
Crosstalk and signaling switches in mitogen-activated protein kinase cascades
Upload
independentCategory
view
3download
0
ARTHRITIS & RHEUMATISMVol. 48, No. 6, June 2003, pp 1582–1593DOI 10.1002/art.11014© 2003, American College of Rheumatology
In Vivo Selective Inhibition ofMitogen-Activated Protein Kinase Kinase 1/2 in
Rabbit Experimental Osteoarthritis Is Associated Witha Reduction in the Development of Structural Changes
Jean-Pierre Pelletier,1 Julio C. Fernandes,1 Julie Brunet,1 Florina Moldovan,1 Denis Schrier,2
Craig Flory,2 and Johanne Martel-Pelletier1
Objective. The primary aim of this study was toinvestigate, using an experimental rabbit model of os-teoarthritis (OA), the effect of a selective mitogen-activated protein kinase kinase 1/2 (MEK-1/2) inhibitor,PD 198306, on the development of structural changes.Additional aims were to assess the effects of the inhib-itor on levels of phosphorylated extracellular signal–regulated kinase 1/2 (phospho–ERK-1/2) and matrixmetalloproteinase 1 (MMP-1; collagenase 1) in OAchondrocytes.
Methods. After surgical sectioning of the anteriorcruciate ligament of the right knee joint, rabbits withOA were separated into 3 experimental groups: oraltreatment with placebo or with PD 198306 at a thera-peutic concentration of 10 mg/kg/day or 30 mg/kg/day.Each treatment started immediately after surgery. Theanimals were killed 8 weeks after surgery. Macroscopicand histologic studies were performed on the cartilageand synovial membrane. The levels of phospho–ERK-1/2 and MMP-1 in OA cartilage chondrocytes wereevaluated by immunohistochemistry. Normal, untreatedrabbits were used as controls.
Results. OA rabbits treated with the highest dos-age of MEK-1/2 inhibitor showed decreases in thesurface area (size) of cartilage macroscopic lesions (P <0.002) and in osteophyte width on the lateral condyles(P � 0.05). Histologically, the severity of synovial in-flammation (villous hyperplasia) was also reduced (P <0.02). In cartilage from placebo-treated OA rabbits, asignificantly higher percentage of chondrocytes in thesuperficial layer stained positive for phospho–ERK-1/2and MMP-1 compared with normal controls. Rabbitstreated with the highest dosage of PD 198306 demon-strated a significant and dose-dependent reduction inthe level of phospho–ERK-1/2 and a lower level ofMMP-1.
Conclusion. This study demonstrates that, in vivo,PD 198306, a selective inhibitor of MEK-1/2, can par-tially decrease the development of some of the structuralchanges in experimental OA. This effect was associatedwith a reduction in the level of phospho–ERK-1/2 in OAchondrocytes, which probably explains the action of thedrug.
The degradation of osteoarthritic (OA) cartilageis related to a complex interaction of mechanical andbiochemical factors (1–3). Among the latter, a numberof catabolic factors, including proinflammatory cyto-kines, nitric oxide (NO), and proteases, have beendemonstrated to play major roles (1,4).
OA cells, at both the cartilage and synovialmembrane levels, are actively involved in producing anexcess amount of catabolic factors (1). The increase inthe metabolic activity of these cells is related to thestimulation of proinflammatory cytokines, such asinterleukin-1 (IL-1), tumor necrosis factor � (TNF�),IL-6, and leukemia inhibitory factor (1,4–6). After bind-
Supported by grants from Pfizer Global Research & Devel-opment, Ann Arbor, Michigan, and from Le Fonds de la Recherche enSante du Quebec (FRSQ).
1Jean-Pierre Pelletier, MD, Julio C. Fernandes, MD, JulieBrunet, PhD, Florina Moldovan, MD, PhD, Johanne Martel-Pelletier,PhD: Hopital Notre-Dame, Centre Hospitalier de l’Universite deMontreal, Montreal, Quebec, Canada; 2Denis Schrier, MD, CraigFlory, PhD: Pfizer Global Research & Development, Ann Arbor,Michigan.
Address correspondence and reprint requests to Jean-PierrePelletier, MD, Osteoarthritis Research Unit, Hopital Notre-Dame,Centre Hospitalier de l’Universite de Montreal, 1560 rue SherbrookeEst, Montreal, Quebec H2L 4M1, Canada. E-mail: [email protected].
Submitted for publication March 1, 2002; accepted in revisedform February 13, 2003.
1582

ing to a specific membrane-bound receptor, these cyto-kines initiate the activation of a number of intracellularsignaling pathways that lead to the synthesis of transcrip-tion factors that will induce the expression of multiplegenes, including those from catabolic factors (7).
The protein kinase pathways are believed to be ofkey importance among the different signaling systemsactivated by proinflammatory cytokines (7–12). Thesepathways comprise a number of signaling cascades. Thethree most predominant cascades culminate in the acti-vation of the extracellular signal–regulated kinase 1/2(ERK-1/2), c-Jun N-terminal kinase (JNK), and p38families of mitogen-activated protein kinases (MAPKs).These cascades have been demonstrated in several stud-ies to be essential to the synthesis of a number ofcatabolic factors responsible for inducing the structuralchanges seen in OA (7).
ERK-1/2 is activated by MAPK kinase 1/2 (MEK-1/2) as part of the MAPK pathway (7,13). In thispathway, the Raf kinases phosphorylate and activateMEK-1/2, which in turn phosphorylates and activatesERK-1/2. Activated ERK-1/2 can then translocate in thenucleus and activate transduction factors by phosphory-lation, thus altering specific gene expression. In addition,ERK-1/2 has a number of cytosolic substrates that caninfluence gene expression directly or indirectly (13).
The MAPK pathway has also been demonstratedto be a key factor in the induction of matrix metallopro-teinases (MMPs) by cytokines (14), production of cyclo-oxygenase 2 (COX-2) by chondrocytes, and chondrocyteapoptosis (15). The JNK and p38 signaling cascades areimplicated in the synthesis of MMP by chondrocytes(7,14). Moreover, p38 has been shown to be involved inthe expression of the inducible NO synthase (iNOS) (16)and proinflammatory cytokines (17). A recent study hasdemonstrated that an inhibitor of p38 very effectivelyreduced the synthesis in vitro of cytokines and iNOS(16). In another recent study, treatment with a specificp38 inhibitor in a rat model of inflammatory arthritiswas found to reduce the progression of structural dam-age while simultaneously reducing the synthesis of proin-flammatory cytokines (18).
The primary goal of the present study was toexamine, using a rabbit experimental model of OA, theeffect of a specific inhibitor of MEK-1/2, PD 198306, onthe development of structural changes in articular tis-sues. We also examined the effect of the inhibitor on thephosphorylation level of ERK-1/2, as well as on some ofthe major pathophysiologic pathways of OA.
PD 198306 is a potent, selective, non-ATP com-petitive inhibitor of MEK-1/2. It inhibits the isolated
enzyme at a concentration of 8 nM and inhibits MEKactivity in synovial fibroblasts at concentrations of 30–100 nM, depending on the species. The compound ishighly selective for MEK and has a 50% inhibitionconcentration of �1 �M for ERK, �4 �M for c-Src, �10�M for phosphatidylinositol 3-kinase �, and �4 �M forcyclin-dependent kinases. PD 198306 has a bioavailabil-ity of 62% when taken orally and is active in severalanimal models of rheumatoid arthritis, including ratstreptococcal cell wall–induced arthritis (median effec-tive dose [ED50] 11.2 mg/kg) and rat adjuvant arthritis(ED50 6.6 mg/kg) according to an internal file of PfizerGlobal Research & Development (Ann Arbor, MI).Figure 1 shows the chemical structure of PD 198306.
