
Role of invariant natural killer T cells in lipopolysaccharide-induced pregnancy loss
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Role of invariant natural killer T cells in lipopolysaccharide-induced pregnancy loss

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Role of invariant natural killer T cells in lipopolysaccharide-inducedpregnancy loss
Liping Li a, Lijuan Shi b, Xinying Yang b, Lihua Ren a, Jing Yang a, Yi Lin c,⇑a Department of Obstetrics and Gynecology, Guangzhou Medical University Affiliated Guangzhou First People’s Hospital, Guangzhou 510180, Chinab Department of Obstetrics and Gynecology, Renji Hospital, School of Medicine, Shanghai Jiaotong University, Shanghai 200001, Chinac Bio-X Institutes, Shanghai Jiao Tong University, Shanghai 200240, China
a r t i c l e i n f o
Article history:Received 30 October 2012Accepted 25 October 2013Available online 6 November 2013
Keywords:InflammationInvariant natural killer T cellsLipopolysaccharidePregnancy lossToll-like receptor
a b s t r a c t
We aimed to investigate the role of invariant natural killer T (iNKT) cells in infection-associated preg-nancy loss. Wild-type (WT) C57BL/6 mice and iNKT cell-deficient Ja18�/� mice were treated with lipo-polysaccharide (LPS). Embryo resorption rates (ERRs), decidual costimulatory molecule and activationmolecule expression, and cytokine production were determined. WT and Ja18�/� mice were adoptivelytransferred with purified iNKT cells. ERRs, decidual costimulatory molecule and activation moleculeexpression, and cytokine production were assessed. LPS-treated Ja18�/� mice showed markedly reducedERRs, decreased CD40, CD80, CD86, and CD69 expression, and reduced Th1 cytokine production at thematernal-fetal interface compared with WT mice. ERRs, expression of CD40, CD80, CD86, and CD69,and Th1 cytokine production in LPS-injected Ja18�/� mice following iNKT cell adoptive transfer wereremarkably upregulated compared with control mice that did not receive adoptively transferred iNKTcells. Our results suggest that iNKT cells play an important role in LPS-induced pregnancy loss.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction
Many factors are thought to be involved in recurrent spontane-ous abortion. Besides chromosomal and structural abnormalities,inflammation processes are an important trigger of miscarriage[1,2]. The overwhelming majority of mid-trimester pregnancy losscases are associated with ascending infection from the lower gen-ital tract [3,4], such as chorioamnionitis is a common cause of sec-ond trimester pregnancy loss [4].
An efficient defense against invading pathogenic microorgan-isms is achieved through coordination of a complex network ofboth innate and acquired immune responses. The first step in theelimination of a pathogenic bacteria or viruses is reliable detection.Toll-like receptors (TLRs) are the major class of signaling receptor,which recognize conserved structures of microbes called patho-gen-associated molecular patterns (PAMPs) in bacteria, viruses,fungi, and parasites [5,6]. TLRs are present not only on uterine leu-kocytes, but also on trophoblast cells, thus implying active cross-talk between the placenta and local immunity [7,8]. TLRs play anessential role in immune recognition, cell signal transduction, up-stream events of immune response cascades, and inflammation-in-duced pregnancy loss [9,10]. Despite a growing associationbetween TLRs and inflammation-induced pregnancy loss, the
precise mechanisms of TLR-mediated inflammation at the mater-nal-fetal interface are largely unknown.
Natural killer T (NKT) cells constitute a highly conserved T lym-phocyte subpopulation that was originally identified as an unusualT cell population co-expressing T cell receptors (TCRs) and somereceptors characteristic of NK cells. Most NKT cells express aninvariant TCR a chain (Va14-Ja18 in mice and Va24-Ja18 in hu-mans), and are referred to as invariant NKT (iNKT) cells or type INKT cells [11]. In contrast with conventional T cells, which recog-nize peptide antigens presented by major histocompatibility com-plex (MHC) class I or II proteins, iNKT cells recognize glycolipidantigens presented by the MHC class I-like antigen presenting mol-ecule, CD1d [12]. The hallmark of iNKT cell activation is the rapidand robust secretion of a variety of cytokines immediately follow-ing TCR engagement [13,14].
iNKT cells directly recognize a-linked glycosphingolipids anddiacylglycerol antigens that are expressed by bacteria suchas Sphingomonas, Ehrlichia, and Borrelia burgdorferi in aCD1d-dependent manner [15–18]. Furthermore, LPS-positivebacteria (such as Salmonella and Escherichia coli) activateTLR4-expressing antigen presenting cells (APCs) through LPS andcan indirectly induce iNKT cell activation through presentation ofself-glycolipids in conjunction with IL-12 co-stimulation, or by anIL-12/IL-18-dependent, CD1d-independent mechanism [19,20].These results suggest that a vast array of microorganisms mightbe able to induce iNKT cell activation indirectly through APC
0008-8749/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.cellimm.2013.10.007
⇑ Corresponding author. Fax: +86 21 50390610.E-mail addresses: [email protected], [email protected] (Y. Lin).
Cellular Immunology 286 (2013) 1–10
Contents lists available at ScienceDirect
Cellular Immunology
journal homepage: www.elsevier .com/locate /yc imm

Author's personal copy
stimulation. Therefore, iNKT cells at the maternal-fetal interfacemay be activated by self-lipid antigens in conjunction with solublefactors from dendritic cells (DCs) upon TLR signaling, and play a rolein inflammation-induced pregnancy loss.
Conventional T cells do not express the invariant Va14-Ja18antigen receptor [21], indicating their selective use in Va14 NKTcells. Thus, disruption of the invariant Va14-Ja18 receptor resultsin the selective loss of Va14 NKT cells, leaving other types of lym-phoid cells, including T, B, and NK cells, intact [22,23]. Ja18�/�
mice, which are fertile and healthy in appearance [24], have nochanges in the total lymphocyte populations compared withwild-type (WT) mice, with the exception of a complete loss ofthe Va14 NKT cell population [22,23,25].
The potential involvement of iNKT cells in the pathogenesis ofinfection-associated pregnancy loss remains to be determined. Inour present study, we used an adoptive transfer system inJa18�/� mice to elucidate the role that iNKT cells may playin LPS-induced pregnancy loss.
2. Materials and methods
2.1. Animals
Eight-week-old female and male WT C57BL/6 (B6) mice andiNKT cell-deficient Ja18�/� mice on a C57BL/6 background werepurchased from The Jackson Laboratory (Bar Harbor, Maine) andsubsequently maintained under pathogen-free conditions in theLaboratory Animal Facility of Guangzhou Medical College. Animalswere acclimated in our facility for at least 2 weeks before use inthese experiments. All animal procedures followed the guidelinesof the Chinese Council for Animal Care. Syngeneic B6 � B6 andJa18�/� � Ja18�/� mating combinations were established. Each fe-male mouse was co-caged with one male. The point at which a vag-inal plug was detected was designated as day 0 gestation.
