
Induction of serum- and glucocorticoid-induced kinase-1 (SGK1) by cAMP regulates increases in...
11
Induction of Serum- and Glucocorticoid-Induced Kinase-1 (SGK1) by cAMP Regulates Increases in a-ENaC MARGARITA M. VASQUEZ, * ROBERT CASTRO, STEVEN R. SEIDNER, BARBARA M. HENSON, DANIEL J. ASHTON, AND SHAMIMUNISA B. MUSTAFA Department of Pediatrics/Division of Neonatology, University of Texas Health Science Center, San Antonio, Texas a-ENaC expression and activity is regulated by a variety of hormones including b-adrenergic agonists via the second messenger cAMP. We evaluated the early intermediate pathways involved in the up-regulation of SGK1 by DbcAMP and whether SGK1 is a prerequisite for induction of a-ENaC expression. Submandibular gland epithelial (SMG-C6) cells treated with DbcAMP (1 mM) induced both SGK1 mRNA and protein expression. DbcAMP-stimulated SGK1 mRNA expression was decreased by actinomycin D and mRNA and protein expressions were attenuated by PKA inhibitors (H-89 and KT5720). Inhibition of PI3-K with either LY294002 or dominant negative PI3-K reduced DbcAMP-stimulated SGK1 protein and mRNA levels, attenuated the phosphorylation of CREB (a cAMP-activated transcription factor) and decreased a-ENaC protein levels and Na þ transport. In addition, the combination of PKA inhibitors with dominant negative PI3-K synergistically inhibited DbcAMP-induced Na þ transport. Inhibition of SGK1 expression by siRNA decreased but did not obliterate DbcAMP-induced a-ENaC expression. Thus, in a cell line which endogenously exhibits minimal a-ENaC expression, induction of SGK1 by DbcAMP occurs via the PI3-K and PKA pathways. Increased a-ENaC levels and function are partly dependent upon the early induction of SGK1 expression. J. Cell. Physiol. 217: 632–642, 2008. ß 2008 Wiley-Liss, Inc. Na þ transport in various cells or tissues via epithelial Na þ channels (ENaC) is an important regulator of fluid homeostasis. In fluid absorptive epithelia, ENaC is located in apical membranes and comprises the rate-limiting step for Na þ entry into the cell (Matalon and O’Brodovich, 1999). ENaC has been identified in the epithelia of the lung, distal nephron, colon, and salivary glands (North, 1996; Garty and Palmer, 1997; Eaton et al., 2004). Although three homologous protein subunits (a, b, and g) form this amiloride-sensitive ion channel, the a-subunit is essential for Na þ transport particularly in the lung (Canessa et al., 1994; Snyder, 2000). ENaC activity and expression of each subunit is tightly regulated by a variety of hormones, including b 2 -adrenergic agonists, vasopressin, mineralocorticoids, and glucocorticoids (Berthiaume et al., 1987; Finley et al., 1998; Dagenais et al., 2001; Itani et al., 2002; Matthay et al., 2002). b 2 -adrenergic-stimulated ENaC expression and function are partly regulated via the second messenger, cAMP (Walters et al., 1990; Berthiaume, 1991; Stephens et al., 1998; Niisato et al., 1999; Matthay et al., 2002). Exogenous cAMP stimulates amiloride-sensitive Na þ transport in part by modulating ENaC subunit gene expression (Dagenais et al., 2001; Chen et al., 2002; Itani et al., 2002; Thomas et al., 2004). cAMP also stimulates Na þ transport by increasing the translocation of ENaC subunits, in particular the a-subunit, to the apical membrane surface (Snyder, 2000; Morris and Schafer, 2002; Thomas et al., 2004). Similar to the effects by aldosterone in renal epithelial cells, cAMP-stimulated increase in ENaC activity likely involves additional regulatory protein(s) induced early during the cascade of signaling events. One such candidate protein is the serum- and glucocorticoid-inducible kinase 1 (SGK1), a serine/ threonine-specific protein kinase originally described as an immediate early response gene in rat mammary epithelial cells (Webster et al., 1993; Alvarez et al., 1999). SGK1 co-expression with a-, b-, and g-ENaC subunits in Xenopus oocytes or A6 renal epithelial cells increased Na þ transport by increasing the apical membrane expression of ENaC (Alvarez et al., 1999; Chen et al., 1999; Naray-Fejes-Toth et al., 1999; Alvarez and Canessa, 2003). Inhibition of endogenous SGK1 by antisense oligonucleotide specific for SGK1 or dominant negative SGK1 protein decreased steroid-induced stimulation of Na þ transport (Faletti et al., 2002; Boyd and Naray-Fejes-Toth, 2005). Furthermore, in the presence of a SGK1 kinase-dead dominant negative construct, a-ENaC mRNA expression was decreased in renal cells (Boyd and Naray-Fejes-Toth, 2005). Previously, we have reported that rat submandibular gland (SMG-C6) epithelial cells cultured in modified media devoid of hydrocortisone, endogenously express negligible levels of a-ENaC mRNA and protein and demonstrate minimal amiloride-sensitive transepithelial Na þ transport (Mustafa et al., 2008). In DbcAMP-treated SMG-C6 cells, increased a-ENaC mRNA expression, protein formation, and amiloride-sensitive transepithelial Na þ transport was measured at 24 h (Mustafa et al., 2008). Since elevating intracellular cAMP concentration induces SGK1 protein expression in granulosa cells and increases activation of recombinant SGK1 expressed in COS7 cells (Gonzalez-Robayna et al., 2000; Perrotti et al., 2001), we examined the effects of DbcAMP on the expression of SGK1 and specific signaling pathways that in turn may regulate a-ENaC expression and function. Our present findings indicate that treatment of SMG-C6 epithelial cells with DbcAMP induces SGK1 mRNA and protein expression in a time-dependent manner via the protein kinase A (PKA) and phosphatidyl inositol 3-kinase (PI3-K) pathways and inhibition of SGK1 expression by *Correspondence to: Margarita M. Vasquez, Department of Pediatrics/Division of Neonatology, University of Texas Health Science Center, 7703 Floyd Curl Drive, San Antonio, TX 78229. E-mail: [email protected] Received 15 October 2007; Accepted 10 June 2008 Published online in Wiley InterScience (www.interscience.wiley.com.), 9 July 2008. DOI: 10.1002/jcp.21534 ORIGINAL ARTICLE 632 Journal of Journal of Cellular Physiology Cellular Physiology ß 2008 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Induction of serum- and glucocorticoid-induced kinase-1 (SGK1) by cAMP regulates increases in...

ORIGINAL ARTICLE 632J o u r n a l o fJ o u r n a l o f
CellularPhysiologyCellularPhysiology
Induction of Serum- andGlucocorticoid-Induced Kinase-1(SGK1) by cAMP RegulatesIncreases in a-ENaC
MARGARITA M. VASQUEZ,* ROBERT CASTRO, STEVEN R. SEIDNER, BARBARA M. HENSON,DANIEL J. ASHTON, AND SHAMIMUNISA B. MUSTAFA
Department of Pediatrics/Division of Neonatology, University of Texas Health Science Center, San Antonio, Texas
a-ENaC expression and activity is regulated by a variety of hormones includingb-adrenergic agonists via the second messenger cAMP. Weevaluated the early intermediate pathways involved in the up-regulation of SGK1 by DbcAMP and whether SGK1 is a prerequisite forinduction ofa-ENaC expression. Submandibular gland epithelial (SMG-C6) cells treated with DbcAMP (1 mM) induced both SGK1 mRNAand protein expression. DbcAMP-stimulated SGK1 mRNA expression was decreased by actinomycin D and mRNA and proteinexpressions were attenuated by PKA inhibitors (H-89 and KT5720). Inhibition of PI3-K with either LY294002 or dominant negative PI3-Kreduced DbcAMP-stimulated SGK1 protein and mRNA levels, attenuated the phosphorylation of CREB (a cAMP-activated transcriptionfactor) and decreased a-ENaC protein levels and Naþ transport. In addition, the combination of PKA inhibitors with dominant negativePI3-K synergistically inhibited DbcAMP-induced Naþ transport. Inhibition of SGK1 expression by siRNA decreased but did not obliterateDbcAMP-induceda-ENaC expression. Thus, in a cell line which endogenously exhibits minimala-ENaC expression, induction of SGK1 byDbcAMP occurs via the PI3-K and PKA pathways. Increased a-ENaC levels and function are partly dependent upon the early induction ofSGK1 expression.
