
Sparse Envelope Spectra for Feature Extraction of Bearing ...
Upload
independentCategory
view
2download
0
Plant Science 128 (1997) 169–180
In vitro and in organello phosphorylation of envelope proteinsand phosphoglucomutase in spinach chloroplasts
Lucien Bovet *, Benedicte L’Eplattenier, Paul-Andre Siegenthaler
Laboratoire de Physiologie Vegetale, Uni6ersite de Neuchatel, Emile Argand 13, CH-2007 Neuchatel, Switzerland
Received 6 February 1997; received in revised form 23 June 1997; accepted 24 June 1997
Abstract
In this investigation, we found that the main source of ATP necessary for envelope protein phosphorylation isgenerated by photophosphorylation. Indeed, seven envelope polypeptides exhibiting Mr of about 60, 51, 35, 33, 26,14 and 13 kD were found to be fully labelled in organello in the presence of [32P]Pi and light. The majority of thesephosphopolypeptides were also detected when envelopes were phosphorylated in vitro in the presence of [g-32P]ATP.This supports the idea that the chloroplast stroma significantly supplies envelope protein-kinases with ATP in thelight. Preincubation of intact chloroplasts in darkness or with the phosphate translocator inhibitor 4,4%-diisothio-cianato-stilbene-2,2%-disulfonate (DIDS) strongly prevented chloroplast protein phosphorylation in the presence of[32P]Pi, thereby attesting the involvement of chloroplast ATP in the envelope protein phosphorylation process. In thedark, two chloroplast soluble polypeptides of about 94 and 67 kD were also labelled, when intact chloroplasts werefed with [32P]Pi, thus indicating that these two polypeptides are able to use dark-phosphorylation pathways,independent of the photophosphorylation process, to become phosphorylated. Our results show also that the 67 kDdark-phosphorylated protein is very likely to be a phosphoglucomutase, since its labelling was markedly reduced bythe addition of exogenous glucose 1- or 6-phosphate. © 1997 Elsevier Science Ireland Ltd.
Keywords: Chloroplast envelopes; Phosphoprotein; Protein-kinase activities; Phosphoglucomutase; DIDS; Spinaciaoleracea L.
Abbre6iations: BSA, Bovine serum albumin; Chl, Chloro-phyll; DIDS, 4,4%-diisothiocyanato-stilbene-2,2%-disulfonic acid;LHCII, Light-harvesting chl a/b proteins of PSII; PAGE,Polyacrylamide gel electrophoresis; Mops, 3-[N-mor-pholino]propanesulfonic acid; PGM, Phosphoglucomutase;Rubisco, Ribulose-1,5-bisphosphate carboxylase/oxygenase;SDS, Sodium dodecyl sulfate; Tricine, N-[2-hydroxy-1.1-bis(hydroxymethyl)ethyl]glycine; 2D, Two dimensional.
* Corresponding author. Fax: +41 32 7182271; e-mail:[email protected]
1. Introduction
Chloroplast envelope membranes are involvedin several biosynthesis and photosynthesis regulat-ing pathways [1,2]. The control of some of theactivities catalyzed by envelope enzymes requiresposttranslational modifications, such as protein
0168-9452/97/$17.00 © 1997 Elsevier Science Ireland Ltd. All rights reserved.
PII S 0168 -9452 (97 )00160 -X

L. Bo6et et al. / Plant Science 128 (1997) 169–180170
phosphorylation, that is indeed well-known tooccur when isolated envelope vesicles are incu-bated in the presence of [g-32P]ATP or [g-32P]GTP [3,4]. The extent of this phosphateincorporation in envelope membrane proteins isclearly emphasized by the fact that the numberof substrate proteins is greater in envelopes thanin the stroma and thylakoids [5]. However, verylittle is known to date about the significance ofthe envelope protein phosphorylation, since onlythree proteins associated with envelope vesiculeshave been so far identified to be phosphorylablesubstrates: a phosphoglucomutase [3,6,7], a poolof LHCII and the Rubisco small subunit [8].
To better understand the role of protein phos-phorylation in envelope membranes, it is notonly necessary to identify the target proteins,but also to characterize the associated protein-kinase(s). Unfortunately, the information con-cerning the envelope protein-kinases is still verypoor, since no sequence coding for a typical en-velope protein-kinase has yet been reported inthe literature. The potential candidates havebeen found to be associated with the chloroplastouter envelope membrane [9,10].
Another possible approach is to determine therespective involvement of cytoplasmic andchloroplastic ATP in the phosphorylation of en-velope proteins, since photosynthetic plant cellshave two main ATP producing organelles: mito-chondria and chloroplasts. Some envelopeproteins have been reported to incorporate inorganello labelled phosphate from exogenous [g-32P]ATP [11], as a like-cytoplasmic ATP source.However, this has not yet been demonstratedfor a chloroplastic ATP source, although thelight-generated ATP is well-known to be partlyused as a substrate for thylakoid and stromalprotein phosphorylation [12–14].
The aim of this paper is to compare the pat-tern of phosphorylated proteins in envelopemembranes of spinach chloroplasts after la-belling either in vitro in the presence of [g32-P]ATP or in organello in the presence of[g-32P]ATP or [32P]Pi and light. It is concludedthat ATP generated by photophosphorylation isone of the major sources used in catalyzing thephosphorylation of envelope proteins (e.g. the
60, 51, 35, 33, 26, 14 and 13 kD polypeptides).In the dark, two chloroplast soluble polypep-tides of about 94 and 67 kD are however alsoable to incorporate phosphate in organello justfrom supplied [32P]Pi. As judged by its ability toexchange phosphate with glucose 1- or 6-phos-phate, the 67 kD phosphopolypeptide is mostlikely a phosphoglucomutase.