2.2. LPS-induced abortion model
The average gestation period for mice ranged from 19 to21 days. To induce embryo loss, on day 9 gestation, pregnantJa18�/� mice or B6 mice were injected intraperitoneally (i.p.) with200 ll LPS (Sigma, Saint Louis, MO, USA) saline solution (50 lg/kgbody weight) taking care to ensure that none of the solution en-tered the amniotic cavity. Pregnant WT mice and Ja18�/� mice in-jected i.p. with 200 ll PBS served as negative controls. Ten femalemice were used in each group. Animals were observed frequently(at least four times every day) for any signs of morbidity (piloerec-tion, decreased movement, and vaginal bleeding). Pregnant micewere sacrificed on gestational day 12. At this stage of gestation,feto-placental units undergoing resorption can be clearly distin-guished from their viable counterparts on the basis of size andthe presence of extensive tissue wasting and hemorrhage. Thenumber of implantation sites together with the number of re-sorbed and viable embryos per mouse was counted. The embryoresorption rate (ERR) was calculated as: ERR (%) = number of re-sorbed embryos/number of total embryos � 100 [23,26].
2.3. Isolation of decidual mononuclear cells
For decidual tissue harvest, the uterine horns of pregnant micewere opened longitudinally, and the whole placental and decidualunits were separated individually from the respective embryo andits implantation site. After washing with cold PBS, pooled decidualtissues were cut into small pieces (1 mm3). Selected tissue was di-gested three times with 1 mg/ml Dispase II (Roche, Mannheim,Germany) at 37 �C for 20 min in each cycle in a shaking water bath.
When single or clumped cells were observed under a microscope,the released cells were separated from undigested tissue piecesby filtering through a 50 lm-pore nylon mesh. Mononuclear cellswere purified with Ficoll-Hypaque density medium (density,1.077 ± 0.002 g/ml) by centrifugation at 800 � g for 20 min at22 �C. Any contaminating red blood cells in decidual single-cellsuspensions were eliminated by incubation with red cell lysis buf-fer (GenMed, Arlington, MA, USA) [25].
2.4. Flow cytometry analysis
Cells (106) in 50 ll PBS were incubated with PE/Cy5-conjugatedCD45 (0.25 lg), PE-conjugated CD11c (0.25 lg), FITC-conjugatedCD3 (0.5 lg), allophycocyanin/Cy7-conjugated CD19 (0.5 lg), allo-phycocyanin-conjugated CD49b (0.5 lg), and PE/Cy7-conjugatedCD69 (0.25 lg) for 30 min at 4 �C. In some cases, cells (106) in50 ll PBS were incubated with PE/Cy5-conjugated CD45(0.25 lg), PE-conjugated CD11c (0.25 lg), FITC-conjugated CD40(0.5 lg), allophycocyanin-conjugated CD80 (0.5 lg), and PE/Cy7-conjugated CD86 (0.5 lg) for 30 min at 4 �C. After washing twicewith PBS, cells were fixed in 10 g/l paraformaldehyde. Immuno-stained cells were analyzed on a FACSCanto flow cytometer usingFACSDiva software (BD Biosciences, San Jose, CA, USA). Ten thou-sand cells were detected in each sample. Isotype controls fornon-specific staining were established using matched fluores-cence-labeled isotype control Abs. All of the fluorescence-labeledAbs and isotype controls were purchased from BioLegend (San Die-go, CA, USA). CD45+ cells were gated. The percentages of CD11c+-
CD69+ cells in the CD11c+ cell population, CD3+CD69+ cells in theCD3+ cell population, CD19+CD69+ cells in the CD19+ cell popula-tion, and CD49b+CD69+ cells in the CD49b+ cell population werecalculated. In other cases, the percentages of CD11c+CD40+ cellsin the CD11c+ cell population, CD11c+CD80+ cells in the CD11c+ cellpopulation, and CD11c+CD86+ cells in the CD11c+ cell populationwere calculated in the same way.
2.5. Cytokine assays
Decidual mononuclear cells were cultured in a 5% CO2 atmo-sphere. After 24 h, culture supernatants were collected and storedat �80 �C for cytokine assays. The levels of mouse cytokines,including IFN-c, IL-12p70, IL-4, and IL-10 were assessed usingcommercially available ELISA kits from BD Pharmingen (San Diego,CA, USA). All assays were conducted according to the manufac-turer’s instructions.
For intracellular cytokine analysis, isolated decidual mononu-clear cells (106 cells/ml) from PBS or LPS-treated WT mice wereadded with 50 ng/ml phorbol myristate acetate (PMA) (Sigma,Saint Louis, MO, USA), 500 ng/ml ionomycin (Sigma, Saint Louis,MO, USA) and 5 lg/ml brefeldin A (Sigma, Saint Louis, MO, USA),and cultured at 37 �C in 5% CO2 for 4 h. Stimulated cells were incu-bated with PE-Cy7-conjugated CD11c (0.25 lg), FITC-conjugatedCD3 (0.5 lg), allophycocyanin-Cy7-conjugated CD49b (1.0 lg)and PE-Cy5-conjugated F4/80 (1.0 lg) for 30 min at 4 �C in PBScontaining 2% fetal calf serum and 0.01% NaN3. After washing twicewith PBS, cells were fixed in 10 g/l paraformaldehyde for 10 min.Cells were resuspended in PBS plus 2% fetal calf serum and 0.1%saponin (permeabilization buffer) and incubated with PE-conju-gated anti-IL-12 (1.0 lg) and allophycocyanin-conjugated anti-IFN-c (1.0 lg) in permeabilization buffer. CD11c+, CD3+, CD49b+,and F4/80+ cells were gated and screened for intracellular IL-12and IFN-c production. Isotype controls were established usingmatched fluorescence-labeled isotype control Abs to avoid nonspe-cific staining. All of the fluorescence-labeled Abs and isotype con-trols were purchased from BioLegend (San Diego, CA, USA).
2 L. Li et al. / Cellular Immunology 286 (2013) 1–10

Author's personal copy
2.6. Isolation of spleen iNKT cells
Single-cell suspensions were prepared from spleens using a cellstrainer followed by RBC lysis. iNKT cells were isolated from femaleB6 mouse spleens by labeling with PE-conjugated a-galactosylcera-mide (a-GalCer)-loaded CD1d tetramer. The tetrameric CD1d/a-GalCer complexes bind to the TCR of iNKT cells. iNKT cells weresorted with anti-PE-MicroBeads, according to the manufacturer’sprotocol (Miltenyi Biotec, Teterow, Germany). Because of unspecificbinding of the tetramer to B cells, B cells were depleted prior to iNKTcell enrichment by using B220 MicroBeads. In brief, single-cell sus-pension was prepared. Dead cells were removed by using the DeadCell Removal Kit. The splenic single-cell suspension was passedthrough a 30 lm nylon mesh to remove cell clumps which may clogthe column. The cells were labeled with B220 MicroBeads andpassed through a LD column and an autoMACS™ Separator. Theunlabeled B cell-depleted fraction was collected. FcR Blocking Re-agent was added to prevent Fc receptor-mediated antibody labeling.The B cell-depleted fraction was labeled with PE-conjugated a-Gal-Cer-loaded CD1d tetramer, washed and tetramer+ iNKT cells en-riched by using anti-PE-Microbeads, two MS Columns and anautoMACS™ Separator. The percentage of iNKT cells in the enrichedfraction was determined by labeling with FITC-conjugated CD3e andPE-conjugated a-GalCer/CD1d tetramer. The purity of sorted iNKTcells routinely exceeded 97%, as determined by flow cytometry [27].