J. Cell. Physiol. 217: 632–642, 2008. � 2008 Wiley-Liss, Inc.
*Correspondence to: Margarita M. Vasquez, Department ofPediatrics/Division of Neonatology, University of Texas HealthScience Center, 7703 Floyd Curl Drive, San Antonio, TX 78229.E-mail: [email protected]
Received 15 October 2007; Accepted 10 June 2008
Published online in Wiley InterScience(www.interscience.wiley.com.), 9 July 2008.DOI: 10.1002/jcp.21534
Naþ transport in various cells or tissues via epithelial Naþ
channels (ENaC) is an important regulator of fluid homeostasis.In fluid absorptive epithelia, ENaC is located in apicalmembranes and comprises the rate-limiting step for Naþ entryinto the cell (Matalon and O’Brodovich, 1999). ENaC has beenidentified in the epithelia of the lung, distal nephron, colon, andsalivary glands (North, 1996; Garty and Palmer, 1997; Eatonet al., 2004). Although three homologous protein subunits(a, b, and g) form this amiloride-sensitive ion channel, thea-subunit is essential for Naþ transport particularly in the lung(Canessa et al., 1994; Snyder, 2000).
ENaC activity and expression of each subunit is tightlyregulated by a variety of hormones, including b2-adrenergicagonists, vasopressin, mineralocorticoids, and glucocorticoids(Berthiaume et al., 1987; Finley et al., 1998; Dagenais et al., 2001;Itani et al., 2002; Matthay et al., 2002).b2-adrenergic-stimulatedENaC expression and function are partly regulated via thesecond messenger, cAMP (Walters et al., 1990; Berthiaume,1991; Stephens et al., 1998; Niisato et al., 1999; Matthay et al.,2002). Exogenous cAMP stimulates amiloride-sensitive Naþ
transport in part by modulating ENaC subunit gene expression(Dagenais et al., 2001; Chen et al., 2002; Itani et al., 2002;Thomas et al., 2004). cAMP also stimulates Naþ transport byincreasing the translocation of ENaC subunits, in particular thea-subunit, to the apical membrane surface (Snyder, 2000;Morris and Schafer, 2002; Thomas et al., 2004).
Similar to the effects by aldosterone in renal epithelial cells,cAMP-stimulated increase in ENaC activity likely involvesadditional regulatory protein(s) induced early during thecascade of signaling events. One such candidate protein is theserum- and glucocorticoid-inducible kinase 1 (SGK1), a serine/threonine-specific protein kinase originally described as animmediate early response gene in rat mammary epithelial cells(Webster et al., 1993; Alvarez et al., 1999). SGK1 co-expressionwith a-, b-, and g-ENaC subunits in Xenopus oocytes or A6renal epithelial cells increased Naþ transport by increasing theapical membrane expression of ENaC (Alvarez et al., 1999;Chen et al., 1999; Naray-Fejes-Toth et al., 1999; Alvarez and
� 2 0 0 8 W I L E Y - L I S S , I N C .
Canessa, 2003). Inhibition of endogenous SGK1 by antisenseoligonucleotide specific for SGK1 or dominant negativeSGK1 protein decreased steroid-induced stimulation of Naþ
transport (Faletti et al., 2002; Boyd and Naray-Fejes-Toth,2005). Furthermore, in the presence of a SGK1 kinase-deaddominant negative construct, a-ENaC mRNA expression wasdecreased in renal cells (Boyd and Naray-Fejes-Toth, 2005).
Previously, we have reported that rat submandibular gland(SMG-C6) epithelial cells cultured in modified media devoidof hydrocortisone, endogenously express negligible levels ofa-ENaC mRNA and protein and demonstrate minimalamiloride-sensitive transepithelial Naþ transport (Mustafa et al.,2008). In DbcAMP-treated SMG-C6 cells, increased a-ENaCmRNA expression, protein formation, and amiloride-sensitivetransepithelial Naþ transport was measured at 24 h (Mustafaet al., 2008). Since elevating intracellular cAMP concentrationinduces SGK1 protein expression in granulosa cells andincreases activation of recombinant SGK1 expressed in COS7cells (Gonzalez-Robayna et al., 2000; Perrotti et al., 2001), weexamined the effects of DbcAMP on the expression of SGK1and specific signaling pathways that in turn may regulatea-ENaC expression and function. Our present findings indicatethat treatment of SMG-C6 epithelial cells with DbcAMP inducesSGK1 mRNA and protein expression in a time-dependentmanner via the protein kinase A (PKA) and phosphatidyl inositol3-kinase (PI3-K) pathways and inhibition of SGK1 expression by

I N D U C T I O N O F S G K 1 A N D a - E N A C B Y D b c A M P 633
siRNA decreases a-ENaC expression and amiloride-sensitivetransepithelial Naþ transport.
Materials and MethodsMaterials
Dulbecco’s modified Eagle’s medium (DMEM), Ham F-12 media,fetal bovine serum (FBS), and trace element mix were obtainedfrom Gibco (Grand Island, NY). Transferrin and epidermal growthfactor (EGF) were obtained from JRH Biosciences (Lenexa, KS).Insulin was obtained from Novo Nordisk Pharmaceuticals(Princeton, NJ). Polycarbonate Snapwell1 filters were obtainedfrom Costar (Cambridge, MA). Amiloride, actinomycin D, anddibutyryl-cAMP (DbcAMP) were purchased from Sigma ChemicalCo. (St. Louis, MO). KT5720, H-89, and LY294002 were purchasedfrom Calbiochem (San Diego, CA). The anti-rat a-ENaC antibody,an affinity-purified antibody directed against the L20-C42NH2-terminal portion of the rat a-ENaC subunit was obtainedfrom Affinity BioReagents (Golden, CO). The sheep polyclonalanti-SGK1 antibody was purchased from Upstate Biotechnology(Lake Placid, NY). This antibody has since been discontinued. Allother antibodies used in this study were purchased from SantaCruz Biotechnology (Santa Cruz, CA). The pSRaDp85 plasmidencoding for a dominant negative regulatory subunit PI3-K wasgenerously provided by Dr. N. Ghosh-Choudhury, Departmentof Pathology, University of Texas Health Science Center, SanAntonio, TX.
Cell culture
SMG-C6 epithelial cells (Quissell et al., 1997) have been previouslyutilized to characterize ENaC function (Castro et al., 2000). In ourstudy SMG-C6 epithelial cells were cultured in modified media, thatis, in the absence of hydrocortisone. The modified culture mediumused was DMEM/F12 (1:1 mixture) with 2.5% FBS supplementedwith trace element mix (5 ml/500 ml of media), 5mg/ml transferrin,0.1 mM retinoic acid, 2.0 nM triiodothyroxine (T3), 5 mg/ml insulin,80 ng/ml epidermal growth factor (EGF), 100 U/ml penicillin and100 mg/ml streptomycin (Quissell et al., 1997). Cells were seededat a density of 50,000–100,000 cells/cm2 on Snapwell1 tissueculture treated filters (diameter 12 mm, pore size 0.4 mm) coatedwith 1.0 mg/cm2 of human collagen type-I (Becton Dickinson,Franklin Lakes, NJ). For Northern and Western blot analysis cellswere cultured in 100 mm dishes and treatments were performedwhen cells were 90–100% confluent.
Transepithelial transport studies
Transepithelial resistances (Rt) of the SMG-C6 monolayers grownon permeable filter supports were measured daily followingseeding utilizing an EVOM-G, (World Precision Instrument,Sarasota, FL). Filters exhibiting a Rt> 600 V.cm2 were placedin a modified Ussing chamber, (Costar) equipped with Ag/AgClelectrodes and bathed with Kreb’s-Ringer bicarbonate (KRB)media containing (in mM): 120 NaCl, 2.5 K2HPO4, 0.6 KH2PO4,1.2 CaCl2, 1.2 MgCl2, 20 NaHCO3, and 10 glucose at pH 7.4. Themedia were air-lifted with 5% CO2/21% O2 and temperature wasmaintained at 378C. Using a DVC-1000 Voltage/Current Clamp(World Precision Instrument) with automatic fluid resistancecompensation, stable baseline potential difference (PD) wasmeasured. After equilibration (10–15 min), the monolayer’s PDwas clamped to 0 mV and short-circuit current (Isc) wascontinuously recorded. Every 30 sec, the monolayer was clampedto 1 mV for 0.5 sec so that measured changes in Isc enabledcalculation of Rt using Ohm’s law (Rt¼DPD/DIsc). Reported Iscvalues refer to the movement of positive charges from the apical tothe basolateral side of the membrane, and the PD is referenced tothe apical side (negative relative to the basolateral side). Afterreaching steady state Isc values, amiloride, an ENaC inhibitor(10 mM), was instilled in the apical reservoir and the change in
JOURNAL OF CELLULAR PHYSIOLOGY
current was measured as the amiloride-sensitive Isc. All reagentsused in this study were dissolved in either media or DMSO with thefinal dilution in the Ussing chamber fluid being 1:1,000. This amountof DMSO did not alter bioelectric properties and previous studiesestablished the concentrations of amiloride where maximal effectwas observed without altering Rt, indicating that changes intransepithelial Isc was a direct effect of the specific agent studied(Castro et al., 2000). Neither basal bioelectric properties nor theresponse to specific agents varied following each cell passage.
