When nouns and verbs degrade: Facilitating communication in semantic dementia

September 9, 2004 originally published onlinedoi:10.1182/blood-2004-03-1216
2005 105: 600-608
MayeuxPierre Walrafen, Frédérique Verdier, Zahra Kadri, Stany Chrétien, Catherine Lacombe and Patrick receptorBoth proteasomes and lysosomes degrade the activated erythropoietin
http://bloodjournal.hematologylibrary.org/content/105/2/600.full.htmlUpdated information and services can be found at:
(1930 articles)Signal Transduction (3208 articles)Hematopoiesis and Stem Cells
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom

HEMATOPOIESIS
Both proteasomes and lysosomes degrade the activated erythropoietin receptorPierre Walrafen, Frederique Verdier, Zahra Kadri, Stany Chretien, Catherine Lacombe, and Patrick Mayeux
Activation of the erythropoietin receptor(EpoR) after Epo binding is very transientbecause of the rapid activation of strongdown-regulation mechanisms that quicklydecrease Epo sensitivity of the cells.Among these down-regulation mecha-nisms, receptor internalization and degra-dation are probably the most efficient.Here, we show that the Epo receptor wasrapidly ubiquitinated after ligand stimula-tion and that the C-terminal part of theEpo receptor was degraded by the protea-somes. Both ubiquitination and receptor
degradation by the proteasomes occurredat the cell surface and required Januskinase 2 (Jak2) activation. Moreover, Epo-EpoR complexes were rapidly internal-ized and targeted to the lysosomes fordegradation. Neither Jak2 nor protea-some activities were required for internal-ization. In contrast, Jak2 activation wasnecessary for lysosome targeting of theEpo-EpoR complexes. Blocking Jak2 withthe tyrphostin AG490 led to some recy-cling of internalized Epo-Epo receptorcomplexes to the cell surface. Thus, acti-
vated Epo receptors appear to be quicklydegraded after ubiquitination by 2 proteo-lytic systems that proceed successively:the proteasomes remove part of the intra-cellular domain at the cell surface, andthe lysosomes degrade the remaining partof the receptor-hormone complex. Theefficiency of these processes probablyexplains the short duration of intracellu-lar signaling activated by Epo. (Blood.2005;105:600-608)
© 2005 by The American Society of Hematology
Introduction
The kidney-produced hormone erythropoietin (Epo) is the majorregulator of red blood cell production. Epo inhibits apoptosis of thelate erythroid progenitors and stimulates their proliferation, allow-ing the completion of their differentiation program.1 The Eporeceptor (EpoR) is a type 1 transmembrane protein that belongs tothe cytokine receptor family. It is synthesized as a 62 kDa precursorthat is quickly modified by a high-mannose glycosylation thatincreases its molecular mass to 64 kDa. The mature EpoR exhibitsa 66 kDa molecular mass and a complex endoglycosidase H–resistantglycosylation pattern.2,3 The cell surface expression of the EpoRappears to be tightly controlled by mechanisms that are only partlyunderstood. The EpoR undergoes dimerization and associationwith the Jak2 effector kinase during its maturation process, andJanus kinase 2 (Jak2) association with the EpoR is required forEpoR maturation and cell surface expression.4 Epo binding inducesa conformational change in the receptor complex that activates theassociated Jak2 kinases, leading to the initiation of intracellularsignaling. Several intracellular relays are then activated, includingsignal transducer and activator of transcription 5 (STAT5), Ras/mitogen-associated protein (Ras/MAP) kinase, and phosphatidyl-inositol-3 (PI3) kinase/Akt pathways (reviewed by Constantinescuet al5). All Epo-dependent responses including proliferation andsurvival are abrogated when Jak2 activation is disrupted. Simulta-neously, mechanisms of down-regulation are turned on, and theylead to the rapid decrease of Epo responsiveness of the target cells.These mechanisms involve the recruitment of the SHP-1 (domain-containing protein-tyrosine phosphatase-1) phosphatase to the
receptor6 and to the Jak2 kinase,7 the production and receptorassociation of inhibitor proteins of the CIS (Cytokine InducibleSH2-containing protein)/suppressor of cytokine signaling (CIS/SOCS) family, 8-10 and the internalization of the receptors.11
Moreover, we have shown that the maturation and cell surfaceexpression of newly synthesized EpoRs are blocked by a mecha-nism involving the proteasomes during Epo stimulation.12
EpoR and the thrombopoietin (TPO) receptor (Mpl) exhibitstrong functional similarities. They are homodimeric cytokinereceptors that activate Jak2 to initiate intracellular signaling, andtheir main target cells belong to close differentiation lineages. Incontrast to Epo (ErythroPOietin), TPO (ThromboPOietin) induceslong-lasting signaling in target cells, and the duration of this signalis required for megakaryocytic differentiation.13 It has been re-cently shown that after TPO binding, 65% to 75% of the TPO-Mplcomplexes are internalized and that at least some of the internalizedreceptors rapidly recycle to the cell surface.14 Like TPO-Mplcomplexes, Epo-EpoR complexes are rapidly internalized, and thisprocess is not dependent on Jak2.11 However, the fate of theinternalized Epo-EpoR complexes has not been examined.
The internalization and degradation of ligand-stimulated cyto-kine receptors have been extensively studied for the growthhormone (GH) receptor and the interleukin-2 (IL-2) receptor. AsMpl and the EpoR, the former is a receptor homodimer associatedwith the tyrosine kinase Jak2 and the latter is composed of at least 5proteins, including 2 tyrosine kinases of the Jak family (Jak1 andJak3) and 3 bona fide receptor chains (IL-2 receptor � [IL-2R�],
From the Departement d’Hematologie, Institut Cochin, Institut National de la Sante etde la Recherche Medicale U567 (Paris, France), Centre National de la RechercheScientifique UMR 8104, Universite Rene Descartes; and Laboratoired’Hematologie, Assistance Publique—Hopitaux de Paris, Hopital Cochin, Paris,France.
Submitted April 1, 2004; accepted August 26, 2004. Prepublished online as BloodFirst Edition Paper, September 9, 2004; DOI 10.1182/blood-2004-03-1216.
Supported by the Comite de Paris of the Ligue Nationale Contre le Cancer(LNCC; laboratoire associe no. 8) and the Fondation pour la Recherche
Medicale (F.V.).
P.W. and F.V. contributed equally to this study.
Reprints: Patrick Mayeux, Departement d’Hematologie, Institut Cochin, 27,rue du Faubourg Saint-Jacques F-75014, Paris, France; e-mail: [email protected].
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. section 1734.
© 2005 by The American Society of Hematology
600 BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom

IL-2R�, and IL-2R�). In both cases, endocytosis of the receptors isconstitutive although their cognate ligands increase to some extentthe rate of receptor disappearance.15,16 Endocytosis of the GHreceptor occurs through clathrin-coated pits and depends on anactive ubiquitination system and proteasome activity.17 The internal-ized receptor is then routed to the lysosome and degraded togetherwith the hormone. This step also requires the activity of theubiquitin/proteasome pathway.18 In contrast, internalization of theIL-2 receptor complex mainly occurs through clathrin-independentmechanisms involving detergent-resistant membrane domains.19
The subunits of the IL-2 receptor are then differentially sorted: the� chain recycles back to the plasma membrane, while the � and� chains are sorted through the degradation pathway to thelysosomes.20 IL-2 induces monoubiquitination21 or polyubiquitina-tion22 of the � chain of its receptor. Ubiquitin is clearly involved inIL-2 receptor down-regulation although the steps controlled by theubiquitin/proteasome system remain controversial. According toYu and Malek,23 proteasome inhibitors impair internalization ofIL-2–IL-2 receptor complexes. In contrast, Rocca et al havereported that the ubiquitin/proteasome system is not required forinternalization of the IL-2 receptor although the � chain of the IL-2receptor is monoubiquitinated and ubiquitination is necessary for� routing to the lysosomes.21
In this study, we investigate the mechanisms of Epo and EpoRinternalization and degradation. We show that Epo strongly in-creases the rate of receptor internalization and degradation thatinvolves both proteasomes and lysosomes. Lysosome targeting ofEpo and EpoR requires Jak2 activity and not proteasome activity.Moreover, Jak2 activation induces polyubiquitination of the EpoRs,leading to the degradation at the cell surface of the C-terminal partof the intracellular domain of the receptor by proteasomes.