2.7. Adoptive transfer of pure iNKT cells
Va14 NKT cells accumulate in the liver, spleen, and bone mar-row and reach steady-state levels by approximately 3 weeks ofage [28]. BrdUrd incorporation studies have shown that peripheralVa14 NKT cells are a slowly dividing population [28]. However, theNKT cell pool is labile because of NKT cell redistribution, differen-tiation, and homeostatic expansion to allow the maintenance ofperipheral NKT cell homeostasis [29]. In this study, we used anadoptive transfer system to assess the role of iNKT cells in infec-tion-associated pregnancy loss. For iNKT cell reconstitution ofJa18�/� mice, 2 � 105 purified iNKT cells or PBS were injectedintravenously (i.v.) into the tail vein of Ja18�/� mice 3 d prior toLPS or PBS injection.
Pregnant mice were sacrificed on d 12 gestation. ERRs were cal-culated, decidual mononuclear cells were isolated and cultured for24 h, and the levels of IFN-c, IL-12p70, IL-4, and IL-10 in the culturesupernatants were assayed using commercially available ELISA kitsfrom BD Pharmingen (San Diego, CA, USA) according to the manu-facturer’s instructions. Costimulatory molecule expression ondecidual DCs and decidual immune cell activation were also exam-ined by flow cytometry.
2.8. Statistical analysis
All statistical analyses were performed using SPSS 19.0 soft-ware. Data were determined by one-way ANOVA with Bonferronitest or independent-samples t test. Ten female mice were used ineach group in induced pregnancy loss experiments. In moleculeand cytokine expression assays, experiments were repeated inde-pendently eight times. Results were presented as means ± SD. A Pvalue of <0.05 was considered to indicate statistical significancebetween analyzed groups.
3. Results
3.1. Comparison of ERRs between WT and Ja18�/� mice
To elucidate the intrinsic role of iNKT cells in LPS-induced preg-nancy loss, WT, and Ja18�/�mice were injected i.p. with LPS or PBS
on d 9 gestation, and the rates of embryo resorption were analyzed.As shown in Table 1, there were no significant differences in themean implantation sites per mouse for WT and Ja18�/� mice trea-ted with PBS or LPS. However, the number of viable embryos permouse for WT mice treated with LPS was lower than that forPBS-treated WT or Ja18�/� mice. Moreover, ERRs in WT mice trea-ted with LPS were significantly higher than in WT or Ja18�/� micetreated with PBS. In contrast, ERRs in Ja18�/�mice treated with LPSwere markedly lower than in WT mice, with no difference ob-served in ERRs in WT or Ja18�/� mice injected with PBS.
3.2. Comparison of CD40, CD80, and CD86 expression by decidual DCs
To explore the mechanism underlying the decreased ERR iniNKT cell-deficient Ja18�/� mice, we examined the expression lev-els of costimulatory molecules CD40, CD80, and CD86 on decidualDCs from WT and Ja18�/�mice. No significant differences were de-tected in expression levels of these markers on decidual DCs inPBS-treated WT and Ja18�/� mice. However, expression levels ofthese markers on decidual DCs from LPS-injected WT mice werenotably higher than those from PBS-injected WT or Ja18�/� mice.The expression levels of CD40, CD80, and CD86 on decidual DCsfrom LPS-treated Ja18�/� mice were remarkably reduced in com-parison with those from LPS-treated WT mice (Fig. 1).
3.3. Comparison of CD69 expression by decidual DCs, T, B, and NK cells
To further examine the possible role of iNKT cells on decidualimmune cell activation, we analyzed expression of the early activa-tion marker CD69 on decidual DCs, T, B, and NK cells identified byexpression of the relevant surface markers CD11c, CD3, CD19, andCD49b, respectively. There were no striking differences in CD69expression by decidual DCs, T cells, and NK cells from PBS-treatedWT mice and Ja18�/�mice. The expression levels of CD69 on decid-ual DCs, T cells, and NK cells were markedly upregulated from LPS-injected WT mice compared with PBS-injected WT mice or Ja18�/�
mice. CD69 expression levels on decidual DCs, T cells, and NK cellswere notably decreased from LPS-treated Ja18�/� mice in compar-ison with LPS-treated WT mice. However, there were no significantdifferences in the expression levels of CD69 on decidual B cellsamong PBS-treated or LPS-treated WT and Ja18�/� mice (Fig. 2).
3.4. Comparison of cytokine levels in the culture supernatants ofdecidual mononuclear cells
To investigate the functions of iNKT cells in inflammation-in-duced embryo resorption, the levels of Th1 and Th2 cytokines inthe culture supernatants of decidual mononuclear cells from WTmice and Ja18�/� mice were measured. There were no significantdifferences in the concentrations of IFN-c, IL-12p70, IL-4, and IL-10 between PBS-treated WT and Ja18�/� mice. However, the con-centrations of the Th1 cytokines IFN-c and IL-12p70 in the culturesupernatants of decidual mononuclear cells from LPS-injected WTmice were apparently upregulated in comparison with those fromPBS-treated WT or Ja18�/� mice. The concentrations of IFN-c andIL-12p70 in the culture supernatants from LPS-treated Ja18�/�
mice were markedly reduced compared with LPS-treated WT mice.In contrast, the levels of IL-4 and IL-10 in the culture supernatantsshowed no marked differences among PBS-treated or LPS-treatedWT and Ja18�/� mice (Fig. 3).
3.5. Comparison of intracellular cytokine levels of decidual lymphocytesubsets
To further explore which decidual lymphocyte subsets in-creased the production of the Th1 cytokines after LPS treatment.
L. Li et al. / Cellular Immunology 286 (2013) 1–10 3

Author's personal copy
Table 1Comparison of embryo resorption rates (ERRs) between wild-type and Ja18�/� mice.
Group Mice Stimulator N Implantation sites per mouse Viable embryos per mouse ERR (%)
A WT PBS 10 8.4 ± 1.1 7.2 ± 1.2 8.4 ± 5.9B Ja18�/� PBS 10 8.6 ± 1.1 7.8 ± 1.4 9.4 ± 5.1C WT LPS 10 8.4 ± 1.3 6.1 ± 1.0a,b 25.8 ± 6.7c,d,e
D Ja18�/� LPS 10 8.2 ± 1.3 7.3 ± 1.3 11.1 ± 6.5
a P < 0.05 versus group A.b P < 0.05 versus group B.c P < 0.01 versus group A.d P < 0.01 versus group B.e P < 0.01 versus group D.
Fig. 1. Comparison of CD40, CD80, and CD86 expression on decidual DCs. Decidual mononuclear cells were incubated with anti-CD45, anti-CD11c, anti-CD40, anti-CD80 andanti-CD86 mAbs and isotype controls as indicated. CD45+ cells were gated and the percentages of decidual CD40+ cells in the CD11c+ cell population (A), CD80+ cells in theCD11c+ cell population (B), and CD86+ cells in the CD11c+ cell population (C) were calculated. Data represent means ± SD (n = 8 per group).
