
In vitro kinetic studies on the mechanism of oxygen-dependent cellular uptake of copper...
17
IOP PUBLISHING PHYSICS IN MEDICINE AND BIOLOGY Phys. Med. Biol. 54 (2009) 2103–2119 doi:10.1088/0031-9155/54/7/017 In vitro kinetic studies on the mechanism of oxygen-dependent cellular uptake of copper radiopharmaceuticals Jason P Holland 1,3,4 , Jeffrey H Giansiracusa 2 , Stephen G Bell 1 , Luet-Lok Wong 1 and Jonathan R Dilworth 1 1 Department of Chemistry, University of Oxford, Chemistry Research Laboratory, 12 Mansfield Road, Oxford,OX1 3TA, UK 2 Department of Mathematics, Mathematical Institute, University of Oxford, 24-29 St Giles’, Oxford, OX1 3LB, UK E-mail: [email protected] and [email protected] Received 2 December 2008, in final form 22 January 2009 Published 12 March 2009 Online at stacks.iop.org/PMB/54/2103 Abstract The development of hypoxia-selective radiopharmaceuticals for use as therapeutic and/or imaging agents is of vital importance for both early identification and treatment of cancer and in the design of new drugs. Radiotracers based on copper for use in positron emission tomography have received great attention due to the successful application of copper(II) bis(thiosemicarbazonato) complexes, such as [ 60/62/64 Cu(II)ATSM] and [ 60/62/64 Cu(II)PTSM], as markers for tumour hypoxia and blood perfusion, respectively. Recent work has led to the proposal of a revised mechanism of hypoxia-selective cellular uptake and retention of [Cu(II)ATSM]. The work presented here describes non-steady-state kinetic simulations in which the reported pO 2 -dependent in vitro cellular uptake and retention of [ 64 Cu(II)ATSM] in EMT6 murine carcinoma cells has been modelled by using the revised mechanistic scheme. Non-steady-state (NSS) kinetic analysis reveals that the model is in very good agreement with the reported experimental data with a root-mean-squared error of less than 6% between the simulated and experimental cellular uptake profiles. Estimated rate constants are derived for the cellular uptake and washout (k 1 = 9.8 ± 0.59 × 10 −4 s −1 and k 2 = 2.9 ± 0.17 × 10 −3 s −1 ), intracellular reduction (k 3 = 5.2 ± 0.31 × 10 −2 s −1 ), reoxidation (k 4 = 2.2 ± 0.13 mol −1 dm 3 s −1 ) and proton-mediated ligand dissociation (k 5 = 9.0 ± 0.54 × 10 −5 s −1 ). Previous mechanisms focused on the reduction and reoxidation steps. However, the data suggest that the origins of hypoxia-selective retention may reside with the stability of the copper(I) anion with respect to protonation and ligand dissociation. In vitro 3 Present address: Memorial Sloan-Kettering Cancer Center, 1275 York Avenue, NY 100 65, USA. 4 Author to whom any correspondence should be addressed. 0031-9155/09/072103+17$30.00 © 2009 Institute of Physics and Engineering in Medicine Printed in the UK 2103
Transcript of In vitro kinetic studies on the mechanism of oxygen-dependent cellular uptake of copper...

IOP PUBLISHING PHYSICS IN MEDICINE AND BIOLOGY
Phys. Med. Biol. 54 (2009) 2103–2119 doi:10.1088/0031-9155/54/7/017
In vitro kinetic studies on the mechanism ofoxygen-dependent cellular uptake of copperradiopharmaceuticals
Jason P Holland1,3,4, Jeffrey H Giansiracusa2, Stephen G Bell1,
Luet-Lok Wong1 and Jonathan R Dilworth1
1 Department of Chemistry, University of Oxford, Chemistry Research Laboratory,12 Mansfield Road, Oxford,OX1 3TA, UK2 Department of Mathematics, Mathematical Institute, University of Oxford, 24-29 St Giles’,Oxford, OX1 3LB, UK
E-mail: [email protected] and [email protected]
Received 2 December 2008, in final form 22 January 2009Published 12 March 2009Online at stacks.iop.org/PMB/54/2103
Abstract
The development of hypoxia-selective radiopharmaceuticals for use astherapeutic and/or imaging agents is of vital importance for both earlyidentification and treatment of cancer and in the design of new drugs.Radiotracers based on copper for use in positron emission tomographyhave received great attention due to the successful application of copper(II)bis(thiosemicarbazonato) complexes, such as [60/62/64Cu(II)ATSM] and[60/62/64Cu(II)PTSM], as markers for tumour hypoxia and blood perfusion,respectively. Recent work has led to the proposal of a revised mechanismof hypoxia-selective cellular uptake and retention of [Cu(II)ATSM]. Thework presented here describes non-steady-state kinetic simulations in whichthe reported pO2-dependent in vitro cellular uptake and retention of[64Cu(II)ATSM] in EMT6 murine carcinoma cells has been modelled by usingthe revised mechanistic scheme. Non-steady-state (NSS) kinetic analysisreveals that the model is in very good agreement with the reported experimentaldata with a root-mean-squared error of less than 6% between the simulatedand experimental cellular uptake profiles. Estimated rate constants are derivedfor the cellular uptake and washout (k1 = 9.8 ± 0.59 × 10−4 s−1 and k2 =2.9 ± 0.17 × 10−3 s−1), intracellular reduction (k3 = 5.2 ± 0.31 × 10−2 s−1),reoxidation (k4 = 2.2 ± 0.13 mol−1 dm3 s−1) and proton-mediated liganddissociation (k5 = 9.0 ± 0.54 × 10−5 s−1). Previous mechanisms focusedon the reduction and reoxidation steps. However, the data suggest that theorigins of hypoxia-selective retention may reside with the stability of thecopper(I) anion with respect to protonation and ligand dissociation. In vitro
3 Present address: Memorial Sloan-Kettering Cancer Center, 1275 York Avenue, NY 100 65, USA.4 Author to whom any correspondence should be addressed.
0031-9155/09/072103+17$30.00 © 2009 Institute of Physics and Engineering in Medicine Printed in the UK 2103

2104 J P Holland et al
kinetic studies using the nicotimamide adenine dinucleotide (NADH)-dependent ferredoxin reductase enzyme PuR isolated from the bacteriumRhodopseudomonas palustris have also been conducted. NADH turnoverfrequencies are found to be dependent on the structure of the ligand andthe results confirm that the proposed reduction step in the mechanism ofhypoxia selectivity is likely to be mediated by NADH-dependent enzymes.Further understanding of the mechanism of hypoxia selectivity may facilitatethe development of new imaging and radiotherapeutic agents with increasedspecificity for tumour hypoxia.
(Some figures in this article are in colour only in the electronic version)
1. Introduction
The transition of a solid tumour from a benign to a more malignant phenotype is associatedwith fundamental changes in the tumour microenvironment and tissue physiology. Phenotypicchanges often include irregular growth in tumour vasculature via stimulation of angiogenesisand vasculogenesis, decreased tissue perfusion, altered gene expression, tumour acidosis(lowered extracellular tissue acidity, pHe) and tumour hypoxia (reduced oxygen tension,pO2/mmHg). Bunn and Poyton have reviewed the biochemical pathways of oxygen sensingin both single-cell and multi-cellular organisms (Bunn and Poyton 1996).
The presence of tumour hypoxia represents a major problem for conventional cancertherapies including both radiation therapy and chemotherapy. Oxygen is the most electronaffinic molecule found within cells. Radiation therapy relies on the presence of molecularoxygen to ‘fix’ the damage caused by ionizing radiation. Radiotherapy induces DNA-basedradical formation and subsequent electron transfer to oxygen leads to the generation of highlyreactive oxygen species (ROS) including superoxide and hydroxyl radicals which resultin permanent damage to the cell. In the absence of oxygen, cellular defence and repairmechanisms can restore the damaged DNA by hydrogen atom donation from non-proteinsulfhydryls. Chemotherapeutic resistance correlates with tumour hypoxia and develops dueto changes in both tumour physiology and tissue geometry. Hypoxia induces specific geneexpression which produces proteins that cause a decrease in the rate of progression throughthe normal cell cycle (Heacock and Sutherland 1986). As most anticancer drugs target rapidlyproliferating cells, the decreasing rate of cell cycle progression with increasing distance fromtumour vasculature causes decreased cell kill in hypoxic tissue. In addition, due to the limitedrange of diffusion of most anticancer drugs from capillaries (a consequence of their highreactivity and molecular weight), the proximity of hypoxic tissue from tumour vasculature(typically >100 μm) confers a passive resistance to chemotherapy. Hypoxia also confersresistance to drugs such as cisplatin through expression of various hypoxic stress proteins,(Murphy et al 1994, Kelley et al 1988, Kelley and Rozencweig 1989, Satoh et al 1993) and hasbeen linked to increased malignant progression and metastasis. Therefore, early identificationof tumour hypoxia and understanding the physiological consequences are essential both inanticancer drug discovery and in the effective clinical management of cancer treatment (Brownand Wilson 2004).
