
Induction of specific T cell immunity in patients with prostate cancer by vaccination with...
10
ORIGINAL ARTICLE Supriya Perambakam Sigrun Hallmeyer Samarth Reddy Nadim Mahmud Linda Bressler Phillip DeChristopher Delores Mahmud Rafael Nunez Jeffrey A. Sosman David J. Peace Induction of specific T cell immunity in patients with prostate cancer by vaccination with PSA146–154 peptide Received: 25 May 2005 / Accepted: 3 October 2005 / Published online: 10 November 2005 Ó Springer-Verlag 2005 Abstract T cell immunotherapy of prostate cancer (CaP) offers the potential for less toxic, more effective outcomes. A clinical trial was conducted in 28 patients with locally advanced or metastatic CaP to determine whether an HLA-A2 binding epitope of prostate-specific antigen, PSA146–154 (PSA-peptide), can induce specific T cell immunity. Patients were vaccinated either by intradermal injection of PSA-peptide and GM-CSF or by intravenous administration of autologous dendritic cells pulsed with PSA-peptide at weeks 1, 4 and 10. Delayed-type hyper- sensitivity (DTH) skin testing was performed at weeks 4, 14, 26 and 52. Fifty percent of the patients developed positive DTH responses to PSA-peptide. The size of the DTH induration progressively increased over time in the majority of responding patients. Skin biopsies from seven DTH-positive patients were available and T cells that developed in situ were also characterized. The phenotype of recovered T cells demonstrated variable proportions of CD4 + CD8 À , CD4 À CD8 + and CD4 + CD8 + T cell pop- ulations. Cytokine analysis of PSA-peptide stimulated T cells per bead array assay exhibited specific IFN-c and TNF-a response in six of seven patients. Specific IL-4 response was observed in five patients, while IL-10 response was detected in one patient. Purified CD4 À CD8 + T cells isolated from four patients demon- strated specific cytolytic activity per chromium release assay. In conclusion, immunization with PSA-peptide induced specific T cell immunity in one-half of the patients with locally advanced and hormone-sensitive, metastatic CaP. DTH-derived T cells exhibited PSA- peptide-specific cytolytic activity and predominantly expressed a type-1 cytokine profile. Keywords Cytolytic T cells Dendritic cells DTH PSA-peptide Type-1 and type-2 cytokines Tumor immunity Vaccine Abbreviations CaP: Prostate cancer CBA: Cytokine bead array CRA: Chromium release assay DC: Dendritic cells DMSO: Dimethyl sulfoxide DTH: Delayed-type hypersensitivity PSA: Prostate-specific antigen PSA: Peptide-PSA146–154 peptide Introduction Prostate cancer (CaP) is the most common malignancy and the second leading cause of cancer-related mortality among the male population in the United States [22]. Patients who present with locally advanced or dissemi- nated metastases usually succumb with hormone- refractory disease. Most affected individuals are elderly with limited tolerance for conventional treatment regi- mens. Current therapeutic approaches for metastatic CaP are limited to a palliative role. Hence, there is a strong need for the development of novel therapeutic strategies that are effective and well tolerated. In patients with CaP, similar to other human malig- nancies, the immune system retains the potential to recognize native self-determinants associated with tumor S. Perambakam (&) S. Hallmeyer S. Reddy N. Mahmud D. Mahmud R. Nunez D. J. Peace Department of Medicine, Section of Hematology/Oncology, University of Illinois, Room 3150, Molecular Biology Research Building, 900 South Ashland Avenue, Chicago, IL 60607, USA E-mail: [email protected] L. Bressler Department of Pharmacy, University of Illinois, Chicago, IL 60607, USA P. DeChristopher R. Nunez Department of Pathology, University of Illinois, Chicago, IL 60607, USA J. A. Sosman Department of Medicine, Vanderbilt University, Nashville, TN 37232, USA Cancer Immunol Immunother (2006) 55: 1033–1042 DOI 10.1007/s00262-005-0090-x
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Induction of specific T cell immunity in patients with prostate cancer by vaccination with...

ORIGINAL ARTICLE
Supriya Perambakam Æ Sigrun Hallmeyer
Samarth Reddy Æ Nadim Mahmud Æ Linda Bressler
Phillip DeChristopher Æ Delores Mahmud
Rafael Nunez Æ Jeffrey A. Sosman Æ David J. Peace
Induction of specific T cell immunity in patients with prostate cancerby vaccination with PSA146–154 peptide
Received: 25 May 2005 / Accepted: 3 October 2005 / Published online: 10 November 2005� Springer-Verlag 2005
Abstract T cell immunotherapy of prostate cancer (CaP)offers the potential for less toxic, more effective outcomes.A clinical trial was conducted in 28 patients with locallyadvanced or metastatic CaP to determine whether anHLA-A2 binding epitope of prostate-specific antigen,PSA146–154 (PSA-peptide), can induce specific T cellimmunity. Patients were vaccinated either by intradermalinjection of PSA-peptide andGM-CSF or by intravenousadministration of autologous dendritic cells pulsed withPSA-peptide at weeks 1, 4 and 10. Delayed-type hyper-sensitivity (DTH) skin testing was performed at weeks 4,14, 26 and 52. Fifty percent of the patients developedpositive DTH responses to PSA-peptide. The size of theDTH induration progressively increased over time in themajority of responding patients. Skin biopsies from sevenDTH-positive patients were available and T cells thatdeveloped in situ were also characterized. The phenotypeof recovered T cells demonstrated variable proportions ofCD4+CD8�, CD4�CD8+ and CD4+CD8+ T cell pop-ulations. Cytokine analysis of PSA-peptide stimulated Tcells per bead array assay exhibited specific IFN-c andTNF-a response in six of seven patients. Specific IL-4
response was observed in five patients, while IL-10response was detected in one patient. PurifiedCD4�CD8+ T cells isolated from four patients demon-strated specific cytolytic activity per chromium releaseassay. In conclusion, immunization with PSA-peptideinduced specific T cell immunity in one-half of thepatients with locally advanced and hormone-sensitive,metastatic CaP. DTH-derived T cells exhibited PSA-peptide-specific cytolytic activity and predominantlyexpressed a type-1 cytokine profile.
