
Individual prey specialization in wasps: predator size is a weak predictor of taxonomic niche width...
21
Transworld Research Network 37/661 (2), Fort P.O. Trivandrum-695 023 Kerala, India Predation in the Hymenoptera: An Evolutionary Perspective, 2011: 101-121 ISBN: 978-81-7895-530-8 Editor: Carlo Polidori 5. Individual prey specialization in wasps: Predator size is a weak predictor of taxonomic niche width and niche overlap Carlo Polidori 1 , Davide Santoro 1 , Josep Daniel Asís 2 and José Tormos 2 1 Dipartimento di Biologia, Sezione di Zoologia e Citologia, Università degli Studi di Milano via Celoria 26, 20133 Milano, Italy; 2 Unidad de Zoología, Facultad de Biología Universidad de Salamanca 37071 Salamanca, Spain Abstract. Intuitively, larger predators in a population may be favoured in subduing, handling and carrying to the nest a wider range of prey compared to smaller ones. In absence of any individual chemical or behavioural bias in prey choice, and independently from size-biased preference, it may be predicted: (1) that smaller wasps would have both narrower niches and lower niche overlap than larger wasps, (2) a positive correlation between individual niche width and niche overlap, and (3) a positive correlation between predator size and prey size. We tested these predictions using available data in literature on individual prey spectrum for 10 populations/generations of apoid wasps (Hymenoptera: Crabronidae). The analysis showed that individual specialization is widespread. Smaller individuals were more specialized than larger ones, as we hypothesized, in only one population, while the opposite was found in two cases; on the other side, niche width correlated (positively) with wasp size only in one population. In five out of the 10 examined cases predator size Correspondence/Reprint request: Dr. Carlo Polidori, Dipartimento di Biologia, Sezione di Zoologia e Citologia, Università degli Studi di Milano, via Celoria 26, 20133 Milano, Italy E-mail: [email protected]
Transcript of Individual prey specialization in wasps: predator size is a weak predictor of taxonomic niche width...

Transworld Research Network 37/661 (2), Fort P.O. Trivandrum-695 023 Kerala, India
Predation in the Hymenoptera: An Evolutionary Perspective, 2011: 101-121 ISBN: 978-81-7895-530-8
Editor: Carlo Polidori
5. Individual prey specialization in wasps: Predator size is a weak predictor of
taxonomic niche width and niche overlap
Carlo Polidori1, Davide Santoro1, Josep Daniel Asís2 and José Tormos2 1Dipartimento di Biologia, Sezione di Zoologia e Citologia, Università degli Studi di Milano
via Celoria 26, 20133 Milano, Italy; 2Unidad de Zoología, Facultad de Biología Universidad de Salamanca 37071 Salamanca, Spain
Abstract. Intuitively, larger predators in a population may be favoured in subduing, handling and carrying to the nest a wider range of prey compared to smaller ones. In absence of any individual chemical or behavioural bias in prey choice, and independently from size-biased preference, it may be predicted: (1) that smaller wasps would have both narrower niches and lower niche overlap than larger wasps, (2) a positive correlation between individual niche width and niche overlap, and (3) a positive correlation between predator size and prey size. We tested these predictions using available data in literature on individual prey spectrum for 10 populations/generations of apoid wasps (Hymenoptera: Crabronidae). The analysis showed that individual specialization is widespread. Smaller individuals were more specialized than larger ones, as we hypothesized, in only one population, while the opposite was found in two cases; on the other side, niche width correlated (positively) with wasp size only in one population. In five out of the 10 examined cases predator size
Correspondence/Reprint request: Dr. Carlo Polidori, Dipartimento di Biologia, Sezione di Zoologia e Citologia, Università degli Studi di Milano, via Celoria 26, 20133 Milano, Italy E-mail: [email protected]

Carlo Polidori et al. 102
correlated with prey size. Overall, the predictions were all true for just one population. In conclusion, different factors probably influence often more than predator size the dynamics of individual prey specialization in wasps. Circumstantial evidences suggest that individual-biased exploitation of different hunting areas, some behavioural traits of prey, and different ratios between prey and wasp size may partially account for the observed patterns. Introduction Despite a lot of published models of predator-prey dynamics assume that conspecific individuals are identical and that they use the same resources [e.g. 1, 2], it is now clear that many apparently generalist species are composed of individual specialists that use small subsets of the population’s niche [e.g. 3, 4, 5]. Such individual specialization may even exceed differences between conventional species [6, 7], and may have critical ecological and evolutionary implications [8]. For example, individual specialization has been thought to reduce intraspecific competition [9, 10, 11]. Individual specialization is generally related to constraints on an individual’s ability to efficiently exploit a wide variety of resources. Constraints generally arise from functional trade-offs in which consumers efficiently exploiting one type of resource are inefficient using another type of resource [8]. In central-place foragers, like predatory wasps, for example, the size of a predator may prevent the use of those prey taxa which are too large to be successfully subdued, handled, and carried to the nest [12]. However, most studies attempting to explain foraging behaviour in animals have assumed that there is no size variation among individuals within populations (for dissenting views, see [13, 14]), despite variation in individual body size can be large within populations [15]. Concerning Hymenoptera, only recently a central role of size dynamics was recognized in explaining host selection behaviour [e.g. 16]. Intuitively, larger individuals in a given population may be capable of using a broader range of prey species than smaller ones, which would be forced to hunt only small prey. Such pattern commonly occurs in other animal taxa and could be seen even through individuals’ growth: for example, many marine fish predators undergo considerable increases in body size during their development, resulting in ontogenetic niche shift [17]. In wasps, such prediction is tentatively supported by recent studies. For example, in the digger wasp Cerceris arenaria L., a weevil-hunting specialist, larger females hunt both small and large weevils, while smaller females hunt only small ones (thus including in their diet a lower number of prey species) [18]. Such a study, however, based this suggestion on a