Tumour oxygen levels can be measured directly using a polarographic oxygen electrode(Eppendorf) mounted on a fine needle (Parker et al 2004, Nordsmark et al 2001, 2003,Bentzen et al 2003). However, the technique is invasive, subject to sampling errors, is only

Kinetics studies on Cu-ATSM 2105
Scheme 1. Schematic representation of the first mechanism of hypoxia-selective uptake andretention proposed by Fujibayashi et al (1997) and supported by the work of Obata et al (2001).
suitable for accessible tumours and has not yet been approved for clinical use (Dehdashti et al2003). Functional molecular imaging is an alternative, non-invasive method for locating andquantifying tumour hypoxia in vivo. In particular, positron emission tomography (PET)offers the advantage of correlating radiopharmaceutical uptake with tumour physiologyand has the potential to be used for quantitative delineation of tumour hypoxia. A rangeof hypoxia-selective radiopharmaceuticals have been developed and the most promisingcompounds include [18 F]-fluoromisonidazole ([18 F]-FMISO)(Couturier et al 2004) and copperradionuclide complexes of the diacetyl-2,3-bis(4-N-methyl-3-thiosemicarbazonato) ligand,[∗Cu(II)ATSM] (where ‘∗’ represents a positron emitting radionuclide of copper including,60Cu, 61Cu, 62Cu or 64Cu) (Vavere and Lewis, 2007).
Hypoxia-selective uptake of [62Cu(II)ATSM] was first reported in 1997 (Fujibayashiet al 1997). Over recent years, several research groups have investigated the origins ofhypoxia selectivity of copper bis(thiosemicarbazonato) complexes. The first mechanism wasproposed by Fujibayashi et al and supported by the work of Obata et al (scheme 1) (Fujibayashiet al 1997, Obata et al 2001). Experiments showed that [62Cu(II)ATSM] accumulates inhypoxic myocardium via a bioreductive retention mechanism involving nicotinamide adeninedinucleotide (NADH)-dependent enzymes of the electron transport chain (ETC) found inmitochondria (Fujibayashi et al 1993, 1997). It was suggested that upon intracellular reduction[Cu(II)ATSM] (half-wave potential, E1/2 = −0.646 V versus the saturated calomel referenceelectrode)(Holland et al 2008b) becomes trapped irreversibly. Reduction of [Cu(II)ATSM]only occurs in hypoxic cells and involves electron transfer from hyper-reduced complex I(ubiquinone oxidoreductase) using NADH as a two-electron donor. In normoxic cells, complexI is incapable of reducing [Cu(II)ATSM]. In contrast, the non-hypoxia-selective copper(II)complex of pyruvaldehyde bis(4-N-methyl-3-thiosemicarbazonato) ligand, [Cu(II)PTSM], hasa less negative one-electron reduction potential (E1/2(SCE) = −0.565 V) and may be reducedin all cells by complex I in its normal state (i.e., not hyper reduced), leading to irreversibleintracellular trapping. In this mechanism, pO2-dependence of one-electron reduction is thediscriminating factor which controls the reversibility of cellular uptake.

2106 J P Holland et al
Scheme 2. Schematic representation of the revised mechanism of hypoxia selectivity of[Cu(II)ATSM] (Holland et al 2008b). Copper complexes within the outer and inner boxes representintracellular species which are reversibly and irreversibly trapped within cells, respectively. Therate constants are discussed in the main text and correspond to cellular uptake (k1), washout (k2),enzyme-mediated reduction (k3), reoxidation by molecular oxygen (k4), protonation/deprotonation(k5 and k6) and irreversible ligand dissociation (kirr).
In 2001, Obata et al (2001) demonstrated that in subcellular fractions of Erhlichascites tumour cells, [Cu(II)ATSM] reduction was mediated by enzymes located in themicrosome/cytosol fraction, rather than the mitochondria. Reduction was heat sensitiveand was enhanced when the culture medium concentration of either reduced nicotinamideadenine dinucleotide (NADH) or nicotinamide adenine dinucleotide phosphate (NADPH) wasincreased. Control experiments showed that neither NADH nor NADPH, are capable ofreducing [Cu(II)ATSM] alone. In addition, specific reductase inhibitor studies showed thatinhibition NADH-dependent cytochrome b5 reductase and NADPH-dependent cytochromeP450 reductase microsomal enzymes, by phenylthiourea (PTU) and adenosine monophosphate(AMP), respectively, caused a 25–50% decrease in [Cu(II)ATSM] reduction, compared withcontrol experiments.
Subsequent reports by Dearling et al (1998, 2002) and Maurer et al (2002) noted thatthis first mechanism is inconsistent with cellular uptake and washout studies (Lewis et al1999). They proposed an alternative mechanism in which reversible [Cu(II)ATSM] reductionmay occur in both normoxic and hypoxic tissue generating an unstable, anionic copper(I)species which becomes reversibly trapped by virtue of the negative charge. Once formed,the copper(I) species may undergo either dissociation, leading to irreversible release andintracellular trapping of copper(I) ions in the cell or oxidation by molecular oxygen to give theinitial, neutral [Cu(II)ATSM] which could diffuse back out of the cell. In this mechanism, theorigins of hypoxia selectivity focused on the relative stability of the reduced copper(I) aniontowards ligand dissociation or reoxidation as the discriminating factor between, for example,the different observed biodistribution of [Cu(II)ATSM] and the non-hypoxia-selective bloodperfusion tracer, [Cu(II)PTSM] (Okazawa et al 1994).
Recently, we reported extensive spectroelectrochemical and computational studieswhich led to the proposal of a revised mechanism of hypoxia selectivity of copperbis(thiosemicarbazonato) complexes (scheme 2) (Holland et al 2008b). These data builtupon the second mechanism and provided the first experimental evidence on the nature of the

Kinetics studies on Cu-ATSM 2107
reduced copper(I) species, the feasibility of reoxidation and the crucial role of protonationand ligand dissociation in the pathway leading to irreversible intracellular trapping of thecopper(I) ion.
This paper describes the mathematical simulation of pO2-dependent in vitro cellularuptake data by using non-steady-state (NSS) kinetic analysis based on our revised mechanisticmodel (scheme 2). In addition, further in vitro kinetic experiments investigating the reversible,enzyme-mediated reduction of copper(II) bis(thiosemicarbazonato) complexes and turnoverof NADH in aqueous solution are also reported. These studies provide support for theinvolvement of an enzyme-mediated reduction process and, for the first time, demonstrate thedependence of the rate of reduction on the structure of the copper(II) complex. NSS kineticmodelling suggests that a shift in emphasis from the reduction/reoxidation step to the relativerate of proton-mediated ligand dissociation is required to explain the origins and differencesin the observed hypoxia selectivity of copper(II) bis(thiosemicarbazonato) complexes.
