were first included using a full model. No significant 3-way interactions existed, and these were therefore excluded from the final model. For the crop content analysis, in-dividual observations were available, thus including the sex of the broilers. The data were checked and found not normally distributed. The test was then performed with the natural log-transformed value to obtain nor-mality and homogeneity of variance with plots consid-ered as the error term. Weight gain, feed intake, and FCR (feed intake/weight gain) were analyzed with plot as the experimental unit, and data were checked and found to be normally distributed.
Where transformation was used, results are presented with back-transformed values. Feed conversion rate was estimated by the total amount of food supplemented (mean) divided by the averaged weight gain (mean). In addition to the above tests, behavior information was subjected to analysis of covariance to identify possible correlations between broilers found outdoors, their ac-tivity, and age. The P-values less than or equal to 0.05 were considered statistically significant.
RESULTS AND DISCUSSIONDuring the entire experimental period (aged 80 to
113 d), birds showed excellent health (no signs of foot-pad burns and good plumage state). Mortality was re-stricted to 2 of the K8R and 1 of the L40 genotypes. Causes of death were either unknown or culled because of injuries. There were no reports of predation because the 12 experimental plots were electrically fenced to keep out foxes and were covered with nylon nets to avoid avian predation.
Table 1. Nutritional contents of different feed ingredients1 used in the study (% DM)
Item
Supplementary feed Type of vegetation2
Commercial broiler feed
Whole wheat
Grass+ clover
Chicory+ weeds
Total DM (%) 88.4 86.5 15.1 16.1Percent of DM CP (N × 6.25) 19.7 11.6 15.9 11.2 Crude fat 6.3 2.0 2.6 2.4 Ash 6.3 1.5 9.9 12.4 Starch 46.0 67.9 4.0 3.7 Sugar 3.8 2.3 6.6 6.2 Cellulose 5.8 2.1 29.8 28.6ME (MJ/kg of DM) 13.4 14.1 4.2 3.6Amino acids (g/kg of DM) Lysine 10.5 3.5 9.5 8.5 Methionine 3.0 1.8 3.2 1.6 Threonine 7.4 3.5 9.0 7.7 Cysteine 3.5 2.8 1.5 1.4
1Two samples from each supplementary feed were analyzed with the mean values reported. Six samples from each type of vegetation were submitted for analyses, and the mean values are presented.
2Forage sampled before the introduction of broilers to the plots.
2108 ALMEIDA ET AL.
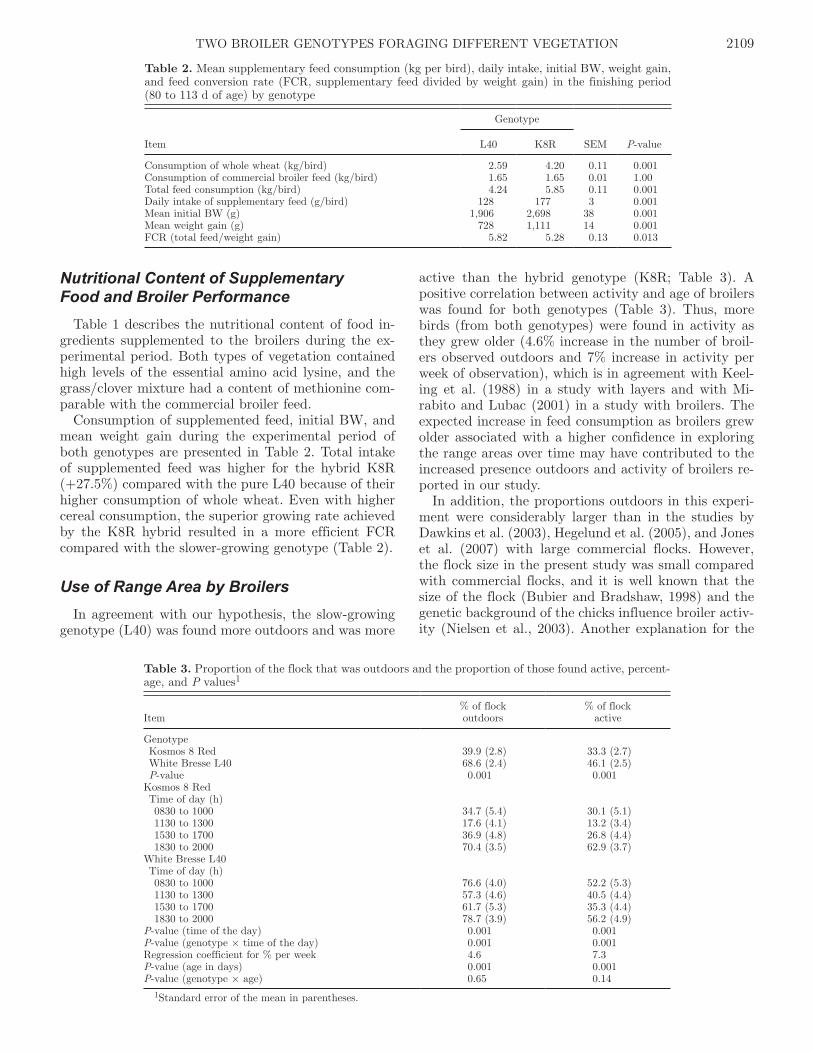
Nutritional Content of Supplementary Food and Broiler Performance
Table 1 describes the nutritional content of food in-gredients supplemented to the broilers during the ex-perimental period. Both types of vegetation contained high levels of the essential amino acid lysine, and the grass/clover mixture had a content of methionine com-parable with the commercial broiler feed.
Consumption of supplemented feed, initial BW, and mean weight gain during the experimental period of both genotypes are presented in Table 2. Total intake of supplemented feed was higher for the hybrid K8R (+27.5%) compared with the pure L40 because of their higher consumption of whole wheat. Even with higher cereal consumption, the superior growing rate achieved by the K8R hybrid resulted in a more efficient FCR compared with the slower-growing genotype (Table 2).
Use of Range Area by BroilersIn agreement with our hypothesis, the slow-growing
genotype (L40) was found more outdoors and was more
active than the hybrid genotype (K8R; Table 3). A positive correlation between activity and age of broilers was found for both genotypes (Table 3). Thus, more birds (from both genotypes) were found in activity as they grew older (4.6% increase in the number of broil-ers observed outdoors and 7% increase in activity per week of observation), which is in agreement with Keel-ing et al. (1988) in a study with layers and with Mi-rabito and Lubac (2001) in a study with broilers. The expected increase in feed consumption as broilers grew older associated with a higher confidence in exploring the range areas over time may have contributed to the increased presence outdoors and activity of broilers re-ported in our study.
In addition, the proportions outdoors in this experi-ment were considerably larger than in the studies by Dawkins et al. (2003), Hegelund et al. (2005), and Jones et al. (2007) with large commercial flocks. However, the flock size in the present study was small compared with commercial flocks, and it is well known that the size of the flock (Bubier and Bradshaw, 1998) and the genetic background of the chicks influence broiler activ-ity (Nielsen et al., 2003). Another explanation for the
Table 2. Mean supplementary feed consumption (kg per bird), daily intake, initial BW, weight gain, and feed conversion rate (FCR, supplementary feed divided by weight gain) in the finishing period (80 to 113 d of age) by genotype
Item
Genotype
SEM P-valueL40 K8R
Consumption of whole wheat (kg/bird) 2.59 4.20 0.11 0.001Consumption of commercial broiler feed (kg/bird) 1.65 1.65 0.01 1.00Total feed consumption (kg/bird) 4.24 5.85 0.11 0.001Daily intake of supplementary feed (g/bird) 128 177 3 0.001Mean initial BW (g) 1,906 2,698 38 0.001Mean weight gain (g) 728 1,111 14 0.001FCR (total feed/weight gain) 5.82 5.28 0.13 0.013
Table 3. Proportion of the flock that was outdoors and the proportion of those found active, percent-age, and P values1
Item% of flock outdoors
% of flock active
Genotype Kosmos 8 Red 39.9 (2.8) 33.3 (2.7) White Bresse L40 68.6 (2.4) 46.1 (2.5) P-value 0.001 0.001Kosmos 8 Red Time of day (h) 0830 to 1000 34.7 (5.4) 30.1 (5.1) 1130 to 1300 17.6 (4.1) 13.2 (3.4) 1530 to 1700 36.9 (4.8) 26.8 (4.4) 1830 to 2000 70.4 (3.5) 62.9 (3.7)White Bresse L40 Time of day (h) 0830 to 1000 76.6 (4.0) 52.2 (5.3) 1130 to 1300 57.3 (4.6) 40.5 (4.4) 1530 to 1700 61.7 (5.3) 35.3 (4.4) 1830 to 2000 78.7 (3.9) 56.2 (4.9)P-value (time of the day) 0.001 0.001P-value (genotype × time of the day) 0.001 0.001Regression coefficient for % per week 4.6 7.3P-value (age in days) 0.001 0.001P-value (genotype × age) 0.65 0.14
1Standard error of the mean in parentheses.
2109TWO BROILER GENOTYPES FORAGING DIFFERENT VEGETATION

present result might be the restriction of protein-rich commercial diet, which may have stimulated the birds to explore the forage area.
In our experiment broilers showed a diurnal rhythm that differed between the 2 genotypes in the study. The hybrid K8R was found much more inside and closer to the broiler houses during the day but with increased ac-tivity observed during evenings. In contrast, the slow-growing genotype (L40) showed a more uniform activ-ity pattern during the day. This confirms what other studies have found, namely that foraging behavior var-ies according to genotype (Kjaer and Mench, 2003), and most important for farm management, that there are preferences for specific times of the day.
According to our results and in agreement with Dawkins et al. (2003), during the periods close to noon (from 1100 to 1500 h) there was limited foraging activ-ity by broilers of both genotypes (Table 3), but espe-cially the K8R broilers. Restriction of the commercial diet to a daily feed at 1100 h may have influenced and even increased the need for exploring the range, espe-cially of hybrid K8R broilers in the evening periods.
Feed Intake Based on Crop ContentTable 4 gives the level of significance of factors af-
fecting the content of grass, clover, chicory, wheat, and commercial broiler feed in the crops of the broilers. In particular, genotype, sex, and slaughter date—re-flecting the age of the broilers—as well as time of the day influenced crop content of broilers. These effects are shown in Table 5, which for each genotype gives the amounts of total plant material, wheat, and com-mercial broiler feed found in the broiler crops for the morning and evening periods and for the 3 different harvesting dates in the experiment. Amount of plant material (total plants) in the crop was much higher in the afternoon than in the morning. The hybrid geno-type K8R had higher amounts of plant material in the crops than the L40 genotype and males had a higher content than females. The effect of genotype and sex, however, interacted with harvesting date in that to-tal plant material decreased over time in K8R broilers (most pronounced for males), whereas this pattern was not seen in L40 birds. This is probably a reflection of a reduction in plant availability at the end of the experi-ment (Figure 3). Looking at the grass and chicory sepa-rately, the amount of grass found in the crops increased linearly over time in contrast to the amount of clover and chicory, which linearly decreased over time (data not shown). This shows the reduction in availability of the highly nutritious clover and chicory (especially in chicory plots) during the period even at a low stocking rate (>4.2 m2/head outdoor area), as also illustrated in Figure 3.
The amount of whole wheat in the crops was highest in the afternoon and highest for the K8R genotype (Ta-ble 5), highest for males compared with females, and the differences between sexes were most pronounced for T
able
4. Si
gnifi
canc
e (P
val
ues)
of fa
ctor
s af
fect
ing
the
crop
con
tent
of br
oile
rs o
f bo
th g
enot
ypes
Food
typ
eV
eget
atio
n
type
Gen
otyp
eSe
xH
arve
stin
g da
teH
arve
stin
g tim
eV
eget
atio
n ty
pe
× g
enot
ype
Veg
etat
ion
type
×
sex
Veg
etat
ion
type
×
har
vest
ing
date
Gra
ss0.
910.
630.
0002
0.00
010.
0010
0.19
0.68
0.47
Clo
ver
0.00
010.
0058
0.00
010.
0031
0.00
330.
0217
0.01
020.
18C
hico
ry0.
0001
0.00
220.
0063
0.00
030.
0088
0.00
890.
0003
0.00
19Tot
al p
lant
10.
120.
0001
0.00
010.
140.
0001
0.83
0.78
0.26
Whe
at0.
730.
0225
0.00
010.
0114
0.00
760.
950.
450.
86C
omm
erci
al b
roile
r fe
ed0.
970.
980.
0002
0.00
890.
0001
0.27
0.12
0.58
Veg
etat
ion
type
×
har
vest
ing
tim
eG
enot
ype
× s
exG
enot
ype
×
har
vest
ing
date
Gen
otyp
e ×
har
vest
ing
tim
eSe
x
× h
arve
stin
g da
teSe
x
× h
arve
stin
g tim
eH
arve
stin
g tim
e
× h
arve
stin
g da
te
Gra
ss0.
440.
120.
740.
580.
290.
250.
0130
Clo
ver
0.01
630.
940.
0125
0.96
0.30
0.28
0.75
Chi
cory
0.08
0.22
0.01
620.
500.
130.
140.
0340
Tot
al p
lant
10.
890.
780.
0014
0.44
0.03
740.
410.
14W
heat
0.38
0.04
370.
250.
0397
0.35
0.38
0.24
Com
mer
cial
bro
iler
feed
0.67
0.30
0.02
120.
0001
0.00
360.
0015
0.55
1 Tot
al p
lant
: th
e to
tal am
ount
of gr
ass,
clo
ver,
and
chi
cory
acc
ount
ed in
the
broi
ler
crop
.
2110 ALMEIDA ET AL.

L40. In addition, an interaction existed between geno-type and harvesting date (age), where K8R had a higher increase in crop content with time than L40, probably reflecting the expected higher requirement for growth of the hybrid genotype when compared with the pure slow-growing breed. Apparently, due to a higher nutri-tional requirement, K8R broilers have compensated for a comparatively smaller availability of the commercial broiler feed by consuming more wheat compared with the slow-growing L40 genotype (daily allocation of con-centrate was 50 g/bird, for both genotypes).
Genotype did not appear to influence the content of concentrate in the crops. However, a difference was observed for sex (males having higher content than fe-males) probably reflecting the competitiveness of the males in the situation with the restriction in supple-mented commercial broiler feed. As already explained, all food items found in the crops were influenced by the time of the day in agreement with results of Horsted et al. (2007), with more items found in the crops for the evening than for the morning period.
Forage Contribution to Broiler NutritionThe analysis of broilers weight gain during the ex-
periment shows that L40 broilers grew, on average, 22 g/d, which agrees with the findings by Hermansen et al. (2004) for L40 broilers slaughtered at 130 d. The K8R broilers grew 33.6 g/d, which is approximately 10% more than the growing rate advertised by the breeding company for the same growing period (30.3 g/d), indi-cating that growth rate was not substantially reduced by the restricted supply of commercial broiler feed. As expected and in agreement with Leeson et al. (1999), both genotypes in our study had a high feed consump-tion per kilogram of gain (Table 2) in the finishing pe-riod. However, the interpretation of these results is lim-ited because we did not compare groups restricted with groups unrestricted in supplemented feed with appro-
Table 5. Amount of food items found in the crops based on genotype and sex of the broilers for each period of the day (morning and evening), for each of the 3 dates of harvesting and the estimated daily intake of herbage, by least squares means
Item
Genotype
SEM
White Bresse L40 Kosmos 8 Ross
Males Females Males Females
Total plant1 (mg) Time of day: morning 627 192 1,343 423 120 Time of day: afternoon 933 560 2,797 1,171 284 Harvest age (average morning + evening) Age of broilers (d) 84 711 350 2,394 744 162 100 802 219 3,117 1,140 425 112 827 560 701 507 87Wheat (g) Time of day: morning 7.3 3.2 12.1 11.6 0.8 Time of day: afternoon 10.4 7.4 13.1 10.4 0.9 Harvest age (average morning + evening) Age of broilers (d) 84 d 8.6 3.4 9.7 7.7 0.7 100 d 8.4 6.0 14.1 17.3 1.2 112 d 9.4 6.5 13.7 12.0 1.1Commercial broiler feed (g) Time of day: morning 2.1 1.4 0.2 0.2 0.2 Time of day: afternoon 15.7 7.2 18.7 13.5 1.2 Harvest age (average morning + evening) Age of broilers (d) 84 8.5 2.5 6.6 5.0 1.1 100 11.6 4.9 12.6 3.0 1.6 112 6.4 5.3 9.0 12.5 1.5Intake of total plant2 (g DM/d) 7.7 5.1 20.7 9.4 3.2
1Total plant: the total amount of grass, clover, and chicory in the crop.2Values calculated by the regression equation y = 0.144x − 0.178 from Antell and Ciszuk (2006) where y is the total amount of plant in the crop on
an air-dried based for the evening period and x is the estimated herbage intake per day, in grams of DM.
Figure 3. Estimated forage availability and degradation during the occupation of the forage area (from 74 to 113 d of age).
2111TWO BROILER GENOTYPES FORAGING DIFFERENT VEGETATION

priate replicates. Nevertheless, it remains unclear from our own measurements if the consumption of herbage affected FCR.
As discussed earlier, the herbage available to the broilers (Table 1) showed a relatively high nutritional content of essential amino acids. However, we were not able to estimate the possible intake hereof based on crop content because soil-living organisms also have a high content of essential amino acids (Pokarzhevskii et al., 1997). The insignificant amounts of soil particles found in broiler crops suggest that the dietary restric-tion applied in our study was not sufficient to stimulate animals to seek their amino acids from elsewhere as reported by Horsted et al. (2007), who found larger amounts of soil in the crops of layers with no access to a commercial diet compared with layers with access. Nevertheless, to estimate the possible contributions of amino acids from the herbage found in the broiler crops in our study, we tried to contrast their typical require-ments with the amount of these essential nutrients sup-plied in the feed (whole wheat + commercial broiler feed).
The daily requirements for lysine and methionine were taken from Chwalibog (1993), who estimated these according to the genotype, growth rate, and age of broilers. Thus, L40 broilers required 340 mg of ly-sine and 275 mg of methionine, and K8R broilers 495 mg of lysine and 385 mg of methionine. By comparing the requirements presented by Chwalibog (1993) with the content available in the supplementary feed (whole wheat + commercial broiler feed) corrected for DM, both genotypes apparently received sufficient lysine but not methionine. Thus, a proximate deficit in methio-nine supplementation in the order of 10% for L40 and 15% for K8R hybrid broilers was found.
To determine whether this shortfall would be met by its availability in herbage, we estimated the forage nutritional content by using a method first proposed by Antell and Ciszuk (2006) and later applied by Horsted et al. (2007), who investigated differences in herbage intake of layers foraging different types of vegetation. By applying the regression proposed by Antell and Cis-zuk (2006), the herbage found in the crops for the eve-ning periods reflects the daily herbage consumption in grams of DM per day (Table 5) for both genotypes and sexes. On the basis of the mean daily herbage intake from each genotype (Table 2) and the mean nutritional content of the herbages given in Table 1, the nutritional requirements seemed to be fulfilled considering those proximate calculations.
We also tried to estimate the contribution of energy from the forage crops, although it was difficult to ob-tain good estimates for the energy requirements based on the genotypes and ages of broilers used in our study. We finally settled on a method proposed by Sakomura et al. (2003) for parent stock of fast-growing broilers combined with a method proposed by NRC (1994) for slow-growing layers, correcting for the requirements as-sociated with egg production. By comparing the mean
values from both formulas with what was available from the supplemented feed, we propose that the sup-plemented feed was able to cover approximately 90% of the daily energy required for growth and maintenance of both genotypes. It is important to remember that neither method includes energy requirements for forag-ing activity.
In conclusion, the slow-growing genotype (L40) was found to be more active in exploring the range com-pared with the hybrid genotype K8R, which confirms our hypothesis. Conversely, restricted access to a com-mercial diet may have encouraged broilers to explore the range areas, especially the hybrid K8R genotype, presumably because it had a higher deficit of the amino acid methionine. However, the supply of nutrients from forage to free-range broilers is more uncertain if large flocks are targeted, which limits the prospects of up-scaling our findings. From a farm design point of view, it should be borne in mind that broilers can consume considerable amounts of forage as part of their nutri-tional needs and this is mainly influenced by genotype, age of the broilers and time of the day. Further studies are necessary to elucidate if a more restricted feeding of commercial broiler feed to large flocks, supplemented especially during morning periods, can stimulate forag-ing activity and increase the herbage intake during the evenings as part of a strategy to achieve 100% on-farm self-sufficiency in the nutrient supply to broilers in or-ganic systems.
ACKNOWLEDGMENTSAarhus University (Denmark) and SOAR (Research
School for Organic Agriculture and Food Systems, Tjele, Denmark) are thanked for financing our study. The authors thank Orla Nielsen (Department of Agro-ecology, Research Center Foulum, Aarhus University) for his great technical support during the study. Thanks are also due to Margit Schacht (Aeces Flyter Ind, Agro Business Park, Tjele, Denmark) for her valuable help on the English revision of this manuscript. Finally, we wish to thank the two anonymous reviewers who con-structively contributed to increasing the quality of the paper.
REFERENCESAlmeida, G. F. d., K. Horsted, S. M. Thamsborg, N. C. Kyvsgaard,
J. F. S. Ferreira, and J. E. Hermansen. 2012. Use of Artemisia annua as a natural coccidiostat in free-range broilers and its ef-fects on infection dynamics and performance. Vet. Parasitol. 186:178–187.
Antell, S., and P. Ciszuk. 2006. Forage consumption of laying hens—The crop content as an indicator of feed intake and AME content of ingested forage. Arch. Geflugelkd. 70:154–160.
Bubier, N. E., and R. H. Bradshaw. 1998. Movement of flocks of lay-ing hens in and out of the hen house in four free range systems. Br. Poult. Sci. 39(Suppl.):S5–S10.
Chwalibog, A. 1993. Husdyrernæring–Bestemmelse af næringsværdi og næringsbehov 2. Udgave. Jordbrugsforlaget, København.
Dawkins, M. S., P. A. Cook, M. J. Whittingham, K. A. Mansell, and A. E. Harper. 2003. What makes free-range broiler chickens
2112 ALMEIDA ET AL.

