
In vivo CTL induction with point-substituted ovalbumin peptides: immunogenicity correlates with...
8
Vaccine, Vol. 13, No. 3, pp. 313320, 1995 Copyright <c, 1995 Elsevier Science Ltd Printed in Great Britain. All rights reserved 0264410x/95 $10.00+0.00 In viva CTL induction with point-substituted ovalbumin peptides: immunogenicity correlates with peptide-induced MHC class I stability Grayson B. Lipford *, Stefan Bauer, Herrman Wagner and Klaus Heeg Class I molecules are conformationally sensitive to peptide binding, prolonging the complex’s half-life on the surface of the cell. By making a series of H2-Kh anchor motif amino acidpoint substitutions in the ovalbumin 257-264 octamer, we were able to analyse subtle changes in peptide binding, Kb stabilization and in vivo immunogenicity. The cell line RMA-S was used to determine peptide-dependent Kb stabilization under equilibrium and non-equilibrium binding conditions. Sixteen conservative and non-conservative amino acid substitutions were made at positions 3,5 or 8 of the peptide. At 37”C, Kb stabilization was dlrerentially aflected by these substitutions, with several substitutions severely aflecting Kb surface expression. When the substituted peptides were used as immunogens to prime cytotoxic T lymphocytes (CTL) in vivo, each peptide’s ability to stabilize K!’ directly correlated with the intensity of spec@ CTL activation. We conclude that peptide class I stabilization is an important influencing factor in determining cell surface steady-state expression of these peptides and thus the breadth of CTL recruitment. These concepts may relate the phenomenon of immunodominance to cell surface-presented peptide steady-state levels and may also aid in peptide vaccine design. Keywords: Vaccination: peptide; cytolytic T cells; immunogenicity CD8+ cytotoxic T lymphocytes (CTL) recognize short peptides that are the product of intracellularly processed ‘self’ and ‘foreign’ proteins and that are presented at the surface by MHC class I molecules’,‘. The repertoire of peptides presented for immunosurveillance is dependent on the complement of several different coexpressed MHC class I molecules, and on the ability of each MHC molecule to bind many peptides of diverse sequence. As a first approximation, the specific priming of CTL is thus limited to MHC class I-binding peptides. The polymorphism among class I molecules is found primarily within the peptide-binding groove, specifically at amino acids that line the groove which form interactions with peptide moieties3. The binding groove is closed at each end, limiting peptide length to 8-10 amino acids. Evidence has been accumulating rapidly which suggests that three-dimensional pockets within the groove accommodate peptide amino acids of conserved size and chemical character, thus forming allele-specific anchor motifs for a given MHC class I molecule3-‘. These Institute for Medical Microbiology, Technical University of Munich, Trogerstrasse 9, Munich 81675, Germany. *To whom correspondence should be addressed. (Received 6 June 1994; revised 27 July 1994; accepted 2 August 1994) motifs can serve as a minimal search rule for identifying MHC class I-binding peptides within a given protein sequence, thus accelerating CTL epitope definitions,9. However, knowledge of the parameters limiting the pool of binding peptides for a given class I molecule is still rudimentary. The murine H-2Kb molecule is the most thoroughly investigated MHC class I allele. The peptide-binding motif for Kb was first postulated to be tyrosine or phenylalanine at position 5 and leucine at the carboxy terminal, position 8’. A second report, which focused on the Kb-binding peptide vesicular stomatitis virus NP-52-59 (VSV 59-59), determined that tyrosine at position 3 was essential for binding and T-cell receptor recognition”. Crystallization of Kb together with the peptides Sendai virus NP-322-330 (SV9), VSV 52-59, or ovalbumin (OVA) 257-264, bound in the groove, was utilized for X-ray crystallographic measurements and has yielded stuctural models of this molecule5.6.’ I. The amino acids in peptide positions 5 and 8 appear to fit into deep pockets, the C and F pockets respectively, forming secure anchors. However, the amino acid at position 3 appears to be nested within a more shallow groove, the D pocket. As revealed by crystallography, local conformational change within Kb can occur around the D pocket in Vaccine 1995 Volume 13 Number 3 313
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of In vivo CTL induction with point-substituted ovalbumin peptides: immunogenicity correlates with...

Vaccine, Vol. 13, No. 3, pp. 313320, 1995 Copyright <c, 1995 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0264410x/95 $10.00+0.00
In viva CTL induction with point-substituted ovalbumin peptides: immunogenicity correlates with peptide-induced MHC class I stability
Grayson B. Lipford *, Stefan Bauer, Herrman Wagner and Klaus Heeg
Class I molecules are conformationally sensitive to peptide binding, prolonging the complex’s half-life on the surface of the cell. By making a series of H2-Kh anchor motif amino acidpoint substitutions in the ovalbumin 257-264 octamer, we were able to analyse subtle changes in peptide binding, Kb stabilization and in vivo immunogenicity. The cell line RMA-S was used to determine peptide-dependent Kb stabilization under equilibrium and non-equilibrium binding conditions. Sixteen conservative and non-conservative amino acid substitutions were made at positions 3,5 or 8 of the peptide. At 37”C, Kb stabilization was dlrerentially aflected by these substitutions, with several substitutions severely aflecting Kb surface expression. When the substituted peptides were used as immunogens to prime cytotoxic T lymphocytes (CTL) in vivo, each peptide’s ability to stabilize K!’ directly correlated with the intensity of spec@ CTL activation. We conclude that peptide class I stabilization is an important influencing factor in determining cell surface steady-state expression of these peptides and thus the breadth of CTL recruitment. These concepts may relate the phenomenon of immunodominance to cell surface-presented peptide steady-state levels and may also aid in peptide vaccine design.
Keywords: Vaccination: peptide; cytolytic T cells; immunogenicity
CD8+ cytotoxic T lymphocytes (CTL) recognize short peptides that are the product of intracellularly processed ‘self’ and ‘foreign’ proteins and that are presented at the surface by MHC class I molecules’,‘. The repertoire of peptides presented for immunosurveillance is dependent on the complement of several different coexpressed MHC class I molecules, and on the ability of each MHC molecule to bind many peptides of diverse sequence. As a first approximation, the specific priming of CTL is thus limited to MHC class I-binding peptides.
