
Activated T cells recruit exosomes secreted by dendritic cells ...
Upload
independentCategory
view
0download
0
In vivo spermatotoxic effect of chromium as reflectedin the epididymal epithelial principal cells, basalcells, and intraepithelial macrophages of a nonhumanprimate (Macaca radiata Geoffroy)
Mariajoseph Michael Aruldhas, Ph.D.,a Senthivinayagam Subramanian, Ph.D.,a,b
Pasupathi Sekhar, Ph.D.,a Ganapathi Vengatesh, Ph.D.,a Peranaidu Govindarajulu, Ph.D.,a
and Mohammad Abdulkader Akbarsha, Ph.D.c
a Department of Endocrinology, Dr. ALM Post-Graduate Institute of Basic Medical Sciences, University of Madras, Taramani
Campus, Chennai, India; b Present address: Texas A & M University System Health Science Center, Temple, Texas; andc Department of Animal Science, School of Life Sciences, Bharathidasan University, Tiruchirappalli, India
Objective: To understand, through a simulation experiment in a nonhuman primate model, the potential in vivo
spermatotoxic toxic effect of hexavalent chromium (CrVI) in men who are occupationally or environmentally
exposed to it.
Design: Controlled laboratory study.
Setting: Research laboratory in a department of endocrinology in a university in India.
Animal(s): Male bonnet monkey, Macaca radiata Geoffroy.
Intervention(s): Monkeys were exposed ad libitum to 100, 200, and 400 ppm CrVI, dissolved in drinking water,
for a chronic period of 180 days.
Main Outcome Measure(s): Examination of epididymis with a transmission electron microscope and assessment
of the effect of CrVI in terms of accumulation of sperm-derived lipofuscin (LF) material in the principal cells,
basal cells, and intraepithelial macrophages of the epithelium.
Result(s): The abundance of basal cells and intraepithelial macrophages and the content of LF material in these
cell types increased. The principal cells phagocytosed from the lumen the dead sperm resulting from CrVI
exposure and processed them partially into LF material, which was acquired by the basal cells and intraepithelial
macrophages and processed further. The LF material–laden basal cells and intraepithelial macrophages appeared
to leave the epithelium, accompanied by recruitment of fresh basal cells and intraepithelial macrophages.
Conclusion(s): Occupational or environmental exposure to CrVI, as would occur in the tannery, soap, and other
industries in developing and underdeveloped countries, can be toxic in vivo to spermatozoa. (Fertil Steril� 2006;
86(Suppl 3):1097–105. ©2006 by American Society for Reproductive Medicine.)
Key Words: Basal cell, chromium, epididymis, intraepithelial macrophage, lipofuscin material, principal cell,
spermatotoxicity
Hexavalent chromium (CrVI) is a heavy-metal pollutant that
can cause carcinogenesis and several systemic toxicities
(1–5). A case-controlled study of the male partners of infer-
tile couples who were exposed to CrVI in welding fumes
through the nasal route suggested that low-level exposure to
this heavy metal may not be a major hazard affecting sper-
matogenesis (6). In a recent review, Bonde (7) opined that
evidence to implicate CrVI in male reproductive toxicity is
not adequate. Two recent epidemiological studies, one from
China (8) and another from India (9), also attest poor semen
quality in men who are exposed to Cr in the chrome-plating
and welding industries.
The available information on the male-reproductive toxic
effect of CrVI in human beings is confined to occupational
exposure to CrVI fumes from stainless-steel welding through
the nasal route only. However, occupational exposure to
CrVI varies with the intensity and the route, depending upon
the nature of work. For example, in the case of the tannery,
soap, and a few other industries, the exposure route can be
oral and even through the skin. Tannery effluent in several
developing countries, including India, and in underdevel-
oped countries is let out without much treatment and can
contaminate water bodies, including subsoil water. These
occupational and environmental exposures to CrVI could be
chronic. Therefore, there is pertinent need to investigate the
male-reproductive toxic effect of CrVI in high-level chronic-
exposure groups.
Received December 24, 2005; revised and accepted March 17, 2006.
Supported by the Council of Scientific and Industrial Research, Govern-
ment of India, New Delhi (grant 60(00222)/97/EMR II, to M.M.A.); by the
Department of Science and Technology, Government of India, New
Delhi, under the Fund for Improvement of Science and Technology
program (SR/FST/LSI-206/200 and SR/FST/LSI-112/2002); and by the
University Grants Commission, New Delhi, India, under the Special
Assistance program (F.3-1/99(SAP-III)).
Reprint requests: Mariajoseph Michael Aruldhas, Ph.D., Department of
Endocrinology, Dr. ALM Post-Graduate Institute of Basic Medical Sci-
ences, University of Madras, Taramani Campus, Chennai 600113, India
(FAX: 91-44-24926709; E-mail: [email protected]).
10970015-0282/06/$32.00 Fertility and Sterility� Vol. 86, Suppl 3, October 2006doi:10.1016/j.fertnstert.2006.03.025 Copyright ©2006 American Society for Reproductive Medicine, Published by Elsevier Inc.