Individual prey specialization versus size in wasps 103
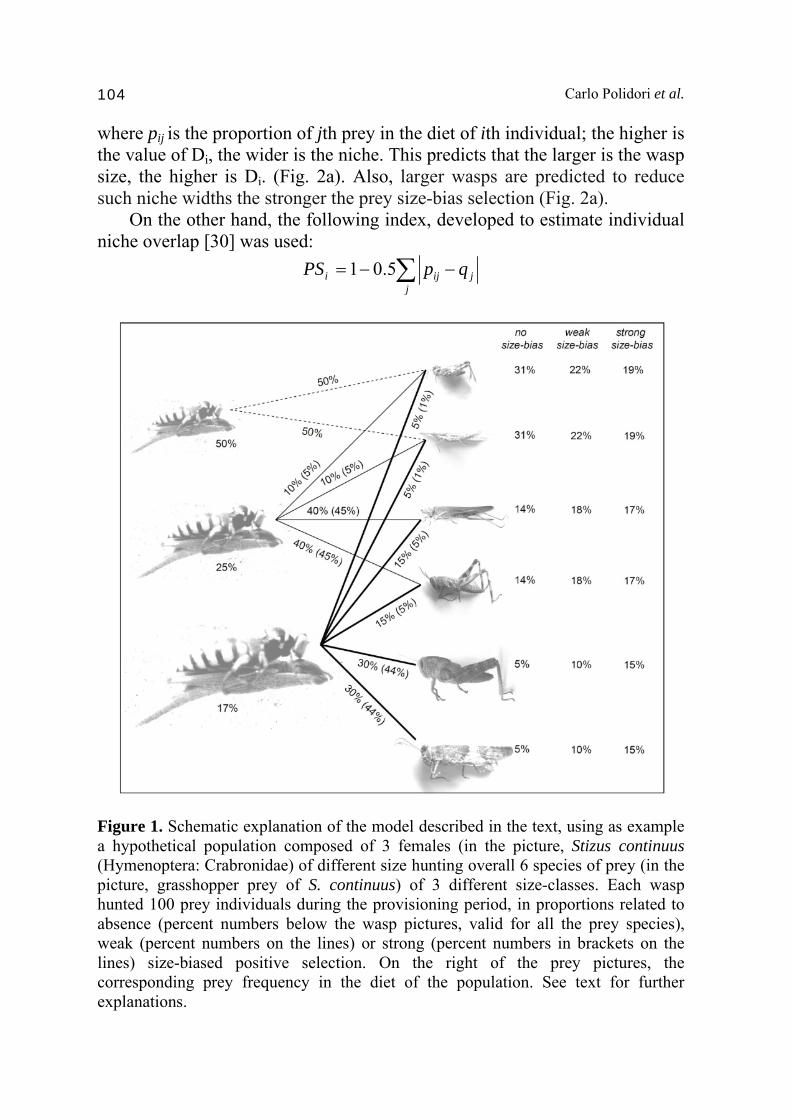
measure, prey species richness, which actually does not identify individual specialization [e.g. 11]. Moreover, larger C. arenaria females tend to prefer larger prey [18]; this positive correlation between the size of the predators and that of their prey has been repeatedly (but not always, see e.g. [19]) found also in various other species of wasps [e.g. 20, 21, 22, 23, 24, 25, 26], and seems to be ubiquitous throughout natural systems [e.g. 27, 28]. In wasps, even in monophagous species, larger females tend to hunt for larger individuals of the unique prey species [29]. Thus, if predator size shapes the prey spectrum at the individual level, one may expect intra-specific, size-dependent variation of the prey niche among individuals (in non-monophagous species). In this chapter, we contrasted available data on individual prey spectrum of selected species of apoid wasps (Hymenoptera: Crabronidae) with these predictions, starting from a simple model of size-niche relationship. Assumptions and predictions of a simple size-niche model Consider a population with n wasps of different size (1/3 small, 1/3 medium-sized, and 1/3 large) hunt on the whole on m prey species (1/3 small, 1/3 medium-sized and 1/3 large). The individual selection may be mediated by wasp size at different degrees (from absence of prey size bias to strong preference of larger wasps for larger prey). As assumptions, encounter rate is identical for all females hunting on any of the prey species, and prey species have identical abundances and availabilities in the environment. In absence of any further chemical or behavioural (e.g. learning) bias in individual prey selection, the small wasps would hunt only the small prey (1/3m), medium-sized wasps both small and medium-sized prey (2/3m), and large wasps all the prey species, in proportions related to the degree of prey size preference (or due to constraints) (Fig. 1). Consider now two concepts related to individual niche: niche width and niche overlap. In the first case, specialists are individuals with foraging niches narrower than the population’s niche, while in the second case specialists have niches that exhibit little overlap with the population niche [30]. These two variables related to niche do not have the same meaning. For example, the use of a rare dietary resource or having little dietary overlap with the population may not reflect niche width [31]. In our simple model, the following broadly used index, developed to estimate individual niche width [32] was used:
∑=
jij
i pD 2
1
Carlo Polidori et al. 104
where pij is the proportion of jth prey in the diet of ith individual; the higher is the value of Di, the wider is the niche. This predicts that the larger is the wasp size, the higher is Di. (Fig. 2a). Also, larger wasps are predicted to reduce such niche widths the stronger the prey size-bias selection (Fig. 2a). On the other hand, the following index, developed to estimate individual niche overlap [30] was used:
∑ −−=j
jiji qpPS 5.01
Figure 1. Schematic explanation of the model described in the text, using as example a hypothetical population composed of 3 females (in the picture, Stizus continuus (Hymenoptera: Crabronidae) of different size hunting overall 6 species of prey (in the picture, grasshopper prey of S. continuus) of 3 different size-classes. Each wasp hunted 100 prey individuals during the provisioning period, in proportions related to absence (percent numbers below the wasp pictures, valid for all the prey species), weak (percent numbers on the lines) or strong (percent numbers in brackets on the lines) size-biased positive selection. On the right of the prey pictures, the corresponding prey frequency in the diet of the population. See text for further explanations.