2. Non-steady-state kinetics
Kinetic modelling of biochemical systems is an important tool used frequently to analysein vitro and in vivo experimental data, establish correlative effects, predict structure activityrelations and develop our theoretical understanding of complex biological processes. The firstmodel to consider the fate of tissue oxygenation as a function of distance from the capillarieswas reported by Krogn (1919a, 1919b,). The simplistic model consisted of two cylindricalcompartments representing the blood vessel (inner compartment) and surrounding tissue(outer compartment). Advanced methods are now capable of simulating three-dimensional(3D) maps of the complex and irregular vasculature found within solid tumours (Kelly andBrady 2006). By using experimental in vivo data acquired from dynamic PET imagingof radiopharmaceuticals such as [18F]-FMISO, the latest mathematical models have beenused to optimize imaging strategies and provide post-imaging analysis of, for example, thequantification of hypoxia in patients with head and neck cancer (Thorwarth et al 2005a,2005b). Further examples reporting the use of kinetic simulations include simulations ofskeletal uptake (Bertoldo et al 2001) and improved quantification of heterogeneous tumouruptake (Wu et al 1995) of the metabolic imaging agent [18F]-2-fluoro-2-deoxy-D-glucose,[18F]-FDG; simulations of glucose uptake and utilization in patients with type-I diabetes(Boroujerdi et al 1995); nonlinear regression analysis of the multicomponent glucose uptakekinetics in Saccharomyces cerevisae yeast (Coons et al 1995); simulations of cancer growth(Jain 2002); kinetic studies on the accumulation of chemotherapeutic anthracyclines in multi-drug resistant tumour cells (Demant and Friche 1998); and kinetic analysis of [18F]-3′-deoxy-3′-fluorothymidine as a cell proliferation marker in patients with gliomas (Muzi et al 2006).The feasibility of multitracer PET imaging using [Cu(II)ATSM], [Cu(II)PTSM], [18F]-FDGand [11C]-Acetate has also been assessed by simulations of in vivo tissue activity curves(Kadrmas and Rust 2005, Rust and Kadrmas 2006).
Although the proposed mechanism (scheme 2) is consistent with all available chemical,electrochemical, spectroscopic and computational data, the tumour microenvironment is verydifferent from the experimental conditions used (Holland et al 2008b). Therefore, the validityof the mechanism was evaluated by performing in silico simulations in which simulated pO2-dependent cellular uptake profiles were compared with experimental in vitro cellular uptakeand washout data of [64Cu(II)ATSM] in EMT6 murine carcinoma cells at varying pO2 (Lewiset al 1999). Experimental cellular uptake (1.2–1.4 × 106 cells mL−1, 37.0 ◦C) was measuredat five different oxygen concentrations corresponding to normoxic ([O2] = 20.0%), hypoxic([O2] = 5.0, 0.5, 0.1%) and anoxic ([O2] = 0.0%) conditions.

2108 J P Holland et al
Scheme 3. Schematic representation of the three-compartment, four-species, five rate constantmodel used in the NSS kinetic simulations of experimental in vitro cellular uptake and retentionof [64Cu(II)ATSM] (Holland et al 2008b, Lewis et al 1999). Compartment 1 represents theextracellular medium and contains species A which corresponds to extracellular [Cu(II)ATSM].Compartment 2 contains species B and C which correspond to the reversibly trapped [Cu(II)ATSM],intracellular reduced copper(I) anion [Cu(I)ATSM]−. Compartment 3 contains the fraction ofcopper species which remain irreversibly trapped within the cell, D.
The kinetic model can be represented by a conventional compartmental model(scheme 3). The evaluation of kinetic schemes, including the calculation of kinetic rateconstants, is usually conducted by fitting the experimental results with the integrated rateequations (Pogliani et al 1996). Solutions for the integrated rate laws often require the use ofassumption to simplify the models. Despite experimental confirmation that either mono- ordi-protonation of the reduced copper(I) ion [Cu(I)ATSM]− is required for ligand dissociation,no experimental measure of pKa of either the copper(II) or copper(I) complexes is available.As a consequence, the rate of protonation/deprotonation, corresponding to the rate constantsk5[H3 O+] and k6 in scheme 2, respectively, was assumed to be fast in comparison to the otherreactions. This assumption allows the rate constant kirr to be replaced by a new irreversiblerate constant (k5 in scheme 3) which is a convolution of the rate constants associated withthe protic equilibria and ligand dissociation reactions. The simplified four-component, five-rate constant model includes three distinct compartments which represent the extracellularmedium and two intracellular compartments corresponding to the reversibly and irreversiblytrapped copper fractions (scheme 3). Species A–D correspond to the extracellular (A) andintracellular (B) [Cu(II)ATSM], the intracellular reduced copper(I) anion [Cu(I)ATSM]− (C)and the irreversibly trapped copper species resulting from ligand dissociation compartment 3contains the fraction of copper species which remain irreversible trapped within the cell(D).
Non-steady-state (NSS) kinetic analysis was achieved by using matrix methods (Poglianiet al 1996). All simulations were performed by using Matlab (MathWorks 2006). In ourmodel, as well as assuming rapid rates of protonation/deprotonation, the concentration ofoxygen was assumed to remain constant during the experiment. The rate constants, k1–k5
are given by the rate laws (equations (1)–(4)) and have been calculated numerically by fittingto experimental pO2-dependent cellular uptake and washout data for [64Cu(II)ATSM] (Lewiset al 1999).
da
dt= k2b − k1a (1)
db
dt= k1a + k4[O2]c − (k2 + k3)b (2)
dc
dt= k3b − (k4[O2] − k5)c (3)
dd
dt= k5c. (4)
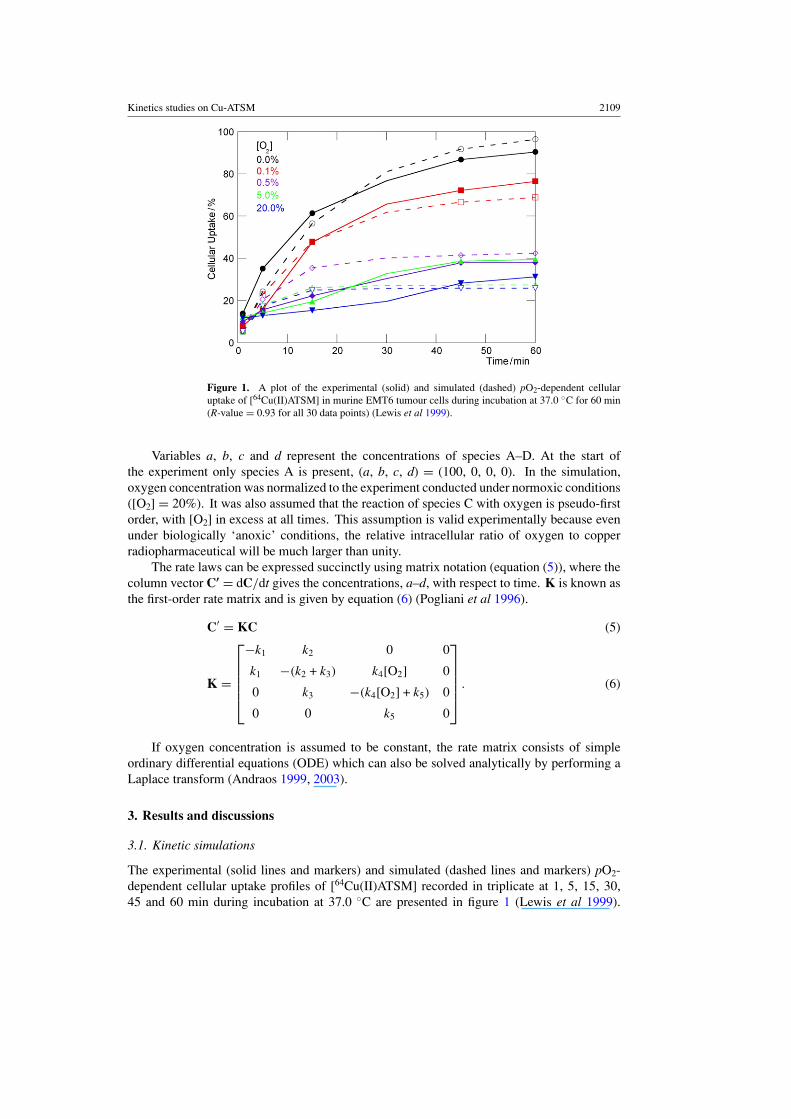
Kinetics studies on Cu-ATSM 2109
Figure 1. A plot of the experimental (solid) and simulated (dashed) pO2-dependent cellularuptake of [64Cu(II)ATSM] in murine EMT6 tumour cells during incubation at 37.0 ◦C for 60 min(R-value = 0.93 for all 30 data points) (Lewis et al 1999).
