
'FREE' HEXANE CONTENT IN Results of a collaborative study ...
Upload
independentCategory
view
1download
0
Food Chemistry 114 (2009) 1245–1250
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Induction of apoptosis in U937 human leukaemia cells by the hexane fraction ofan extract of immature Citrus grandis Osbeck fruits
Hee-Kyung Lim a,1, Jeong Yong Moon b,1, Hana Kim b, Moonjae Cho a, Somi Kim Cho b,c,d,*
a Department of Medicine, Medical School, Cheju National University, Jeju 690-756, Republic of Koreab Faculty of Biotechnology, College of Applied Life Sciences, Cheju National University, 66 Jejudaehakno, Jeju 690-756, Republic of Koreac The Research Institute for Subtropical Agriculture and Biotechnology, Cheju National University, Jeju 690-756, Republic of Koread Subtropical Horticulture Research Institute, Cheju National University, Jeju 690-756, Republic of Korea
a r t i c l e i n f o
Article history:Received 25 August 2008Received in revised form 19 September2008Accepted 31 October 2008
Keywords:ApoptosisU937 human leukemia cellsCitrus grandis OsbeckBcl-2 familyCaspase-3 activity
0308-8146/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.foodchem.2008.10.088
* Corresponding author. Address: Faculty of BioteLife Sciences, Cheju National University, 66 JejudaehakKorea. Tel.: +82 64 754 3348; fax: +82 64 756 3351.
E-mail addresses: [email protected], gtp@korea1 These authors equally contributed to this work.
a b s t r a c t
The antiproliferative effect of an immature Citrus grandis Osbeck fruit extract was investigated usingU937 human leukaemia cells. Maximum cytotoxicity was observed using the hexane fraction (HF) ofthe extract. Cell death was dose-dependent (IC50 = ca. 60 lg/ml) and was characterised by chromatin con-densation, apoptotic body formation, and DNA fragmentation. The induction of apoptosis was confirmedby caspase-3 activity assays and by immunoblotting using antibodies against Bcl-2, Bax, poly(ADP-ribose) polymerase (PARP), caspase-9, and caspase-3. The molecular mechanism underlying HF-inducedapoptosis in U937 cells may involve a mitochondria-mediated signalling pathway, as demonstrated by anincrease in the Bax/Bcl-2 expression ratio. Analyses of the HF by gas chromatography (GC) and GC-massspectrometry (MS) tentatively identified 19 compounds, including c-sitosterol (17.5%), 7-methoxy-8-(2-oxo-3-methylbutyl) coumarin (6.8%), stigmasterol (3.8%), and campesterol (3.4%). Together, our resultsprovide the first evidence that the HF of an immature C. grandis Osbeck fruit extract induces apoptosisin U937 cells.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Several studies have reported that citrus fruits have anticancereffects, including the reduced proliferation of some cancer cells(Arias & Ramon-Laca, 2005) and the induction of apoptosis in hu-man gastric and colon cancer cells (Kim et al., 2005). Most Citrusspecies accumulate large amounts of flavonoids and limonoidsduring development (Castillo, Benavente, & Del Rio, 1992) andmuch is known about the cancer-preventing potential of the die-tary bioactive compounds in citrus fruits (Poulose, Harrie, & Patil,2006; Vanamala et al., 2006). Citrus grandis Osbeck, of the familyRutaceae, is widely cultivated in warm countries. In Korea, C. gran-dis Osbeck (‘dangyuza’ in Korean) is cultivated only on Jeju Island,where it has long been used as a herbal medicine. However, previ-ous studies on C. grandis Osbeck have focused mainly on changes inthe limonoid content during fruit growth and on the antioxidantactivity of the species (Lim, Yoo, Moon, Jeon, & Cho, 2006; Mokbel& Hashinaga, 2006). Studies on the anticancer effects of C. grandisOsbeck are limited.
ll rights reserved.
chnology, College of Appliedno, Jeju 690-756, Republic of
.com (S.K. Cho).
Apoptosis is characterised by distinct morphologic changes,including cell shrinkage, membrane blebbing, chromatin conden-sation, DNA fragmentation, and the formation of apoptotic bodies(Wyllie, 1997). Bcl-2 family proteins determine whether a cell livesor dies, by controlling the release of mitochondrial apoptogenicfactors, which are associated with death proteases called caspases(Oltval, Milliman, & Korsmeyer, 1993). Caspases, a class of cysteineproteases, are central players in the apoptotic process that trigger acascade of proteolytic cleavage events (Shah, Gapor, & Sylvester,2003). The activation of caspase-3 is an important downstreamevent in apoptosis (Earnshaw, Martins, & Kaufmann, 1999). Theregulation of apoptosis in normal and malignant cells has becomean area of intensive study in cancer research (Johnstone, Ruefli, &Lowe, 2002). The therapeutic application of apoptosis is currentlybeing considered as a model for the development of anti-tumourdrugs (Hong et al., 2003). It is therefore essential to identify novelapoptosis-inducing compounds that are candidate anti-tumouragents.