Experiments in laboratory animal models and cell lines
attest impairment of male reproductive function by chro-
mium (10–19), albeit some differences as a result of varia-
tions in the experimental design. It is not possible to carry
out such experiments in human beings, and hence, studies in
an animal model close to human beings, such as nonhuman
primates, that simulate the occupational and/or environmen-
tal exposure to CrVI in the tannery industry for a chronic
period may be pertinent.
Adopting the above rationale, we subjected male bonnet
monkeys (Macaca radiata) to chronic–CrVI toxicity testing.
We reported several adverse changes in the testis, including
cytological changes in different generations of germ cells in
the adluminal compartment of the seminiferous epithelium,
generation of uninucleate and multinucleate giant cells, pre-
mature release of germ cells from the Sertoli cell, occurrence
of macrophages in the lumen of the seminiferous tubules,
and decrease in the activities of enzymatic and nonenzymatic
antioxidants in the testis (20).
Earlier, we reported the presence of undifferentiated
and/or immature uninucleate and multinucleate germ cells in
the lumen of ductus epididymidis, and hypertrophy and
hyperplasia of the cauda epididymal epithelium. These
changes obstructed the distal portions of the male reproduc-
tive tract, resulting in a tendency of sperm to escape through
certain weak patches in the epithelium. To prevent the gen-
eration of auto-antibodies to sperm antigens in sequel to such
an extravasation, microcanaliculi, lined by a hitherto unre-
ported cell type, developed in the epididymal epithelium, in
which these sperm were closeted and secluded from the
circulatory system. It was suggested that these cells were
derived from basal cells (BCs) (21).
In recent years, several reports have ascribed a role to
epididymal epithelial BCs and intraepithelial macrophages
(IEMs) in surveillance of sperm antigens when there is
obstruction to passages of sperm (22–26). Obstruction of
sperm passage leads to increase in the number of BCs and
IEMs and to a dense accumulation of lipofuscin (LF) mate-
rial in their cytoplasm. These are indications of the arrival of
sperm antigens into the epithelium through transcytosis
across the principal cells (PCs) and also through leaky junc-
tions between the tall columnar cells. These changes result in
increased abundance of BCs and IEMs and also in increased
content of LF material in these cell types, which are reflec-
tions of a pathophysiological state of the luminal sperm and
of a defense mechanism to sperm antigens.
Taking a cue from the above background information, we
used a transmission electron microscope to reexamine, using
the same tissue blocks used for our earlier study (21), dif-
ferent segments of the epididymis of monkeys that were
exposed to CrVI. This article reports similar changes in the
BCs and IEMs. The study strongly implicates chronic CrVI
exposure through the oral route in the male-reproductive
toxic effect of bringing about death of epididymal luminal
sperm, but the epididymis responds to the situation with
appropriate changes to process them.
MATERIALS AND METHODS
The animal (Macaca radiata), experimental design, treat-
ment of CrVI, and analysis were the same as reported in our
article published elsewhere (21), except that the recovery
study is not included in the present report. The experimental
design was approved by the institutional animal ethics com-
mittee, which includes a nominee of the Committee for the
Purpose of Control and Supervision of Experiments on An-
imals, Government of India.
Briefly, adult monkeys obtained from the Department of
Forests, Government of Tamil Nadu, India, were subjected
to quarantine and maintained under animal-house conditions.
The monkeys were divided into four groups of three each.
Monkeys in groups 1 to 3 were provided with drinking water
containing 100, 200, and 400 ppm CrVI (potassium dichro-
mate), respectively, for 180 days. Monkeys in group 4
formed the control and received plain water in which CrVI
was not dissolved.
At the end of the experimental period, the monkeys were
subjected to sodium pentobarbital anesthesia, and the testi-
cles and epididymides were autopsied through an incision in
the scrotum, after which the incision was closed with stitches
of nylon thread. After healing of the wound and removal of
the stitches, the monkeys were released into the reserve
forest as per the guidelines of Committee for the Purpose of
Control and Supervision of Experiments on Animals, Gov-
ernment of India. Under this circumstance, perfusion fixation
was not performed. Thin slices of different segments of the
epididymides were fixed in 2.5% glutaraldehyde prepared in
cacodylate buffer and were postfixed in 1% osmium tetrox-
ide. Semithin sections (1 �m), obtained with a Leica (Gena,
Germany) ultramicrotome, were stained with toluidine blue
O and were used to select areas for ultrathin sections. The
latter, cut with the same ultramicrotome, were stained with
uranyl acetate and lead citrate and observed in a Phillips
201C (Amsterdam, Holland) transmission electron micro-
scope. The images were scanned into a Pentium III com-
puter, and the images were edited and processed using Pho-
toshop version 7.0 software (Adobe, San Jose, CA).