Variables a, b, c and d represent the concentrations of species A–D. At the start ofthe experiment only species A is present, (a, b, c, d) = (100, 0, 0, 0). In the simulation,oxygen concentration was normalized to the experiment conducted under normoxic conditions([O2] = 20%). It was also assumed that the reaction of species C with oxygen is pseudo-firstorder, with [O2] in excess at all times. This assumption is valid experimentally because evenunder biologically ‘anoxic’ conditions, the relative intracellular ratio of oxygen to copperradiopharmaceutical will be much larger than unity.
The rate laws can be expressed succinctly using matrix notation (equation (5)), where thecolumn vector C′ = dC/dt gives the concentrations, a–d, with respect to time. K is known asthe first-order rate matrix and is given by equation (6) (Pogliani et al 1996).
C′ = KC (5)
K =
⎡⎢⎢⎢⎢⎣
−k1 k2 0 0
k1 −(k2 + k3) k4[O2] 0
0 k3 −(k4[O2] + k5) 0
0 0 k5 0
⎤⎥⎥⎥⎥⎦
. (6)
If oxygen concentration is assumed to be constant, the rate matrix consists of simpleordinary differential equations (ODE) which can also be solved analytically by performing aLaplace transform (Andraos 1999, 2003).
3. Results and discussions
3.1. Kinetic simulations
The experimental (solid lines and markers) and simulated (dashed lines and markers) pO2-dependent cellular uptake profiles of [64Cu(II)ATSM] recorded in triplicate at 1, 5, 15, 30,45 and 60 min during incubation at 37.0 ◦C are presented in figure 1 (Lewis et al 1999).

2110 J P Holland et al
Overall, NSS kinetic analysis based on scheme 3 gives a very good fit between the observedexperimental and simulated cellular uptake of [64Cu(II)ATSM]. As the concentration of oxygendecreases and the system becomes increasingly hypoxic, the simulations correctly predict theobserved increase in cellular uptake. If all 30 experimental data points are included in thenumerical analysis, the calculated root-mean-squared error (RMSE) between the experimentaland simulated curves is <6%. Considering that Burgman et al have found that cellular uptakeof 64Cu-ATSM and the observed hypoxia selectivity is dependent upon the nature of the cellline, it is not surprising that minor differences are observed between the experimental data andthe simulations (Burgman et al 2005).
The value of k1 was fitted using only the first experiment (black lines) conducted underanoxic conditions (figure 1). If [O2] = 0.0% then all of the [64Cu(II)ATSM] that enters thecell (corresponding to species B) will be converted rapidly to species D, becoming trappedirreversibly. Under these conditions, the rate equations simplify to give a rate of irreversibletrapping that depends only on the value of k1. Rate constants k2–k5 were obtained by a separatenumerical fit using all 30 data points. The optimized rate constants are calculated to be k1 =9.8 ± 0.59 × 10−4 s−1, k2 = 2.9 ± 0.17 × 10−3 s−1, k3 = 5.2 ± 0.31 × 10−2 s−1, k4 = 2.2 ±0.13 mol−1 dm3 s−1 and k5 = 9.0 ± 0.54 × 10−5 s−1.
Interestingly, the value of k3 (0.052 ± 0.0031 s−1) which corresponds to the rate ofreduction is the same order of magnitude as the heterogeneous rate constant for electrontransfer (ks = 0.0125 ± 0.0017 cm s−1) measured from electrochemical cyclic voltammetryexperiments. In addition, if scheme 2 is replaced by a more simplistic three species, three-rate constant model (equation (7)), formally equivalent to the first mechanism proposed byFujibayashi et al and used in the NSS kinetic modelling, the optimized simulated uptake curvesdo not reproduce the correct pO2-dependent behaviour and the value of the calculated RMSEincreases to >16%.
Ak1�k′
2
Xkirr−−→ D (7)
Using the optimized rate constants, simulated pO2-dependent concentration profiles forspecies A–D versus time, t min−1, have been calculated (figures 2(a)–(e)). The value of a(t)can be compared directly with the experimental uptake measurements (100%—experimentalcellular uptake%). For each of the five values of [O2], the calculated cellular uptake profilesfor species A are in excellent agreement with the experimental data. With the exception ofthe experiment conducted at [O2] = 5.0%, the final predicted value of a(t) lies within 5% ofthe measured value. This provides confidence in the further analysis of the pO2-dependentspecies distribution.
From the species distribution profiles (figures 2(a)–(e)), it can be seen that under normoxic([O2] = 20.0%) and borderline hypoxic ([O2] = 5.0%) conditions, low-cellular uptake isobserved (a(t)) and once inside the cell, the high-oxygen concentration displaces the position ofequilibria towards the neutral [Cu(II)ATSM] (species B). Under these conditions the majorityof the intracellular copper concentration lies within compartment 2 and is only reversiblytrapped within the cell. As the oxygen concentration is decreased to 0.5% and the cells becomemore hypoxic, the position of equilibrium between the intracellular [Cu(II)ATSM] and thereduced copper(I) complex, [Cu(I)ATSM]− (species B and C, respectively) shifts towardsthe right. However, both species B and C remain reversibly trapped and may be washed outof the cells by reintroducing oxygen into the experiment (Dearling et al 2002, Lewis et al 1999).The observed experimental washout was confirmed by performing kinetic simulations in whichthe final concentrations of species A–D calculated at 60 min, from the [O2] = 0.5% experiment,were used as initial input concentrations and the [O2] was reset to 20.0%. In accordance with
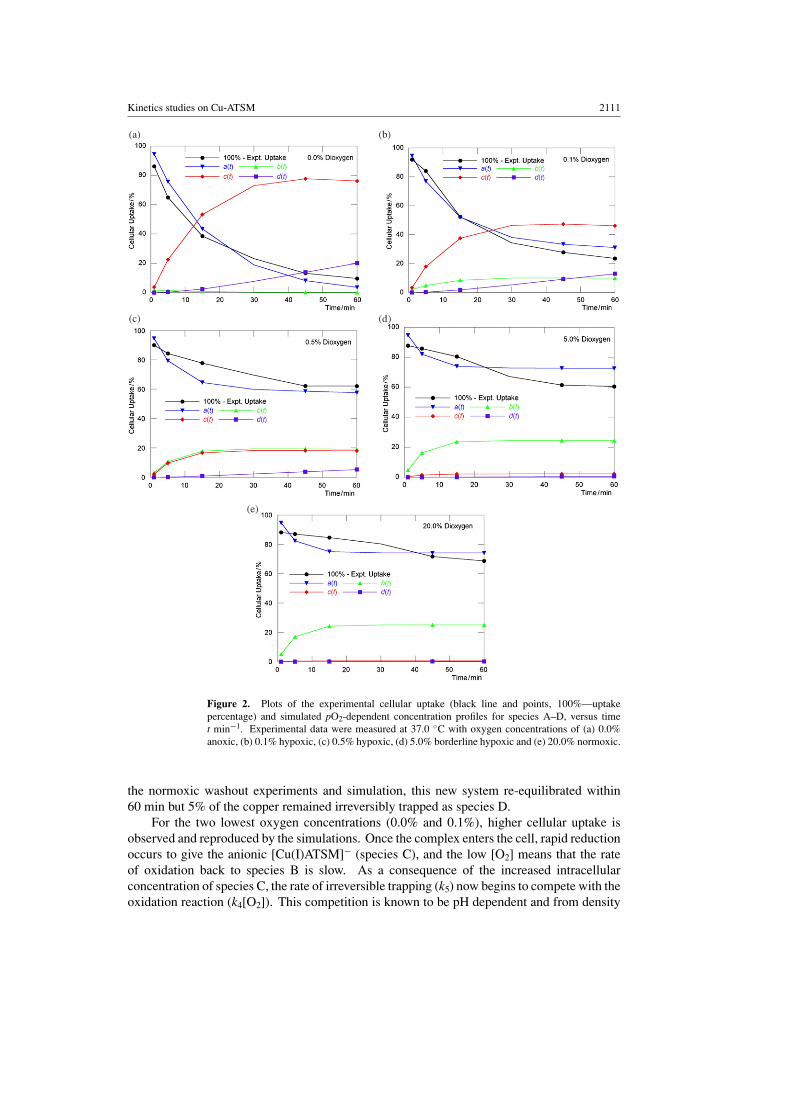
Kinetics studies on Cu-ATSM 2111
(a) (b)
(c)
(e)
(d)
Figure 2. Plots of the experimental cellular uptake (black line and points, 100%—uptakepercentage) and simulated pO2-dependent concentration profiles for species A–D, versus timet min−1. Experimental data were measured at 37.0 ◦C with oxygen concentrations of (a) 0.0%anoxic, (b) 0.1% hypoxic, (c) 0.5% hypoxic, (d) 5.0% borderline hypoxic and (e) 20.0% normoxic.
the normoxic washout experiments and simulation, this new system re-equilibrated within60 min but 5% of the copper remained irreversibly trapped as species D.
