
Trophic ecology of reef sharks determined using stable isotopes and telemetry

M
O
Im
DJa
b
c
a
ARAHA
KTETSA
I
vTtaenuWeta
em
B
h1
ARTICLE IN PRESSG ModelAMBIO-40685; No. of Pages 10
Mammalian Biology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Mammalian Biology
jou rn al hom epage: www.elsev ier .com/ locate /mambio
riginal Investigation
nferring spatial and temporal behavioral patterns of free-ranginganatees using saltwater sensors of telemetry tags
elma Nataly Castelblanco-Martíneza,b,∗, Benjamin Morales-Velab, Daniel H. Slonec,anneth Adriana Padilla-Saldívarb, James P. Reidc, Héctor Abuid Hernández-Aranab
Oceanic Society, 30 Sir Francis Drake Boulevard, P.O. Box 437, Ross, CA 94957, USAEl Colegio de la Frontera Sur, Av. Centenario Km. 5.5, CP 77900 Chetumal, Quintana Roo, MexicoSoutheast Ecological Science Center, U.S. Geological Survey, 2201 NW 40th Terrace, Gainesville, FL 32605, USA
r t i c l e i n f o
rticle history:eceived 30 April 2014ccepted 9 July 2014andled by Frank E. Zachosvailable online xxx
eywords:richechus manatusthology
a b s t r a c t
Diving or respiratory behavior in aquatic mammals can be used as indicator of physiological activityand consequently, to infer behavioral patterns. Five Antillean manatees, Trichechus manatus manatus,were captured in Chetumal Bay and tagged with GPS tracking devices. The radios were equipped witha micropower saltwater sensor (SWS), which records the times when the tag assembly was submerged.The information was analyzed to establish individual fine-scale behaviors. For each fix, we establishedthe following variables: distance (D), sampling interval (T), movement rate (D/T), number of dives (N),and total diving duration (TDD). We used logic criteria and simple scatterplots to distinguish betweenbehavioral categories: ‘Travelling’ (D/T ≥ 3 km/h), ‘Surface’ (↓TDD, ↓N), ‘Bottom feeding’ (↑TDD, ↑N) and
elemetryireniansutoecology
‘Bottom resting’ (↑TDD, ↓N). Habitat categories were qualitatively assigned: Lagoon, Channels, Caye shore,City shore, Chanel edge, and Open areas. The instrumented individuals showed a daily rhythm of bot-tom activities, with surfacing activities more frequent during the night and early in the morning. Moreinvestigation into those cycles and other individual fine-scale behaviors related to their proximity toconcentrations of human activity would be informative.
© 2014 Deutsche Gesellschaft für Säugetierkunde. Published by Elsevier GmbH. All rights reserved.
ntroduction
For the last few decades, biotelemetry has proven to be aery useful tool for marine species tracking (Aarts et al., 2008).hese tools offer increasingly sophisticated means – e.g. large-scaleelemetry arrays, fine-scale positioning, and use of physiologicalnd environmental sensors – of evaluating the behavior, spatialcology, energetics, and physiology of free-living animals in theiratural environment (Cooke, 2008). Telemetry has been widelysed to investigate the behavior of seabirds (Adams and Flora, 2010;ilson et al., 2009; Wood et al., 2000), marine mammals (Bailleul
t al., 2007; Baumgartner and Mate, 2005; Mate et al., 1998), rep-iles (Jonsen et al., 2007; McMahon et al., 2007; Read et al., 2007)nd fishes (Mate et al., 1998; Sims et al., 2009).
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
Most of the broad scale studies of manatee radio tracking havemployed VHF radio-transmitters and/or Platform Terminal Trans-itters (PTTs) with the Argos data service (www.argos-system.org)
∗ Corresponding author at: Oceanic Society, 30 Sir Francis Drake Boulevard, P.O.ox 437, Ross, CA 94957, USA. Tel.: +55 9832850209.
E-mail address: [email protected] (D.N. Castelblanco-Martínez).
ttp://dx.doi.org/10.1016/j.mambio.2014.07.003616-5047/© 2014 Deutsche Gesellschaft für Säugetierkunde. Published by Elsevier Gmb
(Deutsch et al., 2003). A major limitation of VHF and PTT is theinfrequent intervals between fixes limiting their application at finerspatial resolutions (Schofield et al., 2007). Another issue is the spa-tial precision of locations, which lack the high resolution accuracyneeded for fine-scale tracking (Hazel, 2009). However, recent radiotracking studies of manatees in Florida and the Caribbean haveutilized Argos-linked Global Positioning System (GPS) tags, whichprovide much more accurate spatial precision.
A large number of studies have addressed the fine-scale habitatselection by marine megafauna species as turtles (Fossette et al.,2010; Schofield et al., 2007; Senko et al., 2010; Yasuda and Arai,2009), cetaceans (Allen et al., 2001; Hastie et al., 2003; Johnstonet al., 2005; MacLeod and Zuur, 2005; Skov and Thomsen, 2008),seals (Burns et al., 2008; Hindell et al., 2002; Lea and Dubroca,2003), sea lions (Wolf and Trillmich, 2007) and dugongs (Sheppardet al., 2006). However, studies of fine-scale behavior of endangeredmanatees (Order Sirenia: Trichechus spp.) are underrepresented inthe literature. Our understanding of fine-scale behavior may be
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
improved by examining the diving behavior and correlating it withphysical features (Burns et al., 2008).
Data loggers recording depth at specific time interval(Time–Depth recorders, TDRs) have been widely used to describe
H. All rights reserved.

ING ModelM
2 Mamm
fie(e(a2efdob
tooewiG1dm
M
S
t(asH7caaiatosabvaM
C
fmaaAbdelswTc
ARTICLEAMBIO-40685; No. of Pages 10
D.N. Castelblanco-Martínez et al. /
ne-scale behavior of depth-diving mammals as cetaceans (Aokit al., 2007; Baird et al., 2002, 2005; Davis et al., 2007), sea ottersBodkin et al., 2007), elephant seals (Campagna et al., 1999; Costat al., 2003; Le Boeuf et al., 1993; Muelbert et al., 2004), sealsBekkby and Bjorge, 2000; Blix and Nordoy, 2007; Frost et al., 2001),s well as in shallow-depth animals as dugongs (Chilvers et al.,004; Churchward, 2001) and turtles (Fossette et al., 2010; Hayst al., 2001a; Hochscheid et al., 2005; Reina et al., 2004). However,or shallow marine mammals as manatees, the use of TDRs to studyiving patterns has been limited due to issues in the interpretationf data (Hagihara et al., 2009), and alternative technical tools shoulde explored.
