
In vitro effects of methylmercury on ascidian (Styela plicata) immunocyte responses

Induction of anterior neural fates in the ascidian Ciona intestinalis
Clare Hudson*, Patrick Lemaire*
Laboratoire de GeÂneÂtique et Physiologie du DeÂveloppement, Institut de Biologie du DeÂveloppement de Marseille, CNRS-INSERM-Universite de la
Mediterranee-AP de Marseille, Parc Scienti®que de Luminy, Case 907, F-13288, Marseille Cedex 9, France
Received 26 September 2000; received in revised form 24 October 2000; accepted 25 October 2000
Abstract
The sensory vesicle of ascidians is thought to be homologous to the vertebrate forebrain and midbrain (Development 125 (1998) 1113).
Here we report the isolation of two sensory vesicle markers in the ascidian Ciona intestinalis, which are homologs of vertebrate otx and gsx
homeobox genes. By using these markers to analyze the induction of anterior neural tissue in Ciona, we ®nd that the restriction of anterior
neural fate to the progeny of the anterior animal blastomeres is due to a combination of two factors. The vegetal blastomeres show a
differential inducing activity along the anterior-posterior axis, while the competence to respond to this inducing signal is markedly higher in
the anterior animal blastomeres than in the posterior animal blastomeres. This differential competence to respond is also observed in response
to bFGF, a candidate neural inducer in ascidians (J. Physiol. 511.2 (1998) 347) and can be detected by the gastrula stage. Our results,
however, indicate that bFGF can only induce a subset of the responses of the endogenous inducer, suggesting that additional signals in the
embryo are necessary to induce a fully patterned nervous system. q 2001 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Gsx; Gsh; otx; Ciona intestinalis; Ascidians; Urochordate; Sensory vesicle; Neural induction; Neural competence; bFGF; Anterior-posterior pattern
1. Introduction
Ascidians are primitive invertebrate chordates that have
been classically used for developmental biology studies
(Conklin, 1905; Nishida, 1997). They are classed into two
orders, determined by the position of the gonad: the Enter-
ogona, with the gonads in the gut loop, including Ciona,
Ascidia and Phallusia, and the Pleurogona, with paired
gonads, including Styela, Halocynthia and Mogula
(reviewed in Satoh, 1994). During larval stages ascidians
exhibit a remarkably simple chordate body plan, possessing
a notochord and dorsal hollow nerve cord (for a general
review see Satoh, 1994; Nishida, 1997). Both the cleavage
pattern and the cell lineage are invariant in early ascidians.
Gastrulation starts at the 110-cell stage, when the majority of
cells are already restricted to form one tissue fate (Nishida,
1997). By larval stages, Ciona contains approximately 2500
cells of which 266 constitute the central nervous system
(Nicol and Meinertzhagen, 1991). Despite the high degree
of conservation of cleavage pattern and cell lineage between
the different ascidian species and an apparent conservation of
gene expression pro®les, regulation of gene expression may
have diverged signi®cantly in some cases. This was recently
highlighted in a study showing that the upstream regulatory
sequences of brachyury genes in Halocynthia and Ciona are
not conserved (Takahashi et al., 1999).
The nervous system of ascidian tadpoles consists of the
sensory vesicle (possibly equivalent to the forebrain and
midbrain), the neck region (midbrain or anterior hindbrain),
and the visceral ganglion and nerve cord (hindbrain and
spinal cord) (Wada et al., 1998). The sensory vesicle
contains two sense organs, each containing a melanocyte
or pigment cell: the posterior pigment cell is part of the
ocellus within a light-sensitive eye-like structure, and the
anterior pigment cell forms the gravity sensitive otolith.
Fate mapping experiments have determined that the
central nervous system mainly derives from the anterior
half of the eight-cell embryo (the A4.1 and a4.2 pairs;
Fig. 1) (Nishida, 1987; Nicol and Meinertzhagen,
1988a,b) (from henceforth blastomeres will be referred to
simply by their name, rather than as a pair). The sensory
vesicle forms from a4.2 derivatives (Fig. 1). The neck, visc-
eral ganglion and nerve cord predominantly develops from
A4.1 derivatives, with the dorsal-most cells being derived
from the b4.2 blastomere (Fig. 1).
Early experiments in ascidians revealed that the majority
of blastomeres, when isolated from eight-cell embryos,
continue to develop according to their normal lineage, high-
Mechanisms of Development 100 (2001) 189±203
0925-4773/01/$ - see front matter q 2001 Elsevier Science Ireland Ltd. All rights reserved.
PII: S0925-4773(00)00528-1
www.elsevier.com/locate/modo
* Corresponding authors. Tel.: 133-4-91-829248; fax: 133-4-91-
820682.
E-mail addresses: [email protected] (C. Hudson),
[email protected] (P. Lemaire).

lighting the importance of cell autonomous maternal deter-
minants in ascidian development. One notable exception to
this mosaic development was a lack of neural differentiation
in animal half (a4.2 1 b4.2) embryos or animal blastomeres
cultured in isolation. It was further shown that induction of
neural fate in animal blastomeres, monitored by formation
of pigment cells, required an interaction with A4.1 blasto-
mere derivatives (Rose, 1939; Reverberi and Minganti,
1946, 1947). Analyzing membrane excitability in clea-
vage-arrested embryos of Halocynthia aurantium and Halo-
cynthia roretzi con®rmed that a signal was required from the
inducing cell A4.1 for the responding cell a4.2 to adopt a
neural fate and that this induction occurs within a speci®c
time window, being completed around the early gastrula
stage (Okado and Takahashi, 1990a). Although b4.2
normally does not differentiate into pigment cells or
neurons, it was shown that it could respond to the A4.1-
derived signals and develop both neural-type membrane
excitability in Halocynthia and form pigment cells in Styela
and Aspetto (Rose, 1939; Reverberi and Minganti, 1947;
Okado and Takahashi, 1990a). In all cases, however, the
competence of b4.2 to adopt a neural fate was signi®cantly
lower than that of a4.2 (Rose, 1939; Reverberi and
Minganti, 1947; Okado and Takahashi, 1990a). Finally,
more recent experiments have revealed that serine protease
or basic FGF treatment of the a4.2 blastomere (Ortolani et
al., 1979; Okado and Takahashi, 1990a, 1993; Inazawa et
al., 1998) can mimic the inductive properties of A4.1. The
action of bFGF was restricted to a similar time window as
induction via cell contact with the endogenous inducer,
while proteases showed slightly different kinetics, invoking
a neural response in a much shorter time window (Inazawa
et al., 1998).
These studies, which initiated the analysis of neural
induction in ascidians, had two major limitations. First, in
these experiments neural membrane excitability and
pigment cell formation identify only a subset of the a4.2-
derived neural cells. It is therefore important to use addi-
tional neural markers to test whether the results described
above can be extended to all a4.2-derived neural lineages.
Second, these studies focused on terminal differentiation
and therefore little is known about the intermediate steps
of neural tissue formation. For example, it is unclear when
the response of a4.2 and b4.2 to the signal starts to differ. Is
the inducing signal initially interpreted differently by each
cell, or is an identical early response translated later into the
adoption of distinct fates? The response to this important
question requires the isolation of early neural markers.
Here, we have cloned Ciona homologs of vertebrate otx
and gsx which are expressed in the a4.2-derived neural
lineages and which we used to address these issues.
2. Results
2.1. Identi®cation of Ciona homeobox-containing genes
belonging to the gsx/ind and otx families
We have cloned a Ciona intestinalis homeobox gene
encoding for a protein whose homeodomain is most similar
to that of mouse and medaka gsh (78% amino acid identity),
Amphioxus AmphiGsx (75% identity) and Drosophila ind
(73% amino acid identity) (Fig. 2b). Gsh (Genomic Screen-
ing Homeobox) genes were ®rst identi®ed in mouse (gsh-1
and 2; Singh et al., 1991; Valerius et al., 1995; Hsieh-Li et al.,
1995) and subsequently in medaka (Ol-Gsh 1; Deschet et al.,
1998), Amphioxus (AmphiGsx; Brooke et al., 1998) and
Drosophila (intermediate neuroblasts defective (ind);
Weiss et al., 1998) and expression of these molecules is
restricted to the nervous system. Outside of the gsh/ind
family, the most similar homeodomain is that of the
HoxD4 gene (65% amino acid identity). A shorter fragment
of the same cDNA has previously been cloned and named Ci-
Hbox2 (Di Gregorio et al., 1995). On the basis of sequence
comparisons and expression data (see below) we propose to
rename Ci-Hbox2, Ci-gsx. Outside of the homeodomain, the
predicted Ci-gsx amino acid sequence shares little homology
with its homologs in other species and lacks the polyalanine
tracts found in vertebrate gsh proteins (Valerius et al., 1995;
Hsieh-Li et al., 1995; Deschet et al., 1998) (Fig. 2a).
We also cloned a Ciona homolog of vertebrate otx and
Drosophila otd, which are involved in anterior neural
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203190
Fig. 1. Fate map of the ascidian nervous system (taken and adapted from Nishida, 1987). On the right of the ®gure, the central nervous system is shown, and on
the left a lateral view of an eight-cell stage embryo is shown. The anterior nervous system, the sensory vesicle, is derived from a4.2. The nerve cord is derived
from A4.1, with the dorsal-most cells being descendants of b4.2.

formation from Drosophila to mouse (Finkelstein and
Boncinelli, 1994). The protein sequence of Ci-otx is very
similar to Hroth, the Halocynthia counterpart (Wada et al.,
1996), sharing 52% identity over the entire protein
sequence. The homeodomain of Ci-otx shares a high degree
of conservation with other Otx homeodomains (Fig. 2c).