2. Materials and methods
2.1. Plant material and chemicals
Spinach leaves (Spinacia oleracea L.) were pur-chased from the local market. [g-32P]ATP (PB10168) and [32P]Pi orthophosphate (PBS 11) werefrom Amersham-Rhan (Zurich, CH) and 4,4%-diiso-thiocyanato-stilbene-2,2%-disulfonic acid (DIDS)was from Sigma (Chemie Brunswig AG, Basel, CH).
2.2. Preparation of intact chloroplasts, stroma,en6elope and thylakoid membranes
Intact chloroplasts were isolated as describedby Mourioux and Douce [15]. Thylakoids,stroma and envelope membranes were preparedfrom intact chloroplasts according to the meth-ods of Rawyler et al. [16], Douce and Joyard[17] and Siegenthaler and Dumont [18], respec-tively. The quantitative determination of chloro-phylls was carried out according to Bruinsma[19]. The protein concentration was determinedusing the dye-binding method of Bradford [20]or Lowry et al. [21] with a BSA standard.
2.3. In 6itro protein phosphorylation assay
Phosphorylation of proteins (30 mg) was car-ried out in the presence of 4 mM MgCl2, 50mM Mops-NaOH, pH 7.6, and 33 nM [g-32P]ATP (110 TBq mmol-1). After incubation at23°C for 5 min, the enzymatic reaction wasstopped by adding one volume of SDS-samplebuffer. Protein samples were finally boiled for 3min and subjected to SDS-PAGE.

L. Bo6et et al. / Plant Science 128 (1997) 169–180 171
2.4. Phosphorylation of intact chloroplasts in thepresence of [ 32P]Pi
The assay medium and experimental conditionsused for generating chloroplast protein phospho-rylation were adapted from Foyer [13] andSundby et al. [22].
After centrifugation (2000×g for 3 min), pel-leted intact chloroplasts (3 mg Chl) were resus-pended in 10 ml of the following phosphorylationmedium (PMa): 50 mM Mops-NaOH pH 7.6, 1mM NaHCO3, 5 mM MgCl2, 0.33 M sorbitol.The phosphorylation was started by adding 37MBq of [32P]Pi orthophosphate and performed at23°C for 10 min under white light (12 000 lux) andgentle mixing. After incubation, 2 ml of unla-belled intact chloroplasts (1.5 mg Chl ml−1) wereadded as carrier to the 10 ml phosphorylated onesand the mixture was immediately laid on a Percollmedium (40% (v/v) Percoll in PMa supplementedwith 20 mM Na2MoO4 and 30 mM NaF) andcentrifuged at 2700×g for 20 min at 4°C. Thepellet was then resuspended in 12 ml of lysismedium (LMa) containing 10 mM Tricine-NaOHpH 7.6, 5 mM MgCl2, 20 mM Na2MoO4 and 30mM NaF. After incubation at 4°C for 10 min, theisoosmolarity of the medium was restored byadding 1.4 ml of 3 M sorbitol and the lysedchloroplasts centrifuged at 20 000×g for 30 minto isolate envelope membranes and stroma (super-natant) from thylakoid membranes (pellet). Theseparation of envelope membranes from stromawas then achieved under standard procedure, withthe exception that all buffers, including the dis-continuous sucrose gradient, were supplementedwith 20 mM Na2MoO4 and 30 mM NaF asprotein phosphatase inhibitors.
2.5. Phosphorylation of intact chloroplasts in thepresence of [g-32P]ATP
After centrifugation (2000×g for 3 min), pel-leted intact chloroplasts (4 mg Chl) were resus-pended in 2 ml of the following phosphorylationmedium (PMb): 50 mM Mops-KOH pH 7.6, 5mM MgCl2, 0.33 M sorbitol. The phosphoryla-tion was started by adding 50 nM [g-32P]ATP (110TBq mmol−1) and 10 m l of 1 mM ATP under dim
illumination condition. After incubation at 23°Cfor 10 min, the chloroplasts were washed threetimes in PMb supplemented with 20 mMNa2MoO4 and 30 mM NaF, and pelleted at5800×g for 30 s. Intact chloroplasts were lysed ina buffer (LMb) containing 10 mM Tricine-NaOHpH 7.6, 0.8 mM MgCl2, 20 mM Na2MoO4 and 30mM NaF and the thylakoids, stromal and envel-ope fractions were isolated under standard proce-dure.
2.6. Gel electrophoresis and autoradiography
SDS-PAGE was performed according to themethod of Laemmli [23]. The relative molecularmasses of polypeptides separated by SDS-PAGEwere estimated according to the electrophoreticmobilities of the following markers (R): phospho-rylase b (94 kD), BSA (67 kD), ovalbumin (43kD), carbonic anhydrase (30 kD), soybean trypsininhibitor (20.1 kD), a-lactalbumin (14.4 kD). Thegels were stained with Coomassie brilliant blue(Fluka, Buchs, CH) and the labelled polypeptidesdetected by autoradiography using X-OMAT ARfilms (Kodak, Lausanne, CH). In some cases, 32Pincorporation into the polypeptides was deter-mined by the Cerenkov procedure (BETAmaticliquid scintillation counter; Kontron, Zurich, CH)after excision of the labelled bands from the gelsor by quantitative densitometry of the autoradio-grams with the Millipore BioImage System(BioImage, Paris, France).
