
Immunity to HIV M Cell DNA Vaccination for CTL
10
of November 25, 2013. This information is current as to HIV M Cell DNA Vaccination for CTL Immunity T. Shata and David W. Pascual Xinhai Wang, David M. Hone, Asmahan Haddad, Mohamed http://www.jimmunol.org/content/171/9/4717 2003; 171:4717-4725; ; J Immunol References http://www.jimmunol.org/content/171/9/4717.full#ref-list-1 , 30 of which you can access for free at: cites 53 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2003 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on November 25, 2013 http://www.jimmunol.org/ Downloaded from by guest on November 25, 2013 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Immunity to HIV M Cell DNA Vaccination for CTL

of November 25, 2013.This information is current as
to HIVM Cell DNA Vaccination for CTL Immunity
T. Shata and David W. PascualXinhai Wang, David M. Hone, Asmahan Haddad, Mohamed
http://www.jimmunol.org/content/171/9/47172003; 171:4717-4725; ;J Immunol
Referenceshttp://www.jimmunol.org/content/171/9/4717.full#ref-list-1
, 30 of which you can access for free at: cites 53 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2003 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 25, 2013
http://ww
w.jim
munol.org/
Dow
nloaded from

M Cell DNA Vaccination for CTL Immunity to HIV 1
Xinhai Wang,* David M. Hone,† Asmahan Haddad,* Mohamed T. Shata,† andDavid W. Pascual2*
To facilitate invasion, reovirus has evolved to attach to M cells, a specialized epithelium residing within the follicle-associatedepithelium that covers mucosal inductive tissues. Thus, we questioned adapting reovirus protein�1 to ferry DNA vaccines to themucosa to immunize against HIV. Three expression plasmids encoding HIV(Ba-L) gp160, cytoplasmic gp140, and secreted gp140were tested in mice as protein�1-poly-L-lysine-DNA complexes (formulated vaccine) via the intranasal route. Evaluation ofcell-mediated immunity showed that the formulated gp160 DNA vaccine was more effective for stimulating envelope (Env)-specificCTL responses in lungs, lower respiratory lymph nodes (LN), cervical LN, submaxillary gland LN, and spleens. Three doses ofvaccine were required for CTL responses, and intranasal naked DNA immunizations were ineffective. The greatest CTL activitywas observed between weeks 8 and 10 for gp160-vaccinated mice, and activity remained detectable by week 16. These Env-specificCTL responses were perforin dependent in peripheral tissues, but mostly Fas dependent in the lungs. These Env-specific CTLs alsoproduced IFN-�. Mice vaccinated with the formulated gp160 DNA vaccine showed potent antiviral immunity against vacciniavirus-env replication in ovaries. Thus, compared with live vectors, protein�1-mediated DNA delivery represents an alternativemucosal formulation for inducing cellular immunity against HIV-1. The Journal of Immunology, 2003, 171: 4717–4725.
T he mucosal surfaces are the primary sites of transmissionfor most infectious diseases requiring the induction ofmucosal immunity for preventing pathogen entry at these
surfaces. More recently, vaccines, particularly attenuated viruses(1, 2), have been designed for their mucosal application. Althoughthese vaccines are effective for stimulating appropriate host im-mune responses, alternative vaccines and vaccine delivery strate-gies are warranted to avoid the potential side effects of live viralvectors. Such alternatives may lie with subunit vaccines or DNAvaccines, the latter being relatively easy to prepare and administer.Recent evidence shows that DNA vaccination can confer protec-tive immunity against a number of infectious agents including in-fluenza (3, 4), herpes simplex virus (5), HIV (6, 7), and Borreliaburgdorferi (8) infections. However, naked DNA immunizationshave been proved less than optimal for stimulating mucosal im-munity (7, 9, 10) when they are administered peripherally. In con-trast, microencapsulated DNA vaccines effectively elicit protectivemucosal responses in a murine rotavirus challenge model (11, 12).Although it is unclear whether such protection was conferred bythe protective properties of microencapsulation, the ability of mi-crospheres to interact with Peyer’s patches, M cells (13, 14), or acombination of both suggests that more efficient uptake of DNA bymucosal inductive tissues favors enhanced mucosal immunity.
Receptor-mediated gene transfer is effective for delivering trans-genes into tissues by specific cellular receptors. One such method
is to conjugate to a DNA-binding domain a ligand specific for acell surface receptor. This can be facilitated using poly-L-lysine(PL).3 This DNA-ligand complex is internalized by targeted cellswhen the ligand binds to its respective cell surface receptor. Suchreceptor-mediated gene transfer has been accomplished by target-ing a variety of receptors: asialo-orosomucoid (15, 16); transferrin(17–19); lectins (20); and insulin (21).
Reovirus is an enteric pathogen that infects the host followingattachment to intestinal Peyer’s patch M cells (22). Thus, as withother enteric pathogens, reovirus exploits M cells as a means togain entry into the host. Mediating reovirus attachment is its ad-hesin, protein �1, which is expressed and located at the 12 verticesof the viral icosahedron (22). Protein �1 is a 45-kDa protein thatpolymerizes via its N terminus (23) to form either a trimer (24) ora tetramer (25) depending on how it is isolated.
In our previous studies, we have demonstrated that our recom-binant protein �1 binds its receptor on cell lines (26) and nasal-associated lymphoid tissue M cells (27). Using protein �1-PL-DNA for intranasal (i.n.) transgene vaccination, mice weresuccessfully immunized against reporter gene constructs, lucif-erase and �-galactosidase (�gal) (27). Herein, this study focuseson the induction cell-mediated immunity against HIV envelope(Env) by using protein �1-mediated DNA immunization.
Our results show that protein �1-mediated transgene vaccina-tion can be used for immunization against HIV. Three DNA vac-cines were tested, and the data for the mice given the gp160 DNAas protein �1-PL-gp160 DNA (formulated vaccine), but not asnaked DNA, showed long-lasting Env-specific CTLs with potentantiviral immunity.
Materials and MethodsProduction of recombinant reovirus �1 protein
The recombinant protein �1 from reovirus serotype 3 as a maltose-bindingprotein was generated and expressed in Escherichia coli, as previously
*Veterinary Molecular Biology, Montana State University, Bozeman, MT 59717; and†Institute of Human Virology, Baltimore, MD 21201
Received for publication May 23, 2003. Accepted for publication August 19, 2003.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by U.S. Public Health Grants AI-49071 (to D.W.P.),AI-41914, and AI-55367 (to D.M.H.) and in part by the Montana Agricultural Stationand U.S. Department of Agricultural Formula Funds. This is Montana AgriculturalStation Journal Series 2003-11.2 Address correspondence and reprint requests to Dr. David W. Pascual, VeterinaryMolecular Biology, Montana State University, Bozeman, MT 59717-3610. E-mailaddress: [email protected]
3 Abbreviations used in this paper: PL, poly-L-lysine; i.n., intranasal; Env, envelope;LN, lymph nodes; LRLN, lower respiratory LN; CLN, cervical LN; SMLN, submax-illary gland LN; CM, complete medium; CMA, concanamycin A; FAE, follicle-associated epithelium; �gal, �-galactosidase; FasL, Fas ligand.
The Journal of Immunology
Copyright © 2003 by The American Association of Immunologists, Inc. 0022-1767/03/$02.00
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

described (26). This fusion protein was purified by affinity chromatographyusing amylose resin according to the manufacturer’s directions (New En-gland Biolabs, Beverly, MA). The MBP::protein �1 fusion protein is re-ferred to as recombinant protein �1.
Mice
Specific pathogen-free male and female BALB/c mice were purchasedfrom the National Cancer Institute (Frederick Cancer Research Facility,Frederick, MD) at 5–6 wk of age and maintained in the Animal ResourcesCenter at Montana State University (Bozeman, MT). Breeding pairs ofIFN-�-deficient (IFN-��/�) mice on a BALB/c background, as well aswild-type (IFN-��/�) mice on shared background (28, 29), were kindlyprovided by Genentech (San Francisco, CA), and colonies were establishedand maintained in the Animal Resources Center. All mice were kept underpathogen-free conditions in horizontal laminar flow cabinets and were fedsterile food and water ad libitum. The mice were free of bacterial and viralpathogens, as determined by Ab screening and by histopathological anal-ysis of major organs and tissues. The mice used in these experiments werebetween 5 and 8 wk of age.