Northern blot analysis
Following specific treatments total RNA was isolated fromcultured SMG-C6 epithelial cells using TRIzol reagent GIBCO(BRL, Gaithersburg, MD) according to the manufacturer’sinstructions. RNA (20mg) was size-fractionated by electrophoresison a 0.8% agarose/2.2 M formaldehyde gel under denaturingconditions and transferred using a PosiBlot (Stratagene, La Jolla,CA) onto a Magna nylon membrane (Microns Separations, Inc.,Westborough, MA). RNA was immobilized by ultraviolet crosslinking. cDNA probes were labeled with a multiprime DNA labelingsystem using [a-32P] dCTP (specific activity, 3,000 Ci/mmol).Northern blot hybridizations were performed in 50% formamide,1 M NaCl, 10% dextran sulfate, 50 mM Tris–HCl pH 7.5, 0.1%sodium pyrophosphate and 0.2% Denhardt’s solution at 428Cfor 16 h. The membranes were washed once in 2� SSC, 0.1%SDS at room temperature for 10 min and then in 2� SSC, 0.1%SDS at 658C for 5 min. Radioactivity was visualized using aPhosphorImager (Molecular Dynamics, Sunnyvale, CA). Controlhybridizations were performed using a [a-32P] end-labeledoligonucleotide complimentary to rat 18S rRNA. Signal intensitywas quantified by densitometry analysis using the ImageQuantsoftware.
Western blot analysis
Following treatment of cultured SMG-C6 epithelial cells with eitherDbcAMP and/or specific inhibitors for different periods of time, themedium was removed and the cells were homogenized in lysisbuffered (Tris–HCl 50 mM, pH 7.4, TBS) containing 5 mM EDTA,5 mM EGTA, leupeptin (1 mM), pepstatin A (1 mM), aprotinin(1 mM), and phenylmethylsulphonyl fluoride (1 mM). Samplesunderwent three rapid freeze-thaw cycles to ensure cells werelysed. Protein concentrations were determined using the BioRadDC protein assay (BioRad, Hercules, CA), with BSA used as thestandard. Fifty to sixty micrograms of total protein was subjectedto SDS–PAGE. The separated proteins were transferredelectrophoretically to PVDF membranes (Millipore, Bedford, MA)using a semi-dry transfer blot system and blocked in TBS containing5% non-fat dried milk powder for 1 h and then incubated withprimary antibody in TBS containing 0.01% Tween-20 and 5% nonfatdried milk powder for 24 h. The blots were then incubated withsecondary antibody conjugated to horseradish peroxidaseappropriately diluted in the same buffer for 1 h. Peroxidaselabeled-proteins were visualized using an enhancedchemiluminescence (ECL) assay kit (Amersham Pharmacia Biotech,Little Chalfont, England). The relative intensities of the bands werequantified by densitometry using the NIH imaging program. Inseveral instances the blots were re-probed for b-actin tocontrol for equal loading of samples.
Transient transfection studies
SMG-C6 epithelial cells were cultured in 100 mm tissue culturedishes or Snapwell1 filters as described above. Cells weretransfected with a dominant negative 85 kDa subunit of PI3-K usingLipofectamine 2000 reagent (Invitrogen, Carlsberg, CA) accordingto the manufacturer’s instructions. Lipofectamine 2000 wascomplexed with 2mg dominant negative PI3-K plasmid in OptiMEMmedium for 1 h and then added to the cells. After 3 h, cells weretreated with DbcAMP.

634 V A S Q U E Z E T A L .
RNA interference
RNA interference studies for gene knock-down were performedusing small interfering double-stranded RNA (SiRNA) molecules.SiRNA specific for rat SGK1 was obtained from Dharmacon(Lafayette, CO). SMG-C6 cells were transfected with 0.1 mM ofsiRNA using Lipofectamine reagent according to themanufacturer’s instructions for 24 h prior to exposure to DbcAMP.The effectiveness of the siRNA for silencing SGK1 gene expressionwas assessed by Northern blot analysis.
Statistical analysis
All data are presented as means� SE. All experiments wereperformed at least five times unless otherwise stated. ANOVAwith Bonferroni post-hoc analysis was used to evaluate significantdifferences between experimental groups. A P-value of <0.05 wasconsidered statistically significant.
ResultsCycloheximide inhibits DbcAMP-induced a-ENaCmRNA expression
SMG-C6 epithelial cells cultured in modified mediaendogenously express minimal amounts ofa-ENaC mRNA andprotein and exhibit little amiloride-sensitive transepithelial Naþ
transport (Mustafa et al., 2008). We have previously reportedthat treatment of SMG-C6 cells with exogenous DbcAMPmarkedly induced a-ENaC mRNA expression by 24 h andremained elevated for up to 48 h (Mustafa et al., 2008). Giventhe fact that a-ENaC mRNA is abundantly expressed at 24 h, itis very likely that intermediary protein(s) are actively involvedin the regulation of a-ENaC mRNA transcription. To this end,SMG-C6 cells were treated with both DbcAMP and a proteinsynthesis inhibitor, cycloheximide for 24 h. As shown inFigure 1, DbcAMP-induced a-ENaC mRNA expression was
Fig. 1. Effect of protein synthesis inhibition on a-ENaC mRNAsynthesis. Confluent cells were treated with DbcAMP andcycloheximide and total RNA was isolated and subjected to Northernblot hybridization with a 32P-labeled a-ENaC cDNA probe. Arepresentative image demonstrating 2 cycloheximide treatmentsalone and 4 with cycloheximide and DbcAMP is shown. Thecorresponding densitometric analysis is shown with mRNAnormalized to 18S. Data is expressed as mean W SE (n U 2–4determinations at each time point).
M
P < 0.05 versus DbcAMP treatedcells.
JOURNAL OF CELLULAR PHYSIOLOGY
significantly decreased in the presence of cycloheximide. Wealso observed that cycloheximide did not alter the negligibleendogenous levels of a-ENaC mRNA. This finding indicatedthat the DbcAMP-dependent increase in a-ENaC mRNAexpression in SMG-C6 epithelial cells initially requires thesynthesis of intermediary protein(s).
Effect of DbcAMP on SGK1 expression in SMG-C6epithelial cells
In renal M1 cells, SGK1 regulates endogenous anddexamethasone-induced transcription of a-ENaC (Boyd andNaray-Fejes-Toth, 2005). To investigate the role of SGK1 in theinduction of a-ENaC by DbcAMP, we first examined the effectof DbcAMP on SGK1 mRNA expression. Minimal constitutiveSGK1 mRNA expression was present in untreated or controlcells (Fig. 2A). However, exposure of SMG-C6 epithelial cellsto 1 mM of DbcAMP elevated SGK1 mRNA expression,peaking between 0.5 and 2 h, and returning to basal levelsby 3 h (Fig. 2A). To test the hypothesis that the increase inDbcAMP-induced SGK1 mRNA occurred as a result of elevatedtranscription of the SGK1 gene, cells were treated with thetranscriptional inhibitor actinomycin D in the presence ofDbcAMP for 1 h. Figure 2B shows that DbcAMP-inducedincrease in SGK1 transcript was inhibited after 1 h of treatment;indicative of a transcriptional-dependent regulation of DbcAMPon SGK1 expression. However, treatment of cells withcycloheximide did not alter the DbcAMP-stimulatedexpression of SGK1 mRNA (Fig. 2C); suggesting that synthesisof intermediary protein(s) is not required for DbcAMP-inducedSGK1 mRNA expression in SMG-C6 epithelial cells. Toevaluate whether DbcAMP-induced increase in SGK1 mRNAwas associated with an increase in SGK1 protein, whole celllysates were subjected to immunoblot analysis using acommercially available anti-SGK1 antibody. Protein levels ofSGK1 were quite low in control cells, but increased after 2 h oftreatment with DbcAMP and remained elevated at 24 h(Fig. 2D). The anti-SGK1 antibody recognized a protein band atapproximately 48–50 kDa as previously reported (Tong et al.,2004).
Inhibition of PKA reduces DbcAMP-induced SGK1mRNA and protein expression
cAMP-induced activation of protein kinase A (PKA) is necessaryto initiate several different cellular functions including geneexpression (Richards, 2001; Schwartz, 2001). Activation of PKAis involved in cAMP-mediated increases in a-ENaC protein inSMG-C6 cells and H441 epithelial cells (Thomas et al., 2004;Mustafa et al., 2008). To evaluate the role of PKA duringinduction of SGK1, SMG-C6 epithelial cells were separatelyexposed to the PKA inhibitors KT5720 (0.5 mg/ml) or H-89(10 mM) simultaneously with 1 mM DbcAMP for 1 h. As shownin Figure 3A, DbcAMP-mediated SGK1 mRNA levels weremarkedly reduced in the presence of either PKA inhibitorcompared to DbcAMP exposed cells alone. Furthermore, wealso observed that H-89 and KT5720 significantly attenuated,but not abolished, DbcAMP-induced SGK1 protein expressionfollowing 4 h of treatment (Fig. 3B).