3. Results
3.1. Protein phosphorylation in thylakoid,en6elope and stromal fractions in the presence of[g-32P]ATP
Several polypeptides were phosphorylated afterincubation of thylakoid membrane, envelopemembrane and stromal fractions in the presence[g-32P]ATP (Fig. 1). In thylakoids, the phosphory-lated polypeptide profile after SDS-PAGE (Fig.1(B), lane T) is comparable to that reported else-where [24]. In this case, thylakoid membraneswere rapidly isolated without extensive washing to

L. Bo6et et al. / Plant Science 128 (1997) 169–180172
remove the excess of Rubisco (Fig. 1(A), lane T).In envelope membranes, five polypeptides ofabout 67, 51, 26, 15 and 14 kD were the majorphosphorylated substrates (Fig. 1(B), lane E), the67 kD phosphopolypeptide being also present inthe stroma fraction (Fig. 1(B), lane S). The iden-tity of three phosphorylated protein substrateshave been recently elucidated: the 26 and 14 kDphosphorylated bands are phosphorylated LHCIIand Rubisco small subunit pools, respectively [8],whereas the 67 kD phosphoprotein is a phospho-glucomutase [25]. Concerning the 51 and 15 kDenvelope phosphopolypeptides, only little infor-mation is available: the 51 kD one might beinvolved in the protein import process intochloroplasts [11] while the 15 kD one is constitu-tive of the outer envelope membrane [3]. It has tobe pointed out that an unidentified 94 kD stromalphosphopolypeptide often appeared on other au-toradiograms [3].
3.2. Protein phosphorylation of intact chloroplastsin the presence of [g-32P]ATP and isolation ofthylakoid, en6elope and stromal fractions
Envelope membrane proteins also underwentphosphorylation when intact chloroplasts were in-cubated with [g-32P]ATP. However, the profile ofthe generated envelope phosphopolypeptides wasquite different (compare Fig. 1(B) with Fig. 2(B)).Under these conditions, the major phosphorylatedenvelope proteins exhibited Mr of about 60, 56,51, 47 and 39 kD (Fig. 2(B)), as already reportedby Hinz and Flugge [11]. Interestingly, a same 51kD polypeptide was also phosphorylated in vitro(Fig. 1), as attested by 2D gels (IEF×SDS-PAGE) analyses (data not shown). As shown inFig. 2(B), this envelope phosphorylated polypep-tide slightly contaminated the thylakoid fraction,despite the good-partitioning of the three chloro-plast compartments (Fig. 2(A)). In fact, this is dueto the absence of extensive thylakoid washings toremove bound envelope membranes [16]. Con-cerning the chloroplast soluble phosphorylatedproteins, only two labelled bands having Mr of 94and 67 kD were slightly phosphorylated underthese conditions (Fig. 2(B)). In thylakoids, LHCII(particularly the form b) was phosphorylated.This is not surprising since we observed thatLHCIIb and LHCIIa incorporated labelled phos-phate in intact chloroplasts after incubation withcarrier free [g-32P]ATP (data not shown). Thesedata indicate that small amounts of exogenousATP have the ability to quickly reach the stromacompartment, probably by counterexchange withADP, Pi or PEP through the chloroplast ATP/ADP translocator [26].
3.3. Protein phosphorylation of intact chloroplastsin the presence of [ 32P]Pi orthophosphate
Several chloroplast proteins incorporated la-belled phosphate when intact chloroplasts wereincubated in the presence of light, NaHCO3 and[32P]Pi orthophosphate (Fig. 3(B)). Under thesephosphorylation conditions, the major phospho-polypeptides exhibited Mr of about 94, 67, 60, 36,28 (LHCIIa), 26 (LHCIIb), 20, 15, 14 and 12 kD.Some of these phosphorylated bands have already
Fig. 1. In vitro protein phosphorylation of the three majorchloroplast compartments in the presence of [g-32P]ATP.Proteins (30 mg) of thylakoids (T), envelope vesicles (E) andstroma (S) were separated by 13% SDS-PAGE, as well as Mr
protein markers (R). The gel was stained by Coomassie blue(A) and revealed by autoradiography (B). The position of themajor envelope labelled polypeptides is indicated on the rightside of the gel.

L. Bo6et et al. / Plant Science 128 (1997) 169–180 173
Fig. 2. In organello protein phosphorylation in the presence of [g-32P]ATP. Proteins (30 mg) of thylakoids (T), stroma (S) andenvelopes (E) were separated by 13% SDS-PAGE after isolation from lysed chloroplasts, as well as Mr protein markers (R). The gelwas stained by Coomassie blue (A) and revealed by autoradiography (B). The position of the major labelled polypeptides is indicatedon the right side of the gel.
been described [5,12]. By analogy with their elec-trophoretic mobilities, some of these phosphory-lated polypeptides, including LHCIIa and b [14],could be identified and likely corresponded tophosphoglucomutase (67 kD, [6]), D1 or/and D2(36 kD, [12]), and cytochrome b6 (20 kD, [27]).The other phosphoproteins remain to be iden-tified. Unexpectedly, in darkness, the 32P incorpo-ration also occured in the two 67 and 94 kDpolypeptides (Fig. 3(A)) and to a much lesserdegree in LHCIIb, this latter labelling being dueto low light diffusion during the sample collection(Eppendorf opening). No reports published todate mentioned that two polypeptides of 94 and67 kD are dark-phosphorylated in organello. Forall phosphopolypeptides, the 32P incorporation
was complete after 5 min (Fig. 3(A) and (B)) andended after 10 min incubation, as determined byquantitative densitometry or Cerenkov counting(data not shown). These experiments fixed thedark and light conditions under which chloroplastlabelling occurs.