Vaccination
The recombinant protein �1 was covalently linked to PL, as previouslydescribed (17, 26, 27). Groups of five mice each were immunized withnaked DNA or the formulated vaccine containing one of the three DNAvaccine constructs: pOGL6, which expresses HIV-1 Env Ba-L (30);pSF128, which expresses gp140sec; or pSF128-�LP, which expressesgp140cyt. This latter construct was generated by PCR amplification of thegp140-encoding sequence in pSF128 using a 5�-primer that initiated PCRdownstream of the 31-aa CD5 leader peptide and substituted a methioninein its place. The amplified sequence was then inserted into pcDNA3.1using T4 DNA ligase. Each of the described DNA vaccines were assessedin transfection assays and shown to produce equivalent levels of envelopeglycoprotein by SDS-PAGE and semiquantitative capture ELISAs. Indi-vidual mice were anesthetized with isofluorane (Abbott Laboratories,North Chicago, IL) by inhalation and then immunized i.n three times at7-day intervals with 20 �g of plasmid DNA per dose with or without 15 �gof protein �1-PL.
Lymphocyte isolation
Lymphocytes were isolated from spleens, lungs, mediastinal and hilarlymph nodes (LRLN), cervical LN (CLN), and submaxillary gland LN(SMLN). LRLN, CLN, and SMLN were passed through Nitex mesh (Fair-view Fabrics, Hercules, CA) to remove debris. Splenic mononuclear cellsuspensions were obtained by mechanical disruption, followed by Ficoll-Hypaque (Lymphocyte M; Accurate Chemical, Westbury, NY) densitygradient centrifugation (31). Lung mononuclear cells were treated with 0.5mg/ml Clostridium histolyticum type IV collagenase (Worthington, Free-hold, NJ) for 15-min intervals at 37°C. After incubation, the digestionmixtures were passed through Nitex mesh (Fairview Fabrics) to removeundigested material. Mononuclear cells were isolated by Percoll (Pharma-cia, Uppsala, Sweden) density gradient centrifugation; the mononuclearcells interfaced between 40 and 75% Percoll. We noted �95% viability forlymphocytes isolated from each tissue, as determined by trypan blueexclusion.
Lymphocyte culture and 51Cr release assay
Both ex vivo and in vitro restimulated mononuclear cells were assessed forHIV. Env-specific CTL activity using BC-Env, a derivative of the masto-cytoma cell lines P815, stably transfected with the full-length env gene ofHIV-1 and BC-�gal as negative control target cells, which are P815 cells,stably transfected with the E. coli lacZ gene (32). Targets were treated withmitomycin C (50 �g/ml; Sigma-Aldrich, St. Louis, MO) and extensivelywashed with complete medium [CM; RPMI 1640 plus 10% FBS (HyClone,Logan, UT) plus 10 mM HEPES buffer plus 10 mM nonessential aminoacids plus 10 mM sodium pyruvate plus 10 U/ml penicillin/streptomycin].Targets were subsequently labeled with Na2
51CrO4 (DuPont/NEN, Wil-mington, DE) for 1 h at 37°C. The cells were washed three times in CM.Effector cells were incubated with various ratios of labeled targets in U-bottom 96-well microtiter dishes (Corning-Costar, Oneonta, NY) for 4 h at37°C. After incubation, cells were spun, and supernatants were assessed forreleased 51Cr. Specific cytolysis was determined as the amount of 51Crreleased in the presence of effector cells corrected for spontaneous 51Crrelease divided by maximal (detergent-induced) release corrected for spon-taneous 51Cr release. For Ag restimulation, splenic and LN mononuclearcells were restimulated with mitomycin C-treated BC-Env cells at 5:1 inCM for 5 days at 37°C.
To assess cytolytic mechanisms, washed ex vivo or Ag-restimulatedlymphocytes were treated with anti-FasL mAb or 10 nM concanamycin A(CMA; Wako Bioproducts, Richmond, VA) for 2 h at 37°C, similar to thatpreviously described (33). Likewise, some ex vivo or Ag-restimulated lym-phocytes were treated with 10 �g/ml anti-FasL mAb (clone MFL3; BDPharMingen, San Diego, CA). Cells were assessed for CTL activity, asdescribed above.
Cytokine detection essays
Supernatants collected from Env (Bc-Env)-restimulated cultures were an-alyzed for expression of IFN-�, IL-2, IL-4, IL-5, IL-6, and IL-10. Com-mercially produced cytokine-specific mAbs (BD PharMingen) were used,as previously described (31, 34).
Vaccinia virus-Env challenge
The extent of antiviral immunity was determined using a vaccinia-env chal-lenge model, as previously described (35, 36). The vaccinia-env vector,vP1174, was obtained from AIDS Research and Reference Reagent Pro-gram, Division of AIDS, National Institute of Allergy and Infectious Dis-eases, National Institutes of Health (Bethesda, MD). Briefly, vP1174 wasexpanded in BSC-1 cells until 90% of the cells lysed. Culture supernatantswere collected by centrifugation at 4000 � g for 10 min, and aliquots of thesupernatants were stored in liquid nitrogen until used. These supernatantstypically yielded �5 � 109 vP1174 PFU/ml, as determined by a directplaque essay on BSC-1 cells. To assess antiviral protection, groups ofBALB/c mice were vaccinated i.n. or i.m. with naked gp160 DNA vaccineor i.n. with the formulated gp160 DNA vaccine as described above. Allmice were vaccinated at the same time intervals. Mice were given 108 PFUof vP1174 i.p. 16 wk after primary vaccination. The ovaries from the chal-lenged mice were harvested, homogenized, and clarified by centrifugationat 4000 � g for 10 min. Serial 10-fold dilutions were made for infection ofBSC-1 cell monolayers, and plaques were enumerated 2 days after in vitroculture.
Statistical analysis
Statistical differences between experimental parameters were determinedsignificant if p was �0.05 using Student’s t test and one-way ANOVA. Forthe challenge study, Wilcoxon’s two sample test was performed.