Activation of the PI3-K pathway is important forDbcAMP-induced SGK1 mRNA and protein expression
In addition to PKA, the phosphatidylinositol-3 kinase (PI3-K)signaling pathway is activated by cAMP (Perrotti et al.,2001; Richards, 2001; Thomas et al., 2004). Stimulation ofPI3-K either directly by cAMP or through small intermediaryproteins activates downstream proteins, specificallyphosphoinositide-dependent kinases (PDK1) (Park et al., 1999).Activation of the PI3-K/PDK1 pathway is thought to play amajor role during cell proliferation and survival (Richards,
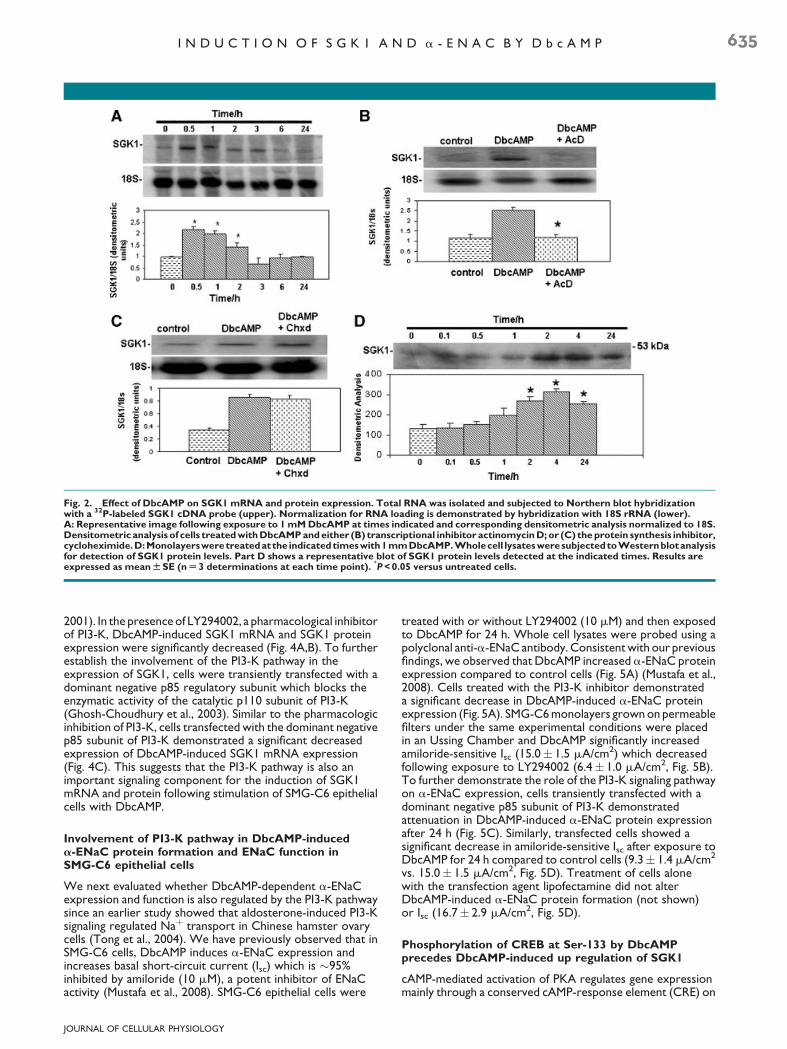
Fig. 2. Effect of DbcAMP on SGK1 mRNA and protein expression. Total RNA was isolated and subjected to Northern blot hybridizationwith a 32P-labeled SGK1 cDNA probe (upper). Normalization for RNA loading is demonstrated by hybridization with 18S rRNA (lower).A: Representative image following exposure to 1 mM DbcAMP at times indicated and corresponding densitometric analysis normalized to 18S.Densitometric analysis of cells treated with DbcAMP and either (B) transcriptional inhibitor actinomycin D; or (C) the protein synthesis inhibitor,cycloheximide.D:Monolayersweretreatedatthe indicatedtimeswith1mmDbcAMP.Wholecell lysatesweresubjectedtoWesternblotanalysisfor detection of SGK1 protein levels. Part D shows a representative blot of SGK1 protein levels detected at the indicated times. Results areexpressed as mean W SE (n U 3 determinations at each time point).
M
P < 0.05 versus untreated cells.
I N D U C T I O N O F S G K 1 A N D a - E N A C B Y D b c A M P 635
2001). In the presence of LY294002, a pharmacological inhibitorof PI3-K, DbcAMP-induced SGK1 mRNA and SGK1 proteinexpression were significantly decreased (Fig. 4A,B). To furtherestablish the involvement of the PI3-K pathway in theexpression of SGK1, cells were transiently transfected with adominant negative p85 regulatory subunit which blocks theenzymatic activity of the catalytic p110 subunit of PI3-K(Ghosh-Choudhury et al., 2003). Similar to the pharmacologicinhibition of PI3-K, cells transfected with the dominant negativep85 subunit of PI3-K demonstrated a significant decreasedexpression of DbcAMP-induced SGK1 mRNA expression(Fig. 4C). This suggests that the PI3-K pathway is also animportant signaling component for the induction of SGK1mRNA and protein following stimulation of SMG-C6 epithelialcells with DbcAMP.
Involvement of PI3-K pathway in DbcAMP-induceda-ENaC protein formation and ENaC function inSMG-C6 epithelial cells
We next evaluated whether DbcAMP-dependent a-ENaCexpression and function is also regulated by the PI3-K pathwaysince an earlier study showed that aldosterone-induced PI3-Ksignaling regulated Naþ transport in Chinese hamster ovarycells (Tong et al., 2004). We have previously observed that inSMG-C6 cells, DbcAMP induces a-ENaC expression andincreases basal short-circuit current (Isc) which is �95%inhibited by amiloride (10 mM), a potent inhibitor of ENaCactivity (Mustafa et al., 2008). SMG-C6 epithelial cells were
JOURNAL OF CELLULAR PHYSIOLOGY
treated with or without LY294002 (10 mM) and then exposedto DbcAMP for 24 h. Whole cell lysates were probed using apolyclonal anti-a-ENaC antibody. Consistent with our previousfindings, we observed that DbcAMP increased a-ENaC proteinexpression compared to control cells (Fig. 5A) (Mustafa et al.,2008). Cells treated with the PI3-K inhibitor demonstrateda significant decrease in DbcAMP-induced a-ENaC proteinexpression (Fig. 5A). SMG-C6 monolayers grown on permeablefilters under the same experimental conditions were placedin an Ussing Chamber and DbcAMP significantly increasedamiloride-sensitive Isc (15.0� 1.5 mA/cm2) which decreasedfollowing exposure to LY294002 (6.4� 1.0 mA/cm2, Fig. 5B).To further demonstrate the role of the PI3-K signaling pathwayon a-ENaC expression, cells transiently transfected with adominant negative p85 subunit of PI3-K demonstratedattenuation in DbcAMP-induced a-ENaC protein expressionafter 24 h (Fig. 5C). Similarly, transfected cells showed asignificant decrease in amiloride-sensitive Isc after exposure toDbcAMP for 24 h compared to control cells (9.3� 1.4 mA/cm2
vs. 15.0� 1.5 mA/cm2, Fig. 5D). Treatment of cells alonewith the transfection agent lipofectamine did not alterDbcAMP-induced a-ENaC protein formation (not shown)or Isc (16.7� 2.9 mA/cm2, Fig. 5D).
Phosphorylation of CREB at Ser-133 by DbcAMPprecedes DbcAMP-induced up regulation of SGK1
cAMP-mediated activation of PKA regulates gene expressionmainly through a conserved cAMP-response element (CRE) on

Fig. 3. Effect of PKA inhibitors on DbcAMP-mediated SGK1 mRNAandproteinexpression.CulturedSMG-C6epithelialcellsweretreatedwith either KT5720 (0.5 mg/ml) or H-89 (10 mM) simultaneously with1 mM DbcAMP. A: Total RNA was isolated after 1 h and subjected toNorthern blot hybridization and normalized to 18S. B: Whole celllysates were isolated after 4 h and then subjected to Western blotanalysis using the anti-SGK1 antibody. A representative blot withdensitometric analysis is shown. n U 3 for each treatment.
M
P < 0.05versus DbcAMP-treated cells.