We sought to investigate the ability of dangyuza to induce apop-tosis and to determine the biochemical mechanisms underlyingapoptosis in U937 human leukaemic cells. To determine the spe-cies’ antiproliferative activity, we examined the effects of variouscitrus solvent fractions on cell viability, using a 3-(4,5-dimethylthi-azol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)-based assay.Annexin V-FITC and propidium iodide (PI) labelling assays, Hoechst

1246 H.-K. Lim et al. / Food Chemistry 114 (2009) 1245–1250
33342 staining, and analyses of DNA fragmentation confirmed theinduction of apoptosis. The expression of the anti-apoptotic pro-tein Bcl-2 and of the pro-apoptotic proteins Bax, caspase-9, andcaspase-3 was assessed by immunoblotting. Furthermore, to deter-mine which constituents in the extract may contribute to its activ-ity, a compositional analysis of the hexane fraction (HF) wasperformed by gas chromatography–mass spectrometry (GC–MS).Our results suggest that the HF contained bioactive compoundscapable of inducing apoptosis in U937 cells via caspase-3 activa-tion and regulation of the apoptosis-related proteins Bcl-2 and Bax.
2. Materials and methods
2.1. Reagents
C. grandis Osbeck fruits were obtained from the National Insti-tute of Subtropical Agriculture in Jeju Province, Korea. Botanicalsamples were taxonomically identified by Doctor Kim HY and avoucher specimen (No. SKC.070531) is deposited at the laboratoryof Professor Somi K. Cho in the Faculty of Biotechnology, College ofApplied Life Sciences, Cheju National University. Immature fruitswere collected from C. grandis Osbeck trees about 30 days afterflowering, while mature fruits were collected about 210 days afterflowering. RPMI 1640 medium, Dulbecco’s modified Eagle’s med-ium (DMEM), trypsin/EDTA, foetal bovine serum (FBS), penicillin,streptomycin, Hoechst 33342 dye, and Trizol were purchased fromInvitrogen Life Technologies Inc. (Grand Island, NY). Dimethyl sulf-oxide (DMSO), MTT, PI, RNase A, and caspase activity assay kitswere purchased from Sigma Chemical Co. (St. Louis, MO). An An-nexin V-FLOUS Staining Kit was purchased from BD Biosciences(Franklin Lakes, NJ). Anti-Bcl-2, -Bax, -cleaved caspase-3, -PARP,and -b-actin antibodies were purchased from Cell Signaling (Dan-vers, MA). A Bradford Protein Assay Kit and polyvinylidene fluoride(PVDF) membranes were purchased from Bio-Rad (Hercules, CA).
2.2. Preparation of the fruit extracts
Air-dried immature C. grandis Osbeck fruits were pulverisedusing a milling machine and extracted with 80% methanol by stir-ring for three days at room temperature (RT). The extract was fil-tered, concentrated with a vacuum rotary evaporator underreduced pressure, and lyophilised. The dried methanol extractswere then suspended in water (1 l) and further fractionated, byadditional extraction with n-hexane, chloroform, ethyl acetate,and n-butanol in a stepwise manner (Lim et al., 2006). Each extractpowder was dissolved in DMSO and diluted with phosphate-buf-fered saline (PBS, pH 7.4) to the desired final concentration.
2.3. Cell culture
HeLa, U937, HCT-15, Hep-G2, MCF-7, NCI-H460, and SNU-16cells obtained from the American Type Culture Collection weremaintained at 37 �C in a humidified atmosphere under 5% CO2 inRPMI 1640 or DMEM (Invitrogen Gibco BRL, Gaithersburg, MD)containing 10% heat-inactivated FBS, 100 U/ml penicillin, and100 lg/ml streptomycin. Exponentially growing cells were treatedwith various concentrations of the solvent fractions, as indicated.
2.4. Cell viability assay
The effect of the HF on the viability of various cancer cell lineswas determined by an MTT-based assay (Carmichael, Degraff, Gaz-dar, Minna, & Mitchell, 1987). Briefly, exponential-phase cells werecollected and transferred to a microtitre plate (104–105 cells perwell). The cells were then incubated for four days in the presence
of various concentrations of the HF. After incubation, 0.1 mg ofMTT were added to each well and the cells were incubated at37 �C for 4 h. The plates were then centrifuged at 1,500 rpm for20 min at RT and the medium was carefully removed. DMSO(150 ll) was then added to each well to dissolve the formazancrystals. The plates were read immediately at 540 nm on a Sunrisemicroplate reader (Tecan, Salzburg, Austria). The concentration atwhich growth was inhibited by 50% (the IC50 value) was deter-mined in triplicate for each well.
2.5. Microscopic observation of cellular morphology and nuclearfragmentation
U937 cells were placed in 24-well plates at 105 cells/ml. Sixteenhours after plating, the cells were treated with an aliquot of the HF.After 16 h, 1.5 ll of 10 mg/ml Hoechst 33342, a DNA-specific fluo-rescent dye, were added to the solution in each well and the plateswere incubated for 10 min at 37 �C. The stained cells were then ob-served under an Olympus fluorescence microscope.
2.6. Analysis of phosphatidylserine externalization
Cells (2 � 105 cells/ml) were treated with 100 lg/ml HF for 3 hthen suspended in 100 ll of Annexin V-binding buffer (10 mMHEPES/NaOH, pH 7.4, 140 mM NaCl, and 5 mM CaCl2). AnnexinV-FITC and PI were then added to a final concentration of 5 lg/ml. The cells were then incubated for 15 min at room temperaturein the dark and diluted with 300 ll of binding buffer. Flow cyto-metric analysis was performed using an EPICS-XL FACScan flowcytometer (Coulter, Miami, FL, USA). Cells positive for Annexin Vbut negative for PI fluorescence were identified as apoptotic(Zhong, Wang, Ho, & Chang, 2003).