ResultsMode of HIV-1 Env expression impacts induction of CTLresponses
Previous results have shown that M cell-targeted DNA deliveryrepresented an effective means to mucosally vaccinate for the stim-ulation of CTL responses (27). To adapt this vaccine vehicle topotentially immunize against HIV, M cell-targeted DNA deliveryusing recombinant reovirus protein �1 was used for i.n. immuni-zation of BALB/c mice. Three groups of BALB/c mice (five pergroup) each received an Env (Ba-L) expression plasmid in con-junction with protein �1 conjugated to PL and were given 20 �gof plasmid per dose on days 0, 7, and 14. Each group received aDNA vaccine that expressed full-length gp160, which is located inthe plasma membrane, a truncated derivative of Env that lacks thetransmembrane and cytoplasmic domains and is secreted(gp140sec), or a leader peptide-deficient derivative of gp140 that isexpressed cytoplasmically (gp140cyt). After 6 wk post-primary im-munization, various respiratory and splenic tissues were isolated,and mononuclear cell fractions were subjected to cytolytic assaysby 51Cr release to determine specific reactivity for Env. Bothfreshly isolated lymphocytes and in vitro-restimulated lympho-cytes showed specific lysis of Env-expressing target cells, but not�gal-expressing targets (Fig. 1). However, variable killing was ob-served and was dependent on the mode of Env expression. Rankorder experiments for lung lymphocytes revealed that the DNAvaccine for gp160 � gp140cyt � gp140sec (Fig. 1, A--C and TableI). Freshly isolated lymphocytes from the other lymphoid com-partments were not as effective in the lysis of Env-expressing tar-gets, varying from 13 to 26% specific lysis (Fig. 1, A–C). How-
4718 TARGETED DNA DELIVERY FOR HIV
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
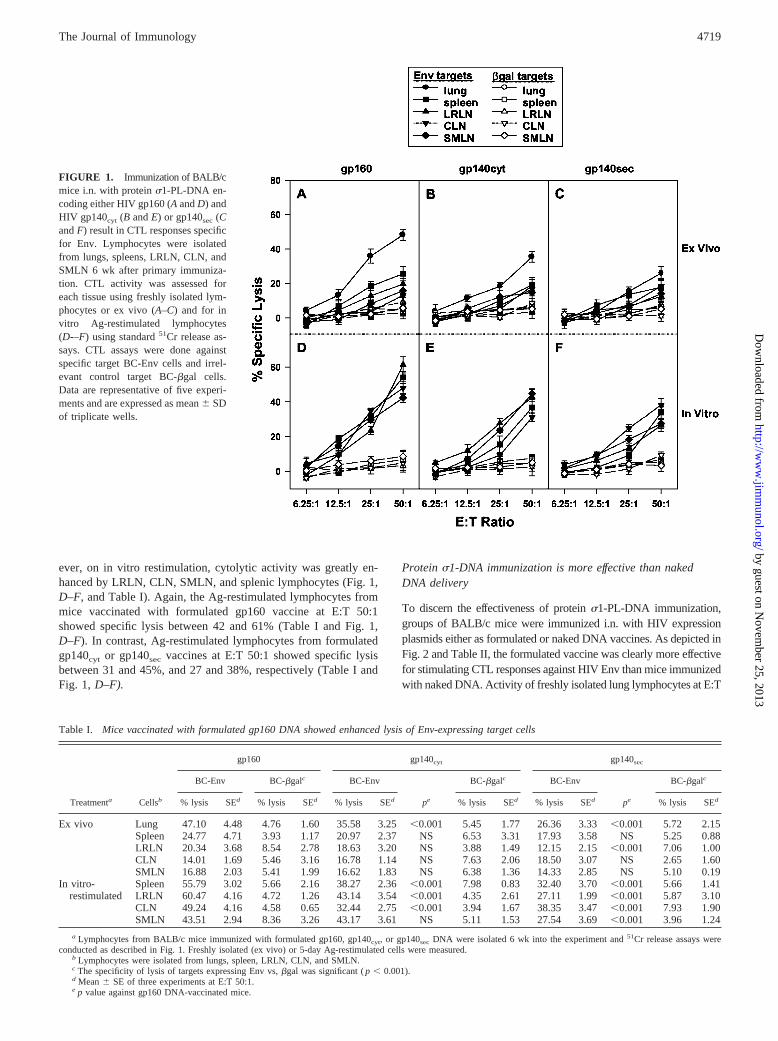
ever, on in vitro restimulation, cytolytic activity was greatly en-hanced by LRLN, CLN, SMLN, and splenic lymphocytes (Fig. 1,D–F, and Table I). Again, the Ag-restimulated lymphocytes frommice vaccinated with formulated gp160 vaccine at E:T 50:1showed specific lysis between 42 and 61% (Table I and Fig. 1,D–F). In contrast, Ag-restimulated lymphocytes from formulatedgp140cyt or gp140sec vaccines at E:T 50:1 showed specific lysisbetween 31 and 45%, and 27 and 38%, respectively (Table I andFig. 1, D–F).
Protein �1-DNA immunization is more effective than nakedDNA delivery
To discern the effectiveness of protein �1-PL-DNA immunization,groups of BALB/c mice were immunized i.n. with HIV expressionplasmids either as formulated or naked DNA vaccines. As depicted inFig. 2 and Table II, the formulated vaccine was clearly more effectivefor stimulating CTL responses against HIV Env than mice immunizedwith naked DNA. Activity of freshly isolated lung lymphocytes at E:T
Table I. Mice vaccinated with formulated gp160 DNA showed enhanced lysis of Env-expressing target cells
Treatmenta Cellsb
gp160 gp140cyt gp140sec
BC-Env BC-�galc BC-Env
pe
BC-�galc BC-Env
pe
BC-�galc
% lysis SEd % lysis SEd % lysis SEd % lysis SEd % lysis SEd % lysis SEd
Ex vivo Lung 47.10 4.48 4.76 1.60 35.58 3.25 �0.001 5.45 1.77 26.36 3.33 �0.001 5.72 2.15Spleen 24.77 4.71 3.93 1.17 20.97 2.37 NS 6.53 3.31 17.93 3.58 NS 5.25 0.88LRLN 20.34 3.68 8.54 2.78 18.63 3.20 NS 3.88 1.49 12.15 2.15 �0.001 7.06 1.00CLN 14.01 1.69 5.46 3.16 16.78 1.14 NS 7.63 2.06 18.50 3.07 NS 2.65 1.60SMLN 16.88 2.03 5.41 1.99 16.62 1.83 NS 6.38 1.36 14.33 2.85 NS 5.10 0.19
In vitro- Spleen 55.79 3.02 5.66 2.16 38.27 2.36 �0.001 7.98 0.83 32.40 3.70 �0.001 5.66 1.41restimulated LRLN 60.47 4.16 4.72 1.26 43.14 3.54 �0.001 4.35 2.61 27.11 1.99 �0.001 5.87 3.10
CLN 49.24 4.16 4.58 0.65 32.44 2.75 �0.001 3.94 1.67 38.35 3.47 �0.001 7.93 1.90SMLN 43.51 2.94 8.36 3.26 43.17 3.61 NS 5.11 1.53 27.54 3.69 �0.001 3.96 1.24
a Lymphocytes from BALB/c mice immunized with formulated gp160, gp140cyt, or gp140sec DNA were isolated 6 wk into the experiment and 51Cr release assays wereconducted as described in Fig. 1. Freshly isolated (ex vivo) or 5-day Ag-restimulated cells were measured.
b Lymphocytes were isolated from lungs, spleen, LRLN, CLN, and SMLN.c The specificity of lysis of targets expressing Env vs, �gal was significant ( p � 0.001).d Mean SE of three experiments at E:T 50:1.e p value against gp160 DNA-vaccinated mice.
FIGURE 1. Immunization of BALB/cmice i.n. with protein �1-PL-DNA en-coding either HIV gp160 (A and D) andHIV gp140cyt (B and E) or gp140sec (Cand F) result in CTL responses specificfor Env. Lymphocytes were isolatedfrom lungs, spleens, LRLN, CLN, andSMLN 6 wk after primary immuniza-tion. CTL activity was assessed foreach tissue using freshly isolated lym-phocytes or ex vivo (A–C) and for invitro Ag-restimulated lymphocytes(D-–F) using standard 51Cr release as-says. CTL assays were done againstspecific target BC-Env cells and irrel-evant control target BC-�gal cells.Data are representative of five experi-ments and are expressed as mean SDof triplicate wells.