RESULTS
Irrespective of the segments, that is, initial segment (IS),
caput (CA), corpus (CO), and cauda (CU), the epididymal
epithelium of control monkeys possessed very few BCs and
IEMs (Fig. 1A and B). The BCs had the characteristic
arrangement abutting the basal lamina (BL) and possessed
heterochromatic round or irregular nucleus occupying al-
most the entire cell, leaving only little cytoplasm around it.
The cytoplasm was scarce in organelles, which consisted of
mitochondria, endoplasmic reticulum, lysosomes, and endo-
cytic vesicles. The Golgi apparatus was hardly visible (Fig. 1C).
1098 Aruldhas et al. Spermatotoxic effect of chromium in a nonhuman primate Vol. 86, Suppl 3, October 2006
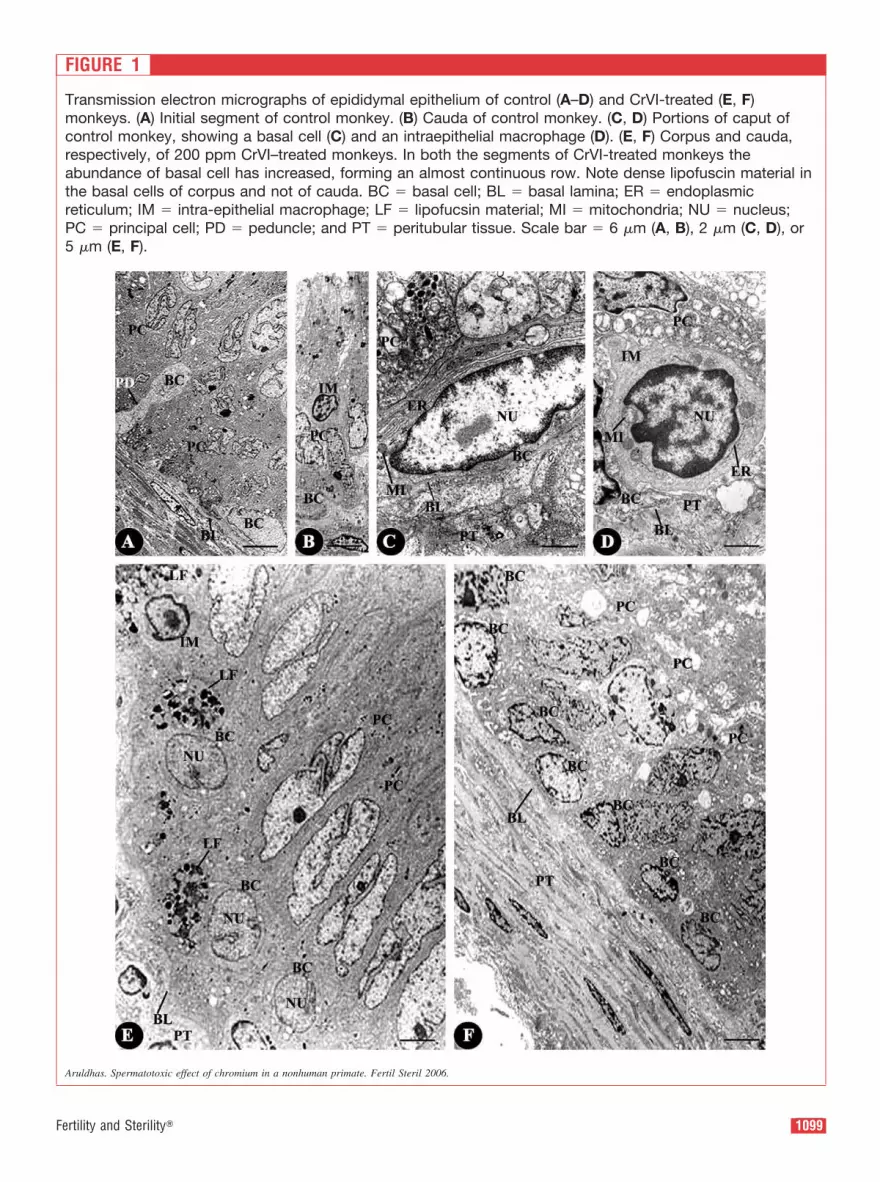
FIGURE 1
Transmission electron micrographs of epididymal epithelium of control (A–D) and CrVI-treated (E, F)
monkeys. (A) Initial segment of control monkey. (B) Cauda of control monkey. (C, D) Portions of caput of
control monkey, showing a basal cell (C) and an intraepithelial macrophage (D). (E, F) Corpus and cauda,
respectively, of 200 ppm CrVI–treated monkeys. In both the segments of CrVI-treated monkeys the
abundance of basal cell has increased, forming an almost continuous row. Note dense lipofuscin material in
the basal cells of corpus and not of cauda. BC � basal cell; BL � basal lamina; ER � endoplasmic
reticulum; IM � intra-epithelial macrophage; LF � lipofucsin material; MI � mitochondria; NU � nucleus;
PC � principal cell; PD � peduncle; and PT � peritubular tissue. Scale bar � 6 �m (A, B), 2 �m (C, D), or
5 �m (E, F).