2.7. Analysis of DNA fragmentation
U937 cells (105 cells/ml) were treated with 0, 25, 50, 100, or200 lg/ml HF for 24 h and then harvested. Genomic DNA was thenextracted from the cells using a Wizard Genomic DNA PurificationKit (Promega, Madison, WI). Four micrograms of the DNA wereelectrophoresed on a 1.8% agarose gel containing 0.1 lg/ml ethi-dium bromide and visualised under ultraviolet light (Paris, Berto-glio, & Breard, 2007).
2.8. Immunoblot analysis
After treatment, the cells were collected and washed twice withcold PBS. The cells were then lysed in lysis buffer (50 mM Tris-HCl,pH 7.5, 150 mM NaCl, 1% Nonidet P-40, 2 mM EDTA, 1 mM EGTA,1 mM NaVO3, 10 mM NaF, 1 mM DTT, 1 mM PMSF, 25 lg/ml apro-tinin, and 25 lg/ml leupeptin) and kept on ice for 30 min. The ly-sates were then centrifuged at 12,000g at 4 �C for 20 min; thesupernatants were stored at �70 �C until use. The protein concen-tration was determined by the Bradford method. Aliquots of the ly-sates (30–50 lg of protein) were separated by 8–12% SDS-PAGEand transferred onto a PVDF membrane using a glycine transferbuffer (192 mM glycine, 25 mM Tris-HCl, pH 8.8, and 20% metha-nol [v/v]). After blocking with 5% non-fat dried milk, the membranewas incubated for 2 h with primary antibodies, followed by 30 minwith secondary antibodies in milk containing Tris-buffered saline(TBS) and 0.5% Tween. Anti-human caspase-3, -caspase-9, -PARP,-Bcl-2, and -Bax antibodies were used at a 1:1000 dilution as theprimary antibodies, while horseradish peroxidase-conjugatedhorse anti-rabbit IgG (Vector Laboratories, Burlingame, CA, USA)was used at a 1:5000 dilution as the secondary antibody. Themembrane was then exposed to X-ray film. Protein bands were de-

Fig. 1. Effect of the hexane fraction on U937 cell viability. (A) Cells treated withsolvent fractions of immature dangyuza fruits. Untreated (DMSO alone) cells wereused as controls. Viability as determined by MTT reduction. The values presentedare the means ± SD (n = 4). Fractions: d, hexane (HF); s, chloroform (CF); h, ethylacetate (EF); ., butanol (BF); and j, aqueous residual (ARF).
Table 1Inhibitory effects of the hexane fraction on human cancer cell growth.
Cell line Characteristics IC50 (lg/mL)
U937 Histiocytic lymphoma cells 59.57HepG2 Hepatoblastoma cells 131.45HeLa Cervical cancer cells 286.52HCT-15 Colon cancer cells 86.55MCF-7 Breast cancer cells 143.76NCI-H460 Lung cancer cells 72.67SNU-16 Stomach cancer cells 90.0
Cells were treated with the hexane fraction for four days. IC50 is defined as theconcentration that resulted in a 50% decrease in cell number.
H.-K. Lim et al. / Food Chemistry 114 (2009) 1245–1250 1247
tected using the WEST-ZOL� plus Western Blot Detection System(iNtRON, Gyeonggi-do, Korea).
2.9. Caspase-3 activity assay
Caspase-3 activity was measured using a commercially avail-able kit (Sigma), according to the manufacturer’s protocol. U937cells were lysed in lysis buffer after treatment with the HF (25,50, 100, or 200 lg/ml) for 24 h. Active caspase-3 was used as a po-sitive control. All mixtures were incubated overnight in a humidi-fied environment at 37 �C. The amount of p-nitroaniline (p-NA)released from the substrate was measured at 405 nm.
2.10. GC analysis
GC analysis of the HF was carried out using an Agilent 6850 sys-tem equipped with a flame ionisation detector (FID) and HP-5MScapillary column (30 m � 0.25 mm I.D., 0.25 lm film thickness).The injector and detector were maintained at 220 and 280 �C,respectively. The column oven temperature was set at 100 �C forinjection (held for 5 min), raised at 5 �C/min to 200 �C (held for5 min), raised again at 5 �C/min to 300 �C, and finally held at300 �C for 10 min. Nitrogen was used as the carrier gas at a flowrate of 1.0 ml/min. The splitless injection volume was 1 ll. Quanti-tative data were obtained from the FID area percent data.
2.11. GC–MS
The bioactive HF was analysed by GC–MS using an Agilent6890N gas chromatograph coupled to an Agilent 5975N mass spec-trometer. An HP-5MS capillary column (30 m � 0.25 mm I.D.,0.25 lm film thickness) was used for gas chromatographic separa-tion of the analytes. The injection volume was 2 ll with a split ratioof 5:1; the injector temperature was held constant at 220 �C. He-lium was used as the carrier gas with an inlet pressure of10.48 psi, corresponding to a flow rate of 1.0 ml/min. The columnoven temperature was set at 100 �C (held for 5 min), raised at5 �C/min to 200 �C (held for 5 min), raised at 5 �C/min to 300 �C,and finally held at 300 �C for 10 min. The mass spectrometer wasoperated in the electron impact (EI) mode with an ionisation en-ergy of 70 eV. The transfer line was set at 280 �C, the quadrupoletemperature at 150 �C, and the source temperature at 230 �C. Thechemical constituents of the analyte were identified by comparingthe MS fragmentation patterns with those of Wiley 7th edited li-brary data of the GC–MS system.