2.2. Ciona gsx is expressed in the posterior sensory vesicle
Expression of Ci-gsx in early embryos was ®rst detected
by in situ hybridization at the mid-gastrula stage in two
blastomeres of the neural plate (Fig. 3a). The neural plate
of ascidians is a simple structure consisting of a monolayer
of cells, of which the cleavage pattern is well documented
allowing one to precisely identify each cell and know its
lineage (Nicol and Meinertzhagen, 1988a,b). Expression of
Ci-gsx is restricted to the neural plate (Fig. 3i,j). In order to
determine precisely which cells express Ci-gsx, we
collected embryos at sequential time points during
gastrula-neurula stages and revealed Ci-gsx expression by
in situ hybridization. We then stained the embryos with a
nuclear dye, which allowed us to identify each blastomere
by comparing the cell number and pattern of cell division
with the neural plate fate map established by Nicol and
Meinertzhagen (1988b). Ci-gsx expression was ®rst
detected in two cells in the third row of neural plate cells
(Fig. 3a,e). The precise identity of these cells was con®rmed
by following the pattern of cell cleavages. First the medial
four cells in row II divide to give the 44-cell neural plate
(Nicol and Meinertzhagen, 1988b) (Fig. 3b,f). This is
followed by the medial four cells in row I (Fig. 4c,g).
Next the medial four cells in rows III and V divide, along
with the lateral cells in rows I and II (Fig. 3d,h). From this
analysis, we conclude that Ci-gsx expression is ®rst detected
in the a9.33 pair of blastomeres, which are fated to form
posterior sensory vesicle (Nishida, 1987; Nicol and
Meinertzhagen, 1988b). Expression is absent from the ante-
rior sister of a9.33, the a9.34 blastomere which is also of
sensory vesicle lineage, suggesting that the onset of Ci-gsx
expression may be coincident with cell fate restriction of
posterior sensory vesicle. When the a9.33 precursor divides,
expression of Ci-gsx is maintained in both sister blasto-
meres, the a10.65 and a10.66 (Fig. 3d,h). This expression
pattern persists and by early tailbud stages, expression is
restricted to two domains bordering the midline (Fig. 3k).
By mid-tailbud stages in embryos co-stained with tyrosi-
nase, a marker for pigment cell precursors, the strongest
expression of Ci-gsx can be seen in the posterior sensory
vesicle bordering the posterior pigment cell (Fig. 3l). We
conclude that at neural plate stages Ci-gsx is a marker of
posterior a4.2-derived sensory vesicle precursors and,
following this lineage, subsequently marks the posterior
sensory vesicle during tailbud stages.
2.3. Expression of Ciona otx
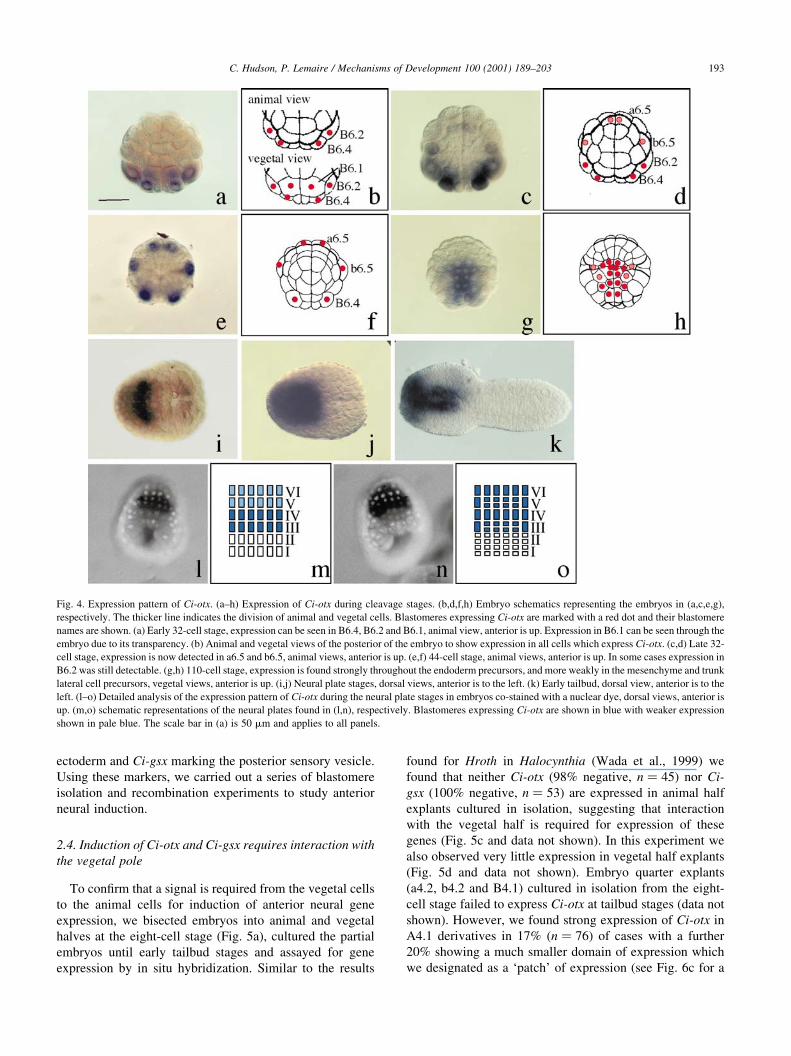
Expression of Ci-otx is essentially the same as described
in Halocynthia with a few differences (Fig. 4 and for a
detailed description in Halocynthia, Wada et al., 1996).
Expression is detected at the early 32-cell stage in two
posterior endoderm blastomeres (B6.1) and the muscle/
mesenchyme/notochord precursors (B6.4, B6.2; Fig. 4a,b).
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 191
Fig. 2. (a) Predicted amino acid sequence of Ci-gsx. An in-frame stop codon was present upstream of the methionine, indicating that the protein sequence is full
length. The homeobox is shown in bold. (b) Sequence comparison between homeoboxes of Ci-gsx, mouse mgsh-1 and -2, medaka olgsh-1, amphioxus
AmphiGsx, and Drosophila ind. (c) Sequence comparisons of the homeobox sequences between Ci-otx, Halocynthia Hroth, Drosophila otd, rat otx2 and
Zebra®sh otx3.

By the late 32-cell stage (Fig. 4c,d), expression begins in
b6.5, the derivatives of which gives rise to epidermis, tail tip
muscle and endoderm and to the most dorsal ependymal cell
of the nerve cord. Expression also appears in a6.5 which
gives rise to the sensory vesicle, palps and pharynx. Forty-
four-cell stage embryos show expression in a6.5, b6.5 and
B6.4 (Fig. 4e,f). In some embryos expression could still be
detected in B6.1 (for example, see Fig. 7o). This is slightly
different to Halocynthia embryos where expression of Hroth
is no longer detectable in B6.4 mesenchyme precursors at
the 44-cell stage (Wada et al., 1996). By the 110-cell stage,
expression of Ci-otx is detected throughout the endoderm,
mesenchyme and trunk lateral cell precursors, but becomes
barely detectable in the neural precursors (Fig. 4g,h). This is
also different from what is observed in Halocynthia and
Herdmania where expression is strongly maintained in the
a4.2-derived neural precursors at the 110-cell stage (Wada
et al., 1996; Hinman and Degnan, 2000). Shortly after
gastrulation commences, Ci-otx is strongly upregulated in
the neural plate precursors (see Fig. 7j) and subsequently
downregulated in the endoderm and mesenchyme. By
neural plate stages, expression is observed as an intense
band across the neural plate (Fig. 4i). As for Ci-gsx, using
double staining with a nuclear dye, we determined that this
site of expression corresponds to the six cells in rows III and
IV of the neural plate, i.e. the sensory vesicle and pharynx
precursors (Nicol and Meinertzhagen, 1988b; Nishida,
1987) (Fig. 4l,m). Subsequently, expression becomes stron-
ger throughout the a4.2-derived neural plate (rows III±VI)
(Fig. 4j,n,o) persisting until tailbud stages when expression
is detected in the forming sensory vesicle and the anterior
epidermal cap (Fig. 4k). Comparison of the expression
patterns of Ci-otx and Ci-gsx indicates that from its onset
of expression, transcripts of Ci-gsx are found within the
most posterior expression domain of Ci-otx.
Therefore, from mid-gastrula stages these two molecules
speci®cally mark the anterior nervous system, with Ci-otx
marking the entire sensory vesicle and part of the anterior
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203192
Fig. 3. Expression pattern of Ci-gsx. (a±d) In situ hybridization of gastrula-neurula stage embryos co-stained with a nuclear dye to identity cell positions. In
each case dorsal views are shown; anterior is up. Cell rows that have just undergone cleavage are marked with an arrow. (e±h) Schematic representation of the
neural plate; anterior is up. Newly cleaved cells are marked with an arrow. For simplicity, the b4.2-derived neural plate and the lateral neural plate cells derived
from A4.1 in rows I and II are omitted. Rows I and II are A4.1 derivatives and rows III±VI are a4.2 derivatives. Cell cleavages were compared to Nicol and
Meinertzhagen (1988b). Cells that express Ci-gsx are coloured blue and cell names are indicated. (i) Lateral view of an early neural plate stage embryo
hybridized with Ci-gsx probe; dorsal is up, anterior is to the left. (j) Schematic representation of the embryo shown in (i); neu.pl., neural plate; noto., notochord;
ecto., ectoderm; endo., endoderm. (k) Early tailbud stage, dorsal view, anterior is to the left. (l) Late tailbud co-stained for tyrosinase (in brown) and Ci-gsx
expression (in blue), dorsal view, anterior is to the left. The scale bar throughout is 50 mm.
ectoderm and Ci-gsx marking the posterior sensory vesicle.