MATERIALS AND METHODS
Surgery. Forty-six white New Zealand male rabbits(ages 16–18 weeks, weighing 2.5–3 kg) were used in this study.Sectioning of the anterior cruciate ligament (ACL) of the rightknee was performed on all rabbits through a medial parapa-tellar incision, as previously described (19,20). Briefly, rabbitswere anesthetized with ketamine (30 mg/kg; Wyeth-Ayerst,Montreal, Quebec, Canada), xylazine (5 mg/kg; Bayer, Etobi-coke, Ontario, Canada), and intramuscular acepromazine mal-eate (1 mg; Wyeth-Ayerst). Under sterile conditions, an an-teromedial incision of the right knee was performed. Thesubcutaneous tissue and retinaculum were incised and re-tracted, along with the articular capsule. The medial compart-ment was visualized, and the ACL was sectioned with a scalpel.The capsule, medial retinaculum, and skin were sutured. Allrabbits were housed in regular individual cages, fed ad libitum,and allowed to exercise.
The rabbits were randomly distributed into three ex-perimental groups. Group 1 (n � 20) consisted of OA rabbitsthat received placebo treatment (equivalent volume of vehicleonly). Groups 2 and 3 (n � 10 per group) consisted of OArabbits given PD 198306 at a dosage of 10 mg/kg/day or 30mg/kg/day, respectively. PD 198306 was administered oncedaily at 8:00 AM as a liquid solution, by oral gavage into thestomach. The drug was dissolved in a 3-ml volume of 1.25%hydroxypropyl cellulose and 0.05% sodium lauryl sulfate,which was prepared fresh daily before administration. Treat-ment was initiated the day after surgery and continued for 8weeks, at which time the rabbits were killed. The animal carepersonnel were blinded to the treatment groups. Four rabbitsdid not complete the study. One animal from group 1 had awound infection. The other 3 rabbits, 1 in group 2 and 2 ingroup 3, had traumatic injuries during the course of the study.Six additional unoperated normal rabbits were used as controlsand were killed at the same time as the animals in theexperimental groups. The study protocol was approved by theInstitutional Review Board.
Drug dosage and pharmacokinetic study. Dosage lev-els for this study were determined by comparing plasma levelsof PD 198306 from a rabbit pharmacokinetic study withefficacious plasma levels in rat models of arthritis. In the
SELECTIVE INHIBITION OF MEK-1/2 IN EXPERIMENTAL OA 1583

pharmacokinetic study, plasma samples were taken 1, 2, 4, 8,and 24 hours after a single oral dose of the compound.Exposures of PD 198306 were also determined during thepresent study by taking plasma samples between 1 hour and 24hours after the last dose.
PD 198306 plasma levels were analyzed by a liquid/liquid extraction procedure using 650 �l of methyl t-butyl ether(95%/5% ethanol), 75 �l of 0.1% acetic acid, and a 75-�lsample. Samples were shaken 15–20 minutes and were thencentrifuged at 4,000 revolutions per minute for 15 minutes.Ether (550 �l) was transferred via a 96-well multipipette(Tomtec, Hamden, CT) to a 96-well block (1 ml/well), thenevaporated under N2. The samples were reconstituted with 100�l of a 60:40 solution of acetonitrile (ACN; Fisher Scientific,Nepean, Ontario, Canada) to 0.1% acetic acid and thenassessed by liquid chromatography/mass spectrometry on aBetasil (Thermo Electron Corporation, Waltham, MA) phenyl2.1 � 10 � 3–�m column (mobile phase 50:50 ACN:0.1%formic acid, flow rate 0.25 ml/minute, ionization electrospraypositive-ion mode, and injection volume 2 �l).
Macroscopic grading of lesions. Immediately after thetreatments were discontinued, the rabbits were killed and theright and left knees were dissected. Each knee was examinedfor gross morphologic changes, including the presence ofosteophyte formation and cartilage lesions, as previously de-scribed (21,22). The schedule for killing the animals was basedon the randomization protocol, and examinations were per-formed by two independent observers who were blinded to thetreatment group. The degree of osteophyte formation wasgraded by measuring the maximum width (in mm) of the spuron the medial and lateral femoral condyles using a Digimaticdigital caliper (Mitutoyo, Kawasaki, Japan). These 2 valuesrecorded for each rabbit were considered separately for thepurpose of statistical analysis. The cartilage changes on themedial and lateral femoral condyles and tibial plateaus weregraded separately, using a dissecting microscope (Stereozoom;Bausch & Lomb, Rochester, NY).
The depth (grade) of erosion was graded on a scale of0–4, where 0 � normal-appearing surface, 1 � minimalfibrillation or a slight yellowish discoloration of the surface,2 � erosion extending into superficial or middle layers, 3 �erosion extending into the deep layers, and 4 � erosionextending to the subchondral bone. The surface area (size) of
the articular surface changes was measured and expressed inmm2. A macroscopic total joint score was also obtained byadding the mean scores of cartilage lesions from the medialand lateral condyles together with those from the medial andlateral plateaus.
Histologic grading. Histologic evaluation was per-formed on sagittal sections of cartilage from the lesion areas ofeach femoral condyle and tibial plateau of the right knees inOA rabbits and from equivalent sites in normal rabbits asdescribed previously (21,22). Specimens were dissected, fixedin 10% neutral buffered formalin, and embedded in paraffinfor histologic evaluation. Serial sections (5 �m) were stainedwith Safranin O. The severity of the OA lesions was graded ona scale of 0–14 by two independent observers under blindedconditions, using the histologic/histochemical scale of Mankinet al (23). This scale evaluates the severity of OA lesions basedon the loss of staining with Safranin O (scale of 0–4), cellularchanges (scale of 0–3), invasion of the tidemark by bloodvessels (scale of 0–1), and structural changes (scale of 0–6,where 0 � normal cartilage structure and 6 � erosion of thecartilage down to the subchondral bone). The scoring wasbased on the most severe histologic changes within eachcartilage section.
Representative specimens of synovial membrane fromthe medial and lateral compartments of the knee were alsodissected from the underlying tissues. The specimens werefixed in 10% buffered formalin, embedded in paraffin, sec-tioned (5-�m sections), and stained with hematoxylin andeosin. Two synovial membrane specimens were examined foreach compartment, and the highest score from each compart-ment was recorded. The average was calculated and consid-ered as a unit for the whole knee. The severity of synovitis wasgraded on a scale of 0–10 (24) by two independent observersunder blinded conditions, adding the scores for 3 histologiccriteria: synovial lining cell hyperplasia (scale of 0–2), villoushyperplasia (scale of 0–3), and degree of cellular infiltration bymononuclear and polymorphonuclear cells (scale of 0–5).
Immunohistochemical studies. Cartilage specimensfrom condyles and plateaus were processed for immunohisto-chemical analysis as previously described (25,26). Briefly,specimens were fixed with TissuFix #2 (Laboratoires GillesChaput, Montreal, Quebec, Canada) for 24 hours and werethen embedded in paraffin. Sections (5 �m) of paraffin-embedded specimens were placed on slides (Superfrost Plus;Fisher Scientific), deparaffinized in toluene, hydrated in aseries of graded dilutions of ethanol, and preincubated withchondroitinase ABC (0.25 units/ml) in phosphate bufferedsaline (PBS) for 60 minutes at 37°C, citrate buffer (0.01 mM,pH 6.0) for 20 minutes, and Triton X-100 (0.3%) for 30minutes. The specimens were washed in PBS and then in 2%H2O2/PBS for 30 minutes. Slides were further incubated withUniversal Blocking Solution with Avidin Blocking Solution(0.2 ml/ml; Vector, Burlingame, CA) in PBS for 15 minutes,blotted, and overlaid with a mouse monoclonal antibodyagainst MMP-1 (100 �g/ml, dilution 1:1,500; Calbiochem-Novabiochem, San Diego, CA) or a mouse monoclonal anti-body against phosphorylated ERK-1/2 (phospho–ERK-1/2)(40 �g/ml, dilution 1:25; Cell Signaling Technology, Beverly,MA) for 18 hours at 4°C in a humidified chamber.