3. Results
3.1. Effect of the HF on cell viability
Four organic solvents were used in a stepwise manner to extractthe anticancer components from a methanol extract of immaturecitrus fruits. The effect of various concentrations of the resultingfractions (n-hexane [HF], chloroform [CF], ethyl acetate [EF], n-butanol [BF], and aqueous residue [ARF]) on the growth of U937cells was subsequently examined by an MTT-based assay. Amongthe extracts tested, the HF appeared to be most potent. The percentviability of U937 cells exposed to the HF at 25, 50, 100, and 200 lg/ml was 68.2 ± 1.7, 27.1 ± 4.7, 14.9 ± 2.1, and 8.2 ± 0.4, respectively,compared with the controls (p < 0.001); moreover, the inhibition ofcell growth was dose-dependent (Fig. 1). We next investigated theeffect of the HF on the growth of other cancer cell lines. Each cellline responded differently to treatment with the HF; notably,HEL299 cells (normal human embryo lung fibroblasts) were notsusceptible to HF-induced toxicity (data not shown). As indicatedin Table 1, the IC50 values varied depending on the cell lines (e.g.,
60 lg/ml for U937 [histiocytic lymphoma cells] and 287 lg/mlfor HeLa [cervical cancer cells]).
3.2. Induction of apoptosis by the HF
To elucidate whether the HF-induced decrease in viability wasattributable to apoptosis, we used Annexin V-FITC and PI staining(Fig. 2A), nuclear staining with Hoechst 33342 (Fig. 2B), and DNAfragmentation assays (Fig. 2C).
Flow cytometric analysis using Annexin V/PI double-stainingrevealed that the percentage of Annexin V-FITC-positive/PI-nega-tive U937 cells that were apoptotic rather than necrotic exceededthe control level after 6 h of treatment with 60 lg/ml HF(Fig. 2A). Nuclear staining with Hoechst 33342, a fluorescentDNA-binding dye, revealed fragmented and condensed nuclei inthe HF-treated cells, which is typical of apoptotic cells (Fig. 2B).Lastly, cells treated with increasing concentrations of the HFshowed a progressive accumulation of fragmented DNA (Fig. 2C).While the HF did not cause significant DNA fragmentation at25 lg/ml, a ladder corresponding to DNA fragmentation by inter-nucleosomal cleavage associated with apoptosis was observed at100 and 200 lg/ml HF. Taken together, these data indicate thatthe HF induced apoptosis in U937 cells.
3.3. Altered Bcl-2 family activity and caspase activation by HFtreatment in U937 cells
To determine the mechanism of HF-induced apoptosis, theexpression of anti- and pro-apoptotic proteins following HF treat-ment was examined by immunoblotting. The time-course analysis
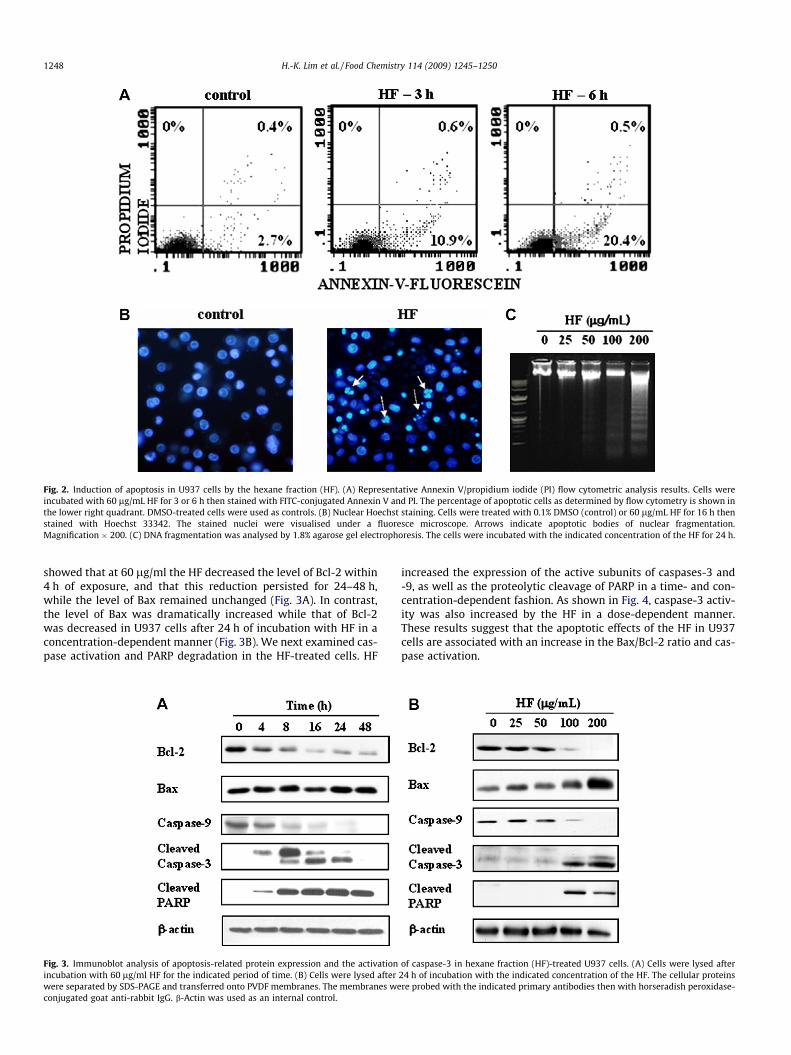
Fig. 2. Induction of apoptosis in U937 cells by the hexane fraction (HF). (A) Representative Annexin V/propidium iodide (PI) flow cytometric analysis results. Cells wereincubated with 60 lg/mL HF for 3 or 6 h then stained with FITC-conjugated Annexin V and PI. The percentage of apoptotic cells as determined by flow cytometry is shown inthe lower right quadrant. DMSO-treated cells were used as controls. (B) Nuclear Hoechst staining. Cells were treated with 0.1% DMSO (control) or 60 lg/mL HF for 16 h thenstained with Hoechst 33342. The stained nuclei were visualised under a fluoresce microscope. Arrows indicate apoptotic bodies of nuclear fragmentation.Magnification � 200. (C) DNA fragmentation was analysed by 1.8% agarose gel electrophoresis. The cells were incubated with the indicated concentration of the HF for 24 h.