The polymorphism among class I molecules is found primarily within the peptide-binding groove, specifically at amino acids that line the groove which form interactions with peptide moieties3. The binding groove is closed at each end, limiting peptide length to 8-10 amino acids. Evidence has been accumulating rapidly which suggests that three-dimensional pockets within the groove accommodate peptide amino acids of conserved size and chemical character, thus forming allele-specific anchor motifs for a given MHC class I molecule3-‘. These
Institute for Medical Microbiology, Technical University of Munich, Trogerstrasse 9, Munich 81675, Germany. *To whom correspondence should be addressed. (Received 6 June 1994; revised 27 July 1994; accepted 2 August 1994)
motifs can serve as a minimal search rule for identifying MHC class I-binding peptides within a given protein sequence, thus accelerating CTL epitope definitions,9. However, knowledge of the parameters limiting the pool of binding peptides for a given class I molecule is still rudimentary.
The murine H-2Kb molecule is the most thoroughly investigated MHC class I allele. The peptide-binding motif for Kb was first postulated to be tyrosine or phenylalanine at position 5 and leucine at the carboxy terminal, position 8’. A second report, which focused on the Kb-binding peptide vesicular stomatitis virus NP-52-59 (VSV 59-59), determined that tyrosine at position 3 was essential for binding and T-cell receptor recognition”. Crystallization of Kb together with the peptides Sendai virus NP-322-330 (SV9), VSV 52-59, or ovalbumin (OVA) 257-264, bound in the groove, was utilized for X-ray crystallographic measurements and has yielded stuctural models of this molecule5.6.’ I. The amino acids in peptide positions 5 and 8 appear to fit into deep pockets, the C and F pockets respectively, forming secure anchors. However, the amino acid at position 3 appears to be nested within a more shallow groove, the D pocket. As revealed by crystallography, local conformational change within Kb can occur around the D pocket in
Vaccine 1995 Volume 13 Number 3 313

Dependence of immunogenicity on peptide-induced MHC class I stability: G.B. Lipford et al.
response to the amino acid at peptide position 3 and thus potentially influence Kb conformation or responding T-cell receptor usage6. A peptide-induced conformational change of MHC class I has been demonstrated serologically’2-‘6. In contrast to VSV 52-59, which utilizes tyrosine for D pocket interactions, OVA 257-264 utilizes isoleucine, while SV9 utilizes proline. The side chains of these three amino acids are of different chemical character and thus could be expected to influence peptide binding or Kb conformation in dissimilar ways.
Vaccination with synthetic peptides which induce CTL has been shown experimentally to protect against bacterial, viral and tumour challenge’7-20. These peptide sequences have been determined from known antigenic proteins by synthesizing short overlapping peptide sequences or deduced through prediction using allele- specific binding motifs. For the purposes of T-cell epitope prediction models, it would be desirable to understand fully what parameters influence class I-binding peptide repertoires and perhaps more importantly how these peptides influence T-cell receptor usage and thus immunorecognition. Studies have been conducted which examined point-substituted Kb-binding peptides for altered binding and also T-cell clone responsiveness2rmz3. In this report, we have analysed point substitution changes in the anchor motif for their influence on Kb peptide binding, class I stabilization and immunogenicity as monitored by primary immune responses induced by in uiuo immunization with peptide entrapped in Quil-A liposomes24. Special emphasis was given to point substitutions at position 3 because of ambiguity as to its importance as an anchor position and its potential effects on Kb conformation. Conclusions were drawn which are imnortant to all neotide-MHC class I interactions and
by Lipford et al. 24. Briefly, 18.0 mg p hosphatidylcholine 2.0 mg phosphatidylglycerol and 5.0 mg cholesterol, in a 2:0.2: 1 molar ratio, were suspended in 5.0 ml chloroform in a 1OOml round-bottomed flask. The mixture was rotary-evaporated under reduced pressure until a thin lipid film formed on the flask wall. Residual chloroform was removed by vacuum desiccation. Each peptide (3 mg) was solubilized in a minimal volume of the appropriate solvent and diluted to l.Oml with phosphate-buffered saline (PBS) containing 0.4 mg Quil-A. This solution was slowly added to the dried lipid and hand shaken until the lipids were resuspended. The crude peptide liposome mixture was allowed to equilibrate for 30min at room temperature, transferred to a microfuge tube and centrifuged at 6000 rev min- ’ for 5 min in an Eppendorf’ microfuge. The mixture was then filter extruded through a 0.2 pm Anotop 10R syringe mount filter (E. Merck, Darmstadt, Germany). Mice were immunized in the hind footpad with 50,nl per foot of peptide liposome preparation. After 4 days, the draining popliteal lymph nodes (LN) were removed and a single-cell suspension was prepared.
Cell culture
Cells were cultured in Click’s RPM1 supplemented with 10% (v/v) heat-inactivated fetal calf serum (Biochrom, Berlin, Germany), 5 x 10m5~ 2-mercaptoethanol, 10 mM L-glutamine, 10 mM HEPES, 50 pg ml- 1 indometha- tine and antibiotics. LN cells were cultured at 3 x 106/ml in 24-well culture plates (Nunc, Wiesbaden, Germany) in the presence of 1OU ml-l rIL-2 for 4 days. The blast cells were harvested by centrifugation over a Ficoll gradient (d= 1.077 kg l- ‘) and tested for lytic activity.
L 1 I
which will potentially influence future subunit vaccine success. “Cr-release assay
MATERIALS AND METHODS
Reagents and cell lines
L-a-phosphatidylcholine (type XI-E), L-a-phosphatidyl- o,L-glycerol, cholesterol and ovalbumin were purchased from Sigma (Munich, Germany). Quil-A (Spikoside) was purchased from Isotec Production AB (Lulea, Sweden). Peptides were custom-synthesized by Neosystem Labora- toire (Strasbourg, France) or by the solid-phase method with an automated peptide synthesizer (430A, Applied Biosystems, Foster City, CA, USA) in the laboratory of Ulrich Koszinowski, University of Ulm, Germany. Human recombinant interleukin-2 (rIL-2) was donated by Eurocetus (Amsterdam, The Netherlands). EL-4 (H-2b) thymoma cells were purchased from ATCC (American Type Culture Collection, Rockville, MD). RMA-S cells were provided by Dr K. Karre (Stockholm, Sweden).
EL-4 target cells were labelled with Na251Cr04 and then washed as previously described25. Peptide was suspended to 1 mM in PBS and further diluted to a working concentration with culture medium. For peptide pulsing, 51Cr-labelled cells were incubated with peptide for 30min at 37°C followed by washing. The OVA 257-264 peptide was pulsed at a concentration of 50 nM, the R-, E- and G-substituted peptides at 10~~ and all remaining peptides at 250 nM. To replicate serial dilutions of effector CTL in 100 ~1, the same volume of target cells (104/ml) was added. Controls contained no effector CTL (spontaneous release), or 100 ~1 of 1% Triton X-100 (maximal release). Effector and target cells were pelleted (509, 2 min) and then incubated for 4 h. Supernatants were removed and y-irradiation was measured. Percent- age specific lysis was calculated as follows: ((experimental release - spontaneous release)/(maximal release - spon- taneous release)) x 100.