4719The Journal of Immunology
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
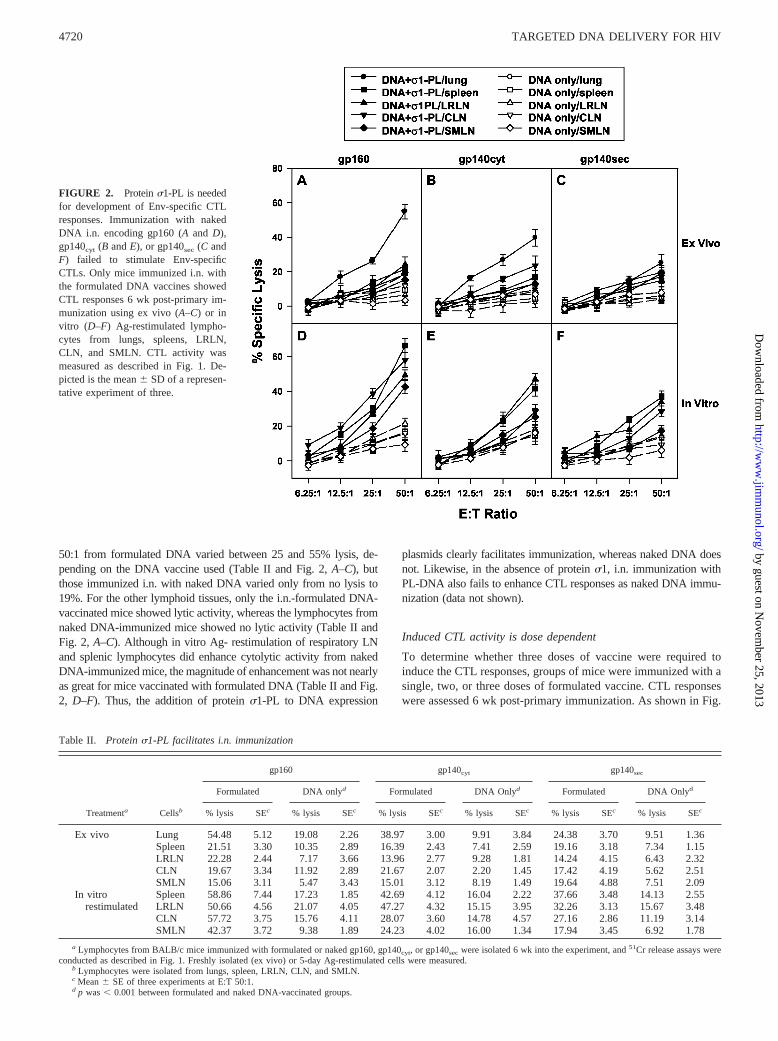
50:1 from formulated DNA varied between 25 and 55% lysis, de-pending on the DNA vaccine used (Table II and Fig. 2, A–C), butthose immunized i.n. with naked DNA varied only from no lysis to19%. For the other lymphoid tissues, only the i.n.-formulated DNA-vaccinated mice showed lytic activity, whereas the lymphocytes fromnaked DNA-immunized mice showed no lytic activity (Table II andFig. 2, A–C). Although in vitro Ag- restimulation of respiratory LNand splenic lymphocytes did enhance cytolytic activity from nakedDNA-immunized mice, the magnitude of enhancement was not nearlyas great for mice vaccinated with formulated DNA (Table II and Fig.2, D–F). Thus, the addition of protein �1-PL to DNA expression
plasmids clearly facilitates immunization, whereas naked DNA doesnot. Likewise, in the absence of protein �1, i.n. immunization withPL-DNA also fails to enhance CTL responses as naked DNA immu-nization (data not shown).
Induced CTL activity is dose dependent
To determine whether three doses of vaccine were required toinduce the CTL responses, groups of mice were immunized with asingle, two, or three doses of formulated vaccine. CTL responseswere assessed 6 wk post-primary immunization. As shown in Fig.
FIGURE 2. Protein �1-PL is neededfor development of Env-specific CTLresponses. Immunization with nakedDNA i.n. encoding gp160 (A and D),gp140cyt (B and E), or gp140sec (C andF) failed to stimulate Env-specificCTLs. Only mice immunized i.n. withthe formulated DNA vaccines showedCTL responses 6 wk post-primary im-munization using ex vivo (A–C) or invitro (D–F) Ag-restimulated lympho-cytes from lungs, spleens, LRLN,CLN, and SMLN. CTL activity wasmeasured as described in Fig. 1. De-picted is the mean SD of a represen-tative experiment of three.
Table II. Protein �1-PL facilitates i.n. immunization
Treatmenta Cellsb
gp160 gp140cyt gp140sec
Formulated DNA onlyd Formulated DNA Onlyd Formulated DNA Onlyd
% lysis SEc % lysis SEc % lysis SEc % lysis SEc % lysis SEc % lysis SEc
Ex vivo Lung 54.48 5.12 19.08 2.26 38.97 3.00 9.91 3.84 24.38 3.70 9.51 1.36Spleen 21.51 3.30 10.35 2.89 16.39 2.43 7.41 2.59 19.16 3.18 7.34 1.15LRLN 22.28 2.44 7.17 3.66 13.96 2.77 9.28 1.81 14.24 4.15 6.43 2.32CLN 19.67 3.34 11.92 2.89 21.67 2.07 2.20 1.45 17.42 4.19 5.62 2.51SMLN 15.06 3.11 5.47 3.43 15.01 3.12 8.19 1.49 19.64 4.88 7.51 2.09
In vitro Spleen 58.86 7.44 17.23 1.85 42.69 4.12 16.04 2.22 37.66 3.48 14.13 2.55restimulated LRLN 50.66 4.56 21.07 4.05 47.27 4.32 15.15 3.95 32.26 3.13 15.67 3.48
CLN 57.72 3.75 15.76 4.11 28.07 3.60 14.78 4.57 27.16 2.86 11.19 3.14SMLN 42.37 3.72 9.38 1.89 24.23 4.02 16.00 1.34 17.94 3.45 6.92 1.78
a Lymphocytes from BALB/c mice immunized with formulated or naked gp160, gp140cyt, or gp140sec were isolated 6 wk into the experiment, and 51Cr release assays wereconducted as described in Fig. 1. Freshly isolated (ex vivo) or 5-day Ag-restimulated cells were measured.
b Lymphocytes were isolated from lungs, spleen, LRLN, CLN, and SMLN.c Mean SE of three experiments at E:T 50:1.d p was � 0.001 between formulated and naked DNA-vaccinated groups.
4720 TARGETED DNA DELIVERY FOR HIV
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
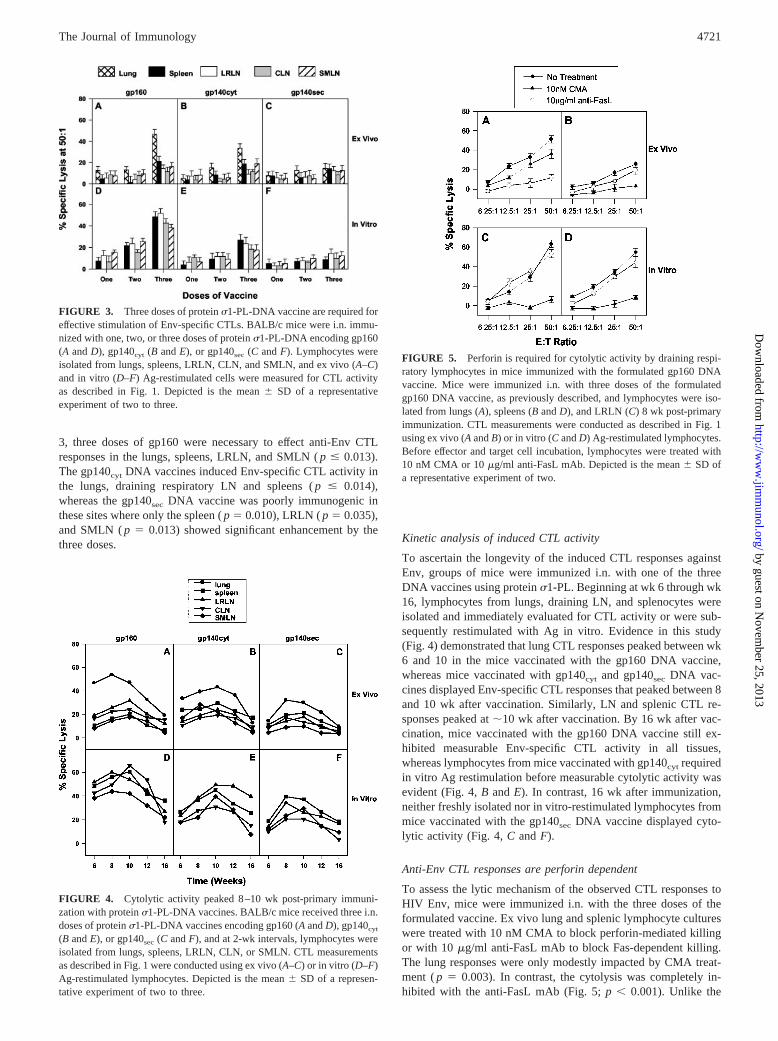
3, three doses of gp160 were necessary to effect anti-Env CTLresponses in the lungs, spleens, LRLN, and SMLN ( p � 0.013).The gp140cyt DNA vaccines induced Env-specific CTL activity inthe lungs, draining respiratory LN and spleens ( p � 0.014),whereas the gp140sec DNA vaccine was poorly immunogenic inthese sites where only the spleen ( p 0.010), LRLN ( p 0.035),and SMLN ( p 0.013) showed significant enhancement by thethree doses. Kinetic analysis of induced CTL activity
To ascertain the longevity of the induced CTL responses againstEnv, groups of mice were immunized i.n. with one of the threeDNA vaccines using protein �1-PL. Beginning at wk 6 through wk16, lymphocytes from lungs, draining LN, and splenocytes wereisolated and immediately evaluated for CTL activity or were sub-sequently restimulated with Ag in vitro. Evidence in this study(Fig. 4) demonstrated that lung CTL responses peaked between wk6 and 10 in the mice vaccinated with the gp160 DNA vaccine,whereas mice vaccinated with gp140cyt and gp140sec DNA vac-cines displayed Env-specific CTL responses that peaked between 8and 10 wk after vaccination. Similarly, LN and splenic CTL re-sponses peaked at �10 wk after vaccination. By 16 wk after vac-cination, mice vaccinated with the gp160 DNA vaccine still ex-hibited measurable Env-specific CTL activity in all tissues,whereas lymphocytes from mice vaccinated with gp140cyt requiredin vitro Ag restimulation before measurable cytolytic activity wasevident (Fig. 4, B and E). In contrast, 16 wk after immunization,neither freshly isolated nor in vitro-restimulated lymphocytes frommice vaccinated with the gp140sec DNA vaccine displayed cyto-lytic activity (Fig. 4, C and F).