Keywords Cytolytic T cells Æ Dendritic cells Æ DTH ÆPSA-peptide Æ Type-1 and type-2 cytokines Æ Tumorimmunity Æ Vaccine
Abbreviations CaP: Prostate cancer Æ CBA: Cytokinebead array Æ CRA: Chromium release assay Æ DC:Dendritic cells Æ DMSO: Dimethyl sulfoxide Æ DTH:Delayed-type hypersensitivity Æ PSA: Prostate-specificantigen Æ PSA: Peptide-PSA146–154 peptide
Introduction
Prostate cancer (CaP) is the most common malignancyand the second leading cause of cancer-related mortalityamong the male population in the United States [22].Patients who present with locally advanced or dissemi-nated metastases usually succumb with hormone-refractory disease. Most affected individuals are elderlywith limited tolerance for conventional treatment regi-mens. Current therapeutic approaches for metastaticCaP are limited to a palliative role. Hence, there is astrong need for the development of novel therapeuticstrategies that are effective and well tolerated.
In patients with CaP, similar to other human malig-nancies, the immune system retains the potential torecognize native self-determinants associated with tumor
S. Perambakam (&) Æ S. Hallmeyer Æ S. Reddy Æ N. MahmudD. Mahmud Æ R. Nunez Æ D. J. PeaceDepartment of Medicine, Section of Hematology/Oncology,University of Illinois, Room 3150,Molecular Biology Research Building,900 South Ashland Avenue, Chicago, IL 60607, USAE-mail: [email protected]
L. BresslerDepartment of Pharmacy, University of Illinois,Chicago, IL 60607, USA
P. DeChristopher Æ R. NunezDepartment of Pathology, University of Illinois,Chicago, IL 60607, USA
J. A. SosmanDepartment of Medicine, Vanderbilt University,Nashville, TN 37232, USA
Cancer Immunol Immunother (2006) 55: 1033–1042DOI 10.1007/s00262-005-0090-x

cells [14]. Prostate-specific antigen (PSA) is a 34 kDakallikrein-like serine protease, which is exclusively pro-duced by ductal and acinar epithelial cells of the prostategland in males [11, 24]. Several lines of evidence supportthe possibility that PSA may be a useful target antigenfor specific T-cell immunotherapy of CaP. Firstly, theexpression of PSA is highly restricted to normal andtransformed prostatic epithelial tissues. Secondly, PSA isexpressed at substantial levels by most adenocarcinomasof the prostate. Thirdly, the potential danger of devel-oping debilitating autoimmune injury to normal tissuesis limited because the prostate gland is not essential forsurvival and is often removed as part of conventionalprimary therapy.
The immunogenic potential of PSA administered as awhole-protein or expressed with DNA and RNA vectorshas been evaluated in several phase 1 clinical trials [1, 6,9, 12, 21]. These studies have shown modest immuno-logical and/or clinical responses, encouraging furtherdevelopment of immunotherapies for patients with CaP.Vaccination with defined peptide epitopes of PSA mayprove advantageous compared to methods that usewhole PSA protein. Peptide-based vaccines are expectedto elicit more focused responses, and thus, limit the risksof unexpected auto-immune injury.
Our laboratory has identified a 9-mer peptide of PSA(amino acid position 146–154, sequence KLQCVDLHV)which binds to the prevalent human leukocyte antigen,HLA-A2, and elicits specific cytotoxic T lymphocyte(CTL) responses in vitro from normal individuals andpatients with CaP [17, 26]. A phase 1b pilot trial wasconducted to determine whether vaccination with PSA146–154 peptide (hereafter called PSA-peptide) can in-duce specific T cell immunity in patients with CaP. A totalof 28 HLA-A2+ patients with high-risk, locally advanced(n=14) or metastatic, hormone-sensitive CaP (n=14)were randomly assigned to two different methods ofadministration of the PSA-peptide. One protocol entailedintradermal injection of PSA-peptide with recombinantGM-CSF. The second protocol consisted of intravenousadministration of autologous dendritic cells (DC) pulsedwith PSA-peptide. Vaccinations were performed on threeoccasions over a 10-week period. Strong delayed-typehypersensitivity (DTH) skin reactions to PSA-peptidewere elicited by both vaccination methods in fifty percentof the patients. Seven of 14 patients with positive specificDTH reaction volunteered to undergo biopsy of thereaction site. Functional analysis of DTH derived T cellsin terms of immuno-phenotyping, cytolytic activity andcytokine profile were examined.
Materials and methods
Patients
A total of 28 HLA-A2+ patients with pathologicallyconfirmed CaP were enrolled in the phase 1b clinicaltrial. All subjects were between 50 and 80 years of age at
the time of enrollment. Informed consent was obtainedfrom all patients with authorization of the InstitutionalReview Board of the University of Illinois. All patientshad undergone radio-therapy or surgical ablation of theprostate and had completed primary therapy a minimumof 6 weeks prior to enrollment in the vaccine study.Fourteen patients had high-risk, locally advanced dis-ease (group A), and fourteen patients had metastatic,hormone-sensitive disease (group B). Group A-patientshad either (1) T3, T4 disease, (2) serum PSA levelsgreater than 10 (ng/ml) or (3) a Gleason grade of at least7. Group B-patients had stage D disease following pri-mary therapy, with a declining serum PSA, and/or astable or improving bone scan or CT scan in response tohormone therapy. All patients were tested against ananergy panel of mumps, measles and candida prior tovaccination and were found to be reactive.
Cytokines and peptides
Recombinant IL-4 and GM-CSF were obtained fromSchering-Plough Research Institute (Kenilworth, NJ,USA) and Berlex Inc. (Richmond, CA, USA), respec-tively. PSA-peptide and Flu-M1 (GILGFVFTL) peptidewere synthesized under GLP conditions by MultiplePeptide Systems (San Diego, CA, USA). The synthe-sized peptides were greater than 97% pure by RP-HPLCand mass spectral analysis. The final vialized dosageform was prepared by the pharmaceutical manufactur-ing facility at the University of Iowa, College of Phar-macy.