Each slide was washed 3 times in PBS (pH 7.4) andstained using the avidin–biotin complex method (Vectastain
Figure 1. Chemical structure of PD 198306.
1584 PELLETIER ET AL
ABC Kit; Dako Diagnostics Canada, Mississauga, Ontario,Canada). This method entails incubation in the presence of thebiotin-conjugated secondary antibody for 60 minutes at roomtemperature, followed by the addition of the avidin–biotin–peroxidase complex for 45 minutes. All incubations werecarried out in a humidified chamber, and color was developedwith 3,3�-diaminobenzidine (Dako Diagnostics Canada) con-taining H2O2.
To determine the specificity of staining, 3 controlprocedures were used, according to the same experimentalprotocol: 1) use of adsorbed immune serum (1 hour at 37°C)with a 20-fold molar excess of recombinant or purified antigen,2) omission of the primary antibody, and 3) substitution of theprimary antibody with an autologous preimmune serum. Theantigens used in our study were purified human MMP-1(Calbiochem-Novabiochem) and the phospho–Thr 202/Thr204 peptide (Cell Signaling Technology). Several sections weremade from each block of cartilage, and slides from eachspecimen were processed for immunohistochemical analysis.Each section was examined under a light microscope (LeitzOrthoplan; Wild Leitz, Ville St. Laurent, Quebec, Canada)and photographed with Ektachrome 64 ASA film (EastmanKodak, Rochester, NY).
Morphometric analysis. Three sections from each fem-oral condyle and tibial plateau specimen were examined usinga Leitz Diaplan microscope (40�; Wild Leitz), and eachsection was scored separately. The number of chondrocytesstaining positive in the upper zone (superficial and upperintermediate layers of cartilage) for MMP-1 or phospho–ERK-1/2 was estimated as previously described (25–27). The analysiswas performed as previously described (28) only in the super-ficial layers of cartilage, since the majority of OA chondrocytesstaining positive for MMP-1 (26,28) or phospho–ERK-1/2 arelocated in these layers. Briefly, each section was divided into 3different areas at the superficial layers of cartilage. For eachspecimen, it was ensured before the evaluation that an intactcartilage surface at the margin of the lesion could be detectedand used as a marker to validate the morphometric analysis.
The cell count scores were determined separately forthe lateral and medial sides of the condyles and plateaus. Thetotal number of chondrocytes and the number of chondrocytesstaining positive were then quantitated separately for thesuperficial zone. The final results were expressed as thepercentage of positive chondrocytes. The maximum score for
each cartilage specimen was 100%. Each slide was evaluated bytwo independent observers under blinded conditions; variationbetween the observers’ findings was �5%.
Statistical analysis. All data were expressed as themean � SEM or as the median and range and analyzed withthe Mann-Whitney U test when appropriate. P values less than0.05 were considered significant.
RESULTS
Exposure data. At a dosage of 10 mg/kg/day, PD198306 gave a maximum concentration (Cmax) and anarea under the curve (AUC; 0–24 hours) of 3.7 �g/mland 24.2 �g/ml, respectively. At 30 mg/kg/day, the Cmaxand AUC (0–24 hours) values were 5.9 �g/ml and 48.7�g/ml, respectively.
Macroscopic and microscopic findings. Osteo-phytes. In placebo-treated OA rabbits, osteophytes werepresent on both sides of the condyles in all animals, withmean � SEM widths were 2.2 � 0.3 mm and 1.4 � 0.2mm for the medial and lateral condyles, respectively. InOA rabbits treated with PD 198306 at 10 mg/kg/day, theincidence of osteophytes was 94% and the medial andlateral condyle widths were 2.1 � 0.3 mm and 1.1 � 0.2mm, respectively. In OA rabbits treated with 30 mg/kg/day, the incidence was 69% and the medial and lateralcondyle widths were 2.4 � 0.4 mm and 0.5 � 0.3 mm.The osteophytes on the lateral condyles of rabbitstreated with the highest dosage of the drug were foundto be smaller (P � 0.05) compared with those of theplacebo-treated group.
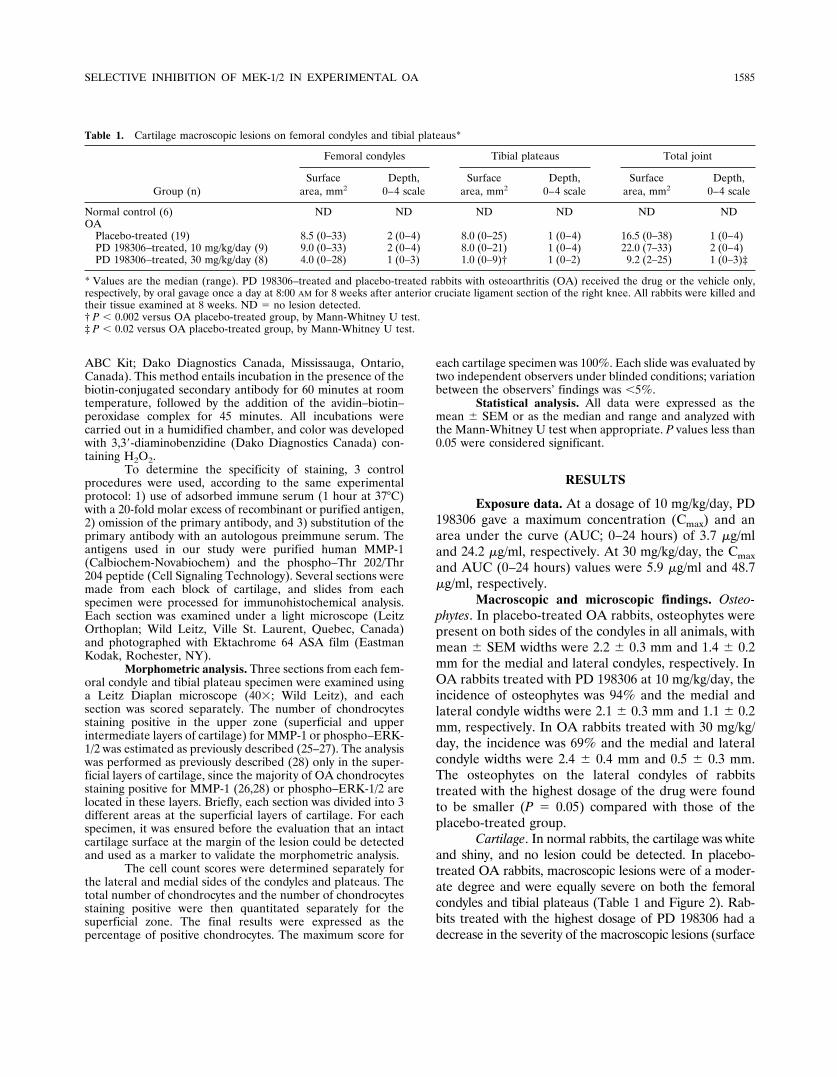
Cartilage. In normal rabbits, the cartilage was whiteand shiny, and no lesion could be detected. In placebo-treated OA rabbits, macroscopic lesions were of a moder-ate degree and were equally severe on both the femoralcondyles and tibial plateaus (Table 1 and Figure 2). Rab-bits treated with the highest dosage of PD 198306 had adecrease in the severity of the macroscopic lesions (surface
Table 1. Cartilage macroscopic lesions on femoral condyles and tibial plateaus*
Group (n)
Femoral condyles Tibial plateaus Total joint
Surfacearea, mm2
Depth,0–4 scale
Surfacearea, mm2
Depth,0–4 scale
Surfacearea, mm2
Depth,0–4 scale
Normal control (6) ND ND ND ND ND NDOA
Placebo-treated (19) 8.5 (0–33) 2 (0–4) 8.0 (0–25) 1 (0–4) 16.5 (0–38) 1 (0–4)PD 198306–treated, 10 mg/kg/day (9) 9.0 (0–33) 2 (0–4) 8.0 (0–21) 1 (0–4) 22.0 (7–33) 2 (0–4)PD 198306–treated, 30 mg/kg/day (8) 4.0 (0–28) 1 (0–3) 1.0 (0–9)† 1 (0–2) 9.2 (2–25) 1 (0–3)‡
* Values are the median (range). PD 198306–treated and placebo-treated rabbits with osteoarthritis (OA) received the drug or the vehicle only,respectively, by oral gavage once a day at 8:00 AM for 8 weeks after anterior cruciate ligament section of the right knee. All rabbits were killed andtheir tissue examined at 8 weeks. ND � no lesion detected.† P � 0.002 versus OA placebo-treated group, by Mann-Whitney U test.‡ P � 0.02 versus OA placebo-treated group, by Mann-Whitney U test.