Mice RMA-S stabilization assay
C57B1/6 (H-2b) mice were obtained from the Zentral- institut fur Versuchstierforschung, Hannover, Germany. Animals were used at an age of 8-12 weeks.
Peptide liposome formation and immunization
Liposomes were manufactured by a rehydration entrapment method according to the method described
RMA-S cells were cultured into mid-log growth phase at 37°C washed, aliquoted into a 96-well plate at a density of lo5 cells/100~l/well and incubated overnight at 26°C. These cells were pelleted, resuspended in RPM1 medium containing the test peptide at various concen- trations and incubated for 30 min at 26°C. The cells were then transferred to an incubator at 37°C and incubated for the indicated times (&180 min) followed by washes at
314 Vaccine 1995 Volume 13 Number 3

Dependence of immunogenicity on peptide-induced MHC class I stability: G.B. Lipford et al.
4°C in RPMI. For a zero time-point determination, cells were immediately pelleted after the 26°C incubation and washed at 4°C. Following washes, the cells were stained at 4°C for Kb surface expression with anti-Kb monoclonal antibodies (mAbs) and detected with an FITC-conjugated goat anti-mouse Ig as the secondary antibody. FACS analysis was performed on an EPICS@-Profile II (Coulter, Krefeld, Germany). The antibodies used primarily were mAb Y3 (HB 176, ATCC, Rockville, MD, USA) and mAb 28.8.6 (HB 51, ATCC), although mAb 28.13.3 (HB 41, ATCC), mAb B8.24.3 (TIB 139, ATCC), mAb AF6-88.5 (Pharmingen, San Diego, CA, USA) and mAb CL 9007-A (Cedarlane, Hornby, Ontario, Canada) were also tested. All peptide concentration pulsings were performed in triplicate. The conditions of this assay allowed equilibrium peptide-class I binding to occur because peptide was continually present.
A second type of assay was performed as described above, except that cells were pulsed with peptide in the absence of serum, and peptide was removed from the medium by washing the cells three times prior to the 37°C incubation period. The cells were then resuspended in medium at 37°C and incubated for the indicated times (O-1 80 min) at 37°C. The zero time point was determined by immediately resuspending in medium at 4°C containing an anti-Kb antibody. The conditions of this assay allowed free peptide dissociation from class I molecules without the opportunity of reassociation.
RESULTS
Point substitutions in SIINFEKL display differing MHC class I-stabilizing affinities
It has been shown previously that cell-surface MHC class I molecules which lack bound peptide are unstable at 37”C, with a half-life (flj2) of 1 h for unloaded molecules versus 6 h for loaded molecules26.
Recently, an assay for peptide-MHC class I binding has been developed based on class I thermostability utilizing the cell line RMA-S14*2’,23,27. Typically, a saturating dose of 100~~ peptide has been employed in these assays. In an effort to determine the plasticity of the Kb-binding motif, we utilized MHC class I stabilization to determine the effect of point substitutions to the anchor residues of the immunodominant Kb-binding peptide SIINFEKL from ovalbumin (OVA 257-264). Since point substitutions could affect peptide affinity for MHC class I molecules, a titration of peptide versus Kb stabilization was necessary to define potentially small differences.
Figure IA demonstrates that the stability of Kb on the surface of RMA-S cells is peptide concentration- dependent. On non-peptide-pulsed cells, the rate of loss for Kb from the surface followed first-order kinetics with a tli2 of 32min, in agreement with published data26,28. The quantity of Kb on the surface came to steady-state levels, i.e. 37°C equilibrium, within 3 h. The tli2 of Kb loss progressively increased with increasing pulse peptide concentration and the system approached saturation near IO ,UM OVA 257-264 peptide, with a tli2 of 726 min. At concentrations between no added peptide and saturating peptide, the observed loss rate was probably an averaged resultant dependent on the relative percentage of class I with bound peptide versus empty.
Peptide
726
32
OF.,, 0 30 60 90 120 150 180
Time (min)
32
Figure 1 Peptide-dependent kinetics of Kb loss under equilibrium binding conditions. The titration of Kb peptide-dependent stabilization with the peptide OVA 257-264 (SIINFEKL, graph A) or the R3 peptide (SIRNFEKL, graph B) was monitored by mAb Y3 staining versus duration of incubation at 37°C. The concentration of incubating peptide is given adjacent to the plotted curve. Also given are the f1,2 of Kb loss calculated through a first-order rate of decay equation utilizing the curve-fitting function of the graphic program Sigma PlotTM. Points represent the mean of triplicate determinations
Figure ZB demonstrates the effect of a substitution at position 3 of arginine for isoleucine, SIRNFEKL (R3). Comparing Figure IA and B, the Kb loss dynamics are similar; however, the peptide concentrations required to . achieve a slmllar t1,2 are as much as lOO-fold different at low peptide concentrations. These results are more easily demonstrated by plotting the mean fluorescence intensity at 3 h versus peptide concentration (Figure 2). These points form a sigmoidal curve with a 50% maximal stabilization concentration for OVA 257-264 of 4.0n~ when mAb Y3 was used for detection. The R3 peptide yielded a 50% maximal stabilization of 400 nM.
