
Melatonin reduces early changes in intramitochondrial cardiolipin during apoptosis in U937 cell line
9
Melatonin reduces early changes in intramitochondrial cardiolipin during apoptosis in U937 cell line F. Luchetti a, * , B. Canonico b , F. Mannello c , C. Masoni c , A. D’Emilio a , M. Battistelli a , S. Papa a,b , E. Falcieri a,d a Istituto di Scienze Morfologiche, Universita ` degli Studi di Urbino ‘‘Carlo Bo’’ Urbino, Italy b Centro di Citometria e Citomorfologia, Universita ` degli Studi di Urbino ‘‘Carlo Bo’’ Urbino, Italy c Istituto di Istologia e Analisi di Laboratorio, Universita ` degli Studi di Urbino ‘‘Carlo Bo’’ Urbino, Italy d ITOI-CNR, Istituto Ortopedici Rizzoli, Bologna, Italy Received 18 April 2006; accepted 28 August 2006 Available online 5 September 2006 Abstract Cardiolipin (CL) is found exclusively in the inner mitochondrial membrane. CL deficiency leads to an alteration in the stability of mitochondrial membranes, to an increased permeability as well as a decreased respiratory rate, and therefore to mitochondria which are completely dysfunctional. It is known that reactive oxygen species (ROS) cause a decrease and a variation in CL content, concom- itantly the formation of the mitochondrial permeability transition pore facilitates the release of cytochrome c (cyt c) into the cytosol. Melatonin (Mel), the secretory product of the pineal gland, is a potent and efficient endogenous radical scavenger. It has been shown to protect, various biomolecules, such as DNA, membrane lipids, and cytosolic proteins from oxidative damage. To evaluate the protective role of Mel, we have studied U937 cells treated with UV-B irradiation. In our model, the administration of 1 mM Mel before UV-B irradiation showed a significant protection from apoptotic cell death, in particular, mitochondrial structure and function were preserved through apoptotic pathways when cells were preincubated with 1 mM Mel before UV-B exposure. The cardiolipin-sensitive probe 10-nonyl acridine orange (NAO) was used to monitor changes in mitochondrial lipids. Our data suggest that the Mel treatment protects CL from ROS and this suggests a possible link with the reduction of the apoptotic phenomenon. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Apoptosis; Cardiolipin; Mitochondria 1. Introduction Cardiolipin (CL), a polyunsaturated acidic phospho- lipid, is found exclusively in bacterial and mitochondrial membranes where it is intimately associated with the enzyme complexes of the respiratory chain (Daum, 1985). In particular, CL is localized in the inner membrane of mitochondria, but small amounts can also be detected in a highly purified preparation of the outer membrane (Hovius et al., 1993). Several authors have reported that CL is concentrated in intermembrane contact sites, where it may have access to the outer membrane and ultimately, to the external surface of the mitochondrion. Another problem is the transmembrane distribution of CL within the inner membrane. Phospholipase digestion of the inner membrane indicates that most of the CL faces are located in the mitochondrial matrix, while binding of creatine kinase, a protein found in the intermembrane spaces, demonstrates that some CL is exposed on the sur- face of the membrane (Schlame et al., 2000). CL interacts with a large number of mitochondrial proteins, most of which are located in the inner membrane (Jacobson et al., 2002). In mitochondria, most cyt c is membrane bound: at least 15% is tightly protein bound (due to both electrostatic and hydrophobic interactions), the remaining is loosely bound (likely electrostatically), interacting with 0887-2333/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2006.08.003 * Corresponding author. Tel.: +39 0722 304316; fax: +39 0722 304242. E-mail address: [email protected] (F. Luchetti). www.elsevier.com/locate/toxinvit Toxicology in Vitro 21 (2007) 293–301
Transcript of Melatonin reduces early changes in intramitochondrial cardiolipin during apoptosis in U937 cell line

www.elsevier.com/locate/toxinvit
Toxicology in Vitro 21 (2007) 293–301
Melatonin reduces early changes in intramitochondrialcardiolipin during apoptosis in U937 cell line
F. Luchetti a,*, B. Canonico b, F. Mannello c, C. Masoni c, A. D’Emilio a,M. Battistelli a, S. Papa a,b, E. Falcieri a,d
a Istituto di Scienze Morfologiche, Universita degli Studi di Urbino ‘‘Carlo Bo’’ Urbino, Italyb Centro di Citometria e Citomorfologia, Universita degli Studi di Urbino ‘‘Carlo Bo’’ Urbino, Italy
c Istituto di Istologia e Analisi di Laboratorio, Universita degli Studi di Urbino ‘‘Carlo Bo’’ Urbino, Italyd ITOI-CNR, Istituto Ortopedici Rizzoli, Bologna, Italy