SELECTIVE INHIBITION OF MEK-1/2 IN EXPERIMENTAL OA 1585
only) on the femoral condyles and tibial plateaus. More-over, the total joint score was found to be significantlylower for the grade of lesions in animals treated with the
highest dosage of PD 198306 (P � 0.02). Histology ofcartilage from normal rabbits demonstrated a normalmicroscopic appearance (Figure 2). In placebo-treated OA
Figure 2. Macroscopic appearance at 8 weeks of cartilage from the femoral condyles and tibial plateaus of normal, placebo-treated osteoarthritic(OA), and PD198306-treated OA rabbits. No lesions were detected in the normal rabbits. Erosions and pitting of the condyles and tibial plateauswere evident in the placebo-treated OA rabbits. In PD198306-treated OA rabbits, a decrease in the severity of lesions on the condyles and plateauswas seen with the highest drug dosage tested (30 mg/kg/day). P � posterior; A � anterior.
1586 PELLETIER ET AL
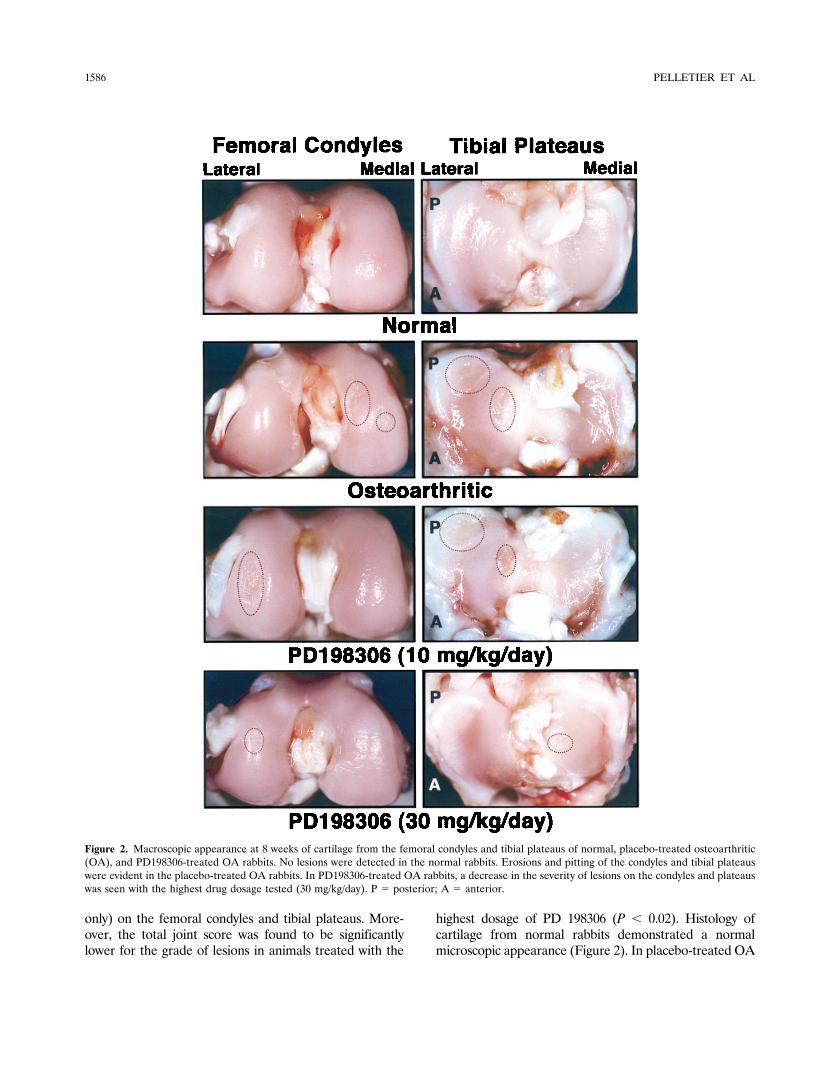
rabbits, the histologic lesions were of moderate degreeand were more severe on the condyles than on theplateaus (mean � SEM scores of 6.2 � 0.6 and 4.1 � 0.6,respectively) (Figure 3). Rabbits treated with the lowestdosage of PD 198306 had a slight increase in the severityof lesions on the condyles and plateaus (scores of 7.8 �0.7 and 4.7 � 0.8, respectively). However, these differ-ences were not statistically significant. In rabbits treatedwith the highest dosage of the inhibitor (30 mg/kg/day),
the severity scores of histologic lesions on condyles andplateaus were 5.3 � 0.8 and 2.5 � 0.5, respectively.
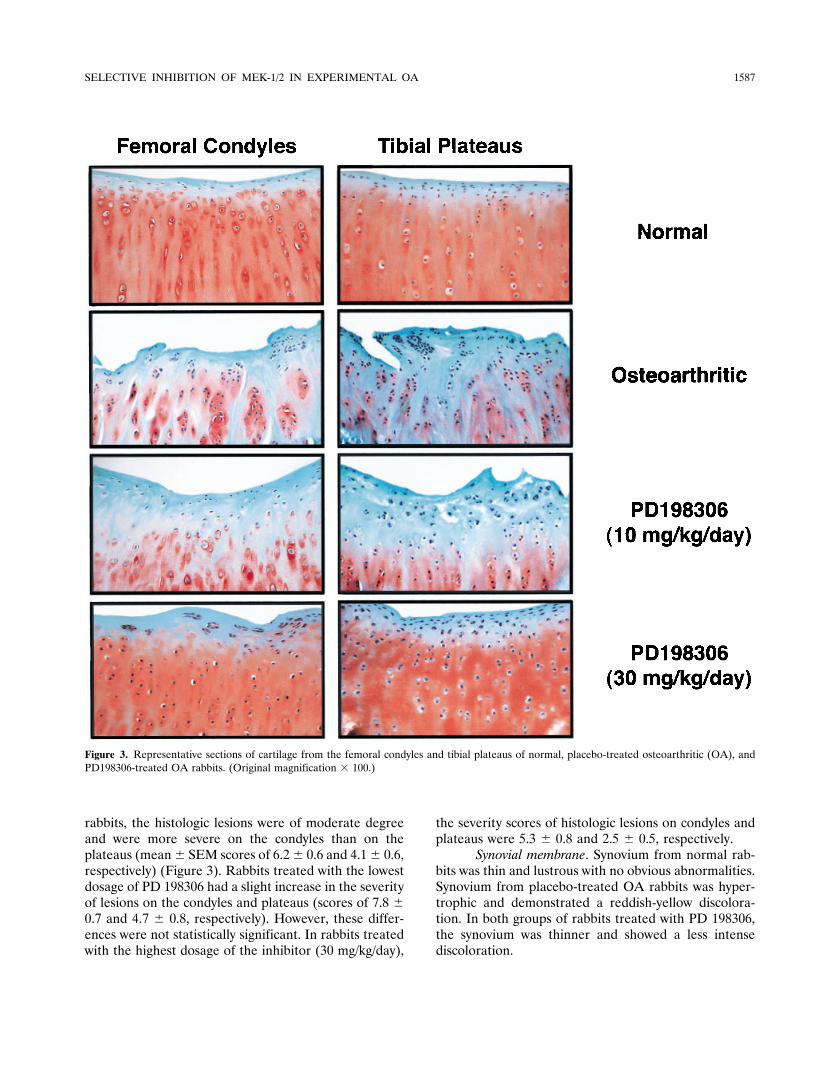
Synovial membrane. Synovium from normal rab-bits was thin and lustrous with no obvious abnormalities.Synovium from placebo-treated OA rabbits was hyper-trophic and demonstrated a reddish-yellow discolora-tion. In both groups of rabbits treated with PD 198306,the synovium was thinner and showed a less intensediscoloration.