For the two lowest oxygen concentrations (0.0% and 0.1%), higher cellular uptake isobserved and reproduced by the simulations. Once the complex enters the cell, rapid reductionoccurs to give the anionic [Cu(I)ATSM]− (species C), and the low [O2] means that the rateof oxidation back to species B is slow. As a consequence of the increased intracellularconcentration of species C, the rate of irreversible trapping (k5) now begins to compete with theoxidation reaction (k4[O2]). This competition is known to be pH dependent and from density

2112 J P Holland et al
Scheme 4. Schematic representation of the experimental design used in the in vitro enzyme-mediated kinetic experiments. Experimentally the rates for electron transfer between NADH andthe reductase enzyme (in the oxidized form, ket(NADH)) and between the reduced copper(I) anion,[Cu(I)L]−, and dissolved oxygen are both faster than the observed rate of electron transfer (seethe main text for discussions) between the reduced reductase enzyme and the neutral [Cu(II)L]complex, ket(Enz-Cu). Reactive oxygen species (ROS) include HO2
− and HO− anions.
functional calculations, calculated pKa values indicate that copper bis(thiosemicarbazonato)complexes with fewer alkyl groups on the ligand backbone (for example, [Cu(II)PTSM]),this rate of ligand dissociation will be increased further by shifting the position of equilibriatowards the more heavily protonated species (scheme 2) (Holland et al 2006). The simulateduptake profiles indicate that after 60 min incubation at 0.0% oxygen, the concentration ofirreversibly trapped species D is around 21% (Lewis et al 1999). From the calculated valuesof the rate constants, it can be seen that irrespective of the relative degree of protonation, therate of irreversibly ligand dissociation and trapping (k5) is two orders of magnitude smallerthan the rate of reduction (k3).
These kinetic simulations indicate that although accepted opinions on the mechanism ofhypoxia selectivity of [Cu(II)ATSM] focus on the rate of reduction and reoxidation as theprimary factor contributing to the observed correlation between tissue uptake and [O2], itis more likely that proton-mediated ligand dissociation plays the major role (Holland et al2008b, Dearling et al 1998, 2002, Fujibayashi et al 1997, Maurer et al 2002, Obata et al 2001,Holland et al 2006). The relative stability of the reduced copper(I) bis(thiosemicarbazonato)species towards ligand dissociation is pH dependent and is governed by the nature of theligand (Holland et al 2008b). Therefore, although the experimental rates of reduction (k3)and reoxidation (k4) differ between the hypoxia-selective [Cu(II)ATSM] and the non-selective[Cu(II)PTSM], the actual origins of hypoxia selectivity may arise from their different rates ofpH-dependent ligand dissociation (k5).
Overall, these kinetic simulations indicate that the proposed mechanism shown inscheme 2 is at least plausible and can account for the observed differences in pO2-dependentcellular uptake and washout of the hypoxia-selective radiopharmaceutical, [Cu(II)ATSM].
3.2. Enzymatic reduction studies
Further support for the proposed mechanism (scheme 2) was obtained by conducting in vitrokinetic studies investigating the rate of enzyme-mediated reduction of a range of copper(II)bis(thiosemicarbazonato) complexes. Structures of the copper(II) complexes studied areshown in figure 3. The rate of electron transfer from an NADH-dependent ferrodoxin reductaseenzyme (PuR) obtained from the bacterium Rhodopseudomonas palustris (R. palustris), tothe copper(II) complexes was measured by monitoring the rate of oxidation of NADH by

Kinetics studies on Cu-ATSM 2113
Figure 3. Structures of the copper(II) bis(thiosemicarbazonato) complexes studied in the in vitroenzyme-mediated kinetic experiments.
using electronic absorption spectroscopy (scheme 4). In Tris buffer at pH 7.4, NADHhas an intense absorption peak at 340 nm (molar absorption coefficient, ε340 = 6.22 ×103 M−1 cm−1). The oxidized species NAD+ does not absorb at or near 340 nm. The[Cu(II)ATSM] and [Cu(I)ATSM]− complexes show a well-defined isosbestic point at 347nm allowing the rate of NADH oxidation (ε347 = 5.95 × 103 M−1 cm−1) to be monitoredat this wavelength without interference from potential changes in absorbance due to thecopper species (Holland et al 2008b). Unfortunately due to the experimental apparatus it wasnot possible to conduct these experiments at varying oxygen concentrations. All data wererecorded under normal atmospheric [O2] conditions.
The proposed reaction (scheme 4) involves an initial two-electron transfer in the formof a hydride ion from NADH to the enzyme. Then, in the presence of a reduciblecopper(II) bis(thiosemicarbazonato) complexes, successive one-electron transfer reactionsoccur generating the reduced copper(I) anionic species, [Cu(I)L]−. Under the experimentalconditions, the high concentration of dissolved oxygen leads to the rapid oxidation of thiscopper(I) anion, regenerating the neutral [Cu(II)L] complex and producing a reactive oxygenspecies. Presumably in the aqueous medium, if the initial ROS was formed as a superoxideanion, it would react rapidly with water to give peroxide and hydroxide anions (Maricle andHodgson 1965). In this mechanism, the rate of electron transfer from NADH to the enzyme(ket(NADH)) is independent of the nature of the copper(II) complex. However, the rate ofconsumption of NADH, as measured by the decrease in absorbance, will depend upon therate of the electron transfer process between the enzyme and the copper complex. Previouslyreported experiments and the kinetic simulations presented above have shown that the rate ofoxidation of the reduced copper(I) anion by dissolved oxygen (ket(Cu–O2), is fast (Hollandet al 2008b). Therefore, any observed difference in the rate of NADH turnover is directlyrelated to changes in the rate of electron transfer from the enzyme to the copper(II) complex.