636 V A S Q U E Z E T A L .
the promoter sequence of target genes (Kwok et al., 1994;Habener et al., 1995; Garty and Palmer, 1997; Hansen et al.,1999). This regulatory effect is mediated via a 43 kDa CREbinding protein (CREB) which functions as a transcriptionfactor (Kwok et al., 1994; Garty and Palmer, 1997; Hansen et al.,1999). Phosphorylation of a serine residue at 133 (Ser-133) isessential for the transcriptional activation of CREB (Kwok et al.,1994; Garty and Palmer, 1997). We have previously shown byWestern blot analysis that in SMG-C6 epithelial cells, DbcAMPinduced phosphorylation of CREB peaks 10 minutes followingstimulation and is markedly decreased by 60 min and PKAinhibition significantly reduced DbcAMP-inducedphosphorylation of CREB protien (Mustafa et al., 2008). Theantibody used to assess the extent of CREB phosphorylationrecognized moieties phosphorylated at Ser-133. Thephosphorylation of CREB may also involve the PI3-K/PDK1pathway (Richards, 2001). To further investigate whether PI3-Ksignaling stimulated by DbcAMP also activates CREB, SMG-C6cells were pretreated with the PI3-K inhibitor LY294002 for 1 hand then exposed to DbcAMP for 10 min. As previouslyreported, DbcAMP increased the expression ofphosphorylated CREB (P-CREB) after 10 min compared tountreated cells, but the level of P-CREB was significantlyreduced in the cells pretreated with the PI3-K inhibitor(Fig. 6A). Cells transfected with dominant negative p85subunit of PI3-K also demonstrated a markedly decreasedDbcAMP-induced phosphorylation of CREB compared toDbcAMP treated cells (Fig. 6B). The addition of lipofectamine(transfection media) had no significant effect on the levels ofP-CREB compared to control cells. Our observations indicate
JOURNAL OF CELLULAR PHYSIOLOGY
that CREB is also phosphorylated by the PI3-K pathway and maybe a contributing component in the signaling pathway evoked byDbcAMP resulting in the up regulation of SGK1.
Effect of PKA and PI3-K pathways on DbcAMP-induceda-ENaC function
Since we previously demonstrated inhibition of PKA markedlyattenuated a-ENaC function (Mustafa et al., 2008), we nextevaluated whether concomitant inhibition of both PKA andPI3-K pathways would block ENaC function. SMG-C6 cellswere grown on permeable filters, transfected with a dominantnegative p85 subunit of PI3-K and then exposed to the PKAinhibitors, H89 or KT5720, followed by the addition ofDbcAMP for 24 h. Monolayers transfected with dominantnegative p85 subunit of PI3-K showed a decrease inamiloride-sensitive Isc when compared to those treated withDbcAMP alone. The presence of either of the PKA inhibitors inaddition to the dominant negative p85 subunit of PI3-Ksynergistically decreased amiloride-sensitive Isc, approximatinguntreated (control) levels (Fig. 7).
Regulation of DbcAMP-induced a-ENaCexpression by SGK1
Our findings indicate that DbcAMP induces both SGK1 anda-ENaC expression via the PKA and PI3-K pathways inSMG-C6 cells (Mustafa et al., 2008). However, SGK1 mRNA issignificantly expressed much earlier than that of a-ENaCmRNA (0.5–2 h vs. 24 h), following stimulation with DbcAMP.Furthermore, inhibition of either the PI3-K or PKA pathwaysdecreases both SGK1 and a-ENaC expression, but, it is notclear ifa-ENaC induction by DbcAMP is dependent on SGK1 orif there is a sharing of common signaling pathways. Therefore,we examined the effects of siRNA inhibition on SGK1expression in SMG-C6 (Meng et al., 2005). In SMG-C6 cellstreated with scrambled siRNA, we did not observe asignificant increase in the endogenous amounts of SGK1mRNA nor did we detect changes in the abundance ofDbcAMP-induced SGK1 mRNA expression (Fig. 8A). Theadequacy of silencing SGK1 mRNA expression is depictedby Figure 8B. Cells transfected with siRNA and then treatedwith DbcAMP for 1 h demonstrated SGK1 mRNA levels similarto control cells. In subsequent experiments, cells transfectedwith SGK1 siRNA and then treated with DbcAMP for 24 hshowed decreased expression of a-ENaC mRNA andprotein (Fig. 9A,B), as well as significantly decreasedamiloride-sensitive Isc (Fig. 9C). These findings demonstratethat SGK1 is indeed an important requirement for induction ofa-ENaC expression by DbcAMP in SMG-C6 cells.
Reduction of existing NaR transport by inhibiting thePI3-K pathway
Phosphorylation of endogenous SGK1 by the PI3-K pathwayincreases Naþ transport by enhancing ENaC expression at theapical membrane through phosphorylation of the ubiquitinligase Nedd4-2, thus inhibiting internalization and degradationof ENaC (Itani et al., 2002; Kamynina and Staub, 2002; Thomasand Itani, 2004). We have shown that DbcAMP initiates SGK1expression prior to the observed induction of a-ENaCexpression, thus implying that SGK1 may be a prerequisite fora-ENaC expression and consequent function in SMG-C6epithelial cells. As DbcAMP-induced SGK1 protein levels arestill apparent after 24 h of treatment, we investigated whetherblocking SGK1 activity (i.e., inhibiting phosphorylation) at thistime would alter DbcAMP-induced Naþ transport. In theseexperiments, cells were grown on permeable filters and treatedwith DbcAMP for 24 h. Filters were mounted on the Ussingchamber and LY294002 was then added to the apical andbasolateral chambers. Following the addition of LY294002,

Fig. 4. Effect of PI3-K inhibition DbcAMP-induced SGK1 mRNA and protein expression. SMG-C6 epithelial cells were treated with the PI3Kinhibitor LY294002 (10 mM) together with 1 mM DbcAMP for 1 h (mRNA analysis) and 4 h (protein analysis). A: Typical Northern blot for SGK1mRNA expression. B: Representative Western blot analysis for SGK1 is shown. C: Monolayers were transfected with a dominant negative PI3-KconstructasdescribedinthemethodssectionofthispaperandsubsequentlytreatedwithDbcAMPfor1h.TotalRNAwasisolatedandsubjectedtoNorthern blot hybridization with a 32P-labeled SGK1 cDNA probe (upper). Normalization for RNA loading is demonstrated by hybridization with18S rRNA (lower). Depicted is a representative Northern blot image and densitometric analysis with normalization to 18S.
M
P < 0.05 versusDbcAMP-treated cells.
I N D U C T I O N O F S G K 1 A N D a - E N A C B Y D b c A M P 637
basal Isc initially decreased by 32% after 1 h (data not shown)with a subsequent decrease of 54.7% compared to baselinecurrent after 2 h (Fig. 10). These results indicate that in SMG-C6cells with very minimal endogenous expression of SGK1 anda-ENaC, DbcAMP induces SGK1 expression which is not onlypartly required for subsequent induction of a-ENaCexpression, but also regulates ENaC function.
Discussion
Published reports indicate that SGK1 regulates ENaC atboth the level of subunit expression and activity at the apicalmembrane. In renal cortical collecting duct epithelial cellstransfected with a kinase dead dominant negative SGK1,transcription of the a- and b-subunits of ENaC weresignificantly lower than that expressed in cells transfectedwith full-length SGK1 protein (Helms et al., 2003; Boyd andNaray-Fejes-Toth, 2005). Co-expression of SGK1 with theENaC subunits in Xenopus oocytes and A6 cells increasedENaC activity (Alvarez et al., 1999; Chen et al., 1999;Naray-Fejes-Toth et al., 1999).
cAMP activates transcription and phosphorylation of SGK1via the PKA and PI3-K pathways (Gonzalez-Robayna et al., 2000;Perrotti et al., 2001). cAMP also induces SGK1 protein
JOURNAL OF CELLULAR PHYSIOLOGY
expression in granulosa cells and increases activation ofrecombinant SGK1 expressed in COS7 cells (Gonzalez-Robaynaet al., 2000; Perrotti et al., 2001). In H441 cells, both theimmediate and sustained response of cAMP on Naþ transportwas associated with increased phosphorylation of SGK1(Thomas et al., 2004).
In this study, epithelial cells derived from the ratsubmandibular gland (SMG-C6) were cultured in modifiedmedia (Castro et al., 2000). As there is minimal endogenousa-ENaC expression and Naþ transport, this cell line serves asan excellent model for investigating the mechanistic/signalingpathways by which agonists increase epithelial Naþ transport.Previously, we have shown that treatment of SMG-C6 epithelialcells with exogenous DbcAMP increased a-ENaC mRNAexpression, protein formation, and Naþ transport following24 h treatment via a PKA-sensitive pathway (Mustafa et al.,2008). The presence of SGK1 is not only critical for enhancingENaC activity, but may also be important for a-ENaCexpression. Therefore, we sought to evaluate the potentialeffect of DbcAMP on the expression of SGK1 in these cells andattempted to identify whether DbcAMP-induced a-ENaCexpression and function is dependent upon the presence ofSGK1.
Exposure of SMG-C6 cells to DbcAMP induces rapidexpression of SGK1 mRNA within 30 min and protein by 2 h.

Fig. 5. Effect of inhibiting the PI3-K pathway on a-ENaC protein formation and function. SMG-C6 cells were exposed to LY294002 (10 mM) and1mMDbcAMPfor24h.A:Wholecell lysatesweresubjectedtoWesternblotanalysisusinganti-a-ENaCantibody.B:Short-circuitcurrent(Isc)wasmeasured at baseline and after the addition of amiloride (10mM) to the apical bath across monolayers grown on filters placed in Ussing chambers.Amiloride-sensitive Isc are depicted in the graph. C: SMG-C6 cells were transfectedwith PI3-K dominantnegative cDNA construct and treated for24 h with DbcAMP. Samples were then subjected to Western blot analysis. D: Amiloride-sensitive Isc in monolayers transfected with PI3-Kdominant negative construct and treated with DbcAMP.