Individual prey specialization versus size in wasps 105
where pij describes the proportion of the jth resource category in individual i’s diet and qj is the proportion of the jth resource category in the population’s niche; PSi varies from 0 (maximum individual specialization) to 1 (no individual specialization). This predicts higher PSi for larger wasps (Fig. 2c). Also, smaller wasps would be more specialized the stronger the prey size-bias selection (Fig. 2c).
Figure 2. Values of niche width (Di) (a), niche overlap (Psi) (b), their relationship (c), and predator-prey size relationship (d) resulting from the model of Fig. 1. In such simple model, the smallest female would have the narrowest niche width (lowest Di) (Fig. 2a) and it would be the more specialized (lowest PSi) (Fig. 2b). This would produce a strongly positive correlation between PSi and Di (Fig. 2c), which would be strongest in absence of individual size-biased selection. Also, an increase of individual size-biased selection would produce a stronger correlation between wasp size and prey size (Fig. 2d); this would decrease Di for larger females and decrease PSi for smaller females. See text for further explanations.

Carlo Polidori et al. 106
This situation will also produce automatically a positive correlation between PSi and Di, strongest in absence of individual size-biased selection (Fig. 2b), and a positive correlation between predator and prey size, even in absence of further size-biased selection (for medium and large wasps), (Fig. 2d). A first consequence of such simple model is that, in case females were all large enough to hunt for any prey species (i.e. very low variance in wasp size) and/or the prey have all the same exploitable size for all wasps, niche width and niche overlap would be very similar or even identical for all wasps (thus absence of individual specialization). A second consequence is that populations with lower variance in females’ size should have higher individual mean niche overlap (IS = mean PSi) when compared to populations composed of individuals with strong size variation. The population-level niche width (D) was calculated, as follows [32]:
∑=
jjq
D 2
1 (p and j as above)
Note that, contrary to IS, D can not be easily compared across populations, because its value is strongly dependent on prey richness [30, 32]. Hence, we compared among populations the ratio Di/D (referred in the following as “relative niche”), which represents how much the individual niche width is narrower than the population niche width. The four predictions of our model are thus summarized as follow: (1) smaller wasps would have narrower taxonomic prey niches (lower Di) than larger wasps, (2) smaller wasps would have lower taxonomic prey overlap (lower PSi) than larger wasps, (3) there would be a positive correlation between niche width and niche overlap per female wasp, and (4) there would be a positive correlation between wasp size and prey size. Data sample and statistical analysis We selected from the literature those works that have information both on prey taxa used by different individuals of a given wasp species and a measure of size of individual wasps and their prey (body length, body weight, thorax width, or head width), in order to verify the four predictions listed above (Fig. 2). In some cases, such information was not explicitly available in these articles, but in these cases we had the raw data. Six works were selected, encompassing the following species: Cerceris arenaria [18], Cerceris rubida Jurine [24], Cerceris californica Cresson [20] Stizus continuus (Klug) [19, 33], Oxybelus lamellatus Olivier [34] and Bembix

Individual prey specialization versus size in wasps 107
merceti Parker [35]. We also used unpublished information on further samples of C. rubida (Polidori, unpublished data) and on one additional species (Bembix sinuata Panzer; Asís et al., unpublished data). All of these species, members of the family Crabronidae, share a number of important traits of their biology: they all provision subterranean nests with previously paralysed nymphs or adults of different insect taxa to feed their developing offspring. For two of these species, we obtained data for more than 1 generation and/or year; these data were analysed separately, because of the common fluctuations across time and space in the prey use by apoid wasps [33,36,37]. Thus, on the whole, data were available for 10 populations/ generations (Table 1). Because the indices used in the analysis may become quite inaccurate with very small sample sizes, only females associated with at least 4 prey items each were considered. The value of D indicates how much the niche is wide and the value of IS if prey spectra of individuals significantly overlap. To test the significance of the IS-values, we used the IndSpec1 software [30], which uses a nonparametric Monte Carlo procedure to generate replicate null diet matrices drawn from the population distribution, from which P values can be computed [30]. We used 10,000 replicates in Monte Carlo bootstrap simulations to obtain each P value. The calculated indices (Di and PSi) were plotted against the size of individual wasps and plotted one against the other, and predator size was plotted against prey size, to search for linear regressions. Pearson correlation test (for n>10; correlation coefficient: r) and Spearman correlation test (for n<10; correlation coefficient: ρ) were employed to quantify the degree to which such variables are related. To avoid pseudo-replication problems, predator-prey size correlations were performed associating with each female the mean size of their prey. Wasp size variance of a population, calculated as the coefficient of variation (CV=SD/mean), was plotted against IS and Di/D across populations. Note that, for some populations, sample size for predator-prey size relationships does not always correspond to sample size used for analysis of taxonomic specialization (depending on how much individuals were associated to either prey spectrum or prey size). In the text and tables, mean values are reported ± standard deviation. Results and discussion In the Results section, species are grouped by their prey type (insect order). For each species, we give the results of the analyses and try to find out possible factors in the foraging biology that may account for the observed patterns.

Carlo Polidori et al. 108
T
able
1. L
ist o
f po
pula
tions
/gen
erat
ions
sel
ecte
d fr
om th
e lit
erat
ure
(plu
s un
publ
ishe
d da
ta)
and
used
for
the
anal
ysis
, with
in
form
atio
n on
sam
ple
size
s. *
Sam
ple
size
s re
fers
to
the
indi
ces’
cal
cula
tions
and
rel
ated
ana
lyse
s; f
or w
asp-
prey
siz
e re
latio
nshi
ps, i
n so
me
case
s sam
ple
was
diff
eren
t (se
e te
xt a
nd T
able
2).