Materials and methods
Reagents and cells
Highly purified recombinant human Epo (specific activity 120 000 U/mg)was a generous gift from Dr M. Brandt (Roche Laboratories, Basel,Switzerland). The protease inhibitors N-Ac-Leu-Leu-norLeucinal (LLnL)and lactacystin were obtained from Calbiochem (Merck Biosciences,Darmstadt, Germany). Other inhibitors were purchased from Sigma (StLouis, MO). Anti-EpoR antibodies used in immunoprecipitation experi-ments (C-236) were produced by immunizing rabbits with a recombinantprotein composed of the full intracellular domain of the human EpoR fusedto glutathione S-transferase (GST). Experiments using various EpoRmutants have shown that these antibodies recognize epitopes included in thefirst 60 amino acids of the intracellular domain of the EpoR, because theyimmunoprecipitate full-length EpoRs and EpoRs truncated after amino acid310. Anti-GST control antibodies were prepared using the same protocol.Anti-EpoR antibodies from Santa Cruz Biotechnology (catalog no. SC-695)(Santa Cruz, CA) were used for immunoblot analysis. These antibodies(C-20) recognize the last 20 amino acids of the EpoR. Antiubiquitinantibodies (catalog no. PW 8810) and anti-Jak2 antibodies (catalog no.SC-278) were from Affiniti Research Products (Exeter, United Kingdom)and Santa Cruz Biotechnology, respectively. Bissulfosuccinimidyl suberate(BS3) was from Pierce (Rockford, IL). Deglycosylation enzymes andproteinase K were from Roche (Mannheim, Germany). Human UT-7 cells24
were cultivated in � minimum essential medium (�MEM) containing 10%fetal calf serum complemented with 2 U/mL Epo. Before each experiment,the cells were growth factor deprived by overnight incubation in IscoveDulbecco MEM containing 0.1% bovine serum albumin treated with AG501-X8 ion exchange resin Bio-Rad (Hercules, CA) 25 �g/mL iron-loadedhuman transferrin.
Whole cell extracts, immunoprecipitation, and Western blotting
Whole cell extracts, immunoprecipitations, and Western blots were per-formed as previously described.25 Extracts from 5 � 105 cells were used ineach assay for the Western blot experiments. Western blots were quantifiedusing the ImageJ 1.32 software after densitometric scanning of the films(http://rsb.info.nih.gov/ij/).
Epo labeling, Epo binding, and Epo internalization studies
Epo labeling using IODO-GEN (Pierce, Rockford, IL) and Epo bindingwere done as previously described.26,27 A saturating 125I-Epo concentration(1 nM; 2 U/mL) was used for these experiments to suppress the effects ofputative modifications of EpoR affinity induced by the different inhibitorsused. Nonspecific binding, determined using a 250-fold excess of unlabeledEpo, was less than 5% in each case. All reported data represent specificbinding. In most experiments, we performed an acidic wash to separate cellsurface–bound and internalized Epo. After incubation with 125I-Epo, thecells were washed twice at 4°C to remove unbound ligand. They wereincubated in 0.5 mL acidic buffer (NaCl 150 mM, sodium acetate 50 mM,pH 3.5) for 3 minutes at 4°C. The pH was then adjusted to 7.4 using 1 MTris (tris(hydroxymethyl)aminomethane)–HCl, pH 9, and the cell suspen-sion was centrifuged. The radioactivities of the supernatant (cell surface–bound Epo) and of the cell pellet (internalized Epo) were determined. When125I-Epo was bound to the cells at 4°C to inhibit Epo internalization, morethan 95% of cell-bound 125I-Epo was recovered in the acidic washsupernatant using this method. Each experiment was performed at least 3 timeswith similar results; 2 � 106 cells were used in each assay for the 125I-Epo bindingexperiments.
Cross-linking studies
To cross-link 125I-Epo to internalized receptors, the cells were incubatedwith 125I-Epo, washed to remove unbound Epo, and solubilized with 25 mMHEPES (N-2-hydroxyethylpiperazine-N�-2-ethanesulfonic acid), 150 mMNaCl, 5 mM EGTA (ethyleneglycoltetraacetic acid), and 1% Nonidet P-40,pH 8.00. The lysate was cleared by centrifugation and incubated for 20minutes in ice with 2 mM BS3. The water-soluble cross-linker BS3 wasused to achieve the high concentration of cross-linker required for theseexperiments. The reaction was stopped by adding ethanolamine, pH 8.00, toa final concentration of 0.1 M, and the EpoR complexes were immunopre-cipitated using C-236 anti-EpoR antibodies. Immunoprecipitated materialwas denatured by boiling in 1% sodium dodecyl sulfate (SDS) and 50 mMdithiothreitol (DTT). DTT was removed by chromatography through G25Sephadex spun columns. Recovered material was then diluted 10-fold withsolubilization buffer and subjected to another immunoprecipitation usingC-236 anti-EpoR antibodies. Lastly, nonprecipitated 125I-Epo–cross-linkedproteins were recovered using anti-Epo antibodies; 50 � 106 to 100 � 106
cells were used in each assay for the cross-linking experiments.
Results
Epo induces the rapid degradation of its receptor
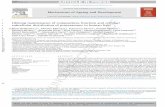
The half-life of the cell surface EpoR in the absence of Epo wasdetermined by blocking protein synthesis with cycloheximide andmeasuring cell surface EpoRs at several time points thereafter byshort incubations with 125I-Epo. Disappearance of EpoR from thecell surface in unstimulated cells followed a first-order kineticsshowing an EpoR half-life of 3 hours � 14 minutes (mean � SDof 3 independent experiments). In contrast, Epo stimulation ledto the disappearance of more than 75% of cell surface EpoRswithin 30 minutes, with kinetics that cannot be fitted by a simplefirst-order mechanism (Figure 1A). During Epo stimulation,125I-Epo transiently appeared inside the cell, with a maximum internal-ization level after 20 minutes of incubation (Figure 1B). The rates ofdisappearance from the cell surface and intracellular accumulation of
EPO RECEPTOR DOWN-REGULATION 601BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom
Epo reflect the combination of internalization and recycling. Monensinhas been shown to allow receptor internalization but to block the trafficof intracellular vesicles, leading to the inhibition of both routing to thelysosomes and recycling to the plasma membrane.28 The initial rate ofEpo accumulation inside the cell was not increased in monensin-treatedcells, indicating that few, if any, Epo-EpoR complexes recycled back tothe cell surface after internalization. After 30 minutes of incubation,intracellular radioactivity decreased in control cells, suggesting that Epowas degraded. In contrast, radioactivity did not decrease in monensin-treated cells, suggesting that Epo degradation could occur in lysosomes.
These results were confirmed by Western blot experimentsusing an antibody (C-20) that recognizes the last 20 amino acids ofthe intracellular domain of the EpoR. This antibody revealed 2forms of EpoR in UT-7 cell extracts (bands 1 and 2 in Figure 2A).In Epo-starved cells, band 1 was most generally detected as adoublet with an apparent molecular mass of 66 kDa. After Epostimulation, most of the lower band of the doublet shifted to theupper band (Figures 2C, 3B, and 6B), most likely because of the
high level of EpoR phosphorylation induced by ligand binding.30
Band 1 disappeared after endoglycosidase F treatment of immuno-precipitated EpoRs but not after digestion by endoglycosidase H(Endo H) (Figure 2A) and was sensitive to proteinase K treatmentof intact cells (Figure 2A). These results show that band 1corresponds to the mature, cell surface form of the EpoR. Band 2(around 64 kDa) was sensitive to Endo H treatment and resistant toproteinase K treatment of intact cells. It represents the maturing,endoplasmic reticulum form of the EpoR. Band 3 (around 62 kDa)was only detected after endoglycosidase treatment and probablycorresponds to the unglycosylated form of the EpoR. In restingUT-7 cells, the half-life of the maturing form (band 2) was 15minutes and the half-life of the mature form (band 1) was around 3hours (Figure 2B). This latter value is in perfect agreement withthat obtained from the experiments depicted in Figure 1A. InEpo-stimulated cells, the half-life of the maturing form did notsignificantly change (Figure 2C), whereas that of the mature formstrongly decreased, confirming the results obtained in 125I-Epobinding experiments. Thus, most of the mature EpoRs are quicklydegraded during Epo stimulation. Cell surface expression of theEpoR has been shown to be controlled by Jak2 association.4 Theintracellular pool of Jak2 was not significantly modified bycycloheximide during the incubation in the presence or absence ofEpo, showing that the half-life of Jak2 was much longer than that ofthe EpoR and that disappearance of the mature form of the EpoRwas not due to the lack of Jak2.