Using these markers, we carried out a series of blastomere
isolation and recombination experiments to study anterior
neural induction.
2.4. Induction of Ci-otx and Ci-gsx requires interaction with
the vegetal pole
To con®rm that a signal is required from the vegetal cells
to the animal cells for induction of anterior neural gene
expression, we bisected embryos into animal and vegetal
halves at the eight-cell stage (Fig. 5a), cultured the partial
embryos until early tailbud stages and assayed for gene
expression by in situ hybridization. Similar to the results
found for Hroth in Halocynthia (Wada et al., 1999) we
found that neither Ci-otx (98% negative, n � 45) nor Ci-
gsx (100% negative, n � 53) are expressed in animal half
explants cultured in isolation, suggesting that interaction
with the vegetal half is required for expression of these
genes (Fig. 5c and data not shown). In this experiment we
also observed very little expression in vegetal half explants
(Fig. 5d and data not shown). Embryo quarter explants
(a4.2, b4.2 and B4.1) cultured in isolation from the eight-
cell stage failed to express Ci-otx at tailbud stages (data not
shown). However, we found strong expression of Ci-otx in
A4.1 derivatives in 17% (n � 76) of cases with a further
20% showing a much smaller domain of expression which
we designated as a `patch' of expression (see Fig. 6c for a
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 193
Fig. 4. Expression pattern of Ci-otx. (a±h) Expression of Ci-otx during cleavage stages. (b,d,f,h) Embryo schematics representing the embryos in (a,c,e,g),
respectively. The thicker line indicates the division of animal and vegetal cells. Blastomeres expressing Ci-otx are marked with a red dot and their blastomere
names are shown. (a) Early 32-cell stage, expression can be seen in B6.4, B6.2 and B6.1, animal view, anterior is up. Expression in B6.1 can be seen through the
embryo due to its transparency. (b) Animal and vegetal views of the posterior of the embryo to show expression in all cells which express Ci-otx. (c,d) Late 32-
cell stage, expression is now detected in a6.5 and b6.5, animal views, anterior is up. (e,f) 44-cell stage, animal views, anterior is up. In some cases expression in
B6.2 was still detectable. (g,h) 110-cell stage, expression is found strongly throughout the endoderm precursors, and more weakly in the mesenchyme and trunk
lateral cell precursors, vegetal views, anterior is up. (i,j) Neural plate stages, dorsal views, anterior is to the left. (k) Early tailbud, dorsal view, anterior is to the
left. (l±o) Detailed analysis of the expression pattern of Ci-otx during the neural plate stages in embryos co-stained with a nuclear dye, dorsal views, anterior is
up. (m,o) schematic representations of the neural plates found in (l,n), respectively. Blastomeres expressing Ci-otx are shown in blue with weaker expression
shown in pale blue. The scale bar in (a) is 50 mm and applies to all panels.

description of the scheme used for scoring expression
domains). In these studies we observed many vegetal half
embryos (n � 355); these showed an average positive score
of 25% and a patch score of 10% (Fig. 6d). Expression in
A4.1 derivatives may represent residual expression in the
anterior endoderm, which is obscured by the neural expres-
sion in whole embryos.
As mentioned above, the earliest expression of Ci-otx in
a4.2 derivatives was detected at the late 32-cell stage. To
determine whether the absence of Ci-otx expression
detected in animal explants at tailbud stages was due to a
lack of initiation of Ci-otx expression or to a lack of main-
tenance of this expression, we bisected embryos into animal
and vegetal halves at the eight-cell stage and assayed at the
44-cell stage for gene expression. No expression of Ci-otx
was observed in the animal explants (94% negative,
n � 101), whereas the expression in the vegetal explants
appeared unaffected (81% positive, n � 121); Fig. 5e±g).
This implies a failure of initiation of Ci-otx expression in
the neural lineage and demonstrates that vegetal to animal
cell signalling is required prior to the 44-cell stage for the
initiation of Ci-otx expression. Similar results were found in
a4.2 or b4.2 explants isolated at the eight-cell stage and
cultured until the 44-cell stage, gastrula stage, and tailbud
stage (see Fig. 7).
These experiments show that cell contacts between
animal and vegetal blastomeres are required for the induc-
tion of anterior neural markers. Analysis at the 44-cell stage
demonstrates that this cell interaction is required for the
early induction of Ci-otx in the anterior neural precursors.
2.5. Anterior neural induction involves a differential
inductive capacity of the vegetal blastomeres and a
difference in the competence of the responding animal
blastomeres
The next question asked was whether restriction of Ci-otx
and Ci-gsx to the anterior neural plate relies on a differential
inducing ability of the vegetal blastomeres or a differential
competence of the animal blastomeres to respond. This was
addressed using a series of blastomere co-isolations and
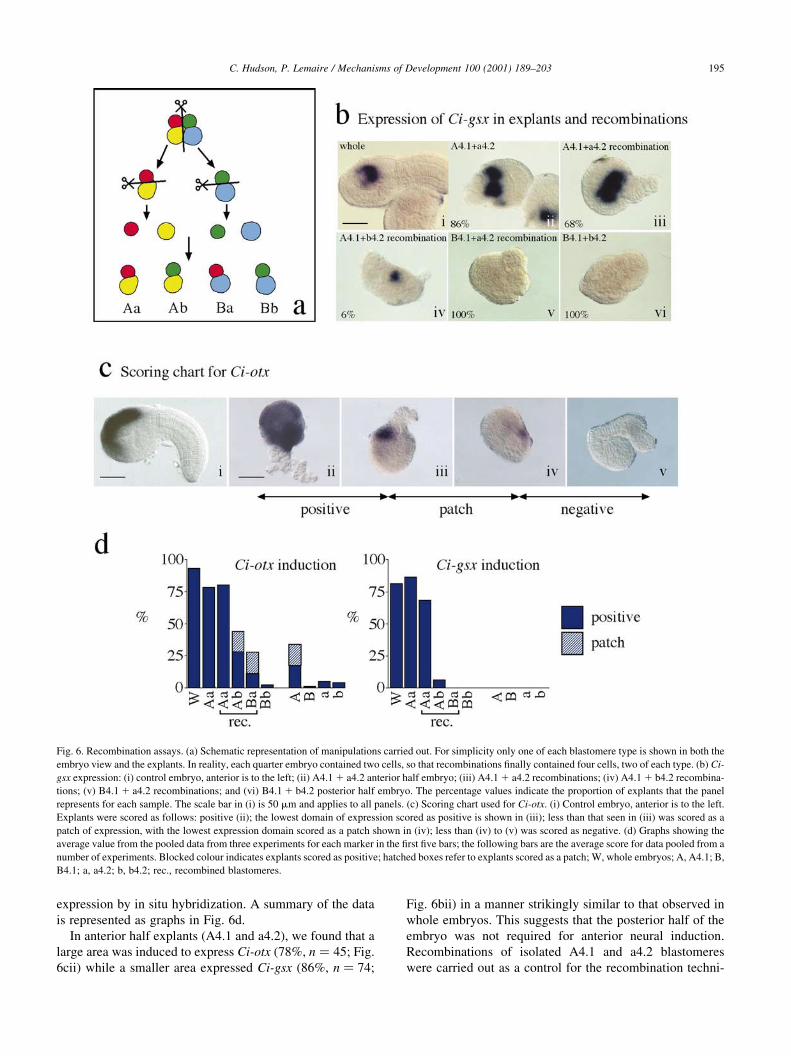
recombinations (Fig. 6a), in which partial embryos were
cultured until the tailbud stage and then assayed for gene
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203194
Fig. 5. (a) Schematic representation of the manipulation carried out: bisection into animal (a4.2 1 b4.2) and vegetal (A4.1 1 B4.1) halves. For simplicity only
one of each blastomere type is shown both for the embryo and explants. In reality, the half embryos contained four cells, two of each type. (b±j) Animal±vegetal
halves, as in (a), assayed for expression of Ci-otx at tailbud stages (b±d), or at the 44-cell stage (e±g). The percentage values indicate the proportion of explants
that the panel represents for each sample. The scale bar in (b) is 50 mm and applies to all panels.
expression by in situ hybridization. A summary of the data
is represented as graphs in Fig. 6d.
In anterior half explants (A4.1 and a4.2), we found that a
large area was induced to express Ci-otx (78%, n � 45; Fig.
6cii) while a smaller area expressed Ci-gsx (86%, n � 74;
Fig. 6bii) in a manner strikingly similar to that observed in
whole embryos. This suggests that the posterior half of the
embryo was not required for anterior neural induction.
Recombinations of isolated A4.1 and a4.2 blastomeres
were carried out as a control for the recombination techni-
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 195
Fig. 6. Recombination assays. (a) Schematic representation of manipulations carried out. For simplicity only one of each blastomere type is shown in both the
embryo view and the explants. In reality, each quarter embryo contained two cells, so that recombinations ®nally contained four cells, two of each type. (b) Ci-
gsx expression: (i) control embryo, anterior is to the left; (ii) A4.1 1 a4.2 anterior half embryo; (iii) A4.1 1 a4.2 recombinations; (iv) A4.1 1 b4.2 recombina-
tions; (v) B4.1 1 a4.2 recombinations; and (vi) B4.1 1 b4.2 posterior half embryo. The percentage values indicate the proportion of explants that the panel
represents for each sample. The scale bar in (i) is 50 mm and applies to all panels. (c) Scoring chart used for Ci-otx. (i) Control embryo, anterior is to the left.