Vaccination
Patients were randomly assigned to treatment protocol 1or protocol 2 as defined below. Protocol 1: PSA-peptide(100 lg) and GM-CSF (500 lg) mixed in 33% clinicalgrade DMSO (Edwards Lifesciences, Irvine, CA, USA)in a total volume of 1 ml were administered intrader-mally as five (0.2 ml aliquot) injections on the volaraspect of the right arm. Patients were immunized con-currently with the HLA-A2 binding Flu-M1 peptide inthe same manner on the left arm at the same concen-tration. Protocol 2: Patients underwent 7–9 l leukaphe-resis at the Blood Donor Center of the University ofIllinois. Monocyte-derived DC were cultured as per themethod of Lau et al. [10] in a clinical grade sterile lam-inar airflow hood. PBMC were obtained by centrifuga-tion over Ficoll-Hypaque (Amersham Biosciences,Uppsala, Sweden). 2·108 PBMC were cultured in T-150flasks (Corning, Big Flats, NY, USA) containing 15 mlof AIM-V medium (Life Technologies, Grand Island,NY, USA) and incubated at 37�C for 2 h. Nonadherentcells were removed by gentle flushing and media wasdecanted and replaced with 25 ml of fresh mediumcontaining IL-4 (1,000 U/ml) and GM-CSF (1,000 U/ml) and incubated for 7 days. On day 7, PSA-peptide
1034

and Flu-M1 peptide (20 lg/ml of each peptide) wereadded and incubated further for 16–18 h. On day 8,floating DC were recovered by pipetting, while adherentDC were harvested by gentle mechanical agitation. DCpopulations were pooled and washed three times withDulbecco’s PBS (BioWhittaker, Walkersville, MD,USA) and then re-suspended in 50 ml of normal salineand aseptically transferred to an infusion bag (Baxter,Irvine, CA, USA). DC were then irradiated at 2,000 radsand aseptically transferred to freezing bags (Baxter)using a 10 cc syringe with a 19-gauge needle andcryopreserved in a freezing mixture containing 30%plasmalyte, 10% DMSO and 10% human serum albu-min and stored in liquid N2. Release criteria for the finalDC product included sterile bacterial, fungal andmycoplasma cultures, negative endotoxin per LimulusAmoebacyte lysate assay, viability of at least 90% andgreater than 50% CD86, CD80, HLA-DR or CD1apositive cells and less than 10% CD14 positive cells byflow cytometric analysis. At the time of infusion, DCwere rapidly thawed at 37�C, again checked for sterilityand viability and administered intravenously to patients.Dose of DC was in the range of 0.94 to 2.02·108 cellsper vaccine (average 1.499, median 1.555). Patients werevaccinated on weeks 1, 4, and 10 and were monitored for4 h after every vaccination. One patient (UPIN88) re-ceived only a single vaccination due to fungal contami-nation of the second infusion DC product.
DTH skin testing
Immune responses were monitored by DTH skin testingon weeks 4, 14, 26, and 52 by intradermal injection of 0(carrier only), 1, 10 and 20 lg of peptide in a total volumeof 200 ll (sterile water with 33% DMSO). The FluM-1,positive control peptide, was injected on the volar aspectof the left arm, while the PSA-peptide was injected on theright arm. DTH reactions was measured at 48–72 hfollowing injection and scored as 0 (<10 mm), 1(10–14 mm), 2 (15–20 mm), or 3 (>20 mm). An indu-ration score of 2 and 3 were considered as a positivereaction. A stringent cut-off value of 14 mm was takeninto consideration for measuring true DTH skin re-sponses and to avoid false positives.
A (10·10 mm2) punch biopsy of the skin wasobtained from a strongly positive DTH site for each ofthe seven responding patients who volunteered to bebiopsied. The biopsy was divided into two parts. Onepart was used for isolation of lymphocytes as describedbelow and the second part was fixed in 10% bufferedformalin for histology and further studies.
Isolation of lymphocytes from skin biopsies of DTHreaction sites
The dermal tissue was minced into small fragments andco-cultured in RPMI-1640 medium (BioWhittaker,
Walkersville, MD, USA) containing 10% human ABserum (complete medium) and IL-2 (100 U/ml) alongwith CD3/CD28 beads (Dynal, Oslo, Norway) at aconcentration of 2.5·105 per well in 1 ml of medium in48-well plates (Nunc, Naperville, IL, USA). Out-growthof T cells was observed within 3 days. On day 4, 500 llof the spent medium was aspirated and replenished withfresh medium plus IL-2 and cultured further for a totalof 6–8 days, until the culture wells became confluent.Recovered T cells were tested directly (cycle 0) or after1–4 cycles of in vitro stimulation with irradiated autol-ogous PBMC pulsed with PSA-peptide and low-dose IL-2 (20 U/ml). Each cycle of stimulation was 7±1 days.
Cytokine bead array (CBA) analysis
Cytokines released into the culture supernatant,including IFN-c, TNF-a, IL-4, IL-6 and IL-10, weremeasured concurrently by CBA analysis (BD Bio-sciences, San Diego, CA, USA) as described earlier [18].Briefly, the antigen-presenting cell line, T2 (ATCC,Manassas, VA, USA) was used as a stimulator and waspulsed with 20 lg/ml of PSA-peptide or control HLA-A2 binding peptide, HIV-RT476-484 or diluent alone(0.4% volume by volume). T2 cells (25,000/well) werecultured with T cells (100,000/well) in complete mediumcontaining 30 U/ml of IL-2 in a total volume of 1 ml perwell in 48-well plates. This particular stimulator toresponder ratio was found to be optimal for culture in48-well plates. Cells were incubated at 37�C for 24 h in5% CO2 atmosphere. Supernatants were harvested andstored in sterile vials at –80�C. At the time of assay,samples were thawed and cytokines were measured usinga CBA kit as per the manufacturer’s protocol with aCalibure flow cytometer (Becton Dickinson, MountainView, CA, USA). Results are represented as net cytokinelevels (pg/ml) which was obtained by subtracting non-specific background responses (T2 cells pulsed withHIV-RT476-484 or diluent).
Immunophenotyping and FACS analysis
DTH-derived T cells were phenotyped for CD4 andCD8 markers. Briefly, 1·105 cells were labeled withCD4-PE and CD8-FITC and gated on side-scatter (SSC)and forward-scatter (FSC) to include lymphocyte pop-ulation. Propidium iodide (Sigma, St. Louis, MO, USA)was added (1 lg/ml, final concentration) to each sampleto exclude dead cells. All incubations were carried out at4�C for 15 min in phosphate-buffered saline containing2% normal mouse serum and 0.01% sodium azide(Sigma). The relative log fluorescence of viable cells wasmeasured at 495 nm and the percentage of CD4+ andCD8+ T cells were quantified using the Calibure.MIgG1-FITC and MIgG2a-PE served as isotype con-trols. All antibodies and isotype controls were purchasedfrom PharMingen (San Diego, CA, USA). In a separate
1035

experiment, CD4�CD8+ T cells were sorted and col-lected aseptically using a Vantage flow cytometer (Bec-ton Dickinson, Mountain View, CA, USA).