At this point it is noted that the rate constant from electron transfer between the enzyme andthe copper(II) complex (ket(Enz–Cu) is complex and may consist of contributions from the rateof binding of the copper(II) complexes to the enzyme (to give the bound (enzyme-substrate)complex), the rate of electron transfer within the (enzyme-substrate) complex and the rate ofdissociation of the reduced copper(I) anion from the oxidized enzyme. For small moleculeelectron acceptors the initial association to form a competent complex and dissociation ofthe products after electron transfer are generally fast. However, the experiments reported
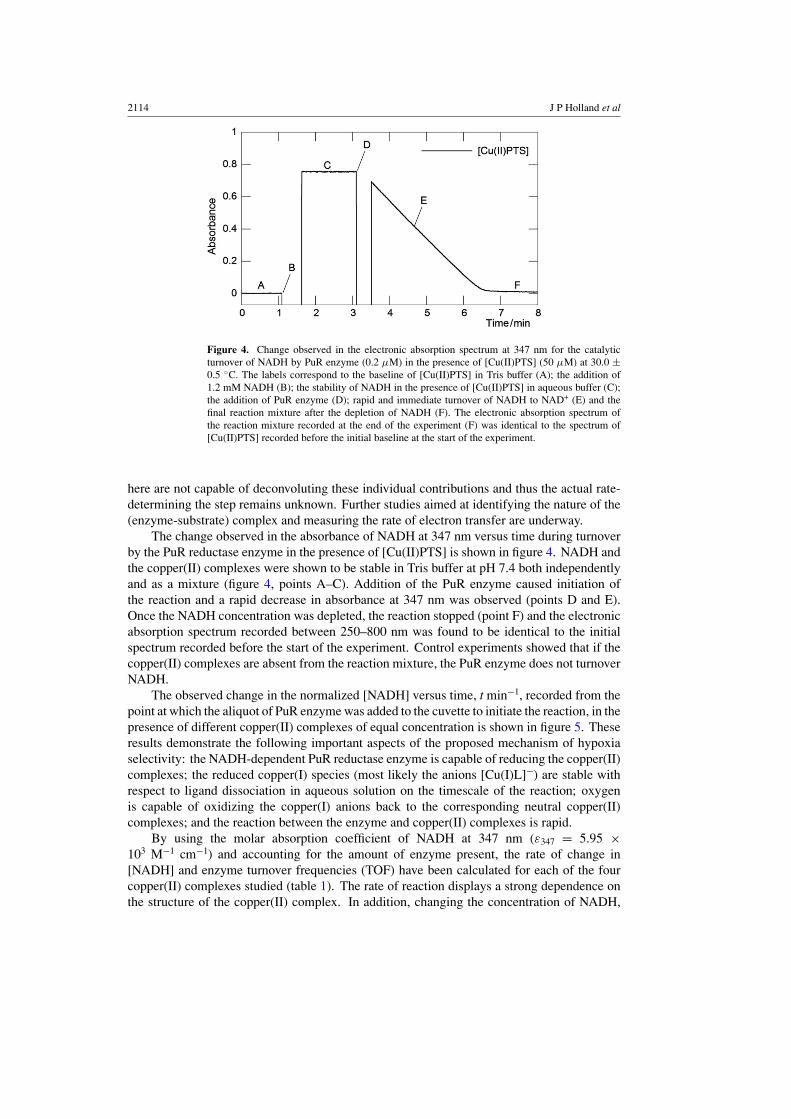
2114 J P Holland et al
Figure 4. Change observed in the electronic absorption spectrum at 347 nm for the catalyticturnover of NADH by PuR enzyme (0.2 μM) in the presence of [Cu(II)PTS] (50 μM) at 30.0 ±0.5 ◦C. The labels correspond to the baseline of [Cu(II)PTS] in Tris buffer (A); the addition of1.2 mM NADH (B); the stability of NADH in the presence of [Cu(II)PTS] in aqueous buffer (C);the addition of PuR enzyme (D); rapid and immediate turnover of NADH to NAD+ (E) and thefinal reaction mixture after the depletion of NADH (F). The electronic absorption spectrum ofthe reaction mixture recorded at the end of the experiment (F) was identical to the spectrum of[Cu(II)PTS] recorded before the initial baseline at the start of the experiment.
here are not capable of deconvoluting these individual contributions and thus the actual rate-determining the step remains unknown. Further studies aimed at identifying the nature of the(enzyme-substrate) complex and measuring the rate of electron transfer are underway.
The change observed in the absorbance of NADH at 347 nm versus time during turnoverby the PuR reductase enzyme in the presence of [Cu(II)PTS] is shown in figure 4. NADH andthe copper(II) complexes were shown to be stable in Tris buffer at pH 7.4 both independentlyand as a mixture (figure 4, points A–C). Addition of the PuR enzyme caused initiation ofthe reaction and a rapid decrease in absorbance at 347 nm was observed (points D and E).Once the NADH concentration was depleted, the reaction stopped (point F) and the electronicabsorption spectrum recorded between 250–800 nm was found to be identical to the initialspectrum recorded before the start of the experiment. Control experiments showed that if thecopper(II) complexes are absent from the reaction mixture, the PuR enzyme does not turnoverNADH.
The observed change in the normalized [NADH] versus time, t min−1, recorded from thepoint at which the aliquot of PuR enzyme was added to the cuvette to initiate the reaction, in thepresence of different copper(II) complexes of equal concentration is shown in figure 5. Theseresults demonstrate the following important aspects of the proposed mechanism of hypoxiaselectivity: the NADH-dependent PuR reductase enzyme is capable of reducing the copper(II)complexes; the reduced copper(I) species (most likely the anions [Cu(I)L]−) are stable withrespect to ligand dissociation in aqueous solution on the timescale of the reaction; oxygenis capable of oxidizing the copper(I) anions back to the corresponding neutral copper(II)complexes; and the reaction between the enzyme and copper(II) complexes is rapid.
By using the molar absorption coefficient of NADH at 347 nm (ε347 = 5.95 ×103 M−1 cm−1) and accounting for the amount of enzyme present, the rate of change in[NADH] and enzyme turnover frequencies (TOF) have been calculated for each of the fourcopper(II) complexes studied (table 1). The rate of reaction displays a strong dependence onthe structure of the copper(II) complex. In addition, changing the concentration of NADH,
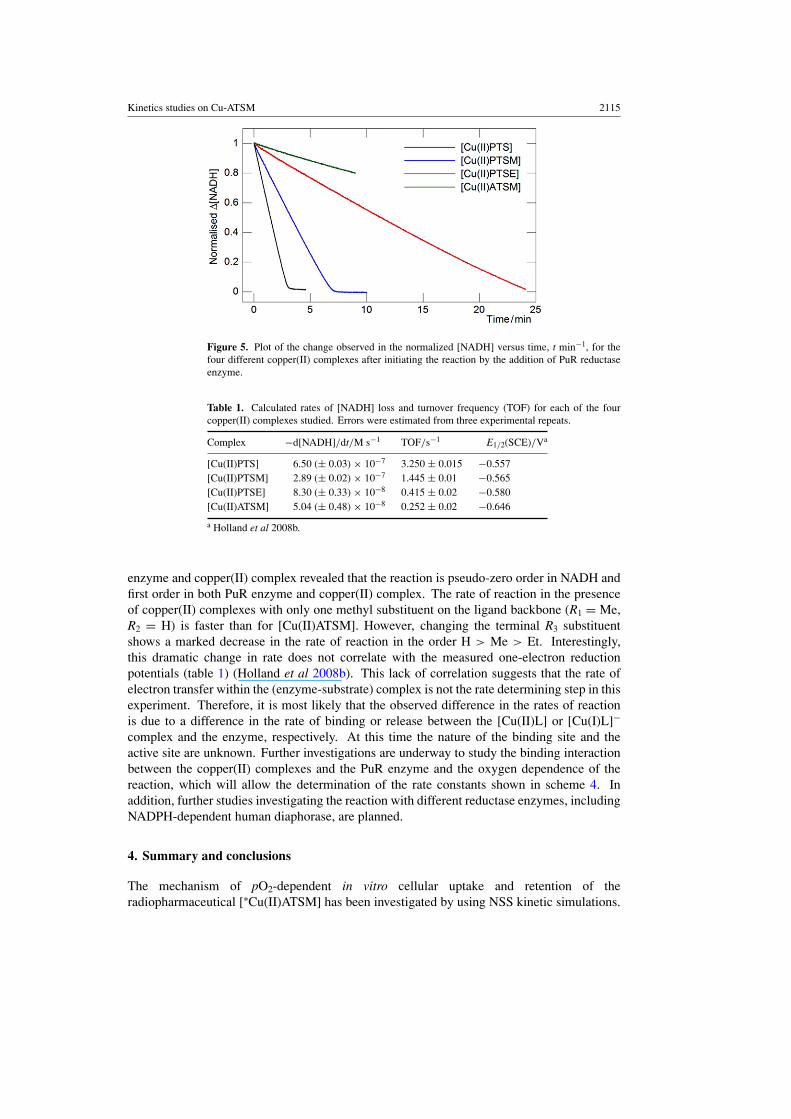
Kinetics studies on Cu-ATSM 2115
Figure 5. Plot of the change observed in the normalized [NADH] versus time, t min−1, for thefour different copper(II) complexes after initiating the reaction by the addition of PuR reductaseenzyme.
Table 1. Calculated rates of [NADH] loss and turnover frequency (TOF) for each of the fourcopper(II) complexes studied. Errors were estimated from three experimental repeats.