Explants were scored as follows: positive (ii); the lowest domain of expression scored as positive is shown in (iii); less than that seen in (iii) was scored as a
patch of expression, with the lowest expression domain scored as a patch shown in (iv); less than (iv) to (v) was scored as negative. (d) Graphs showing the
average value from the pooled data from three experiments for each marker in the ®rst ®ve bars; the following bars are the average score for data pooled from a
number of experiments. Blocked colour indicates explants scored as positive; hatched boxes refer to explants scored as a patch; W, whole embryos; A, A4.1; B,
B4.1; a, a4.2; b, b4.2; rec., recombined blastomeres.
que and gave essentially the same result as co-isolation, with
high induction of Ci-otx (80%, n � 65) and Ci-gsx (68%,
n � 69; Fig. 6biii), showing that the recombination techni-
que was reliable.
In order to investigate if the animal blastomeres (a4.2 and
b4.2) showed differential ability to respond to the anterior
neural inducers emitted from A4.1 descendants, we recom-
bined A4.1 with b4.2 blastomeres. In this experiment, strong
Ci-otx expression was observed in 28% (n � 86) of cases
with a further 19% showing a small patch of expression
(see Fig. 6c for description of scoring procedure). Taking
into account that around 20% of A4.1 explants cultured
alone expressed Ci-otx, we conclude that b4.2 can only acti-
vate Ci-otx in response to signals from A4.1 descendants in a
small number of cases. Ci-gsx was expressed in 6% (n � 70)
of these recombinations and in these cases the area expres-
sing Ci-gsx was small (Fig. 6biv). The marked reduction in
gene expression in these recombinations compared to
A4.1 1 a4.2 recombinations shows that b4.2 has a much
lower competence than a4.2 to adopt an anterior neural fate
in response to the A4.1-derived inducer, thus supporting and
extending the work of Rose (1939), Reverberi and Minganti
(1946, 1947) and Okado and Takahashi (1990a).
The next question addressed was whether there was a
difference in ability to induce anterior markers between
the two vegetal blastomere descendants (A4.1 and B4.1).
In this set of experiments, we recombined the responding
blastomere a4.2 with B4.1. Ci-otx was strongly induced in
11% (n � 95) of cases with a further 20% showing a patch
of expression. Ci-gsx was never observed in these recombi-
nations (n � 79; Fig. 6bv). We conclude that B4.1 deriva-
tives have some ability to induce anterior markers but much
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203196
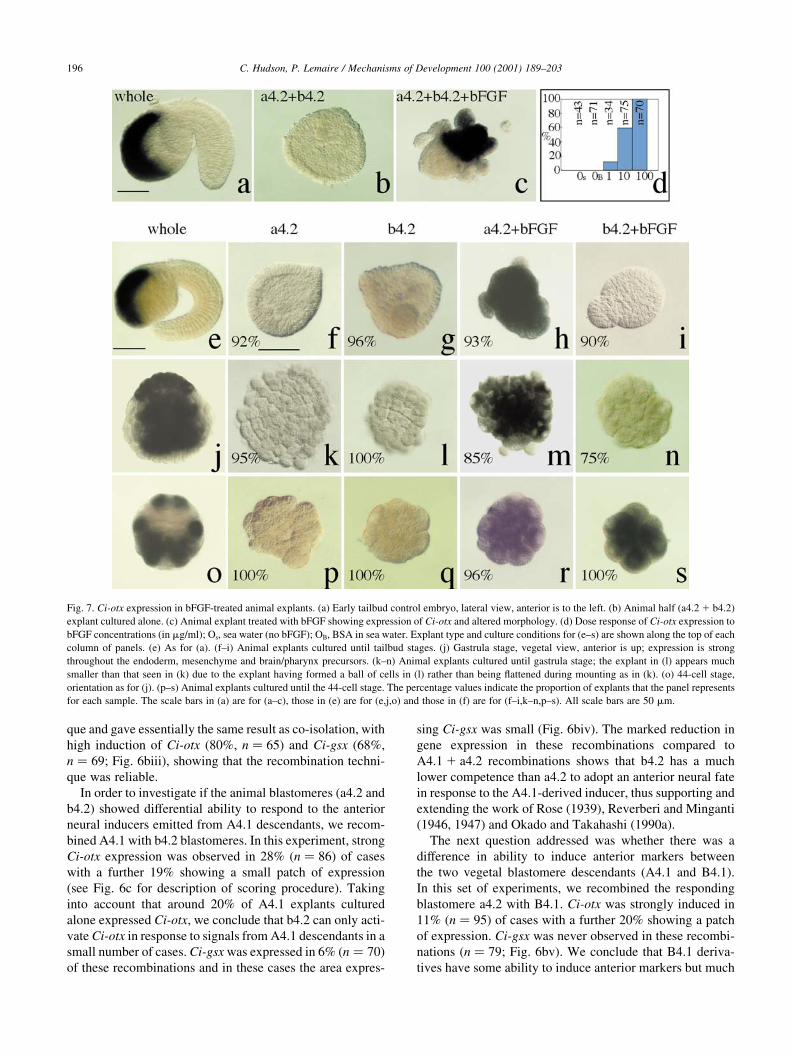
Fig. 7. Ci-otx expression in bFGF-treated animal explants. (a) Early tailbud control embryo, lateral view, anterior is to the left. (b) Animal half (a4.2 1 b4.2)
explant cultured alone. (c) Animal explant treated with bFGF showing expression of Ci-otx and altered morphology. (d) Dose response of Ci-otx expression to
bFGF concentrations (in mg/ml); Os, sea water (no bFGF); OB, BSA in sea water. Explant type and culture conditions for (e±s) are shown along the top of each
column of panels. (e) As for (a). (f±i) Animal explants cultured until tailbud stages. (j) Gastrula stage, vegetal view, anterior is up; expression is strong
throughout the endoderm, mesenchyme and brain/pharynx precursors. (k±n) Animal explants cultured until gastrula stage; the explant in (l) appears much
smaller than that seen in (k) due to the explant having formed a ball of cells in (l) rather than being ¯attened during mounting as in (k). (o) 44-cell stage,
orientation as for (j). (p±s) Animal explants cultured until the 44-cell stage. The percentage values indicate the proportion of explants that the panel represents
for each sample. The scale bars in (a) are for (a±c), those in (e) are for (e,j,o) and those in (f) are for (f±i,k±n,p±s). All scale bars are 50 mm.

less than is found in A4.1. The lack of expression of Ci-gsx
in B4.1 1 a4.2 recombinations may indicate that the induc-
tion of Ci-otx in these explants represents anterior ectoderm,
rather than sensory vesicle. Posterior half (B4.1 and b4.2)
explants showed no expression of Ci-otx (98% negative,
n � 111) or Ci-gsx (100% negative, n � 94; Fig. 6bvi).
These results show that there is a difference in the compe-
tence of the animal blastomeres to respond to anterior neural
inducers as well as a difference in the ability of the vegetal
blastomeres to induce anterior neural markers, and we
propose that both of these factors play an important role
in restricting anterior neural induction to the a4.2 descen-
dants. Our results indicate that the conclusions drawn from
the work of Rose (1939), Reverberi and Minganti (1946,
1947) and Okado and Takahashi (1990a,b) may be generally
applicable to anterior neural induction in ascidians.
2.6. bFGF induces Ci-otx in a4.2, but not b4.2 derivatives
In the recombination experiments described in the
previous section, we analyzed expression of Ci-otx and
Ci-gsx at the tailbud stage. It was therefore not possible to
distinguish between a loss of maintenance of anterior neural
induction in b4.2 derivatives or a lack of induction. This
would require analysis at earlier time points, but the endo-
dermal expression of Ci-otx in A4.1 and B4.1 derivatives
during the cleavage and gastrula stages would render this
experiment dif®cult to interpret. To overcome this dif®culty,
we tested, at different time points after the onset of treat-
ment, the response of a4.2 and b4.2 blastomeres to bFGF, a
growth factor which has been shown to mimic the neural
inducing activity of A4.1 derivatives (Inazawa et al., 1998)
and to be critical for neural development in chick embryos
(Wilson et al., 2000; Streit et al., 2000). To con®rm that
bFGF mimicked the activity of the endogenous inducer,
we ®rst tested whether bFGF could induce expression of
Ci-otx in animal half explants by the tailbud stage. Animal
half explants were isolated at the eight-cell stage and treated
with sea water or various concentrations of bFGF (1, 10 or
100 ng/ml) until the early tailbud stages. Animal explants
cultured to the tailbud stage in sea water alone have a
rounded, disc-shape morphology (Fig. 7b). bFGF-treated
explants appeared more compact and had striking bumps
and protrusions (Fig. 7c). These morphological changes
were accompanied by induction of Ci-otx expression and
were dose-dependent, occurring more frequently with
increasing doses of bFGF (Fig. 7d). Half of the explants
were induced to express Ci-otx at around 10 ng/ml bFGF
and almost 100% expressed Ci-otx after treatment with 100
ng/ml bFGF, a result very similar to that seen in Halo-
cynthia (Inazawa et al., 1998). For all subsequent experi-
ments, unless otherwise stated, 100 ng/ml bFGF was used
because this dose gave the most robust induction of Ci-otx.
As expected, bFGF, like the endogenous inducer, elicited
different responses by the tailbud stage in a4.2 and b4.2.
