
Both proteasomes and lysosomes degrade the activated erythropoietin receptor
Upload
independentCategory
view
3download
0
ABSTRACT
Introduction: Erythropoietin (EPO) is a haematopoietic stimulatory protein that is used to treat anaemia in patients on dialysis. In addition, EPO has been shown to have anti-inflammatory properties, which may be important as dialy-sis patients tend to exist within a chronic, low-grade inflammatory state, and tend to be more susceptible to infections. It has been suggested that EPO has direct immunomodulatory potency on monocytes/macrophages. Methods: In this study, we aimed to clarify the effects of EPO during the in-flammatory processes in human mononuclear phagocytic cells by monitoring the secretion of the following cytokines; midkine, tumour necrosis factor alpha (TNF-α), and interleukin-6 (IL-6). For this purpose, U937 human histiocytic lymphoma cell lines were used. Time-dependent effects of varying doses of EPO (0.1 to 50 IU/ml) treatment during lipopolysaccharide (LPS)-mediated cytotoxicity was measured by 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyltetra-zolium bromide test. LPS-stimulated midkine secretion was measured immu-
Erythropoietin Reduces Lipopolysaccharide-Induced Cell Damage and Midkine Secretion in U937 Human Histiocytic Lymphoma Cells Nuray YazihanAnkara University, Faculty of Medicine, Pathophysiology Department, and Molecular Biology Research and Development Unit, Ankara, Turkey
Ozlem KarakurtAnkara University, Molecular Biology Research and Development Unit, and Ankara Ministry of Health Diskapi Education and Research Hospital, Cardiology Department, Ankara, Turkey
Haluk AtaogluAnkara University, Molecular Biology Research and Development Unit, and Microbiology Department, Ankara, Turkey
Address correspondence to: Nuray Yazihan, Ankara University, Faculty of Medicine, Pathophysiology Department, Morfoloji Binasi, 06100, Sihhiye, Ankara, Turkey. Email: [email protected]
Adv Ther. 2008;25(5):502–514.DOI:10.1007/s12325-008-0055-5
502
ORIGINAL RESEARCH

Yazihan et al.
503
INTRODUCTION
Erythropoietin (EPO) is a haematopoi-etic cytokine frequently administered for the treatment of anaemia in dialysis pa-tients. It has a proven anti-apoptotic and antioxidant effect,1 but has also been re-ported to have an anti-inflammatory effect on various pathological conditions2 such as during ischaemia-reperfusion injuries and chronic inflammation conditions like arthritis.3 As dialysis patients have greater risk of sepsis and other infectious diseases, the novel pro-inflammatory effects of EPO during infection and/or inflammation may be life-saving. Many studies have in-vestigated the cytoprotective role of EPO and have led to its use in the treatment of a number of different disease models such as stroke, surgery and diabetes.4–6 However, the potential role of EPO in the regulation of inflammatory cytokines, in particular by macrophages, has yet to be elucidated.
The capacity of cells to secrete cytokines is an important functional determination of immunological status and is critical to the
nohistochemically and quantification of TNF-α, IL-6 and midkine secretion was achieved by ELISA. Results: EPO treatment prevented the direct toxic effects of LPS on the U937 cells. TNF-α, IL-6 and midkine secretions were found to increase in the U937 cells in response to LPS treatment. Interleukin-6 response was varied in a dose- and time-dependent manner. Conclusion: Treatment with EPO significantly inhibits the LPS-induced secre-tion of midkine and TNF-α regardless of the dosage. The data presented here provide the first evidence to indicate that EPO treatment directly reverses some of the cytotoxic and secretory effects of LPS in mononuclear cells. Inhibition of midkine secretion might be responsible for the anti-inflammatory role of EPO.