Received 18 April 2006; accepted 28 August 2006Available online 5 September 2006
Abstract
Cardiolipin (CL) is found exclusively in the inner mitochondrial membrane. CL deficiency leads to an alteration in the stability ofmitochondrial membranes, to an increased permeability as well as a decreased respiratory rate, and therefore to mitochondria whichare completely dysfunctional. It is known that reactive oxygen species (ROS) cause a decrease and a variation in CL content, concom-itantly the formation of the mitochondrial permeability transition pore facilitates the release of cytochrome c (cyt c) into the cytosol.Melatonin (Mel), the secretory product of the pineal gland, is a potent and efficient endogenous radical scavenger. It has been shownto protect, various biomolecules, such as DNA, membrane lipids, and cytosolic proteins from oxidative damage. To evaluate theprotective role of Mel, we have studied U937 cells treated with UV-B irradiation. In our model, the administration of 1 mM Mel beforeUV-B irradiation showed a significant protection from apoptotic cell death, in particular, mitochondrial structure and function werepreserved through apoptotic pathways when cells were preincubated with 1 mM Mel before UV-B exposure. The cardiolipin-sensitiveprobe 10-nonyl acridine orange (NAO) was used to monitor changes in mitochondrial lipids. Our data suggest that the Mel treatmentprotects CL from ROS and this suggests a possible link with the reduction of the apoptotic phenomenon.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Apoptosis; Cardiolipin; Mitochondria
1. Introduction
Cardiolipin (CL), a polyunsaturated acidic phospho-lipid, is found exclusively in bacterial and mitochondrialmembranes where it is intimately associated with theenzyme complexes of the respiratory chain (Daum, 1985).In particular, CL is localized in the inner membrane ofmitochondria, but small amounts can also be detected ina highly purified preparation of the outer membrane(Hovius et al., 1993). Several authors have reported thatCL is concentrated in intermembrane contact sites, where
0887-2333/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tiv.2006.08.003
* Corresponding author. Tel.: +39 0722 304316; fax: +39 0722 304242.E-mail address: [email protected] (F. Luchetti).
it may have access to the outer membrane and ultimately,to the external surface of the mitochondrion.
Another problem is the transmembrane distribution ofCL within the inner membrane. Phospholipase digestionof the inner membrane indicates that most of the CL facesare located in the mitochondrial matrix, while binding ofcreatine kinase, a protein found in the intermembranespaces, demonstrates that some CL is exposed on the sur-face of the membrane (Schlame et al., 2000). CL interactswith a large number of mitochondrial proteins, most ofwhich are located in the inner membrane (Jacobsonet al., 2002). In mitochondria, most cyt c is membranebound: at least 15% is tightly protein bound (due to bothelectrostatic and hydrophobic interactions), the remainingis loosely bound (likely electrostatically), interacting with

294 F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301
the membrane from which it can be readily detached withhigh ionic salt solutions (Cortese et al., 1995). Cyt c isbound to the mitochondrial inner membrane by means ofits association with CL which facilitates electron transportbetween complexes III and IV of the respiratory chain.Complexes III and IV associate to form a supercomplexwhose functional activity requires inner mitochondrialmembrane lipid CL. They generate an electrochemical pro-ton gradient across the inner membrane in which CL mayact as a proton trap (Schagger, 2002).
CL deficiency leads to alterations in the stability of mito-chondrial membranes, increased permeability, decreasedrespiratory rate, and to totally dysfunctional mitochondria.The consequences of CL oxidation are as follows:
(a) Oxidized CL does not tightly bind cyt c and isreleased into the intermembrane space.
(b) Mitochondrial permeability transition pore (MPTP)formation occurs (Kagan et al., 2004).
It is known that the point of no-return of apoptotic celldeath is mainly determined by two interrelated phenomena,namely mitochondrial membrane permeabilization (MMP)and caspase activation. MMP culminates in the completeloss of the barrier function of the outer mitochondrialmembrane and the consequent release of potentially toxicproteins that are normally secured in the intermembranespace between the inner and the outer mitochondrial mem-branes. Such cytotoxic proteins include caspase-indepen-dent death effectors, nuclease and protease, as well ascaspase activators, namely cyt c, Smac/DIABLO andOmi/Htr2 (Zanzamani and Kroemer, 2003).