Treatment with bFGF induced Ci-otx in a4.2 derivatives
(93%, n � 54), whereas b4.2 derivatives showed a much
reduced response (10%, n � 31; Fig. 7e±i). These results
indicate that bFGF can be used as a replacement to the
A4.1-derived inducer to study the regulation of Ci-otx. This
enabled us to look at an earlier stage at the response of a4.2
and b4.2 to bFGF. bFGF-treated explants were ®rst analyzed
at the early gastrula stages. Eighty-®ve percent (n � 46) of
a4.2 explants and 24% (n � 38) of b4.2 explants expressed
Ci-otx (Fig. 8m,n). Expression was not seen in a4.2 explants
(95% negative, n � 40) or b4.2 explants (100% negative,
n � 40) cultured in sea water alone (Fig. 7k,l). To analyze
this further, we looked at the response to bFGF at the 44-cell
stage. We found that incubation in the presence of bFGF
induced expression of Ci-otx in a4.2 (96%, n � 27) and
b4.2 (100%, n � 27) derivatives when assayed at the 44-
cell stage (Fig. 7r,s), whereas isolated a4.2 and b4.2 blasto-
meres cultured in sea water alone showed no expression
(a4.2, 100% negative, n � 26; b4.2, 100% negative,
n � 19; Fig. 7p,q). It appears that both a4.2 and b4.2 animal
blastomere derivatives can respond to bFGF signalling by
expressing Ci-otx at the 44-cell stage, but that by the early
gastrula stage, this expression is only detected in a4.2 deri-
vatives. There are two possibilities to account for the expres-
sion of Ci-otx at the 44-cell stage in b4.2 explants treated with
bFGF; either the blastomeres are partially converted to an
anterior neural fate which they cannot maintain until the early
gastrula stage, or the expression is indicative of the b4.2
derivative which normally expresses Ci-otx at the 44-cell
stage, i.e. the b6.5 blastomere (Figs. 3e,f and 7o). The
absence of a b6.5 marker does not allow us to con®rm
which of these events is occurring. This result suggests,
however, that FGF can act very early (prior to the 44-cell
stage) to induce Ci-otx in the a4.2 and b4.2 lineages.
Although bFGF seems to mimic the endogenous signal in
its ability to induce the expression of Ci-otx, a signi®cant
difference was seen when analyzing Ci-gsx expression. We
found no induction of Ci-gsx in animal half embryos
isolated at the eight-cell stage and incubated in bFGF until
the tailbud stage (10 ng/ml bFGF 0% positive, n � 38; 100
ng/ml 0% positive, n � 19). Ci-gsx was also not observed in
bFGF-treated or untreated a4.2 or b4.2 explants (data not
shown). This indicates that bFGF is able to elicit in animal
cap cells only part of the genetic programme activated by
the signals emitted by the progeny of A4.1, which suggests
the existence of additional inducing molecules.
Importantly, in these experiments we could not distin-
guish between induction of sensory vesicle or anterior ecto-
derm since both tissues express Ci-otx in whole embryos.
This prompted us to characterize in more detail the fate of
bFGF-treated blastomeres.
2.7. Fate of bFGF-treated a4.2 explants
We ®rst looked for neural tissue in bFGF-treated a4.2
blastomere derivatives by looking for the presence of differ-
entiated neurons at swimming larvae stages. To do this, we
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 197
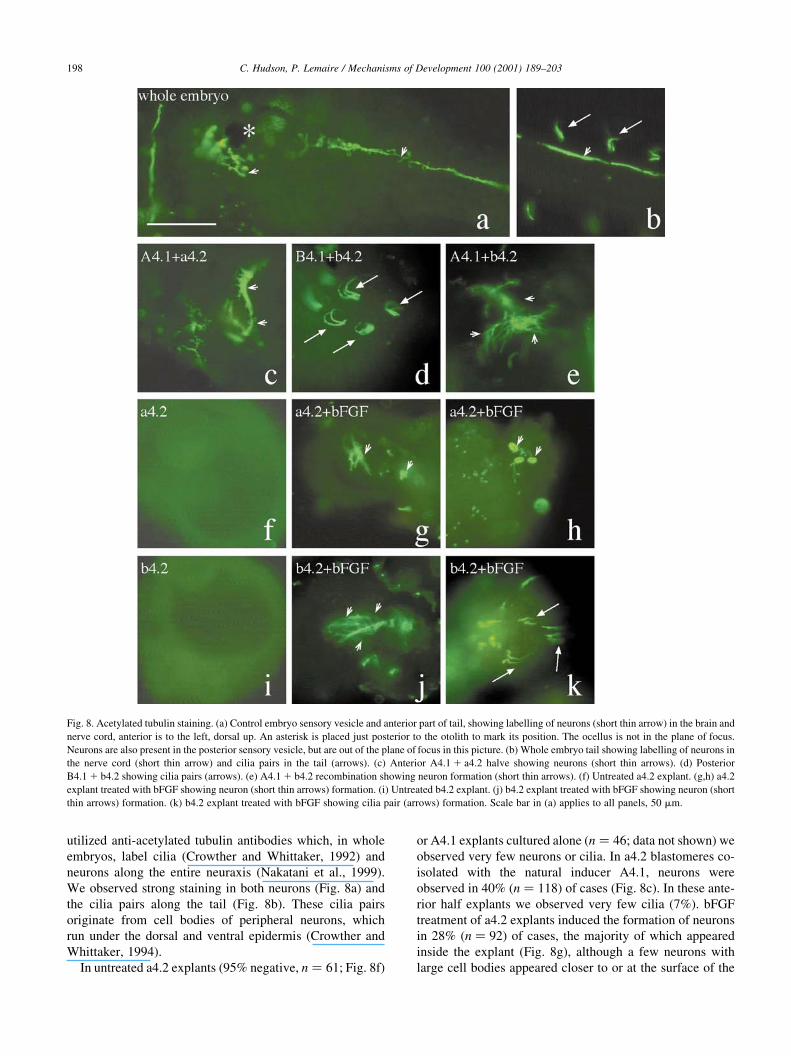
utilized anti-acetylated tubulin antibodies which, in whole
embryos, label cilia (Crowther and Whittaker, 1992) and
neurons along the entire neuraxis (Nakatani et al., 1999).
We observed strong staining in both neurons (Fig. 8a) and
the cilia pairs along the tail (Fig. 8b). These cilia pairs
originate from cell bodies of peripheral neurons, which
run under the dorsal and ventral epidermis (Crowther and
Whittaker, 1994).
In untreated a4.2 explants (95% negative, n � 61; Fig. 8f)
or A4.1 explants cultured alone (n � 46; data not shown) we
observed very few neurons or cilia. In a4.2 blastomeres co-
isolated with the natural inducer A4.1, neurons were
observed in 40% (n � 118) of cases (Fig. 8c). In these ante-
rior half explants we observed very few cilia (7%). bFGF
treatment of a4.2 explants induced the formation of neurons
in 28% (n � 92) of cases, the majority of which appeared
inside the explant (Fig. 8g), although a few neurons with
large cell bodies appeared closer to or at the surface of the
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203198
Fig. 8. Acetylated tubulin staining. (a) Control embryo sensory vesicle and anterior part of tail, showing labelling of neurons (short thin arrow) in the brain and
nerve cord, anterior is to the left, dorsal up. An asterisk is placed just posterior to the otolith to mark its position. The ocellus is not in the plane of focus.
Neurons are also present in the posterior sensory vesicle, but are out of the plane of focus in this picture. (b) Whole embryo tail showing labelling of neurons in
the nerve cord (short thin arrow) and cilia pairs in the tail (arrows). (c) Anterior A4.1 1 a4.2 halve showing neurons (short thin arrows). (d) Posterior
B4.1 1 b4.2 showing cilia pairs (arrows). (e) A4.1 1 b4.2 recombination showing neuron formation (short thin arrows). (f) Untreated a4.2 explant. (g,h) a4.2
explant treated with bFGF showing neuron (short thin arrows) formation. (i) Untreated b4.2 explant. (j) b4.2 explant treated with bFGF showing neuron (short
thin arrows) formation. (k) b4.2 explant treated with bFGF showing cilia pair (arrows) formation. Scale bar in (a) applies to all panels, 50 mm.

explant and may represent a different type of neuron (Fig.
8h). We also observed induction of cilia in a small number
of cases (18%). Neurons formed only rarely in B4.1 1 a4.2
recombinations (8%, n � 153), although cilia formed in
30% of cases (data not shown). Neither neurons nor cilia
formed B4.1 explants cultured alone (n � 66; data not
shown).
These results show that bFGF can induce neurons in a4.2
albeit to a lesser extent than the endogenous A4.1-derived
inducer. a4.2 can also respond in a number of cases to B4.1
and bFGF by producing cilia.
The use of acetylated beta tubulin antibodies did not
allow us to distinguish between the formation of neurons
from the central or peripheral nervous systems. To test for
the presence of sensory vesicle-derived tissue in the bFGF-
treated explants, we looked in treated blastomeres for the
formation of pigment cells, a late differentiation marker for
the sensory vesicle lineage, which originates from the same
anterior-posterior position as Ci-gsx during neural plate
stages. Pigment cells did not form in untreated isolated
a4.2 blastomeres cultured to the swimming larval stage
(0%, n � 71; Fig. 9d). However, bFGF treatment induced
pigment cells in 36% of a4.2 explants (one to four pigment
cells per explant, n � 137; Fig. 9f), similar to the frequency
observed in A4.1-a4.2 explants (35%, one to three pigment
cells, n � 60; Fig. 9b). The majority of pigment cells which
formed in these experiments contained large pigment gran-
ules, suggesting that they are of the otolith type. Recombi-
nations of a4.2 with B4.1 led to induction of pigment cells in
only 4% (n � 153) of cases.