range? In situ measurement of habitat preference. Anim. Behav. 66:151–160.
EU. 1991. Commission regulation N.1538/91 of 15 June 1991 in-troducing detailed rules for implementing regulation 1906/90 on certain marketing standards for poultry. Off. J. Eur. Communi-ties (L143):11–22.
EU. 1999. Council Regulation 1804/1999 supplementing Regula-tion 2092/91 on organic production. Off. J. Eur. Communities 42(L222):1–28.
EU. 2007. Commission regulation N.834/07 of 28 June 2007 on or-ganic production and labelling of organic products and repeal-ing Regulation (EEC) No 2092/91. Off. J. Eur. Communities (L189):1–23.
Gordon, S. H., B. Cottrill, M. Omiczek, T. Verhoeven, R. Weight-man, R. Safford, F. Nicholson, S. Holmes, T. Turner, and H. A. van de Weerd. 2006. Organic egg production—A desk study on sustainable and innovative methods for meeting the hen’s protein requirements. ADAS report on Defra project OF0357. Accessed Feb. 2011. http://randd.defra.gov.uk/Default.aspx?Menu=Menu&Module=More&Location=None&ProjectID=13671&FromSearch=Y&Publisher=1&SearchText=OF0357&SortString=ProjectCode&SortOrder=Asc&Paging=10#Description.
Hegelund, L., J. T. Sørensen, J. B. Kjær, and I. S. Kristensen. 2005. Use of the range area in organic egg production systems: Effect of climatic factors, flock size, age and artificial cover. Br. Poult. Sci. 46:1–8.
Hermansen, J. E., K. Strudsholm, and K. Horsted. 2004. Integration of organic animal production into land use with special reference to swine and poultry. Livest. Prod. Sci. 90:11–26.
Horsted, K., M. Hammershøj, and J. E. Hermansen. 2006. Short-term effects on productivity and egg quality in nutrient-restricted versus non-restricted organic layers with access to different for-age crops. Acta Agric. Scand. 56:42–54.
Horsted, K., and J. E. Hermansen. 2007. Whole wheat versus mixed layer diet as supplementary feed to layers foraging a sequence of different forage crops. Animal 1:575–585.
Horsted, K., J. E. Hermansen, and H. Ranvig. 2007. Crop content in nutrient-restricted versus non-restricted organic laying hens with access to different forage vegetations. Br. Poult. Sci. 48:177–184.
IFOAM. 2005. Norms for organic production and processing. Basic standards. Accessed Sep. 2010. http://www.ifoam.org/about_if-oam/standards/norms/norm_documents_library/Norms_ENG_V4_20090113.pdf.
Jensen, G. H., and L. J. Korschgen. 1947. Contents of crops, giz-zards, and droppings of bobwhite quail force-fed known kinds and quantities of seeds. J. Wildl. Manage. 11:37–43.
Jones, T., R. Feber, G. Hemery, P. Cook, K. James, C. Lamberth, and M. Dawkins. 2007. Welfare and environmental benefits of integrating commercially viable free-range broiler chickens into newly planted woodland: A UK case study. Agric. Syst. 94:177–188.
Keeling, L. J., B. O. Hughes, and P. Dunn. 1988. Performance of free-range laying hens in a polythene house and their behavior on range. Farm Build. Progr. 94:21–28.
Kjaer, J. B., and J. A. Mench. 2003. Behaviour problems associated with selection for increased Production. Pages 67–82 in Poultry Genetics, Breeding and Biotechnology. W. M. Muir and S. E. Aggrey, ed. CAB International, Wallingford, UK.
Leeson, S., L. J. Caston, J. D. Summers, and K. H. Lee. 1999. Per-formance of male broilers to 70 days when fed diets of varying nu-trient density as mash or pellets. J. Appl. Poult. Res. 8:452–464.
Leeson, S., and J. D. Summers. 1997. Commercial Poultry Nutrition. 2nd ed. University Books, Guelph, Ontario, Canada.
Lomu, M. A., P. C. Glatz, and Y. J. Ru. 2004. Metabolizable energy of crop contents in free-range hens. Int. J. Poult. Sci. 3:728–732.
Martin, P., and P. Bateson. 2007. Measuring Behaviour. An Intro-ductory Guide. Cambridge University Press, Cambridge, UK.
Mirabito, L., and S. Lubac. 2001. Descriptive study of outdoor run occupation by ‘Red Label’ type chickens. Br. Poult. Sci. 42:16–17.
Nielsen, B. L., M. G. Thomsen, P. Sorensen, and J. F. Young. 2003. Feed and strain effects on the use of outdoor areas by broilers. Br. Poult. Sci. 44:161–169.
NRC. 1994. Nutrient Requirements of Poultry. NRC’s Subcommit-tee on Poultry Nutrition. 9th rev. ed. Natl. Acad. Press, Wash-ington, DC.
Pedersen, M. A., S. M. Thamsborg, C. Fisker, H. Ranvig, and J. P. Christensen. 2003. New production systems: Evaluation of organic broiler production in Denmark. J. Appl. Poult. Res. 12:493–508.
Permin, A., M. Bisgaard, F. Frandsen, M. Pearman, J. Kold, and P. Nansen. 1999. Prevalence of gastrointestinal helminths in dif-ferent poultry production systems. Br. Poult. Sci. 40:439–443.
Pokarzhevskii, A. D., D. P. Zaboyev, G. N. Ganin, and S. A. Gordi-enko. 1997. Amino acids in earthworms: Are earthworms ecosys-temivorous? Soil Biol. Biochem. 29:559–567.
Prothero, R. E. 1936. English Farming Past and Present. 1961 ed. First published in 1912. Heinemann and Frank Cass and Co., London, UK.
Quinn, G. P., and M. J. Keough. 2002. Experimental design and data analysis for biologists. 6th rev. ed. Cambridge University Press.
Robinson, L. 1961. Modern Poultry Husbandry. Crosby Lockwood, London, UK.
Sakomura, N. K., R. Silva, H. P. Couto, C. Coon, and C. R. Pa-checo. 2003. Modeling metabolizable energy utilization in broiler breeder pullets. Poult. Sci. 82:419–427.
SAS Institute. 2000. SAS OnlineDoc®. Version 9.2. SAS Institute Inc., Cary, NC.
Van de Weerd, H. A., R. Keatinge, and S. Roderick. 2009. A review of key health-related welfare issues in organic poultry production. World’s Poult. Sci. J. 65:649–684.
Walker, A., and S. Gordon. 2003. Intake of nutrients from pasture by poultry. Symposium on Nutrition of Farm Animals Outdoors. Proc. Nut. Soc. 62:253–256.
2113TWO BROILER GENOTYPES FORAGING DIFFERENT VEGETATION

100
Paper III
The effects of cassava (Manihot esculenta), annual wormwood (Artemisia annua)
and mugwort (A. vulgaris) on coccidiosis in organically reared pullets in Brazil
Almeida, G. F. d.; Thamsborg, S. M.; Horsted, K.; Magalhães, P. M.; Ferreira, J. F. S. and
Hermansen, J. E. (2012) Manuscript submitted to Asian-Australasian Journal of Animal Sciences
Published as original manuscript with permission of Asian-Australasian Journal of Animal
Sciences

101
The effects of cassava (Manihot esculenta), annual wormwood (Artemisia annua) and
mugwort (A. vulgaris) on coccidiosis in organically reared pullets in Brazil
Gustavo F. de Almeida a *; Stig M. Thamsborg b; Klaus Horsted a; Pedro M. de Magalhães c; Jorge
F. S. Ferreira d; John E. Hermansen a
a Department of Agroecology, Faculty of Sciences and Technology, Aarhus University. Research
Centre Foulum, Blichers Allé 20, P.O. Box 50 DK-8830 Tjele, Denmark.
b Danish Centre for Experimental Parasitology, Faculty of Health and Medical Sciences,
University of Copenhagen, Dyrlægevej 100, DK-1870 Frederiksberg C, Denmark.
c Chemical, Biological and Agricultural Pluridisciplinary Research Centre (CPQBA), University of
Campinas – UNICAMP, P.O. Box 6171, BR-13081-970 Campinas, SP, Brazil.
d USDA-ARS, US Salinity Laboratory, 450 W Big Spring Rd., Riverside, CA 92507-4962.
* Corresponding author: Gustavo Fonseca de Almeida; Tel. +45 8715 4757; Fax. +45 8715 4798.
E-mail address: [email protected]

102
ABSTRACT: With the aim to identify alternatives to chemical coccidiostats, an on farm
experiment was carried out to investigate potential herbal candidates and strategies of delivery.
Pullets (n=2,300) were randomly allocated to 7 treatments in two replicates. One group of pullets
was supplemented with 3% cassava dried leaves during the first 8 days of life while two groups
were supplemented with 3% Artemisia annua either the first 8 weeks or from week 4 to 8. One
group was supplemented with a concentrated ethanolic extract of A. vulgaris via drinking water
before expected appearance of clinical symptoms while another group was supplemented with A.
annua extract via drinking water after the expected appearance of clinical symptoms. The response
of the above five treatments was compared to a vaccinated group and to the farmer routine, which
was the supplementation of an ethanolic extract of A. vulgaris via drinking water after appearance
of clinical symptoms. Weight gain, feed intake and coccidia (Eimeria spp.) oocyst excretion (OE)
were monitored weekly from 1 to 14 weeks of age. The supplementation of A. annua at 3% in feed
reduced the growth rates of pullets (p<0.05) without affecting OE. While the effects of ethanolic
extracts were limited, the supplementation of 3% cassava dried leaves markedly suppressed OE
(p<0.05) compared to vaccinated pullets without affecting growth rate. In conclusion, cassava may
be an effective strategy to control coccidiosis in small scale systems. Further investigation
focusing on specific species of coccidia, mixing bioactive compounds and testing different periods
of supplementation are warranted.
Keywords: chicken; natural anti-protozoa drug; welfare; cassava; ethanolic plant extract; organic
farming.

103
INTRODUCTION
Coccidiosis remains one of the most important constraints in the poultry industry
worldwide. In the last decades, the disease has been prevented through the addition of anti-
coccidial drugs in the feed. However, the risk of drug residues in poultry products (Ungemach et
al., 2006) has raised consumers’ demand for food with minimal drug use (Herrmann and Roder,
1995). In this respect, new methods to control the disease through food supplements described as
“natural” are likely to play an increasing role (Orengo et al., 2012) since they have been well
accepted by consumers (Brenes and Roura, 2010).
The production of poultry in organic systems is regulated: emphasizing animal welfare,
health and the quality of its products (EU, 1991; EU 2007; BRASIL, 2003; BRASIL, 2007).
National regulations prohibit the use of drugs for preventive treatment of infectious diseases,
including parasites. As a result, treatment of coccidiosis depends on innovative substitutes to
currently-used drugs. The use of live attenuated vaccines (Willians, 2002) is a useful option
available in the market, although not always affordable to the small scale farmer. Thus, the use
of plants and their derivatives may represent a viable alternative (Abbas et al., 2012).
One relevant feed source in this respect is cassava (Manihot esculenta), a tropical crop
that is traditionally cultivated in developing countries as a source of starch. Apart from the main
product, the root, cassava also produces large amounts of leaves that typically are left in the
fields (Sokerya et al., 2007). Fresh cassava forage, including the tender stems, can be used
directly for feeding growing chicks (Ravindran, 1991). However, in diets based on maize-
soybeans grains, the leaf meal was first suggested to be limited to 5 g kg-1 in the diet (Ross and
Enriquez, 1969) due to possible anti-nutritional effects of condensed tannins (CT). Condensed
tannins in cassava leaves may vary from 0.6 to 3.8 g/kg dry matter (DM) according to the
cultivar, period of harvest, drying method, and leaf age (Oni et al., 2011). Condensed tannins are
known to kill pathogenic bacteria (Barry et al., 1986), to have anthelmintic effects in parasitized
dairy cattle and buffaloes (Netpana et al., 2001), and to have coccidiostatic effects in parasitized
goats (Seng and Rodriguez, 2001) and sheep (Marie-Magdeleine et al., 2010). Thus, we
hypothesized that supplementing cassava leaf meal for a short period of time would protect
young pullets against the effects of early coccidia infections.

104
Also, other candidate plants have been investigated as natural food supplements to
control coccidiosis (Naidoo et al., 2008; Wang et al., 2008). Plants from the genus Artemisia
were reported to be effective against a variety of parasites, including apicomplexans (Wright,
2005; Liu et al., 2006) and A. annua (annual or sweet wormwood) was previously reported to be
effective against at least two species of Eimeria spp., the causative agent of coccidiosis (Allen et
al, 1998; Arab et al. 2006). Artemisia annua dried leaves mixed in feed was suggested to be
supplemented prior to coccidia challenge to be effective against E. acervulina (Allen et al., 1997).
For this reason, we found relevant to investigate both early (before the appearance of clinical
symptoms) and late (after the appearance of the clinical symptoms) administration of herbal
coccidiostats. It has been suggested that artemisinin, the main bioactive sesquiterpene lactone
biosynthesized in A. annua leaves is the main anti-coccidial compound. Its mode of action may
affect the intracellular reproduction mechanism of Eimeria spp. (Allen et al, 1997; Arab et al.,
2006) and protein degradation during formation of the oocyst wall (Del Cacho et al., 2010). The
plant is also considered a rich source of antioxidants, being one of the Chinese medicinal herbs
with the highest ORAC (oxygen radical absorbance capacity) level (Zheng and Wang, 2001).
Another plant from the genus Artemisia, A. vulgaris (common mugwort) has been
reported to have anthelmintic and anti-inflammatory properties in humans (Tigno et al., 2000;
Tigno and Gumila, 2000) supposedly due to high levels of antioxidants (Temraz and El-Tantawy,
2008), since this species does not contain artemisinin (Mannan, 2010).
On this background, the objective of this work was to investigate possible benefits of
Artemisia annua, A. vulgaris, and Manihot esculenta as potential coccidiostats provided with
daily feed or drinking water on feed intake, growth rate and oocyst excretion.
MATERIALS AND METHODS
The experimental farm and the current farm routine against coccidiosis
The study was performed in an organic commercial farm located in Jaguariúna, São
Paulo state, Brazil (22°38′ S, 47°00′ W). The production of eggs at Yamaguishi farm is
organized in four stables with capacity of 2500 chickens/stable. Each stable is subdivided in 25
pens of 4 x 8 m with concrete floor and access to an outdoor chicken run/corridor (4x20 m) with

105
Brachiaria decumbens grass and spontaneous weeds. The subdivision of the stables to a capacity
of no more than 100 layers per sub-flock is believed to provide opportunities for better
management and animal welfare. A farm routine is used to identify and treat Eimeria spp.
infections at clinical stage. Following the appearance of blood on faeces and depression of young
pullets in a sub-flock basis, 15 drops of a concentrated ethanolic extract of A. vulgaris was mixed
with 25 L drinking water per day for seven consecutive days.
Experimental design, treatments and management of pullets
Our experiment was linked to the farm routine, following its production schedule. In
December 2010, approximately 2,300 one-day-old pullets (plus 200 males allocated in a
separated pen) of the Isa Brown breed were randomly allocated to one of 14 pens (165 pullets
per pen) thus allowing two replicates for each of the 7 treatments. Table 1 and Figure 1 describe
the treatments, age of the pullets when fed the herbal supplements and the route of delivery.
The pullets were introduced to a non-disinfected stable; visible manure was removed and
new wood shavings were provided. Pullets were supplemented with water and a starter diet ad
libitum, feed supplied in movable tray feeders and the water in grooves available in each pen.
Pullets were vaccinated against Marek disease, Gumboro, infectious Bronchitis, coryza, Fowl
pox, Encephalomyelitis and New castle as demanded by the Brazilian regulation for production
of layer hens (BRASIL, 2011).
Heat was provided to the pullets during the first two weeks of life by an electrical lamp
placed inside chicken brooders in each pen. By the end of week 7, pullets were allowed to
explore the outdoor runs and perches were disposed in the top of the brooders. From One-day-
old to 7 weeks of age, all groups were daily supplemented with a maize-soybean basal starter
diet ad libitum. From week 8 to the end of week 14, all groups received a basal growing diet with
a higher content of energy and protein and a small amount of basaltic gravel to stimulate the
development of the gizzards (Table 2). When the pullets reached 30 days of age, they were
supplemented daily 10 g/head of Napier grass to stimulate the pullets’ interest in the
consumption of herbage.
Plant material and determination of plant anti-parasitic components

106
New top leaves of cassava were harvested at Yamaguishi farm and the stems were
removed. Leaves were dried in the shade (22-23°C) for one month. Approximately 1.5 kg of
dried leaves was stored in a plastic bag in a controlled environment until needed. Just before
formulating the initial diet, the leaves were crushed by hand while sifted through a five-mm
mesh sieve, and these particles were used in the diet formulation for the Ca8d group.
The A. annua L. (var. CPQBA) used in our study was improved to produce high
artemisinin content. Plants were grown on an oxisoil at the Chemical, Biological and
Agricultural Pluridisciplinary Research Centre - CPQBA, University of Campinas, São Paulo,
Brazil (22° 47′ S, 47° 06′ W) and harvested in February 2010, just before flowering, when
artemisinin reaches its peak. The whole plant was dried in the sun during one week and leaves
were separated from secondary stems (long branches) by hand. Before storage, the leaves were
separated from tertiary stems (small branches) by hand and by the use a coffee sieve. After 10
month of storage in a controlled environment (room temperature, 22ºC in shade), leaves were
ground by a hammer mill and sifted through a five-mm mesh sieve, stored in two 20-L carton
bags and transported to the Yamaguishi farm in the same day.
The ethanolic extracts of A. annua and A. vulgaris were prepared at CPQBA - University
of Campinas - using the same extraction procedure. Leaves of A. vulgaris cultivated also at
CPQBA were harvested in September 2010 and dried in the shade for one month in a controlled
environment just before preparation of the ethanolic extract. In a 1 L Erlenmeyer flask wrapped
with aluminium foil to prevent oxidation and degradation by light, 100 g of dried leaves (without
small branches and crushed by hand) were mixed with 1 L ethanol 70% (700 mL ethanol 97° +
300 mL distilled water). The mixture was kept in a dark room with controlled temperature (22°C)
and homogenised with circular movements twice per day for 20 days. The content was filtered
by using paper filters and transferred to 50mL flasks with dropping glass dispensers. Flasks were
identified and stored in the fridge until needed. One week before the trial, the flasks with the
Ethanolic extracts of A. annua and A. vulgaris were transported to the farm and kept in the fridge
before its use.
Chemical evaluation of artemisinin, dihydroartemisinic acid, and artemisinic acid from
the A. annua and A. vulgaris leaves and also from its ethanolic extracts were performed as
suggested by Ferreira and Gonzalez (2009). In addition, tests were conducted to identify the

107
oxidative absorbance capacity level - ORAC of the M. esculenta dried leaves and of the Napier
grass, both based on the DM according to Ferreira and Luthria (2010).
Monitoring coccidia infection and speciation
Faecal samples were collected weekly for each pen from week 1 to week 14 for coccidial
oocysts counting in a method adapted from Long and Rowell (1975). The farmer was trained on
the collection of fresh faeces in 5 different areas of each pen, recording of colour and textures,
and to avoid contamination with wood shavings by the use of disposable chopsticks. Every
Tuesday, one pooled sample per pen of approximately 15 grams (10 to 15 faecal sub-samples)
were stored in plastic disposable containers (n=14/week) and kept in the fridge until next day.
Every Wednesday, samples were transported in a refrigerated box to a private laboratory of avian
diseases accredited by the Brazilian Ministry of Agriculture (www.avipa.com.br) for faecal
oocyst counts. Faeces were examined by a modified McMaster technique using saturated NaCl
solution with 50% glucose monohydrate as flotation fluid with a sensitivity of 20 oocysts per g
of faeces (OPG) (Almeida et al., 2012). For Eimeria speciation, oocysts were purified from
pooled faecal samples from weeks 3, 4, 5 and 6 from each pen and analysed by multiplex PCR
(Fernandez et al., 2003) in the Laboratory of Parasitology, Institute of Biological Sciences -
University of São Paulo, Brazil. The infection dynamics for the treated groups is discussed for
the period pullets were indoor (early infection) and for the period pullets accessed the outdoor
(late infections).
Production parameters
Consumption of diet was recorded for each pen every week and the mean diet
consumption was calculated based on the previous 7 days and estimated for individual pullets. In
the same day, 16 pullets from each group (representing approximately 10% of the sub-flocks)
were randomly selected and weighed. Due to changes in management in the end of week 7 – diet
was changed and the pullets were able to access the outdoor runs - we reported weight gain, feed
intake and feed conversion rate for the first period – week 1 to 7 and for the complete period in
which the infection dynamic was monitored – from week 1 to 14.