Individual prey specialization versus size in wasps 109
Tab
le 2
. Pre
y sp
ecie
s ric
hnes
s (R
), ni
che
over
lap
(IS)
, nic
he w
idth
(D),
and
aver
age
indi
vidu
al n
iche
wid
th (D
i) fo
r the
10
pop
ulat
ions
/gen
erat
ions
use
d in
the
ana
lysi
s, to
geth
er w
ith t
he r
esul
ts o
f lin
ear
corr
elat
ion
test
s (in
bol
d th
e si
gnifi
cant
tes
ts).
R=p
rey
spec
ies
richn
ess.
Size
exp
ress
ed a
s: 1 bo
dy w
eigh
t (m
g), 2 he
ad w
idth
(m
m),
3 body
len
gth
(mm
). N
iche
ove
rlap
sign
ifica
nce:
*P<
0.05
, **
P<0.
01,
***P
<0.0
01.
Hor
izon
tal
lines
mar
k th
e th
ree
grou
ps o
f po
pula
tions
/gen
erat
ions
whi
ch h
unt f
or th
ree
diff
eren
t ins
ect o
rder
s (O
rthop
tera
, Col
eopt
era,
Dip
tera
).

Carlo Polidori et al. 110
Orthoptera-hunting wasps Stizus continuus is a Mediterranean solitary wasp whose populations are bivoltine in Southern Europe (whole adult activity from early June to end of September) [38]; females dig multicellular nests in large aggregations, and hunt grasshoppers, mainly in the family Acrididae, to feed their immature offspring [39, 19]. The data used for this analysis come from three generations (the first one of 2005 and both those of 2007) at a single nesting site (Table 1). This was placed in the area of “Mallada Llarga”, located in “Dehesa del Saler”, part of the “Parque Natural de la Albufera” (Valencia, Spain). Two main microhabitats characterized the nesting site: a bared sandy soil, covered by a crust of salt (typical during the hottest months) and small and thick bushes composed by Salicornia ramosissima (J. Woods) and Sarcocornia fruticosa (L.). The individual prey specialization of this species varied through generations: diet overlap between individuals was significantly reduced and lower in the two generations of 2007 than in that of 2005 (Table 2). In this way, prey species richness was greatly lower (only 2 species hunted, with one accounting for >90% of prey specimens) in 2005 than in both 2007 generations, which showed similar values (although wasps of the two years hunted for different taxa) (Table 2). On the contrary, the niche width (D) and the average Di were more similar between the first generation of 2007 and 2005 than between the two generations of the same year; in particular, niche was widest in the second generation of 2007 (Table 2). Supporting our predictions, more specialized females (lower PSi) had narrower niches (lower Di) in the second generation of 2007. However, they had wider niches in 2005, disagreeing to predictions (Table 2). Also contrary to predictions, size of females did not correlate with individual niche overlap and niche width in both 2005 and the second generation of 2007, and the larger females had smaller diet overlap in the first generation of 2007 (Table 2). Moreover, the relationship between wasp and prey size varied through generations partially agreeing with our predictions, as it was significantly positive in both the two generations of 2007 (though in the first one the result was driven by an outlier, see [33]) (agreeing), but did not so in 2005 (disagreeing; Table 2 and Fig. 3). In this species, thus, size of females only partially, and sometimes not in the direction suggested by our model, seems to affect the degree of intra-specific diet variation. The observed patterns may be due to the factors that influence the overall prey preference at population level and probably also to the much lower number of prey species hunted in 2005 than in 2007. In 2005, S. continuus

Individual prey specialization versus size in wasps 111
primarily hunted for grasshoppers of the genus Heteracris, revealing, at least in the period of the study and in this area, this wasp to be almost monophagous. In contrast, grasshoppers of the genus Acrotylus were ignored by the wasps, in spite of their high abundance in the environment. Polidori et al. [19] found evidence that this bias occurred because Acrotylus is found more often on the soil and on grasses, while Heteracris is nearly only found on Sarcocornia bushes, which probably represents the habitat mostly exploited by the wasps for hunting. Very few females hunted for non-Heteracris prey, maybe because searching for prey on different microhabitats is not common. Also in 2007 S. continuus expressed selective predation, females hunting basically on species encountered at large bushes [33]. For example, the largely hunted Calliptamus (in the first generation) and Tropidopola (in the second generation) live mostly on bushes, whether the usually ignored Acrotylus and Sphingonotus lived preferentially on the ground or on short grass [33]. Once again, prey ecology largely accounts for the observed prey preferences. In the end of the 2007 summer, some prey species were excluded from the diet because they became too large to be carried in flight, so that variation in selectivity across wasp generations was probably weakly related to change in prey community composition [33]. Intraspecific niche variation in S. continuus seems thus a result of complex interactions between ecological prey preference and size constraints. In particular, a possible individual-based learning of hunting sites (bushes) would have stronger effect on shaping individual specialization than the female size. Moreover, in this wasp, the greater variance in size of the prey collected by larger females (that is, weak size-biased selection) may account for the absence of wasp size-prey size correlation in 2005; in this year, this would have been an additional factor that reduced the size-niche correlation, thus following our model. Interestingly, the ratio between prey and wasp size vary considerably among generations, being 0.47±0.23 in the first generation of 2007 and 0.88±0.24 in the second one (comparisons could not be done with 2005, because, in that year, only lengths of prey were available). This may force to a stronger individual prey specialization in the second generation and a stronger correlation between predator and prey size. At last, S. continuus females from both the first 2007 generation and the 2005 generation showed larger variation in size than those from the second generation of 2007 (Fig. 4). According to our model, we would expect an according variation in overlap among individuals (IS). This was not true: wasps, in fact, were not specialized in 2005, but exhibit strong specialization in 2007 (Table 2). A parallel situation was found for the relative niche width (Fig. 4). These results may be explained, at least in part, by distinct environmental abundances of potential prey in the two study years, roughly