Keywords: erythropoietin; lipopolysaccharide; midkine; tumour necrosis factor alpha; interleukin-6
host defence. Among the various cytokines produced by monocytes/macrophages, stimulation of acute inflammation using the endotoxin lipopolysaccharide (LPS) has been seen to induce tumour necrosis factor alpha (TNF-α) and interleukin-6 (IL-6) and also to stimulate interferon gamma (IFN-γ) production, which is considered to have an immunomodulatory effect.7 Furthermore, these cytokine secre-tions can be increased when these cells are in a macrophage form. The growth factor mid-kine may also take part in inflammation and survival of various cell types, be involved in the migration of inflammatory leukocytes, and has been observed to be secreted by macrophages.8–11
In this study, we examined the effects of EPO treatment at different doses on cell survival, pro-inflammatory cytokine and midkine secretion in a human histiocytic lymphoma cell line U937 in undifferentiat-ed (monocytes) and phorbol 12-myristate 13-acetate (PMA)-induced differentiated (macrophage) forms under normal and LPS-stimulated infection-like conditions.

Advances in Therapy® Erythropoietin and Pro-inflammatory Cytokines
504
MATERIALS AND METHODS
A human histiocytic lymphoma cell line (U937) was obtained from the American Type Culture Collec-tion (ATCC, Middlesex, UK). The effect of EPO (rHuEPO, Eprex® 4000 IU/ml flacon; Janssen-Cilag, Beerse, Belgium) treatment during LPS (E. Coli O111, Sigma, MO, USA) application was studied in these cells by assessing cellular viabil-ity and inflammatory cytokine se-cretion. Cellular viability was tested using 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) (Sigma). Inflammatory status was deter-mined by measuring TNF-α, IL-6 and midkine levels from cell culture super-natants using ELISA development kits (Cytolab/PeproTech, Rehovot, Israel).
Cell Culture and Experimental Protocol The U937 cell line was grown in RPMI-1640 medium, supplemented with 10% v/v foetal calf serum (PAA, Linz, Austria), 2 mM L-glutamine (Sigma), 100 µg/ml streptomycin (Sigma) and 100 IU/ml penicillin (Sigma), in a hu-midified atmosphere containing 5% CO2 at 37°C.
The cells were differentiated from monocytes into macrophages by using 100 ng/ml PMA-containing medium for 2 days. One day before the experiment commenced, PMA-containing medium was replaced by fresh RPMI-supplemented medium without PMA. Cells were seeded in 24-well plates (NUNC, Roskilde, Den-mark) as 3×105 cells/ml. Depending on the groups, different concentrations of
EPO (0.1, 1, 10, and 50 IU/ml) and/or LPS (1 µg/ml, E. Coli O111) were added to the medium resulting in the following groups: • EPO0.1IU/ml(n=6)• EPO1IU/ml(n=6)• EPO10IU/ml(n=6)• EPO50IU/ml(n=6)• EPO0.1IU/ml+1µg/mlLPS(n=6)• EPO1IU/ml+1µg/mlLPS(n=6)• EPO10IU/ml+1µg/mlLPS(n=6)• EPO50IU/ml+1µg/mlLPS(n=6)• Control(n=6)• LPS(n=6).
The levels of TNF-α, IL-6 and midkine in cell culture supernatants were measured by ELISA at 2, 6, 24 and 48 hours.
Evaluation of Cellular Toxicity
The MTT test, a colourimetric cy-totoxicity assay which is based upon the ability of living cells to reduce MTT into formazan, was used to evaluate the effects of LPS and EPO on cellular proliferation (2, 24 and 48 hours).
Determination of Differentiation of U937 Cells to Macrophages
Surface expression of CD14 has been a proposed marker of cell activation and myelomonocytic cell differentiation.12 Consequently, levels of CD14 expression in the U937 cells, either unstimulated (monocytes) or PMA-stimulated (mac-rophages), were assessed by flow cytom-etry (Beckman-Coulter FC 500, CA, USA). To achieve this cells were incu-bated with phycoerythrin (PE)-labelled CD14 antibody. Labelling was performed

Yazihan et al.