Chromium release assay (CRA)
Specific cytolytic activity was analyzed by standard 4-hCRA as previously described [26]. Briefly, 1·106 targets,SW480-PSA+, SW480-PSA� and T2 cells, a peptidetransport-deficient B-lymphoblastoid · T-lymphoblas-toid cell line were labeled with 100 lCi of Na51CrO3
(Amersham Pharmacia Biotech, Piscataway, NJ, USA).SW480-PSA+ and SW480-PSA� target cells were gen-erated as previously described [17]. Labeled T2 cells werethen pulsed with PSA-peptide or HIV-RT476-484 con-trol peptide (Research Genetics, Huntsville, AL, USA)or with diluent. Sorted CD4�CD8+ T cells (effectorcells) were plated at indicated concentrations in 96-well‘V’-bottom plates (Nunc, Naperville, IL, USA) in trip-licates and incubated along with 1·103 target cells perwell for 4 h. Supernatants recovered from CRA wereassayed for gamma emission using a Top-count NXTscintillation counter (Packard, Meriden, CT, USA), andthe percent-specific lysis was calculated as previouslydescribed [26].
Results
Specific DTH responses induced by vaccinationwith PSA-peptide
A total of 28 patients were vaccinated with PSA-peptideat three time points, i.e., weeks 1, 4 and 10 and DTHskin testing was performed on weeks 4, 14, 26 and 52.Increasing doses of peptide from 1 to 20 lg elicitedincreasing levels of DTH induration in respondingpatients (Fig. 1). FluM-1 peptide, a positive controlpeptide, showed comparable levels of DTH induration.Injection of carrier only, i.e. 33% DMSO, did not elicitsignificant induration.
Patients with both locally advanced and metastaticCaP responded to the vaccination (Table 1). Fifty per-cent of the patients (14 of 28) developed positive DTHresponses to PSA-peptide. In 13 of 14 patients withpositive DTH reactions, no response was detected at theinitial testing on week 4, and specific responses onlybecame evident at week 14 or later. Multiple cycles ofpeptide vaccination, therefore appeared to augment therecruitment of specific T cells to the DTH sites in themajority of the patients.
The size of the DTH induration to PSA-peptideprogressively increased over time in 9 of 14 respondingpatients. However, the size of the DTH indurationdecreased in four patients (UPIN28, UPIN51, UPIN53,UPIN81) while it was maintained steady in one patient(UPIN69).
The corresponding DTH responses to Flu-M1 pep-tide, a control peptide over various study time points areshown in Table 2. Thirteen of 14 patients who showedpositive DTH responses to PSA-peptide were also po-sitive for Flu-M1 peptide. On the other hand, 9 of 14patients who showed negative DTH responses to PSA-peptide were positive for Flu-M1 peptide.
Phenotype of T cells recovered from DTH sites
Skin biopsy of the DTH site was obtained from sevenpositive patients and provided the basis for the currentreport. T cells were recovered from the dermal tissue aspreviously described in ‘‘Materials and methods’’ usingthe CD3/CD28 micro-bead technique. Outgrowth of Tcells was observed within 3 days, and wells of the cultureplates became confluent by 6–8 days. Recovered T cellswere phenotyped for CD4 and CD8 markers directly(cycle 0) or after stimulation with irradiated autologousPBMC pulsed with PSA-peptide and supplemented withlow-dose IL-2 (20 U/ml) for 1–4 additional cycles.
Variable proportions of CD4+CD8�, CD4�CD8+
and CD4+CD8+ T cells were recovered per flow cyto-metric analysis. The number of CD4�CD8+ T cells wasgreater than CD4+CD8� T cells in four of seven pa-tients at cycles 0, i.e., without peptide priming in vitro(Table 3). Following in vitro stimulation with PSA-peptide, the number of CD4�CD8+ T cells becamegreater than CD4+CD8� T cells in six of seven patients.However, in one patient, UPIN45, CD4+CD8� T cellswere higher than CD4�CD8+ T cells. It is of interest tonote that a significant percentage of CD4+CD8+ T cells(11–16%) was seen in one patient, UPIN55, and thesecells were consistently observed from cycle 0 throughcycle 4 of stimulation (Table 3). Attempts were made toisolate CD4+CD8+ T cells for further analysis, butsufficient cell numbers could not be recovered.
Cytokine profile of T cells derived from DTH sites
T cells recovered from skin biopsies of DTH-positivepatients were examined to determine whether specific Tcells develop in situ. Sufficient T cells were procuredfrom one patient, UPIN55 for direct analysis at cycle 0,i.e., without peptide priming in vitro. T cells from thispatient exhibited specific IFN-c (174.4 pg/ml) and IL-4(34.5 pg/ml) cytokines per CBA analysis.
The cytokine profile following in vitro stimulationwith PSA-peptide for 1–3 cycles was studied in all theseven patients. As shown in Fig. 2, T cells derived fromsix out of seven patients showed specific IFN-c (85–1,032 pg/ml) and TNF-a (23–96 pg/ml) responses. Spe-cific IL-4 (18–486 pg/ml) response was observed in fivepatients, while IL-10 was detected in only one patient(37 pg/ml). Nonspecific expression of IL-6 was seen inone patient (data not shown).
1036

A skin biopsy of a DTH reaction to the FluM-1peptide, a positive control peptide, was also available forone patient, UPIN69. Flu-M1-peptide-specific T cells
generated IFN-c levels (1,090 pg/ml) that were compa-rable to that produced by PSA-peptide-specific T cells(1,032 pg/ml).