All aerobic cells produce ROS as intermediate productsof various metabolic processes. The special feature of theseagents is their high oxidative reactivity, having a significanteffect on redox-sensitive cellular components, primarilyproteins, lipids and nucleic acids (Zorov et al., 2005).Apparently, pro-oxidative and anti-oxidative systems coex-ist in a state of dynamic balance: it is known that an excessof ROS results in programmed cell death and thus mito-chondria operate not only as a power stations for aerobiclife, but also as amplifiers or, sometimes even sources ofapoptotic signals. These two alternative functions are inter-related because electron transfer from respiratory sub-strates to oxygen, involved in energy conservation, isaccompanied by the formation of ROS that act as inducersor secondary messengers of apoptosis. The unsaturatednature of the acyl chains of CL and its proximity toROS-producing sites in the inner membrane of mitochon-dria makes this biomolecule susceptible to peroxidationby ROS (Iver_son and Orrenius, 2004). The decrease inCL content affects cyt c binding to the inner mitochondrialmembrane, thus leading to higher levels of soluble cyt c inthe mitochondrial intermembrane space and to its releasein the cytosol, which represents a critical event in the initi-ation of mitochondria-mediated apoptosis (Kroemer andReed, 2000). The natural substances protecting against an
excess of ROS are reduced glutathione, vitamins, albumins,globulins, and free fatty acids.
Mel was originally characterized as a molecule producedand secreted by the pineal gland, but in recent years itssynthesis also has been revealed in the gastrointestinaltract, eye, bone marrow, immune system, (especially inlymphocytes), the ovary and even in plants. Mel acts as areceptor-independent autocrine and paracrine antioxi-dant, a direct and indirect radical scavenger (Fischeret al., 2006).
Being both an endogenous antioxidant and a non-toxiccompound, it may have an important role in protectingcells from genetic damage due to free radicals.
In this work, we have evaluated whether Mel pretreat-ment prevents the oxidation of CL caused by UV-B expo-sure, its involvement in the apoptotic phenomenon, and itseffect on specific components of the inner mitochondrialmembrane.
2. Materials and methods
2.1. Cell culture
Human myelomonocytic cell line, U937, was grown inRPMI 1640 supplemented with 10% heat-inactivated fetalbovine serum, 2 mM glutamine, 25 mM HEPES pH 7.5,and 1% antibiotics and was maintained at 37 �C in humid-ified air with 5% CO2. Cell viability was assessed by trypanblue exclusion test. For the induction of apoptosis, U937cells (seeded at 1 · 106 cells/ml) were exposed to UV-B(312 nm) irradiation for 30 min at room temperature, fol-lowed by postincubation for 1, 2, 3 and 4 h at 37 �C inhumidified air with 5% CO2. The treatment of U937 cellswith 1 mM Mel was performed immediately before UV-Birradiation (Luchetti et al., 2006).
2.2. Measurement of the production of superoxide anions
The production of superoxide anions (O��2 ) was deter-mined according to the nitro blue tetrazolium (NBT) testpreviously described (Mochida et al., 1998). Briefly, U937cells after exposure to UV-B were cultured for the appro-priate post-incubation time in the absence or presence of1 mM Mel; after washing, 1 · 106 cells were incubated for30 min at 37 �C with 0.2% of NBT solution. The opticaldensity of insoluble formazan produced by O��2 was readat 630 nm by UV–vis spectrophotometer against an appro-priate blank and standard curve.
2.3. Light and electron microscopy
Differently treated U937 pellets were immediately fixedin 2.5% glutaraldehyde in 0.1 M Sorensen phosphate buffer(pH 7.3). The cells were then postfixed in 1% OsO4 in thesame buffer, dehydrated with ethanol and embedded inaraldite as previously described (Battistelli et al., 2004).Semithin sections, collected on slides, were stained with

F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301 295
1% toluidine blue at 60 �C and observed with a LeicaDMLB microscope (Leica Microsystems, Wetzlar, Ger-many). For the ultrastructural analysis, thin sections weredeposited on 300-mesh nickel grids and stained with uranylacetate and lead citrate. They were observed with a PhilipsCM10 transmission electron microscope (FEI Company,Italy) at 80 KV.
2.4. Flow cytometry
Changes in DW and early apoptosis were evaluated bydual staining and evaluated with Mitotracker CMXRos(MT) and Annexin-V (FITC ANX-V). Thirty minutesbefore the end of each incubation period, 500 ll of cell sus-pension were collected from each flask, transferred into aFACS tube and the volume brought up to 1 ml with com-plete medium. These samples were stained with 50 nM MTfor 30 min at 37 �C. At the end of incubation times, theywere washed once in PBS and resuspended in 300 ll ofFITC ANX-V binding buffer (10 mM HEPES/NaOH,pH 7.4, 140 mM NaCl, 2.5 mM CaCl2), stained with 5 llFITC ANX-V and incubated for 10 min at room tempera-ture. Then, to remove excess FITC ANX-V, cells werewashed with binding buffer, resuspended in PBS and ana-lyzed by a FACScan flow cytometer equipped with Cell-Quest software (Salvioli et al., 2000).
2.5. Measurement of CL content
The CL sensitive probe, 10-nonyl-acridine orange(NAO), was used to monitor changes in mitochondrialCL (Ferry et al., 2005). After exposure to UV-B and appro-priate post-incubations, cells were collected by centrifuga-tion, washed in PBS, resuspended in PBS containing100 nM NAO and incubated for 30 min. FL1 fluorescencewas quantified by flow cytometry.