Fig. 2. Comparison of CD69 expression on decidual DC, T, B, and NK cell populations. Decidual CD45+ cells were gated, and the percentages of CD11c+CD69+ cells in theCD11c+ cell population (A), CD3+CD69+ cells in the CD3+ cell population (B), CD19+CD69+ cells in the CD19+ cell population (C), and CD49b+CD69+ cells in the CD49b+ cellpopulation (D) were calculated. Values represent means ± SD (n = 8 per group).
4 L. Li et al. / Cellular Immunology 286 (2013) 1–10
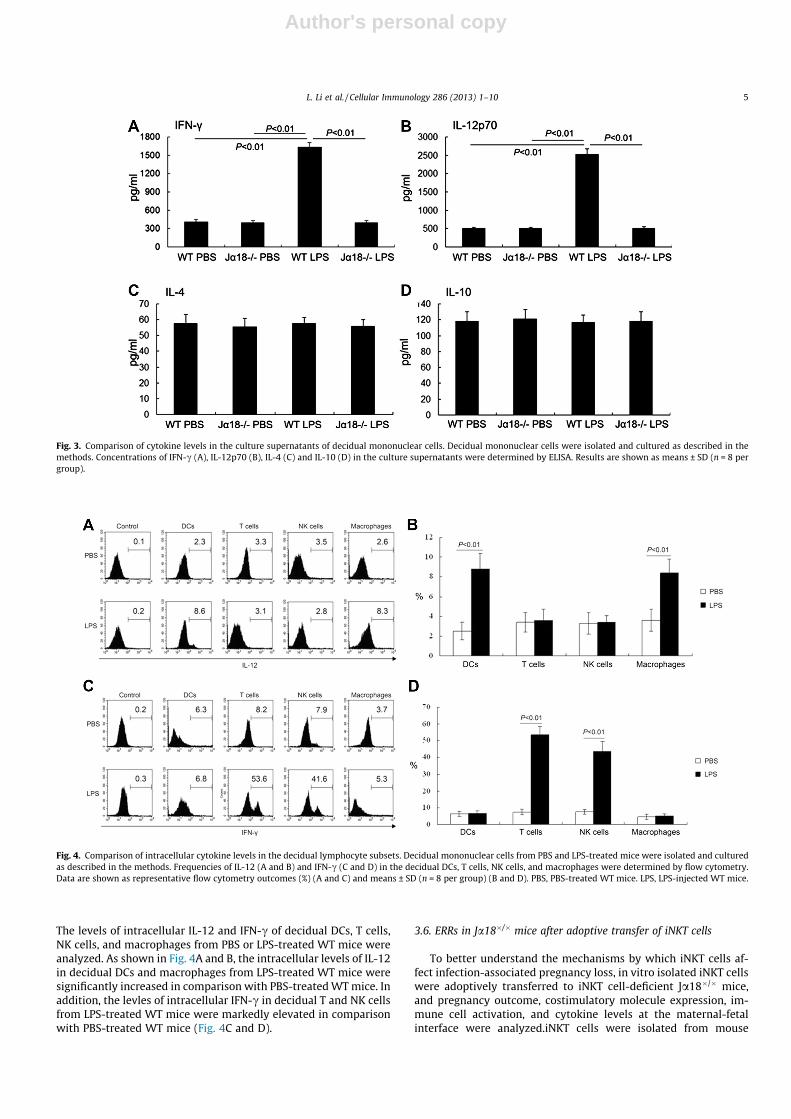
Author's personal copy
The levels of intracellular IL-12 and IFN-c of decidual DCs, T cells,NK cells, and macrophages from PBS or LPS-treated WT mice wereanalyzed. As shown in Fig. 4A and B, the intracellular levels of IL-12in decidual DCs and macrophages from LPS-treated WT mice weresignificantly increased in comparison with PBS-treated WT mice. Inaddition, the levles of intracellular IFN-c in decidual T and NK cellsfrom LPS-treated WT mice were markedly elevated in comparisonwith PBS-treated WT mice (Fig. 4C and D).
3.6. ERRs in Ja18�/� mice after adoptive transfer of iNKT cells
To better understand the mechanisms by which iNKT cells af-fect infection-associated pregnancy loss, in vitro isolated iNKT cellswere adoptively transferred to iNKT cell-deficient Ja18�/� mice,and pregnancy outcome, costimulatory molecule expression, im-mune cell activation, and cytokine levels at the maternal-fetalinterface were analyzed.iNKT cells were isolated from mouse
Fig. 3. Comparison of cytokine levels in the culture supernatants of decidual mononuclear cells. Decidual mononuclear cells were isolated and cultured as described in themethods. Concentrations of IFN-c (A), IL-12p70 (B), IL-4 (C) and IL-10 (D) in the culture supernatants were determined by ELISA. Results are shown as means ± SD (n = 8 pergroup).
Fig. 4. Comparison of intracellular cytokine levels in the decidual lymphocyte subsets. Decidual mononuclear cells from PBS and LPS-treated mice were isolated and culturedas described in the methods. Frequencies of IL-12 (A and B) and IFN-c (C and D) in the decidual DCs, T cells, NK cells, and macrophages were determined by flow cytometry.Data are shown as representative flow cytometry outcomes (%) (A and C) and means ± SD (n = 8 per group) (B and D). PBS, PBS-treated WT mice. LPS, LPS-injected WT mice.
L. Li et al. / Cellular Immunology 286 (2013) 1–10 5

Author's personal copy
spleen using a PE-conjugated CD1d tetramer loaded with a-GalCerand anti-PE Microbeads after depletion of B cells using B220 Micro-Beads. The purity of isolated iNKT cells routinely exceeded 97%, asdetermined by flow cytometry (Fig. 5).
As shown in Table 2, the implantation sites per mouse forJa18�/� mice treated with PBS or LPS were not significantly differ-ent with or without adoptive transfer of iNKT cells. However, the
number of viable embryos per mouse for LPS-treated Ja18�/� micein the presence of adoptively transferred iNKT cell was reduced incomparison with LPS-treated Ja18�/� mice in the absence of theadoptive transfer, and PBS-treated Ja18�/� mice with or withoutiNKT cell adoptive transfer, although this reduction did not reachthe level of statistical significance. Moreover, ERRs for LPS-treatedJa18�/� mice following iNKT cell adoptive transfer significantly in-
Fig. 5. Comparison of the frequencies of iNKT cells with or without purification. Spleen mononuclear cells were incubated with fluorescence-conjugated anti-CD3e, anti-CD1d/a-GalCer, and isotype control Abs. The percentages of CD1d/a-GalCer+ cells in the CD3e+ cell population were calculated. (A) Spleen iNKT cells without purification. (B)Spleen iNKT cells with purification. (C) Double negative control. Data are the mean values of eight mice per group. Numbers in the upper right corner of each group representthe frequencies of iNKT cells.
Table 2Comparison of embryo resorption rates (ERRs) in Ja18�/� mice with or without adoptive transfer of iNKT cells.