Fig. 1 Delayed-typehypersensitivity (DTH) skinreactions to PSA-peptidepostvaccination. Increasingdoses of peptide elicitedincreasing levels of DTHinduration in respondingpatients (a–g, left to rightindicate 1, 10 and 20 lg ofpeptide injected). FluM-1peptide, a positive controlpeptide, showed comparablelevels of DTH induration (h,left to right indicate 20, 10 and1 lg of peptide injected) as thatexhibited by PSA-peptide in thesame patient (UPIN16).Injection of carrier only, i.e.33% DMSO (200 ll), did notelicit significant induration(i, bold arrow )
1037

Specific cytotoxicity of T cells derived from DTH sites
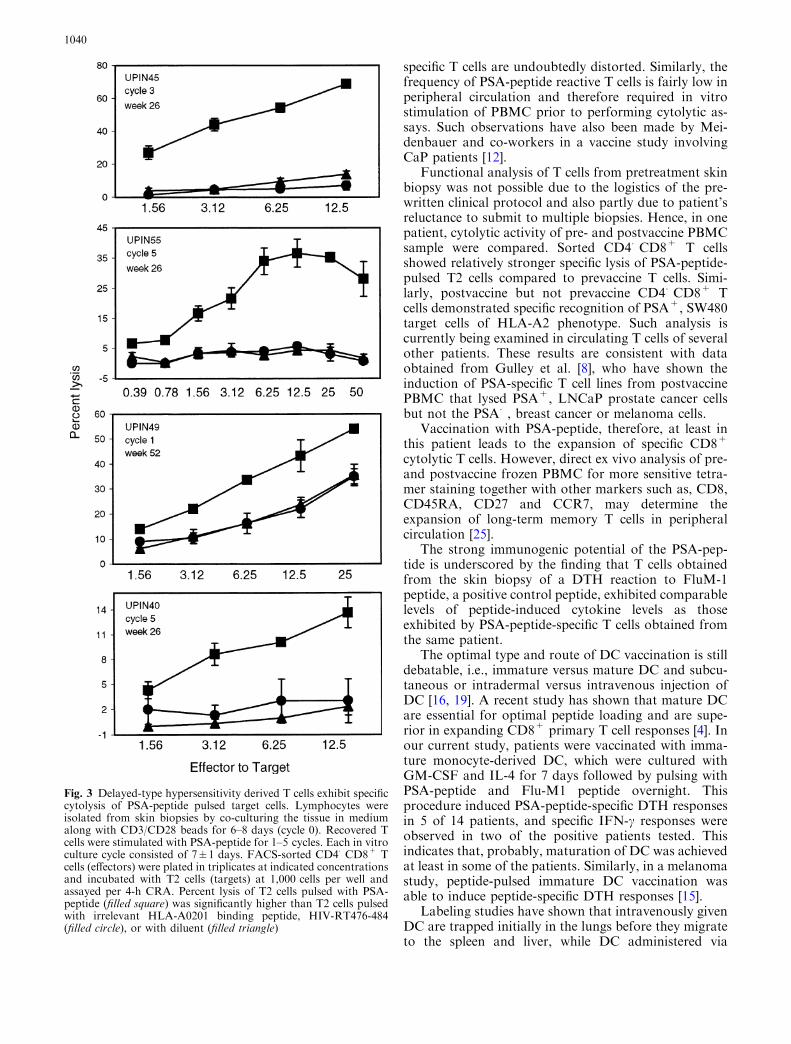
CRA were performed to determine whether T cells de-rived from positive DTH reaction sites also exhibitedspecific cytotoxicity. Following the culture with CD3/CD28 beads, recovered T cells were further stimulated invitro with irradiated autologous PBMCpulsed with PSA-peptide for 1–5 cycles. Since flow cytometric analysis re-vealed mixed phenotypic populations (see Table 3),CD4�CD8+ T cells were FACS sorted in four patients.Sorted CD4�CD8+ T cells showed peptide-specific lysisof PSA-peptide-pulsed T2 cells compared to T2 cellspulsed with irrelevant HLA-A2 binding peptide, HIV-RT476-484, or T2 cells pulsed with diluent. As shown inFig. 3, T cells induced from patients UPIN45, UPIN55,UPIN49 were strongly cytolytic, while T cells from pa-tient UPIN40 demonstrated modest cytolytic activity.
In patient UPIN55, a comparison of cytolytic activityof prevaccine (Week 0) and postvaccine (Week 26)
PBMC-derived T cells was also performed. PBMCsamples were stimulated with PSA-peptide in vitro for5 cycles before proceeding to CRA. Sorted postvaccineCD4�CD8+ T cells (upper panel) showed strongerspecific lysis of PSA-peptide-pulsed T2 cells, especially atlower effector to target ratios compared to prevaccine Tcells (lower panel). Similarly, postvaccine, but not pre-vaccine CD4�CD8+ T cells, demonstrated specific rec-ognition of PSA+, SW480 target cells of the HLA-A2phenotype (Fig. 4).
Discussion
A pilot clinical trial involving a previously identifiedHLA-A2 restricted epitope of PSA was conducted in 28
Table 2 Delayed-type hypersensitivity skin reaction to Flu-M1, acontrol peptide, over various study time points
Patient Protocol Group DTH induration at weeks
4 14 26 52
UPIN13 1 B 0 2 2 2UPIN16 1 A 0 2 3 3UPIN28 1 B 0 1 2 0UPIN40 1 B 1 3 3 3UPIN45 1 A 1 2 0 2UPIN49 1 B 0 2 2 1UPIN50 1 A 1 2 3 3UPIN51 1 A 0 1 0 0UPIN53 2 A 2 1 0 1UPIN55 1 A 1 2 3 1UPIN69 2 B 1 a2 1 1UPIN71 2 A 3 3 1 3UPIN81 2 B 0 3 3 0UPIN88 2 A 1 0 2 2
UPIN2 2 B ND 3 2 0UPIN21 2 A 0 2 2 1UPIN26 2 A 0 1 2 1UPIN27 2 B 0 2 2 NDUPIN32 1 A 1 0 0 0UPIN35 2 B 2 3 2 1UPIN37 1 B 0 0 0 0UPIN38 1 A 0 1 0 0UPIN43 2 A 1 0 0 0UPIN67 1 B 2 1 0 0UPIN70 2 A 0 0 0 1UPIN82 2 B 0 1 2 1UPIN85 1 B 0 2 0 2UPIN89 2 B 0 3 3 3
DTH reactions obtained at the highest dose of Flu-M1 peptide(20 lg) were measured and scored as 0 (<10 mm), 1 (10–14 mm), 2(15–20 mm), 3 (>20 mm). Indurations of 2 and 3 were consideredpositive reactions. ND not done. UPIN2 did not show up for week4, UPIN27 was lost to f/u after week 26. aDTH skin biopsy wastaken at this study time point. Overall, 78.6% of the patients (22 of28) developed positive DTH responses to Flu-M1 peptide. Upperpanel represents patients who developed positive DTH responses toPSA-peptide and lower panel represents patients who showednegative DTH reaction to PSA-peptide (see also Table 1). Group Arepresents patients with high risk, locally advanced disease andgroup B represents patients with hormone-sensitive metastaticdisease
Table 1 Delayed-type hypersensitivity skin reaction to PSA-peptideover various study time points
Patient Protocol Group DTH induration at weeks
4 14 26 52
UPIN13 1 B 0 0 1 2UPIN16 1 A 0 2 a3 3UPIN28 1 B 0 0 3 2UPIN40 1 B 0 2 a2 3UPIN45 1 A 0 1 a2 3UPIN49 1 B 0 0 0 a2UPIN50 1 A 1 1 3 3UPIN51 1 A 0 2 0 0UPIN53 2 A 0 3 3 0UPIN55 1 A 1 1 a2 3UPIN69 2 B 3 a3 3 3UPIN71 2 A 1 a2 2 3UPIN81 2 B 0 2 0 0UPIN88 2 A 0 0 2 2
UPIN2 2 B ND 1 0 0UPIN21 2 A 0 0 0 1UPIN26 2 A 0 0 0 0UPIN27 2 B 0 1 1 NDUPIN32 1 A 0 1 0 0UPIN35 2 B 0 0 0 0UPIN37 1 B 0 0 0 1UPIN38 1 A 0 1 0 0UPIN43 2 A 0 0 0 0UPIN67 1 B 0 0 0 0UPIN70 2 A 0 0 0 1UPIN82 2 B 0 0 0 1UPIN85 1 B 0 1 0 0UPIN89 2 B 0 1 1 0
DTH reactions obtained at the highest dose of PSA-peptide (20 lg)were measured and scored as 0 (<10 mm), 1 (10–14 mm), 2 (15–20 mm), 3 (>20 mm). Indurations of 2 and 3 were consideredpositive reactions. ND not done. UPIN2 did not show up for week4, UPIN27 was lost to f/u after week 26. aDTH skin biopsy wastaken at these study time points. Fifty percent of the patients (14 of28) developed positive DTH responses (upper panel) to PSA-pep-tide that progressively increased over time in majority (9 of 14) ofthe responding patients. Group A represents patients with highrisk, locally advanced disease and group B represents patients withhormone-sensitive metastatic disease