Aruldhas. Spermatotoxic effect of chromium in a nonhuman primate. Fertil Steril 2006.
1099Fertility and Sterility�

FIGURE 2
Aruldhas. Spermatotoxic effect of chromium in a nonhuman primate. Fertil Steril 2006.
1100 Aruldhas et al. Spermatotoxic effect of chromium in a nonhuman primate Vol. 86, Suppl 3, October 2006

Although a few BCs were flat, a few others were elongated
and had a vertical orientation, contacting the BL through a
broad cytoplasmic peduncle (Fig. 1A).
A few IEMs, which were present at different heights along
the epithelium, possessed densely heterochromatic nucleus,
large mitochondria, and scattered endoplasmic reticulum.
The cells were characterized by irregular shape as a result
of the pseudopodia (Fig.1A and B; see also Fig. 3A). Neither
the BCs nor the IEMs possessed any perceptible LF material
(Fig. 1C and D).
In the epididymis of CrVI-treated monkeys, the BCs were
abundant and formed an almost continuous row on top of the
BL in such a way that the epithelium appeared pseudostrati-
fied (Fig. 1E and F). The cytoplasm of every BC of IS, CA,
and CO contained a dense accumulation of LF material,
which was more voluminous than in the control monkeys
(Fig. 1E). Though the BCs were abundant in the CU as in the
proximal segments, the cytoplasm had only sparse LF
material (Fig. 1F).
In several BCs of IS, CA, and CU, the LF material
occupied almost the entire cytoplasm (Figs. 1E and 2A), and
the nucleus was indented at one pole to accommodate the
large mass of LF material (Fig. 2A). The LF material of BCs
occurred as small to large membrane-bound vesicles, each
containing a material of varying electron densities. In some
vesicles, the content at the periphery was more electron
dense than that at the core (Fig. 2B). Cells matching BCs in
organization and loaded with LF material were present in the
peritubular tissue (Fig. 2C). Leukocytic cells matching BCs
but not containing LF material were observed to make access
into the epithelium from the peritubular tissue through the
BL (Fig. 2D).
The IEMs also increased in abundance in the IS, CA, and
CO. In a few cases, the IEMs were engaged in phagocytotic
uptake of cell debris including sperm, which in all probabil-
ity were dead (Fig. 3A). Most of the IEMs, however, con-
tained LF material, some of which were more electron dense
than the BCs (Figs. 2B and 3B). The IEMs acquired the LF
material from outside through phagocytosis (Fig. 3C).
The PCs of IS, CA, and CO also contained LF material in
the supranuclear as well as perinuclear cytoplasm (Fig. 3D
and E). The cytoarchitecture of the PCs was invariably
normal, containing clathrin-coated vesicles in the apical
plasma membrane indicating endocytotic activity (Fig. 3D),
prolific Golgi stacks in the supranuclear cytoplasm, and
vacuolated perinuclear cytoplasm (Fig. 3E). Several PCs
contained spermatozoa in the cytoplasm at various locations
from apical to the base (Fig. 4A–C). The spermatozoa were
in the process of disintegration inside large membrane-
bound vesicles, and LF material was found in the vicinity of
such vacuoles (Fig. 4D). These were spermatozoa that were
phagocytosed at the apical plasma membrane of the PCs
(Fig. 4E). Once inside the cytoplasm, the spermatozoa were
closeted in membrane-bound vesicles (Fig. 4F).
The responses increased with the dose of CrVI treatment;
that is, from 100 to 400 ppm (data not presented).
DISCUSSION
The present study indicates that CrVI exposure leads to
increase in abundance of BCs and IEMs in the monkey
epididymal epithelium and that the cytoplasm of both the
cell types in the IS, CA, and CO is laden with LF material.
Both the cell types appeared to acquire the LF material from
the PCs, produced as a result of partial digestion of sperma-
tozoa, which were phagocytosed through the apical plasma
membrane. The spermatozoa thus phagocytosed are to be
considered dead because live sperm are not phagocytosed,
excepting in the case of autoimmune response to sperm
antigens (27). The results support our earlier findings (20,
21) that exposure of monkey to CrVI through the oral route
for a chronic period of 180 days can bring about toxic effects
on the testis and epididymis, including spermatotoxicity.