Carlo Polidori et al. 112
reflected in the diet spectrum of the wasp [19, 33]. In 2005, the great variation in size among females could not reflect the great variation in individual diets, since the vast majority of potential prey was constituted by only one genus (Heteracris) and, possibly, by one single species (H. littoralis). On the other hand, in the first generation of 2007, the great variation in size among females could reflect the greater variation in individual diets. The discordance found in the second generation, however, could not be explained with this hypothesis. Coleoptera-hunting wasps Cerceris arenaria is a Palearctic solitary digger wasp which nests in large and dense aggregations [40, 41]. It is a weevil-hunting specialist [37]. The studied nesting site was situated in the Regional Park of Adda Sud (Lombardy, northern Italy), which is mainly composed of cultivated fields (maize, soybean, and tomatoes). The nest aggregation of C. arenaria was located in a farm of a small town. At this site, females were active from the middle of June to the end of July [18]. Nine of the most abundant species recorded as prey for the marked females studied in 1999, belong to the genera Sitona and Otiorhynchus. Diet overlap between individuals was small, while niche width was around 3.6 for the population (D) and 2, on average, per individual (Di) (Table 2). There was no correlation between PSi and Di (Table 2). On the other hand, size of females negatively correlated with individual niche overlap, but did not with the niche width (Table 2). Thus, in this species, contrary to predictions of the model, the larger females were those more individually specialized (lower PSi). This parallels to the fact that the relationship between wasp and prey size was strong (Table 2 and Fig. 3). One possible explanation for this pattern may invoke the correlation between the size of prey and that of predator [18]. Such correlation is so strong that the larger wasps eventually discarded small prey species and prey mainly (or only) upon large prey, further increasing their specialization. Such situation resembles that found for S. continuus of the second generation of 2007, when larger wasps had lower diet overlap and predator-prey size relationship was very strong. Moreover, for 7 small wasps, all prey items were Sitona species. Such a genus was well represented in overall prey spectrum, so that, not being a rare resource, it does not increase specialization in such females. In addition, several evidences suggest that sometimes wasps exploit target hunting sites (e.g. trees) only for brief periods (from 1 day to 5 days), although some prey species were detected from wasps incoming to the nests for the whole study period (about 1.5 months) [18]. This may be a result

Individual prey specialization versus size in wasps 113
of temporary fidelity to particular, restricted hunting sites, and may be facilitated by the fact that Sitona and Otiorhynchus are associated with quite different plant hosts (basically grasses and trees/bushes, respectively). At last, the C. arenaria population showed the highest size variation among the study wasp populations (Fig. 4). Thus, according to our model, we expect low overlap among individuals. This holds true, with IS for C. arenaria population being the fourth lowest among the wasp populations (Table 2). Also the relative niche width (0.55) was moderately low (Fig. 4). Cerceris rubida is a small digger wasp confined to southern Europe, and its biology is remarkable in that it is the only European species of the genus known to be social [42]. Females dig nests in compact soils and fill them with previously paralysed small beetles as food for the larvae. This species is of interest to study diet preference dynamics because, in contrast to most Cerceris species, it is one of the few species that hunt for many beetle families (Chrysomelidae, Curculionidae, Nitidulidae and Phalacridae) [24]. Data for this species come from the Maremma Regional Park (Grosseto Province, Tuscany, Central Italy), from the middle of June to August (summer of 2005) [24]. In addition, unpublished data come from the same population studied in 2006. The Park is characterised by the Uccellina Mountains (a chain of hills parallel to the coast and covered by the thick Mediterranean maquis), Mediterranean pine-woods and extended cultivated fields (in particular maize, tomatoes, olives, and sunflowers). The nesting area of C. rubida consisted of a trail bounded by two cultivated fields (alfalfa Medicago sativa and wheat Triticum sp.). At this site, the wasp population hunted for phytophagous beetles mainly belonging to families Chrysomelidae, Curculionidae, and Phalacridae that were common around the wasp nesting site and that altogether represented almost the 95% of all the hunted beetles [24]. Diet overlap between individuals was small in both years, niche width (D) was about 1.6 and 4.6, and the average Di were about 1.3 and 2.3 in the two years (Table 2). There was no correlation between PSi and Di in any year (Table 2). Size of females neither correlated with the individual niche overlap nor with the niche width in any year (Table 2). The relationship between wasp and prey size was absent in 2006 but present in 2005 (Table 2 and Fig. 3). As to what found in S. continuus, and probably differently from C. arenaria, prey of C. rubida seemed to be hunted almost only in one specific environment (cultivated fields), thus suggesting that habitat accounted for most of the observed diet. However, inside these hunting sites, a further stronger selection appeared: beetle species differed markedly in their probability to be hunted, also after controlling for prey body size and prey body shape [24]. Chaetocnema scheffleri (Kutschera) (Chrysomelidae) had