505
at +4°C for 60 minutes. After washingwith phosphate buffered saline (PBS), the cells were suspended in RPMI medium. The CD14-expressing cells were assessed by flow cytometry (Beckman-Coulter FC 500). Results were analysed by displaying the samples’ PE log fluorescence against the background staining of cells with an isotype control antibody (mouse IgG1) and the secondary, PE-conjugated, anti-body (Sigma). Fluorescence was measured at an excitation wavelength of 488 nm and an emission wavelength of 575 nm. To en-sure specificity of the CD14 staining pro-cedures, the binding of each PE-labelled monoclonal antibody was blocked with an excess of unlabelled monoclonal antibody (Sigma). All experiments were performed in triplicate.
For morphologic examination, cells were incubated in 6-cm diameter plastic cell culture petri dishes. Undifferentiated U937 cells and differentiated macrophage cells were examined under phase contrast microscopy (Leica DMR epifluorescence microscope, Germany).
Fluorescent Staining of Permeabilised U937 Cells Treated with LPS
Cells were grown in RPMI 1640 me-dium supplemented with 10% foetal calf serum. They were washed with PBS in ster-ile 0.5 ml eppendorf tubes and incubated with PBS buffered 4% paraformaldehyde (Sigma) at room temperature for 10 min-utes.Afteraspirating,fixingsolutioncellswere rinsed three times with PBS, perme-abilised and blocked, in a solution con-taining 0.05% (v/v) Nonidet P-40, 5% (v/v) goat serum and 1% (w/v) bovine serum albumin prepared in PBS at pH 7.4,
for 15 minutes at room temperature (in NGB buffer). Cells were then incubated with human midkine polyclonal antibody (Cytolab/PeproTech) diluted in NGB buffer at a concentration of 10 μg/ml for 2hoursatroomtemperature.Afterwash-ing the cells with PBS, they were incubated with a FITC-conjugated anti-rabbit goat antibody (Sigma) at a dilution of 1:2000 in NGB for 1 hour at room temperature. Permeabilised cells were examined by Leica DMR epifluorescence microscope.
Measurement of Cytokine Levels
Cell culture supernatants were harvested and analysed for TNF-α, IL-6 and midkine levels in triplicate, using ELISA kits (Peprotech). The lower detection limits of the assays were 16 pg/ml for TNF-α, 32 pg/ml for IL-6 and 300 pg/ml for midkine.
For midkine measurements, mid-kine levels were measured by an ELISA system in which polyclonal anti- human midkine was used as capture an-tibody (Peprotech) and detection was made by biotinylated polyclonal anti-human midkine antibody (Peprotech) followed by streptavidin HRP (Sigma) and TMB enzyme substrate system (Sigma). The reaction was stopped by 1 M H2SO4 and readings were made at 450 nm by a spectrometer (M2, Molecu-lar devices, CA, USA).
Statistical Analysis
Values of LPS and EPO treatments were compared to untreated controls by one-way ANOVA, followed by a mul-tiple comparison test using SPSS 10.0.

Advances in Therapy® Erythropoietin and Pro-inflammatory Cytokines
506
A P value of <0.05 was accepted as statis-tically significant. Data were expressed as mean±standard error of the mean.