Epo and EpoR degradation
Figure 1B shows that 125I-Epo was quickly internalized after EpoRbinding. However, Epo dissociation from its receptor is also veryrapid at 37°C. In a 2-step protocol in which Epo is bound at 4°C andinternalization triggered by a shift to 37°C after removal ofunbound ligand, most 125I-Epo dissociates from the cell surfacebefore internalization (data not shown). Thus, to test for Epodegradation after internalization, UT-7 cells were first incubated for30 minutes at 37°C with iodinated Epo to accumulate radiolabeledhormone inside the cells and washed to remove free Epo. The cellswere resuspended in fresh medium, and the appearance of Epodegradation products in the culture medium was tested by trichloro-acetic acid (TCA) precipitation. As shown in Figure 3A, mostinternalized 125I-Epo was degraded after 1 hour of incubation in
Figure 1. Internalization and degradation kinetics of the cell surface EpoRs.(A) Degradation kinetics of the cell surface EpoRs in resting (f) and Epo-stimulatedcells (Œ). Resting UT-7 cells were incubated with 500 �M cycloheximide (CHX) for theindicated times, and cell surface EpoRs were quantified by a 10-minute incubationwith 1 nM 125I-Epo. Quantification of cell surface EpoRs during Epo stimulation wasperformed by preincubating resting UT-7 cells for 15 minutes with cycloheximidebefore Epo stimulation. At the indicated times, cells were sampled for the determina-tion of cell surface–associated radioactivity as described in “Materials and methods.”(B) Kinetics of 125I-Epo internalization. Resting UT-7 cells were preincubated for 15minutes with 100 �M monensin (closed symbols) or with solvent alone (opensymbols) before stimulation with 125I-Epo. At the indicated times, cells were sampledfor the determination of cell surface–associated radioactivity (squares, thin lines) andinternalized radioactivity (triangles, dashed lines). Total cell-associated radioactivityis also presented on the graph (circles, thick lines).
Figure 2. Degradation kinetics of the EpoR in Epo-starved cells and Epo-stimulated cells. (A) Characterization of the EpoR forms. (i) EpoR immunoprecipitates fromEpo-starved UT-7 cells were incubated for 18 hours at 37°C with endoglycosidase H (Endo H), endoglycosidase F (Endo F), or deglycosylation buffer alone (control) as previouslydescribed.29 (ii) Whole UT-7 cells were incubated for 45 minutes at 4°C without or with 100 �g/mLproteinase K (Prot K), then were washed, and solubilized in electrophoresis sample buffer.Proteins were then analyzed by Western blot using C-20 anti-EpoR antibodies. (B) Stability of the EpoR proteins in Epo-starved cells. UT-7 cells were incubated withcycloheximide to block protein synthesis. Cells were sampled at the indicated times, and whole cell extracts were analyzed by Western blot using C-20 anti-EpoR or anti-Jak2antibodies. (C) Stability of the EpoR proteins in Epo-stimulated cells. UT-7 cells were preincubated with cycloheximide for 15 minutes. Cells were then stimulated with 10 U/mLEpo for the indicated times, and whole cell extracts were analyzed by Western blot using C-20 anti-EpoR (top) or anti-Jak2 (bottom) antibodies. The number 1 indicates matureform of the EpoR; IgG, the heavy chains of the immunoprecipitating antibodies; 2, maturing form of the EpoR; 3, deglycosylated EpoR; and arrowhead, nonspecific bands.
602 WALRAFEN et al BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom

control cells. We then tested the effect of methylamine, an inhibitorof lysosomal degradation,31 and lactacystin, a specific proteasomeinhibitor. None of these inhibitors significantly modified 125I-Epointernalization after 30 minutes of Epo stimulation (legend toFigure 3A). 125I-Epo degradation was inhibited by methylamine butnot by lactacystin. Therefore, we conclude that internalized Epowas degraded by the lysosomes and that proteasomes are notinvolved in Epo degradation.
In striking contrast with Epo, the EpoR was only partiallyprotected by methylamine, as assessed by Western blot experi-ments using the C-20 antibody (Figure 3B-C). Moreover, as wereported previously,12 blocking proteasome activity with lactacys-tin only slightly protected EpoR from degradation. A stronginhibition of EpoR degradation was observed only when bothlysosome and proteasome activities were blocked by a combinationof methylamine and lactacystin or by LLnL, which inhibitsproteasome and lysosome proteases such as cathepsin B. Theseresults suggested that EpoR could be degraded by both lysosomeproteases and the proteasomes and that each degradation processcould be independent of the other. However, because our blottingantibody (C-20) recognizes the last 20 amino acids of the receptor,we could not exclude that only the C-terminal part of the receptorwas degraded by the proteasomes. To test this hypothesis, we testedhow 125I-Epo remained associated with the EpoR when Epodegradation was inhibited by methylamine. To do this, UT-7 cellswere incubated for 90 minutes with 125I-Epo in the presence ofmethylamine. At this time point, most specifically bound 125I-Epowas internalized and protected from degradation by methylamine(Figure 3A). As a control, cells were incubated for 8 minutes with125I-Epo. At this time point, no significant degradation of Epo(Figure 1B) or EpoR (Figure 2C) was detectable. The cells werethen solubilized, and 125I-Epo-EpoR complexes were immunopre-cipitated using C-236 anti-EpoR antibodies. Similar proportions ofradioactivity were immunoprecipitated from cells incubated for 8minutes with Epo alone (51% � 11%) and from cells incubated for90 minutes with Epo and methylamine (48% � 3%). These resultsshow that most 125I-Epo molecules that were protected fromdegradation by lysosome inhibition remained associated with thereceptors, strongly suggesting that only the C-terminal part of theEpoR carrying the epitope recognized in Western blot experimentswas degraded in methylamine-treated cells.
Partial degradation of the EpoR intracellular domainby proteasomes
Because the simultaneous inhibition of proteasomes and lysosomesfully protected EpoR from degradation (Figure 3B), whereaslysosome inhibitors alone protected the N-terminal part of theEpoR only, we hypothesized that the C-terminal part of the EpoRwas degraded by the proteasomes. None of the available antibodiesrecognizing the extracellular domain of the EpoR are able to detectendogenous human EpoRs in Western blots. To overcome thisproblem and to probe whether the C-terminal part of the EpoR wasindeed degraded, we used chemical cross-linking to covalently link125I-Epo to its receptor. UT-7 cells were incubated for 30 minuteswith 125I-Epo, washed to remove unbound radioactivity, andsolubilized. Solubilized extracts were then cross-linked using BS3.In a first step, EpoRs were immunoprecipitated and precipitateswere denatured by boiling in SDS. Precipitates were resolubilizedand first submitted to control immunoprecipitation using anti-GSTantibodies and then to an immunoprecipitation using C-236anti-EpoR antibodies. Lastly, unprecipitated 125I-Epo–cross-linkedproteins were recovered by immunoprecipitation with an anti-Epo
Figure 3. Degradation of internalized Epo and of EpoR. (A) UT-7 cells werepreincubated for 15 minutes at 37°C with 10 mM methylamine (Œ), 25 �Mlactacystin (f), or vehicles alone (E). 125I-Epo was then added to a finalconcentration of 1 nM, and the incubation was continued for 30 minutes. The cellswere then chilled with 10 vol ice-cold phosphate-buffered saline (PBS) andwashed to remove unbound 125I-Epo. Internalized 125I-Epo was determined byacid wash of the cells. For this experiment, this value was 2541 cpm for 106
control cells, 2485 cpm for 106 lactacystin-treated cells, and 2196 cpm for 106
methylamine-treated cells. After washing, the cells were further incubated at 37°Cwith the previously used inhibitors and 50 nM unlabeled Epo to prevent 125I-Eporeassociation with the EpoR. At the indicated times, incubation medium aliquotswere sampled, TCA was added to a final concentration of 15%, and theradioactivity of TCA-soluble fractions was measured. (B) Kinetics of EpoRdegradation in Epo-stimulated cells. UT-7 cells were preincubated for 15 minuteswith cycloheximide alone (control) or in combination with methylamine and/orlactacystin or LLnL as indicated and were stimulated with 10 U/mL Epo for theindicated times. Whole cell extracts were then analyzed by Western blot usingC-20 anti-EpoR antibodies. Arrowheads indicate nonspecific bands. (C) Densito-metric scanning of the experiment presented in panel B. After scanning, theintensity of band 1 was determined using the ImageJ software. For each inhibitor,the intensity of band 1 at t � 0 was set at 100%, and the intensity of this band atlater incubation times was expressed relative to this value.