Basal cells are the second largest population of epithelial
cell type that is common to all segments of the epididymis,
next only to PCs (22). Consistently found in the epididymis
of the different mammalian species, the BCs are established
only after the other cell types (23). For example, in the
mouse epididymis, the BCs are not present before days 12,
14, and 16 postpartum in the CU, CA, and CO epididymides,
respectively. The BC number increases in all segments be-
tween days 20 and 27 postpartum (23). The temporal pattern
in the first appearance of BCs in the epididymal epithelium
FIGURE 2 Continued
Transmission electron micrographs showing basal cells in the epididymal epithelium (A, B) and a basal cell–
like cell (C) and leukocytes (D) in the peritubular tissue of CrVI-treated monkeys. (A) Basal cell of corpus
showing dense accumulation of lipofuscin material and, also, indentation of nucleus at the apical pole to
accommodate it (arrowhead). (B) Portion of a basal cell of initial segment showing the heterogeneous nature
of lipofuscin material. (C) A basal cell–like cell, loaded with lipofuscin material (arrowhead), in the peritubular
tissue. (D) Entry of leukocytes from the peritubular tissue into the epithelium (arrowheads). BC � basal cell;
BL � basal lamina; CA � capillary; EN � endothelial cell around capillary; LE � peritubular tissue
leukocytes; LF � lipofuscin material; NU � nucleus; PC � principal cell; and PT � peritubular tissue. Scale
bar � 5 �m (A), 2 �m (B, D), or 3 �m (C).
1101Fertility and Sterility�
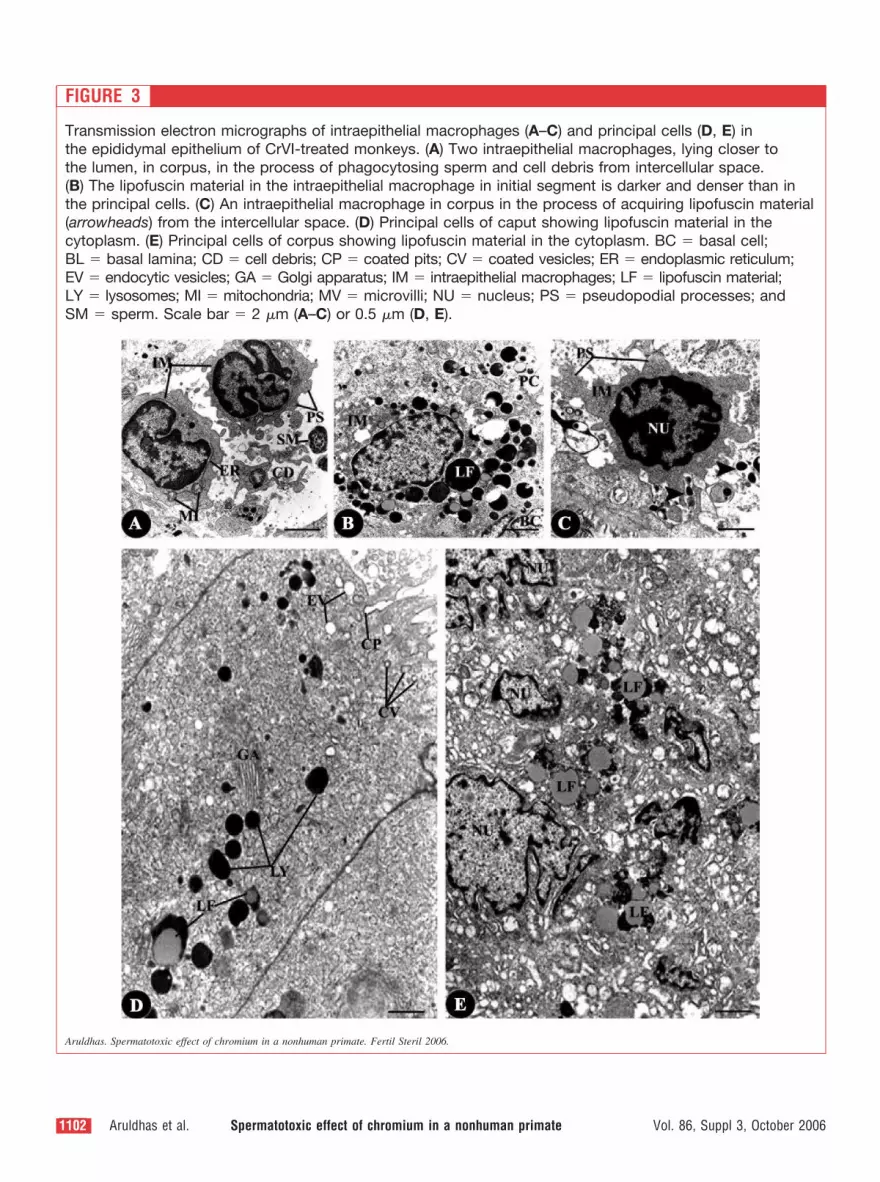
FIGURE 3
Transmission electron micrographs of intraepithelial macrophages (A–C) and principal cells (D, E) in
the epididymal epithelium of CrVI-treated monkeys. (A) Two intraepithelial macrophages, lying closer to
the lumen, in corpus, in the process of phagocytosing sperm and cell debris from intercellular space.