1248 H.-K. Lim et al. / Food Chemistry 114 (2009) 1245–1250
showed that at 60 lg/ml the HF decreased the level of Bcl-2 within4 h of exposure, and that this reduction persisted for 24–48 h,while the level of Bax remained unchanged (Fig. 3A). In contrast,the level of Bax was dramatically increased while that of Bcl-2was decreased in U937 cells after 24 h of incubation with HF in aconcentration-dependent manner (Fig. 3B). We next examined cas-pase activation and PARP degradation in the HF-treated cells. HF
Fig. 3. Immunoblot analysis of apoptosis-related protein expression and the activationincubation with 60 lg/ml HF for the indicated period of time. (B) Cells were lysed afterwere separated by SDS-PAGE and transferred onto PVDF membranes. The membranes weconjugated goat anti-rabbit IgG. b-Actin was used as an internal control.
increased the expression of the active subunits of caspases-3 and-9, as well as the proteolytic cleavage of PARP in a time- and con-centration-dependent fashion. As shown in Fig. 4, caspase-3 activ-ity was also increased by the HF in a dose-dependent manner.These results suggest that the apoptotic effects of the HF in U937cells are associated with an increase in the Bax/Bcl-2 ratio and cas-pase activation.
of caspase-3 in hexane fraction (HF)-treated U937 cells. (A) Cells were lysed after24 h of incubation with the indicated concentration of the HF. The cellular proteinsre probed with the indicated primary antibodies then with horseradish peroxidase-
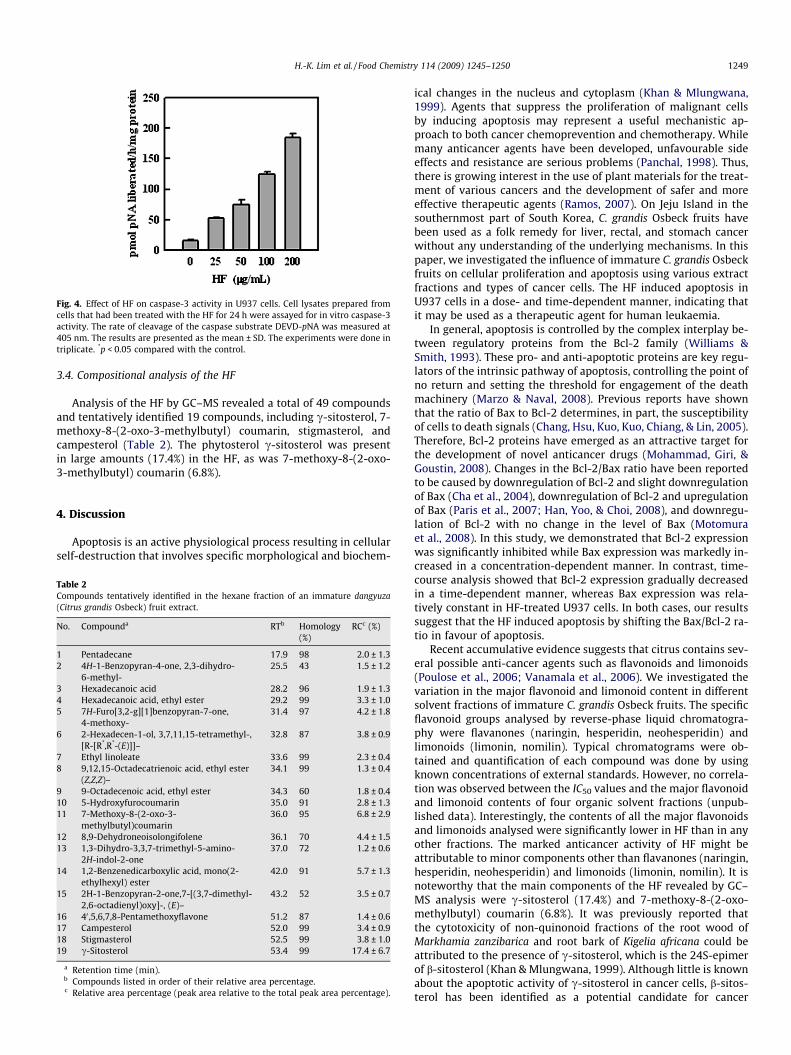
Fig. 4. Effect of HF on caspase-3 activity in U937 cells. Cell lysates prepared fromcells that had been treated with the HF for 24 h were assayed for in vitro caspase-3activity. The rate of cleavage of the caspase substrate DEVD-pNA was measured at405 nm. The results are presented as the mean ± SD. The experiments were done intriplicate. *p < 0.05 compared with the control.