EPO RECEPTOR DOWN-REGULATION 603BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom
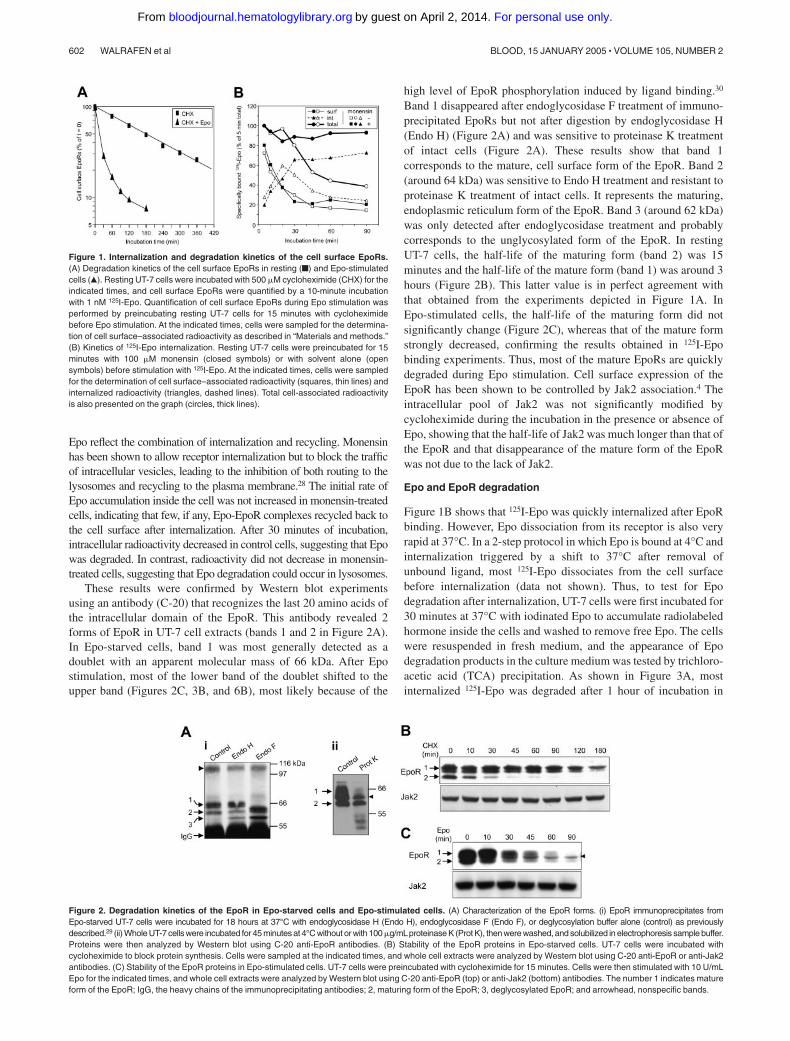
antibody. Several protein bands were detected by autoradiographyafter the first anti-EpoR immunoprecipitation (Figure 4, lane 1).None of these bands were reprecipitated by a nonspecific antibody(anti-GST antibody; Figure 4, lane 2). Only bands A and Bcorresponding to the association of 125I-Epo (34 kDa) with 71 and
40 kDa proteins were reprecipitated by C-236 anti-EpoR antibody(Figure 4, lane 3) and thus corresponded to the receptor itself. Theother bands were recovered after anti-Epo precipitation. The highlyradioactive band with an apparent molecular mass of 34 kDacorresponded to non–cross-linked 125I-Epo, whereas the otherbands should correspond to 125I-Epo cross-linked to EpoR-associated proteins that remain to be identified. When C-20anti-EpoR antibody was substituted for C-236 in the secondimmunoprecipitation, band A but not band B was detected (data notshown). These experiments strongly suggest that band A corre-sponded to the full-length EpoR, whereas band B could be atruncated form of the EpoR. We then tested the effects of proteaseinhibitors on the accumulation of these proteins (Figure 4, rightpanel). When cells were pretreated with methylamine, band Bintensity significantly increased with a simultaneous decrease ofband A. In contrast, when cells were incubated with LLnL, aninhibitor of both proteasome and lysosome activity, no band B wasdetected, whereas band A was strongly increased. Lastly, apulse-chase experiment using unlabeled Epo indicated that theintensity ratio of band B–band A increased during incubation time(data not shown). These results strongly suggest that band B couldbe a degradation product of the EpoR produced by proteasomeaction. From the apparent molecular mass observed in cross-linking experiments (40 kDa after subtracting the 34 kDa of Epo),we estimate that more than half of the intracellular domain wasremoved from the EpoR by proteasomal degradation.
Ubiquitination of the EpoR
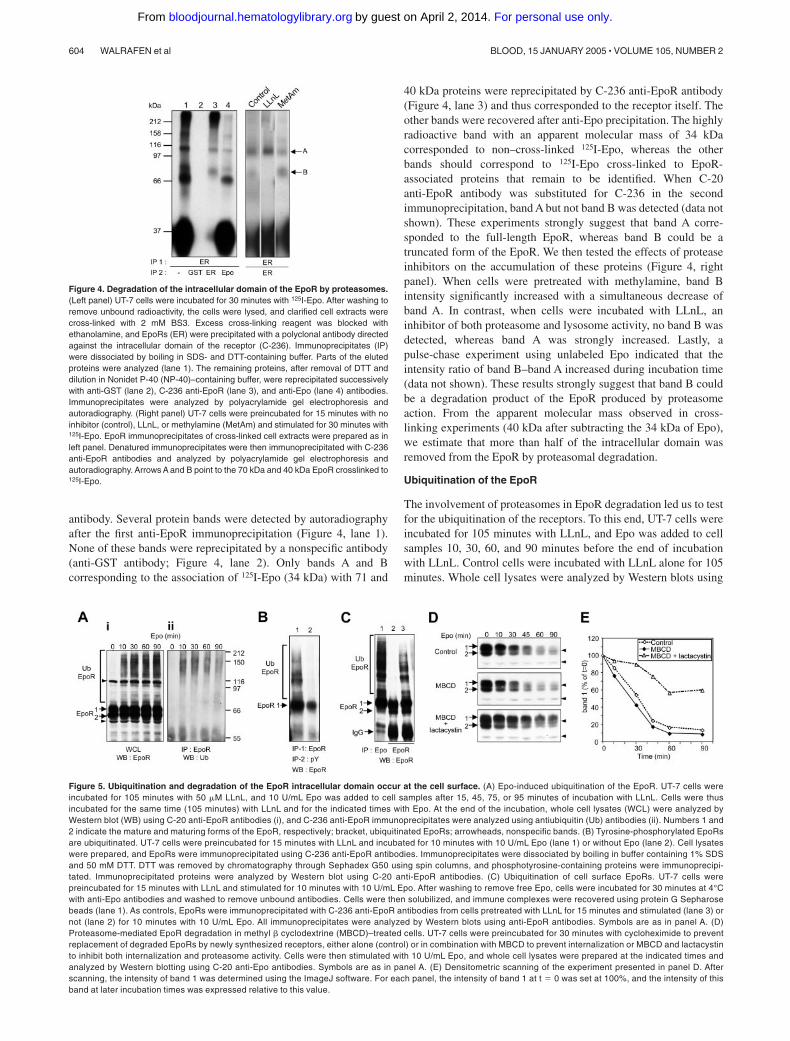
The involvement of proteasomes in EpoR degradation led us to testfor the ubiquitination of the receptors. To this end, UT-7 cells wereincubated for 105 minutes with LLnL, and Epo was added to cellsamples 10, 30, 60, and 90 minutes before the end of incubationwith LLnL. Control cells were incubated with LLnL alone for 105minutes. Whole cell lysates were analyzed by Western blots using
Figure 5. Ubiquitination and degradation of the EpoR intracellular domain occur at the cell surface. (A) Epo-induced ubiquitination of the EpoR. UT-7 cells wereincubated for 105 minutes with 50 �M LLnL, and 10 U/mL Epo was added to cell samples after 15, 45, 75, or 95 minutes of incubation with LLnL. Cells were thusincubated for the same time (105 minutes) with LLnL and for the indicated times with Epo. At the end of the incubation, whole cell lysates (WCL) were analyzed byWestern blot (WB) using C-20 anti-EpoR antibodies (i), and C-236 anti-EpoR immunoprecipitates were analyzed using antiubiquitin (Ub) antibodies (ii). Numbers 1 and2 indicate the mature and maturing forms of the EpoR, respectively; bracket, ubiquitinated EpoRs; arrowheads, nonspecific bands. (B) Tyrosine-phosphorylated EpoRsare ubiquitinated. UT-7 cells were preincubated for 15 minutes with LLnL and incubated for 10 minutes with 10 U/mL Epo (lane 1) or without Epo (lane 2). Cell lysateswere prepared, and EpoRs were immunoprecipitated using C-236 anti-EpoR antibodies. Immunoprecipitates were dissociated by boiling in buffer containing 1% SDSand 50 mM DTT. DTT was removed by chromatography through Sephadex G50 using spin columns, and phosphotyrosine-containing proteins were immunoprecipi-tated. Immunoprecipitated proteins were analyzed by Western blot using C-20 anti-EpoR antibodies. (C) Ubiquitination of cell surface EpoRs. UT-7 cells werepreincubated for 15 minutes with LLnL and stimulated for 10 minutes with 10 U/mL Epo. After washing to remove free Epo, cells were incubated for 30 minutes at 4°Cwith anti-Epo antibodies and washed to remove unbound antibodies. Cells were then solubilized, and immune complexes were recovered using protein G Sepharosebeads (lane 1). As controls, EpoRs were immunoprecipitated with C-236 anti-EpoR antibodies from cells pretreated with LLnL for 15 minutes and stimulated (lane 3) ornot (lane 2) for 10 minutes with 10 U/mL Epo. All immunoprecipitates were analyzed by Western blots using