We aimed to investigate the fine-scale behavior of satellite-agged Antillean manatees in Chetumal Bay. This technology notnly enables animals to be located but can also provide informationn their diving activities (Bailleul et al., 2007). The GPS radios werequipped with a saltwater sensor (SWS), which records the timeshen the tag assembly is submerged. Diving or respiratory behav-
or can be used as an indicator of physiological activity (Read andaskin, 1985), and consequently, to infer behavioral patterns (Boyd,997). Therefore, we used the information obtained from SWS toescribe the diving behavior, and to infer behavioral patterns ofanatees captured in Chetumal Bay.
ethods
tudy area
Chetumal Bay is a large estuary located between Belize andhe southern section of the state of Quintana Roo, Mexico17◦52′–18◦50′ N, 87◦50′–88◦25′ W) (Fig. 1). It covers an area ofbout 2450 km2. The connection with the sea is located in theoutheast section of the bay, and it receives fresh water from theondo River and many smaller creeks. Its depth ranges from 1 to
m, with several nearly circular depressions known as “pozas” thatan attain up to 40 m depth (Carrillo et al., 2009b). With an aver-ge salinity of 13 psu it is considered hyposaline (Gasca et al., 1994)nd has low productivity (Gasca and Castellanos, 1993). The climates tropical with seasonal rains between June and September andn annual precipitation of 1245 mm on average; the mean annualemperature is 27 ◦C with a maximum of 31 ◦C and a minimumf 20 ◦C (Carrillo et al., 2009a). The region has three climatic sea-ons typified by drought (February to May), rains (June to October),nd north winds (November to January) (Carrillo et al., 2009b). Theay is characterized by a low abundance and variety of sub-aquaticegetation (Espinoza-Ávalos et al., 2009), although manatees canlso forage for mangroves and other riparian plants (Castelblanco-artínez et al., 2009b).
apture, tagging and radio-tracking
From March 2006 to May 2007, five Antillean manatees (threeemales and two males) were captured in northwestern Chetu-
al Bay and fitted with radio tags (Table 1). Capture proceduresre explained in Morales-Vela and Padilla-Saldívar (2009). The tagssembly consisted of a buoyant, cylindrical housing containing anrgos-linked GPS tag (TMT-460, Telonics, Inc.) that was attachedy a flexible nylon tether to a belt that fit snugly around the cau-al peduncle (Reid et al., 1995). The tags collected GPS position datavery 20 or 30 min with a measured accuracy of <5 m for >95% of theocations. In this research, the tag also incorporated micro-power
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
altwater sensor (SWS) that functions to suppress transmissionshile the unit is submerged thereby saving power (Telonics, Inc.).
he SWS synchronize uplink transmissions to resume at the pre-ise time when the animal actually breaks the sea surface following
PRESSalian Biology xxx (2014) xxx–xxx
a dive. By transmitting immediately upon resurfacing, the unithas the best chance of successfully getting uplink transmissions toavailable orbiting satellites. The units were recovered at the com-pletion of the study, the GPS locations and sensor data (number andduration of individual tag submergences/dives) were downloadedfrom memory for analysis.
Data analysis
Each manatee was treated as a sampling unit to avoid pseudoreplication (Hurlbert, 1984). The telemetry data were edited toeliminate telemetry fixes before and after tracking time andon land. Points were also eliminated based on excessive speedbetween consecutive fixes. Each fix was associated to a depth valueusing the nearest-neighbor interpolation of ArcMap. Bathymetricinformation was obtained by Laboratory of Physic Oceanographyof ECOSUR (Chetumal), and complementary depth values for fresh-water system of Guerrero Lagoon were collected by the authors.To reduce the effect of shallow depths on data interpretation, fixesobtained at depth values lesser than 2 m were discarded from theanalysis.
The information obtained by the SWS was used to record sub-mergence frequency and duration. The start of the dive was definedby the time that the SWS perceived that the transmitter wassubmerged, and the end of the dive was defined when the SWSrecorded the transmitter breaking the surface (Hays et al., 2004).For each 20 or 30 min interval between the remaining consecu-tive fixes, distance (D) between coordinates, number of dives (N),and total diving duration in seconds (TDD) were used to infer thebehavioral category performed by the individual during the inter-val. Four behavioral categories were broadly defined based on priorknowledge of the species behavior. Long distances between twoconsecutive fixes – i.e. >1000 m (for 20 min interval) or >1500 m(for 30 min interval) – suggested that the manatee was traveling>3 km/h, so the second fix was classified as ‘traveling fast’. For allintervals that were shorter than this distance criterion, the inter-val end point was labeled as idling activities, which were furthersubdivided among behavioral categories using simple logic crite-ria, as shown in Table 2, Fig. 1. Bottom behaviors can be interpretedas resting or feeding. When a manatee is resting on the bottom, itcomes up only a few times to breathe (↓N◦ dives), and the under-water time is high (↑dive duration). When a manatee is feeding onthe bottom, the total dive duration is also high, but since the indi-vidual is more active, it comes to the surface to breathe more often(↑N◦ dives). When a manatee is resting at the surface, the numberof dives is low, and the duration of those dives is shorter (often 0).This pattern could also correspond to feeding or other behaviorsin shallow water. All behaviors calculated from analysis of thesetime intervals were assigned to the second, or ending point of theinterval (Table 2, Fig. 2).
Scatter plots of the data were created first to allow visual assess-ment of the data and to establish cut points for behavioral typesbased on the appearance of the data. The occurrence of behav-ioral categories was expressed as the proportion of fixes whereeach behavior pattern occurred (Martin and Bateson, 1993). Totest if there is a correlation between behavior and ecological vari-ables, habitat categories were qualitatively assigned according tothe locality and general geomorphologic characteristics, and weremutually exclusive, as follow: Lagoon, Channels, Caye shore, Cityshore, Chanel edge, and Open areas. Behavioral frequency wasrepresented in terms of percentages [Fb1 = (nb1/N1) × 100; whereFb1 = frequency of the behavior b in the habitat 1, nb1 = number
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
of fixes assigned to the behavior b in the habitat 1, andN1 = total of fixes in the habitat 1]. The Pearson rank correlation(R2 statistic) method was used to determine a level of corre-lation between the rough behavior category (bottom, surface

Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Inferring spatial and temporal behavioral patterns of free-rangingmanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
ARTICLE IN PRESSG ModelMAMBIO-40685; No. of Pages 10
D.N. Castelblanco-Martínez et al. / Mammalian Biology xxx (2014) xxx–xxx 3
Fig. 1. Area of study.
Table 1Manatees Trichechus manatus manatus tagged with Argos-linked GPS tags in Chetumal Bay.
ID Length (cm) Beginning Ending Tracking time (days) Fixes/day
ID35-M 267 26/03/2006 21/04/2006 26 54.5ID36-M 292 27/03/2006 22/01/2007 301 46.4ID51-F 293 26/04/2007 05/05/2007 9 52.4ID52-F 312 18/05/2007 04/10/2007 139 27.4ID53-F 285 24/05/2007 20/06/2007 27 4.1
Table 2Logical interpretation of the variables (average duration and distance) based on prior knowledge of the species behavior.