(B) The lipofuscin material in the intraepithelial macrophage in initial segment is darker and denser than in
the principal cells. (C) An intraepithelial macrophage in corpus in the process of acquiring lipofuscin material
(arrowheads) from the intercellular space. (D) Principal cells of caput showing lipofuscin material in the
cytoplasm. (E) Principal cells of corpus showing lipofuscin material in the cytoplasm. BC � basal cell;
BL � basal lamina; CD � cell debris; CP � coated pits; CV � coated vesicles; ER � endoplasmic reticulum;
EV � endocytic vesicles; GA � Golgi apparatus; IM � intraepithelial macrophages; LF � lipofuscin material;
LY � lysosomes; MI � mitochondria; MV � microvilli; NU � nucleus; PS � pseudopodial processes; and
SM � sperm. Scale bar � 2 �m (A–C) or 0.5 �m (D, E).
Aruldhas. Spermatotoxic effect of chromium in a nonhuman primate. Fertil Steril 2006.
1102 Aruldhas et al. Spermatotoxic effect of chromium in a nonhuman primate Vol. 86, Suppl 3, October 2006

FIGURE 4
Transmission electron micrographs of principal cells of proximal epididymal segments of CrVI-treated
monkeys showing spermatozoa in the cytoplasm, their acquisition and processing. (A) Sperm heads and
tails in the cytoplasm. (B) The spermatozoa are found closer to the basal lamina. (C) A spermatozoon,
in longitudinal section, in the cytoplasm. (D) Spermatozoa partially processed by lysosomal activity.
(E) Acquisition of spermatozoa through deep indentations between microvilli (arrowheads). (F) The
spermatozoa are closeted in a membrane-bound vesicle in the apical cytoplasm. BC � basal cells;
BL � basal lamina; ER � endoplasmic reticulum; IM � intraepithelial macrophages; LU � lumen;
LF � lipofuscin material; MI � mitochondria; MV � microvilli; NU � nucleus; PC � principal cells;
SM � sperm; and VA � membrane-bound vesicle. Scale bar � 4 �m (A, B), 0.5 �m (C, D, F), or 1 �m (E).
Aruldhas. Spermatotoxic effect of chromium in a nonhuman primate. Fertil Steril 2006.
1103Fertility and Sterility�

has been shown in the rat, ram, rabbit, goat, cat, and human
(23, 28). The hypotheses that the BCs are progenitors of PCs
(29) and that the columnar cells give rise to the BCs (30)
have been set aside (26). The BCs have been shown to
express macrophage antigens in human (22) and in monkey
and mouse (23–26), suggesting that the BCs may be derived
from monocytes, which arrive at the epididymal epithelium
for transformation into macrophages (26). The ultrastructural
similarities between BCs and interstitial macrophages also
attest this suggestion (22).
Several lines of evidence suggest that the BCs are a part of
the immune defense system of epididymis (22–26). A link
between macrophage antigen expression in the BCs and
presence of damaged spermatozoa in the epididymis has
been suggested. In the normal male, the proportion of dam-
aged spermatozoa and the expression of macrophage anti-
gens in the BCs are low in the CU (23). Experimentally
induced sperm cell damage in situ demonstrated that the BCs
respond to the presence of sperm antigens in the lumen with
increased number and expression of macrophage antigens.
This would prevent interaction of sperm antigens with the
immune system and abrogate the risk of immunological
infertility (25).
The relative abundance of LF material in the BCs is a
reflection of the extent of scavenging of sperm antigens. The
LF material in BCs increases with age (22), and it depends
on the arrival of sperm antigens in the epithelium under
experimental or pathophysiological conditions (25). The in-
crease of LF material in the PCs and its prolific occurrence
in the BCs of CrVI-treated monkeys indicates the arrival of
sperm antigens, which could have been processed already by
the PCs. Because the release of antigens from the sperma-
tozoa occurs in pathophysiological states (25, 26), the results
of the present study indicate CrVI-induced pathophysiolog-
ical state of the spermatozoa. This is substantiated by the
increase in the abundance of IEMs with LF material. Active
uptake of LF material by the IEMs from the intercellular
spaces has been documented (22). It also has been shown
that the LF material arrives at the intercellular spaces from
the PCs and thus becomes available to the BCs and IEMs
(22). The PCs acquire sperm antigens from the damaged
spermatozoa in lumen through adsorptive and/or fluid-phase
endocytosis (27).
The present observation in CrVI-treated monkeys shows
that the spermatozoa may be in a pathophysiological state
and are acquired by the PCs and processed in their cytoplasm
by lysosomal activity, resulting in LF material. According to
Gadhially (31), the LF material is the residual body that is
left behind in the cells after lysosomal activity. The abun-
dance of this material increases in the cells of older individ-
uals, and therefore it has been referred to as age pigments,
wear-and-tear pigments, and womb-to-tomb pigments. The
LF material is looked upon as a conglomerate of undigested
residues from lysosomal hydrolysis and modification of se-
questrated material before and after digestion. The power of
lysosomal hydrolases in degrading lipids is limited, and
hence, the lipids are left behind and undergo stepwise oxi-
dation, resulting in overloading of LF material. The LF accu-
mulation is engendered by various noxious influences (31).