RESULTS
Flowcytometric and Morphological Examinations
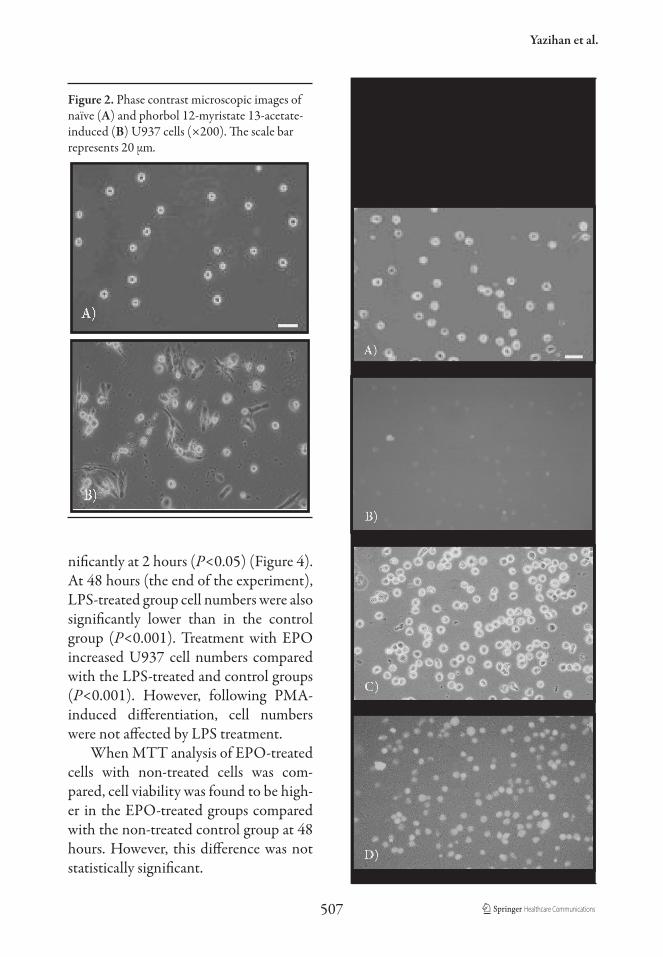
Expression of CD14 on undifferenti-ated U937 cells was low, but was observed to significantly increase following 2 days’ treatment with PMA (Figure 1). Follow-ing PMA treatment, the differentiated cells were spindle-shaped with cellular extensions, had an increased nuclear/
cytoplasmic ratio, paler cytoplasms and more prominent granules. Vacuolations were also present in the cytoplasm of these differentiated cells (Figure 2).
Phase contrast images of un-stimulated and CPS-treated U937 cells are shown in Figures 3A and C, respectively. Figures 3B and D illustrate midkine secretion levels before and after 48 hours of CPS treatment.
Cellular Viability
Compared with the control group, LPS treatment decreased cell number sig-
B)
Figure 1. CD14 expression of undi�erentiated (A) and PMA-induced di�erentiation (B) U937 cells were examined by �ow cytometry. All cells were stained with a PE-conjugated anti-CD14 antibody and analysed by �ow cytometry. FS Lin=forward scatter parameter; PE=phycoerythrin, PMA=phorbol 12-myristate 13-acetate; SS Lin=side scatter parameter.
00
1023
1023
SS Lin1023
FS L
in
00
SS Lin1023
FS L
in
00
205
100 101 102 103
Anti-CD14 PE
00
101
100 101 102 103
Anti-CD14 PE PMA-pulsed
A)
Yazihan et al.
507
nificantly at 2 hours (P<0.05) (Figure 4). At 48 hours (the end of the experiment), LPS-treated group cell numbers were also significantly lower than in the control group (P<0.001). Treatment with EPO increased U937 cell numbers compared with the LPS-treated and control groups (P<0.001). However, following PMA- induced differentiation, cell numbers were not affected by LPS treatment.
When MTT analysis of EPO-treated cells with non-treated cells was com-pared, cell viability was found to be high-er in the EPO-treated groups compared with the non-treated control group at 48 hours. However, this difference was not statistically significant.
Figure 2. Phase contrast microscopic images ofnaïve (A) and phorbol 12-myristate 13-acetate-induced (B) U937 cells (×200). The scale barrepresents 20 µm.
a)
b)
Figure 3. Phase contrast microscopic images of non-treated (A) and lipopolysaccharide (LPS; 1 μg/ml)-treated (C) U937 cells (×200). Fluores-cent staining images with a midkine antibody using FITC-conjugated secondary antibody of permeabi-lised, non-treated (B) and LPS-treated (D) U937 cells after 48 hours. The scale bar represents 20 µm.