Figure 3. Representative sections of cartilage from the femoral condyles and tibial plateaus of normal, placebo-treated osteoarthritic (OA), andPD198306-treated OA rabbits. (Original magnification � 100.)
SELECTIVE INHIBITION OF MEK-1/2 IN EXPERIMENTAL OA 1587
Histology of synovium from normal rabbitsshowed no abnormalities. The synovium from placebo-treated OA rabbits showed a moderate degree of inflam-
mation with synovial lining cell hypertrophia, villoushyperplasia, and a moderate mononuclear cell infiltra-tion of the sublining tissue (Table 2 and Figure 4). The
Figure 4. Representative sections of synovial membrane from normal, placebo-treated osteoarthritic (OA), and PD198306-treated OA rabbits.(Original magnification � 250.)
Table 2. Histologic/histochemical grading of synovial membrane*
Group (n)Synovial lining,
0–2 scale
Villoushyperplasia,
0–3 scale
MNCinfiltration,0–4 scale
PMNinfiltration,0–1 scale
Total,0–10 scale
Normal control (6) ND ND ND ND NDOA
Placebo-treated (19) 1 (0–1) 2 (1–3) 1 (0–2) 0 4 (2–6)PD 198306–treated, 10 mg/kg/day (9) 1 (0–1) 1 (1–2)† 0 (0–1) 0 2 (1–4)‡PD 198306–treated, 30 mg/kg/day (8) 1 (0–1) 1 (1–2)§ 1 (0–1) 0 2 (2–4)§
* Values are the median (range). MNC � mononuclear cell; PMN � polymorphonuclear cell (see Table 1 for other definitions and explanations).† P � 0.001 versus OA placebo-treated group, by Mann-Whitney U test.‡ P � 0.003 versus OA placebo-treated group, by Mann-Whitney U test.§ P � 0.02 versus OA placebo-treated group, by Mann-Whitney U test.
1588 PELLETIER ET AL

two groups of rabbits treated with the PD 198306showed significant reductions in the severity of synovialinflammation. The reductions resulted primarily from aless marked degree of synovial hyperplasia (Table 2).
Immunohistochemistry. Experiments performedwith the antibody that specifically recognized phospho–ERK-1/2 (Table 3 and Figure 5) demonstrated that incartilage specimens from normal rabbits, only a few cells
Figure 5. Detection of phosphorylated extracellular signal–regulated kinase 1/2 in representative sections of cartilage from the femoral condylesand tibial plateaus of normal, placebo-treated osteoarthritic (OA), and PD198306-treated OA rabbits. The reaction revealed that in normal cartilage,only a very small number of cells in the superficial layers stained positive. In contrast, a large number of chondrocytes in placebo-treated OA rabbitsstained positive. Specific nuclear staining was seen in chondrocytes, with maximum intensity in the superficial and upper intermediate layers(superficial zone). In this zone, there was a noticeable decrease in the number of positive chondrocytes in rabbits treated with the highest drug dosagetested (30 mg/kg/day). No specific staining was detected in OA cartilage treated with immunoadsorbed serum. (Original magnification � 250.)
Table 3. Phosphorylated ERK-1/2, MMP-1, and TUNEL reaction levels in rabbit knee cartilage*
Group (n)
ERK-1/2–positive cells, % MMP-1–positive cells, %
Femoral condyles Tibial plateaus Femoral condyles Tibial plateaus
Normal control (6) 3.6 � 0.6† 3.0 � 0.3 6.8 � 0.9† 6.5 � 0.6†OA
Placebo-treated (6) 31.0 � 3.8 30.7 � 1.9 46.9 � 6.1 49.2 � 6.8PD 198306–treated, 10 mg/kg/day (6) 19.5 � 2.2‡ 24.4 � 1.5§ 41.9 � 6.9 43.5 � 6.8PD 198306–treated, 30 mg/kg/day (6) 14.1 � 1.0¶ 15.2 � 1.0† 34.5 � 4.5 37.9 � 1.9
* Values are the mean � SEM. ERK-1/2 � extracellular signal–regulated kinase 1/2; MMP-1 � matrix metalloproteinase 1; OA � osteoarthritis(see Table 1 for explanations).† P � 0.0001 versus OA placebo-treated group, by Mann-Whitney U test.‡ P � 0.04 versus OA placebo-treated group, by Mann-Whitney U test.§ P � 0.06 versus OA placebo-treated group, by Mann-Whitney U test.¶ P � 0.001 versus OA placebo-treated group, by Mann-Whitney U test.
SELECTIVE INHIBITION OF MEK-1/2 IN EXPERIMENTAL OA 1589
within the superficial layers stained positive for thekinase. In specimens from placebo-treated OA rabbits, alarge number of cells, mainly from the superficial upperand upper middle zones of cartilage, showed positivestaining that was located largely in the chondrocytenuclei. Specimens from rabbits treated with PD 198306demonstrated significant decreases in the numbers ofchondrocytes staining positive for phospho–ERK-1/2 atboth dosages tested; however, the decrease was morepronounced in the group treated with the highest dosageof the drug. OA cartilage treated with immunoadsorbedserum demonstrated only background staining (Figure5). For the morphometric analysis, the average totalnumber of cells counted was 190 for the femoral con-dyles and 242 for the tibial plateaus.