We have shown elsewhere that CrVI exposure leads to
impairment of spermatogenesis as a result of the develop-
ment of oxidative stress in the testis of monkeys (20). In
another study published elsewhere, we showed that CrVI
treatment causes obstruction of the distal CU epididymidis
and vas deferens, leading to stress-induced sperm death (21).
Such stress-induced sperm death in the epididymis was at-
tributed to excess free radicals generated and poor scaveng-
ing of the same. Our observation of increased abundance of
LF material in the PCs, BCs, and IEMs attests the above
conclusions.
It is to be inferred that the sperm death caused by CrVI
intoxication occurs in such a large number that the luminal
macrophages do not adequately cleanse them. Therefore, the
PCs take over the role of phagocytosing dead spermatozoa
(32). The digestion of the spermatozoa acquired into the PC
cytoplasm is incomplete and results in LF material. Because
the PCs do not possess the mechanism to process the huge
quantity of LF material, the latter is discharged into the inter-
cellular spaces to be acquired by the BCs and IEMs. The LF
material–laden BCs and IEMs may leave the epithelium and
arrive at the peritubular tissue (22–26).
The poor content of LF material in the BCs and IEMs of
CU makes an interesting observation. Paracrine factors in-
duce macrophage antigen expression once the BCs have
taken up residence in the epithelium and the spermatozoa
and testicular fluid arrive at the epididymis (22). Thus, there
is greater expression of Mac-1 in the BCs of adult CA, and
its expression in the CU occurs only much later, suggesting
a link between macrophage antigen expression and the pres-
ence of spermatozoa and/or testicular fluid in the epididymis.
In the normal male, the proportion of damaged spermatozoa
in the CU was found to be low, whereas in the CrVI-treated
monkeys, the entire epithelium at the CU underwent hyper-
trophy and hyperplasia, contributing to obstruction to the
passage of spermatozoa (21). The consequent limited occur-
rence of spermatozoa in the CU may be responsible for the
fewer LF inclusions in the BCs and IEMs.
Thus, this article provides further strong evidence for the
male-reproductive toxicity of chronic exposure of a nonhu-
man primate to CrVI. The results indicate damage to the
epididymal luminal spermatozoa, as a consequence of which
the PCs take up the role of phagocytosis of the damaged
spermatozoa, which is followed by an increase in the number
of BCs and IEMs with dense accumulation of LF inclusions.
This may be another mechanism by which the epididymis
overcomes the adverse effects of chronic CrVI exposure on
the sperm apart from microcanalization to closet the viable
sperm that might get access into the epithelium (21). This
and our earlier articles on the testis and epididymis of
CrVI-treated monkeys (20, 21), supported by the two recent
1104 Aruldhas et al. Spermatotoxic effect of chromium in a nonhuman primate Vol. 86, Suppl 3, October 2006

reports of increased infertility in men with occupational
exposure to Cr (8, 9), strongly suggest Cr as a vulnerable
disrupter of male fertility.
Acknowledgments: The authors thank Anna Pulimood, M.D., for permis-
sion to use the transmission electron microscope facility of the Welcome
Trust Research Laboratory, Christian Medical College and Hospital, Vel-
lore, India, and also thank B. Kadalmani, M.D. for helping in image
processing.
REFERENCES
1. Burrows D. Metabolism and toxicity. In: Burrows D, ed. Toxicological
review of hexavalent chromium. Boca Raton, FL: CRC Press: 1998:1–
70.
2. Von Burg R, Liu D. Chromium and hexavalent chromium. J Appl
Toxicol 1993;13:225–30.
3. Barceloux D. Chromium. Clin Toxicol 1999;37:173–94.
4. Shi X, Chiu A, Cheu CT, Halliwall B, Castranova V, Vallyathan V.
Reduction of chromium (VI) and its relationship to carcinogenesis.
J Toxicol Environ Health B Crit Rev 1999;12:87–104.
5. Bagchi D, Stohs SJ, Downs BW, Bagchi M, Preuss HG. Cytotoxicity
and oxidative mechanisms of different forms of chromium. Toxicology
2002;180:5–22.
6. Bonde JP, Ernst E. Sex hormones and semen quality in welders exposed
to hexavalent chromium. Hum Exp Toxicol 1992;11:259–63.
7. Bonde JP. Occupational risk to male reproduction. G Ital Med Lav
Ergon 2002;24:112–7.
8. Li H, Chen Q, Yao W, Li L, Shi X, Wang L, et al. Effect of CrVI
exposure on sperm quality: human and animal studies. Ann Occup Hyg
2001;45:505–11.
9. Danadevi K, Rozati R, Reddy PP, Grover P. Semen quality of Indian
welders occupationally exposed to nickel and chromium. Reprod Toxi-
col 2003;17:451–6.
10. Behari J, Chandra SV, Tandon SK. Comparative toxicity of trivalent
and hexavalent chromium to rabbits. III. Biochemical and histological
changes in testicular tissue. Acta Biol Med Ger 1978;37:463–8.