Advances in Therapy® Erythropoietin and Pro-inflammatory Cytokines
508
0
150
300
450
100
250
400
50
200
350
Cel
l num
ber/
ml (
x100
0)Figure 4. MTT test results of U937 cells a�er di�erent dosages of EPO and/or LPS treatment. Time-dependent cell number per group shown in the graphic. Results are depicted as mean±SEM. *P<0.05 and **P<0.001 di�erences between LPS-treated and control groups, respectively. EPO=erythropoietin; LPS=lipopolysaccharide; MTT=3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyltetrazolium bromide; SEM=standard error of the mean.
EPO 0.1 IU/m
l
EPO 1 IU/m
l
EPO 10 IU/m
l
EPO 50 IU/m
l
EPO 0.1 IU/m
l+LPS
EPO 1 IU/m
l+LPS
EPO 10 IU/m
l+LPS
EPO 50 IU/m
l+LPS
Control
LPS
***
**
2 hours 24 hours 48 hours
Figure 5. Effects of different doses of EPO and/or LPS on the time-dependent TNF-α secretion in undifferentiated U937 cells. Results are depicted as mean±SEM. *P<0.001 differences betweenLPS-treated and control at 2, 6, 24 and 48 hours, EPO-treated groups. EPO=erythropoietin;LPS=lipopolysaccharide; SEM=standard error of the mean; TNF-α=tumour necrosis factor alpha.
0
30
40
70
80
20
60
10
50
TN
F-α
leve
ls, p
g/m
l/10
0,00
0 ce
lls
EPO 0.1 IU/m
l
EPO 1 IU/m
l
EPO 10 IU/m
l
EPO 50 IU/m
l
EPO 0.1 IU/m
l+LPS
EPO 1 IU/m
l+LPS
EPO 10 IU/m
l+LPS
EPO 50 IU/m
l+LPS
Control
LPS
*
*
*
6 hours 24 hours2 hours 48 hours

Yazihan et al.
509
0
150
300
350
100
250
50
200
TN
F-α
leve
ls, p
g/m
l/10
0,00
0 ce
llsFigure 6. E�ects of di�erent doses of EPO and/or LPS on the time-dependent TNF-α secretionin PMA-induced di�erentiated U937 cells. Data are depicted as mean±SEM. *P<0.001 di�erencesbetween LPS-treated and control, EPO-treated groups. EPO=erythropoietin; LPS=lipopolysaccharide;PMA=phorbol 12-myristate 13-acetate; SEM=standard error of the mean; TNF-α=tumour necrosis factor alpha.
EPO 0.1 IU/m
l
EPO 1 IU/m
l
EPO 10 IU/m
l
EPO 50 IU/m
l
EPO 0.1 IU/m
l+LPS
EPO 1 IU/m
l+LPS
EPO 10 IU/m
l+LPS
EPO 50 IU/m
l+LPS
Control
LPS
*
6 hours 24 hours2 hours 48 hours
Figure 7. Effects of different doses of EPO and/or LPS on the time-dependent IL-6 secretion inundifferentiated U937 cells. Results are depicted as mean±SEM. *P<0.05 differences betweenLPS-treated and EPO-treated groups at 6 hours. EPO=erythropoietin; IL-6=interleukin-6;LPS=lipopolysaccharide; SEM=standard error of the mean.
0
150
200
350
400
100
300
50
250
IL-6
, pg/
ml/
100,
000
cells
EPO 0.1 IU/m
l
EPO 1 IU/m
l
EPO 10 IU/m
l
EPO 50 IU/m
l
EPO 0.1 IU/m
l+LPS
EPO 1 IU/m
l+LPS
EPO 10 IU/m
l+LPS
EPO 50 IU/m
l+LPS
Control
LPS
*
6 hours 24 hours2 hours 48 hours

Advances in Therapy® Erythropoietin and Pro-inflammatory Cytokines
510
Figure 8. Effects of different doses of EPO and/or LPS on the time-dependent IL-6 secretion in PMA-induceddifferentiated U937 cells. Results are depicted as mean±SEM. *P<0.001 differences between LPS-treated and control, EPO-treated groups. EPO=erythropoietin; IL-6=interleukin-6; LPS=lipopolysaccharide; PMA=phorbol 12-myristate 13-acetate.