Group Mice Adoptive transfer N Implantation sites per mouse Viable embryos per mouse ERR (%)
E Ja18�/� PBS No 10 8.1 ± 1.4 7.2 ± 1.2 11.0 ± 6.5F Ja18�/� PBS Yes 10 8.0 ± 1.2 7.1 ± 0.9 10.8 ± 6.2G Ja18�/� LPS No 10 8.2 ± 1.3 7.2 ± 1.3 11.3 ± 4.5H Ja18�/� LPS Yes 10 8.4 ± 1.4 6.4 ± 0.8 22.8 ± 7.1a,b,c
a P < 0.01 versus group E.b P < 0.01 versus group F.c P < 0.01 versus group G.
Fig. 6. Comparison of CD40, CD80, and CD86 expression on decidual DCs in Ja18�/� mice with or without adoptive transfer of iNKT cells. Ja18�/� mice were transferred withor without purified iNKT cells as indicated in Section 2. Expression of CD40 (A), CD80 (B), and CD86 (C) on decidual CD11c+ cells was calculated. Results are shown asmeans ± SD (n = 8 per group). PBS, PBS-injected Ja18�/� mice without iNKT cell adoptive transfer. PBS + iNKT, PBS-injected Ja18�/� mice with iNKT cell adoptive transfer. LPS,LPS-injected Ja18�/� mice without iNKT cell adoptive transfer. LPS + iNKT, LPS-injected Ja18�/� mice with iNKT cell adoptive transfer.
6 L. Li et al. / Cellular Immunology 286 (2013) 1–10
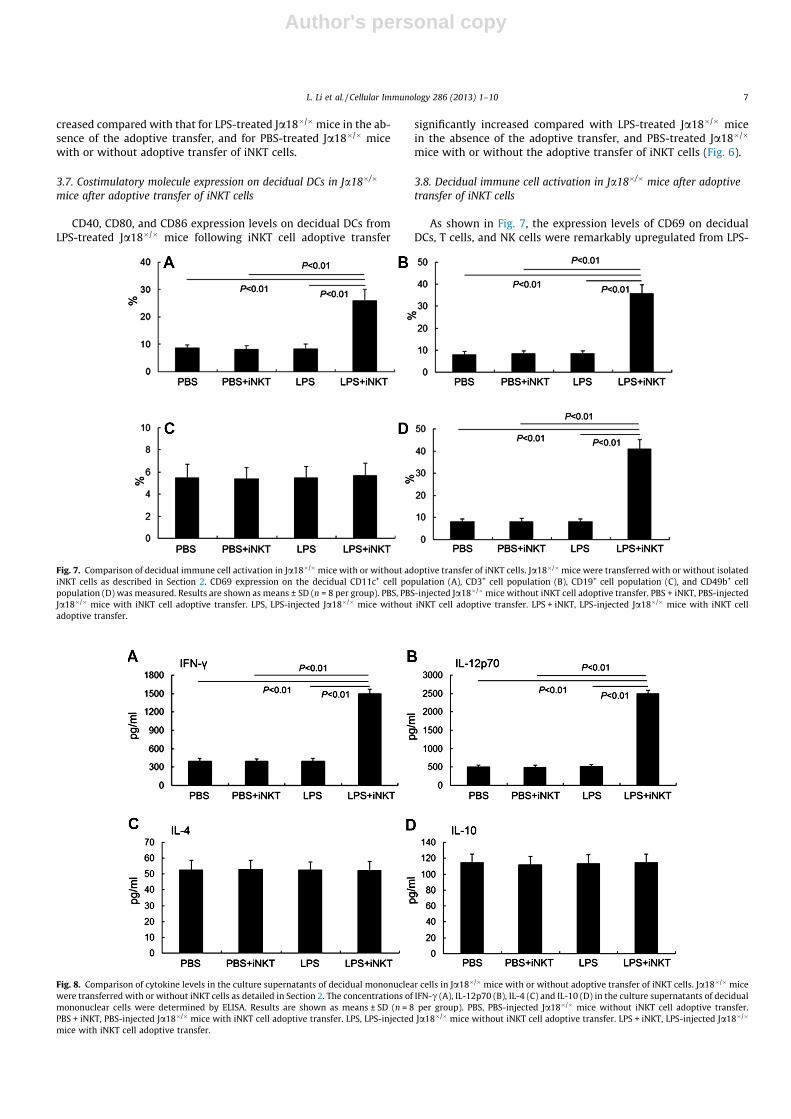
Author's personal copy
creased compared with that for LPS-treated Ja18�/�mice in the ab-sence of the adoptive transfer, and for PBS-treated Ja18�/� micewith or without adoptive transfer of iNKT cells.
3.7. Costimulatory molecule expression on decidual DCs in Ja18�/�
mice after adoptive transfer of iNKT cells
CD40, CD80, and CD86 expression levels on decidual DCs fromLPS-treated Ja18�/� mice following iNKT cell adoptive transfer
significantly increased compared with LPS-treated Ja18�/� micein the absence of the adoptive transfer, and PBS-treated Ja18�/�
mice with or without the adoptive transfer of iNKT cells (Fig. 6).
3.8. Decidual immune cell activation in Ja18�/� mice after adoptivetransfer of iNKT cells
As shown in Fig. 7, the expression levels of CD69 on decidualDCs, T cells, and NK cells were remarkably upregulated from LPS-
Fig. 7. Comparison of decidual immune cell activation in Ja18�/�mice with or without adoptive transfer of iNKT cells. Ja18�/�mice were transferred with or without isolatediNKT cells as described in Section 2. CD69 expression on the decidual CD11c+ cell population (A), CD3+ cell population (B), CD19+ cell population (C), and CD49b+ cellpopulation (D) was measured. Results are shown as means ± SD (n = 8 per group). PBS, PBS-injected Ja18�/�mice without iNKT cell adoptive transfer. PBS + iNKT, PBS-injectedJa18�/� mice with iNKT cell adoptive transfer. LPS, LPS-injected Ja18�/� mice without iNKT cell adoptive transfer. LPS + iNKT, LPS-injected Ja18�/� mice with iNKT celladoptive transfer.
Fig. 8. Comparison of cytokine levels in the culture supernatants of decidual mononuclear cells in Ja18�/� mice with or without adoptive transfer of iNKT cells. Ja18�/� micewere transferred with or without iNKT cells as detailed in Section 2. The concentrations of IFN-c (A), IL-12p70 (B), IL-4 (C) and IL-10 (D) in the culture supernatants of decidualmononuclear cells were determined by ELISA. Results are shown as means ± SD (n = 8 per group). PBS, PBS-injected Ja18�/� mice without iNKT cell adoptive transfer.PBS + iNKT, PBS-injected Ja18�/� mice with iNKT cell adoptive transfer. LPS, LPS-injected Ja18�/� mice without iNKT cell adoptive transfer. LPS + iNKT, LPS-injected Ja18�/�
mice with iNKT cell adoptive transfer.
L. Li et al. / Cellular Immunology 286 (2013) 1–10 7

Author's personal copy
treated Ja18�/� mice following iNKT cell adoptive transfer in com-parison with LPS-treated Ja18�/� mice in the absence of the adop-tive transfer, and PBS-treated Ja18�/� mice with or withoutadoptive transfer of iNKT cells. However, there were no significantdifferences in the expression levels of CD69 on decidual B cellsamong PBS-treated or LPS-treated Ja18�/� mice with or withoutiNKT cell adoptive transfer.