Variable Traveling Idling
Bottom resting Surface resting/surface feeding Bottom feeding
Distance (D) ↑ ↓ ↓ ↓Number of dives (ND) – ↓ ↓ ↑Total diving duration (TDD) – ↑ ↓ ↑
Symbols are interpreted as: ↑ = above average duration or distance, ↓ = below average duration, and — = not applicable.

ARTICLE IN PRESSG ModelMAMBIO-40685; No. of Pages 10
4 D.N. Castelblanco-Martínez et al. / Mammalian Biology xxx (2014) xxx–xxx
Fig. 2. Examples of diving patterns corresponding to idling behaviors, obtained from the SWS for a 20-min period. Diving time (s) is showing in black and surface time inwhite. (a) Bottom resting, (b) bottom feeding and (c) surface resting.
oupiacafpu
R
l
r traveling) and the habitat categories. We used the individ-al behavioral pattern as a proxy for the proportion of timeerforming a behavior over the 24-h cycle. Data were stored
n 6 h periods (A = 06:00–12:00; B = 12:00–18:00; C = 18:00–00nd D = 00:00–06:00) roughly corresponding to light/dark dailyycles. Differences between daily segments were compared using
Kruskal–Wallis one-way non-parametric analysis of variance,ollowing by pairwise Mann–Whitney U-testing for multiple com-arisons (Bonferroni-corrected). The analyses were performedsing the software PAST 2.17 (Harper and Ryan, 2001)
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
esults
Manatees were tracked in depths <6 m, often in very shal-ow waters (1–2 m). Depth ranges and manatee’s fixes percentage
for each range were: <2 m, 51.6%; 2–4 m, 34%, and 4–6 m, 14.4%.There were a total of 14,334 fix attempts in Chetumal Bay, withan average failure rate of 44.29 ± 29.05% (x ± SD). In 19.04% ofthe fixes we could not assess the behavior. It was due to the factthat in those cases, the second fix of the segment failed; there-fore the distance between the points was unknown. For the restof the points, manatees performed Surface behaviors 42.9% of thetime, Bottom Resting 32.2%, Traveling 3.94% and Bottom Feed-ing 1.88% of the time. The categories of behavior were variablebetween individuals and were not influenced by the sex (Fig. 3,Table 3).
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
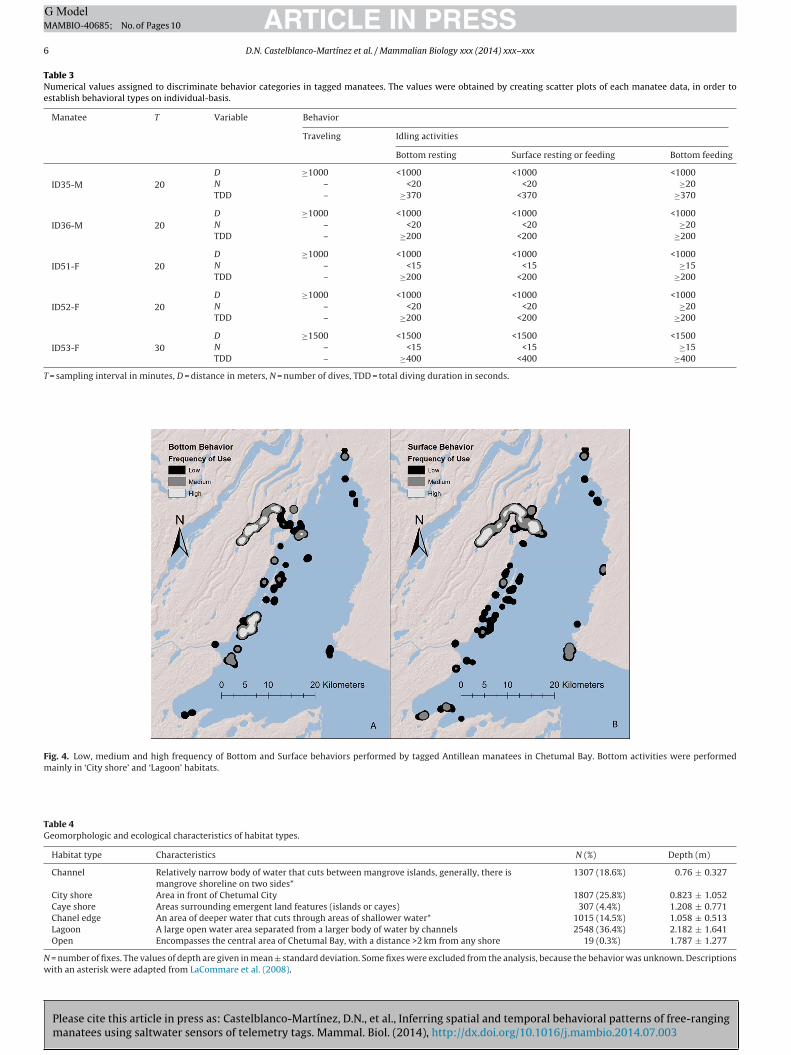
The studied animals performed all four behavioral pat-terns across their home ranges, but some areas showed ahigher frequency of bottom behaviors, specifically in Guer-rero Lagoon (‘Lagoon’), La Barra (‘Chanel edge’) and Chetumal

ARTICLE IN PRESSG ModelMAMBIO-40685; No. of Pages 10
D.N. Castelblanco-Martínez et al. / Mammalian Biology xxx (2014) xxx–xxx 5
Fig. 3. Assignment of behavioral categories for five satellite-tagged manatees, using diving variables and distance between consecutive points. Each point represents ac
CGbtaa(
w1n
oordinate that was assigned a behavioral category.
ity (‘City shore’). Surface behaviors were concentrated in theuerrero Lagoon system (Fig. 4). Nevertheless, interestinglyottom behaviors were relatively more frequent in habi-ats known for highest subaquatic vegetation abundance suchs Caye shores and Channels. In the other hand, in openreas traveling behaviors were relatively more abundant (81%)Table 4).
Manatees showed an apparent daily rhythm of activities in
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
hich bottom behaviors were more frequent between 12:00 and8:00 pm, and surfacing activities occurred more often during theight and early in the morning (Fig. 5).
Discussion
The knowledge of fine-scale behavior is fundamental to under-standing many key aspects of a mammal’s ecology. Even animalswith large home ranges often concentrate their activities on smallpatches where they locate food, mates and shelter (Burt, 1943).When large numbers of animals overlap in range, foraging patchesare likely to be subject to high levels of use, and may be sites
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
of potential competition. Obtaining quantitative measures of fine-scale behavior is therefore the first step to evaluate the extent ofintra-specific resource competition (Hindell et al., 2002). Sirenians
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Inferring spatial and temporal behavioral patterns of free-rangingmanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
ARTICLE IN PRESSG ModelMAMBIO-40685; No. of Pages 10
6 D.N. Castelblanco-Martínez et al. / Mammalian Biology xxx (2014) xxx–xxx
Table 3Numerical values assigned to discriminate behavior categories in tagged manatees. The values were obtained by creating scatter plots of each manatee data, in order toestablish behavioral types on individual-basis.