As inorganic pyrophosphate can function as aphosphate donor, like ATP, in the phosphoryla-tion process of thylakoid proteins [28], the effectof 3 mM sodium pyrophosphate was tested on thein organello chloroplast protein phosphorylationin the presence of [32P]Pi. A labelling decrease,varying from 70 to 80%, of the total 32P incorpo-rated in chloroplast proteins was observed in thelight and darkness (data not shown), indicatingthat PPi competes with ATP as phosphate donor.

L. Bo6et et al. / Plant Science 128 (1997) 169–180174
Fig. 3. Protein phosphorylation of intact chloroplasts in the presence of [32P]Pi orthophosphate. Intact chloroplasts (0.3 mg ml−1
Chl) were incubated in the presence of 7.4 MBq ml−1 [32P]Pi orthophosphate under dark (A) or light (B) conditions. Totalchloroplast proteins corresponding to 3 mg Chl were sampled after 15, 30, 60, 300 and 600 s, separated by 13% SDS-PAGE, andthen subjected to autoradiography. The position of the major labelled polypeptides is indicated on the right side of the gel.
These results could be explained by the presenceof a plastidic pyrophosphatase whose activityfeeds the chloroplast with unlabelled inorganicphosphate [29] resulting in a decrease of the 32Pincorporation into proteins.
The phosphate import into chloroplasts can beimpeded by 4,4%-diisothiocianato-stilbene-2,2%-disulfonate (DIDS) acting as an effective inhibitorof the phosphate translocator [30]. Rumpho et al.[31] showed that DIDS was able to bind itself tothe envelope phosphate translocator of mesophyllchloroplasts from C3 and C4 plants. The effect ofDIDS was tested on the 32P incorporation inchloroplast proteins after preincubation of intactchloroplasts with 5 mM DIDS for 8 min and inorganello labelling with [32P]Pi (data not shown).Chloroplast protein phosphorylation wasmarkedly inhibited by DIDS indicating that a lowconcentration of the inhibitor was sufficient toreduce the pool of chloroplastic [g-32P]ATP byhindering the phosphate translocation intochloroplasts. A Cerenkov counting of the phos-
phorylated bands revealed that a more pro-nounced DIDS-inhibitory effect was observed onthe LHCIIb subpopulation of stroma lamellae(47% of inhibition) compared to the LHCIIagrana form (38% of inhibition), after 5 min la-belling. But this is not surprising since LHCIIb isknown to be more rapidly phosphorylated thanLHCIIa, under similar experimental conditions[32].
3.4. Protein phosphorylation of intact chloroplastsin the presence of [ 32P]Pi orthophosphate andisolation of thylakoid, en6elope and stromalfractions
The phosphorylated polypeptides constitutiveof the three major chloroplast compartments wereanalysed by SDS-PAGE after phosphorylation inorganello in the presence of [32P]Pi. TheCoomassie blue staining of the gel revealed, withthe exception of the Rubisco large and smallsubunits in envelopes and stroma [18], that en-
L. Bo6et et al. / Plant Science 128 (1997) 169–180 175
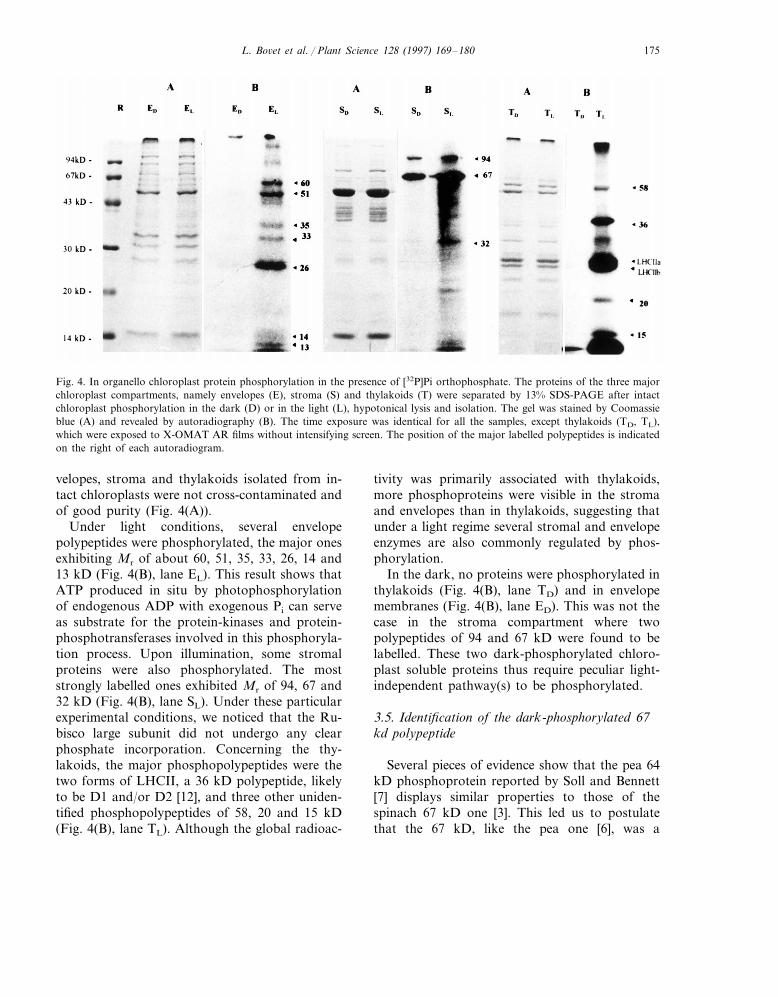
Fig. 4. In organello chloroplast protein phosphorylation in the presence of [32P]Pi orthophosphate. The proteins of the three majorchloroplast compartments, namely envelopes (E), stroma (S) and thylakoids (T) were separated by 13% SDS-PAGE after intactchloroplast phosphorylation in the dark (D) or in the light (L), hypotonical lysis and isolation. The gel was stained by Coomassieblue (A) and revealed by autoradiography (B). The time exposure was identical for all the samples, except thylakoids (TD, TL),which were exposed to X-OMAT AR films without intensifying screen. The position of the major labelled polypeptides is indicatedon the right of each autoradiogram.
velopes, stroma and thylakoids isolated from in-tact chloroplasts were not cross-contaminated andof good purity (Fig. 4(A)).