3. Results
Mel is known to be a potent endogenous radical scaven-ger. As shown in Fig. 1, UV-B exposure induces an increase
Fig. 1. Effects of Mel on the production of superoxide anions by NBT teston UV-B exposed U937 cells; irradiation in the presence or absence ofmelatonin was followed by different lengths of post-incubation time. Dataare means ± ES of three independent experiments carried out in triplicate.
Fig. 2. Control U937 cells in semithin (A) and thin (B, C) sections. Celland mitochondrial morphology appears well preserved. A, Bar = 20 lm;B, C, Bar = 1 lm.
296 F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301
in the production of O��2 . The increase in the generation ofO��2 is correlated with the time of postincubation. In fact,
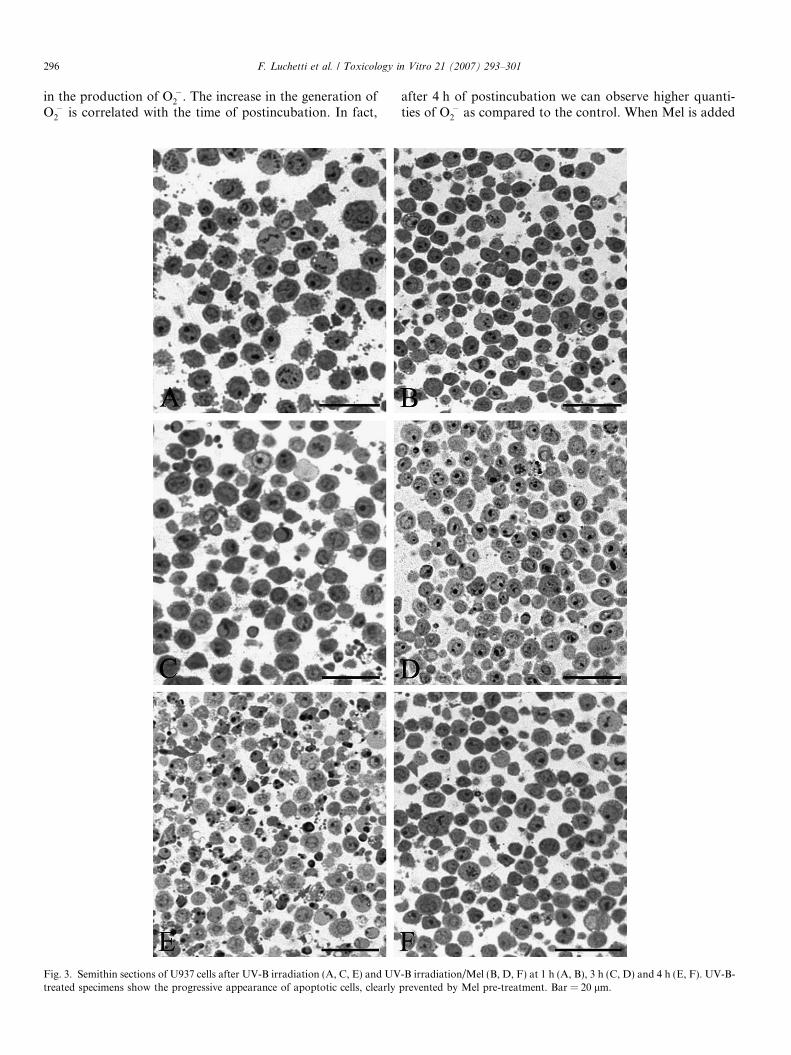
Fig. 3. Semithin sections of U937 cells after UV-B irradiation (A, C, E) and UVtreated specimens show the progressive appearance of apoptotic cells, clearly
after 4 h of postincubation we can observe higher quanti-ties of O��2 as compared to the control. When Mel is added
-B irradiation/Mel (B, D, F) at 1 h (A, B), 3 h (C, D) and 4 h (E, F). UV-B-prevented by Mel pre-treatment. Bar = 20 lm.
F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301 297
immediately before UV-B treatment, a reduction in theproduction of O��2 is observed: in particular there is adecrease in O��2 production up to 2 h of postincubation,after which we can observe an increase in the generationof O��2 at 3 and 4 h of postincubation. It is important tonote that in all conditions, Mel pre-treatment induces adecrease in O��2 generation when compared to UV-B treat-ment alone.
Fig. 2 shows control U937 cells under light (A) and elec-tron microscopy (B, C). Cell morphology appears well pre-served both in shape and organelle features. In particular,mitochondria show a regular distribution of matrix densityand christae.