3.9. Cytokine levels of decidual mononuclear cells in Ja18�/�mice afteradoptive transfer of iNKT cells
The concentrations of the Th1 cytokines IFN-c and IL-12p70 inthe culture supernatants of decidual mononuclear cells from LPS-injected Ja18�/� mice following iNKT cell adoptive transfer wereremarkably higher than those from LPS-injected Ja18�/� mice inthe absence of the adoptive transfer, and PBS-injected Ja18�/�
mice with or without iNKT cell adoptive transfer. In contrast, nonotable differences were observed in the concentrations of theTh2 cytokines IL-4 and IL-10 among PBS-treated or LPS-treatedJa18�/� mice with or without iNKT cell adoptive transfer (Fig. 8).
4. Discussion
The presence of iNKT cells in both the mouse and the humandecidua [23,26], and the expression of CD1d on both the villousand extravillous trophblasts [30,31], indicates that iNKT cellshave an immunoregulatory role at the maternal-fetal interface.Moreover, murine iNKT cells accumulate in the decidua duringpregnancy and provoke abortion upon stimulation with a-Gal-Cer, a specific ligand for Va14 NKT cells by perforin-dependentkilling and the secretion of TNF-a and IFN-c [23]. Similarly, ithas been demonstrated that a-GalCer efficiently induces preg-nancy loss during early pregnancy in a perforin-dependent man-ner, and pregnancy loss during mid-gestation by a perforin-independent, cytokine-dependent mechanism [26]. Therefore,we hypothesize that decidual iNKT cells are activated throughindirect recognition of pathogen-related molecules such as bac-terial LPS upon TLR stimulation at the maternal-fetal interface[19,20], subsequently producing massive amounts of cytokinesthat play a role in infection-associated pregnancy loss. In thisstudy, we investigated the effects of iNKT cell deficiency onLPS-induced pregnancy loss.
Ito et al. [23] and Boyson et al. [26] investigated the role of iNKTcells in miscarriage by using a-GalCer, the exogenous marinesponge glycosphingolipid that does not exist in vertebrates. Inour current study, LPS was used to establish a murine model mim-icking infection-associated pregnancy loss. LPS is an importantcomponent of the cell wall of Gram-negative bacteria that are com-monly present in the normal vaginal flora of pregnant women.Pregnant mice were administered LPS (50 lg/kg body weight) ongestational d 9 with the aim of establishing a mid-trimester infec-tion-associated pregnancy loss model [3,4]. This dose of LPS waschosen because it resulted in an elevated rate of embryo resorptioncomparable to that in the CBA/J � DBA/2 abortion-prone mousemodel [32], with no maternal mortality or obvious morbidity. Inthis study, we first showed that the number of viable embryosper mouse was significantly increased and ERRs remarkably de-creased for LPS-treated Ja18�/� mice compared with LPS-treatedWT mice. These data indicate that depletion of iNKT cells downreg-ulated LPS-induced embryo resorption.iNKT cell adoptive transferexperiments were conducted using 2 � 105 cells based on theobservation that approximately 105 NKT cells promote rejectionof OVA-expressing skin grafts [33], and reduce the bacterial burdenin the liver and spleen of the hosts after enteric L. monocytogenesinfection [34]. The iNKT cells were adoptively transferred 3 d prior
to LPS injection based on the observation that adoptive transfer re-sults in homeostatic expansion of Va14+ NKT cells, and approxi-mately 90% of cells divide in the recipient mice by 3 d aftertransfer [29].
Uterine DCs are crucial for decidua formation during embryoimplantation [35]. Nevertheless, these cells possess the capacityto contribute to proinflammatory responses upon pathogenic acti-vation. Driven by pathogens and inflammatory signals, DCs under-go a complex maturation process, which not only leads toenhanced expression of costimulatory molecules and increasedformation of stable MHC/peptide complexes but also to secretionof cytokines that modulate T cell activation and expansion, as wellas synthesis of chemokines and chemokine receptors, and regula-tion of T cell and DC trafficking [36]. In the present study, we dem-onstrated that iNKT cells deficiency reduced the expression of thecostimulatory molecules CD40, CD80, and CD86 by decidual DCsand the number of activated decidual DCs, suggesting that iNKTcells have an impact on infection-associated decidual DC matura-tion and activation.
Productive activation of T cells occurs after concomitantengagement of the TCR with antigen presented on DCs in associa-tion with MHC molecules and the delivery of costimulatory signalsresulting from the interaction of CD80, CD86 with CD28, and CD40with CD40L on the cell surface of T cells [37]. As expected, de-creased expression of the costimulatory molecules CD40, CD80,CD86, and the early activation marker CD69 on decidual DCs wasconsistent with downregulated CD69 expression on decidual Tcells. In our study, the absence of iNKT cells reduced the numberof activated T cells in decidua indicating that iNKT cells play a rolein LPS-stimulated T cell activation.
Accumulating evidence indicates that uterine NK (uNK) cellscontribute to successful implantation and maintenance of preg-nancy [38,39]. However, these cells become cytotoxic in naturein the presence of certain pathogens [40,41]. In our study, the num-ber of activated NK cells in the decidua of LPS-treated Ja18�/� micewas remarkably lower than that in LPS-treated WT mice, demon-strating that iNKT cells have an effect on infection-mediated decid-ual NK cell activation.
Healthy pregnancy is the result of a tightly regulated system ofcrosstalk between the mother and the fetus. Implantation and par-turition are specifically characterized by states of inflammation[42,43]. In contrast, the gestation period, comprising decidualiza-tion, placentation, and fetal development, requires uterine quies-cence guided by high levels of progesterone and the productionof anti-inflammatory cytokines from both maternal and fetal cells[44,45]. Abnormal cytokine production may initiate and intensifythe cascade of inflammatory cytokine production involved in ad-verse pregnancy outcomes. In our experiments, the levels of IL-12 in decidual DCs and macrophages and IFN-c in decidual T andNK cells were significantly increased after LPS treatment, suggest-ing that LPS stimulation inceases IL-12 production in APCs and mayfurther promote T and NK cells IFN-c secretion. The levels of theTh1 cytokines IFN-c and IL-12p70 in the culture supernatants ofdecidual mononuclear cells from LPS-treated Ja18�/� mice wereobviously decreased compared with LPS-treated WT mice, whereasconcentrations of the Th2 cytokines IL-4 and IL-10 were not signif-icantly different. Our results suggest that activation of iNKT cellshelp to shift the Th1/Th2 balance to a Th1 predominant state.These results are consistent with those reported by Ito et al. [23],indicating that the Th1 cytokines IFN-c and TNF-a are essentialfor the Va14 NKT cell-mediated abortion.