1038

patients with high risk, localized (group A) or hormone-sensitive metastatic CaP (group B). Patients were vac-cinated on three occasions over a 10-week period byintradermal injection of PSA-peptide plus GM-CSF orby intravenous administration of autologous DC pulsedwith peptide. Both methods of vaccination were immu-nogenic. Strong DTH skin reactions to the PSA-peptidebecame detectable in 50% (14 of 28) of patients overtime. While DTH responses to Flu-M1 peptide, a posi-tive control peptide, was observed in 78.6% (22 of 28) ofthe patients.
DTH reactions to immunizing peptides have beenobserved in other peptide-vaccination protocols, par-ticularly melanoma vaccine studies, and are often usedas an indicator of antitumor immunity and vaccineefficacy [5, 15, 23]. In the current study, 13 of 14 patientswith positive DTH reactions showed no response wheninitially tested at week 4, and specific responses becameevident at week 14 or later. The size of the DTH indu-ration progressively increased over time in the majority(9 of 14) of the responding patients. These resultstherefore indicate the induction of specific T cellimmunity in CaP patients post vaccination.
Although, injection of PSA-peptide for DTH skintesting might immunologically resemble another roundof vaccination, low doses (1–20 lg) of naked peptidealone were unlikely to have a confounding impact on thevaccine outcome. This argument is further supported bythe observation that, in 4 of 14 responding patients, thesize of DTH induration to PSA-peptide either decreasedor positive DTH responses that were initially seen totallydisappeared by week 52 (Table 1).
Direct recovery of large number of T cells from smallclinical specimens, such as punch biopsy of the skin, isoften very difficult with traditional culture techniquesinvolving only IL-2. In the current study, we, therefore,cultured dermal tissues in the presence of CD3/CD28micro-beads and low-dose IL-2 for 6–8 days. T cellswere tested directly or following further stimulation withPSA-peptide in vitro. The expanded T cells exhibitedspecificity to PSA-peptide per CRA and CBA analysis,presumably reflecting corresponding responder T cellsthat prevail in vivo although the actual frequency of
Fig. 2 Cytokine profile of T cells derived from DTH sites.Lymphocytes were isolated from skin biopsies by co-culturing thetissue inmedium alongwith CD3/CD28 beads for 6–8 days (cycle 0).Additionally, T cells were stimulated with PSA-peptide for1–3 cycles. Each in vitro culture cycle consisted of 7±1 days. Tcells at 100,000 cells per well were incubated with T2 cells pulsed withPSA-peptide or HIV-RT476-484 or with diluent. Specific cytokinesthat secreted into the medium were assayed by CBA, and the netcytokine levels (pg/ml) after subtracting backgrounds are shown
Table 3 Phenotype of DTH-derived T-cells
Patient Cycle of culture % Positive cells
CD4+CD8� CD4�CD8+ CD4+CD8+
UPIN16 0 32 62 83 20 75 3
UPIN40 0 39 48 102 21 69 6
UPIN45 0 78 12 73 56 26 7
UPIN49 0 46 27 62 3 80 4
UPIN55 0 50 29 132 9 59 164 12 65 11
UPIN 69 0 19 53 2UPIN71 0 39 46 6
3 14 69 2
Skin biopsies from seven positive patients were available for invitro analysis. Lymphocytes were isolated from biopsies by co-culturing the tissue in medium along with CD3/CD28 beads for 6–8 days (cycle 0). Additionally, T-cells were stimulated with PSA-peptide pulsed irradiated autologous PBMC for 1–4 cycles. Each invitro culture cycle consisted of 7±1 days. 1·105 cells were labeledwith CD4-PE and CD8-FITC and gated on SSC and FSC to in-clude lymphocyte population. Percent positive cells were quantifiedby two-color staining as described in the Methods section
1039
specific T cells are undoubtedly distorted. Similarly, thefrequency of PSA-peptide reactive T cells is fairly low inperipheral circulation and therefore required in vitrostimulation of PBMC prior to performing cytolytic as-says. Such observations have also been made by Mei-denbauer and co-workers in a vaccine study involvingCaP patients [12].
Functional analysis of T cells from pretreatment skinbiopsy was not possible due to the logistics of the pre-written clinical protocol and also partly due to patient’sreluctance to submit to multiple biopsies. Hence, in onepatient, cytolytic activity of pre- and postvaccine PBMCsample were compared. Sorted CD4�CD8+ T cellsshowed relatively stronger specific lysis of PSA-peptide-pulsed T2 cells compared to prevaccine T cells. Simi-larly, postvaccine but not prevaccine CD4�CD8+ Tcells demonstrated specific recognition of PSA+, SW480target cells of HLA-A2 phenotype. Such analysis iscurrently being examined in circulating T cells of severalother patients. These results are consistent with dataobtained from Gulley et al. [8], who have shown theinduction of PSA-specific T cell lines from postvaccinePBMC that lysed PSA+, LNCaP prostate cancer cellsbut not the PSA�, breast cancer or melanoma cells.