Included within Figure 2 are titration curves collected using mAb 28.8.6. It has been shown that mAb 28.8-6 binding is sensitive to peptide-induced conformational change within Kb in a way distinct from that seen with mAb Y316. In this instance, however, these peptides did not influence Kb detectability. To address the issue of whether the conformation of the MHC class I molecules affected the results of stabilization studies based on serological detection, multiple antibodies were tested for Kb monitoring in the presence of OVA peptides (see Materials and methods). The titration curves of the OVA 257-264 versus Kb stabilization did not deviate
Vaccine 1995 Volume 13 Number 3 315
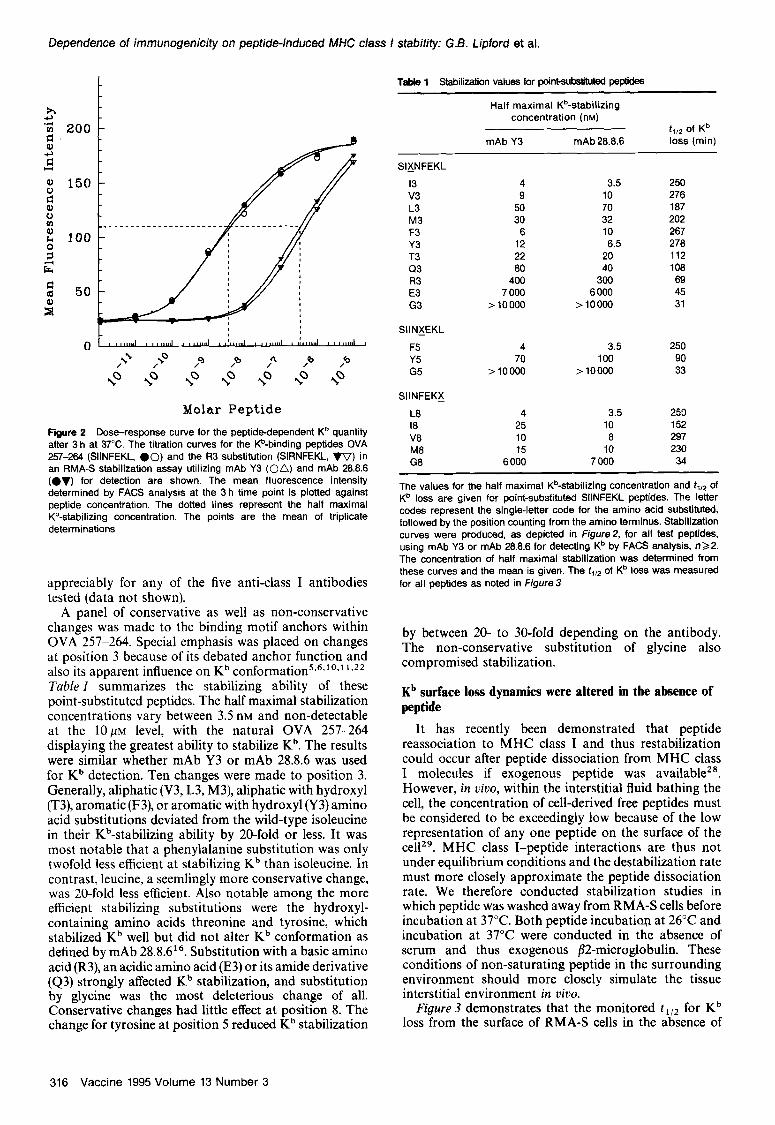
Dependence of immunogenic&y on peptide-induced MHC class f stability: G.B. Lipford et al.
Molar Peptide
Figure 2 Dose-response curve for the peptidedependent K” quantity after 3 h at 37°C. The titration curves for the Kb-binding peptides OVA 257-264 (SIINFEKL, 00) and the R3 substitution (SIRNFEKL, VV) in an RMA-S stabilization assay utilizing mAb Y3 (on) and mAb 28.8.8 (ev) for detection are shown. The mean fluorescence intensity determined by FACS analysis at the 3 h time point is plotted against peptide concentration. The dotted lines represent the half maximal Kb-&bilking concentration. The points are the mean of triplicate determinations
appreciably for any of the five anti-class I antibodies tested (data not shown).
A panel of conservative as well as non-conservative changes was made to the binding motif anchors within OVA 257-264. Special emphasis was placed on changes at position 3 because of its debated anchor function and also its apparent influence on Kb conformation5.6*‘0.1 ‘J* Tuble 1 summarizes the stabilizing ability of these point-substituted peptides. The half maximal stabilization concentrations vary between 3.5 nM and non-detectable at the 10~~ level, with the natural OVA 257-264 displaying the greatest ability to stabilize Kb. The results were similar whether mAb Y3 or mAb 28.8.6 was used for Kb detection. Ten changes were made to position 3. Generally, aliphatic (V3, L3, M3), aliphatic with hydroxyl (T3), aromatic (F3), or aromatic with hydroxyl (Y3) amino acid substitutions deviated from the wild-type isoleucine in their Kb-stabilizing ability by 20-fold or less. It was most notable that a phenylalanine substitution was only twofold less efficient at stabilizing Kb than isoleucine. In contrast, leucine, a seemlingly more conservative change, was 20-fold less efficient. Also notable among the more efficient stabilizing substitutions were the hydroxyl- containing amino acids threonine and tyrosine, which stabilized Kb well but did not alter Kb conformation as defined by mAb 28.8.616. Substitution with a basic amino acid (R3), an acidic amino acid (E3) or its amide derivative (43) strongly affected Kb stabilization, and substitution by glycine was the most deleterious change of all. Conservative changes had little effect at position 8. The change for tyrosine at position 5 reduced Kb stabilization
TaMe 1 Stabilization values for point-substituted peptides
Half maximal K%tabilizing concentration (nM)
f,,2 of Kb mAb Y3 mAb 28.8.8 loss (min)
SIXNFEKL
I3 4 3.5 250 v3 9 10 278 L3 50 70 187 M3 30 32 202 F3 8 10 287 Y3 12 8.5 278 T3 22 20 112 Q3 80 40 108 R3 400 300 89 E3 7 000 6 000 45 G3 >10000 > 10000 31
SIINXEKL
F5 4 3.5 250 Y5 70 100 90 G5 >10000 > 10000 33
SIINFEKX
L8 4 3.5 253 18 25 10 152 V8 10 8 297 M8 15 10 230 G8 6 000 7 000 34
The values for the half maximal K%tabilizing concentration and f1,2 of Kb loss are given for point-substituted SIINFEKL peptides. The letter codes represent the single-letter code for the amino acid substituted. followed by the position counting from the amino terminus. Stabilization curves were produced, as depicted in Figure& for all test peptides, using mAb Y3 or mAb 28.8.6 for detecting Kb by FACS analysis, n22. The concentration of half maximal stabilization was determined from these curves and the mean is given. The f,,* of Kb loss was measured for all peptides as noted in Figure 3
by between 20- to 30-fold depending on the antibody. The non-conservative substitution of glycine also compromised stabilization.
Kb surface loss dynamics were altered in the absence of peptide
It has recently been demonstrated that peptide reassociation to MHC class I and thus restabilization could occur after peptide dissociation from MHC class I molecules if exogenous peptide was available”. However, in uioo, within the interstitial fluid bathing the cell, the concentration of cell-derived free peptides must be considered to be exceedingly low because of the low representation of any one peptide on the surface of the ce1129. MHC class I-peptide interactions are thus not under equilibrium conditions and the destabilization rate must more closely approximate the peptide dissociation rate. We therefore conducted stabilization studies in which peptide was washed away from RMA-S cells before incubation at 37°C. Both peptide incubation at 26°C and incubation at 37°C were conducted in the absence of serum and thus exogenous flZmicroglobulin. These conditions of non-saturating peptide in the surrounding environment should more closely simulate the tissue interstitial environment in duo.