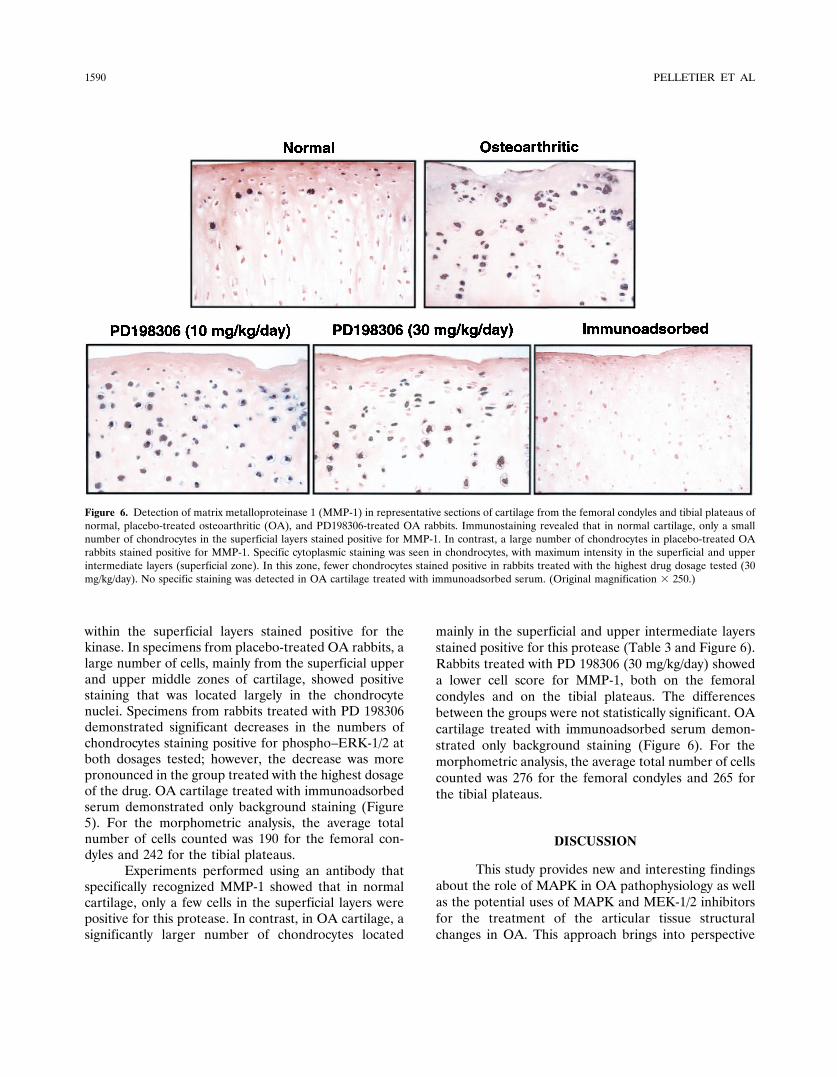
Experiments performed using an antibody thatspecifically recognized MMP-1 showed that in normalcartilage, only a few cells in the superficial layers werepositive for this protease. In contrast, in OA cartilage, asignificantly larger number of chondrocytes located
mainly in the superficial and upper intermediate layersstained positive for this protease (Table 3 and Figure 6).Rabbits treated with PD 198306 (30 mg/kg/day) showeda lower cell score for MMP-1, both on the femoralcondyles and on the tibial plateaus. The differencesbetween the groups were not statistically significant. OAcartilage treated with immunoadsorbed serum demon-strated only background staining (Figure 6). For themorphometric analysis, the average total number of cellscounted was 276 for the femoral condyles and 265 forthe tibial plateaus.
DISCUSSION
This study provides new and interesting findingsabout the role of MAPK in OA pathophysiology as wellas the potential uses of MAPK and MEK-1/2 inhibitorsfor the treatment of the articular tissue structuralchanges in OA. This approach brings into perspective
Figure 6. Detection of matrix metalloproteinase 1 (MMP-1) in representative sections of cartilage from the femoral condyles and tibial plateaus ofnormal, placebo-treated osteoarthritic (OA), and PD198306-treated OA rabbits. Immunostaining revealed that in normal cartilage, only a smallnumber of chondrocytes in the superficial layers stained positive for MMP-1. In contrast, a large number of chondrocytes in placebo-treated OArabbits stained positive for MMP-1. Specific cytoplasmic staining was seen in chondrocytes, with maximum intensity in the superficial and upperintermediate layers (superficial zone). In this zone, fewer chondrocytes stained positive in rabbits treated with the highest drug dosage tested (30mg/kg/day). No specific staining was detected in OA cartilage treated with immunoadsorbed serum. (Original magnification � 250.)
1590 PELLETIER ET AL

the possibility of pharmacologic intervention with regardto specific pathophysiologic targets.
The last few decades of research in the field ofOA have allowed us not only to better characterize themorphologic changes taking place during the course ofthe disease, but also to discover a number of veryimportant mechanisms responsible for such changes(1–4). For instance, the participation of synovial inflam-mation in the progression of cartilage changes at theclinical stage of the disease is becoming increasinglyobvious (4). There are now a number of clinical studiesthat demonstrate a clear association between inflamma-tion and disease progression (29–32). A number offactors synthesized within the inflamed synovium areresponsible for stimulating the synthesis of a largernumber of catabolic/anabolic factors, which in turnmediate the structural changes (1–4). Among the differ-ent synovial factors, cytokines, such as IL-1� and TNF�,are believed to play a prime role (6). The activation ofcells by these cytokines is mediated by the binding to aspecific cell membrane surface receptor, which triggersthe activation of a number of complex intracellularsignaling pathways (7,33–36). Other inflammatory fac-tors (e.g., NO) have also been demonstrated to activateintracellular signaling pathways (37). Among these, theprotein kinase pathways are of major importance in theinduction of the synthesis of transcription factors thatmediate the up-regulation of a number of very impor-tant catabolic factors, such as MMP and NO (16,38).
The findings of the present study suggest that PD198306 could effectively reduce the phosphorylation ofMEK-1/2 in situ, since this kinase is immediately up-stream from ERK-1/2 and is the only kinase that hasthus far been demonstrated to be able to phosphorylateERK-1/2 (7,13). The effect of PD 198306 was dosedependent and more pronounced in the highest-dosagetreatment group, which also provides additional evi-dence of this inhibition. Since the MAPK pathway wasinvolved in the synthesis of a number of catabolic factorsparticipating in the development of the structuralchanges in OA, it is plausible that the action of PD198306 was mediated through the inhibition of theirsynthesis. Moreover, the reduction in the severity ofsynovial inflammation, which was related to a reductionin the level of synovial hyperplasia, could probably havealso contributed to this phenomenon. However, whenconsidering the reduction in the histologic score of thesynovial membrane, one must take into account that thesynovial scoring system used in the study gave moreweight to villous hyperplasia and cellular infiltration,
which represented 8 of the 10 points (total score) in thesystem.
We have previously demonstrated in the OAexperimental dog model that agents that could effec-tively reduce synovial inflammation (such as a selectiveiNOS inhibitor) could also simultaneously reduce theappearance/progression of structural changes in OA,including cartilage lesions and osteophyte formation(39). This phenomenon was associated with a concomi-tant reduction in the level of synthesis of IL-1� andinflammatory mediators such as prostaglandin E2(PGE2) and NO (39,40). This finding is most interesting,since the synthesis of PGE2, a well-known factor alsocapable of inducing bone remodeling (41), can beblocked by MEK-1/2 inhibition, which reduces COX-2expression.
Treatment with therapeutic concentrations of PD198306 was also found to be capable of reducing theprogression of cartilage damage. This effect was notedon the surface of the tibial plateaus and the depth oflesions in the total joint, both of which were significantlyreduced. This protective effect was also associated witha better preservation of the cartilage histologic structure.The lower score on the Mankin scale observed in thisstudy was in great part related to this particular finding.However, it should be noted that this scoring systemgives a much greater emphasis to matrix changes, since10 of the 14 points (total score) were for structuralchanges and loss of Safranin O staining. Moreover, theseOA lesions were not always in the same topographiclocation; therefore, the sampling of cartilage specimenscould also explain some of the variation observed in thestudy.
Concerning the role of cartilage catabolic factors,a number of biochemical factors that participate signif-icantly in the degradation of cartilage macromoleculeshave been identified (1–4). Among these factors, theproteases, and more specifically MMP, have been dem-onstrated to be major factors in the catabolism ofcartilage macromolecules (42). In the MMP family,collagenases are believed to be key players in theproteolysis of cartilage type II collagen (43).