0
150200
350
450400
100
300
50
250
IL-6
, pg/
ml/
100,
000
cells
EPO 0.1 IU/m
l
EPO 1 IU/m
l
EPO 10 IU/m
l
EPO 50 IU/m
l
EPO 0.1 IU/m
l+LPS
EPO 1 IU/m
l+LPS
EPO 10 IU/m
l+LPS
EPO 50 IU/m
l+LPS
Control
LPS
*
6 hours 24 hours2 hours 48 hours
Table 1. Effects of different doses of EPO and/or LPS on the time-dependent midkine secretion in naïveand PMA-induced U937 cells. Results are depicted as mean±SEM.
Midkine levels, pg/ml
U937 cells PMA-induced U937 cells
Groups 2 hours 24 hours 48 hours 2 hours 24 hours 48 hoursEPO 0.1 IU/ml <300 <300 <300 <300 <300 478±23
EPO 1 IU/ml <300 <300 <300 <300 <300 510±56
EPO 10 IU/ml <300 <300 <300 <300 530±53 545±55
EPO 50 IU/ml <300 <300 <300 <300 565±57 690±68
EPO 0.1 IU/ml+LPS 434±22 <300 <300 <300 421±23 643±34
EPO 1 IU/ml+LPS 514±42 <300 <300 <300 380±38 745±75
EPO 10 IU/ml+LPS 633±62 <300 <300 <300 445±45 435±34
EPO 50 IU/ml+LPS 483±73 <300 <300 <300 510±51 545±55
Control <300 <300 <300 <300 475±48 535±113
LPS 419±61* 777±83* 1084±29* 525±58* 650±69* 1045±85*
*P<0.001 differences between LPS-treated and control, EPO-treated groups. EPO=erythropoietin; LPS=lipopolysaccharide; PMA=phorbol 12-myristate 13-acetate;SEM=standard error of the mean.

Yazihan et al.
511
Cytokine Levels of U937 Cell Cultures Exposed to Different Doses of EPO and/or LPS
The addition of LPS to the culture me-dium resulted in an increase in the secretion of TNF-α, IL-6 and midkine in undifferen-tiated and differentiated cells (Figures 5, 6, 7 and 8 and Table 1).
Specifically, compared with the con-trol group the level of TNF-α secretion was observed to increase between 2 and 24 hours in undifferentiated cells, and in-crease significantly between 2 to 6 hours in differentiated cells (P<0.001). As for IL-6 secretion, following LPS application, levels increased in both cell types. How-ever, in the undifferentiated cells, levels decreased at 6 hours whereas in the differ-entiated PMA-induced cells levels contin-ued to rise until the end of the experiment (48 hours). Midkine levels increased in both undifferentiated and PMA-induced cells at 2 hours after LPS stimulation (Table 1). However, PMA treatment alone did not induce midkine secretion (Figures 5, 6, 7 and 8 and Table 1).
As for the effect of EPO, in both cell types EPO treatment during LPS applica-tion decreased TNF-α secretion compared with LPS-treated alone (P<0.001) (Figures 5 and 6). Treatment with EPO decreased TNF-α secretion in undifferentiated U937 cells (especially at lower doses), but not in PMA-induced differentiated cells. IL-6 secretion from U937 cells was lower at the 10 IU/ml dosage of EPO. During LPS application, IL-6 secretion was significant-ly depressed at higher dosages of EPO in the U937 cell line and PMA-differentiated cells (P<0.05; Figures 7 and 8). Further-more, EPO treatment decreased LPS-
induced midkine secretion significantly in both naïve and PMA-induced U937 cells (Table 1).
DISCUSSION
This present study focused on the ef-fects of EPO on LPS-induced cytokine se-cretion in monocyte/macrophage (U937) cells. Our results suggest that EPO may modify the cytokine response to LPS by decreasing secretions of TNF-α and IL-6. In addition, LPS had a cytotoxic effect on undifferentiated U937 cells, whereas PMA-induced differentiated cells were not affected significantly. Thus, EPO may have a cytoprotective role against LPS toxicity.