108
Statistical analysis
Data on feed intake, growth rate and feed conversion rate were analysed by one-way
ANOVA in SAS (version 9.2) (SAS Institute, 2000) where the statistical unit was the pen
average. The infection dynamics was analysed by a mixed procedure (PROC MIX) where the
statistical unit was the ln transformed value of each pen observation per week in 14 weeks,
according to the following model:
Yijk = μ + αi + βj +(αβ)ij + Ak(ij) + εijk
where Yijk = was the natural logarithm of the number of oocysts per gram of faeces (OPG) for
each pen in each week; μ = the mean value; αi = the treatment, i = 1,2,3,4,5,6,7 (Vaccination ; 3%
A. annua dried leaves from day 1; 3% A. annua dried leaves from day 30; 3% M. esculenta dried
leaves; A. vulgaris ethanolic extract in the drinking water before appearance of clinical
symptoms; A. vulgaris ethanolic extract in the drinking water after appearance of clinical
symptoms and A. annua ethanolic extract in the drinking water after appearance of clinical
symptoms ); βj = the weeks OPG were accounted; j = (1 to 14); (αβ)ij = the interaction treatment
× weeks; Ak = the random effect of plot number (1 to 14) and εijk = the residual; Week was
defined as repeated measurement and εijk was assumed to have a multivariate distribution where
observations from different treatments were uncorrelated while observations from different
weeks were assumed to have a toeplitz (Type= TOEP) correlation structure. P-values less than or
equal to 0.05 were considered statically significant.
RESULTS
Chemical composition of leaves and ethanolic extracts
Leaves and ethanolic extract of A. vulgaris showed no traces of bioactive sesquiterpenes
compounds. A. annua leaves contained high levels of artemisinin (1.03g/100g DM). They also
contained dihydroartemisinic acid (0.28g/100g DM) and artemisinic acid (0.04g/100g DM), all

109
with a standard deviation of 0.01. For the ethanolic extract of A. annua, high concentrations of
artemisinin (1.15mg/mL), dihydroartemisinic acid (0.41mg/mL) and artemisinic acid
(0.06mg/mL) were found. From the ORAC tests, dried leaves of M. esculenta showed high levels
of antioxidant capacity from two different replicate samples (827 µM Trolox equivalents (TE)
per gram) while the Napier grass showed comparatively smaller levels (174 µM TE/g).
Artemisia effects on performance
Pullets fed dried leaves of A. annua during the first 8 weeks showed lower body weight
gain (BWG) when compared to all other treatment combinations, which did not differ in growth
rate (Table 3). This lower body weight gain was maintained during the entire 14-week period.
The feed intake for this treatment was numerical lower in the first 7 weeks, but nevertheless feed
conversion rate was significantly impaired compared with pullets in the other treatments.
Infection dynamics
The OPG counts from pooled samples of each pen along the 14 weeks period were
significantly influenced by treatment (F6,8= 3.8; P< 0.05), week (F13,25 = 3462; P< 0.0001) and
the interaction week x treatment (F78,25 = 22; P<0.0001). The strong interaction observed
between treatment and week illustrates that the peaks for OPG counts appeared at different times
for the individual treatments (Figure 2).
The prevalence of coccidia infection in the vaccinated group increased from nil to 100%
in the first four weeks of pullet’s life (Figure 2A). Pullets in this treatment were challenged with
the available species in the commercial vaccine: Eimeria tenella, E. acervulina, E. maxima and E.
necatrix. This group was also infected with oocysts from E. praecox during early infection (1-7
weeks) (columns 13 and 14 - Figure 3). This was the only treated group that did not show an
infection peak at week 7 (Figure 2). After accessing the outdoor runs, pullets in this group were
challenged like the other groups and this can be observed in the dynamic of oocyst excretion
from weeks 9 to 13 (Figure 2).
The severity of coccidia infections in the studied farm is observed by interpreting the
infection dynamics from Figure 2 F representing the farmer routine (Group EAv2). Pullets were

110
most likely challenged from oocysts available in the pen at weeks 4-5 leading to an infection
peak approximately 2 weeks later, at week 7. Signals of blood on faeces were observed at week 8,
when treatment was supplemented in drinking water following the farmer routine.
An earlier infection was observed for the second group of pullets treated after appearance
of clinical symptoms (EAa1). Pullets were challenged in an early stage and a mean peak was
observed at week 5 (Figure 2G). In addition to the initial coccidial challenge, both groups treated
with ethanolic extracts after the appearance of clinical symptoms (EAv2 and EAa1) were
challenged at a late stage, after accessing the outdoor runs, observed by the small peaks reported
on week 11 (Figures 2F and 2G).
Pullets supplemented with 3% A. annua since one-day-old and pullets supplemented with
4 weeks of age showed a different infection dynamic compared to the other treatment
combinations. Infection peaks were observed in these two groups without clear patterns (Figures
2B and 2C). In addition, pullets supplemented with 3% A. annua were also challenged in the late
period, after accessing the chicken runs. Oocysts of Eimeria praecox (Lane 5 on Figure 3) were
coincidently found (Columns 1 and 2 on Figure 3) in both groups.
The species of Eimeria identified by the multiplex PCR analysis consisted of E.
acervuliva; E. tenella and E. mitis oocysts, which were the most common species during early
infection. Eimeria brunetti; E. maxima, E. preacox and E. necatrix were also found during early
infection, however less prevalent among the groups (Figure 3).
The mean OE was lower for the groups supplemented with 3% cassava leaves (Table 3).
Thus, cassava dried leaves supplemented in the diet during the first 8 days of pullet’s life
suppressed oocyst excretion (OE) to almost half of what was accounted for the vaccinated pullets
and less than one third of what was accounted for the pullets supplemented with A. annua from
week 4 to 8 (Aad30). By observing Figure 2D, pullets supplemented with 3% cassava dried
leaves showed smaller infection peaks at week 7 when compared to all other treatments in our
study.
Pullets in the group supplemented with the ethanolic extract of A. vulgaris via drinking
water (EAv1) before the appearance of clinical symptoms presented similar OE compared to the
cassava group but not different from the vaccinated group (Vac) and from the two groups
supplemented with ethanolic extracts after appearance of clinical symptoms (EAv2 and EAa1)
(Table 3). By observing the infection dynamics for all pullets in our study (Fig. 2), it is possible

111
to identify that at week 11, when a small infection peak is apparently observed in all groups at a
late infection, the EAv1 treatment (Figure 2E) showed nothing.
DISCUSSION
Dried leaves of Artemisia annua, supplemented at 3% of the dietary inclusion from day
one of pullets life, limited weight gain and feed conversion rate for the initial and total growing
periods under investigation (from 1 to 7 weeks and from 1 to 14 weeks). These results agree with
our previous findings where dried leaves of A. annua decreased feed intake and growth rates for
slow growing broilers in free range systems (Almeida et al., 2012). However, in our previous
study, the botanical supplementation resulted in a lower OE in broilers consuming A. annua for
two weeks before coccidia challenge, thus using a prophylactic strategy recommended for a
positive effect of artemisinin against at least two species of Eimeria (Allen et al. 1997). The
strategy of supplementing A. annua at two different stages in the current study was an attempt to
follow a similar approach. Nevertheless, in both cases in this study, pullets were challenged
before a reasonable period of artemisinin consumption what may have lead to the failure of
suppressing OE (Figures 2B and 2C).
The implications of these findings highlight the necessity of high hygiene procedures
when allocating new pullets to avoid early infections as their immunological responses are
highly influenced by the age of the chicks (Swinkels et al., 2006; 2007). In addition, it is
necessary to target on strategies for delivering herbal supplements to reduce severity of
coccidiosis without compromising feed intake as herbal products may often have unpalatable
components (Caygill and Mueller-Harvey, 1999). The post harvesting processes should also be
considered to preserve/eliminate different components of interest in the leaf biomass when
exploring potential herbal supplements (Ferreira and Luthria, 2010)
Based on the dynamics of the coccidial infections, the mean OE for the pullets receiving
herbal treatments rose to a maximum at week 7 (early infection) and were followed by a sharp
decline two to three weeks when animals were aged 8 to 10 weeks old. Subsequently, a smaller
peak on week 11 (late infection) was seen, probably reflecting the access to the outdoor area.
To our knowledge, this is the first evidence showing that supplementation of 3% cassava
dried leaves to pullets naturally infected with Eimeria spp. oocysts markedly suppressed OE.

112
Thus, the reduced number of oocysts for the pullets receiving cassava dried leaves suggests that
oocysts, initially ingested and latter excreted in the environment, were not as viable and infective
as oocysts from other experimental units, perhaps due to the presence of CT and other
antioxidant ingredients (Molan et al., 2003; Horigome et al., 1988).
In order to benefit from medicinal properties of plant components to improve animal
health and suppress parasite reproduction without interfering on growing capacity of the
chickens, we tested different delivering strategies based on the visionary knowledge of a
collaborating chicken producer. We showed that supplementing drinking water with an ethanolic
extract of Artemisia vulgaris 10 days before the expected appearance of the clinical symptoms of
the disease suppressed the oocyst output at a late stage (Figure 2E) leading this group to be
compared with pullets supplemented with cassava dried leaves (Table 3). Infective oocysts were
available in the pens what caused challenges at early stages. In this farm study, we hypothesize
that perhaps starting supplementation in the beginning of production instead of supplementing
before the farm chronology for the clinical symptoms, the herbal component (without artemisinin)
could have provided protection, suppressed OE at a higher degree by improving health status of
the flocks.
Chickens rapidly develop protective immunity after exposure to Eimeria spp. but none
stimulates cross-protection to other species (Tyzzer, et al., 1932). On this respect, oocysts of E.
praecox - the Eimeria species with shortest development – 83 to 90 hrs post infection
(McDougald et al., 1997) - were found in pens with pullets receiving A. annua in the diet at
different ages (Figures 2 B, 2 C and Figure 3). However, even E. praecox have been reported to
cause depression in growth (Long and Johnson, 1988), we cannot attribute its early infectivity as
the main cause of reduced weight gain in pullets consuming A. annua from the first day of life
(Aad1). Pullets in other pens that did not consumed A. annua (Figures 2 E and 2 G) had infection
with E. praecox (Figure 3, Lane 5) and performance attributes were not impaired (Table 3).
We evaluated the antioxidant capacity of cassava dried leaves, and they showed up to 50%
higher ORAC values than alfafa, a potential source of high antioxidants also for human
consumption (Xiea et al., 2008). In addition to antioxidant properties, cassava leaves are rich in
CT (Oni et al., 2011). Aqueous extracts of pine bark (Pinus radiata) was tested on sporulation of
coccidia oocysts and abnormalities in sporocysts exposed to two different dosages were reported
(Molan et al., 2009). They suggested that CT may act by penetrating the wall of the oocyst

113
during the sporulation thus damaging its cytoplasm (sporont). In addition, CT have also been
documented to inhibit endogenous enzyme activities (Horigome et al., 1988) leading to reduced
oocyst sporulation by enzyme inactivation cause by CT (Molan et al., 2003).
During the first 8 days of life, pullets in the Ca8d group consumed an approximated
amount of 30 g diet/head per day (based on DM). Assuming an average level of CT of 2.2 g/kg
DM of cassava dried leaves as previously presented by Oni et al., (2011), we estimated that
pullets consumed individually an average of 2 mg CT on a daily basis. In association with the
high ORAC values, the supplementation of cassava to the level of 3% leaded to 50-60%
suppression in oocyst excretion as compared to the Vaccinated group. However, it is unknown to
what degree the reduction in oocysts output observed in our study may positively impact the
epidemiology of the disease in a case of a higher infection level. Unfortunately, our study could
not benefit from comparisons with uninfected or untreated pullets as we have adapted the
experimental design with the farmer management and structure. Nevertheless, by observing
Figure 2F – representing the farmer´s routine and considering that blood on faeces was observed
after the main infection peak reported at week 7, the infection dynamic occurring in the studied
farm was mild perhaps subclinical. This low infection is maybe attributed to the size of the
flocks thus supposedly affecting performance at a late stage (De Gussem, 2007) in which our 14
weeks study could not capture.
It is believed that CT found in cassava and artemisinin found in A. annua may act against
coccidiosis by suppressing the parasite reproduction while A. vulgaris and other plants extracts
with antioxidant ingredients may contribute to a better equilibrium in the body anti-oxidant
defenses thus leading to a fast immunization. The supplementation of ethanolic extract of A.
vulgaris before expected appearance of clinical symptoms did not influence the main infection
peak at week 7. However, it contributed to suppress the oocyst output at a late stage (Figure 2E).
Harvesting the top leaves and removing stems of cassava plants to be included in the diet
was labour-intensive and may influence negatively on the yield of its main product, the tubers. In
the small scale perspective, those characteristics may restrict its use, unless the leaves are
collected, dried and stored after the tubers are harvested but it will certainly negatively influence
the levels of CT and also its antioxidant capacity.
Dietary inclusion of A. annua leaves may not work as a coccidiostat for the Isa Brown
pullets managed under organic systems, and challenged with coccidia oocysts at an early stage.

114
Supplementation of ethanolic extracts in drinking water sounds practical and promising with
regards to usefulness at commercial farm level. However, more research is necessary on mixing
herbal extracts containing artemisinin, condensed tannins and antioxidant ingredients delivered
via drinking water.
IMPLICATIONS
While the ethanolic extracts presented a limited effect, a daily supplementation of 3%
cassava dried leaves in the first 8 days of pullets life suppressed oocyst excretion (OE) by 50-60%
compared to a vaccinated group. Condensed tannins (estimated daily intake of 2 mg/pullet) may
perhaps affect oocyst infectivity thus reducing its capacity to proliferate in the environment
while antioxidants compounds may have boosted the host immune system by reducing the
effects of oxidative stress caused by coccidial infections. Further investigations targeting specific
species of coccidia, mixture of different bioactive compounds, times of supplementation and
delivery routes are warranted.
ACKNOWLEDGEMENTS
The authors wish to thank the group of farmers leaded by Romeu M. Leite from the
Yamaguishi farm for his visionary knowledge and technical support. Aarhus University and
SOAR - Research School for Organic Agriculture and Food Systems - are thanked for financing
our study. Dr Alda B. N. Madeira from the Institute of Biomedical Sciences from USP is thanked
for Eimeria speciation by multiplex PCR. Thanks are also due to Mr. Barry Harter for his
valuable help on HPLC-UV analysis.

115
REFERENCES
Abbas, R. Z., Colwell, D. D. and Gilleard, J., 2012. Botanicals: an alternative approach for the
control of avian coccidiosis. World Poultry Sci J, 68, 203-215.
Allen, P.C., Lydon, J. and Danforth, H. D., 1997. Effects of components of Artemisia annua on
coccidia infections in chickens. Poultry Sci, 76, 1156-1163.
Allen, P.C., Danforth, H. D. and Augustine, P. C., 1998. Dietary modulation of avian coccidiosis.
Int J Parasitol, 8, 1131-1140.
Almeida, G. F. d., Horsted, K., Thamsborg, S. M., Kyvsgaard, N. C., Ferreira, J. F. S. and
Hermansen, J. E., 2012. Use of Artemisia annua as a natural coccidiostat in free-range
broilers and its effects on infection dynamics and performance. Vet Parasitol, 186, 178-187.
Arab, H.A., Rahbari, S., Rassouli, A., Moslemi, M.H. and Khosravirad, F.D.A., 2006.
Determination of artemisinin in Artemisia sieberi and anticoccidial effects of the plant
extract in broiler chickens. Trop Anim Health Prod, 38, 497-503.
Barry, T. N., Manley, T. R. and Duncan, S. J., 1986. The role of condensed tannin in the
nutritional value of Lotus pedunculatus for sheep: Site of carbohydrate and protein
digestion as influenced by dietary reactive tannin concentration. Brit J Nutr, 55, 123-137.
BRASIL, 2003. Federal Law n. 10.831. Organic agriculture and sets other provisions. Available
in English at: http://www.prefiraorganicos.com.br/media/45792/lei_10.831_ingles.pdf.
Access Jan 2012.
BRASIL, 2007. Federal Decree n. 6.323. Regulates Law No. 10.831 on organic agriculture and
sets other provisions. Available in English at:
http://www.prefiraorganicos.com.br/media/45812/decreto_6.323_ingles.pdf. Access Jan
2012. Access Jan 2012.
BRASIL, 2011. Federal Normative Instruction n. 46. Regulates the vegetable and animal
production in organic systems. Available in Portuguese at:
http://www.in.gov.br/visualiza/index.jsp?data=07/10/2011&jornal=1&pagina=4&totalAr
quivos=248 Access Jan 2012.
Brenes, A. and Roura, E., 2010. Essential oils in poultry nutrition: main effects and modes of
action. Anim Feed Sci Tech, 158, 1-14.
Caygill, J. C. and Mueller-Harvey, I. (Eds.), 1999. Secondary Plant Products - Antinutritional

116
and beneficial actions in animal feeding. Nottingham University Press, Nottingham, UK,
129 pp.
De Gussem, M., 2007. Coccidiosis in poultry: Review on diagnosis, control, prevention and
interaction with overall gut health. In: Proceedings of the 16th European Symposium on
Poultry Nutrition. pg 253–261. World’s Poultry Science Association, Beekbergen, the
Netherlands.
Del Cacho, E., Gallego, M., Francesch, M., Quílez, J. and Sánchez-acedo, C., 2010. Effect of
artemisinin on oocyst wall formation and sporulation during Eimeria tenella infection.
Parasitol Intern, 4, 506-511.
Duke, J. A., Godwin, M. J. B., du Cellier, J., and Duke, P. N. K., 2002. Handbook of Medicinal
Herbs, 2nd edn. CRC Press, Washington, DC.
EU, 1991. Commission regulation N.1538/91 of 15 June 1991 introducing detailed rules for
implementing regulation 1906/90 on certain marketing standards for poultry. Official
Journal of the European Communities (L143), 11-22.
EU, 2007. Commission regulation N.834/07 of 28 June 2007 on organic production and labeling
of organic products and repealing Regulation (EEC) No 2092/91. Official Journal of the
European Communities (L189), 1-23.
Fanatico, A., 2006. Organic poultry production in the United States. ATTRA - National
Sustainable Agriculture Information Service. Available at: http://www.attra.org/attra-
pub/PDF/coccidiosis.pdf. Access Jan 2012.
Fernandez, S., Pagotto, A.H., Furtado, M.M., Katsuyama, A. M., Madeira, A.M.B.N and Gruber,
A., 2003. A multiplex PCR assay for the simultaneous detection and discrimination of the
seven Eimeria species that infect domestic fowl. Parasitol, 127, 317-325.
Ferreira, J. F. S. and Gonzales, J. M., 2009. Analysis of underivatized artemisinin and related
sesquiterpene lactones by high-performance liquid chromatography with ultraviolet
detection. Phytochem Analysis, 2, 91-97.
Ferreira, J. F. S. and Luthria D. L., 2010. Drying Affects Artemisinin, Dihydroartemisinic Acid,
Artemisinic Acid, and the Antioxidant Capacity of Artemisia annua L. Leaves. J Agr
Food Chem, 58, 1691-1698.
Liu, C., Zhao, Y. and Wang, Y., 2006. Artemisinin: current state and perspectives for
biotechnological production of an antimalarial drug. Appl Microbiol Biot, 72, 11-20.