Manatee T Variable Behavior
Traveling Idling activities
Bottom resting Surface resting or feeding Bottom feeding
ID35-M 20D ≥1000 <1000 <1000 <1000N – <20 <20 ≥20TDD – ≥370 <370 ≥370
ID36-M 20D ≥1000 <1000 <1000 <1000N – <20 <20 ≥20TDD – ≥200 <200 ≥200
ID51-F 20D ≥1000 <1000 <1000 <1000N – <15 <15 ≥15TDD – ≥200 <200 ≥200
ID52-F 20D ≥1000 <1000 <1000 <1000N – <20 <20 ≥20TDD – ≥200 <200 ≥200
ID53-F 30D ≥1500 <1500 <1500 <1500N – <15 <15 ≥15TDD – ≥400 <400 ≥400
T = sampling interval in minutes, D = distance in meters, N = number of dives, TDD = total diving duration in seconds.
Fig. 4. Low, medium and high frequency of Bottom and Surface behaviors performed by tagged Antillean manatees in Chetumal Bay. Bottom activities were performedmainly in ‘City shore’ and ‘Lagoon’ habitats.
Table 4Geomorphologic and ecological characteristics of habitat types.
Habitat type Characteristics N (%) Depth (m)
Channel Relatively narrow body of water that cuts between mangrove islands, generally, there ismangrove shoreline on two sides*
1307 (18.6%) 0.76 ± 0.327
City shore Area in front of Chetumal City 1807 (25.8%) 0.823 ± 1.052Caye shore Areas surrounding emergent land features (islands or cayes) 307 (4.4%) 1.208 ± 0.771Chanel edge An area of deeper water that cuts through areas of shallower water* 1015 (14.5%) 1.058 ± 0.513Lagoon A large open water area separated from a larger body of water by channels 2548 (36.4%) 2.182 ± 1.641Open Encompasses the central area of Chetumal Bay, with a distance >2 km from any shore 19 (0.3%) 1.787 ± 1.277
N = number of fixes. The values of depth are given in mean ± standard deviation. Some fixes were excluded from the analysis, because the behavior was unknown. Descriptionswith an asterisk were adapted from LaCommare et al. (2008).

ARTICLE IN PRESSG ModelMAMBIO-40685; No. of Pages 10
D.N. Castelblanco-Martínez et al. / Mammalian Biology xxx (2014) xxx–xxx 7
F ata wD y cycl
or(
S
i(tdsati0epsp(tardtbCmbt
antd
ig. 5. Daily rhythm of bottom activities of five manatees in Chetumal Bay. D = 00:00–06:00) with periods C and D (in gray) roughly corresponding to dark dail
ften clearly make use of multiple, distinct, core habitats sepa-ated by areas apparently only used for traveling between themSheppard et al., 2006).
patial pattern of activities
Bottom behaviors can be interpreted as resting or feeding activ-ties. Manatees in Chetumal Bay are mainly seagrasses eatersCastelblanco-Martínez et al., 2009b), and it is plausible to assumehat bottom feeding could be driven by the presence and abun-ance of benthic plants. In the near area of Drowned Cayes (Belize),eagrass abundance was an important variable explaining the prob-bility of sighting a manatee (LaCommare et al., 2008). It is knownhat seagrasses, the main component of manatee diet, are foundn small, isolated patches along Chetumal Bay, representing only.07% of the total biomass of the ecosystem (Castelblanco-Martínezt al., 2012). When forage is only available in small, low densityatches, herbivores dedicate more time traveling among feedingites, which results in a low probability of encounter within theatches of vegetation and a dilution of the observed correlationBailey et al., 1996). Heithaus et al. (2006) stated that expected habi-at use under random movement is difficult to assess if resourcesre irregularly distributed and patchy, animals are tracked overelatively short periods, have ill-defined home ranges, or showirectional biases to their movements that are related to habi-at preference. Manatees in estuarine areas are supposed to beottom feeders, but, due to the low abundance of seagrasses inhetumal Bay, they might also forage on riparian vegetation, likeangrove (Castelblanco-Martínez et al., 2009b); and in this case
ottom behaviors not necessarily will be related to feeding activi-ies.
All manatees showed a “bottom” activity peak during, near and
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
fter noon, while surface activities were more frequent during theight and early morning. (Table 5). Our data indicate that the mana-ees showed more surface activities during the night, but we cannotetermine from our data whether the manatees were feeding,
ere stored in 6 h periods (A = 06:00–12:00; B = 12:00–18:00; C = 18:00–00 andes.
resting or socializing during those times. Though we did not rigor-ously test it because of the few animals in our data set, this diurnalpattern is evidence that manatee behavior is not random in time ashas been suggested in earlier publications (Bernier, 1985; Hartman,1979; Parker, 1922). Reynolds (1979) suggested that as manateeslack natural predators and have abundant food, they may spendtheir time feeding, resting and socializing in a random fashion. Inthe other hand, it has been found that dugongs (Dugon dugong)learn diurnal behavioral patterns in response to hunting or harass-ment (Anderson, 1981). Manatees in the Drowned Cayes (Belize)use resting holes more frequently during the day than at night(Bacchus et al., 2009), suggesting that manatees are more activeduring the night; so the authors hypothesized that manatees mightchange their behavioral state in response to approaching vessels.Therefore, the need of energy conservation in response to seasonalchanges in temperature, food availability or even hunting pressureor boating activity, can alter the lack of pattern (Miksis-Olds et al.,2007a,b). In Chetumal Bay, human pressures on manatees are rel-atively scarce, and more research is needed to clarify the origin ofthe suggested daily pattern of activities.
Depth selection
In general, Caribbean and Florida manatees have been reportedin waters shallower than 8 m (Áxis-Arroyo et al., 1998; de Thoisyet al., 2003; Hartman, 1979; Kochman et al., 1985; Morales-Velaet al., 2000; Olivera-Gómez and Mellink, 2005; Paludo, 1997),though this may be reflective of the available depth range wherethey are found rather than a preference. For instance, mana-tees have been spotted in Orinoco Basin in water depths of21.2 m, but probably only during movements between core areas(Castelblanco-Martínez et al., 2009a). Previous research in Chetu-
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
mal Bay showed that habitat use is spatially and temporallyheterogeneous, with a preference for shallow water. However,those studies employed aerial (Morales-Vela et al., 1996; Morales-Vela and Olivera-Gómez, 1994; Olivera-Gómez and Mellink, 2002,

ARTICLE IN PRESSG ModelMAMBIO-40685; No. of Pages 10
8 D.N. Castelblanco-Martínez et al. / Mammalian Biology xxx (2014) xxx–xxx
Table 5Differences in bottom behavior frequency between daily segments (A = 06:00–12:00; B = 12:00–18:00; C = 18:00–00 and D = 00:00–06:00), when compared using aKruskal–Wallis one-way non-parametric analysis of variance (H), following by pairwise Mann–Whitney U-testing for multiple comparisons. Values with asterisk showed tobe significantly different (Bonferroni-corrected *p < 0.017, **p < 0.0033, ***p < 0.00033).