Anti-Env CTL responses are perforin dependent
To assess the lytic mechanism of the observed CTL responses toHIV Env, mice were immunized i.n. with the three doses of theformulated vaccine. Ex vivo lung and splenic lymphocyte cultureswere treated with 10 nM CMA to block perforin-mediated killingor with 10 �g/ml anti-FasL mAb to block Fas-dependent killing.The lung responses were only modestly impacted by CMA treat-ment ( p 0.003). In contrast, the cytolysis was completely in-hibited with the anti-FasL mAb (Fig. 5; p � 0.001). Unlike the
FIGURE 3. Three doses of protein �1-PL-DNA vaccine are required foreffective stimulation of Env-specific CTLs. BALB/c mice were i.n. immu-nized with one, two, or three doses of protein �1-PL-DNA encoding gp160(A and D), gp140cyt (B and E), or gp140sec (C and F). Lymphocytes wereisolated from lungs, spleens, LRLN, CLN, and SMLN, and ex vivo (A–C)and in vitro (D–F) Ag-restimulated cells were measured for CTL activityas described in Fig. 1. Depicted is the mean SD of a representativeexperiment of two to three.
FIGURE 4. Cytolytic activity peaked 8–10 wk post-primary immuni-zation with protein �1-PL-DNA vaccines. BALB/c mice received three i.n.doses of protein �1-PL-DNA vaccines encoding gp160 (A and D), gp140cyt
(B and E), or gp140sec (C and F), and at 2-wk intervals, lymphocytes wereisolated from lungs, spleens, LRLN, CLN, or SMLN. CTL measurementsas described in Fig. 1 were conducted using ex vivo (A–C) or in vitro (D–F)Ag-restimulated lymphocytes. Depicted is the mean SD of a represen-tative experiment of two to three.
FIGURE 5. Perforin is required for cytolytic activity by draining respi-ratory lymphocytes in mice immunized with the formulated gp160 DNAvaccine. Mice were immunized i.n. with three doses of the formulatedgp160 DNA vaccine, as previously described, and lymphocytes were iso-lated from lungs (A), spleens (B and D), and LRLN (C) 8 wk post-primaryimmunization. CTL measurements were conducted as described in Fig. 1using ex vivo (A and B) or in vitro (C and D) Ag-restimulated lymphocytes.Before effector and target cell incubation, lymphocytes were treated with10 nM CMA or 10 �g/ml anti-FasL mAb. Depicted is the mean SD ofa representative experiment of two.
4721The Journal of Immunology
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

lung, the splenic CTL were mostly inhibited by the CMA treatment(Fig. 5B; p � 0.001). Ag-restimulated splenic and LRLN lympho-cytes showed the expected increased cytolytic activity, which wasinhibited entirely by CMA treatment ( p � 0.001), whereas theanti-FasL treatment was partially inhibitory for splenic responses( p 0.012; Fig. 5, C and D). Collectively, these data demonstratethat perforin is important for immunity induced by formulatedgp160 vaccination in the draining respiratory LN.
IFN-� is important for initial CTL response to Env
To assess the contribution of IFN-� to cytolytic activity induced byprotein �1-PL-gp160 vaccination, both BALB/c and IFN-��/�
mice were vaccinated i.n. Freshly isolated lymphocytes from IFN-��/� lungs, respiratory LN, and spleen failed to exhibit cytolyticactivity when compared with BALB/c responses ( p � 0.010; Fig.6, A and B). In contrast, in vitro Ag restimulation showed vigorouskilling by IFN-��/� lymphocytes, indicating that the Env-specificCTLs did not require IFN-� for lytic activity.
To assess the cytokine profiles subsequent to Ag restimulation,supernatants from Ag-restimulated lymphoid cultures from spleen,LRLN, CLN, and SMLN were assayed by cytokine-specificELISAs. The three vaccines showed preferential stimulation ofIFN-� and IL-2 production when compared with IL-4, IL-5, IL-6,or IL-10 levels (Fig. 7).
Vaccinia-Env challenge of mice vaccinated with formulatedgp160 DNA vaccine
The vaccinia virus challenge bearing an HIV protein is generallyused to assess levels of cell-mediated immunity (35). To assess thepotential efficacy of our formulated gp160 DNA vaccine, BALB/c
mice were given the formulated or naked gp160 DNA vaccine viathe i.n. route as previously described. As a positive control, onegroup of mice were immunized i.m. with the same gp160 DNAvaccine at the same time intervals and with the same doses. Thelast group of mice were left unimmunized to serve as a naivecontrol. Approximately 16 wk after primary immunization, eachgroup was challenged with vaccinia-env, and ovaries were as-sessed for levels of vaccinia PFU. It was quite evident that thegroup of mice given the formulated gp160 DNA vaccine showedpotent antiviral immunity when compared with challenged naivemice or other immunized groups (Fig. 8). Thus, these data show
FIGURE 6. Env-specific CTLs are induced in IFN-��/� mice. BALB/c(A and C) and IFN-��/� (B and D) mice were i.n. immunized with three dosesof formulated gp160 DNA vaccine as previously described. Lymphocyteswere isolated from lungs, respiratory LN, and spleens 6 wk post-primaryimmunization. CTL measurements were conducted as described in Fig. 1using (A and B) ex vivo or in vitro (C and D) Ag-restimulated lympho-cytes. Depicted is the mean SD of a representative experiment of three.
FIGURE 7. Ag-restimulated respiratory LN and splenic lymphocytecultures showed elevated levels of IFN-� and IL-2. Cytokine-specificELISAs were performed on supernatants collected from Ag-restimulatedcultures from the mice immunized with gp160 (A), gp140cyt (B), orgp140sec (C) DNA vaccines. Each vaccine showed preferential stimulationof IFN-� and IL-2 after immune lymphocytes were cultured for 5 days withBC-Env cells. Minimal to no Th2-type cytokines were produced. Depictedis the mean SEM of three experiments. Statistical differences betweengp160 DNA vs gp140cyt and gp140sec DNA-vaccinated groups are: �, p �
0.001; ��, p � 0.018; ���, p � 0.05.