Under light conditions, several envelopepolypeptides were phosphorylated, the major onesexhibiting Mr of about 60, 51, 35, 33, 26, 14 and13 kD (Fig. 4(B), lane EL). This result shows thatATP produced in situ by photophosphorylationof endogenous ADP with exogenous Pi can serveas substrate for the protein-kinases and protein-phosphotransferases involved in this phosphoryla-tion process. Upon illumination, some stromalproteins were also phosphorylated. The moststrongly labelled ones exhibited Mr of 94, 67 and32 kD (Fig. 4(B), lane SL). Under these particularexperimental conditions, we noticed that the Ru-bisco large subunit did not undergo any clearphosphate incorporation. Concerning the thy-lakoids, the major phosphopolypeptides were thetwo forms of LHCII, a 36 kD polypeptide, likelyto be D1 and/or D2 [12], and three other uniden-tified phosphopolypeptides of 58, 20 and 15 kD(Fig. 4(B), lane TL). Although the global radioac-
tivity was primarily associated with thylakoids,more phosphoproteins were visible in the stromaand envelopes than in thylakoids, suggesting thatunder a light regime several stromal and envelopeenzymes are also commonly regulated by phos-phorylation.
In the dark, no proteins were phosphorylated inthylakoids (Fig. 4(B), lane TD) and in envelopemembranes (Fig. 4(B), lane ED). This was not thecase in the stroma compartment where twopolypeptides of 94 and 67 kD were found to belabelled. These two dark-phosphorylated chloro-plast soluble proteins thus require peculiar light-independent pathway(s) to be phosphorylated.
3.5. Identification of the dark-phosphorylated 67kd polypeptide
Several pieces of evidence show that the pea 64kD phosphoprotein reported by Soll and Bennett[7] displays similar properties to those of thespinach 67 kD one [3]. This led us to postulatethat the 67 kD, like the pea one [6], was a

L. Bo6et et al. / Plant Science 128 (1997) 169–180176
phosphoglucomutase [3]. In order to substantiatethis hypothesis, intact chloroplasts were incubatedin the dark with [32P]Pi in the presence of glucose1-P or glucose 6-P (Fig. 5). The addition of phos-phorylated glucose to the incubation medium pre-vented almost completely the phosphateincorporation in the 67 kD polypeptide withoutaffecting labelling in the 94 kD one. This indicatesthat glucose 1-P and glucose 6-P are competitivesubstrates for the 67 kD protein, and that, invivo, its dark-phosphorylation likely occurred viasuch labelled sugar phosphate and not directlywith [32P]Pi orthophosphate. Similar results wereobtained when stromal proteins, isolated fromintact chloroplasts phosphorylated in the presenceof [32P]Pi and light, were post-incubated with glu-cose 1-P or glucose 6-P (data not shown). Fromthese experiments we conclude that the 67 kD
Fig. 6. Effect of DIDS on the dark-phosphorylation of the 67kD protein. Intact chloroplasts (0.3 mg ml−1 Chl) were phos-phorylated in the dark with 7.4 MBq ml−1 [32P]Pi orthophos-phate, after preincubation with (�) or without (�) 5 mMDIDS for 8 min. Chloroplast proteins corresponding to 3 mgChl were collected after 1, 5, 10 and 15 min, separated by 13%SDS-PAGE and subjected to autoradiography. 32P incorpora-tion into the 67 kD polypeptide was determined by quantita-tive densitometry (IOD: integrated optic density).
Fig. 5. Effects of glucose 1-P (Glu 1-P) and glucose 6-P (Glu6-P) on the dark-phosphorylation of the 94 kD and 67 kDpolypeptides. Intact chloroplasts (0.3 mg ml−1 Chl) werephosphorylated in the dark with 7.4 MBq ml−1 [32P]Pi or-thophosphate, in the absence or presence of 100 mM Glu 1-Por Glu 6-P. Chloroplast proteins corresponding to 3 mg Chlwere collected after 1, 5 and 10 min, separated by 13%SDS-PAGE and subjected to autoradiography. 32P incorpora-tion into the polypeptides was determined by quantitativedensitometry (IOD: integrated optic density) of the autoradio-gram bands (upper panel).
phosphorylated protein is a phosphoglucomutase(PGM), as suggested earlier [3].
The effect of the phosphate translocator in-hibitor DIDS was analysed on the dark[32P]phosphate incorporation of PGM (Fig. 6). Inthe presence of 5 mM DIDS, the PGM labellingdecreased markedly, indicating that blocking[32P]Pi import into chloroplasts prevented itsphosphorylation in the dark. This observationwas also valid for the 94 kD phosphopolypeptide(data not shown). Interestingly, the extent ofDIDS inhibitory effect on PGM labelling wassimilar to that observed on LHCII in the light(data not shown). Thus, this result may indicatethat the dark-phosphorylated substrate(s) forPGM is probably generated in the stroma and isnot directly provided by [32P]Pi, which cannotserve as a substrate for PGM [33], or by [g-32P]ATP.