117
Long P. L. and Rowell, J. G., 1975. Sampling broiler house litter for coccidial oocysts. British
Poultry Sci, 16, 583-592.
Long, P. L. and Johnson, J. K., 1988. Eimeria of American chickens: characteristics of six
attenuated strains produced by selection for precocious development. Avian Pathol, 17,
305-314.
Herrmann, R. and Roder, C., 1995. Does food consumption converge internationally?:
measurement, empirical tests and determinants. Eur Rev Agric Econ, 22, 400 - 414.
Horigome, T., Kumar R. and Okamoto, K., 1988. Effects of condensed tannins prepared from
leaves of fodder plants on digestive enzymes in vitro and in the intestine of rats. Brit J
Nutri, 60, 275-285.
Mannan, A., Ahmed, I., Arshad, W., Asim, M. F., Qureshi, R. A., Hussain, I. and Mirza, B.,
2010. Survey of artemisinin production by diverse Artemisia species in northern Pakistan.
Malaria Journal, 9, 310. http://dx.doi.org/10.1186/1475-2875-9-310.
Marie-Magdeleine, C., Mahieu, M., Philibert, L., Despois, P., Archimède, H., 2010. Effect of
cassava (Manihot esculenta) foliage on nutrition, parasite infection and growth of lambs.
Small Ruminant Res, 93, 10-18.
McDougald, L. R., Fuller, L. and Mattiello, R. 1997. A Survey of Coccidia on 43 Poultry Farms
in Argentina. Avian Dis, 41, 923-929.
Molan, A. L., Meagher, L. P., Spencer, P. A., and Sivakumaran, S., 2003. Effect of flavan-3-ols
on in vitro egg hatching, larval development and viability of infective larvae of
Trichostrongylus colubriformis. Intern J Parasitol, 33, 1691-1698.
Molan, A. L., Liu, Z., and De, S., 2009. Effect of pine bark (Pinus radiata) extracts on
sporulation of coccidian oocysts. Folia Parasit. 56, 1-5.
Naidoo, V., McGaw, L.J., Bisschop, S.P.R., Duncan, N. and Eloff, J.N., 2008. The value of plant
extracts with antioxidant activity in attenuating coccidiosis in broiler chickens. Vet
Parasitol, 153, 214-219.
Netpana, N., Wanapat, M., Poungchompu, O., Toburan, W., 2001. Effect of condensed tannins in
cassava hay on fecal parasitic egg counts in swamp buffaloes and cattle. In: Preston, T.R.,
Ogle, B., Wanapat, M. (Ed.), Proceedings of the International Workshop on Current
Research and Development on Use of Cassava as Animal Feed, Khon Kaen University,
Thailand. Available at: http://www.mekarn.org/procKK/wana1.htm. Access Nov 2011.

118
Oni, A. O., Onwuka, C. F. I., Arigbede, O. M., Anele, U. Y., Oduguwa, O. O., Onifade, O. S.
and Tan, Z. L., 2011. Chemical composition and nutritive value of four varieties of
cassava leaves grown in South-Western Nigeria. J Anim Physiol Anim Nutri, 95, 583-
590.
Orengo, J., Buendía, A. J., Ruiz-Ibáñez, M. R., Madrid, J., Del Río, L., Catalá-Gregori, P.,
García, V. and Hernández, F., 2012. Evaluating the efficacy of cinnamaldehyde and
Echinacea purpurea plant extract in broilers against Eimeria acervulina. Vet Parasitol,
185, 158-163.
Ravindran, V., 1991. Preparation of cassava leaf products and their use as animal feed. In: Roots,
tubers, plantains and bananas in animal feeding. APHP Paper 95: 81-98, FAO: Rome.
Available at:
http://www.fao.org/WAICENT/FAOINFO/AGRICULT/AGA/AGAP/FRG/AHPP95/95-
111.pdf. Access Feb 2012.
Ross, E. and Enriquez, F. Q., 1969. The nutritive value of cassava leaf meal. Poultry Science,
48, 846-853.
SAS Institute, 2000. SAS Online Doc®, Version 9.2. SAS Institute, Cary, NC.
Seng, S. and Rodriguez, L., 2001. Foliage from cassava, Flemingia macrophylla and bananas
compared with grasses as forage sources for goats: effects on growth rate and intestinal
nematodes. Livest Res Rural Develop, 13, 1-6.
Swinkels, W. J., Post, J., Cornelissen, J. B., Engel, B., Boersma, W. J. and Rebel, J. M., 2006.
Immune responses in Eimeria acervulina infected one-day-old broilers compared to
amount of Eimeria in the duodenum, measured by real-time PCR. Vet Parasitol., 138:
223–233.
Swinkels, W. J., Post, J., Cornelissen, J. B., Engel, B., Boersma, W. J. and Rebel, J. M., 2007.
Immune responses to an Eimeria acervulina infection in different broilers lines. Vet
Immunol. Immunop., 117: 26–34.
Sokerya, S., Waller, P. J., Ledin, I. and Höglund, J., 2007. The effects of short-term feeding of
fresh cassava (Manihot esculenta) foliage on gastrointestinal nematode parasite infections
in goats in Cambodia. Trop Biomed, 24, 47-54.
Temraz, A. and El-Tantawy, W. H., 2008. Characterization of antioxidant activity of extract from
Artemisia vulgaris. Pakistan J Pharm Sci, 21, 321-326.

119
Tigno, X.T. and Gumila, E., 2000. In vivo microvascular actions of Artemisia vulgaris L. in a
model of ischemia-reperfusion injury in the rat intestinal mesentery. Clin Hemorheol
Micro, 23, 159-165.
Tigno, X.T., De Guzman, F. and Flora, A. M., 2000. Phytochemical analysis and hemodynamic
actions of Artemisia vulgaris L. Clin Hemorheol Micro, 23, 167-175.
Tyzzer, E. E., Theiler, H., and Jones, E. E.,1932. Coccidiosis in gallinaceous birds. II. A
comparative study of species of Eimeria of the chicken. Am J Hyg, 15, 319-393.
Ungemach, F. R., Müller-Bahrdt, D. and Abraham, G., 2006. Guidelines for prudent use of
antimicrobials and their implications on antibiotic usage in veterinary medicine. Int J
Med Microbiol, 296, 33-38.
Wang, M.L., Suo, X., Gu, J.H., Zhang, W.W., Fang, Q. and Wang, X., 2008. Influence of grape
seed proanthocyanidin extract in broiler chickens: effect on chicken coccidiosis and
antioxidant status. Poult Sci, 87, 2273-2280.
Williams, R.B., 2002. Fifty years of anticoccidial vaccines for poultry, 1952-2002. Avian
Dis, 46, 755-802.
Wright, C.W., 2005. Traditionals antimalarials and the development of novel antimalarial drugs.
J Ethnopharmacol, 100, 67-71.
Xiea, Z., Huangb, J., Xua, X. and Jin, Z., 2008. Antioxidant activity of peptides isolated from
alfalfa leaf protein hydrolysate. Food Chem, 111, 370-376.
Zheng, W. and Wang, Y. 2001. Antioxidant activity and phenolic compounds in selected herbs.
J Agric Food Chem, 49, 5165-5170.

120
Table 1 – Description of treatments, supplementation strategy of the herbal product and age of the pullet’s when treatments were
performed.
Group
Treatment
Supplementation
Strategy*
Period of
Treatment**
Dosing period (age of the
pullets in days)***
Vac Vaccination1 Drinking water Before 3-8
Aad1 A. annua dried leaves 3% of diet Before 1-56
Aad30 A. annua dried leaves 3% of diet Before 30-56
Ca8d M. esculenta dried leaves 3% of diet Before 1-8
EAv1 A. vulgaris ethanolic extract Drinking water Before 48-58
EAv2 A. vulgaris ethanolic extract2 Drinking water After 63-70
EAa1 A. annua ethanolic extract Drinking water After 65-72
*Diet supplemented was prepared by reducing 3% of the basal starter diet (based on its weight) and substituting the same
amount by dried leaves of the plant materials.
** Type of treatment consisted of supplementing the botanical coccidistat before or after expected appearance of clinical
symptoms of the disease.
***Interval between the beginning and conclusion of the treatment administrated based on the age of the pullets. 1 Livacox® Q (Merial Animal Health Ltd. Brazil) -positive control; 2 farmer’s routine.

121
Table 2 – Ingredients and diet composition supplied to the pullets from the beginning to the end
of the experiment, when chicks were 14 weeks old.
Dietary ingredients, % of diet Starter diet Growth diet
Maize grains 53.5 64.2 Soybean meal (44.5%) 38.5 26.7 Quinoa 1.0 0.6 Wheat meal (9.8%) 0.0 4.0 PMIX_growth_P3251 0.4 0.4 Yes-Minerals_Fix3 0.2 0.2 Yes-Minerals_AVES3604 0.2 0.2 Fosfocalcio 205 2.0 2.0 Basaltic gravel 0.0 0.1 Limestone, pulverized 1.0 1.6 Soybean oil (kg) 2.8 0.0 Sodium bicarbonate 0.4 0.4 Chemical composition (% DM) Dry Matter 91.2 89.2 Ash 10.1 6.1 Crude protein (N*6,25) 17.1 19.2 Crude fiber 3.2 3.5 Ether extract 4.9 5.0 NDF (Neutral Detergent Fiber) 14.6 19.3 ADF(Acid Detergent Fiber) 4.3 5.8 Lignin 0.4 1.1 Metabolizable energy (MJ/kg DM) 15.9 16.7 1Supplying per kg of diet: vit. A, 11,500 IU, vit. D3, 6,000 IU, vit. E, 10 mg, vit. K3, 1.6 mg, vit. B1, 1.4 mg, vit. B2, 4 mg, vit. B6, 2 mg, vit. B12, 8 µg, niacin, 25 mg, panthotenic acid, 11 mg, folic acid, 4 mg, choline chloride, 28 mg, methionine, 99 mg, Cu, 8 mg, Fe, 49 mg, Mn, 62 mg, Zn, 49 mg, I, 1.2 mg, Se, 0.2 mg, antioxidant, 12.3 mg. 2Supplying per kg of diet: vit. A, 12,100 IU, vit. D3, 6,000 IU, vit. E, 5 mg, vit. K3, 1.6 mg, vit. B2, 3 mg, vit. B12, 8 mcg, niacin, 20 mg, panthotenic acid, 5 mg, choline chloride, 28 mg, methionine, 680 mg, Cu, 8 mg, Fe, 49 mg, Mn, 62 mg, Zn, 49 mg, I, 1.2 mg, Se, 0.2 mg. 3Supplying per kg of diet: β-glucan, 0.36 g, Al2O3, 0.12 g, Ca(OH)2, 14 mg, SiO2, 0.22 g. 4Supplying per kg of diet: Cu amino acid Chelate, 20 mg, Fe amino acid Chelate, 0.14 g, Mn amino acid Chelate, 0.16 g, crude protein, 0.22 g, selenium protein, 0.64 mg, Zn amino acid Chelate, 0.22 g. 5Supplying in % per kg product: phosphorus, 20%, soluble phosphorus in citric acid 2%, 90%, calcium, 21%, fluor, 0.20%, max humidity, 5%.

122
Table 3 – Effect of treatments on performance (body weight gain, feed intake and feed conversion rate - FCR) and on the mean oocyst output along the 14 weeks
of pullets life (back transformed values from least square means of ln-transformed counts are presented).
P
value
Treatment
Attribute Vac Aad1 Aad30 Ca8d EAv1 EAv2 EAa1
BWG ¹ 1-7 weeks 0.05 456±06a 331±11b 405±10a 426±41a 461±33a 448±18a 432±20a
FI² 1-7 weeks 0.30 1.4±0.1 1.2±0.1 1.4±0.1 1.4±0.1 1.4±0.1 1.3±0.1 1.5±0.1
FCR³ 1-7 weeks 0.03 3.2±0.1a 3.9±0.0b 3.5±0.1ab 3.3±0.3a 3.1±0.2a 3.0±0.0a 3.4±0.0a
BWG 1-14 weeks 0.06 1151±11 993±23 1062±17 1046±04 1079±30 1121±19 1057±40
FI 1-14 weeks 0.08 5.9±0.0 6.3±0.1 5.9±0.1 5.8±0.3 5.7±0.1 5.5±0.1 6.0±0.0
FCR 1-14 weeks 0.01 5.1±0.0b 6.3±0.2a 5.5±0.2b 5.6±0.2b 5.3±0.2b 4.9±0.3b 5.8±0.2ab
k OPG 4 0.04 16.8±1.5ab 18.5±1.7ab 27.6±2.5a 7.4±0.7c 12.4±1.1bc 12.6±1.1bc 16.6±1.5abc
a,b Values in a same row marked with different letters are significantly different (p<0.05). Values are presented by the LS means ± SEM, n = 2 replicates. 1BWG = Body weight gain (grams) for the specified period. 2FI = Feed Intake (Kg diet/period) for the specified period. 3FCR = Kg feed supplied/kg weight gain for the specified period. 4k OPG = Mean oocyst count (x 1000) for the 14 weeks period.

123
Figure title and Legends
Figure 1: Time line and description of treatments supplemented to the pullets.
Treatments consisted of: Vaccination (Vac - positive control), 3% A. annua dried
leaves from day 1 (Aad1); 3% A. annua dried leaves from day 30 (Aad30); 3% M.
esculenta dried leaves (Ca8d); A. vulgaris ethanolic extract in the drinking water
before appearance of clinical symptoms (EAv1); A. vulgaris ethanolic extract in the
drinking water after appearance of clinical symptoms (EAv2 - farmer’s standard
routine) and A. annua ethanolic extract in the drinking water after appearance of
clinical symptoms (EAa1).

124
Figure 2: Effect of treatments on coccidia oocyst excretion during the first 14 weeks of the
pullets’ life.
Oocyst excretion was based on weekly fresh samples collected from the ground of each of two
replicate pens per treatment (see Table 1 for list of explanations of treatments).

125
Figure 3: Agarose gel electrophoresis of multiplex PCR products using DNA samples*
Eimeria acervulina (Lane 1), E. brunetti (Lane 2), E. tenella (Lane 3), E. mitis (Lane 4), E.
praecox (Lane 5), E. maxima (Lane 6), E. necatrix (Lane 7). Molecular size markers are L-
Ladder 100pb Invitrogen. Species found for each experimental unit are presented from columns 1
to 14. Column 15 is the positive control (DNA from the 7 Eimeria species) and column 16 is the
negative control.
*Oocysts were purified for speciation from pooled samples collected at weeks 3, 4, 5 and 6.
Columns 13 and 14 represent replicates 1 & 2 of the Vaccination group (Vac - positive control).
Columns 1 and 5 represent replicates 1 & 2 of the 3% A. annua dried leaves from day 1 (Aad1).
Columns 2 and 9 represent replicates 1 & 2 of 3% A. annua dried leaves from day 30 (Aad30).
Columns 3 and 7 represent replicates 1 & 2 of 3% M. esculenta dried leaves (Ca8d).
Columns 4 and 11 represent replicates 1 & 2 of A. vulgaris ethanolic extract supplemented in the
drinking water before the appearance of clinical symptoms of coccidiosis (EAv1).
Columns 6 and 8 represent replicates 1 & 2 of A. vulgaris ethanolic extract in the drinking water
after the appearance of clinical symptoms of coccidiosis (EAv2 - farmer’s standard routine).
Columns 10 and 12 represent replicates 1 & 2 of A. annua ethanolic extract in the drinking water
after the appearance of clinical symptoms of coccidiosis (EAa1).

126
Paper IV
The effects of combining Artemisia annua and Curcuma longa ethanolic
extracts in broilers challenged with infective oocysts of Eimeria acervulina and
E. maxima
Almeida, G. F. d.; Thamsborg, S. M.; Madeira, A. M. B. N.; Ferreira, J. F. S.; Magalhães, P. M.;
Horsted, K. and Hermansen, J. E. (2012) Manuscript submitted to Parasitology
Published as original manuscript with permission of Parasitology

127
The effects of combining Artemisia annua and Curcuma longa ethanolic extracts in broilers
challenged with infective oocysts of Eimeria acervulina and E. maxima
Gustavo F. d. Almeidaa*; Stig M. Thamsborgb; Alda M. B. N. Madeirac; Jorge F. S. Ferreirad;
Pedro M. Magalhãese; Luiz C. Demattê Filhof; Klaus Horsteda; John E. Hermansena
a Department of Agroecology, Faculty of Sciences and Technology, Aarhus University. Research
Centre Foulum, Blichers Allé 20, P.O. Box 50 DK-8830 Tjele, Denmark.
b Danish Centre for Experimental Parasitology, Faculty of Life Sciences, University of
Copenhagen, Dyrlægevej 100, DK-1870 Frederiksberg C, Denmark.
c Department of Parasitology, Institute of Biomedical Sciences, University of São Paulo, Av.
Prof. Lineu Prestes, 1374. São Paulo, SP, Brazil.
d USDA-ARS, US Salinity Laboratory, 450 W. Big Springs Rd., Riverside, CA 92507-4617.
e Chemical, Biological and Agricultural Pluridisciplinary Research Centre (CPQBA), University
of Campinas – UNICAMP, P.O. Box 6171, BR-13081-970 Campinas, SP, Brazil.
f Department of Economy and Rural Sociology, Luiz de Queiroz College of Agriculture
(ESALQ), University of São Paulo, Avenida Pádua Dias 11, Piracicaba, SP, Brazil. * Corresponding author: Gustavo Fonseca de Almeida; Tel. +45 8715 4757; Fax. +45 8715 4798.
E-mail address: [email protected]

128
SUMMARY
Due to an increasing demand for natural products to control parasites in animal production we
investigated the effects of combining ethanolic extracts of Artemisia annua and Curcuma longa
supplemented in drinking water to prevent coccidosis in broilers. Three different dosages of the
herbal mixture were compared with a negative control (uninfected), a positive control (infected
and untreated), chemical coccidiostats (nicarbazin+narazin and later salinomycin), vaccination,
and a product based on oregano. Differences in performance (weight gain, feed intake, and feed
conversion rate), mortality, gross intestinal lesions and oocyst excretion were investigated.
Broilers supplemented with chemical coccidiostats presented superior performance attributes
compared to all other groups. A dose-dependent response for scoring of lesions was observed for
the herbal extracts. In average, broilers supplemented with the two highest dosages of the
mixture presented intermediate scores, higher than broilers supplemented with coccidiostats but
less than broilers supplemented with vaccination, oregano and negative controls. A trend for
lower mortality (p=0.08) in the last part of growing period (23-43d) was also observed in broilers
supplemented with the highest dosage when compared to broilers supplemented with chemical
coccidiostats. In conclusion, the delivery strategy is practical for implementation at farm level
but further studies on dose levels and modes of action are needed.
Keywords: Broiler; Natural anti-protozoa drugs; coccidiostats; drug combination; plant extract;
herbal medicine; coccidiosis.

129
INTRODUCTION
Coccidiosis is a very important parasitic disease in the broiler industry (Shirley et al.,
2005) and it is caused by highly host-specific protozoan parasites belonging to the genus Eimeria
(Williams, 1999). Parasites multiply in the intestines damaging gut tissues which, in turn,
reduces feed intake and absorption of nutrients (Morris et al., 2007). Infections caused
simultaneously by E. acervulina, E. maxima, and E. tenella are frequently diagnosed in intensive
poultry farming systems (McDougald et al., 1997), with their control usually at a higher priority
over the other Eimeria species affecting broilers (Shirley et al., 2005). The disease is primarily
controlled by medication and a range of limitations have been reported in the past few years
(Martin et al., 1997). Underdose of anticoccidials used in feed could lead to resistance
(Daugschies et al. 1998) and the extensive use of drugs has led to the development of resistance
to such products (Chapman, 1993), which also could be phased out as feed additives before
January 1st 2013 for public health and food safety reasons (COM, 2008).
Research has been carried out to find effective, but non-pathogenic, vaccines (Lillehoj
and Trout, 1993) to be used in substitution to anticoccidial products. In addition, the use of plants
and extracts as therapeutics might be an option (Akhtara et al., 2012). The substitution of
ionophore drugs is unlikely to be matched by a single approach, but a comprehensive
combination of strategies could help to reduce the prevalence of coccidiosis in intensive
production systems (Pinard-van der Laan et al., 1998).
In organic systems, the use of live attenuated vaccines (Williams, 2002) is a useful option
available in the market but these are expensive to produce (Abbas et al., 2012). Cost effective
alternatives are demanded for safer and effective strategies to control the disease, and several
candidate medicinal plants have been investigated (Naidoo et al., 2008; Wang et al., 2008). New
natural ingredients to substitute chemicals products, e.g. plants, extracts and their combinations
are believed to play an important role in the near future because they are usually residue free and
well accepted by consumers (Orengo et al., 2012). For example, Artemisia annua was earlier
suggested to control Eimeria acervulina in prophylactic supplementation strategies before
coccidial challenges (Allen et al., 1997). However, different levels of inclusion of artemisinin
(main bioactive component of A. annua) in feed and oral supplementation had no effect against E.
maxima, suggesting a species-specific mode of action (Arab et al., 2006; Allen et al., 1997).

130
Another relevant plant with known anti-protozoa properties (Shahiduzzaman et al., 2009)
is the spice turmeric Curcuma longa, suggested as a potential candidate to treat human malaria
(Cui et al., 2007; Reddy et al., 2005). In Pakistan, small farmers supplement turmeric powder as
a feed additive for the control of coccidiosis in broilers (Abbas et al., 2012). Allen et al. (1998)
reported reduction on intestinal lesions and suppression in oocyst excretion when 1% dietary
inclusion was available to broilers challenged with E. maxima occysts. The bioactive ingredient
responsible for efficacy against the parasites is curcumin (diferuloylmethane), a phenolic
compound with high antioxidant (Subramanian et al., 1994), anti- inflammatory (Huang et al.,
1997), anti-tumor properties (Rao et al., 1995), and found in C. longa roots in concentrations
ranging from 1 to 5% (Conney et al., 1991). Souza and Glória (1998) screened a range of
samples from important commercial sites in Brazil and reported an average concentration of 4%
w/w curcumin in the powder.
Artemisinin was reported to have a rapid metabolism in vivo (Chen et al., 2009) with low
stability and fast decomposition following ingestion (Klaiman, 1985). Curcumin has been tested
as an attractive partner for artemisinin in combination therapy against malaria as the molecule
has a short half-life similar to artemisinin (Nandakumar et al., 2006). With similar
pharmacokinetics an optimum protection in a resistance prevention perspective may be achieved
(Nosten and White, 2007).
With reported anti-protozoan activity of both artemisinin and curcumin, we hypothesized
that the combination of these ingredients available in A. annua and C. longa herbal extracts
could provide synergistic effects and act against dual infections caused by E. maxima and E.
acervulina oocysts in fast growing broilers, similar to chemical coccidiostats.
Regarding strategies for delivering herbal extracts to broilers, 3% dried leaves of A.
annua was supplemented in the feed three weeks before a challenge with E. acervulina and E.
maxima (Almeida et al., 2012). Feed intake was impaired supposedly due to unpalatable
components available in the A. annua leaves even though oocyst excretion was suppressed by
60-70% (Almeida et al., 2012). In another study comparing different strategies of delivery of A.
annua to chickens naturally infected with Eimeria spp., supplementation of ethanolic extracts via
drinking water, as recommended by an organic farmer, did not impair feed intake but the effect
on oocyst excretion was limited (Almeida et al., unpublished data).