ID35-M ID36-M ID51-F ID52-F ID53-F
H = 30.97; p = 8.47E−07 H = 40.43; p = 6.059E−12 H = 41.78, p = 3.307E−09 H = 40.43, p = 8.604E−09 H = 17.34, p = 0.0002821A–B 0.3258 1.37E−06*** 0.8192 0.3671 0.04417A–C 7.04E−06*** 3.21E−07*** 4.68E−07*** 5.28E−07*** 0.001227*A–D 0.02629 1.39E−06*** 2.765E−04*** 4.82E−06*** 0.0001152***
1E−0758E−01542*
2c(ipsaiuoo
M
dobG(gbsftbgaataroAlimiftcwpwd
dmdriib
B–C 1.33E−05*** 7.85E−07*** 4.2B–D 0.03953 0.006172* 1.6C–D 0.02759 1.133E−04*** 0.0
005) and boat surveys (Áxis-Arroyo et al., 1998). In thoseases, statistical tests were performed only for the shallow zones<4.5 m), because manatees in deeper waters had lower detectabil-ty (Olivera-Gómez and Mellink 2002, 2005), suggesting that thereference for shallow water may have been confounded with aampling bias. In the present study, the same bias may be presents most of the GPS fix failures occurred when the manatees weren deeper water, most probably because the tags would be pullednderwater more often there. The addition of time–depth recordersn future tracked animals would address this potential bias thatccurs with current sampling methodologies.
ethodological constraints
Satellite tracking has proved successfully to document long-istance animal movements. However, once more detailed aspectsf animal behavior are considered, the location accuracy is likely toecome a much more important issue (Hays et al., 2001b). WhilePS loggers are routinely used for terrestrial and aerial animals
Steiner et al., 2000), tracking marine vertebrates with GPS log-ers can be more problematic. This is because infrequent surfacingehaviors limits the time when loggers are available for acquiringatellite signals (Schofield et al., 2007), therefore increasing the fixailure (Vincent et al., 2002). The limited available bandwidth ofhe Argos system severely constrains the amount of data that cane transmitted so that, for example, complete dive profiles cannotenerally be obtained for marine animals (Hays et al., 2001a). Ournalysis is limited to the coordinates’ acquisition, since positionsre needed to estimate distances between points, and conversely,o discriminate between traveling and idling behaviors. Fine-scalenalyses of manatee habitat use and movements may requireestricting the data set to the highest location quality or devel-ping new analytical techniques to incorporate locational error.dditionally, when a tagged manatee occupies shallow areas, satel-
ites are more likely to acquire coordinates because the housings frequently floating. An empirical approach is needed to deter-
ine if fine-scale behaviors in very shallow waters can reliably bedentified. By the other hand, when a manatee is undertaking aast traveling movement, the satellite housing is pulled horizon-ally through the water, and the underwater antenna is unable toommunicate with the satellites (Sheppard et al., 2006). Here, salt-ater switch data is used to infer diving behavior. An additionalroblem is the switch triggering without an actual dive (e.g. a waveashed over the tag). Evaluating and correcting biases affectingata collection is needed.
Argos system’s constraints have led to the use of artificialisplacements of animals carrying data loggers, rather than trans-itters, so that their behavior can be recorded in even more
etail as they travel (Webb et al., 1998). Archival time–depth
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
ecorders (Time–Depth Recorders, TDRs) are suitable for gather-ng large quantities of continuous dive-related behavioral datan air-breathing diving vertebrates. Interpreting dive data in aehavioral context requires identifying specific dive types (feeding,
*** 2.40E−05*** 0.04594*** 0.001248** 0.01723
0.01998 0.7738
traveling, interacting, etc.) based on dive profiles (Bodkin et al.,2007). For shallow marine mammals as sirenians, the use ofTDRs has been limited due to issues in the interpretation of data(Hagihara et al., 2009). However, improving TDR technologies anddata analysis could help in answering unresolved questions aboutmanatee behavior at very fine spatial level.
Acknowledgements
This research was funded by SEMARNAT-CONACyT (Project2002-C01-1128). CONACyT also provided Ph.D. funding for DNC-M.We thank Hilda Chavez for her valuable help in data processing, andto Holger Weissenberger and Laura Carrillo for helping in variousdatabase analyses.
References
Aarts, G., MacKenzie, M., McConnell, B., Fedak, M., Matthiopoulos, J., 2008. Estimatingspace-use and habitat preference from wildlife telemetry data. Ecography 31,140–160.
Adams, J., Flora, S., 2010. Correlating seabird movements with ocean winds: linkingsatellite telemetry with ocean scatterometry. Mar. Biol. 157, 915–929.
Allen, M.C., Read, A.J., Gaudet, J., Sayigh, L.S., 2001. Fine-scale habitat selection offoraging bottlenose dolphins Tursiops truncatus near Clearwater, Florida. Mar.Ecol. Prog. Ser. 222, 253–264.
Anderson, P.K., 1981. The behavior of the dugong (Dugong dugon) in relation toconservation and management. Bull. Mar. Sci. 31, 640–647.
Aoki, K., Amano, M., Yoshioka, M., Mori, K., Tokuda, D., Miyazaki, N., 2007. Diel divingbehavior of sperm whales off Japan. Mar. Ecol. Prog. Ser. 349, 277–287.
Áxis-Arroyo, J., Morales-Vela, B., Torruco-Gómez, D., Vega-Cendejas, M.E., 1998. Fac-tors associated with habitat use by the Caribbean manatee (Trichechus manatus),in Quintana Roo, Mexico (Mammalia). Rev. Biol. Trop. 46, 791–803.
Bacchus, M.L.C., Dunbar, S.G., Self-Sullivan, C., 2009. Characterization of resting holesand their use by the Antillean manatee (Trichechus manatus manatus) in theDrowned Cayes, Belize. Aquat. Mamm. 35, 62–71.
Bailey, D.W., Gross, J.E., Laca, E.A., Rittenhouse, L.R., Coughenour, M.B., Swift, D.M.,Sims, P.L., 1996. Mechanisms that result in large herbivore grazing distributionpatterns. J. Range Manage. 49, 386–400.
Bailleul, F., Charrassin, J.B., Monestiez, P., Roquet, F., Biuw, M., Guinet, C., 2007.Successful foraging areas of southern elephant seals from Kerguelen Islands inrelation to oceanographic conditions. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 362,2169–2181.
Baird, R.W., Borsani, J.F., Hanson, M.B., Tyack, P.L., 2002. Diving and night-timebehavior of long-finned pilot whales in the Ligurian Sea. Mar. Ecol. Prog. Ser.237, 301–305.