Two additional experiments were conducted toshow the involvement of internal ATP on chloro-plast protein phosphorylation in the dark. In afirst set of experiments, ATP was generated inside

L. Bo6et et al. / Plant Science 128 (1997) 169–180 177
the chloroplasts (0.3 mg ml−1 Chl) in the dark bythe addition of 1 mM dihydroxyacetone-P, 1 mMoxaloacetate and 0, 0.1, 1 and 10 mM PPi, respec-tively [34]. Sodium pyrophosphate was added inthe dark at 23°C, exactly 2 min before the addi-tion of 5 MBq ml−1 [32P]Pi orthophosphate, tosupply chloroplasts with the inorganic phosphateresulting from the activity of endogenous py-rophosphatases [29]. After 5 min incorporation,samples (1.8 mg Chl) were collected, separated bySDS-PAGE and subjected to autoradiography.Under these conditions, LHCII labelling was notstimulated, indicating that [g-32P]ATP possiblygenerated in the stroma was not used as substrateby LHCII kinase (data not shown). On the otherhand, increasing concentration of pyrophosphatemarkedly reduced 32P incorporation in PGM andthe 94 kD polypeptide (87 and 60% of the controlin the presence of 10 mM PPi, data not shown). Ina second set of experiments, stromal ATP levelswere lowered by the addition of increasing con-centrations (0, 1, 5 and 10 mM) of glycerate, asATP trap [34]. Glycerate was added to the chloro-plasts (0.3 mg ml−1 Chl) in the dark at 23°C,exactly 2 min before the addition of 5 MBq ml−1
[32P]Pi orthophosphate. After 5 min incorpora-tion, samples (1.8 mg Chl) were collected, sepa-rated by SDS-PAGE and subjected toautoradiography. Increasing concentration ofglycerate strongly reduced 32P incorporation inPGM and the 94 kD polypeptide (73 and 82% ofthe control in the presence of 10 mM glycerate,data not shown). These results support the ideathat the intrachloroplastic dark-ATP pool is notthe substrate for phosphorylating PGM and the94 kD polypeptide, although the stromal ATPsynthesis and hydrolysis in the dark largely affects32P incorporation into both proteins.
4. Discussion
The aim of this investigation was to comparethe rate and extent of envelope protein phospho-rylation, under three different conditions of 32Pincorporation, i.e. in vitro (Fig. 1) and in or-ganello in the presence of [g-32P]ATP (Fig. 2), orin organello in the presence of light and [32P]Pi
(Fig. 4).
When intact chloroplasts were phosphorylatedin the presence of exogenous [g-32P]ATP (Fig. 2),the protein labelling in the three major chloro-plast compartments was significantly reducedcompared to that observed under in vitro condi-tions (Fig. 1). This might be due to: (1) a rela-tively low import of labelled ATP intochloroplasts [35]; (2) the presence of 2 mM unla-belled ATP in the assay medium [3]; and/or (3)some intrachloroplastic protein phosphatase ac-tivities [5,36], despite the presence of proteinphosphatase inhibitors (Fig. 2(B)). In envelopes,only the 51 kD polypeptide was strongly labelledunder in organello conditions (Fig. 2), comparedto the polypeptides phosphorylated under in vitroconditions (Fig. 1). This rather simple envelopephosphopolypeptide pattern cannot be readily as-cribed to the sole addition of micromolar unla-belled ATP, since concentrations of MgATP orATP up to 100 mM stimulate the in vitro incorpo-ration of labelled phosphate into several envelopepolypeptides [3,25]. This suggests that envelopeprotein-kinases and protein-phosphotransferases,and/or substrates are preferentially connected ei-ther with the cytoplasmic or the chloroplasticATP source to allow protein phosphorylation.
In this investigation, we found that not onlythylakoid and stromal proteins [5,13], but alsoenvelope polypeptides, can incorporate [32P]phos-phate when intact chloroplasts are incubated inthe presence of [32P]Pi and light (Fig. 4). Thisshows that ATP generated by photophosphoryla-tion may be a substrate source for envelopeprotein phosphorylation. Interestingly, seven en-velope polypeptides of about 60, 51, 35, 33, 26, 14and 13 kD, which were phosphorylated in or-ganello (Fig. 4), were also found when isolatedinner envelope vesicles were incubated in the pres-ence of [g-32P]ATP [3,25]. This observation sug-gests that most of these envelope phospho-polypeptides are probably constitutive of the in-ner envelope membrane. Moreover, the majorityof the protein-kinases involved in the phosphory-lation process should be located in the vicinity ofthe inner face of the inner envelope membrane inorder to utilize more rapidly and easily the ATPproduced by thylakoids. Concerning the phospho-polypeptide of 67 kD which is strongly labelled in

L. Bo6et et al. / Plant Science 128 (1997) 169–180178
envelope vesicles (Fig. 1), it was not present in inorganello phosphorylated envelopes, but exclu-sively in the stroma (Figs. 2 and 4). This isprobably due to the peculiar hydrophilic characterof its phosphorylated form [3].