M
P < 0.05 versus DbcAMP-treated cells.
638 V A S Q U E Z E T A L .
The induction of the SGK1 transcript is ablated by actinomycinD but not by cycloheximide. The time-course pattern of SGK1mRNA expression induced by DbcAMP and the effects ofcycloheximide are similar to dexamethasone effects on A6
Fig. 6. EffectofPI3-KInhibitorsonDbcAMP-inducedCREBPhosphorylatusingphosphor-specificCREBantibody.A:Cellsweretreatedwith1mMDbexposed to DbcAMP for 10 min. B: Cells transfected with a mutated nonf10 min. Densitometric analysis is expressed as mean values of 3–4 separa
JOURNAL OF CELLULAR PHYSIOLOGY
kidney cells (Pearce et al., 2000). In addition, we observedsignificant amounts of SGK1 protein evident at 24 h followingstimulation with DbcAMP (Fig. 2D). Interestingly, cAMPtreatment did not induce expression of SGK1 mRNA in H441
ion.PhosphorylationofCREBwasdeterminedbyWesternblotanalysiscAMPfor10minorpretreated for1hwithLY294002(10mM)andthen
unctional cDNA construct of PI3-K and then exposed to DbcAMP forte experiments.
M
P < 0.05 versus DbcAMP-treated cells.
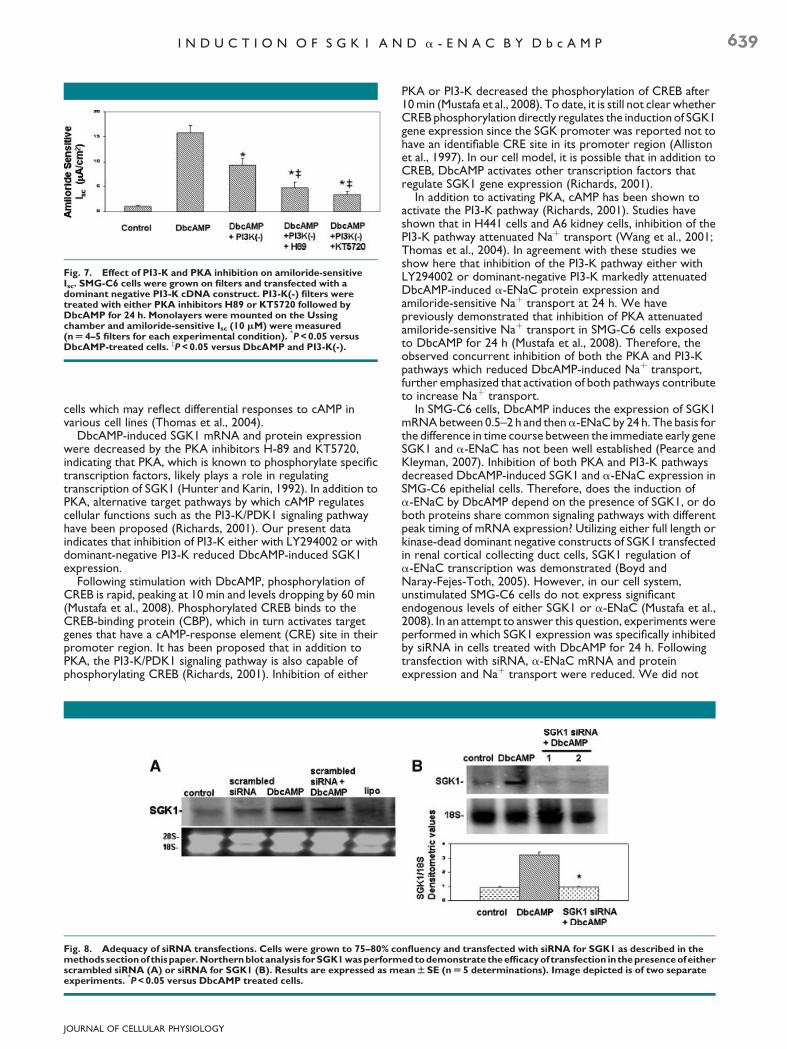
Fig. 7. Effect of PI3-K and PKA inhibition on amiloride-sensitiveIsc. SMG-C6 cells were grown on filters and transfected with adominant negative PI3-K cDNA construct. PI3-K(-) filters weretreated with either PKA inhibitors H89 or KT5720 followed byDbcAMP for 24 h. Monolayers were mounted on the Ussingchamber and amiloride-sensitive Isc (10 mM) were measured(n U 4–5 filters for each experimental condition).
M
P < 0.05 versusDbcAMP-treated cells. zP < 0.05 versus DbcAMP and PI3-K(-).
I N D U C T I O N O F S G K 1 A N D a - E N A C B Y D b c A M P 639
cells which may reflect differential responses to cAMP invarious cell lines (Thomas et al., 2004).
DbcAMP-induced SGK1 mRNA and protein expressionwere decreased by the PKA inhibitors H-89 and KT5720,indicating that PKA, which is known to phosphorylate specifictranscription factors, likely plays a role in regulatingtranscription of SGK1 (Hunter and Karin, 1992). In addition toPKA, alternative target pathways by which cAMP regulatescellular functions such as the PI3-K/PDK1 signaling pathwayhave been proposed (Richards, 2001). Our present dataindicates that inhibition of PI3-K either with LY294002 or withdominant-negative PI3-K reduced DbcAMP-induced SGK1expression.
Following stimulation with DbcAMP, phosphorylation ofCREB is rapid, peaking at 10 min and levels dropping by 60 min(Mustafa et al., 2008). Phosphorylated CREB binds to theCREB-binding protein (CBP), which in turn activates targetgenes that have a cAMP-response element (CRE) site in theirpromoter region. It has been proposed that in addition toPKA, the PI3-K/PDK1 signaling pathway is also capable ofphosphorylating CREB (Richards, 2001). Inhibition of either
Fig. 8. Adequacy of siRNA transfections. Cells were grown to 75–80% comethods sectionof thispaper. Northern blot analysis forSGK1 wasperformscrambled siRNA (A) or siRNA for SGK1 (B). Results are expressed as mexperiments.
M
P < 0.05 versus DbcAMP treated cells.
JOURNAL OF CELLULAR PHYSIOLOGY
PKA or PI3-K decreased the phosphorylation of CREB after10 min (Mustafa et al., 2008). To date, it is still not clear whetherCREB phosphorylation directly regulates the induction of SGK1gene expression since the SGK promoter was reported not tohave an identifiable CRE site in its promoter region (Allistonet al., 1997). In our cell model, it is possible that in addition toCREB, DbcAMP activates other transcription factors thatregulate SGK1 gene expression (Richards, 2001).
In addition to activating PKA, cAMP has been shown toactivate the PI3-K pathway (Richards, 2001). Studies haveshown that in H441 cells and A6 kidney cells, inhibition of thePI3-K pathway attenuated Naþ transport (Wang et al., 2001;Thomas et al., 2004). In agreement with these studies weshow here that inhibition of the PI3-K pathway either withLY294002 or dominant-negative PI3-K markedly attenuatedDbcAMP-induced a-ENaC protein expression andamiloride-sensitive Naþ transport at 24 h. We havepreviously demonstrated that inhibition of PKA attenuatedamiloride-sensitive Naþ transport in SMG-C6 cells exposedto DbcAMP for 24 h (Mustafa et al., 2008). Therefore, theobserved concurrent inhibition of both the PKA and PI3-Kpathways which reduced DbcAMP-induced Naþ transport,further emphasized that activation of both pathways contributeto increase Naþ transport.