anti-EpoR antibodies. Symbols are as in panel A. (D)Proteasome-mediated EpoR degradation in methyl � cyclodextrine (MBCD)–treated cells. UT-7 cells were preincubated for 30 minutes with cycloheximide to preventreplacement of degraded EpoRs by newly synthesized receptors, either alone (control) or in combination with MBCD to prevent internalization or MBCD and lactacystinto inhibit both internalization and proteasome activity. Cells were then stimulated with 10 U/mL Epo, and whole cell lysates were prepared at the indicated times andanalyzed by Western blotting using C-20 anti-Epo antibodies. Symbols are as in panel A. (E) Densitometric scanning of the experiment presented in panel D. Afterscanning, the intensity of band 1 was determined using the ImageJ software. For each panel, the intensity of band 1 at t � 0 was set at 100%, and the intensity of thisband at later incubation times was expressed relative to this value.
Figure 4. Degradation of the intracellular domain of the EpoR by proteasomes.(Left panel) UT-7 cells were incubated for 30 minutes with 125I-Epo. After washing toremove unbound radioactivity, the cells were lysed, and clarified cell extracts werecross-linked with 2 mM BS3. Excess cross-linking reagent was blocked withethanolamine, and EpoRs (ER) were precipitated with a polyclonal antibody directedagainst the intracellular domain of the receptor (C-236). Immunoprecipitates (IP)were dissociated by boiling in SDS- and DTT-containing buffer. Parts of the elutedproteins were analyzed (lane 1). The remaining proteins, after removal of DTT anddilution in Nonidet P-40 (NP-40)–containing buffer, were reprecipitated successivelywith anti-GST (lane 2), C-236 anti-EpoR (lane 3), and anti-Epo (lane 4) antibodies.Immunoprecipitates were analyzed by polyacrylamide gel electrophoresis andautoradiography. (Right panel) UT-7 cells were preincubated for 15 minutes with noinhibitor (control), LLnL, or methylamine (MetAm) and stimulated for 30 minutes with125I-Epo. EpoR immunoprecipitates of cross-linked cell extracts were prepared as inleft panel. Denatured immunoprecipitates were then immunoprecipitated with C-236anti-EpoR antibodies and analyzed by polyacrylamide gel electrophoresis andautoradiography. Arrows A and B point to the 70 kDa and 40 kDa EpoR crosslinked to125I-Epo.
604 WALRAFEN et al BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom
anti-EpoR antibodies (Figure 5A). Immunoprecipitates of EpoRswere also analyzed by Western blots using antiubiquitin antibod-ies. Smears in the high molecular mass region of the gels wereobserved in immunoblots of anti-EpoRs from whole cell lysatesof Epo-stimulated cells. These smears were also recognized byantiubiquitin antibodies in immunoprecipitates of anti-EpoRs.These results show that the EpoRs are polyubiquitinated inEpo-stimulated cells. Sequential immunoprecipitations usinganti-EpoR and antiphosphotyrosine antibodies clearly showedthat the tyrosine-phosphorylated EpoRs were ubiquitinated(Figure 5B). These experiments were performed in the presenceof LLnL to inhibit the degradation of ubiquinated proteins. Inthe absence of LLnL, EpoR ubiquitination was also detectable inEpo-stimulated cells, although very transiently (data not shown).
To examine whether stimulated EpoRs were ubiquinated atthe cell surface, Epo was bound to UT-7 cells, and the cells wereincubated with anti-Epo antibodies before solubilization. Im-mune complexes were then analyzed by Western blot usinganti-EpoR antibodies (Figure 5C). Solubilized extracts fromEpo-stimulated and unstimulated cells were also immunoprecipi-tated using C-236 anti-EpoR antibodies as controls. The matur-ing form of the EpoRs (Figure 5C, band 2) was observed inEpoR precipitates but not in Epo precipitates, showing thatno Epo exchange occurred during solubilization or antibodyrecovery. These results clearly show that the cell surface,Epo-bound EpoRs were ubiquitinated. Methyl � cyclodextrine(MBCD) has been shown to inhibit cell surface protein internal-ization.32,33 Accordingly, 125I-Epo did not internalize and re-mained at the cell surface of UT-7 cells preincubated with 5 mMMBCD (data not shown). Cell surface blockage of the EpoRby MBCD did not prevent its ubiquitination or its tyrosinephosphorylation (data not shown) or the degradation of itsC terminus (Figure 5D-E). Lactacystin inhibited the degradationof EpoR in MBCD-treated cells. These results show that bothubiquitination and proteasomal degradation of the EpoR oc-curred at the cell surface.