FIGURE 8. Immunization of female BALB/c mice i.n. with formulatedgp160 DNA vaccine showed vigorous antiviral immunity subsequent tochallenge with vaccinia virus-env. Groups of mice received i.n. immuni-zations with the formulated or naked gp160 vaccine, i.m. immunizationwith naked gp160 vaccine, or were left unvaccinated (naive controls). Six-teen weeks after primary immunization, mice were given vaccinia-env i.p.,and 6 days later, ovaries were harvested. At the 99% confidence intervals:�, p � 0.0001, ��, p � 0.001, and ���, p � 0.01.
4722 TARGETED DNA DELIVERY FOR HIV
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

that protein �1 targeting is an effective means to stimulate hostresponses via the i.n. route.
DiscussionA unique attribute of mucosal inductive tissues is the specializedepithelium covering these organized mucosal lymphoid tissues,termed follicle-associated epithelium (FAE). Within the FAE is adifferentiated epithelial cell subset referred to as M cells. M cellsare adept at uptake and transport of luminal Ags, including solubleproteins and infectious agents. It is also believed that certain patho-gens, such as Salmonella typhimurium (37), Yersinia enteroco-litica (38), and reoviruses (22), invade the host tissues by bindingto M cells. To take advantage of this specialized niche, we hy-pothesize that targeting M cells using proteins derived from suchinfectious agents would serve as M cell ligands. One such ligand,reovirus protein �1, was previously shown in its recombinant formto bind M cells and facilitate i.n. immunization using expressionplasmid reporter gene constructs (26). Both transfection (26) andimmunization (27) required covalent attachment of protein �1 toPL to enable complexing to DNA vaccine. It was evident in thisprevious study (27) that targeting was necessary to generate mu-cosal responses because naked or PL-complexed DNA failed tostimulate mucosal Ab or CTL responses.
To expand on the earlier observation that the protein �1-PL-DNA delivery system can stimulate CTL responses, work de-scribed here focused on the adaptation of this technique to stim-ulate cell-mediated anti-HIV Env responses. The best results wereobtained using the expression plasmid encoding gp160 rather thangp140cyt or gp140sec. The reason that differences were obtainedwas not resolved, although the mode of expression may have im-pacted the immunogenicity of Env in these constructs. Minimally,we know that it was not attributed to differences in protein pro-duction because transfection assays with the three DNA vaccinesyielded similar levels of expressed protein (data not shown). Wehad expected the secreted gp140 to induce CTL responses resem-bling those induced by membrane-bound gp160, but the magnitudeof CTL responses between the two vaccines was notably different.Although some improvement in host CTL responses was observedbetween 8 and 10 wk post-primary immunization, the gp160 vac-cine was clearly the most effective to stimulate enhanced CTLresponses when given i.n., and these CTL responses continued tobe evident for at least 16 wk after primary immunization. Perhapsone possibility to account for the increased immunogenicitymay be related to the increased size of gp160 making it moreimmunogenic or providing additional target peptides. In con-trast, CTL responses induced by gp140sec waned 16 wk aftervaccination and were undetectable even after in vitro Ag re-stimulation. Finally, these results showed that at least threedoses of vaccine were required to obtain the observed magni-tude in CTL responses.
The potency and longevity of these CTL responses were alsoassessed using the vaccinia-env challenge similar to that previ-ously described (35, 36). As our in vitro analyses suggested, theformulated gp160 DNA vaccine induced potent antiviral immunityas evidenced by the reduced vaccinia virus PFUs when comparedwith mice given the same vaccine by the naked i.n. or i.m. routes.Such data support our contention that targeting vaccines can en-hance immune responses. It is also important that only one previ-ous study (36) has shown a similar extent of cell-mediated re-sponses when a DNA vaccine was given i.n. However, thisrequired a live vector to carry the DNA vaccine but again suggeststhat targeting mucosal inductive tissues may be important for suc-
cessful vaccination (39). Thus, a DNA subunit vaccine is capableof eliciting strong cell-mediated immunity if it can be effectivelytargeted using our approach or by live vectors.
The CTL responses acquired using the gp160 vaccine were ac-complished in the absence of adjuvant. Typically, successful i.n.DNA vaccination against HIV immunogens has required the use ofa live vector, as in the case for adenovirus (40, 41), vesicularstomatitis virus (42), or a bacterial vector (30). Otherwise, adju-vant is coadministered (43) or incorporated into the DNA (44). Inaddition, the enthusiasm for administering adjuvants i.n. has beendampened by recent data showing olfactory bulb apoptosis (45,46). Our data suggest that effective targeting to mucosal inductivetissues may circumvent the need for additional adjuvants and stillbe able to stimulate long-lasting CTL responses; thus, effectivevaccine targeting potentially can supplant the need for addi-tional adjuvants. Current studies are assessing these possibili-ties. Collectively, from this study, as well as those described forlive vector vaccines (30, 40 – 42) and incorporation to lipo-somes, it is evident that for i.n. delivery of DNA vaccines, theDNA must be protected or targeted to mucosal inductive sites tofacilitate vaccination. Otherwise, as shown here, application ofnaked DNA to the nasal passages fails to implement effectivetransgene vaccination.
CD8� T cells do appear to play an active role in the control oflentivirus infections, either by cytolytic mechanisms or productionof �-chemokines. In experimental models, depletion of CD8� Tcells in SIV-infected rhesus monkeys (47) or HIV infection inmice (48) implicate the role of CD8� T cells in the control vire-mia. Perforin, one of the cytotoxins released by immune CTLs,mediates lysis of infected target cells by facilitating entry of pro-teases or granzymes into target cells. Recent data have suggestedthat during chronic HIV infection, HIV-specific CTLs are inducedbut express low levels of perforin (49–52), despite the ability toexpress granzyme A (51, 53). Given these findings, a vaccine thatstimulates perforin-dependent, HIV-specific CTL responses mayprovide a benefit to the vaccinee. Evidence reported above showsthat one attribute of the protein �1-PL-DNA delivery is the ca-pacity to stimulate perforin-dependent CTL responses followingi.n. vaccination.
Coupled to the ability of gp160 DNA vaccine to induce CTLresponses was the capacity of immune T cells to produce of IFN-�.Ag-restimulated T cell cultures showed enhanced production ofIFN-� and IL-2, whereas only low levels of cytokines associatedwith Th2/Tc2 cells were detected in these cultures. In a previousstudy (27), Th2 cytokines were detected after Ag restimulation ofCD4� T cells; however, cytokine responses by immune CD8� Tcells were not evaluated at that time. Future studies will evaluatethe ability of the gp160, as well as other HIV DNA vaccines tostimulate systemic and mucosal Abs against Env, and determinewhich Th cell subsets support these responses. Such evaluationswere beyond the scope of these current studies.
In summary, a method is described for DNA vaccination via thei.n. route. Thus, DNA vaccines induce perforin-dependent CTLresponses after i.n. administration in formulations containing the�1-PL-DNA delivery fusion protein. Of three DNA vaccinestested, the plasma membrane expressed gp160 DNA vaccine wasthe most immunogenic for CTL responses, suggesting that themode of expression influences the immunogenicity of DNA vac-cine-expressed immunogens delivered via the i.n. route. The abil-ity to target respiratory mucosal inductive sites in the respiratorytract aided by the ability to elicit CTL responses in the lungs, as
4723The Journal of Immunology
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

well as in the draining respiratory LN, will provide a useful tool todevelop vaccines against pathogens that target mucosal surfaces,such as HIV-1 and influenza.
AcknowledgmentsSpecial thanks go to Nancy Kommers for her assistance in preparing themanuscript.
References1. Rennels, M. B., R. I. Glass, P. H. Dennehy, D. I. Bernstein, M. E. Pichichero,
E. T. Zito, M. E. Mack, B. L. Davidson, and A. Z. Kapikian. 1996. Safety andefficacy of high-dose rhesus-human reassortant rotavirus vaccines: report of theNational Multicenter Trial. United States Rotavirus Vaccine Efficacy Group. Pe-diatrics 97:7.