Baird, R.W., Hanson, M.B., Dill, L.M., 2005. Factors influencing the diving behaviourof fish-eating killer whales: sex differences and diel and interannual variationin diving rates. Can. J. Zool. 83, 257–267.
Baumgartner, M., Mate, B., 2005. Summer and fall habitat of North Atlantic rightwhales (Eubalaena glacialis) inferred from satellite telemetry. Can. J. Aquat. Sci.62, 527–543.
Bekkby, T., Bjorge, A., 2000. Diving behaviour of harbour seal Phoca vitulina pupsfrom nursing to independent feeding. J. Sea Res. 44, 267–275.
Bernier, B.J., 1985. Comparison of Differences in Behaviors of Captive-Born andNatural-Born Manatees (Trichechus manatus) as a Measure of Adjustment to aContained Natural Environment. Biological Sciences, Florida Institute of Tech-nology, pp. 62.
Blix, A., Nordoy, E., 2007. Ross seal (Ommatophoca rossii) annual distribution, divingbehaviour, breeding and moulting, off Queen Maud Land, Antarctica. Polar Biol.
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
30, 1449–1458.Bodkin, J., Monson, D.H., Esslinger, G.G., 2007. Activity budgets derived from
time–depth recorders in a diving mammal. J. Wildl. Manage. 71, 2034–2044.Boyd, I.L., 1997. The behavioural and physiological ecology of diving. Trends Ecol.
Evol. 12, 213–217.

ING ModelM
Mamm
B
B
C
C
C
C
C
C
C
C
C
C
D
d
D
E
F
F
G
G
H
H
H
H
H
H
ARTICLEAMBIO-40685; No. of Pages 10
D.N. Castelblanco-Martínez et al. /
urns, J.M., Hindell, M.A., Bradshaw, C.J.A., Costa, D.P., 2008. Fine-scale habitat selec-tion of crabeater seals as determined by diving behavior. Deep Sea Res. II: Top.Stud. Oceanogr. 55, 500–514.
urt, W.H., 1943. Territoriality and home range concepts as applied to mammals. J.Mammal. 24, 346–352.
ampagna, C., Fedak, M.A., McConnell, B.J., 1999. Post-breeding distribution and div-ing behavior of adult male southern elephant seals from Patagonia. J. Mammal.80, 1341–1352.
arrillo, L., Palacios-Hernández, E., Ramírez, A.M., Morales-Vela, B., 2009a. Carac-terísticas hidrometeorológicas y batimétricas. In: Espinoza-Ávalos, J., Islebe,G., Hernández-Arana, H. (Eds.), El sistema ecológico de la Bahía de Chetu-mal/Corozal: costa occidental del mar Caribe. El Colegio de la Frontera Sur,Chetumal, Quintana Roo, México, pp. 12–20.
arrillo, L., Palacios-Hernández, E., Yescas, M., Ramírez-Manguilar, A., 2009b. Spatialand seasonal patterns of salinity in a large and shallow tropical estuary of theWestern Caribbean. Estuar. Coasts 32, 906–916.
astelblanco-Martínez, D.N., Barba, E., Schmitter-Soto, J.J., Hernández-Arana, H.A.,Morales-Vela, B., 2012. The trophic role of the endangered Caribbean manateeTrichechus manatus; in an estuary with low abundance of seagrass. Estuar. Coasts35, 60–77.
astelblanco-Martínez, D.N., Bermúdez-Romero, A.L., Gómez-Camelo, I.V., Rosas,F.C.W., Trujillo, F., Zerda-Ordonez, E., 2009a. Seasonality of habitat use, mor-tality and reproduction of the Vulnerable Antillean manatee Trichechus manatusmanatus in the Orinoco River, Colombia: implications for conservation. Oryx 43,235–242.
astelblanco-Martínez, D.N., Morales-Vela, B., Hernández-Arana, H.A., Padilla-Saldívar, J., 2009b. Diet of manatees Trichechus manatus manatus in ChetumalBay, Mexico. Latin Am. J. Aquat. Mammals 7, 39–46.
ooke, S.J., 2008. Biotelemetry and biologging in endangered species research andanimal conservation: relevance to regional, national, and IUCN Red List threatassessments. Endanger. Species Res. 4, 165–185.
osta, D.P., Crocker, D.E., Gedamke, J., Webb, P.M., Houser, D.S., Blackwell, S.B.,Waples, D., Hayek, L.A.C., Boeuf, B.J.L., 2003. The effect of a low-frequency soundsource (acoustic thermometry of the ocean climate) on the diving behavior ofjuvenile northern elephant seal, Mirounga angustirostris. J. Acoust. Soc. Am. 113,1155–1165.
hilvers, B., Delean, S., Gales, N.J., Holley, D.K., Lawler, I.R., Marsh, H., Preen, A.R.,2004. Diving behaviour of dugongs, Dugong dugon. J. Exp. Mar. Biol. Ecol. 304,203–224.
hurchward, C.A., 2001. The Effect of Depth and Activity Type on Dugong (Dugongdugon) Diving Behaviour in Shark Bay, Western Australia. Department of Bio-logical Sciences, The University of Calgary, Calgary, Alberta, pp. 221.
avis, R.W., Jaquet, N., Gendron, Markaida, U., Bazzino, G., Gilly, W., 2007. Div-ing behavior of sperm whales in relation to behavior of a major prey species,the jumbo squid, in the Gulf of California, Mexico. Mar. Ecol. Prog. Ser. 333,291–302.
e Thoisy, B., Spiegelberger, T., Rousseau, S., Talvy, G., Vogel, I., Vie, J.C., 2003. Distri-bution, habitat, and conservation status of the West Indian manatee Trichechusmanatus in French Guiana. Oryx 37, 431–436.
eutsch, C.J., Reid, J.P., Bonde, R.K., Easton, D.E., Kochman, H.I., O’Shea, T.J., 2003. Sea-sonal movements, migratory behavior and site fidelity of West Indian manateesalong the Atlantic Coast of the United States. Wildl. Monogr. 151, 1–77.
spinoza-Ávalos, J., Hernández-Arana, H.A., Álvarez-Legorreta, T., Quan-Young, L.I.,Oliva-Rivera, J.J., Valdez-Hernández, M., Zavalamendoza, A., Cruz-Pinón, G.,López, C.Y., Sepúlveda-Lozada, A., Worumference, P., Villegas-Castillo, A., VanTussenbroek, B.I., 2009. Vegetación acuática sumergida. In: Espinoza-Ávalos,J., Islebe, G., Hernández-Arana, H. (Eds.), El sistema ecológico de la Bahía deChetumal/Corozal: costa occidental del mar Caribe. El Colegio de la FronteraSur, Chetumal, Quintana Roo, México, pp. 148–158.