131
On this background, this study was undertaken to evaluate the effect of three different
dose levels of herbal extracts of artemisinin:curcumin on avian coccidiosis (intestinal lesions,
oocyst excretion and performance attributes). The extracts were administered in the drinking
water and compared to appropriate controls, to broilers supplemented with chemical products
(representing the conventional production system), vaccinated or supplemented with a product
based on oregano, thus representing alternative strategies available in the market to control
coccidiosis in organic production systems.
MATERIALS AND METHODS
Experimental design and broilers
The study was carried out at the experimental stable at Korin Agricultural Ltd.
(www.korin.com.br) located in the municipality of Ipeúna, Sao Paulo, Brazil (22° 24′ S, 47° 41′
W). On 15th of August 2011, one-day-old male broilers (Cobb 500S, n=1440) previously
vaccinated in hatchery against Marek, fowlpox, and Gumboro were randomly allocated to 48
pens distributed in eight treatment combinations with six replicates of 30 broilers totalizing 180
broilers per treatment. Three different dose levels of a combination A. annua and C. longa
ethanolic extracts (LOW; MEDIUM and HIGH) supplemented in drinking water were compared
with a negative control (uninfected, untreated), a positive control (infected and untreated),
treatment with chemical coccidiostats (see next section), vaccination and a product based on
oregano. Table 1 describes the treatments, the supplementation strategies, doses, age and
duration of treatments. Each pen was 3.0 m² (1.5 x 2 m) with food and water supplied ad libitum.
All pens were cleaned and disinfected two days before the beginning of the experiment
and high standards of hygiene were followed during the trial. All practical handling was
performed from outside the 48 pens in lateral corridors and the pens were isolated from each
other by the use of PVC walls. When it was necessary, technicians were trained in using
disposable plastic bags in their shoes to avoid contamination. Broilers were raised on concrete
floor with wood shavings until they were 43 days of age, when the experiment was terminated.
Broilers were raised in the comfort range temperature and humidity recommended by the strain

132
manual and the experimental protocols evolving broilers were in accordance with international
guidelines for animal care and health and with the Brazilian legislation.
Experimental diets, water supply and oral administration of herbal extracts
The feeding program in our study followed the routine of farmers linked to the
collaborative broiler company: initial (1-8 days), growth (9-22 days), fattening I (23-29 days),
fattening II (30-36 days old) and final (37-43 days). Broilers in the control groups (positive - T1
and negative – T4) were supplemented with basal diet without any other type of supplementation.
Nicarbazin + narazin (Maxiban®) 50 ppm was supplemented from 1 to 22 days of age and
salinomycin (Coxistac®) 66 ppm supplemented from 22 to 39 days of age in diet for broilers
receiving the chemical coccidostats (T3). Livacox vaccine at recommended doses (Merial Ltd.
Brazil) was administrated to the broilers in the same day broilers arrived from hatchery by the
use of a movable drinker allocated in each replicated pen (T2). Broilers in the group T5 were
supplemented with a product based on oregano extract with carvacrol + timol (Regano®) and
broilers in T6, T7 and T8 received the herbal mixtures via drinking water since the first day of
life.
Eight water tanks (one tank per treatment) with capacity for 160 L were placed in the top
of the barn and water was distributed by gravity to each replicated pens (n=6 per treatment) by
the use of plastic hoses. Every second day from the beginning of the trial, water from the tanks
with the mixture of herbal extracts, also including T5, was replaced. Treatments were prepared
fresh daily to avoid any confounding factor related to possible instability or oxidation of the
herbal components.
The herbal extracts of A. annua and C. longa were prepared at CPQBA - University of
Campinas - using the same extraction procedure. In summary, in two 1 L Erlenmeyer flasks
wrapped with aluminium foil to prevent oxidation and degradation by light, 100 g of dried leaves
or 100 g curcuma powder were mixed with 1 L ethanol 70% (700 mL ethanol 97° + 300 mL
distilled water). The mixtures were kept in a dark room with controlled temperature (22°C) and
homogenized with circular movements twice per day for 20 days. The contents were filtered by
using paper filters and transferred to 100mL flasks with dropping glass dispensers. Flasks were
properly identified and stored in the fridge until needed.

133
One week before the trial, flasks with the Ethanolic herbal extracts of A. annua and C.
longa were transported to the experimental station at Korin Agropecuária Ltd. and kept in the
fridge before its use. One day before allocating the chicks to the pens, the ethanolic concentrated
solutions were mixed with 155 liters tap water using the same volume depending on the dosage:
(1.5 mL of each ethanolic extract in T6, 2.5 mL for T7 and 3.6 mL for T8). The content of active
ingredients was estimated by HPCL according to Ferreira and Gonzales (2009). 1.15mg
artemisinin/mL was found for the A. annua ethanolic extract (unpublished data) while from
proximal calculations, the ethanolic extract of C. longa had an estimated concentration of 4.6 mg
curcumin/mL. In accordance to relevant literature on curcumin and artemisinin contents in raw
materials and in herbal ethanolic extracts (Souza and Gloria, 1992; Ferreira et al., 2011
respectively) we estimated that the three dosages supplemented represented a 1:4 ratio of
artemisinin:curcumin (Table 1).
Transmission by contact and monitoring strategy
At day 8 three randomly chosen broilers per pen (n= 18 per treatment) were artificially
inoculated by gavage with a 1 mL distillated water suspension containing 5x104 E. maxima +
2x105 E. acervulina sporulated oocysts. These broilers were nominated "seeders" and were
raised with contemporary broilers with the aim of transmitting the disease by contact (Velkers et
al., 2010). “Seeders” were removed from the pens nine days post inoculation and harvested
(cervical dislocation) for scoring of lesions (n= 5 per treatment) following Johnson and Reid
(1970) and supported by illustrations provided by Anon (1990). Results of scoring of lesions are
presented by the mean (± SD) for lesions caused by E. acervulina (duodenum), E. maxima (mid
part of small intestine) and averaged for the total lesions observed.
To assess infection dynamics spread by the “seeders”, five birds were randomly selected
in each pen and nominated as “tracers”. Twice weekly starting three days post inoculation of
“seeders”, the “tracers” were placed in communitarian cages (one cage per pen) during 15
minutes for pooled samplings of fecal samples. In total, eight pooled fecal samples per pen were
examined during the trial. In the same day of faeces collection, pooled samples (n= 48) were
analysed by a modified McMaster technique using saturated NaCl solution with 50% glucose

134
monohydrate as flotation fluid with a sensitivity of 20 oocysts per g of faeces (OPG) (Almeida et
al., 2012). Results are presented as the mean ± SEM of oocyst excretion (OE) for each treatment.
Performance attributes
Consumption of feed per pen was monitored and recorded every week. Individual body
weight of the same ten contemporary broilers in each pen (not “seeders” or “tracers”)
(n=60/treatment) was taken every week. We estimated performance attributes by considering the
mean body weight at 24 hours post hatch, 22 days-old, and when broilers were 43 days of age
before slaughter. With this strategy, we could estimate body weight gain, daily gain, feed intake
and feed conversion rate for two main periods - before and after expected infection and for the
total experiment (1-22d; 23-43d and 1-43 days of age). Results of performance attributes are
presented as the mean ± SEM and averaged for each treatment combination.
Statistical Analysis
Broilers in our study were allocated to pens in a completely randomized design.
Categorical data on lesion score was firstly analysed by linear regression to identify any possible
effects attributed to different dosages and followed by multiple range tests. Data on mortality
were analysed by the Chi-square test. The infection dynamics was analysed by a mixed
procedure (PROC MIX) in SAS (SAS, 2000) where the statistical unit was the logarithmical
transformed value for each OPG observation according to the following model:
Yijk = μ + αi + βj +(αβ)ij + Ak(ij) + εijk
where Yijk = was the natural logarithm of the number of oocysts per gram of faeces (OPG) for
each pen in each sampling date; μ = the mean value; αi = the treatment, i = 1,2,3,4,5,6,7,8
(Uninfected negative control; Vaccinated, infected; Coccidiostat, infected; Untreated, Infected;
Regano®, Infected; Dose 1 (LOW) of herbal extracts, Infected; Dose 2 (MEDIUM) of herbal
extracts, Infected; Dose 3 (HIGH) of herbal extracts, Infected); βj = the OPG sampling date; j =
(1 to 8); (αβ)ij = the interaction treatment × OPG sampling date; Ak = the random effect of pen

135
number (1 to 48) and εijk = the residual; Sampling date was defined as repeated measurement and
εijk was assumed to have a multivariate distribution where observations from different treatments
were uncorrelated while observations from different weeks were assumed to have a toeplitz
(Type= TOEP) correlation structure. Differences in performance attributes were accessed by
analysis of variances with one-way ANOVA tests and the dependent variables were the mean
values for each attribute observed for each pen (body weight gain, feed intake and feed
conversion rate) and the independent variables were the treatments. P-values less than or equal to
0.05 were considered statically significant.
RESULTS
Figure 1A illustrates the mean scores (±SD) of lesions observed in the duodenum of the
“seeders”. As expected, no lesions were observed for the broilers in the negative control group
(T1) and for broilers supplemented with the chemical coccidiostat (T3). However, vaccinated
broilers (T2) presented the highest mean of scores with no differences from broilers in the
positive controlled group (T4).
In addition, broilers supplemented with Regano® (T5) did not differ from vaccinated
broilers (T2) in terms of lesions caused by E. acervulina. Broilers supplemented with LOW dose
of the herbal mixture (T6) presented an intermediary mean score of lesions, not different from
broilers supplemented with Regano® (T5), and not different to the other two doses of the herbal
mixtures (T7 and T8). The two higher doses of the mixture A. annua and C. longa presented less
lesions compared to the vaccinated broilers (T2), to broilers supplemented with Regano® (T5)
and to broilers in the positive control group (T4). However, higher scores were observed for the
mixed treatments (T6, T7 and T8) when compared with broilers supplemented with the chemical
coccidiostat (T3) and with broilers in the negative control group (T1).
The scoring of lesions in the mid-intestinal mucosal surface (Fig 1B) indicated that
broilers receiving the two highest dosages of the herbal combination (T7 and T8) did not differ
from broilers supplemented with the chemical coccidiostats (T3). In addition, vaccinated broilers
(T2) presented the smallest score of lesions in the mid part of the intestine while broilers
supplemented with Regano® (T5) presented the highest score of lesions, similar to the infected

136
non treated broilers (T4) and to broilers supplemented with the LOW dose of the herbal
combination (T6).
When the sum of lesions (duodenum + mid intestine) was considered, a regression
(p=0.04) was observed showing a dose dependent factor when supplementing the mixture A.
annua and C. longa in the ratio 1:4 artemisnin:curcumin (Figure 1C). In summary, for each
increase in dose supplementation (T4–T6–T7–T8), the mean score of lesions was reduced by a
factor of 0.3.
Mortality was low in the first part of the experiment (1-22d) and no records were
observed for broilers supplemented with the chemical coccidiostat. However, in the second
growing period (22-43d), a trend in reduced mortality (p=0.08) was observed for the broilers
treated with the highest dosage of the herbal combination T8 (0.0%) compared to mortality of
birds treated with the chemical coccidiostat T3 (5.6%). Based on the oocyst excreted per g of
faeces (OPG), broilers receiving the smallest dose (LOW) of the herbal combination (T6)
excreted numerically more oocysts, similar to the infected, untreated (positive control) broilers
(T4) and with broilers supplemented with Regano® (T5) (Table 2).
Broilers in the two groups T6 and T4 presented the highest infection peaks at 15 days
post inoculation (PI) and a second peak at 22 days PI compared with broilers supplemented with
the chemical coccidiostat (T3), in the uninfected group (T1) and with broilers receiving the
MEDIUM dose of the combination A. annua and C. longa extracts (T7) (Figure 2). Considering
the total amount of oocysts excreted, broilers supplemented with MEDIUM (T7) and HIGH (T8)
levels of herbal ethanolic extracts presented intermediary OPG not different from the one
recorded for vaccinated broilers (T2) and for broilers in the uninfected group (T1), and not
statistically different from the broilers supplemented with the chemical coccidiostats (Table 2).
Broilers supplemented with the LOW dose of the botanical mixture (T6) and broilers
vaccinated (T2) presented the smaller feed intake for the initial period (1-22d) compared to
broilers in the uninfected group (T1) (Table 2). In the final period investigated (22-43d), broilers
supplemented with Regano® (T5) and broilers vaccinated (T2) presented the highest feed intake
compared with broilers in the negative control group (T1). When all experimental period is
considered, feed intake was higher for the broilers consuming Regano® (T5) and smaller in the
group of broilers consuming the HIGH dose of the botanical mixture (T8).

137
In the initial period, body weight gain (BWG) was higher for the broilers supplemented
with the chemical coccidiostats (T3) but not different compared to broilers in the negative
control group (T1). In addition, BWG in this period (1-22d) was not different when compared to
broilers supplemented with the LOW dose of herbal mixture (T6) (Table 2). Broilers vaccinated
(T2), supplemented with Regano® (T5) and broilers supplemented with the HIGH dose of the
herbal mixture (T8) presented the smallest BWG.
In the second period (22-43d), after the dissemination of oocysts by the “seeders”,
supplementation of chemical coccidiostats in the diet remained as the most efficient treatment in
terms of BWG (Table 2). However, it was not different if compared with the vaccinated broilers
(T2), broilers in the control group (T1) and the ones supplemented with the Regano® (T5). No
differences in FCR were observed in all periods investigated. For the total period (1-43d), BWG
was higher only for the broilers supplemented with the chemical coccidiostats. In addition, feed
intake was impaired for the broilers supplemented with the Regano® (T5) and vaccinated (T2)
when compared to broilers supplemented with the HIGH dose of the botanical mixture (T8) for
the total period investigated (Table 2).
DISCUSSION
To our knowledge this is the first study investigating to combined use A. annua and C.
longa (ethanolic extracts) against coccidiosis in chickens. The study was performed based on
reports on synergistic effects of the combination artemisinin:curcumin against malaria
(Padmanaban et al., 2012; Nandakumar et al., 2006) and that synergistic effects caused by adding
curcumin were not obtained in a ration 1:2 due to a high concentration of artemisinin (Isacchi et
al., 2012). Our results suggest a dose-dependent effect (in a 1:4 ratio) against dual infections
caused by Eimeria acervulina and E. maxima based on reduced lesions score, excreted oocysts,
performance attributes, and mortality.
In general, broilers in the uninfected group (T1) showed no lesions in the mucosal surface
attributed to infections caused by E. acervulina and, apparently, a very low score (mean of 0.4)
attributed to infections caused by E. maxima (Figures 1A and 1B). However, when OPG
counting was considered (Table 2 and Figure 2), “tracers” located in two replicated pens in this
group (T1) started oocyst excretion at 18 to 23 days of age.

138
From one to 22 days of age, broilers in the negative control group (T1) presented no
differences in BWG compared to broilers receiving the chemical coccidiostat (T3) but higher
BWG if compared with broilers supplemented with the SMALL dose of the herbal mixture (T6).
Feed intake and FCR were not impaired when compared with broilers consuming the chemical
coccidiostat (Table 2). However, when broilers were 25 to 30 days old, oocysts were found in
two replicates of this group (T1). Even with the strict hygiene measures applied during the study,
broilers in all six replicates of the uninfected group (T1) were contaminated and excreted oocysts
in the final part of their growing period (22-43d). Even with BWG for the second period not
different compared to broilers consuming chemical coccidiostats (T3), when the total period is
taking in to account (1-43d) this has led to a smaller BWG when comparing T1 with broilers
supplemented with chemical coccidiostat (T3) (Table 2). However, Rosen (1995), in a review
with more than 1200 studies, concluded that supplementation with ionophores improves
performance attributes of fast growing broilers, suggesting that the low infection experienced in
“tracer” broilers for group T1 did not influenced performance attributes and would be compatible
with uninfected broilers. Nevertheless, flies, ants, other insects, and also the technician boots and
hands can serve as vectors and highlight the opportunism and fast dissemination of the Eimeria
parasites (Henken, 1994).
In contrast, broilers in the positive group (T4) presented the highest score of lesions
(Figure 1) and higher excreted amount oocysts, without increasing mortality, compared to other
groups (Table 2). Broiler “seeders” were artificially challenged with dual coccidial infections at
8 days of age and euthanized for scoring of lesions 9 days post inoculation, after sufficient time
to contaminate the litter in each pen in accordance with the pre-patent period of E. acervulina
and E. maxima parasites (Eckert et al., 2005). From this strategy, it was possible to assess the
effects of treatments on intestinal lesions and at the same time allow “seeders” to infect
contemporary broilers by contact (Velkers et al., 2010) imitating the natural transmission
pathway occurring at commercial farms.
Broilers supplemented with chemical coccidiostats (T3) presented no lesions
attributed to E. acervulina (Figure 1A) while few lesions attributed to E. maxima were observed
(Figure 1B). In group T3, the parasites were not completely suppressed. According to Butaye et
al. (2000) and with Martin et al., (1997), and more recently, Abbas et al. (2011), this points to a
build-up of parasite resistance. By observing the infection dynamics from the consecutive OPG

139
counting (Figure 2), a successful suppression in parasites reproduction was observed until
broilers were 22 days of age. From this point ahead, apparently the drug of choice (salinomycin
12%) was not 100% efficient in suppressing the infection (Figure 2). However, even without
complete suppression of oocyst excretion, performance attributes were superior for the group
treated with coccidiostats (T3).
Vaccinated broilers presented the highest mean score of lesions (2.4) in the duodenum
mucosal surface (Figure 1A). Perhaps the time of the E. acervulina challenge, provoked by the
artificial inoculation (2 x 105 oocysts), was too close to vaccination time, suggesting that the time
interval between vaccination and challenge was not sufficient to allow a proper immunization, in
agreement with Oviedo-Rondón et al. (2006). This is also in accordance with the theory that
antibodies released after an initial challenge take time to induce protection by blocking the
development and replication of parasites (Crane et al., 1988; Hafeez et al., 2007; Anwar et al.,
2008).
Vaccinated broilers (T2) presented the lowest mean score of lesions (0.4) in the mid part
of the small intestine, the site of E. maxima infections, comparable to uninfected controls (T1).
On the one hand, vaccination could have been efficient in providing immunization against E.
maxima, quite differently from what was observed with the absence of protection against E.
acervulina. On the other hand, heavier challenges caused by E. acervulina may lead to inhibition
of E. maxima to develop (Mathis, 2005). The former hypothesis sounds more plausible as the
lesions caused by E. acervulina were higher for broilers in T2 (Figure 1A).
Khatafalla et al. (2011) reported a dose-dependent effect of curcumin against E. tenella
sporozoites in vitro. However, there is no literature referring to the use of a mixture of A. annua
and C. longa ethanolic extracts against coccidiosis in chickens. We observed a dose-dependent
response when supplementing ethanolic extracts of A. annua and C. longa equivalent of a 1:4
ratio of artemisinin:curcumin. However, when accessing the scoring of lesions in the mid-
intestinal mucosal surface caused by E. maxima infections (Fig 1B), broilers in T7 and T8 did
not differ from broilers supplemented with the chemical coccidiostats (T3).
In terms of total oocyst excreted by “tracers” among the groups, broilers supplemented
with MEDIUM and HIGH doses were not different compared to broilers supplemented with the
chemical coccidiostats (Table 2). On the one hand, the two higher dosages of the mixtures A.
annua and C. longa (T7 and T8) provided some protection compared to the untreated group (T4),

140
although not statistically different for the total OPG (Table 2). Also, (as supported by Figure 1)
total lesion score was significantly lower for groups T7 and T8 than for group T4 (infected,
untreated). For the total period investigated (1 – 43d), groups T7 and T8 had numerical lower
mortality rates (4.1 and 1.3% respectively) than the group (T3) treated with the chemical
coccidiostat (5.6%). Even though they were not different for total oocyst excreted (Table 2), the
infection dynamics (Figure 2) illustrates that the broilers supplemented with the MEDIUM dose
had a smaller but not statistically different infection peak than broilers supplemented with the
HIGH dose, comparable to broilers supplemented with the chemical coccidiostat.
Although one could argue that the interval between samplings for OPG did not capture
the highest period for occyst extraction, in our opinion, the combination of measured parameters
(OPG, lesion score, and performance attributes) were sufficient to capture the differences
observed among treatments.
For performance attributes, a numerical reduction in BWG was observed in the initial
period (1-22d) suggesting that dose influenced weight gain. It could also be an indication of the
presence of anti palatable components in ethanolic extracts, thus leading broilers to restrict feed
intake as reported when supplementing dried leaves of A. annua to broilers (Almeida et al.,
2012). Even though differences in feed intake were observed between groups, FCR was not
impaired during this period (1-22d) refuting this hypothesis.
Strategies to control coccidiosis in conventional production systems are dependent on
chemical drugs while in organic systems live vaccines or commercial preparations of plant-based
products are used (Abbas et al., 2012). Some studies associate vaccination with botanical
products. For example, Waldenstedt (2003) suggested vaccination in combination with oregano
containing products for increased intestinal health of chickens, and thus reduce the effects caused
by coccidiosis.
Before (1-22d) and after the coccidial challenge (22-43d), broilers supplemented with the
chemical coccidiostat showed higher performance attributes in our study. However, broilers
supplemented with the HIGH dose of the botanical mixtures (T8) presented smaller feed intake
(13 to 22 g/day) compared with vaccinated broilers (T2) and broilers supplemented with
Regano® (T5) without any differences in BWG and FCR (Table 2).