2. Belshe, R. B., C. Stevens, G. J. Gorse, S. Buchbinder, K. Weinhold, H. Sheppard,D. Stablein, S. Self, J. McNamara, S. Frey, et al. 2001. Safety and immunoge-nicity of a canarypox-vectored human immunodeficiency virus type 1 vaccinewith or without gp120: a phase 2 study in higher- and lower-risk volunteers.J. Infect. Dis. 183:1343.
3. Ross, T. M., Y. Xu, R. A. Bright, and H. L. Robinson. 2000. C3d enhancementof antibodies to hemagglutinin accelerates protection against influenza virus chal-lenge. Nat. Immunol. 1:127.
4. Fu, T. M., A. Friedman, J. B. Ulmer, M. A. Liu, and J. J. Donnelly. 1997.Protective cellular immunity: cytotoxic T-lymphocyte responses against domi-nant and recessive epitopes of influenza virus nucleoprotein induced by DNAimmunization. J. Virol. 71:2715.
5. Gallichan, W. S., and K. L. Rosenthal. 1998. Long-term immunity and protectionagainst herpes simplex virus type 2 in the murine female genital tract after mu-cosal but not systemic immunization. J. Infect. Dis. 177:1155.
6. Boyer, J. D., B. Wang, K. E. Ugen, M. Agadjanyan, A. Javadian, P. Frost,K. Dang, R. A. Carrano, R. Ciccarelli, L. Coney, W. V. Williams, andD. B. Weiner. 1996. In vivo protective anti-HIV immune responses in non-humanprimates through DNA immunization. J. Med. Primatol. 25:242.
7. Boyer, J. D., K. E. Ugen, M. Chattergoon, B. Wang, A. Shah, M. Agadjanyan,M. L. Bagarazzi, A. Javadian, R. Carrano, L. Coney, W. V. Williams, andD. B. Weiner. 1997. DNA vaccination as anti-human immunodeficiency virusimmunotherapy in infected chimpanzees. J. Infect. Dis. 176:1501.
8. Simon, M., M. L. Gern, P. Hauser, W. Zhong, P. J. Nielsen, M. D. Kramer,C. Brenner, and R. Wallich. 1996. Protective immunization with plasmid DNAcontaining the outer surface lipoprotein A gene of Borrelia burgdorferi is inde-pendent of an eukaryotic promoter. Eur. J. Immunol. 26:2831.
9. Feltquate, D. M., S. Heaney, R. G. Webster, and H. L. Robinson. 1997. DifferentT helper cell types and antibody isotypes generated by saline and gene gun DNAimmunization. J. Immunol. 158:2278.
10. Noll, A., N. Bucheler, E. Bohn, R. Schirmbeck, J. Reimann, and I. B. Austenite.1999. DNA immunization confers systemic, but not mucosal, protection againstenteroinvasive bacteria. Eur. J. Immunol. 29:986.
11. Chen, S. C., D. H. Jones, E. F. Fynan, G. H. Farrar, J. C. Clegg,H. B. Greenberg, and J. E. Herrmann. 1998. Protective immunity induced byoral immunization with a rotavirus DNA vaccine encapsulated in micropar-ticles. J. Virol. 72:5757.
12. Herrmann, J. E., S. C. Chen, D. H. Jones, A. Tinsley-Bown, E. F. Fynan,H. B. Greenberg, and G. H. Farrar. 1999. Immune responses and protection ob-tained by oral immunization with rotavirus VP4 and VP7 DNA vaccines encap-sulated in microparticles. Virology 259:148.
13. Eldridge, J. H., J. K. Staas, J. A. Meulbroek, J. R. McGhee, T. R. Tice, andR. M. Gilley. 1991. Biodegradable microspheres as a vaccine delivery system.Mol. Immunol. 28:287.
14. Jepson, M. A., N. L. Simmons, D. T. O’Hagan, and B. H. Hirst. 1993. Compar-ison of poly (DL-lactide-co-glycolide) and polystyrene microsphere targeting tointestinal M cells. J. Drug Target 1:245.
15. Wu, C. H., J. M. Wilson, and G. Y. Wu. 1989. Targeting genes: delivery andpersistent expression of a foreign gene driven by mammalian regulatory elementsin vivo. J. Biol. Chem. 264:16985.
16. Wu, G. Y., P. Zhan, L. L. Sze, A. R. Rosenberg, and C. H. Wu. 1994. Incorpo-ration adenovirus into a ligand-based DNA carrier system results in retention oforiginal specificity and enhances targeted gene expression. J. Biol. Chem.269:11542.
17. Wagner, E., M. Zenke, M. Cotten, H. Beug, and. M. L. Birnstiel. 1990. Trans-ferrin-polycation conjugates as carriers for DNA uptake into cells. Proc. Natl.Acad. Sci. USA 87:3410.
18. Wagner, E., K. Zatloukal, M. Cotten, H. Kirlappos, K. Mechtler, D. T. Curiel, andM. L. Birnstiel. 1992. Coupling of adenovirus to transferrin-polylysine/DNAcomplexes greatly enhances receptor-mediated gene delivery and expression oftransfected genes. Proc. Natl. Acad. Sci. USA 89:6099.
19. Lozier, J. N., A. R. Thompson, P. C. Hu, M. Read, K. M. Brinkhous, K. A. High,and D. T. Curiel. 1994. Efficient transfection of primary cells in a canine hemo-philia B model using adenovirus-polylysine-DNA complexes. Hum. Gene Ther.5:313.
20. Batra, R. K., F. Wang-Johanning, E. Wagner, R. I. Garver, Jr., and D. T. Curiel.1994. Receptor-mediated gene delivery employing lectin-binding specificity.Gene Ther. 1:255.
21. Sobolev, A. S., A. A. Rosenkranz, O. A. Smirnova, V. A. Nikitin,G. L. Neugodova, B. S. Naroditsky, I. N. Shilov, I. N. Shatski, and L. K. Ernst.1998. Receptor-mediated transfection of murine and ovine mammary glands invivo. J. Biol. Chem. 273:7928.
22. Lee, P. W., E. C. Hayes, and W. K. Joklik. 1981. Protein �1 is the reovirus cellattachment protein. Virology 108:156.
23. Mah, D. C., G. Leone, J. M. Jankowski, and P. W. Lee. 1990. The N-terminalquarter of reovirus cell attachment protein �1 possesses intrinsic virion-anchoringfunction. Virology 179:95.
24. Strong, J. E., G. Leone, R. Duncan, R. K. Sharma, and P. W. Lee. 1991. Bio-chemical and biophysical characterization of the reovirus cell attachment protein�1: evidence that it is a homotrimer. Virology 184:23.
25. Bassel-Duby, R., M. L. Nibert, C. J. Homcy, B. N. Fields, and D. G. Sawutz.1987. Evidence that the �1 protein of reovirus serotype 3 is a multimer. J. Virol.61:1834.
26. Wu, Y., M. J. Boysun, K. L. Csencsits, and D. W. Pascual. 2000. Gene transferfacilitated by a cellular targeting molecule, reovirus protein �1. Gene Ther. 7:61.
27. Wu, Y., X. Wang, K. L. Csencsits, A. Haddad, N. Walters, and D. W. Pascual.2001. M cell-targeted DNA vaccination. Proc. Natl. Acad. Sci. USA 98:9318.
28. Dalton, D. K., S. Pitts-Meek, S. Keshav, I. S. Figari, A. Bradley, andT. A. Stewart. 1993. Multiple defects of immune cell function in mice withdisrupted interferon-� genes. Science 259:1739.
29. Qian, Q., and J. E. Cutler. 1997. � Interferon is not essential in host defenseagainst disseminated candidiasis in mice. Infect. Immun. 65:1748.