Taken together, these results show that bFGF-treated a4.2
blastomeres do indeed contain sensory vesicle tissue,
suggesting that while bFGF cannot fully replace A4.1 as
an anterior neural inducer, the induction of Ci-otx by
bFGF does not solely re¯ect induction of anterior ectoderm.
Additionally, these results further show that similar to the
results found for Ci-otx and Ci-gsx, B4.1 is a much less
competent inducer of neurons and pigment cells in a4.2
derivatives than A4.1.
2.8. Fate of bFGF-treated b4.2 explants
In the majority of cases involving b4.2 as the responding
blastomere, we observed no induction of Ci-otx or Ci-gsx.
Therefore, we looked to see if b4.2 explants could respond
to bFGF or A4.1 and produce some neural tissue. In both
uninduced a4.2 and b4.2 explants epidermis differentiation
was seen as judged by the presence of the tunic (Fig. 9d,e,
arrows). However, in either a4.2 or b4.2 explants treated
with bFGF, the surface of the whole or part of the explant
was devoid of tunic (Fig. 9f,g), suggesting that in b4.2
explants as in a4.2 explants, bFGF may suppress epidermal
differentiation.
Analysis of acetylated tubulin in b4.2 gave unexpected
results. While neurons did not form in untreated b4.2
explants (97% negative, n � 99; Fig. 8i), recombinations
with A4.1 induced neuronal formation in 40% (n � 129;
Fig. 8e) of recombinations. Posterior (B4.1 1 b4.2) explants
possessed neurons in fewer cases (11%, n � 123) and these
were generally signi®cantly less numerous than those seen
in A4.1 1 b4.2 blastomere recombinations. Explants treated
with bFGF showed neuronal differentiation in 62% (n � 6;
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 199
Fig. 9. Pigment cell formation at hatching/swimming tadpole stage. (a) Control embryo with two pigment cells, ocellus and otolith, dorsal is up, anterior is to
the left. (b) A4.1 1 a4.2 anterior explant with two otolith-type pigment cells. (c) B4.1 1 b4.2 posterior explant. (d) Untreated a4.2 explant. (e) Untreated b4.2
explant. (f) a4.2 treated with bFGF containing three otolith-type pigment cells. (g) b4.2 treated with bFGF. Arrows indicate the presence of tunic. Arrowheads
mark the border between the tunic positive and tunic negative parts of the explant. The scale bar in (a) is 50 mm and applies to all panels.

Fig. 8j) of cases. Both the proportion of explants induced to
form neurons and the numbers of neurons per positive
explants were higher than that found in bFGF-treated a4.2
explants.
In posterior explants, cilia pairs formed (63%, n � 123;
Fig. 8d). Recombinations of A4.1 1 b4.2 could also
produce cilia, although less frequently (24%, n � 129).
We also observed induction of cilia in b4.2 by bFGF
(68%, n � 66; Fig. 8k). We observed very few cases of
pigment cell formation in A4.1 1 b4.2 recombinations or
bFGF-treated b4.2 (less than 4% in both cases, data not
shown) and never in B4.1 1 b4.2 posterior explants.
Taken together, these data show that b4.2 is competent to
respond to bFGF and A4.1 and adopt a neural fate. As judged
by the lack of Ci-otx, Ci-gsx and pigment cell formation in
b4.2, the neural tissue induced is not anterior in character.
While B4.1 does not induce high levels of Ci-gsx, Ci-otx,
pigment cells or neurons in either a4.2 or b4.2, B4.1 appears
to induce peripheral neurons, as characterized by cilia, in
both animal blastomeres at higher levels than A4.1. These
results also show that b4.2 is more competent to produce cilia
in response to B4.1 or bFGF than is a4.2.
3. Discussion
3.1. Expression patterns of Ci-otx and Ci-gsx
Here, we report the cloning and expression patterns of
two sensory vesicle markers in the ascidian, Ciona intesti-
nalis. Ci-otx is expressed dynamically during early cleavage
and is restricted to a4.2-derived neural precursors from mid-
gastrula stages. The expression pattern of Ci-otx in endo-
derm and anterior neural precursors appears somewhat simi-
lar to that reported in vertebrates. In mouse, otx2 is
expressed in the endoderm which is critical for anterior
neural induction (Rhinn et al., 1998) and subsequently in
the fore- and midbrain (Simeone, 1998). A detailed compar-
ison between ascidian and vertebrate otx expression from
gastrula stages can be found in Wada et al. (1996).
Ci-gsx is expressed in a4.2-derived posterior sensory
vesicle precursors from early neurula to tailbud stages
within the posterior expression domain of Ci-otx. In
mouse and medaka, gsh genes are expressed in two bands
of cells running along the anterior-posterior axis of the
central nervous system located in the intermediate neuro-
genic region, with additional expression in speci®c regions
of the fore-, mid- and hindbrain (Hsieh-Li et al., 1995;
Szucsik et al., 1997; Valerius et al., 1995; Deschet et al.,
1998). In Drosophila, ind is expressed in lateral columns of
the ventral nerve cord, in a pattern remarkably similar to
that described for the mouse and medaka Gsh genes (Weiss
et al., 1998; Arendt and Nubler-Jung, 1999). In contrast,
amphioxus was found to have a more limited expression
domain, being restricted to the cerebral vesicle, the equiva-
lent of the vertebrate fore- and midbrain (Brooke et al.,
1998). We ®nd that expression of Ci-gsx is more reminis-
cent of the expression seen in amphioxus, being restricted to
the sensory vesicle (Brooke et al., 1998). Expression of Ci-
gsx appears much earlier in development than its counter-
parts in other chordate species.
3.2. Anterior neural induction in ascidians
Early studies showed that both a4.2 and b4.2 blastomeres
could respond to neural inducers producing pigment cells and
neuron-type membrane excitability, and that b4.2 had a lower
competence to respond than a4.2 (Rose, 1939; Reverberi and
Minganti, 1946, 1947; Okado and Takahashi, 1990a).
However, since these studies looked at differentiation of
speci®c cell types, it was not clear whether the reduced
competence of b4.2 was due to reduced general neural induc-
tion, or if the induced blastomeres lacked the ability to
produce these speci®c neural cell types. Additionally, in clea-
vage-arrested embryos, only one cell type can be adopted and
therefore a partial neural conversion would not be detected.
In this study we looked again at neural induction using
Ci-otx and Ci-gsx which enabled us to look at earlier steps of
neural tissue formation and to focus on anterior neural fates.
In addition, our analysis did not require cleavage arrest and
therefore we could analyze neural induction at a more
physiological level.
Using a series of blastomere co-isolations and recombi-
nations we found that high levels of anterior neural induc-
tion occurred only when both the anterior vegetal and
anterior animal blastomeres were recombined. Anterior
and posterior vegetal cells have a different inductive capa-
city while anterior and posterior animal cells show a differ-
ent competence to respond. Furthermore, we found that a4.2
and b4.2 blastomeres also have a strikingly different compe-
tence to respond to bFGF and that this difference in compe-
tence can be detected as early as the gastrula stage.
Although it seems that only a4.2 can adopt an anterior
neural fate, our experiments with A4.1 1 b4.2 and bFGF-
treated b4.2 suggest that the difference in competence
between a4.2 and b4.2 is not the result of an inability of
b4.2 to respond to neural inducing signals: the morphology
of bFGF-treated b4.2 explants is altered, tunic formation is
decreased and these explants produce many neurons and
cilia. The precise identity of the neurons produced in
response to A4.1 and bFGF demands a much more detailed
neuronal fate map and a collection of neuronal markers
speci®c to each lineage. Very little is known about the
formation of peripheral neurons in ascidians. We were
able to show that cilia pairs, which mark a speci®c type of
peripheral neuron (Crowther and Whittaker, 1994), could be
induced by B4.1 much more readily than by A4.1. Similarly,
b4.2 was more competent to produce cilia in response to
B4.1 and bFGF than a4.2.
3.3. Basis for a difference in competence
It is likely that the difference in competence to form
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203200

anterior neural tissue between a4.2 and b4.2 descendants is
established maternally, since it is already apparent when
blastomeres are isolated at the eight-cell stage. After ferti-
lization, ascidian eggs undergo extensive cytoplasmic rear-
rangement, termed ooplasmic segregation, resulting in the
localization of a region of cytoplasm termed the myoplasm
or posterior cytoplasm to a sub-equatorial position in the
egg, the future posterior pole. This ooplasmic segregation
is responsible for the correct positioning of the embryonic
anterior-posterior axis, for positioning of endoderm, muscle
and ectoderm lineages, and also for the establishment of the
correct early cleavage asymmetries (Nishida, 1992, 1993,
1994a,b). It has been proposed that posterior fate is domi-
nant over anterior and is determined by maternal determi-
nants localized in the posterior cytoplasm. The default
anterior fate would only occur in the absence of these deter-
minants (Nishida, 1994b). This early segregation event may
also be responsible for the difference in competence
between the animal blastomeres in terms of anterior neural
induction. One could imagine that the posterior cytoplasm
inherited predominantly by B4.1 is also present in b4.2 and
confers upon this cell a different responsiveness to neural
inducers than a4.2. In embryos in which the posterior cyto-
plasm is removed at the one-cell stage, the embryo develops
in a radially symmetrical, anteriorized fashion (Nishida,
1994b). If the posterior vegetal cytoplasm is then fused
into the anterior vegetal side, the polarity of the subsequent
embryos is reversed. It would be very interesting to deter-
mine in normal embryos whether b4.2 cells inherit some
posterior vegetal cytoplasm, and whether this is responsible
for the difference in competence that we see across the
animal hemisphere.