Carlo Polidori et al. 114
the major risk to be hunted, followed by Chaetocnema tibialis (Illiger). Other chrysomelid beetles were not hunted, though very abundant in such areas. Thus, rather than an ecology-based selection (as in S. continuus), a taxon-based specialization at population-level is responsible for the observed pattern of prey preference, suggesting that in this species a restricted sensory window, that would exclude some beetle species from the diet, shapes prey choice. At last, C. rubida varied in size greatly in 2005 and moderately in 2006 (Fig. 4). According to our model, we thus expect lower overlap in 2005 and higher overlap in 2006. This seems to be true when comparing such values with those referred to other wasp populations (Table 2). A parallel situation was found for the relative niche width (Fig. 4). In particular, the very low overlap and the narrow relative niche in 2005 may be due to the same reason suggested for C. arenaria: larger females, which showed strong preference for larger prey, may discard smaller prey species; however, because, differently from C. arenaria, C. rubida females probably hunt all in the same hunting site, the segregation in the diet of small and large females would be less pronounced (i.e. PSi does not correlate negatively with size). Cerceris californica is a North American digger wasp nesting in dense aggregations [20]. The data analysed here come from a work carried out at a nesting site placed in the San Gabriel Mountains of Southern California. At that site, wasps provisioned their nests with buprestid beetles, mainly in the genus Acmaeodera. The authors noted that species of this genus are strongly associated with plants of the genera Adenostoma (Rosaceae) and Ceanothus (Rhamnaceae), so that they inferred that these plants are the main hunting sites for C. californica females. Thus, as in C. rubida and S. continuus, the ecology and feeding habits of prey species probably account for the population’s diet preference. Diet overlap between individuals was small, while niche width (D) and the average Di were around 6.7 and 3.8, respectively (Table 2). A positive correlation appeared between PSi and Di (Table 2). In addition, size of females positively correlated both with individual niche overlap and with niche width (Table 2). Thus, in this species, according to predictions of the model, smaller females were those more individually specialized (lower PSi and lower Di). Also the relationship between wasp and prey size was positive and very strong (Table 2 and Fig. 3). At last, C. californica varied moderately in size in the studied population (Fig. 4). According to our model, we thus expect moderate overlap among individuals, and this seems to be true, giving that IS, although still very significant, was the sixth highest recorded (Table 2). Also the relative niche

Individual prey specialization versus size in wasps 115
Figure 3. Predator-prey size relationships for the 10 analysed populations. * body length for prey of C. rubida and S. continuus, pro-mesothorax junction width for prey of C. arenaria, elytra length for prey of C. californica, and head width for prey of B. sinuata; ** body length for C. californica and head width for all the other species. Trend lines are shown only for significant linear correlations. For population codes, see Table 1. width was moderately high (Fig. 4). Therefore, this population almost perfectly agrees with all the predictions of our model. Diptera-hunting wasps Oxybelus lamellatus is a solitary digger wasp distributed throughout the western part of the Mediterranean area. The data analysed here come from a study carried out at a nesting aggregation located in an area of compact sandy soil with little vegetation, surrounded by Arundo donax L., Juncus acutus L., J. maritimus Lamk., Phragmites communis Trin., and Thymelaea hirsuta (L.). This area was located some 100 m from the Mediterranean shoreline in the natural park called El Marjal del Moro, in the municipality of Sagunto (Valencia, Spain).

Carlo Polidori et al. 116
The prey consisted in males and females of Diptera belonging to 9 families, with Pollenia rudis (Fabricius) (Calliphoridae), Fannia scalaris (Fabricius) (Fannidae) and Neomyia comicina (Fabricius) (Muscidae) the most hunted species. Diet overlap between individuals was small and niche (D and Di) quite narrow (Table 2). Thus, despite O. lamellatus was considered opportunistic in regard to the type of prey [34], our analysis highlights that individual females are strongly specialized in prey choice. No correlation appeared between PSi and Di (Table 2), and size of females did not correlate with niche overlap and niche width (Table 2). Predator and prey sizes are not correlated (Table 2 and Fig. 3). Thus, in this species, female size does not seem to affect the intra-specific diet overlap. One of the possible reasons behind this pattern may consider the characteristic behaviour of prey carriage in this species: in fact, pedal carriage (prey hold with legs) was used for lighter prey, whereas abdominal carriage (prey impaled on the sting) was used for those of greater weight. For mid-sized prey, both types of carriage were observed. Thus, all individual wasps (regarding to their size, which, in any case, did not vary too much, see below) may have the possibility to carry to the nest prey of a wide range of sizes (prey weight/wasp weight from 0.6 to 2.4), shifting if this is particularly large from pedal to abdominal carriage. The individual niche width and the niche overlap, however, remain small due to some unknown individual behavioural bias in prey selection. At last, O. lamellatus was the species with lowest variation in size among the analysed populations (Fig. 4). According to our model, we thus expect high overlap among individuals (IS). This does not seem to be true, IS value being the lowest recorded among the wasp populations analysed (Table 2). The relative niche width was also low (Fig. 4). A study on the factors affecting the prey selection, including hunting sites, behavioural ecology of prey types, and chemical discrimination, are necessary to understand such patterns. Bembix merceti is a solitary Diptera-hunting wasp restricted to the Iberian Peninsula. The nests are unicellular and wasps form small aggregations. In a previous study, females of B. merceti were defined as opportunistic in prey selection, since whereas in 1993 nearly 90% of the prey were beeflies (Bombylidae), in 1994 the prey belonged mostly (50%) to Muscidae [43]. The data analysed here come from a study carried out in Soria (Central Spain) during summer of 2008, in a sandy area with sparse bushes in a landscape of cereal crops. Individuals of this species weakly differed in their

Individual prey specialization versus size in wasps 117
diets (P-value was just below the 0.05 level), while the relative niche width was moderate (Table 2, Fig. 4). PSi and Di were not correlated (Table 2), while size of females did not correlate with niche overlap and niche width (Table 2). The relationship between wasp and prey size was not significant (although close to significance) (Table 2 and Fig. 3). However, in this species, female size does not seem to affect greatly the intra-specific diet overlap. At last, B. merceti showed low variation in size (Fig. 4). According to our model, we thus expect high overlap among individuals. This seems to be true, IS showing weaker diet segregation with respect to all the other specialized populations (Table 2). One possible factor accounting for these patterns may be the relative small size of prey compared to wasp size. In fact, on average, prey weighed about 17% of the wasp weight (between 10% and 23%), and this may result in the possibility, for all the females, to hunt upon all the prey types in similar percentages. According to our model, if all wasps are large enough to subdue and carry all the prey types, intra-specific diet overlap will be greater and this seems, to some extent, to be the case for B. merceti. Bembix sinuata is a Mediterranean solitary wasp whose biology is little known. Published data are available only on the nest structure and prey [44]. This species nest in sandy, horizontal soil devoid of vegetation and prey upon flies of different families, such as Calliphoridae, Syrphidae and Muscidae [44]. The data analysed here come from an unpublished study carried out in Spain during 2007 summer, in the same area as previously described for B. merceti. Individuals of this species did not overlap their diet, while niche width (D and Di) was the widest recorded among the wasp populations reviewed here (Table 2). No correlation appeared between PSi and Di (Table 2), while size of females did not correlate with niche overlap and niche width (Table 2). As in the congeneric B. merceti, the relationship between wasp size and prey size was absent (Table 2 and Fig. 3). At last, B. sinuata showed moderate variation in size (Fig. 4). According to our model, we thus expect moderate overlap among individuals (IS). This does not seem to be true, IS being very low among the values obtained for the studied populations (Table 2). Also the relative niche width was quite reduced (Fig. 4). Interestingly, and maybe accounting for the observed difference between the two Bembix species, in B. sinuata the ratio between prey and wasp size was higher than in B. merceti, about 0.54 (between 0.48 and 0.57), i.e. in this case is unlikely that all the females may hunt all the