Semithin sections of U937 cells after UV-B irradiation(A, C, E) and UV-B irradiation/Mel (B, D, F) are shownin Fig. 3, at 1 h (A, B), 3 h (C, D) and 4 h (E, F). UV-B-
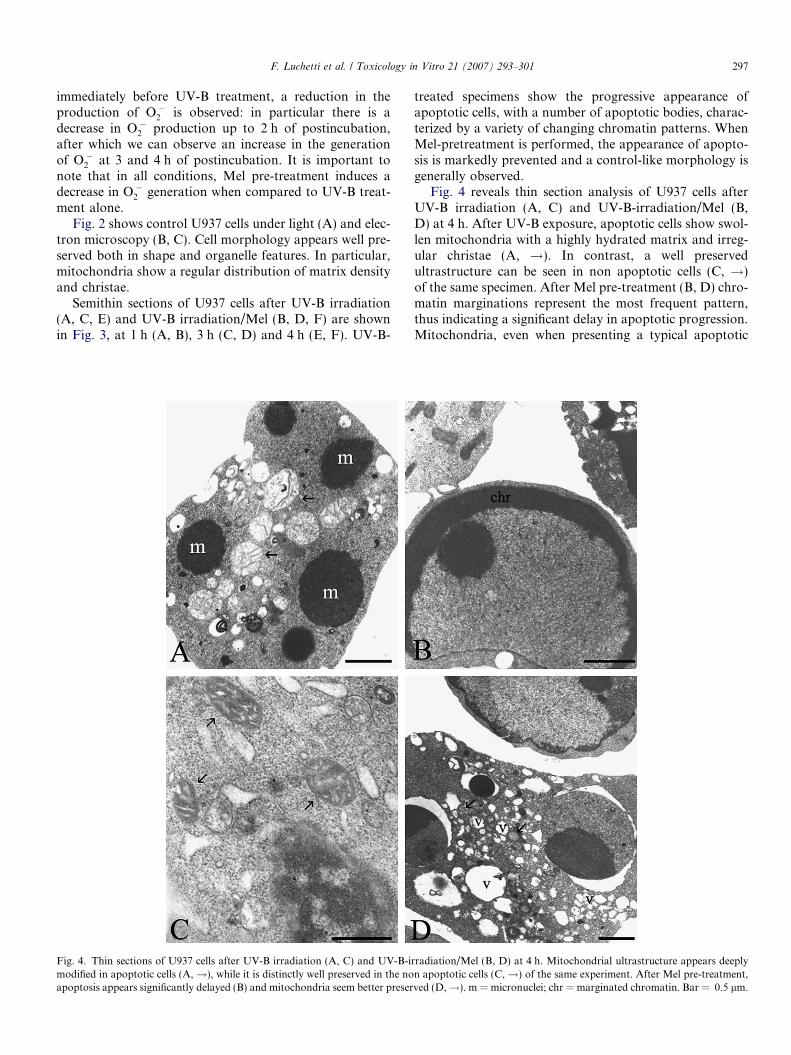
Fig. 4. Thin sections of U937 cells after UV-B irradiation (A, C) and UV-B-imodified in apoptotic cells (A,!), while it is distinctly well preserved in the noapoptosis appears significantly delayed (B) and mitochondria seem better preser
treated specimens show the progressive appearance ofapoptotic cells, with a number of apoptotic bodies, charac-terized by a variety of changing chromatin patterns. WhenMel-pretreatment is performed, the appearance of apopto-sis is markedly prevented and a control-like morphology isgenerally observed.
Fig. 4 reveals thin section analysis of U937 cells afterUV-B irradiation (A, C) and UV-B-irradiation/Mel (B,D) at 4 h. After UV-B exposure, apoptotic cells show swol-len mitochondria with a highly hydrated matrix and irreg-ular christae (A, !). In contrast, a well preservedultrastructure can be seen in non apoptotic cells (C, !)of the same specimen. After Mel pre-treatment (B, D) chro-matin marginations represent the most frequent pattern,thus indicating a significant delay in apoptotic progression.Mitochondria, even when presenting a typical apoptotic
rradiation/Mel (B, D) at 4 h. Mitochondrial ultrastructure appears deeplyn apoptotic cells (C,!) of the same experiment. After Mel pre-treatment,ved (D,!). m = micronuclei; chr = marginated chromatin. Bar = 0.5 lm.

Fig. 5. Flow cytometric analysis of U937 cells. Fluorescein isothiocianate-conjugated FITCANX-V and MT. A: control cells, B: U937 cells incubated withMel alone, C: UV-B irradiated U937 cell, D: Mel-pretreated UV-B exposed cells. Representative images are taken from three independent experiments.
298 F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301
cytoplasmic vacuolization, seem better preserved (D, !)than with UV-B exposition alone.
Changes in cardiolipin structure and distribution occurprior to or concomitantly with membrane exposure tophosphatidilserine, but only after the onset of an overpro-duction of ROS.