A differential competence of animal blastomeres to
respond to neural inducing signals is also observed in verte-
brates. In Xenopus, dorsal (future neural plate) adopts a
neural fate much more readily than ventral (future epider-
mis) ectoderm (Sharpe et al., 1987; Sokol and Melton, 1991;
Otte and Moon, 1992). Recent evidence suggests that a
maternal event, implicating the stabilization of b-catenin,
enhances the competence of the dorsal ectoderm in Xenopus
embryos to become neural, the subsequent induction of
neural tissue relying on zygotic molecules secreted from
the organizer (Baker et al., 1999; Harland, 2000). However,
in Xenopus, this difference in competence is between neural
and epidermal fate, whereas in ascidians our results suggest
that this difference appears to be between anterior and
posterior neural fates. Additionally, in Xenopus, the differ-
ence in competence can be overridden and ventral ectoderm
can be converted to a neural fate. In ascidians, as we show
here, b4.2 explants very rarely adopt an anterior neural fate.
In ascidians, cell fates are determined in embryos with very
few cells, whereas in Xenopus cell fates are determined in
embryos with many more cells. One could imagine that if a
diffusible inducer was responsible for neural induction, in
embryos with a small cell number the competence to
respond would have to be much more tightly regulated
than in much larger embryos with many cells and a highly
regulative mode of development.
3.4. Role of bFGF in neural induction
It has been proposed that bFGF may be the natural neural
inducer in ascidians (Inazawa et al., 1998). The time
window of action and the response of animal blastomeres
to bFGF are similar to those observed with the natural indu-
cer (Inazawa et al., 1998). We con®rmed that in Ciona,
bFGF also has inducing activity and used this as a tool to
further investigate the induction of neural markers. The
early induction of Ci-otx in a4.2 and b4.2 explants by the
44-cell stage suggests that bFGF is acting directly to induce
Ci-otx. The lack of mesoderm (notochord and muscle) and
endoderm in these explants (data not shown) lends support
to this, although we cannot exclude the possibility that a
tissue for which we have no marker as yet is induced
which secondarily induces or maintains neural markers.
a4.2 blastomeres respond to bFGF by expressing Ci-otx
and developing pigment cells at levels comparable to induc-
tion by A4.1. However, high concentrations of bFGF were
required to obtain the same level of Ci-otx induction as that
seen in response to the natural inducer. It is very dif®cult to
imagine that this concentration of bFGF could be achieved
in an embryo, although it is possible that high levels are only
required because recombinant human bFGF used in this
study may be less potent than the endogenous FGF. Testing
this hypothesis will require the cloning and analysis of FGF
molecules in ascidians. Additionally, although a4.2 blasto-
meres responded similarly to bFGF and the endogenous
inducer, in terms of expression of Ci-otx and pigment
cells formation, the much reduced number of neurons
produced and the lack of induction of Ci-gsx in response
to bFGF indicates that even high concentrations of bFGF
cannot fully replace A4.1 as an anterior neural inducer.
Moreover, a recent analysis of the ability of A4.1 and
B4.1 line blastomeres to induce notochord or mesenchyme,
two FGF inducible cell fates, has led to the proposition that
an FGF-like signal is emitted by both A4.1 and B4.1 deri-
vatives (Kim et al., 2000). Our ®ndings that anterior and
posterior vegetal blastomeres have very dissimilar neural
inducing ability lends support to the proposition that FGF
signalling is not the sole requirement for induction of ante-
rior neural fates.
4. Experimental procedures
4.1. Embryos
Adults were obtained from the Station de Biologie
Marine in Roscoff and from the Saumaty industrial harbour
in Marseille. Eggs and sperm were collected by dissection
and cross fertilizations were made in petri dishes. Dechor-
ionated embryos (according to Mita-Miyazawa et al., 1985)
were raised in sea water at 13±188C on 1% agarose-coated
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 201

dishes in arti®cial sea water (420 mM NaCl, 90 mM KCl, 10
mM CaCl2, 24.5 mM MgCl2, 25.5 mM MgSO4, 2.15 mM
NaHCO3, 10 mM Hepes (pH 8.0)) supplemented with 50
mg/ml gentamycin. Explants were dissected with a ®ne glass
needle. Recombinations were carried out by placing blasto-
mere pairs in contact, in 30% PEG(8000) and then washing
in sea water.
bFGF (Sigma, F0291) was diluted in 0.1% BSA (bovine
serum albumin, fraction V, Sigma, A8022) in sea water.
Explants were placed in bFGF solution until the required
stage and ®xed.
4.2. Cloning of Ci-gsx and Ci-otx
Ci-gsx and Ci-otx were ampli®ed from mid-gastrula
embryo cDNA using the degenerate antennapedia class
primers previously described (Moretti et al., 1994) with a
508C annealing temperature and 2 mM MgCl2. Full-length
cDNAs were obtained by screening a tailbud cDNA library
using standard procedures. Sequencing was done by
Genome Express, Genoble. The Genbank database acces-
sion numbers are AF305499 (Ci-otx) and AF305500 (Ci-
gsx).
4.3. In situ hybridization
In situs were carried out as described (Wada et al., 1995).
For Fig. 3 embryos were collected every 15 min from early
gastrula (4.5 h at 188C) until the tailbud began to elongate (8
h). Nuclei were observed using Syto11 or 13 Live Cell
Nucleic Acid Stain (Molecular probes). For photography,
all embryos were mounted in 50±80% glycerol in PBS-
Tween. Often this led the embryos to appear rather bigger
than in unmounted specimens. Embryos co-stained with a
nuclear dye and those in Figs. 3i,k, 4e,g and 9b±e,g were
photographed prior to mounting.
4.4. Staining with anti-acetylated tubulin
Anti-acetylated tubulin (Sigma T6793) antibody staining
was carried out as follows. Brie¯y, embryos or explants
were ®xed in 2208C methanol for 10 min and washed
three times for 10 min in PBS-Triton X (0.05%). Primary
antibody was used at 1:1000 in PBS-Tween (0.1%) and
incubated overnight at 48C. Samples were washed for at
least 10 min three times in PBS-Tween at room temperature,
and then secondary antibody (AlexaFluore 488 goat anti-
mouse IgG, Molecular probes, used at 1:100 in PBS-Tween)
was incubated overnight at 48C. Finally, samples were
washed at least three times for 10 min in PBS-Tween and
mounted in 80% glycerol.
This antibody also stained another structure in explants
on the surface of both a4.2 and b4.2 explants treated with
bFGF or combined with A4.1 (data not shown), which was
not seen in similarly high numbers in whole embryos. These
structures were shorter than the neurons and cilia and may
represent a different kind or partially formed cilia or sensory
neurons. Since we could not de®ne these structures as either
neuron-like or cilia-like, neurons were scored as positive
only if they were present in a group of neurons. Cilia
were scored only if they were present as a pair or if they
could be seen projecting into the tunic.
Note added in proof
Ciona-otx has been independently cloned and reported:
Nanami Utsumi and Hidetoshi Saiga (2001). Comparison of
the Structure and Expression of otx Genes Between Ciona
intestinalis and Halocynthia roretzi. In: Biology of Asci-
dians, subtitle: Proceedings of the First International
Symposium on the Biology of Ascidians, Sappora, Japan
June 26±30, 2000, Springer±Verlag, Tokyo.
Acknowledgements
A big thank you to H. Yasuo for all his help when we
were starting to establish ascidian work in the lab. Many
thanks to A. Spagnuoli for the generous gift of the cDNA
library. We would also like to thank O. Cabaud, J.-P.
Bonamy, S. Marcellini and A. Ribas for their input into
the Ciona culture system. Many thanks to N. Utsumi, H.
Saiga, S. Darras and H. Nishida for discussions. We also
thank K. Dale, L. Kodjabachian, M. Maroto, M. McGrew
and H. Yasuo for their helpful comments on the manuscript.
This work was supported by grants to P.L. from the Centre
National de la Recherche Scienti®que (CNRS), the Associa-
tion pour la Recherche contre le Cancer, the Association
FrancËaise contre les Myopathies, the Human Frontier
Science Program Organisation, and the European Union
(TMR programme). C.H. was a EU-TMR fellow and then
the recipient of a CNRS Poste Rouge.
References
Arendt, D., Nubler-Jung, K., 1999. Comparison of early nerve cord devel-
opment in insects and vertebrates. Development 126, 2309±2325.
Baker, J.C., Beddington, R.S.P., Harland, R.M., 1999. Wnt signalling in
Xenopus embryos inhibits Bmp4 expression and activates neural devel-
opment. Genes Dev. 13, 3149±3159.
Brooke, N.M., Garcia-Fernandez, J., Holland, P.W., 1998. The ParaHox
gene cluster is an evolutionary sister of the Hox gene cluster. Nature
392, 920±922.
Conklin, E.G., 1905. The organisation and cell lineage in the ascidian egg.
J. Acad. Natl. Sci. (Philadelphia) 13, 1±119.
Crowther, R.J., Whittaker, J.R., 1992. Structure of the caudal neural tube in
an ascidian larva: vestiges of its possible evolutionary origin from a
ciliated band. J. Neurobiol. 23, 280±292.
Crowther, R.J., Whittaker, J.R., 1994. Serial repetition of cilia pairs along
the tail surface of an ascidian larva. J. Exp. Zool. 268, 9±16.
Deschet, K., Bourrat, F., Chourrout, D., Joly, J.S., 1998. Expression
domains of the medaka (Oryzias latipes) Ol-Gsh 1 gene are reminiscent
of those of clustered and orphan homeobox genes. Dev. Genes Evol.
208, 235±244.