Complex −d[NADH]/dt/M s−1 TOF/s−1 E1/2(SCE)/Va
[Cu(II)PTS] 6.50 (± 0.03) × 10−7 3.250 ± 0.015 −0.557[Cu(II)PTSM] 2.89 (± 0.02) × 10−7 1.445 ± 0.01 −0.565[Cu(II)PTSE] 8.30 (± 0.33) × 10−8 0.415 ± 0.02 −0.580[Cu(II)ATSM] 5.04 (± 0.48) × 10−8 0.252 ± 0.02 −0.646
a Holland et al 2008b.
enzyme and copper(II) complex revealed that the reaction is pseudo-zero order in NADH andfirst order in both PuR enzyme and copper(II) complex. The rate of reaction in the presenceof copper(II) complexes with only one methyl substituent on the ligand backbone (R1 = Me,R2 = H) is faster than for [Cu(II)ATSM]. However, changing the terminal R3 substituentshows a marked decrease in the rate of reaction in the order H > Me > Et. Interestingly,this dramatic change in rate does not correlate with the measured one-electron reductionpotentials (table 1) (Holland et al 2008b). This lack of correlation suggests that the rate ofelectron transfer within the (enzyme-substrate) complex is not the rate determining step in thisexperiment. Therefore, it is most likely that the observed difference in the rates of reactionis due to a difference in the rate of binding or release between the [Cu(II)L] or [Cu(I)L]−
complex and the enzyme, respectively. At this time the nature of the binding site and theactive site are unknown. Further investigations are underway to study the binding interactionbetween the copper(II) complexes and the PuR enzyme and the oxygen dependence of thereaction, which will allow the determination of the rate constants shown in scheme 4. Inaddition, further studies investigating the reaction with different reductase enzymes, includingNADPH-dependent human diaphorase, are planned.
4. Summary and conclusions
The mechanism of pO2-dependent in vitro cellular uptake and retention of theradiopharmaceutical [∗Cu(II)ATSM] has been investigated by using NSS kinetic simulations.

2116 J P Holland et al
Kinetic analysis using the proposed mechanism reveals that the model is in very goodagreement with the observed experimental cellular uptake. Calculated rate constants suggestthat the origins of hypoxia-selective uptake and trapping may reside with the relative,structure-dependent stability of the reduced copper(I) anion [Cu(I)L]− towards protonation andsubsequent ligand dissociation (k5) and not as previously proposed with the rate of reductionand oxidation (k3 and k4[O2], respectively) (Holland et al 2008b, Dearling et al 1998, 2002,Fujibayashi et al 1997, Maurer et al 2002, Obata et al 2001, Holland et al 2006).
Studies investigating the in vitro turnover of NADH by the reductase enzyme PuRdemonstrate that NADH-dependent enzyme mediated reduction of the copper(II) complexesis likely to occur in vivo. The results also demonstrate that the copper(I) anion [Cu(I)L]−
is stable in aqueous buffer solution with respect to ligand dissociation, and under normoxicconditions (atmospheric [O2]) it is rapidly oxidized by molecular oxygen, back to the neutral[Cu(II)L] complex. Differences in the observed rates of NADH turnover demonstrate that thestructure of the copper(II) complex plays an important role on the relative rates of formationand dissociation of the (enzyme-substrate) complex, and potentially on the reduction rate ofthe copper(II) complex. Further studies are underway aimed at probing the nature of theinteraction between the enzyme and the copper(II) complexes and investigating the affectof using different reductase enzymes, including those derived from humans. An improvedunderstanding of the nature of the mechanism of hypoxia selectivity for this class of copper(II)complexes may facilitate the development of new radiopharmaceutical imaging agents withincreased selectivity for tumour hypoxia.
5. Experimental details
5.1. Synthesis
Copper(II) complexes were synthesized in accordance with previously reported procedures(Holland et al 2008a, 2008b). Characterization data were consistent with the proposedstructures and purity and structural identity were confirmed by microanalyses, high-performance liquid chromatography and high-resolution electrospray mass spectrometry.
5.2. General methods and materials
NADH was obtained from Roche Diagnostics, UK. The genomic DNA of R. palustris CGA009(ATCC BAA-98) was obtained from ATCC-LGC Promochem, UK. The pCWori+ vector wasfrom Richard Dalquist (University of Oregon, USA) and Roland Wolf (University of Dundee,UK). UV/vis spectra and spectroscopic activity assays were recorded at 30 ± 0.5 ◦C on aVarian 1E spectrophotometer.
5.3. PuR enzyme expression
The PuR (RPA3782) protein was produced by using in E coli DH5α transformed witha pCWori+ plasmid containing the RPA3782 gene and purified by using ion-exchangechromatography methods similar to those used for PdR from Pseudomonas putida (Petersonet al 1990, Peng et al 2007). Purified fractions (A280/A454 < 6) were combined and concentratedby ultrafiltration. PuR was stored at −20 ◦C in 50 mM Tris at pH 7.4, containing 50% v/vglycerol. Glycerol was removed immediately before use by gel filtration on a 5 mL PD-10column (GE Healthcare) by eluting with 50 mM Tris, pH 7.4.

Kinetics studies on Cu-ATSM 2117
5.4. Enzyme assays
Steady state NADH turnover rate assays were performed with mixtures (1.2 mL) containing50 mM Tris, pH 7.4 and 0.2 μM PuR. The mixtures were equilibrated at 30 ◦C for 2 min.Substrates were added as a 1.0 mM stock solution in DMSO to a give final concentration of50 μM in Tris buffer with <5% DMSO. Fresh solutions of NADH (20 mgmL−1, 28.2 mM)were prepared and stored at 0 ◦C prior to use. NADH was added in 5 μL aliquots to give a finalconcentration of 0.12 mM in the reaction cuvette. The rate of NADH turnover was monitoredby observing the decrease in absorbance at 347 nm (ε347 = 5.95 mM−1 cm−1) versus time,t min−1.
Acknowledgments
JPH thanks the Engineering and Physical Sciences Research Council (EPSRC) and MertonCollege, Oxford, for funding. JHG thanks Magdalen College, Oxford, for a Junior ResearchFellowship. We thank Dr Jason S Lewis for helpful discussions and for the provision ofexperimental cellular uptake data.