Carlo Polidori et al. 118
Figure 4. Mean niche overlap (IS=mean PSi) and mean relative niche width (Di/D) across the 10 analysed populations. Populations are ordered by increasing wasp size variance (CV=SD/mean, values showed above the bars). The CV values were all calculated from linear measurements (head width, thorax width, or body length), in order to make possible inter-specific comparisons. For population codes, see Table 1. prey species. This would at least partially explain the lower intra-specific diet overlap in this species relative to B. merceti. Thus, in this species, female size does not seem to affect greatly the intra-specific diet overlap. Across-populations comparisons and conclusions of the study This review shows that the degree of intra-specific diet overlap and the individual niche width are related to predator size in a variety of ways which are difficult to predict in absence of additional, detailed information on both the predator and prey populations’ biology. The four predictions of our model (positive correlation between niche overlap/niche width and size, positive relationship between the two indices and positive correlation between predator and prey size) were all accomplished by only one of the populations considered here (C. californica). In this case, as predicted by the model, smaller females weakly overlap their diet, have narrower niches and hunt for smaller prey indicating that predator size is a good predictor of the degree of individual specialization. Only the positive relationship between predator and prey size was found quite commonly throughout the study populations (in five populations). Interestingly, in two cases (S. continuus I and C. arenaria),

Individual prey specialization versus size in wasps 119
we found a negative correlation between size and niche overlap (larger wasps more specialized). Wasps eventually segregate their niches, disagreeing with the main assumption of our model, i.e. that larger wasps hunt for a wider spectrum of prey species than smaller ones. As a consequence of all these results, average individual specialization (IS and mean Di/D) in general did not correlate with the variance of females’ size. On the contrary, populations having moderate size variance seem to be often also those with low individual specialization (Fig. 4). Maybe, in populations with high variance, larger wasps can eventually discard smaller prey species, as maybe occurs in C. arenaria, so that the average individual specialization increases. This situation is in accordance with the optimal foraging theory. On the other side, at least some populations with very small size variation (e.g. O. lamellatus, S. continuus II, C. californica) do not show, contrary to expectations, high overlap; maybe these wasps, to avoid competition, tend to segregate more their prey spectrum. Intra-specific competition has been shown to increase individual specialization in other animal taxa [e.g. 45, 46]. These findings somehow support what has been recently found for a fish, the threespine stickleback (Gasterosteus aculeatus L.): although the similarity in trophic morphology was correlated with dietary similarity between fish individuals, both body size and trophic morphology accounted for low variance in diet overlap [47]. Interestingly, the ratio between prey and wasp size seems to account, to some extent, for the observed differences among populations. In fact, although the ratio was calculated only if the weight was used as a measure for both predator and prey size (for a total of 4 populations), one notes that the two populations with higher prey/wasp weight ratio (S. continuus II: 0.88; O. lamellatus: 1.35) had very high individual specialization (S. continuus II: 0.34; O. lamellatus: 0.45). On the contrary, B. merceti and S. continuus I (0.17 and 0.47 of weight ratio, respectively) had lower specialization (S. continuus I: 0.64; B. merceti: 0.56). A weight ratio well below 1 may indicate that most of females are able to hunt all the (generally small) prey species. In our model, if all wasps could hunt all prey species, individual specialization would be drastically reduced. In conclusion, large females of apoid wasps rarely exploit their theoretical size-dependent advantage enlarging their prey spectrum, probably because different additional factors, such as individual-biased learning for certain prey taxa [e.g. 48], particular prey-subduing techniques [e.g. 49], particular hunting sites [e.g. 50], or even depending on competition’s pressure [e.g. 45, 46], have more influence on the dynamics of prey specialization. In this sense, the niche axes have to be chosen appropriately when measuring niche variation, because, if functionally distinct resources