Figure 3 demonstrates that the monitored tliz for Kb loss from the surface of RMA-S cells in the absence of
316 Vaccine 1995 Volume 13 Number 3

Dependence of immunogenicity on peptideincfuced MHC class I stability: G.6. Lipford et al.
0”““““““““““’ 0 30 80 90 120 150 180
Time (min)
Figure 3 Titration of the peptidedependent rate of loss of Kb in the absence of replacement peptide. The cells were pulsed for 30 min with the indicated concentrations of the SIINFEKL peptide, washed, and the temperature was raised to 37°C. The & of peptidedependent Kb loss is also given. Points represent the mean of triplicate determinations
peptide differs greatly from when peptide remains in solution (see Figure I). The tr,* for OVA 257-264, which apparently saturates at 100 nM, was reduced to 250 min as opposed to > 726 min when peptide was continually present. The observation of saturation near 100n~ was true for all peptides tested, with the exception of G substitutions. Table I lists the fliZ of Kb loss at saturation for all substituted peptides. These values clearly rank the peptides in a similar order to the 50% maximal stabilization values and demonstrate the effect of single amino acid changes. Within this system, the phenomenon of similar saturation quantities and divergent tri2 values of stabilization can be explained by a rapid peptide association rate constant for empty Kb with more peptide-dependent variation in dissociation rate, as has been observed by other$‘.
Point substitutions in SIINFEKL display differing immunogenicities in uiuo
We have previously described an inoculum which entraps peptides in liposomes containing the saponin Quil-Az4. This inoculum was capable of inducing strong primary CTL responses to several immunodominant peptides. We utilized this approach to determine whether differences exist in the immunogenicity of point- substituted OVA 257-264 peptides. C57B1/6 mice were inoculated in the hind footpads with the various peptides and the draining lymph node cells were harvested at day 4 and cultured for an additional 4 days without restimulation. Three CTL lytic activity permutations were tested:
Can the substituted peptide inoculum induce CTL responsive to EL-4 cells pulsed with the immunizing peptide? Can these CTL respond to EL-4 cells pulsed with the wild-type OVA 257-264? and Can OVA 257-264-induced CTL respond to EL-4 cells pulsed with the substituted peptide?
The utilization of wild-type OVA 257-264-induced CTL or OVA 257-264-pulsed targets served as controls for
substituted peptide pulsing efficiency and as a measure of cross-reactivity between peptides.
Table2 shows that OVA 257-264-induced CTL displayed a specific lysis of 48.0+ 2.8% at a 20: 1 effector: target ratio for OVA 257-264-pulsed EL-4 cells. Specific lysis below 10% was considered negative. All position 3 substitutions were immunogenic except E3 and G3. All position 5 and 8 substitutions were immunogenic except G5 and G8. When analysing the three lytic activity permutations, we noticed three groupings. Pattern A included peptides in which CTL induced by that peptide reacted with EL-4 cells pulsed with the substituted peptide or the wild-type OVA peptide equally well and OVA peptide-induced CTL recognized the substituted peptide-pulsed EL-4 cells similarly. Pattern A CTL were apparently blind to the substitution. Pattern B included peptides in which CTL induced by that peptide reacted with substituted peptide-pulsed EL-4 cells with a significantly higher activity than the wild-type OVA peptide. Wild-type OVA peptide-induced CTL did not recognize these peptides well. This implies that pattern B CTL show a specificity for the substitution and this specificity was conversely apparent in the wild-type OVA peptide-induced CTL. Pattern C included peptides in which the substituted peptide-induced CTL did not respond to the immunizing peptide or the wild-type OVA peptide well; however, wild-type OVA peptide-induced CTL responded to these peptides strongly. Pattern C
Table 2 lmmunogenicity of substituted peptides
Substituted peptide- Substituted Wild-type induced peptide- Induced CTL+ induced CTL + substituted CTL + substituted peptide wild-type peptide target target target Pattern
SIXNFEKL
Wild-type v3 L3 M3 F3 Y3 T3 Q3 R3 E3 G3
SIINXEKL
Y5 G5
SIINFEKX
18 VB M6 Ga
47.6& 10.1 22.3k2.8 31.oi5.3 27.5k4.3 44.ak5.3 32.3k5.8 16.7k4.0 12.Ok5.2 5.3fO.l 7.2k2.5
21.5k3.0 4.2k3.3
34.9k2.0 38.5_+ 13.7 36.8+ 1.7
1.9kl.3
46.8112.2 19.7k2.4 22.5i3.2 29.5f3.6 26.3k5.0 16.2k4.4 17.6k1.8 4.4k1.5 a.6k3.a 7.5k3.9
24.O_fl.O 4.7+ 1.7
45.1k5.1 42.6+ 14.8 29.8+ 1.2
5.lk3.0
48.Ok2.8 35.6k4.8 38.4k9.5 37.9i 5.6 24.0 f 5.6 15.9k2.0 9.5k4.5
38.8k9.7 3.7io.4 7.8kl.l 3.8kO.l
27.9k9.4
9.9k2.5
24.4* 1.4 37.2kii.a 47.3f6.8
1.5kO.5
A C A A B 0 C 0 A A
A A
A A A A