Vaccination with PSA-peptide, therefore, at least inthis patient leads to the expansion of specific CD8+
cytolytic T cells. However, direct ex vivo analysis of pre-and postvaccine frozen PBMC for more sensitive tetra-mer staining together with other markers such as, CD8,CD45RA, CD27 and CCR7, may determine theexpansion of long-term memory T cells in peripheralcirculation [25].
The strong immunogenic potential of the PSA-pep-tide is underscored by the finding that T cells obtainedfrom the skin biopsy of a DTH reaction to FluM-1peptide, a positive control peptide, exhibited comparablelevels of peptide-induced cytokine levels as thoseexhibited by PSA-peptide-specific T cells obtained fromthe same patient.
The optimal type and route of DC vaccination is stilldebatable, i.e., immature versus mature DC and subcu-taneous or intradermal versus intravenous injection ofDC [16, 19]. A recent study has shown that mature DCare essential for optimal peptide loading and are supe-rior in expanding CD8+ primary T cell responses [4]. Inour current study, patients were vaccinated with imma-ture monocyte-derived DC, which were cultured withGM-CSF and IL-4 for 7 days followed by pulsing withPSA-peptide and Flu-M1 peptide overnight. Thisprocedure induced PSA-peptide-specific DTH responsesin 5 of 14 patients, and specific IFN-c responses wereobserved in two of the positive patients tested. Thisindicates that, probably, maturation of DC was achievedat least in some of the patients. Similarly, in a melanomastudy, peptide-pulsed immature DC vaccination wasable to induce peptide-specific DTH responses [15].
Labeling studies have shown that intravenously givenDC are trapped initially in the lungs before they migrateto the spleen and liver, while DC administered via
Fig. 3 Delayed-type hypersensitivity derived T cells exhibit specificcytolysis of PSA-peptide pulsed target cells. Lymphocytes wereisolated from skin biopsies by co-culturing the tissue in mediumalong with CD3/CD28 beads for 6–8 days (cycle 0). Recovered Tcells were stimulated with PSA-peptide for 1–5 cycles. Each in vitroculture cycle consisted of 7±1 days. FACS-sorted CD4�CD8+ Tcells (effectors) were plated in triplicates at indicated concentrationsand incubated with T2 cells (targets) at 1,000 cells per well andassayed per 4-h CRA. Percent lysis of T2 cells pulsed with PSA-peptide (filled square) was significantly higher than T2 cells pulsedwith irrelevant HLA-A0201 binding peptide, HIV-RT476-484(filled circle), or with diluent (filled triangle)
1040

intradermal or intranodal injection effectively migrate todraining lymph nodes [3, 13]. However, previous vaccinetrials administered by intravenous route have shownimmunological and clinical responses, and hence weopted for this method [19]. As observed in the currentstudy, 64% of the patients vaccinated by subcutaneousinjection as opposed to 36% vaccinated via intravenousDC arm developed specific DTH responses to PSA-peptide. Therefore, we feel that vaccination by subcu-taneous method is more effective.
Interestingly, the method of vaccination also ap-peared to impact the pattern of cytokine responses.DTH-derived T cells from two patients who exhibitedonly type-1 responses (IFN-c and TNF-a) were vacci-nated by protocol 2. On the other hand, five out of sevenpatients who exhibited type-1 and type-2 cytokine re-sponses (Fig. 2) were vaccinated by protocol 1. Al-though, the study population is small, previous studieshave shown that (1) route of vaccine administration, (2)method of antigen loading and (3) the type of adjuvantadministered influences the polarization and efficacy ofantitumor T cell responses [2, 7, 20].
In conclusion, our results indicate that vaccinationwith soluble peptide or DC-bound peptide elicits strongspecific T cell immunity to the PSA-peptide in one-half
of patients with locally advanced or hormone-sensitivemetastatic CaP. The determinants that distinguishresponding versus nonresponding subjects are currentlybeing examined. The impact of successful immunizationon disease course will be examined in an expanded phase2 study.
Acknowledgements This work was supported by the NationalCancer Institute (CA88062), the Department of Army (DAMD17-98-1-8489), the American Cancer Society (IRG 99-224-01), theIllinois Department of Public Health (prostate cancer research fund4328301), the Cancer Research Institute, New York City, and theGeneral Clinical Research Center of University of Illinois, Chicago,funded by NIH grant M01-RR-13987. The contents in the manu-script are solely the responsibility of the authors and do not nec-essarily reflect the official views of the respective funding agencies.The authors would like to thank Susan Nishimura and SusanCascio for their excellent clinical assistance.