Carlo Polidori et al. 120
are pooled (e.g. simply according to taxa), one may underestimate individual specialization; conversely, high among-individuals variation may not be biologically significant if it is based on short-time sampling [30]. Thus, the use of several sampling schemes is necessary to better understand the predator-prey dynamics in wasps, as it (I) ensures temporal consistency in trophic spectrum variation; and (II) capture all possible behavioural and morphological traits of prey and predators. Nevertheless, predator size distribution and predator-prey size relationship still remain, despite in non-obvious ways, an important factor shaping resource use at the individual and population-level in these insects. Acknowledgements Thanks are due to Riguel Feltrin Contente, which critically reviewed an early manuscript of this chapter, improving it. We are indebted to the Maremma Regional Park offices for the permit provided to study C. rubida biology in the protected territory. References 1. Hutchinson, G.E. 1957, Cold Spring Harbor Symp. Quant. Biol., 22, 415. 2. Pielou, E.C. 1975, Ecological diversity, Wiley, New York. 3. Price, T. 1987, Ecology, 68, 1015. 4. West, L. 1986, Ecology, 67, 798. 5. Durell, S.E.A. le. V. dit. 2000, Biol. Rev., 75, 503. 6. Werner, T.K., and Sherry, T.W. 1986, Proc. Natl. Acad. Sci. USA, 84, 5506. 7. Ehlinger, T.J., and Wilson, D.S. 1988, Proc. Natl. Acad. Sci. USA, 85, 1878. 8. Bolnick, D.I., Svanbäck, R., Fordyce, J.A., Yang, L.H., Davis, J.M., Hulsey,
C.D., and Forister, M.L. 2003, Am. Nat., 161, 1. 9. Van Valen, L. 1965, Am. Nat., 99, 377. 10. Roughgarden, J. 1972, Am. Nat., 106, 683. 11. Svanbäck, R., and Bolnick, D.I. 2007, Proc. R. Soc. Lond. B. Biol. Sci., 274, 839. 12. Coelho, J.R. 2011, in: Predation in the Hymenoptera. An Evolutionary
Perspective, C. Polidori, (Ed.), Research Signpost, India, Trivandrum. 13. Mittelbach, G.G. 1981, Ecology, 62, 1370. 14. Persson, L., and De Roos, A.M. 2003, Ecology, 84, 1129. 15. Bolnick D.I., Svanbäck R., Fordyce J.A., Yang L.H., Davis J.M., Hulsey
C.D., and Forister M.L. Honek, A. 1993, Oikos, 66, 483. 16. Henry, L.M., Ma, B.O., and Roitberg, B.D. 2009, Oecologia, 161. 433. 17. Scharf F.S., Juanes F., and Rountree R.A. 2000. Mar. Ecol. Progr. Ser., 208, 229. 18. Polidori, C., Boesi, R., Isola, F., and Andrietti, F. 2005, Eur. J. Entomol.,
102, 801.

Individual prey specialization versus size in wasps 121
19. Polidori, C., Mendiola, P., Asís, J.D., Tormos, J., Garcia M.D., and Selfa, J. 2009, J. Nat. Hist., 43, 2985.
20. Linsley, E.G., and MacSwain, J.W. 1956, Ann. Entomol. Soc. Am., 49, 71. 21. Lin, N. 1979, J. Washington Acad. Sci., 68, 159. 22. Polidori, C., Boesi, R., Ruz, L., Montalva, J., and Andrietti, F. 2009, Stud.
Neotrop. Fauna E., 44, 55. 23. Hastings, J.M., Holliday, C.W., and Coelho, J.R. 2008, Fla. Entomol., 91, 657. 24. Polidori, C., Gobbi, M., Chatenaud, L., Santoro, D., Montani, O., and Andrietti,
F. 2010, Biol. J. Linn. Soc. 25. Alexander, B. 1985, J. Nat. Hist., 19, 1139. 26. Karsai, I., Somogyi, K., and Hardy, I.C.W., Biol. J. Linn. Soc., 87, 285. 27. Fisher, D.O., and Dickman, C.R. 1993, Ecology, 74, 1871. 28. Roger, C., Coderre, D., and Boivin, G. 2000, Entomol. Exp. Appl., 94, 3. 29. Strohm, E., and Marliani, A. 2002, Behav. Ecol., 13, 52. 30. Bolnick, D.I., Yang, L.H., Fordyce, J.A., Davis, J.M., and Svanbäck, R. 2002,
Ecology, 83, 2936. 31. Sargeant, B.L. 2007, Oikos, 116, 1431. 32. Levins, R. 1968, Evolution in changing environments: some theoretical
explorations, Princeton University Press, Princeton, New Jersey. 33. Santoro, D., Polidori, C., Asís, J.D., and Tormos, J. 2011, J. Animal. Ecol., doi:
10.1111/j.1365-2656.2011.01874.x 34. Tormos, J., Asís, J.D., Gayubo, S.F., Portillo, M., and Torres, F. 2000, Ann.
Entomol. Soc. Am., 93, 326. 35. Asís, J.D., Baños-Picón, L., Tormos, J., Ballesteros, Y., Gayubo, S.F., Alonso,
M. 2011, Behaviour, 148, 191. 36. Brockmann, H.J. 1985, J. Kansas Entomol. Soc., 58, 631. 37. Polidori, C., Boesi, R., Pesarini, C., Papadia, C., Federici, M., Bevacqua, S., and
Andrietti, F. 2007, Zool. Stud., 46, 83. 38. Polidori, C., Mendiola, P., Asís, J.D., Tormos, J., Selfa, J., and Andrietti, F. 2008.
Anim. Behav., 75, 1651. 39. Asís, J.D., Tormos, J., and Jiménez, R. 1988, Ent. News, 99, 199. 40. Field, J., and Foster, W.A. 1995, Anim. Behav., 50, 99. 41. Polidori, C., Casiraghi, M., Di Lorenzo, M., Valarani, B., and Andrietti, F. 2006,
J. Ethol., 24, 155. 42. Polidori, C., Federici, M., Papadia, C., and Andrietti, F. 2006, It. J. Zool., 73, 55. 43. Asís, J.D., Tormos, J., and Gayubo, S.F. 2004, J. Nat. Hist., 38, 1799. 44. Asís, J.D., Gayubo, S.F., and Tormos, J. 1992, Deut. Entomol. Z., 39, 221. 45. Bolnick, D.I. 2001, Nature, 410, 466.a 46. Svänback, R., and Bolnick, D.I. 2007, Proc. R. Soc. Lond. B. Biol. Sci., 274, 839. 47. Bolnick, D.I., and Paull J.E. 2009, Evol. Ecol.Res., 11, 1217. 48. Punzo, F. 2005, J. N. Y. Entomol. Soc., 113, 222. 49. Weiss, M.R., Wilson, E.E., and Castellanos, I. 2004, Anim. Behav., 68, 45. 50. Ehlinger, T.J. 1989, Anim. Behav., 38, 643.




