H.-K. Lim et al. / Food Chemistry 114 (2009) 1245–1250 1249
3.4. Compositional analysis of the HF
Analysis of the HF by GC–MS revealed a total of 49 compoundsand tentatively identified 19 compounds, including c-sitosterol, 7-methoxy-8-(2-oxo-3-methylbutyl) coumarin, stigmasterol, andcampesterol (Table 2). The phytosterol c-sitosterol was presentin large amounts (17.4%) in the HF, as was 7-methoxy-8-(2-oxo-3-methylbutyl) coumarin (6.8%).
4. Discussion
Apoptosis is an active physiological process resulting in cellularself-destruction that involves specific morphological and biochem-
Table 2Compounds tentatively identified in the hexane fraction of an immature dangyuza(Citrus grandis Osbeck) fruit extract.
No. Compounda RTb Homology(%)
RCc (%)
1 Pentadecane 17.9 98 2.0 ± 1.32 4H-1-Benzopyran-4-one, 2,3-dihydro-
6-methyl-25.5 43 1.5 ± 1.2
3 Hexadecanoic acid 28.2 96 1.9 ± 1.34 Hexadecanoic acid, ethyl ester 29.2 99 3.3 ± 1.05 7H-Furo[3,2-g][1]benzopyran-7-one,
4-methoxy-31.4 97 4.2 ± 1.8
6 2-Hexadecen-1-ol, 3,7,11,15-tetramethyl-,[R-[R*,R*-(E)]]–
32.8 87 3.8 ± 0.9
7 Ethyl linoleate 33.6 99 2.3 ± 0.48 9,12,15-Octadecatrienoic acid, ethyl ester
(Z,Z,Z)–34.1 99 1.3 ± 0.4
9 9-Octadecenoic acid, ethyl ester 34.3 60 1.8 ± 0.410 5-Hydroxyfurocoumarin 35.0 91 2.8 ± 1.311 7-Methoxy-8-(2-oxo-3-
methylbutyl)coumarin36.0 95 6.8 ± 2.9
12 8,9-Dehydroneoisolongifolene 36.1 70 4.4 ± 1.513 1,3-Dihydro-3,3,7-trimethyl-5-amino-
2H-indol-2-one37.0 72 1.2 ± 0.6
14 1,2-Benzenedicarboxylic acid, mono(2-ethylhexyl) ester
42.0 91 5.7 ± 1.3
15 2H-1-Benzopyran-2-one,7-[(3,7-dimethyl-2,6-octadienyl)oxy]-, (E)–
43.2 52 3.5 ± 0.7
16 40 ,5,6,7,8-Pentamethoxyflavone 51.2 87 1.4 ± 0.617 Campesterol 52.0 99 3.4 ± 0.918 Stigmasterol 52.5 99 3.8 ± 1.019 c-Sitosterol 53.4 99 17.4 ± 6.7
a Retention time (min).b Compounds listed in order of their relative area percentage.c Relative area percentage (peak area relative to the total peak area percentage).
ical changes in the nucleus and cytoplasm (Khan & Mlungwana,1999). Agents that suppress the proliferation of malignant cellsby inducing apoptosis may represent a useful mechanistic ap-proach to both cancer chemoprevention and chemotherapy. Whilemany anticancer agents have been developed, unfavourable sideeffects and resistance are serious problems (Panchal, 1998). Thus,there is growing interest in the use of plant materials for the treat-ment of various cancers and the development of safer and moreeffective therapeutic agents (Ramos, 2007). On Jeju Island in thesouthernmost part of South Korea, C. grandis Osbeck fruits havebeen used as a folk remedy for liver, rectal, and stomach cancerwithout any understanding of the underlying mechanisms. In thispaper, we investigated the influence of immature C. grandis Osbeckfruits on cellular proliferation and apoptosis using various extractfractions and types of cancer cells. The HF induced apoptosis inU937 cells in a dose- and time-dependent manner, indicating thatit may be used as a therapeutic agent for human leukaemia.
In general, apoptosis is controlled by the complex interplay be-tween regulatory proteins from the Bcl-2 family (Williams &Smith, 1993). These pro- and anti-apoptotic proteins are key regu-lators of the intrinsic pathway of apoptosis, controlling the point ofno return and setting the threshold for engagement of the deathmachinery (Marzo & Naval, 2008). Previous reports have shownthat the ratio of Bax to Bcl-2 determines, in part, the susceptibilityof cells to death signals (Chang, Hsu, Kuo, Kuo, Chiang, & Lin, 2005).Therefore, Bcl-2 proteins have emerged as an attractive target forthe development of novel anticancer drugs (Mohammad, Giri, &Goustin, 2008). Changes in the Bcl-2/Bax ratio have been reportedto be caused by downregulation of Bcl-2 and slight downregulationof Bax (Cha et al., 2004), downregulation of Bcl-2 and upregulationof Bax (Paris et al., 2007; Han, Yoo, & Choi, 2008), and downregu-lation of Bcl-2 with no change in the level of Bax (Motomuraet al., 2008). In this study, we demonstrated that Bcl-2 expressionwas significantly inhibited while Bax expression was markedly in-creased in a concentration-dependent manner. In contrast, time-course analysis showed that Bcl-2 expression gradually decreasedin a time-dependent manner, whereas Bax expression was rela-tively constant in HF-treated U937 cells. In both cases, our resultssuggest that the HF induced apoptosis by shifting the Bax/Bcl-2 ra-tio in favour of apoptosis.