141
From the conditions experienced in this study, we concluded that broilers supplemented
with chemical coccidiostats presented superior performance attributes during the trial due to a
better induction of protection against dual infections caused by E. acervulina and E. maxima
parasites. However, despite lower BWG for the two highest doses of the extracts mixture, the
protection against lesions, and trend in reducing mortality and OPG counts provided by the
HIGH dose of the combined A. annua and C. longa ethanolic extracts, compared to the positive
control, suggests that the supplementation of herbal extracts in drinking water can be a feasible
alternative method for coccidia control in organic production systems. It is affordable, practical
to be implemented at farm level, and is residue free. Other studies to investigate dosages and
modes of action of this combination are warranted. In addition, supplementation of herbal
extracts in combination with other management practices may help farmers engaged in the
production of broilers to reduce the use of synthetic drugs.
ACKNOWLEDGEMENTS
The authors wish to thank Evandro Possamai, Dayana Pereira and Pedro Lorga from
Korin agropecuaria Ltd. for the logistical and technical support during the study.
FINANCIAL SUPPORT
The Danish Ministry of Science, Technology and Innovation is thanked for financing
collaboration among Brazilian and Danish scientists. Aarhus University, SOAR - Research
School for Organic Agriculture and Food Systems and Korin Agropecuária Ltd. - are thanked for
supporting our study.

142
REFERENCES Abbas, R. Z., Colwell, D. D. and Gilleard, J. (2012). Botanicals: an alternative approach for the control of avian coccidiosis. World´s Poultry Science Journal 68, 203-215. Abbas, R. Z., Iqbal, Z. Blake, D., Khan, M. N. and Saleemi, M. K. (2011). Anticoccidial drug resistance in fowl coccidia: the state of play revisited. World´s Poultry Science Journal 67, 337-350. Adams, C., Vahl, H. and Veldman, A. (1996). Interaction between nutrition and Eimeria acervulina infection in broiler chickens: development of an experimental infection model. British Journal of Nutrition 75, 867–873. Allen, P.C., Lydon, J. and Danforth, H.D. (1997). Effects of components of Artemisia annua on coccidia infections in chickens. Poultry Science 76, 1156-1163.
Allen, P.C., Danforth, H. D. and Augustine, P. C. (1998). Dietary modulation of avian coccidiosis. International Journal for Parasitology 8, 1131–1140.
Almeida, G.F d., Horsted, K., Thamsborg, S. M., Kyvsgaard, N. C., Ferreira, J. F. S. and Hermansen, J. E. (2012). Use of Artemisia annua as a natural coccidiostat in free-range broilers and its effects on infection dynamics and performance. Veterinary Parasitology 186, 178-187.
Arab, H.A., Rahbari, S., Rassouli, A., Moslemi, M. H. and Khosravirad, F. D. A. (2006). Determination of artemisinin in Artemisia sieberi and anticoccidial effects of the plant extract in broiler chickens. Tropical Animal Health and Production 38, 497-503.
Akhtara, M., Haia, A., Awaisa, M. M., Iqbala, Z., Muhammadb, F., Haqc, A.u., Anward, M. I., (2012). Immunostimulatory and protective effects of Aloe vera against coccidiosis in industrial broiler chickens. Veterinary Parasitology 186, 170– 177. Anonymous (1990). Diagnosis of coccidiosis in chickens. Janssen Pharmaceutica, Beerse, Belgium. Anwar, M. I., Akhtar, M., Hussain, I., Muhammad, F., Haq, A. U. (2008). Effects of local gametocyte and livacox vaccines on live body weight gain and lymphoid organs in chickens. Pakistan Veterinary Journal 28, 136–138. Brisibe, E. A., Umoren, U. E., Owai, P.U. and Brisibe, F. (2008). Dietary inclusion of dried Artemisia annua leaves for management of coccidiosis and growth enhancement in chickens. African Journal of Biotechnology 7, 4083-4092. Bumstead, J. M., Dunn, P. P. J. and Tomley, F. M. (1995). Nitrocellulose immunoblotting for identification and molecular gene cloning of Eimeria maxima antigens that stimulate lymphocyte proliferation. Clinical and Diagnostic Laboratory Immunology 2, 524–530.

143
Butaye, P., Devriese, L. A. and Haesebrouck, F. (2000). Incomplete cross resistance against ionophores in Enterococcus faecium and Enterococcus faecalis strains from pigs and poultry. Microbial Drug Resistance: Mechanisms Epidemiology and Disease 6, 59-61. Chapman, H. D. (1993). Resistance to anticoccidial drugs in fowl. Parasitology Today 9, 159–162. Chen, Y., Xianfu, L., Hyunjin, P. and Greever, R. (2009). Study of artemisinin nanocapsules as anticancer drug delivery systems, Nanomedicine 5, 316–322. COM (2008). COM/2008/0233. Report from the Commission to the Council and the European Parliament on the use of coccidiostats and histomonostats as feed additives, submitted pursuant to article 11 of regulation (EC) no. 1831/2003 of the European Parliament and of the Council of 22 September, 2003 on additives for use in animal nutrition, http://www.ipex.eu/ipex/cms/home/Documents/dossier_COM20080233. Conney, A. H, Lysz, T., Ferraro, T., Abidi, T. F., Manchand, P. S., Laskin, J. D. and Huang, M. T. (1991). Inhibitory effect of curcumin and some related dietary compounds on tumor promotion and arachidonic metabolism in mouse skin. Advances in Enzyme Regulation 31, 385-396. Crane, M. S., Murray, P. K., Gnozzio, M. J. and MacDonald, T. T. (1988). Passive protection of chickens against Eimeria tenella infection by monoclonal antibody. Infection and Immunity 56, 972–976. Cui, L., Miao, J. and Cui, L. (2007). Cytotoxic effect of curcumin on malaria parasite Plasmodium falciparum: inhibition of histone acetylation and generation of reactive oxygen species. Antimicrobial Agents and Chemotherapy 51, 488–494. Daugschies, A., Gässlein and U., Rommel, M. (1998). Comparative efficacy of anticoccidials under the conditions of commercial broiler production and in battery trials. Veterinary Parasitology 76, 163-171. Del Cacho, E., Gallego, M., Francesch, M., Quílez, J. and Sánchez-acedo, C. (2010). Effect of artemisinin on oocyst wall formation and sporulation during Eimeria tenella infection. Parasitology International 4, 506-511. Eckert, J., Taylor, M., Catchpole, J., Licois, D., Coudert, P. and Bucklar, H. (1995). Morphological characteristics of oocysts. In: Eckert, J., Braun, R., Shirley, M., Coudert, P. (Eds.), COST 89/820. Biotechnology. Guidelines of Techniques in Coccidiosis Research. Office for Official Publications of the European Communities, Luxembourg, pp. 103–119. Ferreira, J. F. S. and Gonzales, J. M. (2009). Analysis of underivatized artemisinin and related sesquiterpene lactones by high-performance liquid chromatography with ultraviolet detection. Phytochemical Analysis 2, 91-97.

144
Ferreira, J. F. S., Peaden, P. and Keiser, J. (2011). In vitro trematocidal effects of crude alcohol-ic extracts of Artemisia annua, A. absinthium, Asimina triloba, and Fumaria officinalis. Par-asitology Research 109, 1585-1592. Hafeez, M. A., Akhtar, M., Javed, M. T. and Haq, A. U. (2007). Maternal immunization by egg propagated gametocyte vaccine to control Eimeria tenella infections in newly hatched chicks. Parasitology Research 100, 1139-1141. Henken, A. M., H. W. Ploeger, E. A. M. Graat, and T. E. Carpenter. (1994). Description of a simulation model for the population dynamics of Eimeria acervulina infection in broilers. Parasitology 108, 503-512. Huang, M. T., Newmark, H. L. and Frenkel, K. (1997). Inhibitory effects of curcumin on tumorigenesis in mice. Journal of Cellular Biochemistry Supplement 27, 26–34. Isacchi, B., Bergonzi, M. C., Grazioso, M., Righeschi, C., Pietretti, A., Severini, C. and Bilia, A. R. (2012). Artemisinin and artemisinin plus curcumin liposomal formulations: Enhanced antimalarial efficacy against Plasmodium berghei-infected mice. European Journal of Pharmaceutics and Biopharmaceutics 80, 528–534. Johnson J. and Reid W. M. (1970). Anticoccidial drugs lesion scoring techniques in battery and floor-pen experiments with chickens. Experimental Parasitology 28, 30-6. Jung, M., Lee, K., Kim, H. and Park, M. (2004). Recent advances in artemisinin and its derivatives as antimalarial and antitumor agents. Current Medical Chemistry 11, 1265-1284. Kettunen, H., Tiihonen, K., Peuranen, S., Saarinen, M. T., Remus and J. C. (2001). Dietary betaine accumulates in the liver and intestinal tissue and stabilizes the intestinal epithelial structure in healthy and coccidia infected broiler chicks. Comparative Biochemistry Physiology Part A: Molecular and Integrative Physiology 130, 759–769. Khalafalla, R. E., Müller, U., Shahiduzzaman, M., Dyachenko, V., Desouky, A. Y., Alber, G. and Daugschies, A. (2011). Effects of curcumin (diferuloylmethane) on Eimeria tenella sporozoites in vitro. Parasitology Research 108, 879-886. Klaiman, D. L. (1985). Quinghaosu (artemisinin): an antimalarial drug from China, Science 228, 1049–1055. Lee , S. H., Lillehoj, H. S., Jang, S. I., Kim, D. K., Ionescu, C. and Bravo, D. (2010). Effect of dietary Curcuma, Capsicum and Lentinus on enhancing local immunity against Eimeria acervulina infection. Journal of Poultry Science 47, 89-95. Lillehoj, H. S., and J. M. Trout, 1993. Coccidia: a review of recent advances on immunity and vaccine development. Avian Pathol. 22:3–31.

145
Mathis, G. F. (2005). Reasons for field problems with E. maxima: E. acervulina vs E. maxima. Proceeding of the IXth International coccidiosis conference, Foz do Iguacú, Brazil. 19-23 Sep 2005. Martin, A., Danforth, H., Barta, J. and Fernando, M. (1997). Analysis of immunological cross-protection and sensitivities to anticoccidial drugs among five geographical and temporal strains of Eimeria maxima. International Journal for Parasitology 27, 527–533. Martinelli, A., Rodrigues, L. A. and Cravo, P. (2008). Plasmodium chabaudi: efficacy of artemisinin + curcumin combination treatment on a clone selected for artemisinin resistance in mice. Experimental Parasitology 199, 304–307. McDougald, L. R., Fuller, L. and Mattiello, R. (1997). A survey of coccidian on 43 poultry farms in Argentina. Avian Diseases 41, 923–929. Min, W., H. S. Lillehoj, C. M. Ashwell, C. P. van Tassell, R. A. Dalloul, L. K. Matukumalli, J. Y. Han, and E. P. Lillehoj. (2005). Expressed sequence tag analysis of Eimeria-stimulated intestinal intraepithelial lymphocytes in chickens. Molecular Biotechnology 30, 143–150. Morris, G. M., Woods, W. G., Richards, D. G. and Gasser, R. B. (2007). Investigating a persistent coccidiosis problem on a commercial broiler–breeder farm utilizing PCR-coupled capillary electrophoresis. Parasitology Research 101, 583-589. Naidoo, V., McGaw, L. J., Bisschop, S. P. R., Duncan, N. and Eloff, J. N. (2008). The value of plant extracts with antioxidant activity in attenuating coccidiosis in broiler chickens. Veterinary Parasitology 153, 214-219. Nandakumar, D. N., Nagaraj, V. A., Vathsala, P. G., Rangarajan, P. and Padmanaban, G. (2006). Curcumin-Artemisinin Combination Therapy for Malaria. Antimicrobial Agents and Chemeotherapy 50, 1859 – 1860. Nosten, F. and White, N. J. (2007). Artemisinin-based combination treatment of Plasmudium falciparum malaria. American Journal Tropical Medicine and Hygiene 77, 181–192. Nwosu, C. O., Yahayah, K. and Igbokwe, I. O. (2011). Toxicity and Anticoccidial Efficacy of Some Plants Used in the Traditional Treatment of Avian Coccidiosis in Semi-arid Northeastern Nigeria. Research Journal of Parasitology 6, 18-30. Orengo, J., Buendía, A. J., Ruiz-Ibáñez, M. R., Madrid, J., Del Río, L., Catalá-Gregori, P., García, V. and Hernández, F. (2012). Evaluating the efficacy of cinnamaldehyde and Echinacea purpurea plant extract in broilers against Eimeria acervulina. Veterinary Parasitology 185, 158-163.
Oviedo-Rondón, E. O., Clemente-Hernández, S., Salvador, F., Williams P., Losa R. and Stephen F. (2006). Essential Oils on Mixed Coccidia Vaccination and Infection in Broilers. International Journal of Poultry Science 5, 723-730.

146
Padmanaban, G., Nagaraj V. A. and Rangarajan, P. N. (2012). Artemisinin-based combination with curcumin adds a new dimension to malaria therapy. Current Science 102, 704-711. Pinard-van der Laan, M.-H., Monvoisin, J.-L., Pery, P., Hamet, N. and Thomas, M. (1998). Comparison of Outbred Lines of Chickens for Resistance to Experimental Infection with Coccidiosis (Eimeria tenella). Poultry Science 77, 185–191. Quinn G.P. and Keough M.J. (2002). Experimental design and data analysis for biologists. 6th printing, 2007. Cambridge University Press. Rao, C. V., Rivenson, A., Simi, B., Reddy, B. S. (1995). Chemoprevention of colon carcinogenesis by dietary curcumin, a naturally occurring plant phenolic compound. Cancer Research 55, 259–266. Reddy, R. C., Vathsala, P. G., Keshamouni, V. G., Padmanaban, G. and Rangarajan, P. N. (2005). Curcumin for malaria therapy. Biochemical and Biophysical Research Communications 326, 472–474. Rosen, G. D. (1995). Antibacterials in poultry and pig nutrition. In: Wallace, R. J. and Chesson, A. (eds), Biotechnology in Animal Feeds and Animal Feeding, VCH Verlagsgesellschaft GmbH, Weinheim. SAS Institute (2000). SAS Online Doc®, Version 9.2. SAS Institute, Cary, NC. Shahiduzzaman, M., Dyachenko, V., Khalafalla, R. E., Desouky, A. Y., Daugschies, A. (2009). Effects of curcumin on Cryptosporidium parvum in vitro. Parasitology Research 105, 1155-1161. Shirley, M. W., Smith, A. L. and Tomley, F. M. (2005). The biology of avian Eimeria with an emphasis on their control by vaccination. Advanced Parasitology 60, 285–330. Souza, C. R. A. and Glória, M. B. A. (1998). Chemical analysis of turmeric from Minas Gerais, Brazil and comparison of methods for flavour free oleoresin. Brazilian Archives of Biology and Technology. 41, 218-224. Subramanian, M., Sreejayan Rao, M. N., Devasagayam, T.P., Singh, B. B. (1994). Diminution of singlet oxygen-induced DNA damage by curcumin and related antioxidants. Mutation Research 311, 249–255. Tsinas, A., Giannenas, I., Voidarou, C.,; Tzora, A., and Skoufos, J. (2011). Effects of an Oregano Based Dietary Supplement on Performance of Broiler Chickens Experimentally Infected with Eimeria Acervulina and Eimeria Maxima. Journal of Poultry Science 48, 194-200. Ungemach, F. R., Müller-Bahrdt, D. and Abraham, G. (2006). Guidelines for prudent use of antimicrobials and their implications on antibiotic usage in veterinary medicine. International Journal of Medical Microbiology 296 (Suppl.) S2, 33-38.

147
Velkers, F. C., Swinkels, W. J. C., Rebel, J. M. J., Bouma, A., Daemen, A. J. J. M., Klinkenberg, D., Boersma, W. J. A., Stegeman, J. A., de Jong, M. C. M. and Heesterbeek, J. A. P. (2010). Effect of Eimeria acervulina infection history on the immune response and transmission in broilers.Veterinary Parasitology 173, 184-192. Waldenstedt, L. (2003). Effect of vaccination against coccidiosis in combination with an antibacterial oregano (Origanum vulgare) compound in organic broiler production. Acta Agriculturae Scandinavica 53, 101 – 109. Wang, M. L., Suo, X., Gu, J. H., Zhang, W. W., Fang, Q. and Wang, X. (2008), Influence of grape seed proanthocyanidin extract in broiler chickens: effect on chicken coccidiosis and antioxidant status. Poultry Science 87, 2273-2280. Williams, R. B. (1999). A compartmentalized model for the estimation of the cost of coccidiosis to the world’s chicken production industry. International Journal for Parasitology 8, 1209-1229. Williams, R. B. (2002). Fifty years of anticoccidial vaccines for poultry, 1952-2002. Avian Disease 46, 755-802. Zaman, M. A., Iqbal, Z., Abbas, R. Z., Khan, M., Muhammad, N. G., Younus, M. and Ahmed, S. (2012). In vitro and in vivo acaricidal activity of a herbal extract. Veterinary Parasitology 186, 431–436.

148
Table 1 – Description of treatments, acronyms, supplementation strategy and age of the broilers
when treatments were supplemented
Treatment Acronym Delivering strategy *
Age of the broilers (days)
Negative Control (untreated, uninfected) T1 - -
Vaccinated¹, infected² T2 Drinking water
1
Coccidiostat³, infected T3 Dietary inclusion
1-39
Positive control (untreated, infected) T4 - -
Regano®4, Infected T5 Drinking water
1-39
Dose 1 of Ethanolic herb extracts of A. annua + C. longa. (11 ppb Artemisinin + 45 ppb Curcumin), Infected
T6 Drinking water
1-42
Dose 2 of Ethanolic herb extracts of A. annua + C. longa. (19 ppb Artemisinin + 74 ppb Curcumin), Infected
T7 Drinking water
1-42
Dose 3 of Ethanolic herb extracts of A. annua + C. longa. (27 ppb Artemisinin + 107 ppb Curcumin), Infected
T8 Drinking water
1-42
¹Livacox vaccine (Merial Ltd. Brazil) consisted of attenuated oocysts of E. acervulina, E. maxima and E. tenella.
² Three (3) “seeders” per pen (10%/pen) aged 8-days-old where orally inoculated with 1ml suspension containing 5 x 104 E. maxima and 2 x 105 E. acervulina sporulated oocysts. “Seeders” were removed from the pens at 17 days-old, 9 days post-inoculation.
³ 50 ppm Nicarbazin + Narazin (Maxiban®) supplemented from 1 to 22 days of age and 66 ppm Salinomycin 12% (Coxistac®) supplemented from 22 to 39 days of age. 4 42 g/Kg Carvacrol + 1.25g/Kg Timol (Regano®).
* Preparation of mixtures were performed every second day. 155 liters of tap water were mixed with dosages of herbal ingredients in T5, T6, T7 and T8.