The primary CTL lytic activity induced by point-substituted SIINFEKL peptides was measured. The letter codes represent the single-letter code for the amino acid substituted followed by the position counting from the amino terminus. The data are given as the mean percentage specific lysisfstandard error of the mean, n 23, at an effector:target ratio of 20:1, as determined by the %r-release assay. Patterns A, 6 and C are explained in the text
Vaccine 1995 Volume 13 Number 3 317
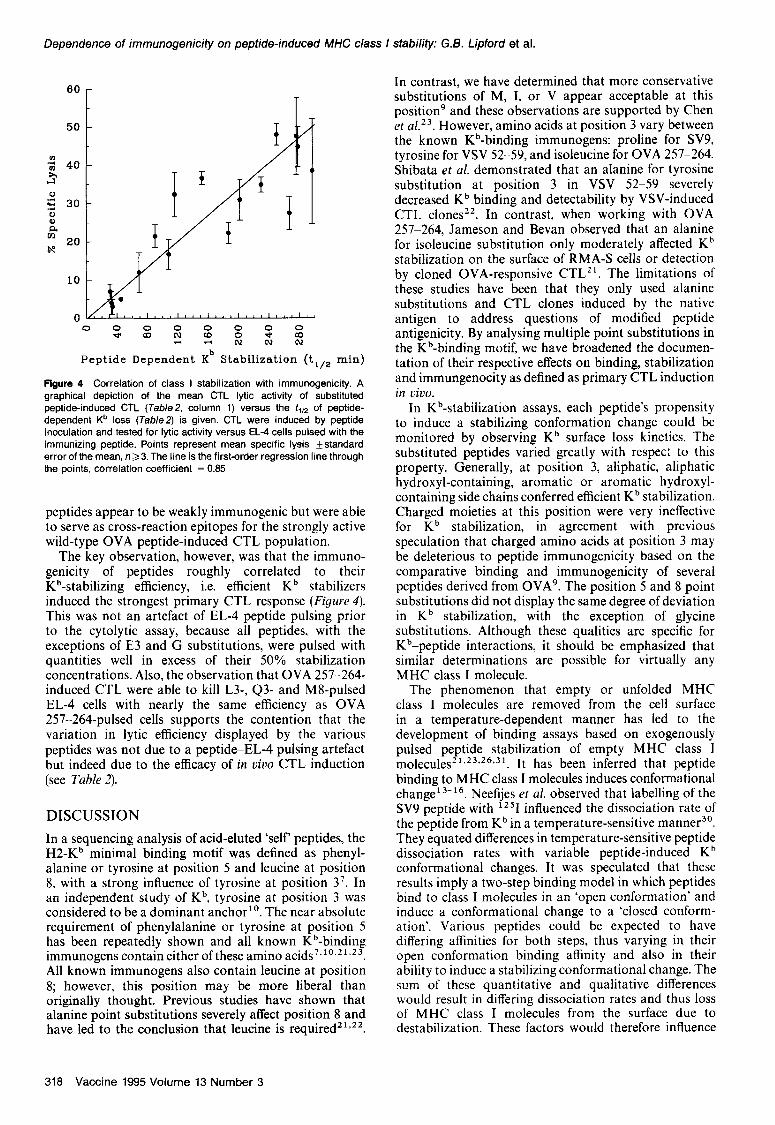
Dependence of immunogenicity on peptide-induced MHC class I stability: G.B. Lipford et al.
60 r
10
Peptide Dependent Kb Stabilization (t,,, rnin)
flgure 4 Correlation of class I stabilization with immunogenicity. A graphical depiction of the mean CTL lytic activity of substituted peptide-induced CTL (Table2, column 1) versus the tin of peptide- dependent Kb loss (Table2) is given. CTL were induced by peptide inoculation and tested for lytic activity versus EL-4 cells pulsed with the immunizing peptide. Points represent mean specific lysis *standard error of the mean, n33. The line is the first-order regression line through the points, correlation coefficient = 0.85
peptides appear to be weakly immunogenic but were able to serve as cross-reaction epitopes for the strongly active wild-type OVA peptide-induced CTL population.
The key observation, however, was that the immuno- genicity of peptides roughly correlated to their Kb-stabilizing efficiency, i.e. efficient Kb stabilizers induced the strongest primary CTL response (Figure 4). This was not an artefact of EL-4 peptide pulsing prior to the cytolytic assay, because all peptides, with the exceptions of E3 and G substitutions, were pulsed with quantities well in excess of their 50% stabilization concentrations. Also, the observation that OVA 257-264- induced CTL were able to kill L3-, Q3- and MS-pulsed EL-4 cells with nearly the same efficiency as OVA 257-264-pulsed cells supports the contention that the variation in lytic efficiency displayed by the various peptides was not due to a peptide-EL-4 pulsing artefact but indeed due to the efficacy of in viva CTL induction (see Table 2).
DISCUSSION
In a sequencing analysis of acid-eluted ‘self’ peptides, the H2-Kb minimal binding motif was defined as phenyl- alanine or tyrosine at position 5 and leucine at position 8, with a strong influence of tyrosine at position 3’. In an independent study of Kb, tyrosine at position 3 was considered to be a dominant anchor”. The near absolute requirement of phenylalanine or tyrosine at position 5 has been repeatedly shown and all known Kb-binding immunogens contain either of these amino acids’.“.’ 1.23. All known immunogens also contain leucine at position 8; however, this position may be more liberal than originally thought. Previous studies have shown that alanine point substitutions severely affect position 8 and have led to the conclusion that leucine is required2’,22.
In contrast, we have determined that more conservative substitutions of M, I, or V appear acceptable at this position’ and these observations are supported by Chen et al 23 However, amino acids at position 3 vary between the known Kb-binding immunogens: proline for SV9, tyrosine for VSV 52-59, and isoleucine for OVA 257-264. Shibata et al. demonstrated that an alanine for tyrosine substitution at position 3 in VSV 52-59 severely decreased Kb binding and detectability by VSV-induced CTL clones22. In contrast, when working with OVA 257-264, Jameson and Bevan observed that an alanine for isoleucine substitution only moderately affected Kb stabilization on the surface of RMA-S cells or detection by cloned OVA-responsive CTL”. The limitations of these studies have been that they only used alanine substitutions and CTL clones induced by the native antigen to address questions of modified peptide antigenicity. By analysing multiple point substitutions in the Kb-binding motif, we have broadened the documen- tation of their respective effects on binding, stabilization and immungenocity as defined as primary CTL induction in vivo.
In Kb-stabilization assays, each peptide’s propensity to induce a stabilizing conformation change could be monitored by observing Kb surface loss kinetics. The substituted peptides varied greatly with respec: to this property. Generally, at position 3, aliphatic, aliphatic hydroxyl-containing, aromatic or aromatic hydroxyl- containing side chains conferred efficient Kb stabilization. Charged moieties at this position were very ineffective for Kb stabilization, in agreement with previous speculation that charged amino acids at position 3 may be deleterious to peptide immunogenicity based on the comparative binding and immunogenicity of several peptides derived from 0VA9. The position 5 and 8 point substitutions did not display the same degree of deviation in Kb stabilization, with the exception of glycine substitutions. Although these qualities are specific for Kb-peptide interactions, it should be emphasized that similar determinations are possible for virtually any MHC class I molecule.