Treatment with EPO alone decreased TNF-α secretion compared with the control and LPS-induced groups in a dose-dependent manner. Depending on the cell type and conditions, TNF-α can induce necrotic and apoptotic cell death, with activated monocytes/macrophages being the major sources of endogenous TNF synthesis.13 Hence, EPO may be involved in preventing LPS-induced cell death by decreasing TNF-α secretion. In turn, TNF-α can cause the secretion of various cytokines such as IL-1, IL-6 and IL-8 from monocytes and macrophag-es.14 Although IL-6 is a major product of monocytes/macrophages upon stimula-tion, there is evidence that IL-6 inhibits the production of IL-1ß and TNF-α.7,14 Kaiser et al. found that during cerebral malaria, TNF-α and IFN-γ mRNA over-expression was inhibited by EPO treat-ment.15 These results could help explain the decrease observed in TNF secretions after24hoursinourstudyinbothundif-

Advances in Therapy® Erythropoietin and Pro-inflammatory Cytokines
512
ferentiated and differentiated cells. The increase in IL-6 concentration could provide an autocrine inhibitory effect on TNF secretion. However, this has to be verified.
EPO treatment decreased TNF-α, IL-6 and midkine secretion when com-pared with LPS and control groups. This suggests that EPO has an anti-inflammato-ry effect under normal conditions. Similar results have been reported by Koroglu et al., who observed that LPS induces dam-age in the lymphoid organs of the rat, and that EPO treatment had a protective ef-fect against LPS-induced toxicity in these tissues.16 In our experiment, the addition of EPO to LPS-stimulated cells dramati-cally reduced cytokine levels. However, as time progressed, cytokine levels increased in a manner similar to that of controls. This suggests that when LPS-exposed in-flammatory cells are under the influence of EPO, they can still sustain cytokine secretion at the 24- and 48-hour periods. This is of great importance as it allows the host defence mechanism to react against pathogens if necessary.
LPS induced the secretion of mid-kine, possibly by a process mediated by TNF-α production.17 In a previous study, we observed that the addition of TNF-α (1 ng/ml) to cell culture medium result-ed in midkine secretion from U937 cells (unpublished data). The present study showed that U937 cells have the capa-bility to continuously secrete midkine under basal conditions and to increase secretion in response to LPS stimula-tion. Midkine secretion was observed to be more prominent in PMA-induced cells. Further studies are necessary to elu-cidate the functional aspects of midkine
secretion in these cells. The midkine se-cretory function of U937 cells has been confirmed by an in-vitro cell culture model.11 The fact that midkine secretion can be induced by LPS treatment may be informative for its involvement in the mechanism of inflammatory conditions. However, the limitation of this study is that the effect of midkine treatment was not evaluated in U937 cells. Fur-ther studies are necessary to elucidate the functional aspects of midkine secretion in these cells either by ‘silencing’ midkine or by using overexpression models. Stud-ies with midkine knockout mice have shown the critical involvement of mid-kine in the pathogenesis of interstitial nephritis, cisplatin-induced renal injury and rheumatoid arthritis.9,10,18 The effects of midkine may be critical in the patho-genesis of inflammatory conditions and may directly recruit inflammatory cells. Neutrophil and macrophage infiltration have been observed to be lower in mid-kine knockout mice, and the induction of chemokines such as macrophage inflam-matory proteins (MIP2) and monocyte chemoattractant protein-1 (MCP1) has also been found to be decreased. It has also been shown that midkine plays a role in calcium mobilisation from neutrophils aside from its chemotactic effects.8
In conclusion, EPO appears to have a cytoprotective effect inhibiting LPS- induced cytokine release. The study has also shown that monocytes and mac-rophages have midkine secretory functions at rest and under stimulatory conditions. Furthermore, EPO can inhibit the LPS- induced midkine secretion. Further work is necessary to elucidate the mechanism of interaction of EPO with midkine.

Yazihan et al.