References
1. Barrou B, Benoit G, Ouldkaci M, Cussenot O, Salcedo M,Agrawal S, Massicard S, Bercovici N, Ericson ML, Thiounn N(2004) Vaccination of prostatectomized prostate cancer pa-tients in biochemical relapse, with autologous dendritic cellspulsed with recombinant human PSA. Cancer Immunol Im-munother 53:453–460
Fig. 4 Specific recognition ofPSA+ and PSA-peptide pulsedtargets of HLA-A2 phenotypeby PBMC-derived T cells. Pre-and postvaccine PBMC(UPIN55) were stimulated invitro with PSA-peptide for 1–5 cycles. Each culture cycleconsisted of 7±1 days. FACS-sorted CD4�CD8+ T cells(effectors) were plated intriplicates at indicatedconcentrations and incubatedwith 51Cr-labeled targets,SW480-PSA+ (filled daimond),SW480-PSA� (open diamond)or T2 cells pulsed with PSA-peptide (filled square), or pulsedwith irrelevant HLA-A0201binding peptide, HIV-RT476-484 (filled circle), or withdiluent (filled triangle) at1,000 cells per well and assayedper 4-h CRA. PostvaccineCD4�CD8+ T cells (upperpanel) showed relativelystronger specific lysis of PSA-peptide pulsed T2 cellscompared to prevaccine T cells(lower panel). Similarly,postvaccine, but not prevaccineCD4�CD8+ T cells,demonstrated specificrecognition of PSA+, SW480target cells of HLA-A2phenotype
1041

2. Constant SL, Bottomly K (1997) Induction of Th1 and Th2CD4+ T cell responses: the alternative approaches. Annu RevImmunol 15:297–322
3. De Vries IJ, Krooshoop DJ, Scharenborg NM, Lesterhuis WJ,Diepstra JH, Van Muijen GN, Strijk SP, Ruers TJ, BoermanOC, Oyen WJ, Adema GJ, Punt CJ, Figdor CG (2003) Effectivemigration of antigen-pulsed dendritic cells to lymph nodes inmelanoma patients is determined by their maturation state.Cancer Res 63:12–17
4. Dieckmann D, Schultz ES, Ring B, Chames P, Held G, Hoo-genboom HR, Schuler G (2005) Optimizing the exogenousantigen loading of monocyte-derived dendritic cells. IntImmunol 17:621–635
5. Disis ML, Schiffman K, Gooley TA, McNeel DG, Rinn K,Knutson KL (2000) Delayed-type hypersensitivity response is apredictor of peripheral blood T-cell immunity after HER-2/neupeptide immunization. Clin Cancer Res 6:1347–1350
6. Eder JP, Kantoff PW, Roper K, Xu GX, Bubley GJ, Boyden J,Gritz L, Mazzara G, Oh WK, Arlen P, Tsang KY, Panicali D,Schlom J, Kufe DW (2000) A phase I trial of a recombinantvaccinia virus expressing prostate-specific antigen in advancedprostate cancer. Clin Cancer Res 6:1632–1638
7. Galea-Lauri J, Wells JW, Darling D, Harrison P, Farzaneh F(2004) Strategies for antigen choice and priming of dendriticcells influence the polarization and efficacy of antitumor T-cellresponses in dendritic cell-based cancer vaccination. CancerImmunol Immunother 53:963–977
8. Gulley JL, Arlen PM, Bastian A, Morin S, Marte J, Beetham P,Tsang KY, Yokokawa J, Hodge JW, Menard C, CamphausenK, Coleman CN, Sullivan F, Steinberg SM, Schlom J, DahutW (2005) Combining a recombinant cancer vaccine with stan-dard definitive radiotherapy in patients with localized prostatecancer. Clin Cancer Res 11:3353–3362
9. Heiser A, Coleman D, Dannull J, Yancey D, Maurice MA,Lallas CD, Dahm P, Niedzwiecki D, Gilboa E, Vieweg J (2002)Autologous dendritic cells transfected with prostate-specificantigen RNA stimulate CTL responses against metastaticprostate tumors. J Clin Invest 109:409–417
10. Lau R, Wang F, Jeffery G, Marty V, Kuniyoshi J, Bade E,Ryback ME, Weber J (2001) Phase I trial of intravenous pep-tide-pulsed dendritic cells in patients with metastatic mela-noma. J Immunother 24:66–78
11. Li TS, Beling CG (1973) Isolation and characterization of twospecific antigens of human seminal plasma. Fertil Steril 24:134–144
12. Meidenbauer N, Harris DT, Spitler LE, Whiteside TL (2000)Generation of PSA-reactive effector cells after vaccination witha PSA-based vaccine in patients with prostate cancer. Prostate43:88–100
13. Morse MA, Coleman RE, Akabani G, Niehaus N, Coleman D,Lyerly HK (1999) Migration of human dendritic cells afterinjection in patients with metastatic malignancies. Cancer Res59:56–58
14. Nair SK, Hull S, Coleman D, Gilboa E, Lyerly HK, MorseMA (1999) Induction of carcinoembryonic antigen (CEA)-specific cytotoxic T-lymphocyte responses in vitro usingautologous dendritic cells loaded with CEA peptide or CEARNA in patients with metastatic malignancies expressing CEA.Int J Cancer 82:121–124
15. Nestle FO, Alijagic S, Gilliet M, Sun Y, Grabbe S, Dummer R,Burg G, Schadendorf D (1998) Vaccination of melanoma pa-tients with peptide- or tumor lysate-pulsed dendritic cells. NatMed 4:328–332
16. Nestle FO, Farkas A, Conrad C (2005) Dendritic-cell-basedtherapeutic vaccination against cancer. Curr Opin Immunol17:163–169
17. Perambakam S., Xue B-H, Sosman JA, Peace DJ (2002)Induction of Tc2 cells with specificity for prostate-specificantigen from patients with hormone-refractory prostate cancer.Cancer Immunol Immunother 51:263–270
18. Perambakam SM, Srivastava R, Peace DJ (2005) Distinctcytokine patterns exist in peripheral blood mononuclear cellcultures of patients with prostate cancer. Clin Immunol 117:94–99
19. Reichardt VL, Brossart P, Kanz L (2004) Dendritic cells invaccination therapies of human malignant disease. Blood Rev18:235–243
20. Romagnani S (1997) The Th1/Th2 paradigm. Immunol Today18:263–266
21. Sanda MG, Smith DC, Charles LG, Hwang C, Pienta KJ,Schlom J, Milenic D, Panicali D, Montie JE (1999) Re-combinant vaccinia-PSA (PROSTVAC) can induce a prostate-specific immune response in androgen-modulated humanprostate cancer. Urology 53:260–266
22. Stewart SL, King JB, Thompson TD, Friedman C, Wingo PA(2004) Cancer mortality surveillance—United States, 1990–2000. MMWR Surveill Summ 53:1–108
23. Waanders GA, Rimoldi D, Lienard D, Carrel S, Lejeune F,Dietrich PY, Cerottini JC, Romero P (1997) Melanoma-reac-tive human cytotoxic T lymphocytes derived from skin biopsiesof delayed-type hypersensitivity reactions induced by injectionof an autologous melanoma cell line. Clin Cancer Res 3:685–696
24. Watt KW, Lee PJ, M’Timkulu T, Chan WP, Loor R (1986)Human prostate-specific antigen: structural and functionalsimilarity with serine proteases. Proc Natl Acad Sci USA83:3166–3170
25. Walker EB, HaleyD,MillerW, FloydK,Wisner KP, SanjuanN,Maecker H, Romero P, Hu HM, Alvord WG, Smith JW II, FoxBA, Urba WJ (2004) Gp100(209-2 M) peptide immunization ofhuman lymphocyte antigen-A2+ stage I-III melanoma patientsinduces significant increase in antigen-specific effector and long-term memory CD8+ T cells. Clin Cancer Res 10:668–680
26. Xue B-H, Zhang Y, Sosman JA, Peace DJ (1997) Induction ofhuman cytotoxic T lymphocytes specific for prostate-specificantigen. Prostate 30:73–78
1042




