Labelling with FITC ANX-V and MT was carried outto evaluate the apoptotic cell-population and its correla-tion with mitochondrial function. In Fig. 5 we can observethat UV-B exposure induces a high percentage of apoptoticcells (40%) revealed as FITC ANX-V positivity (C), absentboth in controls (3%) (A) and Mel (2%) treatment alone(B). The UV-B treatment induces a decrease in MT positiv-ity; in fact in control conditions, about 93% of cells are MTpositive. This percentage decreases to 55% in UV-B condi-tions and is restored at 70% in Mel treated cells. It is inter-esting to observe that when FITC ANX-V positivityincreases, there is a concomitant decrease in MT positivity.This correlation between FITC ANX-V and MT is impor-tant because it allows us to establish a relationship betweenthe apoptotic phenomenon and the mitochondrialpathway.
To evaluate whether a variation in CL constitutes anearly apoptotic event, all the cells were treated withUV-B for 30 min and postincubated for 4 h. Two gateswere performed on apoptotic/necrotic cells and those stillliving. For this purpose NAO, a specific cardiolipin dye,was used.
In Fig. 6 fluorescence from NAO is visible. Histogram Arepresents control cells, histogram B represents melatoninpretreated cells, histogram C represents UV-B treated cellsand Fig. 6D represents cells to which melatonin was addedimmediately before UV-B treatment. In Fig. 6 the altera-tions in cardiolipin elicited by UV-B exposure in U937 cellswere evaluated using NAO. A difference in fluorescenceintensity can be observed between UV-B and Mel condi-tions. In particular UV-B treatment induces a peroxidationof cardiolipin and a shift in the intensity of fluorescence canbe seen in the histograms. The addition of 1 mM Mel pre-vents this peroxidation and a higher signal in NAO fluores-cence is evident.
Fig. 7 shows the apoptotic/necrotic cell population witha different intensity of fluorescence of NAO, since thesequence of the histograms is the same as in Fig. 6.
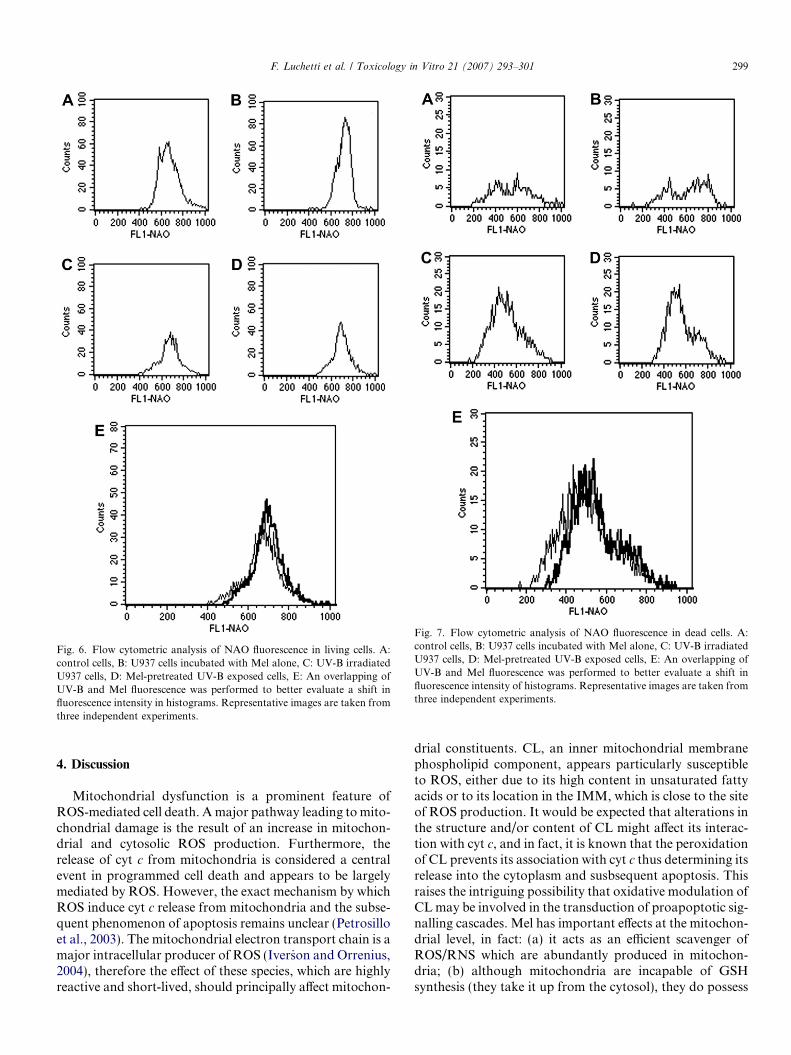
Fig. 6. Flow cytometric analysis of NAO fluorescence in living cells. A:control cells, B: U937 cells incubated with Mel alone, C: UV-B irradiatedU937 cells, D: Mel-pretreated UV-B exposed cells, E: An overlapping ofUV-B and Mel fluorescence was performed to better evaluate a shift influorescence intensity in histograms. Representative images are taken fromthree independent experiments.