In contrast to previous studies [23,26], we used an in vitro iNKTcell adoptive transfer system to further confirm the role of iNKTcells in LPS-induced pregnancy loss. Expression levels of costimu-latory molecules and CD69 and the concentrations of IFN-c andIL-12p70 at the maternal-fetal interface from LPS-injected Ja18�/-
8 L. Li et al. / Cellular Immunology 286 (2013) 1–10

Author's personal copy
� mice following iNKT cell adoptive transfer were notably in-creased in comparison with LPS-injected Ja18�/� mice in the ab-sence of adoptive transfer. These data further support a role foriNKT cells in promoting DC maturation and immune cell activation,and shifting Th1/Th2 balance in pregnancy to a Th1 predominantstate. As expected, the ERRs for LPS-treated Ja18�/� mice followingiNKT cell adoptive transfer were markedly elevated compared withthose without iNKT cell adoptive transfer.
In contrast to previous studies [23,26], we did not explore therole of perforin in iNKT cell-mediated pregnancy loss. While iNKTcells are capable of cytotoxic activity through the expression ofperforin and Fas ligand, their primary effector function is to mod-ulate immune responses by producing large amounts of cytokines[46]. Furthermore, transactivated NK cells following iNKT cellactivation are also likely to mediate direct contact and lysis ofembryonic trophoblasts in a perforin-dependent manner.
Collectively, our results suggest that iNKT cells play a role in LPS-induced pregnancy loss by promoting decidual DC maturation andactivation, decidual T cell and NK cell activation, and proinflamma-tory cytokine production. In future studies we will investigatewhether iNKT cells also play such a role in infection-associatedpregnancy loss in humans.
Acknowledgments
This study was supported by Grants from the National BasicResearch Program of China (2013CB967404), the National NaturalScience Foundation of China (81200478, 31171439), the NationalFunds for Distinguished Young Scientists of China (81125004),Guangdong Medical and Scientific Research Fund (A2012493)and Guangzhou Medical Science and Technology Projects(20121A011020).
References
[1] S.K. Srinivas, L.M. Ernst, A.G. Edlow, M.A. Elovitz, Can placental pathologyexplain second-trimester pregnancy loss and subsequent pregnancyoutcomes?, Am J. Obstet. Gynecol. 199 (2008) 402.e1–402.e5.
[2] S. Haider, M. Knöfler, Human tumour necrosis factor: physiological andpathological roles in placenta and endometrium, Placenta 30 (2009) 111–123.
[3] P. Oakeshott, P. Hay, S. Hay, F. Steinke, E. Rink, S. Kerry, Association betweenbacterial vaginosis or chlamydial infection and miscarriage before 16 weeks’gestation: prospective community based cohort study, Br. Med. J. 325 (2002)1334.
[4] A. Ugwumadu, Chorioamnionitis and mid-trimester pregnancy loss, Gynecol.Obstet. Invest. 70 (2010) 281–285.
[5] M.G. Netea, J.W. Van der Meer, R.P. Sutmuller, G.J. Adema, B.J. Kullberg, Fromthe Th1/Th2 paradigm towards a Toll-like receptor/T-helper bias, Antimicrob.Agents Chemother. 49 (2005) 3991–3996.
[6] K. Takeda, S. Akira, Roles of Toll-like receptors in innate immune responses,Genes Cells 6 (2001) 733–742.
[7] S. Patni, L.P. Wynen, A.L. Seager, G. Morgan, J.O. White, C.A. Thornton,Expression and activity of Toll-like receptors 1–9 in the human termplacenta and changes associated with labor at term, Biol. Reprod. 80 (2009)243–248.
[8] G. Krikun, C.J. Lockwood, V.M. Abrahams, G. Mor, M. Paidas, S. Guller,Expression of Toll-like receptors in the human decidua, Histol. Histopathol.22 (2007) 847–854.
[9] R.L. Goldenberg, C. Thompson, The infectious origins of stillbirth, Am. J. Obstet.Gynecol. 189 (2003) 861–873.
[10] R.L. Gendron, F.P. Nestel, W.S. Lapp, M.G. Baines, Lipopolysaccharide-inducedfetal resorption in mice is associated with the intrauterine production oftumour necrosis factor-alpha, J. Reprod. Fertil. 90 (1990) 395–402.
[11] S. Kim, S. Lalani, V.V. Parekh, L. Wu, L. Van Kaer, Glycolipid ligands of invariantnatural killer T cells as vaccine adjuvants, Expert Rev. Vaccines 7 (2008) 1519–1532.
[12] S.A. Porcelli, R.L. Modlin, The CD1 system: antigen-presenting molecules for Tcell recognition of lipids and glycolipids, Annu. Rev. Immunol. 17 (1999) 297–329.
[13] L. Van Kaer, Regulation of immune responses by CD1d-restricted natural killerT cells, Immunol. Res. 30 (2004) 139–153.
[14] J.M. Coquet, S. Chakravarti, K. Kyparissoudis, F.W. McNab, L.A. Pitt, B.S.McKenzie, S.P. Berzins, M.J. Smyth, D.I. Godfrey, Diverse cytokine production
by NKT cell subsets and identification of an IL-17-producing CD4�NK1.1� NKTcell population, Proc. Natl. Acad. Sci. USA 105 (2008) 11287–11292.
[15] J. Mattner, K.L. Debord, N. Ismail, R.D. Goff, C. Cantu 3rd, D. Zhou, P. Saint-Mezard, V. Wang, Y. Gao, N. Yin, K. Hoebe, O. Schneewind, D. Walker, B.Beutler, L. Teyton, P.B. Savage, A. Bendelac, Exogenous and endogenousglycolipid antigens activate NKT cells during microbial infections, Nature 434(2005) 525–529.
[16] Y. Kinjo, D. Wu, G. Kim, G.W. Xing, M.A. Poles, D.D. Ho, M. Tsuji, K. Kawahara,C.H. Wong, M. Kronenberg, Recognition of bacterial glycosphingolipids bynatural killer T cells, Nature 434 (2005) 520–525.
[17] Y. Kinjo, E. Tupin, D. Wu, M. Fujio, R. Garcia-Navarro, M.R. Benhnia, D.M.Zajonc, G. Ben-Menachem, G.D. Ainge, G.F. Painter, A. Khurana, K. Hoebe, S.M.Behar, B. Beutler, I.A. Wilson, M. Tsuji, T.J. Sellati, C.H. Wong, M. Kronenberg,Natural killer T cells recognize diacylglycerol antigens from pathogenicbacteria, Nat. Immunol. 7 (2006) 978–986.
[18] V. Sriram, W. Du, J. Gervay-Hague, R.R. Brutkiewicz, Cell wallglycosphingolipids of Sphingomonas paucimobilis are CD1d-specific ligandsfor NKT cells, Eur. J. Immunol. 35 (2005) 1692–1701.
[19] N.Y. Crowe, M.J. Smyth, D.I. Godfrey, A critical role for natural killer T cells inimmunosurveillance of methylcholanthrene-induced sarcomas, J. Exp. Med.196 (2002) 119–127.
[20] M. Salio, A.O. Speak, D. Shepherd, P. Polzella, P.A. Illarionov, N. Veerapen, G.S.Besra, F.M. Platt, V. Cerundolo, Modulation of human natural killer T cellligands on TLR-mediated antigen-presenting cell activation, Proc. Natl. Acad.Sci. USA 104 (2007) 20490–20495.