The phenomenon that empty or unfolded MHC class I molecules are removed from the cell surface in a temperature-dependent manner has led to the development of binding assays based on exogenously pulsed peptide stabilization of empty MHC class 1 molecules21.23*26*31. It has been inferred that peptide binding to MHC class I molecules induces conformational change13-16. Neefijes et al. observed that labelling of the SV9 peptide with lz51 influenced the dissociation rate of the peptide from Kb in a temperature-sensitive manner3’. They equated differences in temperature-sensitive peptide dissociation rates with variable peptide-induced Kb conformational changes. It was speculated that these results imply a two-step binding model in which peptides bind to class I molecules in an ‘open conformation’ and induce a conformational change to a ‘closed conform- ation’. Various peptides could be expected to have differing affinities for both steps, thus varying in their open conformation binding affinity and also in their ability to induce a stabilizing conformational change. The sum of these quantitative and qualitative differences would result in differing dissociation rates and thus loss of MHC class I molecules from the surface due to destabilization. These factors would therefore influence
318 Vaccine 1995 Volume 13 Number 3

Dependence of immunogenic@ on peptide-induced MHC class I stability: G.8. Lipford et al.
the surface peptide-MHC class I steady-state composition displayed by the cell.
The driving force behind binding motif analysis is to enhance our understanding of peptide-MHC class I interaction in the hope of determining what is immunogenic. The immune system deals with an immunogen as a polyclonal population response. In contrast to CTL clonal analysis of peptide immuno- genicity, direct peptide immunization and analysis of primary induced CTL populations has the benefit of reflecting the graded peptide-induced response of an intact immune system. Table 2 demonstrates that the intensity of the peptide-specific CTL response was variable depending on the substituted amino acid. The efficiency of response correlated with stabilization efficiency (see Table I, Figure 4). If the two-step binding model of peptide-Kb interaction (as discussed above) is correct, then class I stabilization is an important quality of the peptide. Within the cell, a multitude of peptides may be generated, many of which may be transported into the endoplasmic reticulum and effectively bind MHC class I molecules; but, if peptides vary in their ability to stabilize MHC class I molecules at the cell surface, then the representation of a peptide within the milieu of presented immunogens will depend on its ability to induce a stabilizing conformational change in the class I molecule. For example, for two peptides which enter the endoplasmic reticulum in similar amounts and have the same association rate for the presenting MHC class I molecule, the cell surface steady-state differences between the two peptides would be dependent on stabilizing ability. If production rates are similar but loss rates deviate, then representation on the surface diverges. Thus peptide-class I stabilization, under these circumstances, could greatly influence the repertoire of effective immunogens and thus affect the pool of antigen available for immunosurveillance. This quality could be a partial explanation for the phenomenon of immunodominance, because quantitative increases in peptide representation on the cell surface could potentially increase the avidity of interaction with the T-cell receptor and thus lower the affinity requirements 32 This would effectively expand the activated polyclonal CTL population and thus increase the strength of response.
The various CTL reactivity patterns mentioned in Table2 are intriguing. Pattern B peptides appear to be examples of CTL specificity in which a single amino acid change greatly influences the CTL population response. However, patterns A and C display a blind cross- reactivity between the wild-type OVA peptide and the substituted peptide. This cross-reactive quality is independent of peptide-class I stabilization rankings. In pattern C, the immunogenicity of the peptides was weak due to poor stabilizing efficiency; however, they served as good target peptides for wild-type OVA peptide-induced CTL. This observation implies that pathogen-derived low or sub-immunogenic peptides could be engineered to increase immunogenicity by enhanced class I-stabilizing ability, while at the same time preserving cross-reactivity to the wild-type epitope. Recently, this was achieved in our laboratory with a peptide from human papilloma virus type 16 protein E6 33. By point substituting amino acids at an MHC class I contact point, peptide-dependent class I stability was increased. This resulted in a corresponding enhancement in immunogenicity and
these CTL were capable of killing E6 transfectants where CTL induced with the wild-type peptide failed to kill the transfectant Peptide engineering to increase presentation half-life during inoculation was capable of enhancing the CTL response to a sub-immunogenic epitope naturally processed and presented by a tumour cell.
In conclusion, by using the OVA peptide 257-264 as a template and making amino acid substitutions, we have shown that the degree of peptide-induced MHC class I stablization is an important aspect of peptide immuno- genicity. Utilizing this parameter can potentially aid in the selection of immunogenic epitopes from a pool of possible class I-binding peptides. An understanding of the MHC class I-binding preferences could also assist in engineering better immunogens. Furthermore, the quality of peptide differential stabilization and hence variation in cell surface steady-state levels of antigenic peptides may be key phenomena in the determination of immunodominance.
ACKNOWLEDGEMENTS
The authors are grateful to Dr Carys Evans for critical reading of the manuscript and to Ms Monika Mayer for skilful technical assistance. This work was supported by grants from the Bundesminister fiir Forschung und Technologie and the Wilhelm Sander-Stiftung.
REFERENCES
1
2
3
4
5
6
7
8
9
10
11
12
13
Townsend, A. and Bodmer. H. Antigen recognition by class l-restricted T lymphocytes. Annu. Rev. Immunol. 1989, 7, 601-624 Bjorkman, P.J. and Parham, P. Structure, function, and diversity of class I major histocompatibility complex molecules. Annu. Rev. Biochem. 1990, 59,253-288 Garrett, T.P.. Saper, M.A.. Bjorkman, P.J., Strominger, J.L. and Wiley, D.C. Specificity pockets for the side chains of peptide antigens in HLA-Aw68. Nature 1989, 342, 692-696 Jardetzky. T.S., Lane, W.S.. Robinson, R.A., Madden, DR. and Wiley, D.C. Identification of self peptides bound to purified HLA-B27. Nature 1991, 353, 32&329 Fremont, D.H., Matsumura, M., Stura. E.A., Peterson, P.A. and Wilson, LA. Crystal structures of two viral peptides in complex with murine MHC class I H-2Kb. Science 1992, 257, 919-927 Matsumura, M., Fremont, D.H., Peterson, P.A. and Wilson, LA. Emerging principles for the recognition of peptide antigens by MHC class I molecules. Science 1992, 257, 927-934 Falk, K., Rotzschke. 0.. Stevanovic, S., Jung, G. and Rammensee, H.G. Allele-specific motifs revealed by sequencing of self-peptides eluted from MHC molecules. Nature 1991, 351. 2S0-296 Pamer, E.G., Harty. J.T. and Bevan, M.J. Precise predictlon of a dominant class I MHC-restricted epitope of Listeria monocytogenes. Nature 1991. 353, 852-865 Lipford, G.B., Hoffman, M., Wagner, H. and Heeg, K. Primary in viva responses to ovalbumin. Probing the predictive value of the Kb binding motif. J. Immunol. 1993, 150, 1212-1222 Van Bleek, G.M. and Nathenson, S.G. The structure of the antigen-binding groove of major histocompatibility complex class I molecules determines specific selection of self-peptides. Proc. iVat/ Acad. Sci. US A 1991, 88, 11032-l 1036 Stura, E.A., Matsumura. M., Fremont, D.H., Saito, Y., Peterson, P.A. and Wilson, I.A. Crystallization of murine major histocompatibility complex class I H-2Kb with single peptides. J. Mol. Biol. 1992, 228, 975-982 Lie, M.R.. Myers, N.B., Connolly, J.M., Gorka, J.. Lee, D.R. and Hansen, T.H. The specific binding of peptide ligand to Ld class I major histocompatibility complex molecules determines their antigenic structure. J. Exp. Med. 1991, 173. 449-459 Lie, W.R.. Myers, N.B.. Gorka, J.. Rubocki, R.J., Connolly, J.M. and Hansen, T.H. Peptide ligand-induced conformation and surface expression of the Ld class I MHC molecule. Nature 1990, 344, 439-441
Vaccine 1995 Volume 13 Number 3 319

Dependence of immunogenicity on peptide-induced MHC class I stability: G.6. Lipford et al.