References
Andraos J 1999 A streamlined approach to solving simple and complex kinetic systems analytically J. Chem. Educ.76 1578–83
Andraos J 2003 Quantification and optimization of dynamic kinetic resolution J. Phys. Chem. A 107 2374–87Bentzen L, Keiding S, Nordsmark M, Falborg L, Hansen S B, Keller J, Nielsen O S and Overgaard J 2003 Tumour
oxygenation assessed by 18 F-fluoromisonidazole PET and polarographic needle electrodes in human soft tissuetumours Radiother. Oncol. 67 339–44
Bertoldo A, Peltoniemi P, Oikonen V, Knuuti J, Nuutila P and Cobelli C 2001 Kinetic modeling of [18 F]FDG inskeletal muscle by PET: a four-compartment five-rate-constant model Am. J. Physiol. 281 E524–36
Boroujerdi M A, Umpleby A M, Jones R H and Sonksen P H 1995 A simulation model for glucose kinetics andestimates of glucose utilization rate in type 1 diabetic patients Am. J. Physiol. 268 E766–74
Brown J M and Wilson W R 2004 Exploiting tumor hypoxia in cancer treatment Nat. Rev. Cancer 4 437–47Bunn H F and Poyton R O 1996 Oxygen sensing and molecular adaptation to hypoxia Physiol. Rev. 76 839–85Burgman P, O’Donoghue J A, Lewis J S, Welch M J, Humm J L and Ling C C 2005 Cell line-dependent
differences in uptake and retention of the hypoxia-selective nuclear imaging agent Cu-ATSM Nucl. Med. Biol.32 623–30
Coons D M, Boulton R B and Bisson L F 1995 Computer-assisted nonlinear regression analysis of the multicomponentglucose uptake kinetics of Saccharomyces cerevisiae J. Bacteriol. 177 3251–8
Couturier O, Luxen A, Chatal J-F, Vuillez J-P, Rigo P and Hustinx R 2004 Fluorinated tracers for imaging cancerwith positron emission tomography Eur. J. Nucl. Med. Mol. Imaging 31 1182–206
Dearling J L J, Lewis J S, McCarthy D W, Welch M J and Blower P J 1998 Redox-active metal complexesfor imaging hypoxic tissues: structure-activity relationships in copper(II) bis(thiosemicarbazone) complexesChem. Commun. 2531–2
Dearling J L J, Lewis J S, Mullen G E D, Welch M J and Blower P J 2002 Copper bis(thiosemicarbazone) complexesas hypoxia imaging agents: structure-activity relationships J. Biol. Inorg. Chem. 7 249–59
Dehdashti F, Mintun M A, Lewis J S, Bradley J, Govindan R, Laforest R, Welch M J and Siegel B A 2003 In vivoassessment of tumor hypoxia in lung cancer with 60Cu-ATSM Eur. J. Nucl. Med. Mol. Imaging 30 844–50
Demant E J F and Friche E 1998 Kinetics of anthracycline accumulation in multidrug-resistant tumor cells: relationshipto drug lipophilicity and serum albumin binding Biochem. Pharmacol. 56 1209–17
Fujibayashi Y, Taniuchi H, Yonekura Y, Ohtani H, Konishi J and Yokoyama A 1997 Copper-62-ATSM: a new hypoxiaimaging agent with high membrane permeability and low redox potential J. Nucl. Med. 38 1155–60
Fujibayashi Y, Wada K, Taniuchi H, Yonekura Y, Konishi J and Yokoyama A 1993 Mitochondria-selectivereduction of 62Cu-pyruvaldehyde bis(N4-methylthiosemicarbazone) (62Cu-PTSM) in the murine brain: anovel radiopharmaceutical for brain positron emission tomography (PET) imaging Biol. Pharm. Bull. 16
146–9

2118 J P Holland et al
Heacock C S and Sutherland R M 1986 Induction characteristics of oxygen regulated proteins Int. J. Radiat. Oncol.Biol. Phys. 12 1287–90
Holland J P, Barnard P J, Bayly S R, Betts H M, Churchill G C, Dilworth J R, Edge R, Green J C andHueting R 2008a Synthesis, radiolabelling and confocal fluorescence microscopy of styrene-derivatisedbis(thiosemicarbazonato)zinc and copper complexes Eur. J. Inorg. Chem. 1985–93
Holland J P, Barnard P J, Collison D, Dilworth J R, Edge R, Green J C and McInnes E J L 2008b Spectroelectrochemicaland computational studies on the mechanism of hypoxia selectivity of copper radiopharmaceuticals Chem. AEur. J. 14 5890–907
Holland J P, Green J C and Dilworth J R 2006 Probing the mechanism of hypoxia selectivity of copperbis(thiosemicarbazonato) complexes: DFT calculation of redox potentials and absolute acidities in solutionDalton Trans. 783–94
Jain S 2002 Kinetic model for designing a cancer therapy Cancer Cell Int. 2 13–8Kadrmas D J and Rust T C 2005 Feasibility of rapid multitracer PET tumor imaging IEEE Trans. Nucl. Sci.
52 1341–7Kelley S L, Basu A, Teicher B A, Hacker M P, Hamer D H and Lazo J S 1988 Overexpression of metallothionein
confers resistance to anticancer drugs Science 241 1813–5Kelley S L and Rozencweig M 1989 Resistance to platinum compounds: mechanisms and beyond Eur. J. Cancer
Clin. Oncol. 25 1135–40Kelly C J and Brady M 2006 A model to simulate tumour oxygenation and dynamic [18F]-Fmiso PET data Phys.
Med. Biol. 51 5859–73Krogn A 1919a The number and distribution of capillaries in muscleswith calculations of the oxygen pressure
necessary for supplying tissue J. Physiol. (London) 52 409–15Krogn A 1919b The rate of diffusion of gases through animal tissues, with some remarks on the coefficient of invasion
J. Physiol. (London) 52 391–408Lewis J S, McCarthy D W, McCarthy T J, Fujibayashi Y and Welch M J 1999 Evaluation of 64Cu-ATSM in vitro
and in vivo in a hypoxic tumor model J. Nucl. Med. 40 177–83Maricle D L and Hodgson W G 1965 Reduction of oxygen to superoxide anion in aprotic solvents Anal. Chem.
37 1562–5MathWorks 2006 Matlab 7.1Maurer R I, Blower P J, Dilworth J R, Reynolds C A, Zheng Y and Mullen G E D 2002 Studies on the mechanism of
hypoxic selectivity in copper bis(thiosemicarbazone) radiopharmaceuticals J. Med. Chem. 45 1420–31Murphy B J, Laderoute K R, Chin R J and Sutherland R M 1994 Metallothionein IIA is up-regulated by hypoxia in
human A431 squamous carcinoma cells Cancer Res. 54 5808–10Muzi M, Spence A M, O’Sullivan F, Mankoff D A, Wells J M, Grierson J R, Link J M and Krohn K A 2006 Kinetic
analysis of 3’-deoxy-3’-18 F-fluorothymidine in patients with gliomas J. Nucl. Med. 47 1612–21Nordsmark M et al 2001 Invasive oxygen measurements and pimonidazole labeling in human cervix carcinoma Int.
J. Radiat. Oncol. Biol. Phys. 49 581–6Nordsmark M et al 2003 Measurements of hypoxia using pimonidazole and polarographic oxygen-sensitive electrodes
in human cervix carcinomas Radiother. Oncol. 67 35–44Obata A, Yoshimi E, Waki A, Lewis J S, Oyama N, Welch M J, Saji H, Yonekura Y and Fujibayashi Y
2001 Retention mechanism of hypoxia selective nuclear imaging/radiotherapeutic agent Cu-diacetyl-bis(N4-methylthiosemicarbazone) (Cu-ATSM) in tumor cells Ann. Nucl. Med. 15 499–504
Okazawa H, Yonekura Y, Fujibayashi Y, Nishizawa S, Magata Y, Ishizu K, Tanaka F, Tsuchida T, Tamaki N andKonishi J 1994 Clinical application and quantitative evaluation of generator-produced copper-62-PTSM as abrain perfusion tracer for PET J. Nucl. Med. 35 1910–5
Parker C et al 2004 Polarographic electrode study of tumor oxygenation in clinically localized prostate cancer Int. J.Radiat. Oncol. Biol. Phys. 58 750–7
Peng Y, Xu F, Bell S G, Wong L L and Rao Z 2007 Crystallization and preliminary x-ray diffraction studies of aferredoxin reductase from Rhodopseudomonas palustris CGA009 Acta. Crystallogr. F 63 422–5
Peterson J A, Lorence M C and Amarneh B 1990 Putidaredoxin reductase and putidaredoxin. Cloning, sequencedetermination, and heterologous expression of the proteins J. Biol. Chem. 265 6066–73
Pogliani L, Berberan-Santos M N and Martinho J M G 1996 Matrix and convolution methods in chemical kineticsJ. Math. Chem. 20 193–210
Rust T C and Kadrmas D J 2006 Rapid dual-tracer PTSM+ATSM PET imaging of tumour blood flow and hypoxia:a simulation study Phys. Med. Biol. 51 61–75
Satoh M, Kloth D M, Kadhim S A, Chin J L, Naganuma A, Imura N and Cherian M G 1993 Modulation of bothcisplatin nephrotoxicity and drug resistance in murine bladder tumor by controlling metallothionein synthesisCancer Res. 53 1829–32

Kinetics studies on Cu-ATSM 2119
Thorwarth D, Eschmann S-M, Scheiderbauer J, Paulsen F and Alber M 2005a Kinetic analysis of dynamic[18 F]-fluoromisonidazole PET correlates with radiation treatment outcome in head-and-neck cancer BMCCancer 5 152
Thorwarth D, Eschmann S-M, Paulsen F and Alber M 2005b A kinetic model for dynamic [18 F]-Fmiso PET data toanalyse tumour hypoxia Phys. Med. Biol. 50 2209–24
Vavere A L and Lewis J S 2007 Cu-ATSM: a radiopharmaceutical for the PET imaging of hypoxia Dalton Trans.4893–902
Wu H-M, Huang S-C, Choi Y, Hoh C K and Hawkins R A 1995 A modeling method to improve quantitation offluorodeoxyglucose uptake in heterogeneous tumor tissue J. Nucl. Med. 36 297–306




