11. Ernst E. Testicular toxicity following short-term exposure to tri- and
hexavalent chromium: an experimental study in the rat. Toxicol Lett
1990;51:269–75.
12. Saxena DK, Murthy RC, Lal B, Srivastava RS, Chandra SV. Effect of
hexavalent chromium on testicular maturation in the rat. Reprod Toxi-
col 1990;4:223–8.
13. Acharya UR, Mishra M, Tripathy RR, Mishra I. Testicular dysfunction
and antioxidative defense system of Swiss mice after chromic acid
exposure. Reprod Toxicol 2006;22:87–91.
14. Murthy RC, Saxena DK, Gupta SK, Chandra SV. Ultrastructural ob-
servations in testicular tissue of chromium-treated rats. Reprod Toxicol
1991;5:443–7.
15. Ernst E, Bonde JP. Sex hormones and epididymal sperm parameters in
rats following sub-chronic treatment with hexavalent chromium. Hum
Exp Toxicol 1992;11:255–8.
16. Chowdhury AR, Mitra C. Spermatogenic and steroidogenic impairment
after chromium treatment in rats. Indian J Exp Biol 1995;33:480–4.
17. Sutherland JE, Zhitkovich A, Kluz T, Costa M. Rats retain chromium
in testis following chronic ingestion of drinking water containing
hexavalent chromium. Biol Trace Elem Res 2000;74:41–53.
18. Pereira ML, Santos TM, Das Neves RP, Costa FG, de Jesus OP.
Chromium (V) involvement in the toxicity pathway of testicular dam-
age. Asian J Androl 2002;4:153–5.
19. Cheng RY, Alvord WG, Powell D, Kaspizak KS, Anderson LM.
Microarray analysis of altered gene expression in TM4 Sertoli-like cell
line exposed to chromium III 9 chloride. Reprod Toxicol 2002;16:
223–6.
20. Aruldhas MM, Subramanian S, Sekhar P, Vengatesh G, Chandrahasan
G, Govindarajulu P, et al. Chronic chromium exposure-induced
changes in testicular histoarchitecture are associated with oxidative
stress: study in non-human primate (Macaca radiata Geoffroy). Hum
Reprod 2005;20:2801–13.
21. Aruldhas MM, Subramanian S, Sekhar P, Chandrahasan G, Govindara-
julu P, Akbarsha MA. Microcanalizaiton in the epididymis to overcome
ductal obstruction caused by chronic exposure to chromium—a study in
the mature bonnet monkey (Macaca radiata Geoffroy). Reproduction
2004;128:127–37.
22. Yeung CH, Nashan D, Sorg G, Oberpenning F, Schulze H, Nieschlag E,
et al. Basal cells of the human epididymis—antigenic and ultrastruc-
tural similarities to tissue-fixed macrophages. Biol Reprod 1994;50:
917–26.
23. Seiler P, Wenzel I, Wagenfeld A, Yeung CH, Nieschlag E, Cooper TG.
The appearance of basal cells in the developing murine epididymis and
their temporal expression of macrophage antigens. Int J Androl 1998;
21:217–26.
24. Seiler P, Cooper TG, Yeung CH, Nieschlag E. Regional variation in
macrophage antigen expression by murine epididymal basal cells and
their regulation by testicular factors. J Androl 1999;20:738–46.
25. Seiler P, Cooper TG, Nieschlag E. Sperm number and condition affect
the number of basal cells and their expression of macrophage antigen in
the murine epididymis. Int J Androl 2000;23:65–76.
26. Holschbach C, Cooper TG. A possible extra-tubular origin of epidid-
ymal basal cells in mice. Reproduction 2002;123:517–25.
27. Flickinger CJ, Baran ML, Howards SS, Herr JC. Sperm autoantigens
recognized by auto-antibodies in developing rats following pre-pubertal
obstruction of the vas deferens. J Androl 1996;17:433–42.
28. Atanassova N, McKinnell C, Fisher J, Sharpe RM. Neonatal treatment
of rats with diethylstilbestrol (DES) induces stromal-epithelial abnor-
malities of the vas deferens and cauda epididymis in adulthood follow-
ing delayed basal cell development. Reproduction 2005;129:589–601.
29. Martan J. Epididymal histochemistry and physiology. Biol Reprod
1969;1:134–54.
30. Sun EL, Flickinger CJ. Proliferative activity in the rat epididymis
during postnatal development. Anat Rec 1982;203:273–84.
31. Gadhially FN. Lysosomes. In: Gadhially FN, ed. Ultrastructural pathol-
ogy of the cell and matrix. 3rd revised ed. London: Butterworths,
1988:589–853.
32. Sutovsky P, Moreno R, Ramalho-Santos J, Dominco T, Thompson WE,
Schatten G. A putative, ubiquitin-dependent mechanism for the recog-
nition and elimination of defective spermatozoa in the mammalian
epididymis. J Cell Sci 2001;114:1665–75.
1105Fertility and Sterility�
Copyright © 2022 FDOKUMEN




