The fact that in organello protein phosphoryla-tion in the presence of [32P]Pi was strongly im-peded by the absence of light (Fig. 3) or presenceof DIDS (data not shown), confirms that 32Pincorporation into intact chloroplast proteins isessentially driven by light-induced ATP produc-tion [14]. Nevertheless, an unexpected phosphory-lation of two soluble proteins exhibiting apparentMr of about 94 and 67 kD also occured in thedark. Lowering as well as raising the internal ATPconcentration in the dark strongly reduced 32Pincorporation in PGM and the 94 kD polypeptide(data not shown), thereby excluding the directinvolvement of stromal ATP in the phosphoryla-tion process. This indicates that both polypeptidesutilize dark-phosphorylation pathways, indepen-dent of the photophosphorylation. In contrast tothe 94 kD phosphoprotein (Fig. 5), the 67 kD oneunderwent a severe and specific inhibition withglucose 1-P or glucose 6-P. One can thereforeconsider this protein to be a phosphoglucomutase.In addition, the comparison of the proteolyticdigestion patterns of the 67 kD phosphopolypep-tide obtained after in organello or in vitro la-belling revealed that all of the resulting fragmentswere identical (data not shown). The effect ofDIDS (Fig. 6) and sugar phosphates (Fig. 5) bothsuggest that the dark-phosphorylated substratefor PGM might be [32P]glucose 1-P, which origi-nates from the starch degradation catalysed bythe stromal starch phosphorylase in the dark [37].In our experimental conditions, however, we haveno indication that unlabelled hexose-phosphatesmight be taken up by the chloroplast, althoughQuick et al. [38] have shown recently that au-totrophic chloroplasts isolated from spinach maybe fed by glucose 6-P, thus leaving open thequestion about the intraorganelle location of thechloroplastic PGM [3,7]. Nevertheless, our resultssuggest that the chloroplastic PGM may directlyuse [g-32P]ATP as substrate in the light and an-other phosphorylating substrate (most likely[32P]glucose 1-P) in darkness. Consequently, one
of the main physiological role for PGM in thechloroplast is probably to mute the phosphatefrom the 1 to the 6 position of glucose, in order tosupply the chloroplast with a catabolic carbonenergy source in the absence of light. The identifi-cation of the 94 kD dark-phosphorylated hy-drophilic polypeptide is now under investigationin our laboratory.
This work brings new insight into the study ofprotein phosphorylation in intact chloroplastswith regard to the possible cooperation betweencytoplasmic and stromal ATP pools. Exogenousstresses, drugs, hormones and signals, susceptibleto affect chloroplast biogenesis and developmentcould now also be tested to measure their possibleeffect on chloroplast (envelope) protein phospho-rylation. In this respect, it might be possible thatthe envelope protein phosphorylation process isnecessary for protecting membrane proteinsagainst degradation by light-stress, as for the thy-lakoid D1 protein [39].
Acknowledgements
This work was supported by the Swiss NationalScience Foundation (grants 3100.33693.92 and3100.043297.95 to P.A.S and L.B). The authorsthank Nathalie Simon-Vermot and Marlyse Mey-lan-Bettex for technical assistance and Dr SamuelRoulin (University of Bern) for critical reading ofthe manuscript.
References
[1] R. Douce, J. Joyard, Biochemistry and function of theplastid envelope, Annu. Rev. Cell Biol. 6 (1990) 173–216.
[2] J. Joyard, M.A. Block, R. Douce, Molecular aspects ofplastid envelope biochemistry, Eur. J. Biochem. 199(1991) 489–509.
[3] P.A. Siegenthaler, L. Bovet, A unique protein-kinase ac-tivity is responsible for the phosphorylation of the 26 and14 kDa proteins but not of the 67 kDa protein in thechloroplast envelope membranes of spinach, Planta 190(1993) 231–240.
[4] J. Soll, I. Fisher, K. Keegstra, A guanosine 5%-triphos-phate-dependent protein kinase is localized in the outerenvelope membrane of pea chloroplasts, Planta 176(1988) 488–496.

L. Bo6et et al. / Plant Science 128 (1997) 169–180 179
[5] J. Bennett, Protein phosphorylation in green plant chloro-plasts, Annu. Rev. Plant Physiol. 42 (1991) 281–311.
[6] M.E. Salvucci, R.R. Drake, K.P. Broadbent, B.E. Haley,K.R. Hanson, N.A. McHale, Identification of the 64 kDachloroplast stromal phosphoprotein as phosphoglucomu-tase, Plant Physiol. 93 (1990) 105–109.
[7] J. Soll, J. Bennett, Localization of a 64-kDa phospho-protein in the lumen between the outer and the innerenvelopes of pea chloroplasts, Eur. J. Biochem. 175(1988) 301–307.
[8] L. Bovet, M.O. Muller, P.A. Siegenthaler, The 26- and14-kDa phosphoproteins associated with spinach chloro-plast envelope membranes are distinct membrane-boundpools of the light-harvesting complex of photosystem IIand of the small subunit of the ribulose-1,5-bisphosphatecarboxylase-oxygenase, Planta 195 (1995) 563–569.
[9] J. Soll, Purification and characterization of a chloroplastouter envelope-bound, ATP-dependent protein kinase,Plant Physiol. 87 (1988) 898–903.
[10] K. Waegmann, J. Soll, Phosphorylation of the transitsequence of chloroplast precursor proteins, J. Biol. Chem.(1996) 6545–6554.
[11] G. Hinz, U.I. Flugge, Phosphorylation of a 51-kDa envel-ope membrane polypeptide involved in protein transloca-tion into chloroplasts, Eur. J. Biochem. 175 (1988)649–659.
[12] J. Bennett, Chloroplast protein phosphorylation and theregulation of photosynthesis, Physiol. Plant. 60 (1984)583–590.
[13] C.H. Foyer, Stromal protein phosphorylation in spinach(Spinacia oleracea) chloroplasts, Biochem. J. 231 (1985)97–103.
[14] J.F. Allen, Protein phosphorylation in regulation of pho-tosynthesis, Biochim. Biophys. Acta 1098 (1992) 275–335.
[15] G. Mourioux, R. Douce, Slow passive diffusion of or-thophosphate between intact isolated chloroplasts andsuspending medium, Plant Physiol. 67 (1981) 470–473.
[16] A. Rawyler, M. Meylan, P.A. Siegenthaler, Galactolipidexport from envelope to thylakoid membranes in intactchloroplasts. I. Characterization and involvement in thy-lakoid lipid asymmetry, Biochim. Biophys. Acta 1104(1992) 331–334.