30. Shata, M. T., M. S. Reitz, Jr., A. L. DeVico, G. K. Lewis, and D. M. Hone.2001. Mucosal and systemic HIV-1 Env-specific CD8� T-cells develop afterintragastric vaccination with a Salmonella Env DNA vaccine vector. Vaccine20:623.
31. Pascual, D. W., N. Walters, and P. Hillemeyer. 1998. Repeated intratrachealinstillations of nonreplicating adenovirus 2 vector attenuate CTL responses andIFN-� production. J. Immunol. 160:4465.
32. Chada, S., C. E. DeJesus, K. Townsend, W. T. Lee, L. Laube, D. J. Jolly,S. M. Chand, and J. F. Warner. 1993. Cross-reactive lysis of human targetsinfected with prototypic and clinical human immunodeficiency virus type 1(HIV-1) strains by murine anti-HIV-1IIIB Env-specific cytotoxic T lymphocytes.Virology 67:3409.
33. Kataoka, T., N. Shinohara, H. Takayama, K. Takaku, S. Kondo, S. Yonehara, andK. Nagai. 1996. Concanamycin A, a powerful tool for characterization and es-timation of contribution of perforin- and Fas-based lytic pathways in cell-medi-ated cytotoxicity. J. Immunol. 156:3678.
34. Hillemeyer, P., M. D. White, and D. W. Pascual. 2002. Development of a tran-sient CD4�CD8� T cell subset in the cervical lymph nodes following intratra-cheal instillation with an adenovirus vector. Cell. Immunol. 215:173.
35. Belyakov, I. M., M. A. Derby, J. D. Ahlers, B. L. Kelsall, P. Earl, B. Moss,W. Strober, and J. A. Berzofsky. 1998. Mucosal immunization with HIV-1 pep-tide vaccine induces mucosal and systemic cytotoxic T lymphocytes and protec-tive immunity in mice against intrarectal recombinant HIV-vaccinia challenge.Proc. Natl. Acad. Sci. USA 95:1709.
36. Shata, M. T., and D. M. Hone. 2001. Vaccination with a Shigella DNA vaccinevector induces antigen-specific CD8� T cells and antiviral protective immunity.J. Virol. 75:9665.
37. Jones, B. D., N. Ghori, and S. Falkow. 1994. Salmonella typhimurium initiatesmurine infection by penetrating and destroying the specialized epithelial M cellsof the Peyer’s patches. J. Exp. Med. 180:15.
38. Clark, M. A., B. H. Hirst, and M. A. Jepson. 1998. M-cell surface beta 1 integrinexpression and invasin-mediated targeting of Yersinia pseudotuberculosis tomouse Peyer’s patch M cells. Infect. Immun. 1. 66:1237.
39. Siebers, A., and B. B. Finlay. 1996. M cells and the pathogenesis of mucosal andsystemic infections. Trends Microbiol. 4:22.
40. Lubeck, M. D., R. J. Natuk, M. Chengalvala, P. K. Chanda, K. K. Murthy,S. Murthy, S. Mizutani, S. G. Lee, M. S. Wade, B. M. Bhat, et al. 1994. Immu-nogenicity of recombinant adenovirus-human immunodeficiency virus vaccinesin chimpanzees following intranasal administration. AIDS Res. Hum. Retrovi-ruses 10:1443.
41. Shiver, J. W., T. M. Fu, L. Chen, D. R. Casimiro, M. E. Davies, R. K. Evans,Z. O. Zhang, A. J. Simon, W. L. Trigona, S. A. Dubey, et al. 2002. Replication-incompetent adenoviral vaccine vector elicits effective anti-immunodeficiency-virus immunity. Nature 415:331.
42. Haglund, K., I. Leiner, K. Kerksiek, L. Buonocore, E. Pamer, and J. K. Rose.2002. Robust recall and long-term memory T-cell responses induced by prime-boost regimens with heterologous live viral vectors expressing human immuno-deficiency virus type 1 Gag and Env proteins. J. Virol. 76:7506.
43. Sasaki, S., K. Sumino, K. Hamajima, J. Fukushima, N. Ishii, S. Kawamoto,H. Mohri, C. R. Kensil, and K. Okuda. 1998. Induction of systemic and mucosalimmune responses to human immunodeficiency virus type 1 by a DNA vaccineformulated with QS-21 saponin adjuvant via intramuscular and intranasal routes.J. Virol. 72:4931.
44. Okada, E., S. Sasaki, N. Ishii, I. Aoki, T. Yasuda, K. Nishioka, J. Fukushima,J. Miyazaki, B. Wahren, and K. Okuda. 1997. Intranasal immunization of aDNA vaccine with IL-12- and granulocyte-macrophage colony-stimulatingfactor (GM-CSF)-expressing plasmids in liposomes induces strong mucosaland cell-mediated immune responses against HIV-1 antigens. J. Immunol.159:3638.
45. van Ginkel, F. W., R. J. Jackson, Y. Yuki, and J. R. McGhee. 2000. Cutting edge:the mucosal adjuvant cholera toxin redirects vaccine proteins into olfactory tis-sues. J. Immunol. 165:4778.
4724 TARGETED DNA DELIVERY FOR HIV
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

46. Fujihashi, K., T. Koga, F. W. van Ginkel, Y. Hagiwara, and J. R. McGhee. 2002.A dilemma for mucosal vaccination: efficacy versus toxicity using enterotoxin-based adjuvants. Vaccine 20:2431.
47. Schmitz, J. E., M. J. Kuroda, S. Santra, V. G. Sasseville, M. A. Simon,M. A. Lifton, P. Racz, K. Tenner-Racz, M. Dalesandro, B. J. Scallon, et al. 1999.Control of viremia in simian immunodeficiency virus infection by CD8� lym-phocytes. Science 283:857.
48. Belyakov, I. M., J. D. Ahlers, B. Y. Brandwein, P. Earl, B. L. Kelsall, B. Moss,W. Strober, and J. A. Berzofsky. 1998. The importance of local mucosal HIV-specific CD8� cytotoxic T lymphocytes for resistance to mucosal viral transmis-sion in mice and enhancement of resistance by local administration of IL-12.J. Clin. Invest. 102:2072.
49. Shankar, P., Z. Xu, and J. Lieberman. 1999. Viral-specific cytotoxic T lympho-cytes lyse human immunodeficiency virus-infected primary T lymphocytes by thegranule exocytosis pathway. Blood 94:3084.
50. Appay, V., D. F. Nixon, S. M. Donahoe, G. M. Gillespie, T. Dong, A. King,
G. S. Ogg, H. M. Spiegel, C. Conlon, C. A. Spina, et al. 2000. HIV-specificCD8� T cells produce antiviral cytokines but are impaired in cytolytic function.J. Exp. Med. 192:63.
51. Andersson, J., S. Kinloch, A. Sonnerborg, J. Nilsson, T. E. Fehniger,A. L. Spetz, H. Behbahani, L. E. Goh, H. McDade, B. Gazzard, et al. 2002.Low levels of perforin expression in CD8� T lymphocyte granules in lym-phoid tissue during acute human immunodeficiency virus type 1 infection.J. Infect. Dis. 185:1355.
52. Zhang, D., P. Shankar, Z. Xu, B. Harnisch, G. Chen, C. Lange, S. J. Lee,H. Valdez, M. M. Lederman, and J. Lieberman. 2003. Most antiviral CD8 T cellsduring chronic viral infection do not express high levels of perforin and are notdirectly cytotoxic. Blood 101:226.
53. Andersson, J., H. Behbahani, J. Lieberman, E. Connick, A. Landay, B. Patterson,A. Sonnerborg, K. Lore, S. Uccini, and T. E. Fehniger. 1999. Perforin is not co-expressed with granzyme A within cytotoxic granules in CD8 T lymphocytes presentin lymphoid tissue during chronic HIV infection. AIDS 13:1295.
4725The Journal of Immunology
by guest on Novem
ber 25, 2013http://w
ww
.jimm
unol.org/D
ownloaded from




