513
ACKNOWLEDGEMENTS
This study was supported by TUBI-TAK project no: SBAG-2812-104S329. The authors thank Dr C Aybay for skilful flowcytometric measurements.
REFERENCES
1. Wiese L, Hempel C, Penkowa M, Kirkby N, Kurtzhals JA. Recombinant human erythropoietin increases survival and reduces neuronal apoptosis in a murine model of cerebral malaria. Malar J. 2008;7:3.
2. Brines M, Cerami A. Discovering erythropoietin’s extra-hematopoietic functions: biology and clinical promise. Kidney Int. 2006;70:246–250.
3. Cuzzocrea S, Mazzon E, di Paola R, et al. Erythropoietin reduces the degree of arthritis caused by type II collagen in the mouse. Biochem Soc Trans. 2005;33:1129–1130.
4. Bernaudin M, Marti HH, Roussel S, et al. Potential role for erythropoietin in focal permanent cerebral ischemia in mice. J Cereb Blood Flow Metab. 1999;19:643–651.
5. Sayan H, Ozacmak VH, Guven A, Aktas RG, Ozacmak ID. Erythropoietin stimulates wound healing and angiogenesis in mice. J Invest Surg. 2006;19:163–173.
6. Lipton SA. Erythropoietin for neurologic protection and diabetic neuropathy. N Engl J Med. 2004;10:2516–2517.
7. Mengozzi M, Sironi M, Gadina M, Ghezzi P. Reversal of defective IL-6 production in lipopolysaccharide-tolerant mice by phorbol myristate acetate. J Immunol. 1991;1:899–902.
8. Takada T, Kinkori T, Muramatsu H, Hayakawa A, Torii S, Muramatsu T. Midkine, a retinoic acid-inducible heparin-binding cytokine, is a novel regulator of intracellular calcium in human neutrophils. Biochem Biophys Res Commun. 1997;29:756–761.
9. Sato W, Kadomatsu K, Yuzawa Y, et al. Midkine is involved in neutrophil infiltration into the tubulointerstitium in ischemic renal injury. J Immunol. 2001;15:3463–3469.
10. Kawai H, Sato W, Yuzawa Y, et al. Lack of the growth factor midkine enhances survival against cisplatin-induced renal damage. Am J Pathol. 2004;165:1603–1612.
11. Inoh K, Muramatsu H, Ochiai K, Torii S, Muramatsu T. Midkine, a heparin-binding cytokine, plays key roles in intraperitoneal adhesions. Biochem Biophys Res Commun. 2004;23:108–113.
12. Ziegler-Heitbrock HW, Ulevitch RJ. CD14: cell surface receptor and differentiation marker. Immunol Today. 1993;14:121–125.
13. MacEwan DJ. TNF receptor subtype signalling: differences and cellular consequences. Cell Signal. 2002;14:477–492.
14. Neta R, Sayers TJ, Oppenheim JJ. Relationship of TNF to interleukins. Immunol Ser. 1992;56:499–566.
15. Kaiser K, Texier A, Ferrandiz J, et al. Recombinant human erythropoietin prevents the death of mice during cerebral malaria. J Infect Dis. 2006;193:987–995.
16. Koroglu TF, Yilmaz O, Ozer E, et al. Erythropoietin attenuates lipopolysaccharide-induced splenic and thymic apoptosis in rats. Physiol Res. 2006;55:309–316.

Advances in Therapy® Erythropoietin and Pro-inflammatory Cytokines
514
17. Ma Y, Temkin V, Liu H, Pope RM. NF-kappaB protects macrophages from lipopolysaccharide-induced cell death: the role of caspase 8 and receptor-interacting protein. J Biol Chem. 2005;280:41827–41834.
18. Maruyama K, Muramatsu H, Ishiguro N, Muramatsu T. Midkine, a heparin-binding growth factor, is fundamentally involved in the pathogenesis of rheumatoid arthritis. Arthritis Rheum. 2004;50:1420–1429.
Copyright © 2022 FDOKUMEN




