149
Table 2 – Performance attributes for each treatment* for two distinct periods under investigation and for the entire period investigated.
Periods consisted of 1 to 22 days of age, 22 to 43 days of age and 1 to 43 days of age. Degree of infection is presented by the mean
OPG (oocyst per gram of faeces) for the total period. All results presented are in LS means ± SEM.
Performance attributes
p values
NEGATIVE CONTROL
(T1)
VACCINE
(T2)
CHEMICAL COCCID.
(T3)
POSITIVE CONTROL
(T4)
REGANO
(T5)
LOW DOSE (T6)
MEDIUM DOSE (T7)
HIGH DOSE (T8)
First
Period
1-22 d
BWG1 (g) 0.01 838 ± 15 ab 789 ± 13 cde 873 ± 09 a 818 ± 14 bcd 779 ± 15 de 836 ± 12 ab 821 ± 13 bc 768 ± 16 e
FI2 (g/d) 0.01 68.9 ± 2.3 a 62.6 ± 0.5bc 66.2 ± 0.9 ab 65.1 ± 1.1 abc 65.6 ± 1.2 abc 63.7 ± 0.4 bc 64.5 ± 1.4 bc 61.6 ± 0.9 c
FCR3 0.38 1.71 ± 0.07 1.67 ± 0.04 1.59 ± 0.04 1.67 ± 0.04 1.75 ± 0.05 1.59 ± 0.03 1.65 ± 0.06 1.70 ± 0.03
Mort.4 (%) 0.56 3.3 ± 2.1 1.1 ± 1.1 0.0 ± 0.0 1.1 ± 0.7 2.7 ± 1.2 1.7 ± 1.2 1.1 ± 0.7 1.3 ± 0.8
Second
Period
22-43 d
BWG (g) 0.05 2010 ± 28 ab 2002 ± 35 ab 2103 ± 29 a 1993 ± 44 b 2013 ± 27 ab 1916 ± 35 b 1946 ± 37 b 1964 ± 39 b
FI (g/d) 0.02 177.3 ± 2.9 bc 187.7 ± 5.8ab 180.9 ± 3.2 bc 180.1 ± 2.0bc 194.1 ± 5.1 a 177.4 ± 2.8bc 183.0 ± 2.8 bc 175.1 ± 3.0c
FCR 0.08 1.80 ± 0.03 1.88 ± 0.07 1.71 ± 0.03 1.85 ±0.05 1.91 ± 0.05 1.87 ± 0.05 1.93 ± 0.06 1.87 ± 0.04
Mort. (%) 0.20 2.5 ± 1.3 4.5 ± 2.1 5.6 ± 0.9 3.0 ± 1.2 5.0 ± 1.4 3.1 ± 1.2 3.0 ± 1.1 0.0 ± 0.0
Total
period
1-43 d
BWG (g) 0.01 2848 ± 36 b 2791 ± 41 b 2976 ± 34 a 2811 ± 46 b 2792 ± 34 b 2787 ± 39 b 2767 ± 42 b 2732 ± 47 b
FI (g/d) 0.02 124.4 ± 1.9bc 127.2 ± 4.0ab 125.5 ± 1.7abc 123.6 ± 1.2bc 132.0 ± 3.1a 121.4 ± 1.6bc 125.1 ± 2.3abc 118.4 ± 1.9c
FCR 0.10 1.79 ± 0.03 1.85 ± 0.06 1.71 ± 0.02 1.81 ± 0.04 1.90 ± 0.05 1.80 ± 0.04 1.86 ± 0.06 1.82 ± 0.02
Mort. (%) 0.59 5.9 ± 3.2 5.6 ± 2.6 5.6 ± 0.9 4.1 ± 1.5 7.7 ± 1.3 4.8 ± 1.7 4.1 ± 1.4 1.3 ± 0.8
k OPG5 0.05 7.4 ± 0.9 ab 7.3 ± 0.9 ab 5.3 ± 0.7 b 22.1 ± 2.7 a 15.5 ± 1.9 a 24.6 ± 3.4 a 8.8 ± 1.1 ab 12.7 ± 1.5 ab a,b,c Number in a row marked with different letters are significantly different (p<0.05). *Treatments consisted of: T1= Uninfected negative control (untreated, uninfected); T2= Vaccinated, infected; T3= Coccidiostat, infected; T4= Untreated, Infected (positive control); T5= Regano®, Infected; T6= Dose 1 (LOW) of herbal extracts, Infected; T7= Dose 2 (MEDIUM) of herbal extracts, Infected; T8= Dose 3 (HIGH) of herbal extracts, Infected. 1BWG = Body weight gain for the period specified (g). 2FI = Feed intake for the period specified (g/broiler/day). 3FCR = Kg feed consumed / body weight gain in Kg for the period specified. 4Mort. (%) = Percentage mortality observed for the period specified. 5k OPG = 100 x OPG.

150
Figure 1 – Mean scores of gross lesions ± SD for duodenal (A), mid intestine (B) and total (C)
lesions.
a,b,c Bars in the same figure marked with different letters are significantly different (p<0.05).
* Treatments with different letters are significantly different at P < 0.05. Treatments consisted of:
T1= Negative control (untreated, uninfected); T2= Vaccinated, infected; T3= Coccidiostat,
infected; T4= Untreated, Infected (positive control); T5= Regano®, Infected; T6= Dose 1 (LOW)
of herbal extracts, Infected; T7= Dose 2 (MEDIUM) of herbal extracts, Infected; T8= Dose 3
(HIGH) of herbal extracts, Infected.

151
Figure 2 - Effect of treatments* on coccidia oocyst excretion after 8 consecutive samplings from
“tracer broilers”. Curves are presented representing the variation in the mean OPG (oocyst per
gram of faeces) for the 6 replicates pens for each of the date of faeces collection.
*Treatments consisted of: T1= Negative control (untreated, uninfected); T2= Vaccinated, infected;
T3= Coccidiostat, infected; T4= Untreated, Infected (Positive control); T5= Regano®, Infected;
T6= Dose 1 (LOW) of herbal extracts, Infected; T7= Dose 2 (MEDIUM) of herbal extracts,
Infected; T8= Dose 3 (HIGH) of herbal extracts, Infected.

152
Paper V
Farmer´s attitudes and perceptions on feeding strategies and control of
coccidiosis in alternative broilers systems in Brazil
Almeida, G.F.d.; Figueiredo, E.A.; Molento, M.B.; Demattê Filho, L.C.; Horsted, K.;
Thamsborg, S.M.; Hermansen, J.E. (2012) Manuscript in preparation
Manuscript in preparation for submission to Organic Agriculture

153
Farmer´s attitudes and perceptions on feeding strategies and control of coccidiosis in
alternative broilers systems in Brazil
G. F. d. Almeida a *, E. A. Figueiredo b; M. B. Molento c; L.C. Demattê Filho d; K. Horsted a; S. M.
Thamsborg e and J. E. Hermansen a
a Department of Agroecology, Faculty of Sciences and Technology, Aarhus University. Research
Centre Foulum, Blichers Allé 20, P.O. Box 50 DK-8830 Tjele, Denmark.
b EMBRAPA National Institute of Research in Swine and Poultry Systems. Caixa Postal 21 CEP:
89700-000. Concórdia, SC, Brazil.
c Laboratory of Parasitological Diseases, Federal University of Parana. Rua dos Funcionários,
1540 Curitiba, PR 80035-050, Brazil.
d Department of Economy and Rural Sociology, Luiz de Queiroz College of Agriculture
(ESALQ), University of São Paulo, Avenida Pádua Dias 11, Piracicaba, SP, Brazil.
e Danish Centre for Experimental Parasitology, Faculty of Life Sciences, University of
Copenhagen, Dyrlægevej 100, DK-1870 Frederiksberg C, Denmark.
* Corresponding author: Gustavo Fonseca de Almeida; Tel. +45 8715 4757; Fax. +45 8715 4798.
E-mail address: [email protected]
Correspondence address for proofs:
Department of Agroecology, Faculty of Sciences and Technology, Aarhus University. Research
Centre Foulum, Blichers Allé 20, P.O. Box 50, DK-8830, Tjele, Denmark.

154
Abstract: This paper presents an analysis of family farmers producing broilers in alternative systems in
three different agro ecological zones in Brazil. Two groups of farmers raising free range broilers
were organized in cooperatives in the South of Brazil and a third group of farmers producing
antibiotic free (AF) broilers were linked to a private company in a vertical integration scheme in
the state of São Paulo. Individual assessments of production systems were performed in 21 farms
among the three case studies. The main limitations, advantages, and challenges in production
were investigated. The research was developed through the complementarities provided by a
combination of sociological and agronomic research methods, based on qualitative interviews
and questionnaire with semi-structured questions. Groups producing free range broilers linked to
the two cooperatives in the South revealed high costs of feed ingredients and the organization of
the production chains as the major limiting factors in production. The third group revealed
diseases of the gastrointestinal tract of broilers as the major limiting factor in production due to
the absence of prophylactic treatments in the production of the AF broilers. Free-range broilers
in the south were exposed to permanent paddocks with very little vegetation and plots were not
managed with the aim to supply nutrients to broilers. In São Paulo, the majority of farmers (80%)
producing the AF broilers reported coccidiosis as an important factor affecting production and
75% reported drinking water as the most appropriate delivery strategy to supplement herbal
extracts in strategies to control diseases of the gastrointestinal tract of broilers, especially
coccidiosis. While farmers producing free range broilers in the south were demanding alternative
sources of feed ingredients to reduce production costs and more organization in the production
chains, farmers in São Paulo producing the AF broilers were more preoccupied with the
identification of strategies to control avian coccidosis. Based on farmer’s opinions, this study
presents a list of crops and plant species with potential use to reduce the dependence on maize
and soybeans in addition to forage species with potential use in paddocks cultivation for foraging
broilers. Integration strategies for holistic systems were suggested. Agro-forest systems would be
interesting for farmers in systems with fruits, trees for wood production and free range broilers
could be integrated with small ruminants or dairy cows.
Key words: Family farmer; Broiler; Free range; Antibiotic free; Alternative feed ingredients;
Forage; Animal husbandry.

155
1. Introduction
For many years chickens (Gallus gallus domesticus) have been raised for the production of
eggs and meat for human consumption. From indigenous and less productive genotypes, modern
breeding programs emphasized productivity and specialization, either layers or broilers (Tixer-
Boichard et al., 2012). As a result of intensive selection, production of broilers has markedly
increased in the last 40 years. (FAO, 2012).
The introduction of new technologies for intensive and standardized systems and the
modernization in crop production has favoured countries in the south hemisphere due to cheaper
labour and availability of feed ingredients, especially maize and soybeans (ABEF, 2012). In
2010, Brazil was the third largest world producer of chicken meat accounting for 12% in the
world and 65% of what was produced in South America (FAO, 2012). In 2011, total production
of chicken meat in Brazil reaches 13 million metric tonnes (MT) with 60% of this total volume
produced by industries located in three states: Paraná (28.4%); Rio Grande do Sul (15.2%) and
São Paulo (14.4%) (ABEF, 2012).
On one hand, modernization of the poultry industry has provided cheap sources of proteins
for feeding the population. On the other hand, the industrial system leaded to a range of
limitations (Molento, 2009). For example, coccidiostats and histomonostats could be phased out
as feed additives in Europe by 31st December 2012 (COM, 2008) due to health and food safety
reasons. In recent years, there has been a growing market for antibiotic free, bacteria-free, free-
range and organically-reared chickens and other meat/milk sources due to changes in the
consumers’ behavior (Herrmann and Roder, 1995). Improvements in animal welfare and
environmental protection, health issues - especially the absence of pesticide in the production of
crops and medicine residues in animal products like milk, eggs and meat were the most
important attributes highlighted by consumers (Wier et al., 2008; Martínez-Michell et al., 2011).
Francisco et al. (2007), studying consumer perceptions in metropolitan areas in Brazil, also
reported high concerns with food safety related to risks associated with bacterial contamination.
In free range and organic systems, broilers must be supplemented with diets composed
mainly of organic ingredients which are basically maize and soybeans. Organically produced
crops and cereals are extremely expensive for feeding broilers due to small production and high
competition with human nutrition.

156
In older traditional systems (Prothero, 1936), chickens were normally left to roam seeking
essential parts of their nutritional needs on pasture or below the soil surface. Robinson (1961)
estimated feed savings about 10% when fresh grass was available to hens in free-range systems.
Slow growing broilers raised in a Danish free range system were able to access 10-15% of the
daily methionine requirements from forage (Almeida et al., 2012). In addition, the access to
pasture exerts relevant effects both, product quality and animal health (Lopez-Bote et al., 1998;
Sossidou et al., 2010). However, in practice very little attention has been given to make outdoor
runs attractive and nutritious to broilers (Van de Weerd et al., 2009). In free range systems,
broilers are normally managed in fixed houses and animals are more exposed to parasite
infections because they are raised with access to permanent paddocks (Permin et al., 1999).
Rodrigues et al. (1997) in Mexico, and Luchese et al. (2007) in Brazil, referred to coccidiosis as
the most important cause of chicken mortality also in smallholder systems.
Tovar et al (2005) investigated Mexican family farmers organized producing organic
coffee, Oelofse et al. (2010) studied smallholders producing crops and tropical fruits in Brazil,
China and Egypt and more recently Blanc and Kledal (2012) reported three different smallholder
cooperatives producing organic vegetables and tropical fruits in Brazil. Unfortunately, no studies
were found in respect to smallholder organizations in Brazil producing broilers in organic and
alternative systems.
On this background, three farm case studies of family farmers were searched in Brazil with
two main objectives, first to investigate the extent on which coccidiosis affects production in
drug-free broiler systems, and secondly, exploring farmer’s opinions on alternative ingredients
to substitute the use of maize and soybeans in nutrient supplementation. Future challenges of the
production of broilers in free range systems in Brazil will be discussed.
2. Methods
2.1. Case study areas and data collection
The study was designed as a multiple case study as suggested by Yin (2003). The first
group was formed by contracted farmers in a vertical integration scheme linked to Korin
Agropecuária Ltd., a broiler company in the Estate of São Paulo that produces antibiotic-free

157
broilers without growth enhancers or ingredients of animal origin in the feed. The second group
of farmers was organized in a cooperative located in the Estate of Rio Grande do Sul, producing
mainly oil seed crops and cereals, fruits and free range “caipira” broilers. The third group was
organized in a cooperative in the Estate of Parana producing mainly vegetables and free range
“caipira” broilers. General details of the sites are presented in Table 1.
Each case study consisted of a sample of collaborative farmers. Case areas were selected
following discussions with local researchers and stakeholders based upon the following criteria:
(1) that smallholders were producing broilers without chemical remedies and medicines in
alternative production systems in the regions where most part of the production of table chickens
takes place in Brazil (2) that farmers should be small-scale producers or family owned-farmers,
and (3) linked to an alternative supply markets. Sample sizes and the characteristics of farm sites
are presented in Table 2.
Family farmers from each case study were selected following the provision of the farmer
lists from cooperatives and the broiler company. After preliminary field observations, an
extensive semi-structured questionnaire was conducted over a number of field visits in a method
previously applied by Almeida and Abreu (2009).
The instrument of data collection acquired household information and detailed production
data as well as qualitative information of farmers' perceptions of aspects related to farming
systems. In addition, the focus of the interviews were also to gain knowledge on farmers
opinions and constrains regarding strategies to reduce the use of oil seed crops and cereals in
broilers nutrition by local alternative feed sources and forage crops for foraging broilers.
The questionnaire was basically divided in five main topics. First, the farm description and
the household information on labour use and family members as work power was explored. The
second part described the production and the animal species managed in the current farm
systems. The third part explored the nutritional management including the broilers and the input-
output relation based on the products with economic importance to the farms. The fourth part
explored the most relevant challenges from the family farmers for producing broilers, while the
final part investigated their perspectives for the future in terms of keeping the production of
broilers. We registered the opinion of farmers regarding different feed ingredients to substitute
maize and soybeans and their opinions on appropriate methods to supplement alternative
products in the control of coccidosis. Additionally, qualitative data were collected during follow-

158
up verification visits as it was performed in previous studies with smallholders in Brazil
(Oelofse et al., 2011; Knudsen et al., 2011). Key stakeholders were also interviewed following
Kvale and Brinkmann (2009) and contextual data were collected from relevant organizations
such as Cooperatives, Municipalities, Extension offices and Universities.
2.2 – Analytical strategy
The study was framed by developing a description of production characteristics for each
case study. Initially, the organization of farmers in the case areas was investigated by
interviewing group leaders. It was followed by the description of the regional characteristics in
terms of crop seasons, production and livestock obtained after visiting important stakeholders in
the municipalities.
Categorical data from the questionnaires were used to elaborate a database in Excel®
within different categories including the farmer’s opinions for each case study. Data from
qualitative interviews were transcript and used to explain the main characteristics of the case
areas. Farmer’s opinions were grouped among case studies thus considering the regional
characteristics (environment, neighbours, farmers organization and the supply markets) and used
to identify to what extent the production of broilers was integrated to the farming systems and in
what extent its production was economically important for the farmers.
3. The farmer’s organization
3.1 Ipeúna (SP)
The municipality of Ipeúna is located in the Estate of São Paulo, 208 km of the São Paulo
city and is located in the hydrological basin of Corumbataí river. The main use of soil in the
region is for the production of sugar cane, eucalyptus, and orange. Ipeúna is in the
neighbourhood of mountain areas where small fragments of forests and mosaics of different
crops can be observed in diversified production systems managed by small scale farmers. The
company Korin Agropecuária Ltd. is located in the municipality of Ipeúna and is owned by the
World Messianic Church in Brazil. Farmers linked to the company are placed in the surround of

159
50 km distance from the company. The company’s philosophy considers the Nature Farming for
agricultural practices aiming to produce health food in accordance with principles of nature
(Demattê Filho, 2004).
In 1957, the concept was brought to Brazil by Japanese immigrants and the practice of
Nature Farming was disseminated in their colonies. In 1981 the Mokiti Okada Foundation was
founded in Brazil and a farm in the municipality of Ipeúna became the hub of the Nature
Farming methods. Production started with fruits, legumes and vegetables initially supplied for
the Japanese community. After some years of growth and investments in production, the farm
was structured and the Mokiti Okada Research Centre was launched for investigating the Nuture
Farming system. In 1993 research was initiated in the production of Natural (antibiotic free)
broilers. Korin Agropecuária Ltd. was created in 1995 to be in accordance with the Brazilian
legislation and to market vegetables, fruits and AF broilers.
By the end of 1999, the production of broilers was transferred to integrated and
collaborative family farmers located in the region and in the time of this study the company
interacted with 26 contracted smallholders. Farmers were trained to follow the standardized rule
in the production processes and since 2003 a checklist is verified by a third-party certification
entity. In 2008, due to an increased consumer demand for organic chicken meat, the company
started to produce this type of broilers. By the end of 2010, 5 thousand organic broilers were
slaughtered on a monthly basis in addition to the most part of the AF broilers.
3.2 Tapejara (RS)
The municipality of Tapejara is located in the Estate of Rio Grande do Sul and is included
in the hydrological basin of the rivers Apuaê-Inhandava that delimitates the States of Rio Grande
do Sul and Santa Catarina in the south region of Brazil. The main use of soil is to produce
cereals and oil seed crops and the rotation soybean-maize-wheat the most common strategy used
in this agro ecological zone. Livestock production could also be found with chickens, sheep and
dairy herds as the most relevant species. Only small fragments of set-aside forests could be
observed, especially close to rivers and water springs in accordance to national environmental
legislation. The group of family farmers who participated in this study were linked to the
cooperative Coopervita launched in 1991 after strong economic crises that affected Brazil. In

160
that period, 13 smallholder families initiated the farmer association with the first intention to
reach food security. In common areas, not only food for their own subsistence, family farmers
cultivated fruit plots to supply a neighbour cooperative agro industry.
From 1991 to 1994, farmers structured the cooperative and overcome the previous crises.
In this meantime, farmers were favoured by a governmental program to recover soil fertility
where all farms linked to the cooperative have 1/3 of their land recovered1 . In 1995 the
cooperative begun to compensate and pay for the labour work (i.e. woman was paid by working
in the commune bakery either in the communitarian restaurant while man by taking care of
common fields, livestock and plots). In 1993 a pig house was constructed to benefit from a
private company located in the region as another source of income to member farmers.
In 1998 farmers decided to invest and constructed their own agro industry for processing
the harvested fruits in marmalade jams and sweets. Farmers were conscious that the concurrence
in the market was high and to be competitive, they would succeed only if supply high quality
food products. From that period, the farmers begun to market ecological products (however not
certified as organic) in a specific governmental program2 for products originated from family
agriculture.
In 2002, after a basic course on production of broilers in free range systems organized by
the state extension office (EMATER), some farmers from the cooperative begun to produce free
range broilers. After two years producing the broilers, in 2004 the cooperative constructed its
own, small scale slaughter house to benefit from an increased market demand for their products.
3.3 Quatro Barras (PR)
The municipality of Quatro Barras is located in the metropolitan region of Curitiba, capital
of the State of Parana. The municipality is part of an environmental protection area due to the
potential for water catchment to supply the metropolitan area of Curitiba and its neighbour
1 Recovering soil fertility was basically achieved by the application of lime to correct pH; fertilization strategies based on agronomic recommendations and use of agronomic practices to avoid soil erosion like cultivation under the straw and construction of terraces. 2 In 2011 more than 70% of the cooperative income was provided by the supply the national program of Food acquisition (PAA) which guarantees a minimum price for products originated in smallholder production systems and supplemented to municipal schools and hospitals. A premium price of 30% of the market price for a specific product is paid if the product is certified as organic.