In SMG-C6 cells, DbcAMP induces the expression of SGK1mRNA between 0.5–2 h and thena-ENaC by 24 h. The basis forthe difference in time course between the immediate early geneSGK1 and a-ENaC has not been well established (Pearce andKleyman, 2007). Inhibition of both PKA and PI3-K pathwaysdecreased DbcAMP-induced SGK1 and a-ENaC expression inSMG-C6 epithelial cells. Therefore, does the induction ofa-ENaC by DbcAMP depend on the presence of SGK1, or doboth proteins share common signaling pathways with differentpeak timing of mRNA expression? Utilizing either full length orkinase-dead dominant negative constructs of SGK1 transfectedin renal cortical collecting duct cells, SGK1 regulation ofa-ENaC transcription was demonstrated (Boyd andNaray-Fejes-Toth, 2005). However, in our cell system,unstimulated SMG-C6 cells do not express significantendogenous levels of either SGK1 or a-ENaC (Mustafa et al.,2008). In an attempt to answer this question, experiments wereperformed in which SGK1 expression was specifically inhibitedby siRNA in cells treated with DbcAMP for 24 h. Followingtransfection with siRNA, a-ENaC mRNA and proteinexpression and Naþ transport were reduced. We did not
nfluency and transfected with siRNA for SGK1 as described in theedtodemonstrate theefficacy of transfection inthe presence ofeither
ean W SE (n U 5 determinations). Image depicted is of two separate
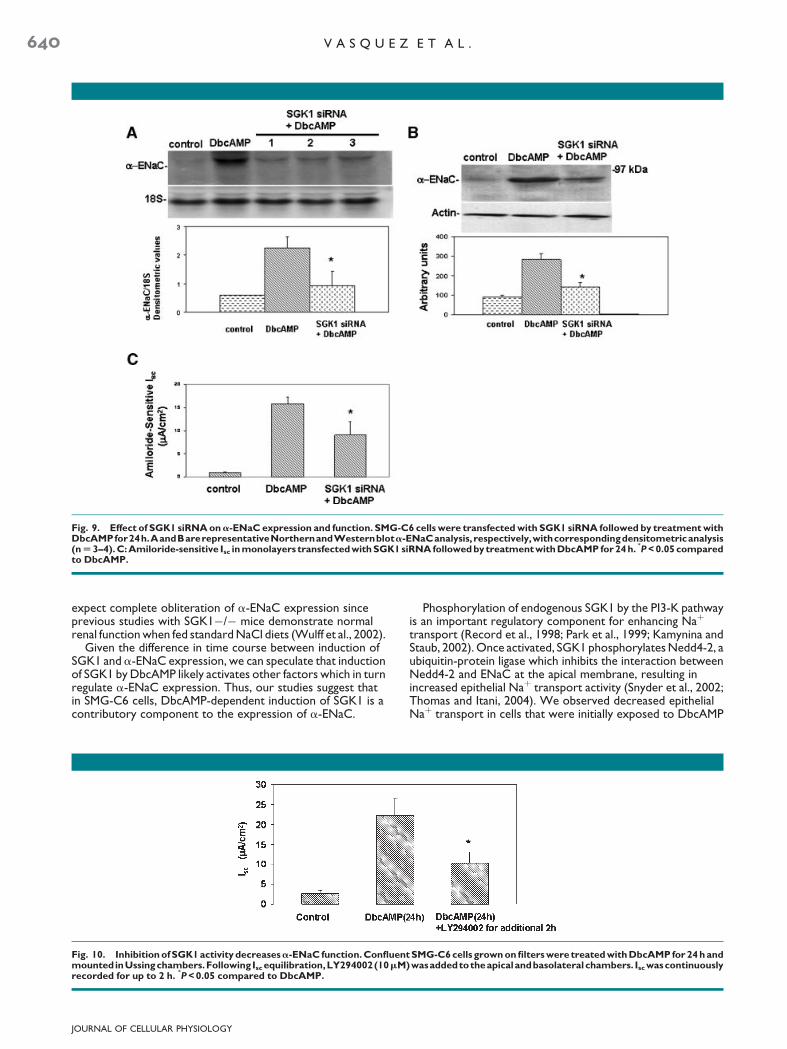
Fig. 9. Effect of SGK1 siRNA ona-ENaC expression and function. SMG-C6 cells were transfected with SGK1 siRNA followed by treatment withDbcAMPfor24h.AandBarerepresentativeNorthernandWesternblota-ENaCanalysis, respectively,withcorrespondingdensitometricanalysis(n U 3–4). C: Amiloride-sensitive Isc in monolayers transfectedwith SGK1 siRNA followed by treatment with DbcAMP for 24h.
M
P < 0.05 comparedto DbcAMP.
640 V A S Q U E Z E T A L .
expect complete obliteration of a-ENaC expression sinceprevious studies with SGK1�/� mice demonstrate normalrenal function when fed standard NaCl diets (Wulff et al., 2002).
Given the difference in time course between induction ofSGK1 and a-ENaC expression, we can speculate that inductionof SGK1 by DbcAMP likely activates other factors which in turnregulate a-ENaC expression. Thus, our studies suggest thatin SMG-C6 cells, DbcAMP-dependent induction of SGK1 is acontributory component to the expression of a-ENaC.
Fig. 10. Inhibition of SGK1 activity decreasesa-ENaC function. Confluenmounted inUssingchambers.Following Isc equilibration,LY294002(10mMrecorded for up to 2 h.
M
P < 0.05 compared to DbcAMP.
JOURNAL OF CELLULAR PHYSIOLOGY
Phosphorylation of endogenous SGK1 by the PI3-K pathwayis an important regulatory component for enhancing Naþ
transport (Record et al., 1998; Park et al., 1999; Kamynina andStaub, 2002). Once activated, SGK1 phosphorylates Nedd4-2, aubiquitin-protein ligase which inhibits the interaction betweenNedd4-2 and ENaC at the apical membrane, resulting inincreased epithelial Naþ transport activity (Snyder et al., 2002;Thomas and Itani, 2004). We observed decreased epithelialNaþ transport in cells that were initially exposed to DbcAMP
t SMG-C6 cells grown on filters were treated with DbcAMP for 24 h and)wasaddedtotheapicalandbasolateralchambers. Isc wascontinuously

Fig. 11. Working model of cAMP-induced SGK1 anda-ENaC expression and function. Left Part: Exposure of SMG-C6 cells to DbcAMP inducesexpression of SGK1 mRNA within 0.5–1 h and SGK1 protein by 2–24 h via the stimulation of the PKA and PI3-K pathways and involvesphosphorylation of the CREB transcription factor. We speculate that there is a further interaction of phosphorylated CREB with othertranscription factors (TFs) to induceSGK1mRNAexpression. The latercomponent of theseevents initiallyevokedbyDbcAMPcontributes to theinduction ofa-ENaC expression and consequent increased NaR transport. Right Part: Phosphorylation of existing SGK1 via the PI3-K pathway isrequired for maintaining NaR transport.
I N D U C T I O N O F S G K 1 A N D a - E N A C B Y D b c A M P 641
for 24 h and then treated for an additional 2 h with LY294002.Therefore, this reduction in ENaC function may be due todecreased phosphorylation of SGK1 since DbcAMP-inducedSGK1 protein is still present at 24 h (Fig. 2D).
The proposed mechanism by which DbcAMP inducesa-ENaC expression and activity is summarized in Figure 11. It isinteresting to note that in SMG-C6 cells, DbcAMP-inducedactivation of the PI3-K pathway is not only involved in theup-regulation of both SGK1 mRNA and protein and a-ENaCprotein expression, but also plays a regulatory functional roleduring Naþ transport. Further characterization of the upstreampathways that are activated following stimulation with DbcAMPwill provide additional evidence for the regulatory effects ofSGK1 on a-ENaC expression.
Literature Cited
Alliston TN, Maiyar AC, Buse P, Firestone GL, Richards JS. 1997. Follicle stimulatinghormone-regulated expression of serum/glucocorticoid-inducible kinase in rat ovariangranulosa cells: A functional role for the Sp1 family in promoter activity. Mol Endocrinol11:1934–1949.
Alvarez de la Rosa D, Canessa CM. 2003. Role of SGK in hormonal regulation of epithelialsodium channel in A6 cells. Am J Physiol 284:C404–C414.
Alvarez de la Rosa D, Zhang P, Naray-Fejes-Toth A, Fejes-Toth G, Canessa CM. 1999. Theserum and glucocorticoid kinase sgk increases the abundance of epithelial sodium channelsin the plasma membrane of Xenopus oocytes. J Biol Chem 274:37834–37839.
Berthiaume Y. 1991. Effect of exogenous cAMP and aminophylline on alveolar and lung liquidclearance in anesthetized sheep. J Appl Physiol 70:2490–2497.
Berthiaume Y, Staub NC, Matthay MA. 1987. Beta-adrenergic agonists increase lung liquidclearance in anesthetized sheep. J Clin Invest 79:335–343.
Boyd C, Naray-Fejes-Toth A. 2005. Gene regulation of ENaC subunits by serum- andglucocorticoid-inducible kinase-1. Am J Physiol 288:F505–F512.
Canessa CM, Schild L, Buell G, Thorens B, Gautschi I, Horisberger JD, Rossier BC. 1994.Amiloride-sensitive epithelial Naþ channel is made of three homologous subunits. Nature367:463–467.
Castro R, Barlow-Walden L, Woodson T, Kerecman JD, Zhang GH, Martinez JR. 2000. Iontransport in an immortalized rat submandibular cell line SMG-C6. Proc Soc Exp Biol Med225:39–48.
Chen SY, Bhargava A, Mastroberardino L, Meijer OC, Wang J, Buse P, Firestone GL, Verrey F,Pearce D. 1999. Epithelial sodium channel regulated by aldosterone-induced protein sgk.Proc Natl Acad Sci 96:2514–2519.
Chen XJ, Eaton DC, Jain L. 2002. Beta-adrenergic regulation of amiloride-sensitive lungsodium channels. Am J Physiol 282:L609–L620.
Dagenais A, Denis C, Vives MF, Girouard S, Masse C, Nguyen T, Yamagata T, Grygorczyk C,KotharyR, Berthiaume Y. 2001. Modulationof alpha-ENaC and alpha1-Naþ-Kþ-ATPase bycAMP and dexamethasone in alveolar epithelial cells. Am J Physiol 281:L217–L230.