14
15
16
17
16
19
20
21
22
23
Elliott, T., Cerundolo, V., Elvin, J. and Townsend, A. Peptide-induced role for indirect conformational alterations in the MHC class I or conformational change of the class I heavy chain. Nature 1991, 361, bound peptide in determining T cell recognition. J. Exp. Med. 1993, 462* ITT,669673 Bluestone, J.A., Jameson, S., Miller, S. and Dick, R. Peptide-induced conformational changes in class I heavy chains alter major histocompatibilty complex recognition. J. Exp. Med. 1992, 176, 1757-1761
24 Lipford, G.B., Wagner, H. and Heeg, K. Vaccination with immunodominant peptides encapsulated in Quil A-containing liposomes induces peptide-specific primary CD6+ cytotoxic T cells. Vaccine 1994,12,73-66
Catipovic, B., Dal Porto, J., Mage, M., Johansen, T.E. and Schneck, J.P. Major histocompatiblity complex conformational epitopss are peptide specific. J. Exp Med. 1992, 176, 1611-1616 Lipford, G.B., Lehn, N., Bauer, S., Heeg, K. and Wagner, H. The immunodominant peptide from listerolysin in Quil A liposomes vaccinates CD6+ CTL and confers protection to infection. Immunol. Leff. 1994,40, 101-104 Kast, W.M., Roux, L., Curren. J., Blom, H.J., Voordouw, A.C.. Meloen, R.H. et al. Protection against lethal Sendai virus infection by in viva priming of virus-specific cytotoxic T lymphocytes with a free synthetic peptide. Proc. Nafl Acad. Sci. US A 1991,88,2263-2267 Schulz, M., Zinkernagel, R.M. and Hengartner, H. Peptide-induced antiviral protection by cytotoxic T cells. Proc. Nafl Acad. Sci. USA 1991,66,991-993 Feltkamp, M.C., Smits, H.L., Vierboom, M.P., Minnaar, R.P., de Jongh, B.M., Drijfhout, J.W. et al. Vaccination with cytotoxic T lymphocyte epitopecontaning peptide protects against a tumor induced by human papillomavirus type %-transformed cells. Eur. J. Immunol. 1993, 23, 2242-2249 Jameson. SC. and Bevan, M.J. Dissection of major histo- compatibility complex (MHC) and T cell receptor contact residues in a Kb-restricted ovalbumin peptide and an assessment of the predictive power of MHC-binding motifs. fur. J. Immunol. 1992, 22, 2663-2667 Shibata, K., Imarai, M., van Bleek, G.M., Joyce, S. and Nathenson, S.G. Vesicular stomatitis virus antigenic octapeptide N52-59 is anchored into the groove of the H-2Kb molecule by the side chains of three amino acids and the main-chain atoms of the amino terminus. Proc. Nafl Acad. Sci. USA 1992,69, 31363139 Chen. W.. McCluskey, J., Rodda, S. and Carbone, F.R. Changes at peptide residues buried in the major histocompatibility complex (MHC) class I binding cleft influence T cell recognition: a possible
25
26
27
26
29
30
31
32
33
Heeg, K., Kuon, W. and Wagner, H. Vaccination of class I major histocompatibilitycomplex (MHC)-restricted murine CD6+ cytotoxic T lymphocytes towards soluble antigens: immunostimulating- ovalbumin complexes enter the class I MHCrestricted antigen pathway and allow sensitization against the immunodominant peptide. Eur. J. Immunol. 1991, 2f, 1523-1527 Ljunggren, H.G., Stam, N.J., Ohlen, C.. Neefjes, J.J., Hoglund, P., Heemels, M.T. et a/. Empty MHC class I molecules come out in the cold. Nature 19% 346, 476-466 Ortiz Navarrete, V. and Hammerling, G.J. Surface appearance and instability of empty H-2 class I molecules under physiological conditions. Proc. Nafl Acad. Sci. USA 1991, 66, 3594-3597 Otten, G.R., Bikoff, E., Ribaudo, R.K., Kozlowski, S., Margulies, D.H. and Germain, R.N. Peptide and beta Pmicroglobulin regulation of cell surface MHC class I conformation and expression J. Immunol. 1992,146,3723-3732 Hunt, D.F., Henderson, R.A., Shabanowitz, J., Sakaguchi, K., Michel, H., Sevilir, N. et al. Characterization of peptides bound to the class I MHC molecule HLA-A2.1 by mass spectrometry. Science 1992, 255, 1261-1263 Neefjes, J.J.. Dierx, J. and Ploegh, H.L. The effect of anchor residue modifications on the stability of major histocompatibility complex class I-peptide interactions. Eur. J. Immunol. 1993, 23, 840845 Townsend, A., Ohlen, C., Bastin, J., Ljunggren, H.G., Foster, L. and Karre, K. Association of class I major histocompatibility heavy and light chains induced by viral peptides. Nature 1969, 340, 443-448 Kane, K.P. and Mescher, M.F. Activation of CDSdependent cytotoxic T lymphocyte adhesion and degranulation by peptide class I antigen complexes. J. Immunol. 1993,156, 4766-4797 Lipford, G.B., Bauer, S., Wagner, H. and Heeg, K. Peptide engineering allows cytotoxic T cell vaccination against human papillomavirus tumor antigen, E6. lmmuno/ogy 1995 (in press)
320 Vaccine 1995 Volume 13 Number 3




