[17] R. Douce, J. Joyard, Purification of the chloroplast envel-ope, in: M. Edelman, R.B. Hallick, N.H. Chua (Eds.),Methods in Chloroplast Molecular Biology, Elsevier, Am-sterdam, 1982, pp. 239–259.
[18] P.A. Siegenthaler, N. Dumont, Characteristics of spinachchloroplast envelope, thylakoid and stroma polypeptidesas revealed by triton X-114 phase partition, Plant CellPhysiol. 31 (1990) 1101–1108.
[19] J. Bruinsma, A comment on spectrophotometric determi-nation of chlorophyll, Biochim. Biophys. Acta 53 (1961)576–578.
[20] M.M. Bradford, A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of protein-dye binding, Anal. Biochem. 72(1976) 248–254.
[21] O.H. Lowry, N.J. Rosenbrough, N.L. Farr, R.J. Randal,Protein measurement with Folin phenol reagent, J. Biol.Chem. 193 (1951) 265–275.
[22] C. Sundby, U.K. Larsson, T. Henrysson, Effects of bicar-bonate on thylakoid protein phosphorylation, Biochim.Biophys. Acta 975 (1989) 277–282.
[23] U.K. Laemmli, Cleavage of structural proteins during theassembly of the head of bacteriophage T4, Nature 227(1970) 680–685.
[24] J. Bennett, E.K. Shaw, S. Bakr, Phosphorylation of thy-lakoid proteins and synthetic peptide analogs, FEBS Lett.210 (1987) 22–26.
[25] L. Bovet, Phosphorylation des proteines de l’enveloppedu chloroplaste d’Epinard et role possible des proteines‘heat-shock’ comme catalyseurs d’activites proteine-ki-nasiques, PhD Thesis, Universite de Neuchatel, Neuchatel(1993), Switzerland.
[26] U.I. Flugge, H.W. Heldt, Metabolite translocators of thechloroplast envelope, Annu. Rev. Plant Physiol. 42 (1991)129–144.
[27] A. Gal, R.G. Hermann, F. Lottspeich, I. Ohad, Phospho-rylation of cytochrome b6 by the LHCII kinase associatedwith cytochrome complex, FEBS Lett. 298 (1992) 33–35.
[28] A.M. Pramanik, S. Bingsmark, M. Lindahl, H.Baltscheffsky, M. Baltscheffsky, B. Andersson, Inorganic-pyrophosphate-dependent phosphorylation of spinachthylakoid proteins, Eur. J. biochem. 198 (1991) 183–186.
[29] H. Baltscheffsky, A.M. Pramanik, M. Lundin, B. An-dersson, Distribution of inorganic pyrophosphatase(PPase) activity in chloroplasts, in: M. Baltscheffsky(Ed.), Current Research in Photosynthesis, vol. 3,Kluwer, Dordrecht, 1990, pp. 201–204.
[30] A. Gross, G. Bruckner, H.W. Heldt, U.I. Flugge, Com-parison of the kinetic properties, inhibition and labellingof the phosphate translocators from maize and spinachmesophyll chloroplasts, Planta 180 (1990) 262–271.
[31] M.E. Rumpho, G.E. Edwards, A.E. Yousif, K. Keegstra,Specific labeling of the phosphate translocator in C3 andC4 mesophyll chloroplasts by tritiated dihydro-DIDS(1,2-ditritio-1,2-[2,2%-disulfo-4,4%-diisothiocyano]diphenylthane), Plant Physiol. 86 (1988) 1193–1198.
[32] U.K. Larsson, C. Sundby, B. Andersson, Characteriza-tion of two different subpopulations of spinach light-har-vesting chlorophyll a/b-protein complex (LHCII):polypeptide composition, phosphorylation pattern andassociation with photosystem II, Biochim. Biophys. Acta894 (1987) 59–68.
[33] W.J. Ray, E.J. Peck, Phosphomutases, in: P.D. Boyer(Ed.), The Enzymes, vol. 6, Academic press, New York,1972, pp. 407–477.
[34] L.J. Olsen, S.M. Theg, B.R. Selman, K. Keegstra, ATP isrequired for the binding of precursor proteins to chloro-plasts, J. Biol. Chem. 264 (1989) 6724–6729.
[35] S.P. Robinson, J.T. Wiskich, Pyrophosphate inhibition ofcarbon dioxide fixation in isolated pea chloroplasts byuptake in exchange for endogenous adenine nucleotides,Plant Physiol. 59 (1977) 422–427.

L. Bo6et et al. / Plant Science 128 (1997) 169–180180
[36] H. Michel, E.K. Shaw, J. Bennett, Protein kinase andphosphatase activities of thylakoid membranes, in: C.J.Leaver, H. Sze (Eds.), Plant Membranes: Structure, Func-tion, Biogenesis, Liss, New York, 1987, pp. 85–102.
[37] E. Beck, P. Ziegler, Biosynthesis and degradation ofstarch in higher plants, Annu. Rev. Plant Physiol. 40(1989) 95–117.
[38] W.P. Quick, R. Scheibe, H.E. Neuhaus, Induction ofhexose-phosphate translocator activity in spinach chloro-plasts, Plant Physiol. 109 (1995) 113–121.
[39] E. Rintamaki, R. Salo, A. Koivuniemi, E.M. Aro, Proteinphosphorylation and magnesium status regulate thedegradation of the D1 reaction centre protein of photo-system II, Plant Sci. 115 (1996) 175–182.
.
Copyright © 2022 FDOKUMEN




