Collagenase 1 (MMP-1) has been identified inOA cartilage (both in human and in experimental mod-els) and shown to be synthesized in an increased amountby OA chondrocytes (25,26,43). Previous studies havedemonstrated that there is a preferential synthesis ofMMP-1 by chondrocytes located in the superficial layersin OA cartilage (25,26). These reports are consistentwith the findings of our study. The involvement of theexcess synthesis and activity of MMP-1 and other colla-
SELECTIVE INHIBITION OF MEK-1/2 IN EXPERIMENTAL OA 1591

genases in the structural changes in OA cartilage hasbeen well established (6,43). In the present study, wedemonstrated that treatment with the inhibitor of MEK-1/2 was capable of reducing the number of chondrocytesthat stained positive for MMP-1. However, it should bepointed out that the differences among the OA PD198306–treated groups and the OA placebo-treatedgroup did not reach statistical significance. It is ofinterest to note that the in situ level of synthesis ofMMP-1 in OA cartilage chondrocytes, as assayed byimmunohistochemistry, has been recently shown by Alt-man and Cheung to reflect the level of MMP activity inrabbit OA (44). The differences in the extent to whichPD 198306 inhibits ERK-1/2 and MMP-1 raise a numberof questions about the relationship between the effect ofthis drug on structural changes and its effect on MMP-1synthesis.
A possible explanation for this finding could bethat, since previous studies have demonstrated thatMMP synthesis depends on the simultaneous activationof many of the protein kinase pathways, including theJNK, MEK-1/2, and p38 pathways (7,11,14), it is there-fore understandable that the selective inhibition of asingle protein kinase pathway could have induced only apartial inhibition of MMP synthesis. However, since theMEK-1/2/ERK-1/2 pathway is also involved in the syn-thesis of other catabolic factors by chondrocytes (includ-ing NO [27,38], which is important in inducing cartilagelesions), it is likely that the effect of PD 198306 wasmediated through the inhibition of synthesis of severalother catabolic factors. Further studies are currentlyunder way to explore additional mechanisms of action ofthis drug.
Preliminary findings from this study provide in-formation about potential mechanisms for mediating thestructure-modifying effects of this MEK-1/2 inhibitor.Treatment with the inhibitor was found to reduce theseverity of synovial inflammation, which was essentiallyrelated to a significant reduction in the villous hyperpla-sia. Therefore, the total number of cells (includingsynovial cells and infiltrating mononuclear cells) thatcould have potentially produced catabolic factors, suchas cytokines and MMP, was reduced by treatment withPD 198306. However, caution must be taken not tooverinterpret these results, and the synovial scoringsystem used in the present study must be taken intoaccount. Moreover, a very important study by Firesteinand Manning (7) recently showed that synovial fibro-blasts from OA patients expressed the 3 main MAPKfamilies. That study also showed that MEK-1/2 wasreadily phosphorylated by IL-1 and was responsible for
increasing MMP-1 gene expression in these cells. There-fore, although this is very speculative, it is possible thatthe inhibition of MEK-1/2 in OA synovium could, byreducing cartilage catabolism, also contribute to a reduc-tion in the level of synovial inflammation (1–4).
In summary, the present study has demonstratedthat the activation of the MAPK pathway could probablycontribute to the pathogenesis of OA. Therapeuticintervention with the goal of MEK-1/2 inhibition mayhave interesting potential for the development of agentsfor the treatment of OA.
REFERENCES
1. Pelletier JP, Martel-Pelletier J, Howell DS. Etiopathogenesis ofosteoarthritis. In: Koopman WJ, editor. Arthritis and allied con-ditions: a textbook of rheumatology. Baltimore: Williams &Wilkins; 2001. p. 2195–245.
2. Williams CJ, Jimenez SA. Genetic and metabolic aspects. In:Reginster JY, Pelletier J-P, Martel-Pelletier J, Henrotin Y, edi-tors. Osteoarthritis: clinical and experimental aspects. Berlin:Springer-Verlag; 1999. p. 134–55.
3. Nuki G. Role of mechanical factors in the aetiology, pathogenesisand progression of osteoarthritis. In: Reginster JY, Pelletier J-P,Martel-Pelletier J, Henrotin Y, editors. Osteoarthritis: clinical andexperimental aspects. Berlin: Springer-Verlag; 1999. p. 101–14.
4. Pelletier J-P, Martel-Pelletier J, Abramson SB. Osteoarthritis, aninflammatory disease: potential implication for the selection ofnew therapeutic targets. Arthritis Rheum 2001;44:1237–47.
5. Attur MG, Patel IR, Patel RN, Abramson SB, Amin AR. Auto-crine production of IL-1 beta by human osteoarthritis-affectedcartilage and differential regulation of endogenous nitric oxide,IL-6, prostaglandin E2, and IL-8. Proc Assoc Am Physicians1998;110:65–72.
6. Martel-Pelletier J, Di Battista JA, Lajeunesse D. Biochemicalfactors in joint articular tissue degradation in osteoarthritis. In:Reginster JY, Pelletier J-P, Martel-Pelletier J, Henrotin Y, edi-tors. Osteoarthritis: clinical and experimental aspects. Berlin:Springer-Verlag; 1999. p. 156–87.
7. Firestein GS, Manning AM. Signal transduction and transcriptionfactors in rheumatic disease. Arthritis Rheum 1999;42:609–21.
8. Bokemeyer D, Sorokin A, Dunn MJ. Multiple intracellular MAPkinase signaling cascades. Kidney Int 1996;49:1187–98.
9. Treisman R. Regulation of transcription by MAP kinase cascades.Curr Opin Cell Biol 1996;8:205–15.
10. Muller-Ladner U, Gay RE, Gay S. Signaling and effector path-ways. Curr Opin Rheumatol 1999;11:194–201.
11. Ridley SH, Sarsfield SJ, Lee JC, Bigg HF, Cawston TE, Taylor DJ,et al. Actions of IL-1 are selectively controlled by p38 mitogen-activated protein kinase: regulation of prostaglandin H synthase-2,metalloproteinases, and IL-6 at different levels. J Immunol 1997;158:3165–73.
12. Geng Y, Valbracht J, Lotz M. Selective activation of the mitogen-activated protein kinase subgroups c-Jun NH2 terminal kinase andp38 by IL-1 and TNF in human articular chondrocytes. J ClinInvest 1996;98:2425–30.
13. Dhillon AS, Kolch W. Untying the regulation of the Raf-1 kinase.Arch Biochem Biophys 2002;404:3–9.
14. Brauchle M, Gluck D, Di Padova F, Han J, Gram H. Independentrole of p38 and ERK1/2 mitogen-activated kinases in the upregu-lation of matrix metalloproteinase-1. Exp Cell Res 2000;258:135–44.
15. Notoya K, Jovanovic DV, Reboul P, Martel-Pelletier J, Mineau F,
1592 PELLETIER ET AL

Pelletier J-P. The induction of cell death in human osteoarthritischondrocytes by nitric oxide is related to the production ofprostaglandin E2 via the induction of cyclooxygenase-2. J Immunol2000;165:3402–10.
16. Badger AM, Cook MN, Lark MW, Newman-Tarr TM, Swift BA,Nelson AH, et al. SB 203580 inhibits p38 mitogen-activatedprotein kinase, nitric oxide production, and inducible nitric oxidesynthase in bovine cartilage-derived chondrocytes. J Immunol1998;161:467–73.
17. Lee JC, Laydon JT, McDonnell PC, Gallagher TF, Kumar S,Green D, et al. A protein kinase involved in the regulation ofinflammatory cytokine biosynthesis. Nature 1994;372:739–46.