[21] M. Taniguchi, H. Koseki, T. Tokuhisa, K. Masuda, H. Sato, E. Kondo, T. Kawano, J.Cui, A. Perkes, S. Koyasu, Y. Makino, Essential requirement of an invariant Va14T cell antigen receptor expression in the development of natural killer T cells,Proc. Natl. Acad. Sci. USA 93 (1996) 11025–11028.
[22] J. Cui, T. Shin, T. Kawano, H. Sato, E. Kondo, I. Toura, Y. Kaneko, H. Koseki, M.Kanno, Taniguchi, requirement for Va14 NKT cells in IL-12-mediated rejectionof tumors, Science 278 (1997) 1623–1626.
[23] K. Ito, M. Karasawa, T. Kawano, T. Akasaka, H. Koseki, Y. Akutsu, E. Kondo, S.Sekiya, K. Sekikawa, M. Harada, M. Yamashita, T. Nakayama, M. Taniguchi,Involvement of decidual Va14 NKT cells in abortion, Proc. Natl. Acad. Sci. USA97 (2000) 740–744.
[24] L. Rogers, S. Burchat, J. Gage, M. Hasu, M. Thabet, L. Willcox, T.A. Ramsamy, S.C.Whitman, Deficiency of invariant Va14 natural killer T cells decreasesatherosclerosis in LDL receptor null mice, Cardiovasc. Res. 78 (2008) 167–174.
[25] L.P. Li, Y.C. Fang, G.F. Dong, Y. Lin, S. Saito, Depletion of invariant NKT cellsreduces inflammation-induced preterm delivery in mice, J. Immunol. 188(2012) 4681–4689.
[26] J.E. Boyson, N. Nagarkatti, L. Nizam, M.A. Exley, J.L. Strominger, Gestationstage-dependent mechanisms of invariant natural killer T cell-mediatedpregnancy loss, Proc. Natl. Acad. Sci. USA 103 (2006) 4580–4585.
[27] N.A. Nagarajan, M. Kronenberg, Invariant NKT cells amplify the innate immuneresponse to lipopolysaccharide, J. Immunol. 178 (2007) 2706–2713.
[28] K. Benlagha, T. Kyin, A. Beavis, L. Teyton, A. Bendelac, A thymic precursor to theNKT cell lineage, Science 296 (2002) 553–555.
[29] T. Ranson, C.A. Vosshenrich, E. Corcuff, O. Richard, V. Laloux, A. Lehuen, J.P. DiSanto, IL-15 availability conditions homeostasis of peripheral natural killer Tcells, Proc. Natl. Acad. Sci. USA 100 (2003) 2663–2668.
[30] J.E. Boyson, B. Rybalov, L.A. Koopman, M. Exley, S.P. Balk, F.K. Racke, F. Schatz, R.Masch, S.B. Wilson, J.L. Strominger, CD1d and invariant NKT cells at the humanmaternal-fetal interface, Proc. Natl. Acad. Sci. USA 99 (2002) 13741–13746.
[31] J. Matsumoto, K. Kawana, T. Nagamatsu, D.J. Schust, T. Fujii, H. Sato, H. Hyodo,T. Yasugi, S. Kozuma, Y. Taketani, Expression of surface CD1d in theextravillous trophoblast cells of early gestational placenta is downregulatedin a manner dependent on trophoblast differentiation, Biochem. Biophys. Res.Commun. 371 (2008) 236–241.
[32] Y. Lin, Y.Y. Zeng, J.X. Zhao, S. Zeng, J.T. Huang, Z. Feng, J.F. Di, M.Y. Zhan, MurineCD45+CD86+ cells isolated from para-aortic lymph nodes in an abortion-pronemodel, J. Reprod. Immunol. 64 (2004) 133–143.
[33] S.R. Mattarollo, M. Yong, L. Tan, I.H. Frazer, G.R. Leggatt, Secretion of IFN-c butnot IL-17 by CD1d-restricted NKT cells enhances rejection of skin graftsexpressing epithelial cell-derived antigen, J. Immunol. 184 (2010) 5663–5669.
[34] T. Ranson, S. Bregenholt, A. Lehuen, O. Gaillot, M.C. Leite-de-Moraes, A.Herbelin, P. Berche, J.P. Di Santo, Invariant V alpha 14+ NKT cells participate inthe early response to enteric Listeria monocytogenes infection, J. Immunol.175 (2005) 1137–1144.
[35] V. Plaks, T. Birnberg, T. Berkutzki, S. Sela, A. BenYashar, V. Kalchenko, G. Mor, E.Keshet, N. Dekel, M. Neeman, S. Jung, Uterine DCs are crucial for deciduaformation during embryo implantation in mice, J. Clin. Invest. 118 (2008)3954–3965.
[36] H.D. Kopcow, S.A. Karumanchi, Angiogenic factors and natural killer (NK) cellsin the pathogenesis of preeclampsia, J. Reprod. Immunol. 76 (2007) 23–29.
[37] W.W. Hancock, M.H. Sayegh, X.G. Zheng, R. Peach, P.S. Linsley, L.A. Turka,Costimulatory function and expression of CD40 ligand, CD80, and CD86 invascularized murine cardiac allograft rejection, Proc. Natl. Acad. Sci. USA 93(1996) 13967–13972.
[38] V. Male, A. Trundley, L. Gardner, J. Northfield, C. Chang, R. Apps, A. Moffett,Natural killer cells in human pregnancy, Methods Mol. Biol. 612 (2010) 447–463.
[39] A. Moffett, S.E. Hiby, How does the maternal immune system contribute to thedevelopment of pre-eclampsia?, Placenta 28 (2007) S51–S56
L. Li et al. / Cellular Immunology 286 (2013) 1–10 9

Author's personal copy
[40] L. Li, J. Kang, W. Lei, Role of Toll-like receptor 4 in inflammation-inducedpreterm delivery, Mol. Hum. Reprod. 16 (2010) 267–272.
[41] S.P. Murphy, N.N. Hanna, L.D. Fast, S.K. Shaw, G. Berg, J.F. Padbury, R. Romero,S. Sharma, Evidence for participation of uterine natural killer cells in themechanisms responsible for spontaneous preterm labor and delivery, Am. J.Obstet. Gynecol. 200 (2008) 308.e1–308.e9.
[42] M.S.M. van Mourik, N.S. Macklon, C.J. Heijnen, Embryonic implantation:cytokines, adhesion molecules, and immune cells in establishing animplantation environment, J. Leukoc. Biol. 85 (2009) 4–19.
[43] C.R. Mendelson, Minireview: fetal-maternal hormonal signaling in pregnancyand labor, Mol. Endocrinol. 23 (2009) 947–954.
[44] R.W. Kelly, A.E. King, H.O.D. Crithley, Cytokine control in human endometrium,Reproduction 121 (2001) 3–19.
[45] J.R. Challis, C.J. Lockwood, L. Myatt, J.E. Norman, J.F. Strauss 3rd, F. Petraglia,Inflammation and pregnancy, Reprod. Sci. 16 (2009) 206–215.
[46] D.I. Godfrey, M. Kronenberg, Going both ways: immune regulation via CD1d-dependent NKT cells, J. Clin. Invest. 114 (2004) 1379–1388.
10 L. Li et al. / Cellular Immunology 286 (2013) 1–10




