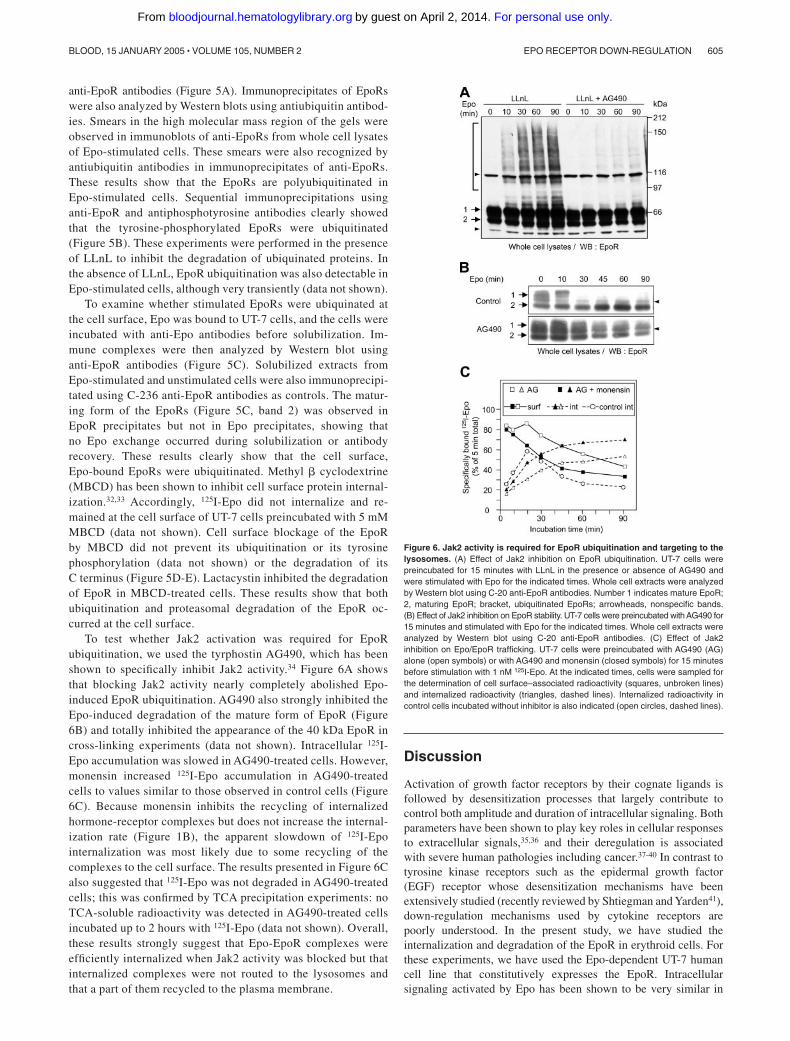
To test whether Jak2 activation was required for EpoRubiquitination, we used the tyrphostin AG490, which has beenshown to specifically inhibit Jak2 activity.34 Figure 6A showsthat blocking Jak2 activity nearly completely abolished Epo-induced EpoR ubiquitination. AG490 also strongly inhibited theEpo-induced degradation of the mature form of EpoR (Figure6B) and totally inhibited the appearance of the 40 kDa EpoR incross-linking experiments (data not shown). Intracellular 125I-Epo accumulation was slowed in AG490-treated cells. However,monensin increased 125I-Epo accumulation in AG490-treatedcells to values similar to those observed in control cells (Figure6C). Because monensin inhibits the recycling of internalizedhormone-receptor complexes but does not increase the internal-ization rate (Figure 1B), the apparent slowdown of 125I-Epointernalization was most likely due to some recycling of thecomplexes to the cell surface. The results presented in Figure 6Calso suggested that 125I-Epo was not degraded in AG490-treatedcells; this was confirmed by TCA precipitation experiments: noTCA-soluble radioactivity was detected in AG490-treated cellsincubated up to 2 hours with 125I-Epo (data not shown). Overall,these results strongly suggest that Epo-EpoR complexes wereefficiently internalized when Jak2 activity was blocked but thatinternalized complexes were not routed to the lysosomes andthat a part of them recycled to the plasma membrane.
Discussion
Activation of growth factor receptors by their cognate ligands isfollowed by desensitization processes that largely contribute tocontrol both amplitude and duration of intracellular signaling. Bothparameters have been shown to play key roles in cellular responsesto extracellular signals,35,36 and their deregulation is associatedwith severe human pathologies including cancer.37-40 In contrast totyrosine kinase receptors such as the epidermal growth factor(EGF) receptor whose desensitization mechanisms have beenextensively studied (recently reviewed by Shtiegman and Yarden41),down-regulation mechanisms used by cytokine receptors arepoorly understood. In the present study, we have studied theinternalization and degradation of the EpoR in erythroid cells. Forthese experiments, we have used the Epo-dependent UT-7 humancell line that constitutively expresses the EpoR. Intracellularsignaling activated by Epo has been shown to be very similar in
Figure 6. Jak2 activity is required for EpoR ubiquitination and targeting to thelysosomes. (A) Effect of Jak2 inhibition on EpoR ubiquitination. UT-7 cells werepreincubated for 15 minutes with LLnL in the presence or absence of AG490 andwere stimulated with Epo for the indicated times. Whole cell extracts were analyzedby Western blot using C-20 anti-EpoR antibodies. Number 1 indicates mature EpoR;2, maturing EpoR; bracket, ubiquitinated EpoRs; arrowheads, nonspecific bands.(B) Effect of Jak2 inhibition on EpoR stability. UT-7 cells were preincubated with AG490 for15 minutes and stimulated with Epo for the indicated times. Whole cell extracts wereanalyzed by Western blot using C-20 anti-EpoR antibodies. (C) Effect of Jak2inhibition on Epo/EpoR trafficking. UT-7 cells were preincubated with AG490 (AG)alone (open symbols) or with AG490 and monensin (closed symbols) for 15 minutesbefore stimulation with 1 nM 125I-Epo. At the indicated times, cells were sampled forthe determination of cell surface–associated radioactivity (squares, unbroken lines)and internalized radioactivity (triangles, dashed lines). Internalized radioactivity incontrol cells incubated without inhibitor is also indicated (open circles, dashed lines).
EPO RECEPTOR DOWN-REGULATION 605BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom

UT-7 cells and in normal erythroid progenitors that are the truetarget cells of Epo.42 However, internalization studies requirenumbers of cells difficult to obtain with normal progenitors.Because UT-7 cells express endogenous EpoRs, they constitute avaluable model for EpoR trafficking studies because artifacts dueto overexpression of transfected receptors are not supposed tooccur. Accordingly, our results show that more than half of theEpoRs in resting UT-7 cells show a complex glycosylation patternand are present at the cell surface, whereas it has been reported thatmost EpoRs accumulate inside EpoR-transfected cells.2,3 There-fore, the artifactual presence of a large intracellular pool of EpoR intransfected cells that is prone to interfere with trafficking studies isavoided by the use of cell lines expressing endogenous receptors.
In the present paper, we show that Epo binding to its receptorinduces ubiquitination, internalization, and degradation of theEpoR. Figure 7 summarizes our results and describes the down-regulation mechanisms that are turned on by Epo binding to itsreceptor. In sharp contrast with the GH receptor (GHR), for whichinternalization is constitutive16 (and GHR degradation is onlyslightly increased by GH),43 internalization and degradation of theEpoR are strongly increased by ligand binding. It has been shownpreviously that EpoRs unable to bind and activate Jak2 wereefficiently internalized after Epo binding, suggesting that Jak2activation was dispensable for EpoR internalization.11 In agreementwith these results, we observed that blocking Jak2 activity did notinhibit Epo-induced EpoR internalization. Because EpoR ubiquiti-nation was inhibited when Jak2 activity was blocked, we concludethat EpoR ubiquitination is not required for internalization. Over-all, these results suggest that Epo binding to its receptor induces aconformational change of the EpoR that is sufficient to promoteEpoR internalization.
In contrast to internalization, Epo and EpoR degradation isdependent on Jak2 activity, and when Jak2 activity was blocked,some Epo-EpoR complexes recycled back to the cell surface.Lysosome inhibitors such as methylamine (Figure 3), ammo-nium chloride, or chloroquine (data not shown) protectedinternalized Epo from degradation, indicating that Epo isdegraded in the lysosomes. The cathepsin B–specific inhibitor44
CA-074-Me largely inhibited 125I-Epo degradation, showingthat cathepsin B is involved in Epo degradation (data notshown). In contrast, blocking proteasome activity using lactacys-tin did not inhibit Epo degradation.
The EpoRs are rapidly ubiquitinated after Epo binding. Thisprocess occurs at the cell surface and requires Jak2 activation. TheE3 ligase proteins responsible for EpoR ubiquitination are notidentified. We have observed that c-Cbl associates with thetyrosine-phosphorylated form of the EpoR (F.V., unpublishedobservation, December 2003). However, overexpression in UT-7 cellsof a mutated form of c-Cbl devoid of E3 ligase activity or blocking c-Cblsynthesis using small interfering RNA (siRNA) did not significantlydecrease Epo-induced ubiquitination of the EpoR (data not shown).SOCS-3 also associates with the activated EpoR10 and could belong to aubiquitination complex. However, the kinetics of SOCS-3 inductionafter Epo stimulation is poorly compatible with the rapid ubiquitinationof the activated EpoR. Moreover, overexpression in UT-7 cells of aSOCS-3 mutant deleted of the SOCS domain did not decrease Epo-induced ubiquitination of the EpoR (data not shown), confirming thatSOCS-3 was probably not involved in EpoR ubiquitination. Recently,the ubiquitin ligase Rul (p33) has been shown to associate to the EpoRand to induce its ubiquitination.45 However, at least in COS cells,p33-induced ubiquitination of the EpoR is constitutive and not regulatedby Epo stimulation of the cells. Whether Rul mediates Epo-inducedubiquitination of the EpoR in Epo-responsive cells has to be determined.