18. Badger AM, Griswold DE, Kapadia R, Blake S, Swift BA,Hoffman SJ, et al. Disease-modifying activity of SB 242235, aselective inhibitor of p38 mitogen-activated protein kinase, in ratadjuvant-induced arthritis. Arthritis Rheum 2000;43:175–83.
19. Yoshioka M, Coutts RD, Amiel D, Hacker SA. Characterizationof a model of osteoarthritis in the rabbit knee. OsteoarthritisCartilage 1996;4:87–98.
20. Chang DG, Iverson EP, Schinagl RM, Sonoda M, Amiel D, CouttsRD, et al. Quantitation and localization of cartilage degenerationfollowing the induction of osteoarthritis in the rabbit knee. Osteo-arthritis Cartilage 1997;5:357–72.
21. Pelletier J-P, Di Battista JA, Raynauld JP, Wilhelm S, Martel-Pelletier J. The in vivo effects of intraarticular corticosteroidinjections on cartilage lesions, stromelysin, interleukin-1 and on-cogene protein synthesis in experimental osteoarthritis. Lab Invest1995;72:578–86.
22. Fernandes JC, Martel-Pelletier J, Otterness IG, Lopez-Anaya A,Mineau F, Tardif G, et al. Effects of tenidap on canine experi-mental osteoarthritis. I. Morphologic and metalloprotease analy-sis. Arthritis Rheum 1995;38:1290–303.
23. Mankin HJ, Dorfman H, Lippiello L, Zarins A. Biochemical andmetabolic abnormalities in articular cartilage from osteoarthritichuman hips. II. Correlation of morphology with biochemical andmetabolic data. J Bone Joint Surg Am 1971;53:523–37.
24. Pelletier J-P, Martel-Pelletier J, Ghandur-Mnaymneh L, HowellDS, Woessner JF Jr. Role of synovial membrane inflammation incartilage matrix breakdown in the Pond-Nuki dog model ofosteoarthritis. Arthritis Rheum 1985;28:554–61.
25. Moldovan F, Pelletier J-P, Hambor J, Cloutier J-M, Martel-Pelletier J. Collagenase-3 (matrix metalloprotease 13) is preferen-tially localized in the deep layer of human arthritic cartilage in situ:in vitro mimicking effect by transforming growth factor �. ArthritisRheum 1997;40:1653–61.
26. Fernandes JC, Martel-Pelletier J, Lascau-Coman V, Moldovan F,Jovanovic D, Raynauld JP, et al. Collagenase-1 and collagenase-3synthesis in early experimental osteoarthritic canine cartilage: animmunohistochemical study. J Rheumatol 1998;25:1585–94.
27. Pelletier J-P, Fernandes JC, Jovanovic DV, Reboul P, Martel-Pelletier J. Chondrocyte death in experimental osteoarthritis ismediated by MEK 1/2 and p38 pathways: role of COX-2 andiNOS. J Rheumatol 2001;28:2509–19.
28. Jovanovic DV, Fernandes JC, Martel-Pelletier J, Jolicoeur F-C,Reboul P, Laufer S, et al. In vivo dual inhibition of cyclooxygenaseand lipoxygenase by ML-3000 reduces the progression of experi-mental osteoarthritis: suppression of collagenase 1 and interleu-kin-1� synthesis. Arthritis Rheum 2001;44:2320–30.
29. Ayral X, Pickering EH, Woodworth TG, Mackillop N, DougadosM. Synovitis predicts the arthroscopic progression of medialtibiofemoral knee osteoarthritis [abstract]. Osteoarthritis Carti-lage 2001;9:S2.
30. Sharif M, Saxne T, Shepstone L, Kirwan JR, Elson CJ, HeinegardD, et al. Relationship between serum cartilage oligomeric matrixprotein levels and disease progression in osteoarthritis of the kneejoint. Br J Rheumatol 1995;34:306–10.
31. Sharif M, Shepstone L, Elson CJ, Dieppe PA, Kirwan JR. In-creased serum C reactive protein may reflect events that precederadiographic progression in osteoarthritis of the knee. Ann RheumDis 2000;59:71–4.
32. Spector TD, Hart DJ, Nandra D, Doyle DV, Mackillop N,Gallimore JR, et al. Low-level increases in serum C-reactiveprotein are present in early osteoarthritis of the knee and predictprogressive disease. Arthritis Rheum 1997;40:723–7.
33. Schett G, Tohidast-Akrad M, Smolen JS, Schmid BJ, Steiner C-W,Bitzan P, et al. Activation, differential localization, and regulationof the stress-activated protein kinases, extracellular signal–regulated kinase, c-Jun N-terminal kinase, and p38 mitogen-activated protein kinase, in synovial tissue and cells in rheumatoidarthritis. Arthritis Rheum 2000;43:2501–12.
34. Winston BW, Lange-Carter CA, Gardner AM, Johnson GL,Riches DW. Tumor necrosis factor alpha rapidly activates themitogen-activated protein kinase (MAPK) cascade in a MAPKkinase kinase-dependent, c-Raf-1-independent fashion in mousemacrophages. Proc Natl Acad Sci U S A 1995;92:1614–-8.
35. Bird TA, Kyriakis JM, Tyshler L, Gayle M, Milne A, Virca GD.Interleukin-1 activates p54 mitogen-activated protein (MAP) ki-nase/stress-activated protein kinase by a pathway that is indepen-dent of p21ras, Raf-1, and MAP kinase kinase. J Biol Chem1994;269:31836–44.
36. Ichijo H. From receptors to stress-activated MAP kinases. Onco-gene 1999;18:6087–93.
37. Beck KF, Eberhardt W, Frank S, Huwiler A, Messmer UK, MuhlH, et al. Inducible NO synthase: role in cellular signalling. J ExpBiol 1999;202:645–53.
38. Martel-Pelletier J, Mineau F, Jovanovic D, Di Battista JA, Pelle-tier J-P. Mitogen-activated protein kinase and nuclear factor �Btogether regulate interleukin-17–induced nitric oxide productionin human osteoarthritic chondrocytes: possible role of transacti-vating factor mitogen–activated protein kinase–activated proteinkinase (MAPKAPK). Arthritis Rheum 1999;42:2399–409.
39. Pelletier J-P, Jovanovic D, Fernandes JC, Manning P, Connor JR,Currie MG, et al. Reduced progression of experimental osteoar-thritis in vivo by selective inhibition of inducible nitric oxidesynthase. Arthritis Rheum 1998;41:1275–86.
40. Pelletier J-P, Lascau-Coman V, Jovanovic D, Fernandes JC,Manning P, Currie MG, et al. Selective inhibition of induciblenitric oxide synthase in experimental osteoarthritis is associatedwith reduction in tissue levels of catabolic factors. J Rheumatol1999;26:2002–14.
41. Kawaguchi H, Pilbeam CC, Harrison JR, Raisz LG. The role ofprostaglandins in the regulation of bone metabolism. Clin Orthop1995;36–46.
42. Martel-Pelletier J, Tardif G, Fernandes JC, Pelletier J-P. Metal-loproteases and their modulation as treatment in osteoarthritis. In:Tsokos GC, editor. Molecular rheumatology. Totowa (NJ): Hu-mana Press; 2000. p. 499–514.
43. Martel-Pelletier J, Pelletier J-P. Wanted—the collagenase respon-sible for the destruction of the collagen network in humancartilage! Br J Rheumatol 1996;35:818–20.
44. Altman RD, Cheung H. A rabbit study of glucosamine sulfate in amodel of osteoarthritis [abstract]. Osteoarthritis Cartilage 2001;9Suppl B:S63.
SELECTIVE INHIBITION OF MEK-1/2 IN EXPERIMENTAL OA 1593
Copyright © 2022 FDOKUMEN




