Fig. 7. Flow cytometric analysis of NAO fluorescence in dead cells. A:control cells, B: U937 cells incubated with Mel alone, C: UV-B irradiatedU937 cells, D: Mel-pretreated UV-B exposed cells, E: An overlapping ofUV-B and Mel fluorescence was performed to better evaluate a shift influorescence intensity of histograms. Representative images are taken fromthree independent experiments.
F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301 299
4. Discussion
Mitochondrial dysfunction is a prominent feature ofROS-mediated cell death. A major pathway leading to mito-chondrial damage is the result of an increase in mitochon-drial and cytosolic ROS production. Furthermore, therelease of cyt c from mitochondria is considered a centralevent in programmed cell death and appears to be largelymediated by ROS. However, the exact mechanism by whichROS induce cyt c release from mitochondria and the subse-quent phenomenon of apoptosis remains unclear (Petrosilloet al., 2003). The mitochondrial electron transport chain is amajor intracellular producer of ROS (Iver_son and Orrenius,2004), therefore the effect of these species, which are highlyreactive and short-lived, should principally affect mitochon-
drial constituents. CL, an inner mitochondrial membranephospholipid component, appears particularly susceptibleto ROS, either due to its high content in unsaturated fattyacids or to its location in the IMM, which is close to the siteof ROS production. It would be expected that alterations inthe structure and/or content of CL might affect its interac-tion with cyt c, and in fact, it is known that the peroxidationof CL prevents its association with cyt c thus determining itsrelease into the cytoplasm and susbsequent apoptosis. Thisraises the intriguing possibility that oxidative modulation ofCL may be involved in the transduction of proapoptotic sig-nalling cascades. Mel has important effects at the mitochon-drial level, in fact: (a) it acts as an efficient scavenger ofROS/RNS which are abundantly produced in mitochon-dria; (b) although mitochondria are incapable of GSHsynthesis (they take it up from the cytosol), they do possess

300 F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301
GPx and GRd for GSH cycling, the enzymes of which arestimulated by Mel and (c) it has been shown to display anti-apoptotic effects, on the apoptotic signals originating inmitochondria.
Unexpectedly, Mel may be present in higher concentra-tions in mitochondria than elsewhere, including bloodserum (Acuna-Castroviejo et al., 2002).
In this work, we have investigated the possible role ofMel in the prevention of CL peroxidation. The resultsobtained demonstrate that MEL pre-treatment reducesthe production of O��2 , and consequently, the number ofapoptotic cells. Furthermore, we have demonstrated thatthe increase in the number of apoptotic cells, revealed asFITC ANX-V positivity, correspond to a decrease in mito-chondrial function, demonstrated by a reduction in the pos-itivity of MT which is accumulated in active mitochondria.Because CL is the most abundant mitochondrial phospho-lipid and helps to maintain the impermeability of the innermitochondrial membrane, CL content has been quantifiedby flow cytometry in U937 cells treated with UV-B aloneand in combination with Mel. Our data demonstrate thatafter 1 and 2 h of postincubation, where few apoptotic cellscan be observed, there is a small modification in CL contentevaluated by NAO fluorescence. At 3 and 4 h of postincuba-tion, in the presence of a high number of apoptotic cells,there is a higher modification in CL content (data notshown). CL is an anionic phospholipid found exclusivelyin the inner mitochondrial membrane of eukaryotic cells(Sorice et al., 2004). Its unique structure among phospholip-ids confers fluidity and stability both to the mitochondrialmembrane and to contact sites between the inner and outermitochondrial membranes. Several studies strongly suggestthat oxidative modification of CL could represent an earlyevent in apoptosis and that ROS can cause a decrease inCL content. Therefore, we have evaluated NAO fluores-cence in treated cells, distinguishing cells which are still alivefrom apoptotic/necrotic cells. Our data suggest that
(1) melatonin pre-treatment decreases the formation ofO��2 ;
(2) melatonin pre-treatment affects apoptosis;(3) this prevention also occurs through the mitochon-
drial pathway;(4) some components of the inner mitochondrial mem-
brane are involved;(5) melatonin seems to prevent the peroxidation of cardi-
olipin in the early stages of apoptosis.
In conclusion, data reported in this study together withour previous results, suggest a new function for melatoninin U937.
Acknowledgement
The work was supported by MIUR, COFIN 2004 andby the University of Urbino.
References
Acuna-Castroviejo, D., Escames, G., Carozo, A., Leon, J., Khaldy, H.,Reiter, R.J., 2002. Melatonin, mitochondrial homeostasis and mito-chondrial-related disease. Current Topics in Medicinal Chemistry 2,133–152.