ossette, S., Hobson, V.J., Girard, C., Calmettes, B., Gaspar, P., Georges, J.-Y., Hays,G.C., 2010. Spatio-temporal foraging patterns of a giant zooplanktivore, theleatherback turtle. J. Mar. Syst. 81, 225–234.
rost, K.J., Simpkins, M.A., Lowry, L.F., 2001. Diving behavior of subadult and adultharbor seals in Prince William Sound, Alaska. Mar. Mamm. Sci. 17, 813–834.
asca, R., Castellanos, I., 1993. Zooplancton de la Bahía de Chetumal, Mar Caribe,México. Rev. Biol. Trop. 41, 619–625.
asca, R., Suárez-Morales, E., Vásquez-Yeomans, L., 1994. Estudio comparativo delzooplancton (biomasa y composición) en dos bahías del Mar Caribe Mexicano.Rev. Biol. Trop. 42, 595–604.
agihara, R., Marsh, H., Jones, R.E., 2009. Developing methods to understand divingbehavior of marine mammals in shallow waters. In: 18th Biennial Conferenceon the Biology of Marine Mammals, Quebec, Canada, p. 120.
arper, D., Ryan, P., 2001. PAST: paleontological statistics software package for edu-cation and data analysis. Palaeontol. Electronica 4, 1–9.
artman, D.S., 1979. Ecology and Behavior of the Manatee (Trichechus manatus) inFlorida. Special Publication No. 5, Lawrence, Kansas.
astie, G.D., Wilson, B., Thompson, P.M., 2003. Fine-scale habitat selection by coastalbottlenose dolphins: application of a new land-based video-montage technique.Can. J. Zool. 81.
ays, G.C., Akesson, S., Broderick, A.C., Glen, F., Godley, B.J., Luschi, P., Martin, C.,Metcalfe, J.D., Papi, F., 2001a. The diving behaviour of green turtles undertaking
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
oceanic migration to and from Ascension Island: dive durations, dive profilesand depth distribution. J. Exp. Biol. 204, 4093–4098.
ays, G.C., Arkesson, S., Godley, B.J., Luschi, P., Santidrian, P., 2001b. The implica-tions of location accuracy for the interpretation of satellite-tracking data. Anim.Behav. 61, 1035–1040.
PRESSalian Biology xxx (2014) xxx–xxx 9
Hays, G.C., Houghton, J.D., Isaacs, C., King, R.S., Lloyd, C., Lovell, P., 2004. First recordsof oceanic dive profiles for leatherback turtles, Dermochelys coriacea, indicatebehavioural plasticity associated with long-distance migration. Anim. Behav.67, 733–743.
Hazel, J., 2009. Evaluation of fast-acquisition GPS in stationary tests and fine-scaletracking of green turtles. J. Exp. Mar. Biol. Ecol. 374, 58–68.
Heithaus, M.R., Hamilton, I.M., Wirsing, A.J., Dill, L.M., 2006. Validation of a random-ization procedure to assess animal habitat preferences: microhabitat use of tigersharks in a seagrass ecosystem. J. Anim. Ecol. 75, 666–676.
Hindell, M.A., Harcourt, R., Waas, J.R., Thompson, D., 2002. Fine-scale three-dimensional spatial use by diving, lactating female Weddell seals Leptonychotesweddellii. Mar. Ecol. Prog. Ser. 242, 275–284.
Hochscheid, S., Bentivegna, F., Hays, G.C., 2005. First records of dive durations for ahibernating sea turtle. Biol. Lett. 1, 82–86.
Hurlbert, S.H., 1984. Pseudoreplication and the design of ecological field experi-ments. Ecol. Monogr. 54, 187–211.
Johnston, D.W., Westgate, A.J., Read, A.J., 2005. Effects of fine-scale oceanographicfeatures on the distribution and movements of harbour porpoises Phocoenaphocoena in the Bay of Fundy. Mar. Ecol. Prog. Ser., 295.
Jonsen, I.D., Myers, R.A., James, M.C., 2007. Identifying leatherback turtle foragingbehaviour from satellite telemetry using a switching state-space model. Mar.Ecol. Prog. Ser. 337, 255–264.
Kochman, H.I., Galen, G.B., Powell, J.A., 1985. Temporal and spatial distribution ofmanatees in Kings Bay, Crystal River, Florida. J. Wildl. Manage. 49, 921–924.
LaCommare, K.S., Self-Sullivan, C., Brault, S., 2008. Distribution and habitat use ofAntillean manatees (Trichechus manatus manatus) in the Drowned Cayes area ofBelize, Central America. Aquat. Mamm. 34, 35–43.
Le Boeuf, B.J.L., Crocker, D.E., Blackwell, S.B., Morris, P.A., Thorson, P.H., 1993. Sexdifferences in diving and foraging behaviour of northern elephant seals. In:Symposium of the Zoological Society of London, pp. 149–178.
Lea, M.-A., Dubroca, L., 2003. Fine-scale linkages between the diving behaviour ofAntarctic fur seals and oceanographic features in the southern Indian Ocean.ICES J. Mar. Sci. 60, 990–1002.
MacLeod, C.D., Zuur, A.F., 2005. Habitat utilization by Blainville’s beaked whales offGreat Abaco, northern Bahamas, in relation to seabed topography. Mar. Biol. 147,1–11.
Martin, P., Bateson, P., 1993. Measuring Behaviour. An Introductory Guide.Cambridge University Press, New York.
Mate, B.R., Gisiner, R., Mobley, J., 1998. Local and migratory movements of Hawaiianhumpback whales tracked by satellite telemetry. Can. J. Zool. 76, 863–868.
McMahon, C.R., Bradshaw, C.J., Hays, G.C., 2007. Satellite tracking reveals unusualdiving characteristics for a marine reptile, the olive ridley turtle Lepidochelysolivacea. Mar. Ecol. Prog. Ser. 329, 239–252.
Miksis-Olds, J.L., Donaghay, P.L., Miller, J.H., Tyack, P.L., Nystrue, J.A., 2007a. Noise-level correlates with manatee use of foraging habitats. J. Acoust. Soc. Am. 121,3011–3020.
Miksis-Olds, J.L., Donaghay, P.L., Miller, J.H., Tyack, P.L., Reynolds, J.E., 2007b. Sim-ulated vessel approaches elicit differential responses from manatees. Mar.Mamm. Sci. 23, 629–649.
Morales-Vela, B., Olivera-Gomez, D., Ramirez-García, P., 1996. Conservación de losmanatíes en la región del Caribe de México y Belice. El Colegio de la FronteraSur, Chetumal, Quintana Roo, pp. 131.
Morales-Vela, B., Olivera-Gómez, L.D., 1994. Distribución espacial y estimaciónpoblacional de los manatíes en la Bahía de Chetumal, Quintana Roo, México.Rev. Inv. Cient. 2, 27–34.