Ubiquitination contributes to the control of several cellularprocesses including targeting proteins for proteasomal degradation,internalization, and sorting of internalized proteins to late endo-somes and lysosomes. Polyubiquitination most often leads toprotein degradation by the proteasome, whereas internalization andintracellular routing of proteins generally involve monoubiquitina-tion.46 Polyubiquitination of the EpoR is probably responsible forthe proteolysis of the EpoRs by proteasomes (Figure 4) thatremoves a significant part of the intracellular domain. A similarmechanism of degradation has been reported for the GHR47 as wellas the shared � subunit (�c) of the IL-3, IL-5, and granulocyte-macrophage colony-stimulating factor (GM-CSF) receptors.48 Ago-nistic ligation of these receptors induces proteasomal degradationof the intracellular domain of �c, leading to a signalization-deficient receptor form containing only 25 to 35 intracellular aminoacids. Although the extent of EpoR degradation by proteasomescannot be accurately deduced from our cross-linking experiments,our results suggest that more than half of the intracellular domain isdegraded by these proteases. Because tyrosine residues are locatedin this region of the receptor, this mechanism could hasten thedesensitization process. In contrast to �c, whose proteasomaldegradation of the intracellular domain is necessary for internaliza-tion and lysosome routing,48 inhibiting proteasome activity did notsignificantly modify the degradation of Epo and EpoR by thelysosomes, showing that initial degradation of this part of the EpoRis not required for lysosome targeting and degradation. Epo andEpoR targeting to the lysosomes and their degradation correlatewith EpoR ubiquitination, strongly suggesting that EpoRs behavesimilarly to several receptors, such as EGF receptors (EGFRs),49
interferon-� receptors,50 or IL-2 receptors,21 whose ubiquitinationcontrols targeting to the lysosome.
After internalization, both Epo and the EpoR are degraded bythe lysosomes; and few, if any, EpoRs are recycled back to themembrane. This is in sharp contrast with the TPO receptor thatpromptly recycles onto the cell surface.14 These properties could atleast partly explain why intracellular transduction by the TPOreceptor is sustained for several hours,13 whereas Epo signaling isof short duration.12 Prolonged activation of TPO signaling has beenshown to be required for megakaryocytic differentiation by main-taining a high level of Erk activation.13 In contrast, it is suggestedthat erythroid differentiation could require a decreased level of
Figure 7. Mechanisms of EpoR down-regulation during Epo stimulation. UponEpo binding, Jak2 (not represented) and the EpoR are tyrosine-phosphorylated,triggering intracellular signaling (Ia). At the cell surface, the receptor is ubiquitinated(II), allowing for recognition by the proteasome and degradation of the cytoplasmictail (III). This degradation process removes the part of the receptor carrying all of thephosphorylated tyrosine residues of the intracellular domain, preventing furthersignal transduction. The cleaved receptor is then internalized and degraded in thelysosomes (IV). When Jak2 activation is prevented by the inhibitor AG490 (Ib), thereceptor is neither phosphorylated nor ubiquitinated. Epo-EpoR complexes are stillefficiently internalized, but no degradation occurs and the complexes recycle tothe surface.
606 WALRAFEN et al BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom

intracellular signaling. Indeed, it has been reported that activationof Foxo3A 51 and caspases52 was necessary for terminal erythroiddifferentiation. These activations are inhibited by intracellularsignaling relays such as the phosphatidylinositol-3 kinase pathway,which is strongly activated in Epo-stimulated cells and is requiredfor erythroid cell proliferation.42 Thus, we can propose thatdecreasing Epo receptor signaling could lower cell proliferationand facilitate terminal differentiation. Because the cell surfaceexpression of newly synthesized EpoRs is inhibited by amechanism involving proteasomes in stimulated cells (ourprevious results: Verdier et al12), both this mechanism and the
degradation of activated receptors should lead to the progressivedisappearance of cell surface receptors; and, accordingly, reticu-locytes are devoid of EpoRs.53
Acknowledgments
We are grateful to Dr Veronique Duprez and Dominique Dumenil(Cochin Institute, Paris, France) for critical reading of themanuscript and helpful comments and Odile Muller for techni-cal expertise.
References
1. Lacombe C, Mayeux P. Erythropoietin (Epo) re-ceptor and Epo mimetics. Adv Nephrol NeckerHosp. 1999;29:177-189.
2. Yoshimura A, D’Andrea AD, Lodish HF. Friendspleen focus-forming virus glycoprotein gp55 in-teracts with the erythropoietin receptor in the en-doplasmic reticulum and affects receptor metabo-lism. Proc Natl Acad Sci U S A. 1990;87:4139-4143.
3. Neumann D, Wikstrom L, Watowich SS, LodishHF. Intermediates in degradation of the erythro-poietin receptor accumulate and are degraded inlysosomes. J Biol Chem. 1993;268:13639-13649.
4. Huang LJ, Constantinescu SN, Lodish HF. TheN-terminal domain of Janus kinase 2 is requiredfor Golgi processing and cell surface expressionof erythropoietin receptor. Mol Cell. 2001;8:1327-1338.
5. Constantinescu SN, Ghaffari S, Lodish HF. Theerythropoietin receptor: structure, activation andintracellular signal transduction. Trends Endocri-nol Metab. 1999;10:18-23.
6. Klingmuller U, Lorenz U, Cantley LC, Neel BG,Lodish HF. Specific recruitment of SH-PTP1 tothe erythropoietin receptor causes inactivation ofJAK2 and termination of proliferative signals.Cell. 1995;80:729-738.
7. Jiao H, Berrada K, Yang W, Tabrizi M, PlataniasLC, Yi T. Direct association with and dephos-phorylation of Jak2 kinase by the SH2-domain-containing protein tyrosine phosphatase SHP-1.Mol Cell Biol. 1996;16:6985-6992.
8. Yoshimura A, Ohkubo T, Kiguchi T, et al. Anovel cytokine-inducible gene CIS encodes anSH2-containing protein that binds to tyrosine-phosphorylated interleukin 3 and erythropoietinreceptors. EMBO J. 1995;14:2816-2826.
9. Verdier F, Chretien S, Muller O, et al. Protea-somes regulate erythropoietin receptor and signaltransducer and activator of transcription 5 (STAT5)activation: possible involvement of the ubiquitinatedcis protein. J Biol Chem. 1998;273:28185-28190.
10. Sasaki A, Yasukawa H, Shouda T, Kitamura T,Dikic I, Yoshimura A. CIS3/SOCS-3 suppresseserythropoietin (EPO) signaling by binding theEPO receptor and JAK2. J Biol Chem. 2000;275:29338-29347.
11. Beckman DL, Lin LL, Quinones ME, LongmoreGD. Activation of the erythropoietin receptor isnot required for internalisation of bound erythro-poietin. Blood. 1999;94:2667-2675.
12. Verdier F, Walrafen P, Hubert N, et al. Protea-somes regulate the duration of erythropoietinreceptor activation by controlling down-regulation of cell surface receptors. J Biol Chem.2000;275:18375-18381.
13. Rouyez MC, Boucheron C, Gisselbrecht S,Dusanter-Fourt I, Porteu F. Control of thrombo-poietin-induced megakaryocytic differentiation bythe mitogen-activated protein kinase pathway.Mol Cell Biol. 1997;17:4991-5000.
14. Dahlen DD, Broudy VC, Drachman JG. Internal-ization of the thrombopoietin receptor is regulated
by 2 cytoplasmic motifs. Blood. 2003;102:102-108.
15. Duprez V, Cornet V, Dautry-Varsat A. Down-regulation of high affinity interleukin 2 receptors ina human tumor T cell line: interleukin 2 increasesthe rate of surface receptor decay. J Biol Chem.1988;263:12860-12865.
16. Strous GJ, Gent J. Dimerization, ubiquitylationand endocytosis go together in growth hormonereceptor function. FEBS Lett. 2002;529:102-109.
17. van Kerkhof P, Govers R, Alves dos Santos CM,Strous GJ. Endocytosis and degradation of thegrowth hormone receptor are proteasome-dependent. J Biol Chem. 2000;275:1575-1580.
18. van Kerkhof P, Strous GJ. The ubiquitin-proteasome pathway regulates lysosomal degra-dation of the growth hormone receptor and itsligand. Biochem Soc Trans. 2001;29:488-493.