Di Gregorio, A., Spagnuoli, A., Ristoratore, F., Pischetola, M., Aniello, F.,
Branno, M., Cariello, L., Di Lauro, R., 1995. Cloning of ascidian
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203202

homeobox genes provides evidence for a primordial chordate cluster.
Gene 156, 253±257.
Finkelstein, R., Boncinelli, E., 1994. From ¯y head to mammalian fore-
brain: the story of otd and Otx. Trends Genet. 10, 310±315.
Harland, R., 2000. Neural induction. Curr. Opin. Genes Dev. 10, 357±362.
Hinman, V.F., Degnan, B.M., 2000. Retinoic acid perturbs Otx gene expres-
sion in the ascidian pharynx. Dev. Genes Evol. 210, 129±139.
Hsieh-Li, H.M., Witte, D.P., Szucsik, J.C., Weinstein, M., Li, H., Potter,
S.S., 1995. Gsh-2, a murine homeobox gene expressed in the develop-
ing brain. Mech. Dev. 50, 177±186.
Inazawa, T., Okamura, Y., Takahashi, K., 1998. Basic ®broblast growth
factor induction of neuronal ion channel expression in ascidian ecto-
dermal blastomeres. J. Physiol. 511.2, 347±359.
Kim, G.J., Yamada, A., Nishida, H., 2000. An FGF signal from endoderm
and localized factors in the posterior-vegetal egg cytoplasm pattern the
mesodermal tissues in the ascidian embryo. Development 127, 2853±
2862.
Mita-Miyazawa, I., Ikegami, S., Satoh, N., 1985. Histospeci®c acetylcho-
linesterase development in the presumptive muscle cells isolated from
16-cell-stage ascidian embryos with respect to the number of DNA
replications. J. Embryol. Exp. Morphol. 87, 1±12.
Moretti, P., Simmons, P., Thomas, P., Haylock, D., Rathjen, P., Vadas, M.,
D'Andrea, R., 1994. Identi®cation of homeobox genes expressed in
human haemopoietic progenitor cells. Gene 144, 213±219.
Nakatani, Y., Moody, R., Smith, W.C., 1999. Mutations affecting tail and
notochord development in the ascidian Ciona savignyi. Development
126, 3293±3301.
Nicol, D., Meinertzhagen, I.A., 1988a. Development of the central nervous
system of the larva of the ascidian, Ciona intestinalis L. I. The early
lineages of the neural plate. Dev. Biol. 130, 721±736.
Nicol, D., Meinertzhagen, I.A., 1988b. Development of the central nervous
system of the larva of the ascidian, Ciona intestinalis L. II. Neural plate
morphogenesis and cell lineages during neurulation. Dev. Biol. 130,
737±766.
Nicol, D., Meinertzhagen, I.A., 1991. Cell counts and maps in the larval
central nervous system of the ascidian Ciona intestinalis (L.). Comp.
Neurol. 309, 415±429.
Nishida, H., 1987. Cell lineage analysis in ascidian embryos by intracellular
injection of a tracer enzyme. III. Up to the tissue restricted stage. Dev.
Biol. 121, 526±541.
Nishida, H., 1992. Regionality of egg cytoplasm that promotes muscle
differentiation in embryo of the ascidian, Halocynthia roretzi. Devel-
opment 116, 521±529.
Nishida, H., 1993. Localized regions of egg cytoplasm that promote expres-
sion of endoderm-speci®c alkaline phosphatase in embryos of the asci-
dian Halocynthia roretzi. Development 118, 1±7.
Nishida, H., 1994a. Localization of egg cytoplasm that promotes differen-
tiation to epidermis in embryos of the ascidian Halocynthia roretzi.
Development 120, 235±243.
Nishida, H., 1994b. Localization of determinants for formation of the ante-
rior-posterior axis in eggs of the ascidian Halocynthia roretzi. Devel-
opment 120, 3093±3104.
Nishida, H., 1997. Cell fate speci®cation by localized cytoplasmic determi-
nants and cell interactions in ascidian embryos. Int. Rev. Cytol. 176,
245±306.
Okado, H., Takahashi, K., 1990a. Induced neural-type differentiation in the
cleavage-arrested blastomere isolated from early ascidian embryos. J.
Physiol. 427, 603±623.
Okado, H., Takahashi, K., 1990b. Differentiation of membrane excitability
in isolated cleavage-arrested blastomeres from early ascidian embryos.
J. Physiol. 427, 583±602.
Okado, H., Takahashi, K., 1993. Neural differentiation in cleavage arrested
ascidian blastomeres induced by proteolytic enzyme. J. Physiol. 463,
269±290.
Ortolani, G., Patricolo, E., Mansueto, C., 1979. Trypsin-induced cell
surface changes in ascidian embryonic cells. Exp. Cell Res. 122,
137±147.
Otte, A.P., Moon, R.T., 1992. Protein kinase C isozymes have distinct roles
in neural induction and competence in Xenopus. Cell 68, 1021±1029.
Reverberi, G., Minganti, A., 1946. Fenomeni di evocazione nello sviluppo
dell'uovo di Ascidie. Riulti dell'indagine sperimentale sull'uovo di
Ascidiella aspersa e do Ascidia malaca allo stadio di otto blastomeri.
Pubbl. Stn. Zool. Napoli 20, 199±252.
Reverberi, G., Minganti, A., 1947. La distribuzione delle potenze nel germe
di Ascidie allo stadio di otto blastomeri, analizzata mediante le combi-
nazioni e i trapianti di blastomeri. Pubbl. Stn. Zool. Napoli 21, 1±35.
Rhinn, M., Dierich, A., Shawlot, W., Behringer, R.R., Le Meur, M., Ang,
S.-L., 1998. Sequential roles for Otx2 in visceral endoderm and
neuroectoderm for forebrain and midbrain induction and speci®cation.
Development 125, 845±856.
Rose, S.M., 1939. Embryonic induction in the ascidia. Biol. Bull. 76, 216±
232.
Satoh, N., 1994. Developmental Biology of Ascidians, Cambridge Univer-
sity Press, Cambridge.
Sharpe, C.R., Fritz, A., De Robertis, E.M., Gurdon, J.B., 1987. A homeo-
box-containing marker of posterior neural differentiation shows the
importance of predetermination in neural induction. Cell 50, 749±758.
Simeone, A., 1998. Otx1 and Otx2 in the development and evolution of the
mammalian brain. EMBO J. 17, 6790±6798.
Singh, G., Kaur, S., Stock, J.L., Jenkins, N.A., Gilbert, D.J., Copeland,
N.G., Potter, S.S., 1991. Identi®cation of 10 murine homeobox genes.
Proc. Natl. Acad. Sci. USA 88, 10706±10710.
Sokol, S., Melton, D.A., 1991. Pre-existent pattern in Xenopus animal pole
cells revealed by induction with activin. Nature 351, 409±411.
Streit, A., Berliner, A.J., Papanayotou, C., Sirulnik, A., Stern, C.D., 2000.
Initiation of neural induction by FGF signalling before gastrulation.
Nature 406, 74±78.
Szucsik, J.C., Witte, D.P., Li, H., Pixley, S.K., Small, K.M., Potter, S.S.,
1997. Altered forebrain and hindbrain development in mice mutant for
the Gsh-2 homeobox gene. Dev. Biol. 191, 230±242.
Takahashi, H., Mitani, Y., Satoh, G., Satoh, N., 1999. Evolutionary altera-
tions of the minimal promoter for notochord-speci®c Brachyury expres-
sion in ascidian embryos. Development 126, 3725±3734.
Valerius, M.T., Li, H., Stock, J.L., Weinstein, M., Kaur, S., Singh, G.,
Potter, S.S., 1995. Gsh-1: a novel murine homeobox gene expressed
in the central nervous system. Dev. Dyn. 203, 337±351.
Wada, H., Saiga, H., Satoh, N., Holland, P.W., 1998. Tripartite organisation
of the ancestral chordate brain and the antiquity of placodes: insights
from ascidian Pax-2/5/8, Hox and Otx genes. Development 125, 1113±
1122.
Wada, S., Katsuyama, Y., Yasugi, S., Saiga, H., 1995. Spatially and tempo-
rally regulated expression of the LIM class homeobox gene Hrlim
suggests multiple distinct functions in development of the ascidian,
Halocynthia roretzi. Mech. Dev. 51, 115±126.
Wada, S., Katsuyama, Y., Sato, Y., Itoh, C., Saiga, H., 1996. Hroth, an
orthodenticle-related homeobox gene of the ascidian, Halocynthia
roretzi: its expression and putative roles in the axis formation during
embryogenesis. Mech. Dev. 60, 59±71.
Wada, S., Katsuyama, Y., Saiga, H., 1999. Anteroposterior patterning of the
epidermis by inductive in¯uences from the vegetal hemisphere cells in
the ascidian embryo. Development 126, 4955±4963.
Weiss, J.B., Von Ohlen, T., Mellerick, D.M., Dressler, G., Doe, C.Q., Scott,
M.P., 1998. Dorsoventral patterning in the Drosophila central nervous
system: the intermediate neuroblasts defective homeobox gene speci-
®es intermediate column identity. Genes Dev. 12, 3591±3602.
Wilson, S.I., Graziano, E., Harland, R., Jessell, T.M., Edlund, T., 2000. An
early requirement for FGF signalling in the acquisition of neural cell
fate in the chick embryo. Curr. Biol. 10, 421±429.
C. Hudson, P. Lemaire / Mechanisms of Development 100 (2001) 189±203 203
Copyright © 2022 FDOKUMEN




