Morales-Vela, B., Olivera-Gomez, L.D., Reynolds III, J.E., Rathbun, G.B., 2000. Distri-bution and habitat use by manatees (Trichechus manatus manatus) in Belize andChetumal Bay, Mexico. Biol. Conserv. 95, 67–75.
Morales-Vela, B., Padilla-Saldívar, J., 2009. Demografía, ecología y salud de lapoblación de manatíes (Trichechus manatus manatus) en Quintana Roo, y suvariación y representación genética en México, Informe Técnico Final. ProyectoSEMARNAT/CONACYT 2002-C01-1128, 273 pp.
Muelbert, M.M.C., Robaldo, R.B., Martínes, P.E., Colares, E.P., Bianchini, A., Setzer, A.,2004. Movement of southern elephant seals (Mirounga leonina L.) from ElephantIs. South Shetlands, Antarctica. Braz. Arch. Biol. Technol. 47, 461–467.
Olivera-Gómez, L.D., Mellink, E., 2002. Spatial and temporal variation in counts ofthe Antillean manatee (Trichechus m. manatus) during distribution surveys atBahia de Chetumal, Mexico. Aquat. Mamm. 28, 285–293.
Olivera-Gómez, L.D., Mellink, E., 2005. Distribution of the Antillean manatee(Trichechus manatus manatus) as a function of habitat characteristics, in Bahiade Chetumal, Mexico. Biol. Conserv. 121, 127–133.
Paludo, D., 1997. Estudos sobre a ecologia e conservac ão do peixe-boi marinhoTrichechus manatus manatus no nordeste do Brasil. Centro de Ciências Exatas e daNatureza. Curso de Pós-graduac ão em Ciências Biológicas. Área de Concentrac ãoZoologia. Universidade Federal da Paraíba, João Pessoa, Paraíba, Brasil, pp. 94.
Parker, G.H., 1922. The breathing of the Florida manatee (Trichechus latirostris). J.Mammal. 3, 127–135.
Read, A.J., Gaskin, D.E., 1985. Radio tracking the movements and activities of harborporpoises, Phocoena phocoena (L.) in the Bay of Fundy, Canadá. Fish. Bull. 83,543–552.
Read, M.A., Grigg, G.C., Irwin, S.R., Shanahan, D., Franklin, C.E., 2007. Satellite track-
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
ing reveals long distance coastal travel and homing by translocated estuarinecrocodiles, Crocodylus porosus. PLoS ONE 2, e949.
Reid, J.P., Bonde, R.K., O’Shea, T.J., 1995. Reproduction and mortality of radio-taggedand recognizable manatees on the Atlantic Coast of Florida. In: O’Shea, T.J., Ack-erman, B.B., Percival, H.F. (Eds.), Population Biology of the Florida Manatee.

ING ModelM
1 Mamm
R
RS
S
S
S
S
Wood, A.G., Naef-Daenzer, B., Prince, P.A., Croxall, J.P., 2000. Quantifying habitat
ARTICLEAMBIO-40685; No. of Pages 10
0 D.N. Castelblanco-Martínez et al. /
National Biological Service Information and Report 1 (289 pp.). US Departmentof the Interior, National Biological Service, Washington, DC, USA, pp. 171–191.
eina, R.D., Abernathy, K.J., Marshall, G.J., Spotila, J.R., 2004. Respiratory frequency,dive behaviour and social interactions of leatherback turtles, Dermochelys cori-acea during the inter-nesting interval. J. Exp. Mar. Biol. Ecol. 316, 1–16.
eynolds III, J.E., 1979. The semisocial manatee. Nat. Hist. 88, 44–53.chofield, G., Bishop, C.M., MacLean, G., Brown, P., Baker, M., Katselidis, K.A.,
Dimopoulos, P., Pantis, J.D., Hays, G.C., 2007. Novel GPS tracking of sea turtles asa tool for conservation management. J. Exp. Mar. Biol. Ecol. 347, 58–68.
enko, J., Koch, V., Megill, W.M., Carthy, R.R., Templeton, R.P., Nichols, W.J., 2010. Finescale daily movements and habitat use of East Pacific green turtles at a shallowcoastal lagoon in Baja California Sur, Mexico. J. Exp. Mar. Biol. Ecol. 391, 92–100.
heppard, J.K., Preen, A.R., Marsh, H., Lawler, I.R., Whiting, S.D., Jones, R.E., 2006.Movement heterogeneity of dugongs, Dugong dugon (Muller), over large spatialscales. J. Exp. Mar. Biol. Ecol. 334, 64–83.
Please cite this article in press as: Castelblanco-Martínez, D.N., et al., Imanatees using saltwater sensors of telemetry tags. Mammal. Biol. (2
ims, D.W., Queiroz, N., Doyle, T.K., Houghton, J.D.R., Hays, G.C., 2009. Satellite track-ing of the World’s largest bony fish, the ocean sunfish (Mola mola L.) in the NorthEast Atlantic. J. Exp. Mar. Biol. Ecol. 370, 127–133.
kov, H., Thomsen, F., 2008. Resolving fine-scale spatio-temporal dynamics in theharbour porpoise Phocoena phocoena. Mar. Ecol. Prog. Ser. 373, 173–186.
PRESSalian Biology xxx (2014) xxx–xxx
Steiner, I., Bürgi, C., Werffeli, S., Dell’Omo, G., Valenti, P., Tröster, G., Wolfer, D.P.,Lipp, H.-P., 2000. A GPS logger and software for analysis of homing in pigeonsand small mammals. Physiol. Behav. 71, 589–596.
Vincent, C., Mcconnell, B.J., Ridoux, V., Fedak, M.A., 2002. Assessment of Argos loca-tion accuracy from satellite tags deployed on captive gray seals. Mar. Mamm.Sci. 18, 156–166.
Webb, P., Crocker, D., Blackwell, S., Costa, D., Boeuf, B., 1998. Effects of buoyancy onthe diving behavior of northern elephant seals. J. Exp. Biol. 201, 2349–2358.
Wilson, L.J., McSorley, C.A., Gray, C.M., Dean, B.J., Dunn, T.E., Webb, A., Reid, J.B., 2009.Radio-telemetry as a tool to define protected areas for seabirds in the marineenvironment. Biol. Conserv. 142, 1808–1817.
Wolf, J.B.W., Trillmich, F., 2007. Beyond habitat requirements: individual fine-scalesite fidelity in a colony of the Galapagos sea lion (Zalophus wollebaeki) createsconditions for social structuring. Oecologia 152, 553–567.
nferring spatial and temporal behavioral patterns of free-ranging014), http://dx.doi.org/10.1016/j.mambio.2014.07.003
use in satellite-tracked pelagic seabirds: application of kernel estimation toalbatross locations. J. Avian Biol. 31, 278–286.
Yasuda, T., Arai, N., 2009. Fine-scale tracking of marine turtles using GPS-Argos PTTs.Zool. Sci. 22, 547–553.
Copyright © 2022 FDOKUMEN




