161
municipalities. Quatro Barras is part of the hydrological basin of the Iguaçu River and is mainly
occupied by small scale farmers producing vegetables surrounded by preserved forest areas.
The group of farmers who participated in this study were linked to the cooperative
COOAG-QB (Cooperativa Agropecuaria de Quatro Barras) and were producing food products
with the brand “graciosa” in connection with to the touristic potential and to attend visitors with
organic products. The cooperative was established 2005 after an effort from the public sector -
EMATER (State technical assistance office) and the municipality of Quatro Barras - that initially
supported farmers to produce without chemicals in accordance to environmental regulations in
the municipality. In addition, farmers could supply the municipality with free range broilers for
touristic celebrations, parties and other local attractions.
After 2 years, smallholders were stimulated to be organized and to reach independence
from the public sector as their own source of income. The cooperative produces mainly
vegetables and the free range broilers under the brand “Graciosa”. The logistics for processing
the broilers (i.e. transport, slaughterhouse, store freezers etc) are all rented. The group of farmers
are 20 km close to the metropolitan area of Curitiba, capital of the state of Parana, favouring the
market supply. The cooperative begun with 20 farmers, however, in 2010 only 5 farmers were
producing the free range broilers.
4. Perceived benefits and limitations in the production of broilers as part of the alternative
systems
4.1 General benefits
Farmers linked to the broiler company in Ipeúna were generally satisfied with the
production income and more than 70% of the respondents reported that high prices and security
in sales were the major advantages in production (Figure 1). In the group of farmers producing
the free range broilers in the municipality of Tapejara (RS), 60% of the respondents reported
high quality attributes of their products, thus including animal welfare, as the major advantages
of producing the broilers (Figure 1). Farmers in Quatro Barras reported managing the farms and
animals without chemicals and drugs as the major advantage in production (Figure 1).

162
4.2 General limitations
In addition to the major advantages, we investigated the main limitations affecting the
production of broilers among farmers in different locations and under different production
systems. 100% of the farmers in Ipeúna producing AF broilers reported the management of
gastrointestinal diseases as the most important limitation affecting production (Figure 2A). More
than 80% of farmers in Ipeúna reported coccidiosis as a relevant problem in production (Figure 2
B) and associated with necrotic enteritis - that could be clinically observed. The use of a
commercial product based in Oregano oil extract was not capable to control coccidiosis,
understood as the primary pathogen giving opportunities for secondary bacterial infection.
Farmers in Ipeúna were critical in respect to vaccination agansit coccidiosis due to its
inefficiency.
Interestingly, less than 40% of farmers in the group from Ipeúna were satisfied with the
broiler genotype used in production – Cobb (Table 1, Figure 2 C). It was observed that the
reduction in the size of the flocks among case studies (Table 2) was inversely proportional to the
report of coccidiosis as major problem for production (Figures 2 A and 2 B). In the group of
farmers from Quatro Barras with the smallest size of flocks, only 20% of farmers reported
coccidosis as a relevant problem for production (Figure 2 C). In both cases where farmers
produced the free range broilers, coccidiosis was not treated.
Farmers in Quatro Barras have a common practice to use residues from garlic in the
preparation of the litters before the beginning of new flocks as it was a cheap residue available in
the locality. Garlic may reduce infectivity of Eimeria oocysts that cause avian coccidosis. Dkhil
et al (2011) reported reduction in oocyst output in infected mice and Toula and AL-Rawi (2007)
reported anticoccidial activity of garlic extracts in the production of rabbits.
The group of smallholders located in the municipalities of Quatro Barras and in Tapejara
were satisfied with the slow growing genotype they were using in the free range production
(Table 1; Figure 2 C). However, 50% of farmers in the municipality of Tapejara reported the
high cost of diet as the most important limitation for the production of broilers (Figure 2 A).
Even though less than 40% of farmers in Tapejara reported coccidiosis as a relevant problem, we
hypothesise that a subclinical stage of the disease (Fanatico, 2006) could be contributing to
reduce feed efficiency and thus increasing production costs.

163
Most part (80%) of the smallholders located in the municipality of Quatro Barras reported
the absence of a slaughter house as the most important limiting factor for the production of
broilers (Figure 2 A). It is important to notice that groups linked to both cooperative bodies were
controlling the production factors via cooperatives differently from the group of smallholders
from Ipeúna that were vertically integrated, only responsible for feeding and taking care of the
broilers.
4.3 Main Challenges for production
The majority of the farmers (70%) from Ipeúna (SP) reported the investment in equipments
and control of gastrointestinal diseases (coccidiosis and clostridiosis) as the most challenging
factors (Figure 3). It highlights that the farmers were preoccupied in keeping appropriated
equipments in the industrial and intensive system. However, the absence of anti-coccidials and
antibiotics in the production of broilers to avoid gastrointestinal diseases was highlighted. In
contrary, 50% of farmers in Tapejara reported the high production costs, mainly attributed to the
dependence on maize and soybeans in the diet as the most important challenge.
Farmers from Quatro Barras complained about the high production costs in a similar way
as the Tapejara farmers. However, 80% of the farmers from Quatro Barras suggested that the
amount of financial resources used for renting the slaughter house and the logistics with
transport compromised the profit and reported as the most important challenge for keeping the
production.
In the two locations were coccidiosis was reported a problem for production (Ipeúna and
Tapejara – Figure 2 B), farmers suggested that products to be used in treatment of infection
diseases would be more efficient if delivered via drinking water. The main reasons for the
farmers who choose supplementing via drinking water were that mixing the natural products in
the feed could cause degradation of the material as feed silos are normally located outside the
chicken houses and exposed to high temperature variation. In addition, farmers in Ipeúna were
used to supplement the Oregano oil extract in water to control coccidiosis as a routine. In
contrary, other farmers argued that the best strategy of delivery would be supplementing via diet
as it would not give to farmers another work activity.

164
5. Strategies to substitute the use of maize and soybeans in feeding alternative broilers
Farmers in the three case areas were provoked during questionnaire applications to suggest
alternative sources of ingredients to supply nutrients to broilers in replacement strategies. It was
suggested that due to some uncontrolled factors, maize and soybeans should be substituted by
other ingredients to compose the broilers feed. In the case areas where farmers were controlling
the factors affecting the production systems (Tapejara and Quatro Barras), the challenge for
suggesting substitutes for the traditional crops (maize and soybeans) was easily accepted and
suggestions of alternative ingredients based on the local and traditional knowledge were
recorded. Farmers from Ipeúna also suggested other feed ingredients, but did not give much
attention to the topic perhaps due to the characteristic of the integration scheme were the
company supplied the ready-made diets.
In Table 3, four different types of cereal crops were suggested as ingredients to substitute
the use of maize and soybeans. Four other annual species were suggested. Cassava meal was the
most frequent (57%) suggestion as an alternative ingredient to the broilers feed. Sorghum (38%)
and pumpkin meal (19%) were also mentioned by the smallholders in the three case areas.
Sunflower, peanut and faba beans meals have been also suggested (15%; 10% and 5%
respectively). Even though free range broilers were exposed to permanent paddocks with very
little vegetation and plots were not managed with the aim to supply nutrients, when farmers were
informed that part of the broilers nutrition would be possible to be supplemented by forage, a
range of plant species was suggested based on previous observations and their traditional
knowledge. The species of plants suggested were compiled and Table 3 shows a list of plants
suggested by the interviewed farmers.
5.1 Perspectives for innovative production systems
The farmers were also stimulated to respond if integration with other livestock species for
synergistic effects in the farming systems could be an option for income generation. Table 4
presents the main suggestion for livestock integration at farming systems based on common
answers among the groups.

165
6. Conclusions and suggestion to tackle the future challenges
• Farmers in Ipeúna were satisfied with the income generated by producing the AF broilers
and by the agreements made with the company. However, some limitations in production
demand further investigation, especially in the use of slow growing genotypes and to
identify substitute products to reduce problems related to gastrointestinal diseases,
especially coccidiosis and necrotic enteritis.
• Farmers in Tapejara were satisfied with the genotypes used. Farmers reported coccidosis
as a problem but not as a limitation for the production. However, it was necessary to
investigate options to reduce production costs attributed to high costs of dietary
ingredients. As the broilers did not represent a high part of the farmer’s income, they
would voluntarily apply strategies to substitute the use of maize and soybeans in the feed
supply. Alternative ingredients would be further investigated in collaboration with the
farmers, especially cassava meal and residues from other regional crops.
• Farmers in Quatro Barras were extremely dependent on the municipality support. They
did not depend on the production of broilers as the main source of income but the
maintenance of the production of broilers relies on the local demand.
• Farmers from the three groups supported that alternative sources of feed ingredients
should be investigated and a range of alternatives were recommended.
• The supplementation of natural products as an alternative for controlling infectious
diseases should preferably be offered via the drinking water.
Acknowledgements
The authors wish to thank the farmers for receiving the group of scientists during this
research. Lew Springer and Fabiola Ribeiro from the Federal University of Parana and the
technicians Marcio Saatkamp and Levino Bassi from EMBRAPA Swine and Poultry institute are
thanked for the support with the logistics and visits and also for applying the questionnaires in
the South part of Brazil. Evandro Possamai, from Korin agropecuaria Ltd. is thanked for the
logistical and technical support during the field work in the state of São Paulo. Finally Aarhus

166
University and SOAR - Research School for Organic Agriculture and Food Systems - are
thanked for financing our study.
References
ABEF, 2012. Associação Brasileira dos Exportadores de Frango do Brasil - ABEF. Estatísticas. Disponível em: http://www.abef.com.br/Estatisticas/MercadoMundial/MercadoMundial.asp. Acessed Jun 2012. Almeida, G. F. d. and Abreu. L. S. de., 2009. Estratégias produtivas e aplicação de princípios da agroecologia: o caso dos agricultores familiares da Cooperativa dos agropecuaristas solidários de Itápolis - COAGROSOL. Revista de Economia Agrícola 56 (1): 37-53. Almeida, G. F. d., Hinrichsen, L. K., Horsted, K.; Thamsborg, S. M. and Hermansen, J. E. 2012. Feed intake and activity level of two broiler genotypes foraging different types of vegetation in the finishing period. Poultry Science. 91, (9): 2105-2113. Blanc, J. and Kledal, P.R., 2012 The organic sector of Brazil: prospects and constraints of facilitating smallholder inclusion. Journal of Rural Studies 28, 1: 142-154 COM (2008). COM/2008/0233. Report from the Commission to the Council and the European Parliament on the use of coccidiostats and histomonostats as feed additives, submitted pursuant to article 11 of regulation (EC) no. 1831/2003 of the European Parliament and of the Council of 22 September, 2003 on additives for use in animal nutrition, http://www.ipex.eu/ipex/cms/home/Documents/dossier_COM20080233. Demattê Filho, L.C., 2004. Aditivos em dietas para frangos de corte criados em sistema alternativo. Master thesis. State University of São Paulo. Botucatu, SP, 86p. Dkhil MA, Abdel-Baki AS, Wunderlich F, Sies H, Al-Quraishy S. Anticoccidial and anti-inflammatory activity of garlic in murine Eimeria papillata infections. Veterinary Parasitology. 2011;175: 66-72. Dourado, L. R. B., Sakomura, N.K., Nascimento, D. C. N., Dorigam, J. C., Marcato, S. M. and Fernandes, J. B. K. 2009. Crescimento e desempenho de linhagens de aves de pescoço pelado criadas em sistema semiconfinado. Ciênc. agrotec., Lavras, 33: 875-881 Fanatico, A., 2006. Organic poultry production in the United States. ATTRA - National Sustainable Agriculture Information Service. Available at: http://www.attra.org/attra-pub/PDF/coccidiosis.pdf.

167
FAO 2012. Food and Agriculture Organization of the United Nations. Data on production; Livestock Primary (Total Poultry and total chicken meat from 2010). Available at: http://faostat.fao.org/site/569/DesktopDefault.aspx?PageID=569#ancor Access Jun 2012. Francisco, D. C.; Nascimento, V. P.; Loguercio, A. P. and Camargo L., 2007. Caracterização do consumidor de carne de frango da cidade de Porto Alegre Ciência. Rural v.37 n.1 Santa Maria jan./fev. 2007. Herrmann, R. and Roder, C., 1995. Does food consumption converge internationally?: measurement, empirical tests and determinants. Eur Rev Agric Econ, 22, 400 - 414. Knudsen, M. T., Almeida, G. F. d., Langer, V., Abreu, L. S. de; Halberg, N., 2011. Environmental assessment of organic juice imported to Denmark: a case study on oranges (Citrus sinensis) from Brazil. Organic Agriculture. 1: 167-185. Kvale, S. and Brinkmann, S., 2009. InterViews, Learning the Craft of Qualitative Research Interviewing. SAGE Publications. Lopez-Bote CJ, Sanz Arias R, Rey AI, Castano A, Isabel B and Thos J. 1998. Effect of free-range feeding on n-3 fatty acid and α-tocopherol content and oxidative stability of eggs. Animal Feed Science Technology, 72: 33. Luchese, F. C., Perin, M., Aita, R. S., Mottin, V. D., Molento, M. B., Monteiro, S. G., 2007. Prevalência de espécies de Eimeria em frangos de criação industrial e alternativa. Braz. J. Vet. Res. Anim. Sci. 44, 81-86. Martínez Michel, L., Anders, S. and Wismer ,W. V. 2011. Consumer Preferences and Willingness to Pay for Value-Added Chicken Product Attributes. Journal of Food Science 76(8): 469–S477. Molento, M.B., 2009. Parasite control in the age of drug resistance and changing agricultural practices. Veterinary Parasitology 163, 229–234. Oelofse, M., Høgh-Jensen, H., Abreu, L. S. de, Almeida, G. F. d., El-Araby, A. Hui, Q. Y. de Neergaard, T. A. F., 2010. A comparative study of farm nutrient budgets and nutrient flows of certified organic and non-organic farms in China, Brazil and Egypt. Nutrient Cycling in Agroecosystems, 87 (3), 455-470. Oelofse, M., Høgh-Jensen, H., Abreu, L. S. de, Almeida, G. F. d., El-Araby, A. Hui, Q. Y., Sultan, T., de Neergaard, T. A. F., 2010. Certified organic agriculture in China and Brazil: Market accessibility and outcomes following adoption. Ecological Economics, Vol. 69, Nr. 9, 2010, s. 1785-1793. Permin, A., Bisgaard, M., Frandsen, F., Pearman, M., Kold, J. and Nansen, P., 1999. Prevalence of gastrointestinal helminths in different poultry production systems. Br. Poult. Sci., 40, 439-443.

168
Prothero, R.E. 1936. English farming past and present. Heinemann and Frank Cass and Co. London, 1961 ed. First published in 1912. Robinson, L. 1961. Modern Poultry Husbandry. Crosby Lockwood. London. Rodriguez, J. C., Segura, J. C., Alzina, A. and Guterrez, M. A., 1997. Factors affecting mortality of crossbred and exotic chicken kept under backyard systems in Yucatan, Mexico. Trop. Anim. Health Prod. 29, 151-157. Sossidou, E. N., A. Dal Bosco, H. A. Elson, and C. Fontes. 2010. Pasture-based systems for poultry production: Implications and perspectives. World’s Poult. Sci. J. 67:47–58. Tixer-Boichard, M., Leenstra, F., Flock, D.K., Hocking P.M. and Weigend, S., 2012. A century of poultry genetics. World's Poultry Science Journal 68, 307-321.
Toula FH, AL-Rawi MM. Efficacy of garlic extract on hepatic coccidiosis in infected rabbits (Oryctolagus cuniculus): histological and biochemical studies. J. Egypt. Soc. Parasitol. 2007 37:957–969. Tovar, L.G., Martin, L., Cruz, M.A.G., Mutersbaugh, T., 2005. Certified organic agriculture in Mexico: market connections and certification practices in large and small producers. Journal of Rural Studies 21, 461–474. Wier, M., Jensen, K. O., Andersen, L. M., Millock, K., 2008. The character of demand in mature organic food markets: Great Britain and Denmark compared. Food Policy 33: 406–421. Van de Weerd, H. A., R. Keatinge and S. Roderick. 2009. A review of key health-related welfare issues in organic poultry production. World's Poult. Sci. J. 65: 649-684. Yin, R. K., 2003. Case Study Research: Design and Methods, Third Edition. ed. SAGE Publications Inc.

169
Table 1 – Case study site characteristics
Case Study Ipeúna Tapejara Quatro Barras State in Brazil São Paulo (SP) Rio Grande do Sul (RS) Paraná (PR) Geographic location 22° 33’ S 47° 55’ W 28º 01’S 52º 07 ’ W 25° 02’ S 49° 04’ W Altitude (masl) 613 658 948 Precipitation (mm year1) 1500 1900 1500 Mean annual temp (ºC) 21 18 17 Period of data collection Feb-Mar 2012 Jun-Jul 2012 Jul-Sept 2012 Production system and type of broiler
Natural and Organic using fast growing genotypes (Cobb)
Caipira free range using slow growing
genotypes (label rouge necked neck)
Caipira free range using slow growing
genotypes (label rouge necked neck)
Main product outputs by economic relevance for the smallholders
Broilers/eucalyptus vegetables
Crops and cereals (cassava, fruits),
broilers and sheep
Vegetables/broilers
Type of organization Vertical integration by contracted farmers
Cooperative with own agro-industry slaughter house
Cooperative without slaughter house
Total number of collaborative farmers
26 22 20
Main market channels for the meat
Private stores Food boutiques Supermarkets
Local stores; market network and municipality (schools and hospitals)
Municipality (schools and fairs. ecological touristic attractions) and street fairs
Source: Data obtained from the municipality reports and from the research.

170
Table 2 – Characteristics of the interviewed sample for the three case studies (median values)
Case Study Ipeúna (SP) Tapejara (RS)
Quatro Barras (PR)
Number of farmers interviewed 8 8 5 Median size of farms (ha) 5 30 5 Flock runs/year 6 3-4 3 Size of flocks (heads) 22.000 1.800 400 Mean household size (n)* 3 3 3 Education of the HH head (yrs) 9 4.5 5 Age HH head (yrs) 43.5 54 52 Annual income from broilers (%) 65 10 15
Source: Data obtained from the research. * Family members living in the same farm

171
Table 3 – Potential species for alternative supplementation nutrients to broilers
Type of crop Scientific name Popular name Potential use Cereal crop Avena sativa Oat Ingredient for diet Cereal crop Secale cereale Rye Ingredient for diet Cereal crop Orysa sativa Rice meal Ingredient for diet Cereal crop Brassica napus Rapeseed Ingredient for diet Annual crop Manihot esculenta Cassava meal Ingredient for diet Annual herbaceous Vicia faba Faba beans Ingredient for diet Annual herbaceous Arachis hypogaea Peanut Ingredient for diet Annual crop Helianthus annuus Sunflower meal Ingredient for diet Perennial vine Curcubita pepo Pumpkin meal Ingredient for diet Grass Axonopus affinis Carpet grass Close to chicken houses Grass Melinis minutiflora Honey grass Nutritive forage Hybrid grass Cynodon dactylon Bermuda grass Nutritive forage Hybrid grass Cynodon dactylon Bermuda grass Nutritive forage Annual herb Amaranthus viridis Green amaranthus Forage/protection Perennial grass Phalaris canariensis Canaryseed Forage Perennial grass Sorghum bicolor Sorghum Ingredient for diet Perennial grass Pennisetum purpureum Uganda Grass Cut forage/protection Perennial grass Pennisetum purpureum Napier Grass Cut forage/protection Perennial grass Saccharum Sugar cane Cut forage/protection Perennial grass Pennisetum clandestinum kikuyu grass Nutritive forage Perennial Grass Lolium multiflorum Rye grass Nutritive forage Peren. Herbaceous Boehmeria nivea Ramie Nutritive legume Peren. Herbaceous Linum usitatissimum flaxseed/Lenseed Nutritive legume/seed Perennial Legume Leucaena glaca Leucaena Nutritive legume/hay Perennial Legume Neonotonia wightii Perenial soybean Nutr. legume/protection Perennial Legume Cajanus cajan Guandu beans Nutr. legume/protection Perennial Legume Medicago sativa Alfafa Nutritive legume Perennial Legume Trifolium repens White clover Nutr legume /resistant Perennial Legume Trifolium pratense Red clover Nutr legume /resistant Perennial Legume Vicia sativa Common Vetch Nutritive legume Perennial Legume Neonotonia wightii Perenial soybean Nutr. legume/protection
Data compiled from research after farmers opinions

172
Table 4 – Smallholder suggestions for ideal integrated systems
Case study Ideal production systems
Ipeúna (SP) Broilers should be kept indoors. Integration could be interesting if use
eucalyptus trees for the production of wood and ruminant species
foraging in between the trees. Beef cattle were most cited but Goats or
Sheep were also mentioned, attributed to the small size of farms.
Tapejara (RS) The use of agro forestry systems by integrating fruits, trees for wood
and livestock (broilers + ruminants) was the most appreciated
suggestion. Opened fields for the production of grains was necessary to
be maintained in part of the farms. Broilers would be integrated in free-
range areas with sheep or dairy cows side by side.
Quatro barras (PR) Integration of dairy cows or sheep with fruits trees was the most
common option.

173
Figure 1 - List of major advantages for producing the alternative broilers (in % of respondents).

174
Figure 2 - Limitations affecting the production of alternative broilers (A); the role of coccidiosis
in production (B); and the use of appropriated genotypes in the production (C) (in % of
respondents).

175
Figure 3 - The main challenge for keeping producing broilers in alternative systems
The case studies were composed by smallholders from the municipality of Ipeúna in the state of São Paulo (producing Natural broilers – Antibiotic Free); Smallholders from the municipality of Tapejara in the state of Rio Grande do Sul (producing free range broilers) and smallholders in the municipality of Quatro Brarras in the state of Parana (producing free range broilers).




