Battistelli, M., Burattini, S., Pomini, F., Scavo, M., Caruso, A., Falcieri,E., 2004. An ultrastructural study on human placenta from intra-uterine growth retardation cases. Microscopi Research and Techniques65, 150–158.
Cortese, J., Voglino, A.L., Hackenbrock, C.R., 1995. Persistence ofcytochrome c binding to membranes at physiological mitochondrialintermembrane space ionic strength. Biochimica et Biophysica Acta1228, 216–228.
Daum, G., 1985. Lipids of mitochondria. Biochimica et Biophysica Acta822, 1–42.
Ferry, S., Degli Esposti, M., Ndebele, K., Philimon, G., Knight, D.,Rosenquist, M., Khosravi-Far, R., 2005. Tumor necrosis factor-related apoptosis-inducing ligand alters mitochondrial membranelipids. Cancer Research 65, 8286–8297.
Fischer, T.W., Zbytek, B., Sayre, R.M., Apostolov, E.O., Basnakian,A.G., Sweatman, T.W., Wortsman, J., Elsner, P., Slominski, A., 2006.Melatonin increases survival of HaCaT keratinocytes by suppressingUV-induced apoptosis. Journal of Pineal Research 40, 18–26.
Hovius, R., Thijssen, J., Van Der Linden, P., Nicolay, K., De Kruiff, B.,1993. Phospholipid asymmetry of the outer membrane of rat livermitochondria. Evidence for the presence of cardiolipin on the outsideof the outer membrane. Federation of European Biochemical Societies330, 71–76.
Iver_son, S.L., Orrenius, S., 2004. The cardiolipin–cytochrome c interactionand the mitochondrial regulation of apoptosis. Archivies BiochemistryBiophysics 423, 37–46.
Jacobson, J., Duchen, M.R., Heales, S.J., 2002. Intracellular distributionof the fluorescent dye nonyl acridine orange responds to themitochondrial membrane potential: implications for assays of cardi-olipin and mitochondrial mass. Journal of Neurochemistry 82, 224–233.
Kagan, V.E., Borisenko, G.G., Tyurina, Y., Tyurin, V., Jiang, J.,Potapovich, I., Kini, V., Amoscato, A., Fujii, Y., 2004. Oxidativepipidomics of apoptosis: redox catalytic interactions of cytochrome c
with cardiolipin and phosphatidylserine. Free Radical Biology Med-icine 37, 1963–1985.
Kroemer, G., Reed, J.C., 2000. Mitochondrial control of cell death.Nature Medicine 6, 513–519.
Luchetti, F., Canonico, B., Curci, R., Battistelli, M., Mannello, F., Papa,S., Tarzia, G., Falcieri, E., 2006. Melatonin prevents apoptosis inducedby UV-B treatment in U937 cell line. Journal of Pineal Research 40,158–167.
Mochida, S., Maski, N., Ohta, Y., Ogata, I., Fujiwara, F., 1998. In situdetection of oxidative stress in rat hepatocytes. Journal of Phatology167, 83–89.
Petrosillo, G., Ruggiero, F.M., Paradise, G., 2003. Role of reactive oxygenspecies and cardiolipin in the release of cytochrome c from mitochon-dria. The Federation of American Societies for Experimental Biology17, 2202–2208.
Salvioli, S., Dobrucki, J., Moretti, L., Troiano, L., Fernandez, G.M.,Pinti, M., Pedrazzi, J., Franceschi, C., Cossarizza, A., 2000. Mito-chondrial heterogeneity durino starosporine-induced apoptosis in HL-60 cells: analysis at the single cell and single orfanelle level. Cytometry40, 189–197.
Schagger, H., 2002. Respiratory chain supercomplexes of mitochondriaand bacteria. Biochimica et Biophysica Acta 1555, 154–159.
Schlame, M., Rua, D., Greenberger, M.L., 2000. The biosynthesis andfunctional role of cardiolipin. Progress in Lipid Research 39, 257–288.
Sorice, M., Circella, A., Cristea, I.M., Garofolo, T., Di Renzo, L.,Alessandri, C., Valesini, G., Degli Esposti, M., 2004. Cardiolipin and

F. Luchetti et al. / Toxicology in Vitro 21 (2007) 293–301 301
its metabolites move from mitochondria to other cellular membranesduring death receptor-mediated apoptosis. Cell Death and Differenti-ation 11, 1133–1145.
Zanzamani, N., Kroemer, G., 2003. Apoptosis: mitochondrial membranepermeabilization – the whole story? Current Biology 173, 71–73.
Zorov, D.B., Bannikova, S.Y., Belousov, V.V., Vyssokikh, M.Y., Zorova,L.D., Isaev, N.K., Krasnikov, B.F., Plotnikov, E.Y., 2005. Reactiveoxygen and nitrogen species: friends or foes? Biochemistry 70, 215–221.




