Recent accumulative evidence suggests that citrus contains sev-eral possible anti-cancer agents such as flavonoids and limonoids(Poulose et al., 2006; Vanamala et al., 2006). We investigated thevariation in the major flavonoid and limonoid content in differentsolvent fractions of immature C. grandis Osbeck fruits. The specificflavonoid groups analysed by reverse-phase liquid chromatogra-phy were flavanones (naringin, hesperidin, neohesperidin) andlimonoids (limonin, nomilin). Typical chromatograms were ob-tained and quantification of each compound was done by usingknown concentrations of external standards. However, no correla-tion was observed between the IC50 values and the major flavonoidand limonoid contents of four organic solvent fractions (unpub-lished data). Interestingly, the contents of all the major flavonoidsand limonoids analysed were significantly lower in HF than in anyother fractions. The marked anticancer activity of HF might beattributable to minor components other than flavanones (naringin,hesperidin, neohesperidin) and limonoids (limonin, nomilin). It isnoteworthy that the main components of the HF revealed by GC–MS analysis were c-sitosterol (17.4%) and 7-methoxy-8-(2-oxo-methylbutyl) coumarin (6.8%). It was previously reported thatthe cytotoxicity of non-quinonoid fractions of the root wood ofMarkhamia zanzibarica and root bark of Kigelia africana could beattributed to the presence of c-sitosterol, which is the 24S-epimerof b-sitosterol (Khan & Mlungwana, 1999). Although little is knownabout the apoptotic activity of c-sitosterol in cancer cells, b-sitos-terol has been identified as a potential candidate for cancer

1250 H.-K. Lim et al. / Food Chemistry 114 (2009) 1245–1250
chemotherapy (Moon, Kim, Choi, & Kim, 2008). Thus, we plan to ex-plore the potential of c-sitosterol as a therapeutic agent for the pre-vention of cancer in future studies. In addition, the apoptoticactivity of the HF could be attributed to the presence of 7-meth-oxy-8-(2-oxo-methylbutyl)coumarin, because coumarins and re-lated compounds have been shown to possess antithrombotic(Hoult & Paya, 1996), antimicrobial (Laurin et al., 1999), and anti-tumorigenic (Goel, Prasad, Parmar, Ghosh, & Saini, 2007) activities.
In conclusion, our study shows that the HF inhibited cancer cellproliferation, via the induction of apoptosis. HF-induced U937 celldeath was shown to be due to apoptosis by the demonstration ofthe induction of caspase-3 activity and the observation of cells con-taining fragmented nuclei and DNA. These results indicate that theHF of an immature C. grandis Osbeck fruit extract has anticanceractivity in vitro. Additional studies are needed to determine themolecular mechanisms of the active components and to evaluatethe potential in vivo anticancer activity of the extract.
Acknowledgment
This study was supported by the Technology Development Pro-gram of the Ministry of Agriculture and Forestry, Republic of Korea.
References
Arias, B. A., & Ramon-Laca, L. (2005). Pharmacological properties of citrus and theirancient and medieval uses in the Mediterranean region. Journal ofEthnopharmacology, 97, 89–95.
Carmichael, J., Degraff, W. G., Gazdar, A. F., Minna, J. D., & Mitchell, J. B. (1987).Evaluation of a tetrazolium-based semiautomated colorimetric assay:Assessment of chemosensitivity testing. Cancer Research, 47, 936–942.
Castillo, J., Benavente, O., & Del Rio, J. A. (1992). Naringin and neohesperidin levelsduring development of leaves, flower buds, and fruits of Citrus aurantium. PlantPhysiology, 99, 66–73.
Cha, Y. Y., Lee, E. O., Lee, H. J., Park, Y. D., Ko, S. G., Kim, D. H., Kim, H. M., Kang, I. C., &Kim, S. H. (2004). Methylene chloride fraction of Scutellaria barbata inducesapoptosis in human U937 leukemia cells via the mitochondrial signalingpathway. Clinica Chimica Acta, 348, 41–48.
Chang, J., Hsu, Y., Kuo, P., Kuo, Y., Chiang, L., & Lin, C. (2005). Increase of Bax/Bcl-XL
ratio and arrest of cell cycle by luteolin in immortalized human hepatoma cellline. Life Sciences, 76, 1883–1893.
Earnshaw, W. C., Martins, L. M., & Kaufmann, S. H. (1999). Mammalian caspases:Structure, activation, substrates, and functions during apoptosis. Annual Reviewof Biochemistry, 68, 383–424.
Goel, A., Prasad, A. K., Parmar, V. S., Ghosh, B., & Saini, N. (2007). 7,8-Dihydroxy-4-methylcoumarin induces apoptosis of human lung adenocarcinoma cells byROS-independent mitochondrial pathway through partial inhibition of ERK/MAPK signaling. FEBS Letters, 581, 2447–2454.
Han, M. H., Yoo, Y. H., & Choi, Y. H. (2008). Sanguinarine-induced apoptosis inhuman leukemia U937 cells via Bcl-2 downregulation and caspase-3 activation.Chemotherapy, 54, 157–165.
Hong, M. Y., Chapkin, R. S., Davidson, L. A., Turner, N. D., Morris, J. S., Carroll, R. J., &Lupton, J. R. (2003). Fish oil enhances targeted apoptosis during colon tumorinitiation in part by downregulating Bcl-2. Nutrition and Cancer, 46, 44–51.
Hoult, J. R., & Paya, M. (1996). Pharmacological and biochemical actions of simplecoumarins: Natural products with therapeutic potential. General Pharmacology,27, 713–722.
Johnstone, R. W., Ruefli, A. A., & Lowe, S. W. (2000). Apoptosis: A link betweencancer genetics and chemotherapy. Cell, 108, 153–164.
Khan, M. R., & Mlungwana, S. M. (1999). c-sitosterol, a cytotoxic sterol fromMarkhamia zanzibarica and Kigelia africana. Fitoterapia, 70, 96–97.
Kim, M. J., Park, H. J., Hong, M. S., Park, H. J., Kim, M. S., Leem, K. H., Kim, J. B., Kim, Y.J., & Kim, H. K. (2005). Citrus Reticulata blanco induces apoptosis in humangastric cancer cells SNU-668. Nutrition and Cancer, 51, 78–82.
Laurin, P., Ferroud, D., Schio, L., Klich, M., Dupuis-Hamelin, C., Mauvais, P., Lassaigne,P., Bonnefov, A., & Musicki, B. (1999). Structure-activity relationship in twoseries of aminoalkyl substituted coumarin inhibitors of gyrase B. Bioorganic andMedicinal Chemistry Letters, 9, 2875–2880.
Lim, H. K., Yoo, E. S., Moon, J. Y., Jeon, Y. J., & Cho, S. K. (2006). Antioxidant activity ofextracts from Dangyuja (Citrus grandis Osbeck) fruits produced in Jeju Island.Food Science and Biotechnology, 15, 312–316.
Marzo, I., & Naval, J. (2008). Bcl-2 family members as molecular targets in cancertherapy. Biochemical Pharmacology (Epub ahead of print).
Mohammad, R., Giri, A., & Goustin, A. S. (2008). Small-molecule inhibitors of Bcl-2family proteins as therapeutic agents in cancer. Recent Patents on Anti-cancerDrug Discovery, 3, 20–30.
Mokbel, M. S., & Hashinaga, F. (2006). Evaluation of the antioxidant activity ofextracts from buntan (Citrus grandis Osbeck) fruit tissues. Food Chemistry, 94,529–534.
Moon, D. O., Kim, M. O., Choi, Y. H., & Kim, G. Y. (2008). b-sitosterol induces G2/Marrest, endoreduplication, and apoptosis through the Bcl-2 and PI3k/Aktsignaling pathways. Cancer Letters, 26, 181–191.
Motomura, M., Kwon, K. M., Suh, S. J., Lee, Y. C., Kim, Y. K., Lee, I. S., Kim, M. S., Kwon,D. Y., Suzuki, I., & Kim, C. H. (2008). Propolis induces cell cycle arrest andapoptosis in human leukemic U937 cells through Bcl-2/Bax regulation.Environmental Toxicology and Pharmacology, 26, 61–67.
Oltval, Z. N., Milliman, C. L., & Korsmeyer, S. J. (1993). Bcl-2 heterodimerizes in vivowith a conserved homolog, bax, that accelerates programmed cell death. Cell,74, 609–619.
Panchal, R. G. (1998). Novel therapeutic strategies to selectively kill cancer cells.Biochemical Pharmacology, 55, 247–252.
Paris, C., Bertoglio, J., & Breard, J. (2007). Lysosomal and mitochondrial pathways inmilterfosine-induced apoptosis in U937 cells. Apoptosis, 12, 1257–1267.
Poulose, S. M., Harrie, E. D., & Patil, B. S. (2006). Antiproliferative effects of citruslimonoids against human neuroblastoma and colonic adenocarcinoma cells.Nutrition and Cancer, 56, 103–112.
Ramos, S. (2007). Effects of dietary flavonoids on apoptotic pathways related tocancer chemoprevention. Journal of Nutritional Biochemistry, 18, 427–442.
Shah, S., Gapor, A., & Sylvester, P. W. (2003). Role of caspase-8 activation inmediating vitamin E-induced apoptosis in murine mammary cancer cells.Nutrition and Cancer, 45, 236–246.
Vanamala, J., Leonardi, T., Patil, B. S., Taddeo, S. S., Murphy, M. E., Pike, L. M., Chapkin,R. S., Lupton, J. R., & Turner, N. D. (2006). Suppression of colon carcinogenesis bybioactive compounds in grapefruit. Carcinogenesis, 27, 1257–1265.
Williams, G. T., & Smith, C. A. (1993). Molecular regulation of apoptosis: Geneticcontrols on cell death. Cell, 74, 777–779.
Wyllie, A. H. (1997). Apoptosis: An overview. British Medical Bulletin, 53, 451–465.Zhong, W. B., Wang, C. Y., Ho, K. J., & Chang, T. C. (2003). Magnolol induces apoptosis
in human leukemia cells via cytochrome c release and caspase activation.Anticancer Drugs, 14, 211–217.
Copyright © 2022 FDOKUMEN




