JOURNAL OF CELLULAR PHYSIOLOGY
Eaton DC, Chen J, Ramosevac S, Matalon S, Jain L. 2004. Regulation of Naþ channels in lungalveolar type II epithelial cells. Proc Am Thorac Soc 1:10–16.
Faletti CJ, Perrotti N, Taylor SI, Blazer-Yost BL. 2002. sgk: An essential convergence point forpeptide and steroid hormone regulation of ENaC-mediated Naþ transport. Am J Physiol282:C494–C500.
Finley N, Norlin A, Baines DL, Folkesson HG. 1998. Alveolar epithelial fluid clearance ismediated by endogenous catecholamines at birth in guinea pigs. J Clin Invest 101:972–981.
Garty H, Palmer LG. 1997. Epithelial sodium channels: Function, structure, and regulation.Physiol Rev 77:359–396.
Ghosh-Choudhury N, Abboud SL, Mahimainathan L, Chandrasekar B, Choudhury GG. 2003.Phosphatidylinositol 3-kinase regulates bone morphogenetic protein-2 (BMP-2)-inducedmyocyte enhancer factor 2A-dependent transcription of BMP-2 gene in cardiomyocyteprecursor cells. J Biol Chem 278:21998–22005.
Gonzalez-Robayna IJ, Falender AE, Ochsner S, Firestone GL, Richards JS. 2000. Follicle-stimulating hormone (FSH) stimulates phosphorylation and activation of protein kinase B(PKB/Akt) and serum and glucocorticoid-lnduced kinase (Sgk): Evidence for a kinase-independent signaling by FSH in granulosa cells. Mol Endocrinol 14:1283–1300.
Habener JF, Miller CP, Vallejo M. 1995. cAMP-dependent regulation of gene transcription bycAMP response element-binding protein and cAMP response element modulator. VitamHorm 51:1–57.
Hansen TV, Rehfeld JF, Nielsen FC. 1999. Mitogen-activatedprotein kinase and protein kinaseA signaling pathways stimulate cholecystokinin transcription via activation of cyclicadenosine 3’,5’-monophosphate response element-binding protein. Mol Endocrinol13:466–475.
Helms MN, Fejes-Toth G, Naray-Fejes-Toth A. 2003. Hormone-regulatedtransepithelial Naþ transport in mammalian CCD cells requires SGK1 expression. Am JPhysiol 284:F480–F487.
Hunter T, Karin M. 1992. The regulation of transcription by phosphorylation. Cell 70:375–387.
Itani OA, Auerbach SD, Husted RF, Volk KA, Ageloff S, Knepper MA, Stokes JB, Thomas CP.2002. Glucocorticoid-stimulated lung epithelial Naþ transport is associated with regulatedENaC and sgk1 expression. Am J Physiol 282:L631–L641.
Kamynina E, Staub O. 2002. Concerted action of ENaC, Nedd4-2, and Sgk1 intransepithelial Naþ transport. Am J Physiol 283:F377–F387.
Kwok RP, Lundblad JR, Chrivia JC, Richards JP, Bachinger HP, Brennan RG, Roberts SG,Green MR, Goodman RH. 1994. Nuclear protein CBP is a coactivator for the transcriptionfactor CREB. Nature 370:223–226.
Matalon S, O’Brodovich H. 1999. Sodium channels in alveolar epithelial cells: Molecularcharacterization, biophysical properties, and physiological significance. Annu Rev Physiol61:627–661.
Matthay MA, Clerici C, Saumon G. 2002. Invited review: Active fluid clearance from the distalair spaces of the lung. J Appl Physiol 93:1533–1541.
Meng F, Yamagiwa Y, Taffetani S, Han J, Patel T. 2005. IL-6 activates serum and glucocorticoidkinase via p38a mitogen-activated protein kinase pathway. Am J Physiol 289:C971–C981.
Morris RG, Schafer JA. 2002. cAMP increases density of ENaC subunits in the apicalmembrane of MDCK cells in direct proportion to amiloride-sensitive Naþ transport. J GenPhysiol 120:71–85.
Mustafa SB, Castro R, Falck AJ, Petershack JA, Henson BM, Mendoza YM, Choudary A,Seidner SR. 2008. Protein kinase A and mitogen-activated protein kinase pathways mediatecAMP induction of a-epithelial Naþ Channels (a-ENaC). J Cell Physiol 215:101–110.
Naray-Fejes-Toth A, Canessa C, Cleaveland ES, Aldrich G, Fejes-Toth G. 1999. sgk is analdosterone-induced kinase in the renal collecting duct. Effects on epithelial Naþ channels.J Biol Chem 274:16973–16978.

642 V A S Q U E Z E T A L .
Niisato N, Ito Y, Marunaka Y. 1999. cAMP stimulates Naþ transport in rat fetal pneumocyte:Involvement of a PTK- but not a PKA-dependent pathway. Am J Physiol 277:L727–L736.
North RA. 1996. Families of ion channels with two hydrophobic segments. Curr Opin CellBiol 8:474–483.
Park J, Leong ML, Buse P, Maiyar AC, Firestone GL, Hemmings BA. 1999. Serum andglucocorticoid-inducible kinase (SGK) is a target of the PI 3-kinase-stimulated signalingpathway. EMBO J 18:3024–3033.
Pearce D, Kleyman TR. 2007. Salt, sodium channels, and SGK1. J Clin Invest 117:592–595.Pearce D, Verrey F, Chen SY, Mastroberardino L, Meijer OC, Wang J, Bhargava A. 2000. Role
of SGK in mineralocorticoid-regulated sodium transport. Kidney Int 57:1283–1289.Perrotti N, He RA, Phillips SA, Haft CR, Taylor SI. 2001. Activation of serum- and
glucocorticoid-induced protein kinase (Sgk) by cyclic AMP and insulin. J Biol Chem276:9406–9412.
Quissell DO, Barzen KA, Gruenert DC, Redman RS, Camden JM, Turner JT. 1997.Development and characterization of SV40 immortalized rat submandibular acinar celllines. In Vitro Cell Dev Biol 33:164–173.
Record RD, Froelich LL, Vlahos CJ, Blazer-Yost BL. 1998. Phosphatidylinositol 3-kinaseactivation is required for insulin-stimulated sodium transport in A6 cells. Am J Physiol274:E611–E617.
Richards JS. 2001. New signaling pathways for hormones and cyclic adenosine3’,5’-monophosphate action in endocrine cells. Mol Endocrinol 15:209–218.
Schwartz JH. 2001. The many dimensions of cAMP signaling. Proc Natl Acad Sci 98:13482–13484.
Snyder PM. 2000. Liddle’s syndrome mutations disrupt cAMP-mediated translocation of theepithelial Naþ channel to the cell surface. J Clin Invest 105:45–53.
JOURNAL OF CELLULAR PHYSIOLOGY
Snyder PM, Olson DR, Thomas BC. 2002. Serum and glucocorticoid-regulated kinasemodulates Nedd4-2-mediated inhibition of the epithelial Naþ channel. J Biol Chem277:5–8.
Stephens RH, Benjamin AR, Walters DV. 1998. The regulation of lung liquid absorption byendogenous cAMP in postnatal sheep lungs perfused in situ. J Physiol 511:587–597.
Thomas CP, Itani OA. 2004. New insights into epithelial sodium channel function in thekidney: Site of action, regulation by ubiquitin ligases, serum- and glucocorticoid-induciblekinase and proteolysis. Curr Opin Nephrol Hypertens 13:541–548.
Thomas CP, Campbell JR, Wright PJ, Husted RF. 2004. cAMP-stimulated Naþ transport inH441 distal lung epithelial cells: Role of PKA, phosphatidylinositol 3-kinase, and sgk1. Am JPhysiol 287:L843–L851.
Tong Q, Booth RE, Worrell RT, Stockand JD. 2004. Regulation of Naþ transport byaldosterone: Signaling convergence and cross talk between the PI3-K and MAPK1/2cascades. Am J Physiol 286:F1232–F1238.
Walters DV, Ramsden CA, Olver RE. 1990. Dibutyryl cAMP induces a gestation-dependentabsorption of fetal lung liquid. J Appl Physiol 68:2054–2059.
Wang J, Barbry P, Maiyar AC, Rozansky DJ, Bhargava A, Leong M, Firestone GL, Pearce D.2001. SGK integrates insulin and mineralocorticoid regulation of epithelial sodiumtransport. Am J Physiol 280:F303–F313.
Webster MK, Goya L, Firestone GL. 1993. Immediate-early transcriptional regulation andrapid mRNA turnover of a putative serine/threonine protein kinase. J Biol Chem268:11482–11485.
Wulff P, Vallon V, Huang DY, Volkl H, Yu F, Richter K, Jansen M, Schlunz M, Klingel K, Loffing J,Kauselmann G, Bosl MR, Lang F, Kuhl D. 2002. Impaired renal Naþ retention in the sgk1-knockout mouse. J Clin Invest 110:1263–1268.




