19. Lamaze C, Dujeancourt A, Baba T, Lo CG, Ben-merah A, Dautry-Varsat A. Interleukin 2 receptorsand detergent-resistant membrane domains de-fine a clathrin-independent endocytic pathway.Mol Cell. 2001;7:661-671.
20. Hemar A, Subtil A, Lieb M, Morelon E, Hellio R,Dautry-Varsat A. Endocytosis of interleukin 2 re-ceptors in human T lymphocytes: distinct intracel-lular localization and fate of the receptor alpha,beta, and gamma chains. J Cell Biol. 1995;129:55-64.
21. Rocca A, Lamaze C, Subtil A, Dautry-Varsat A.Involvement of the ubiquitin/proteasome systemin sorting of the interleukin 2 receptor beta chainto late endocytic compartments. Mol Biol Cell.2001;12:1293-1301.
22. Yen CH, Yang YC, Ruscetti SK, Kirken RA,Dai RM, Li CC. Involvement of the ubiquitin-proteasome pathway in the degradation of non-tyrosine kinase-type cytokine receptors of IL-9,IL-2, and erythropoietin. J Immunol. 2000;165:6372-6380.
23. Yu A, Malek TR. The proteasome regulates re-ceptor-mediated endocytosis of interleukin-2.J Biol Chem. 2001;276:381-385.
24. Komatsu N, Nakauchi H, Miwa A, et al. Establish-ment and characterization of a human leukemiccell line with megakaryocytic features: depen-dency on granulocyte-macrophage colony stimu-lating factor, interleukin 3, or erythropoietin forgrowth and survival. Cancer Res. 1991;51:341-345.
25. Verdier F, Chretien S, Billat C, Gisselbrecht S,Lacombe C, Mayeux P. Erythropoietin inducesthe tyrosine phosphorylation of insulin receptorsubstrate-2: an alternate pathway for erythropoi-etin-induced phosphatidylinositol 3-kinase activa-tion. J Biol Chem. 1997;272:26173-26178.
26. Mayeux P, Billat C, Jacquot R. The erythropoietinreceptor of rat erythroid progenitor cell: character-ization and affinity cross-linkage. J Biol Chem.1987;262:13985-13990.
27. Mayeux P, Casadevall N, Lacombe C, Muller O,Tambourin P. Solubilization and hydrodynamiccharacteristics of the erythropoietin receptor: evi-dence for a multimeric structure. Eur J Biochem.1990;194:271-278.
28. Wang Y, Pennock S, Chen X, Wang Z. Internal-ization of inactive EGF receptor into endosomesand the subsequent activation of endosome-associated EGF receptors. Sci STKE. 2002;161:PL17.
29. Casadevall N, Nataf J, Viron B, et al. Pure red-cell aplasia and antierythropoietin antibodies inpatients treated with recombinant erythropoietin.N Engl J Med. 2002;346:469-475.
30. Dusanter-Fourt I, Casadevall N, Lacombe C, etal. Erythropoietin induces the tyrosine phosphory-lation of its own receptor in human erythropoietin-responsive cells. J Biol Chem. 1992;267:10670-10675.
31. Reijngoud DJ, Oud PS, Kas J, Tager JM. Rela-tionship between medium pH and that of the lyso-somal matrix as studied by two independentmethods. Biochim Biophys Acta. 1976;448:290-302.
32. Subtil A, Gaidarov I, Kobylarz K, Lampson MA,Keen JH, McGraw TE. Acute cholesterol deple-tion inhibits clathrin-coated pit budding. Proc NatlAcad Sci U S A. 1999;96:6775-6780.
33. Rodal SK, Skretting G, Garred O, Vilhardt F, vanDeurs B, Sandvig K. Extraction of cholesterol withmethyl-beta-cyclodextrin perturbs formation ofclathrin-coated endocytic vesicles. Mol Biol Cell.1999;10:961-974.
34. Meydan N, Grunberger T, Dadi H, et al. Inhibitionof acute lymphoblastic leukaemia by a Jak-2 in-hibitor. Nature. 1996;379:645-648.
35. Marshall CJ. Specificity of receptor tyrosine ki-nase signaling: transient versus sustained extra-cellular signal-regulated kinase activation. Cell.1995;80:179-185.
36. Murphy LO, Smith S, Chen RH, Fingar DC, BlenisJ. Molecular interpretation of ERK signal durationby immediate early gene products. Nat Cell Biol.2002;4:556-564.
37. Ciesielski MJ, Fenstermaker RA. Oncogenic epi-dermal growth factor receptor mutants with tan-dem duplication: gene structure and effects onreceptor function. Oncogene. 2000;19:810-820.
38. Schmidt MH, Furnari FB, Cavenee WK, Bogler O.Epidermal growth factor receptor signaling inten-sity determines intracellular protein interactions,ubiquitination, and internalization. Proc Natl AcadSci U S A. 2003;100:6505-6510.
39. Hunter MG, Avalos BR. Deletion of a critical inter-nalization domain in the G-CSFR in acute my-elogenous leukemia preceded by severe con-genital neutropenia. Blood. 1999;93:440-446.
40. Ward AC, van Aesch YM, Schelen AM, Touw IP.Defective internalization and sustained activationof truncated granulocyte colony-stimulating factorreceptor found in severe congenital neutropenia/acute myeloid leukemia. Blood. 1999;93:447-458.
41. Shtiegman K, Yarden Y. The role of ubiquitylationin signaling by growth factors: implications to can-cer. Semin Cancer Biol. 2003;13:29-40.
42. Bouscary D, Pene F, Claessens YE, et al. Criticalrole for PI 3-kinase in the control of erythropoi-etin-induced erythroid progenitor proliferation.Blood. 2003;101:3436-3443.
EPO RECEPTOR DOWN-REGULATION 607BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom

43. Alves dos Santos CM, ten Broeke T, Strous GJ.Growth hormone receptor ubiquitination, endocy-tosis, and degradation are independent of signaltransduction via Janus kinase 2. J Biol Chem.2001;276:32635-32641.
44. Buttle DJ, Murata M, Knight CG, Barrett AJ.CA074 methyl ester: a proinhibitor for intracellularcathepsin B. Arch Biochem Biophys. 1992;299:377-380.
45. Friedman AD, Nimbalkar D, Quelle FW. Erythro-poietin receptors associate with a ubiquitin ligase,p33RUL, and require its activity for erythropoietin-induced proliferation. J Biol Chem. 2003;278:26851-26861.
46. Mosesson Y, Shtiegman K, Katz M, et al. Endocy-tosis of receptor tyrosine kinases is driven by mo-
noubiquitylation, not polyubiquitylation. J BiolChem. 2003;278:21323-21326.
47. Strous GJ, van Kerkhof P. The ubiquitin-proteasome pathway and the regulation of growthhormone receptor availability. Mol Cell Endocri-nol. 2002;197:143-151.
48. Martinez-Moczygemba M, Huston DP. Proteaso-mal regulation of betac signaling reveals a novelmechanism for cytokine receptor heterotypic de-sensitization. J Clin Invest. 2001;108:1797-1806.
49. Levkowitz G, Waterman H, Zamir E, et al. c-Cbl/Sli-1 regulates endocytic sorting and ubiquitina-tion of the epidermal growth factor receptor.Genes Dev. 1998;12:3663-3674.
50. Kumar KG, Tang W, Ravindranath AK, Clark WA,Croze E, Fuchs SY. SCF(HOS) ubiquitin ligase
mediates the ligand-induced down-regulation ofthe interferon-alpha receptor. EMBO J. 2003;22:5480-5490.
51. Bakker WJ, Blazquez-Domingo M, Kolbus A, etal. FoxO3a regulates erythroid differentiation andinduces BTG1, an activator of protein argininemethyl transferase 1. J Cell Biol. 2004;164:175-184.
52. Zermati Y, Garrido C, Amsellem S, et al. Caspaseactivation is required for terminal erythroid differ-entiation. J Exp Med. 2001;193:247-254.
53. Broudy VC, Lin N, Brice M, Nakamoto B, Papayan-nopoulou T. Erythropoietin receptor characteristicson primary human erythroid cells. Blood. 1991;77:2583-2590.
608 WALRAFEN et al BLOOD, 15 JANUARY 2005 � VOLUME 105, NUMBER 2
For personal use only.on April 2, 2014. by guest bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN