
Impacts of Macondo oil from Deepwater Horizon spill on the growth response of the common reed...
8
Impacts of Macondo oil from Deepwater Horizon spill on the growth response of the common reed Phragmites australis: A mesocosm study Chad R. Judy a , Sean A. Graham b , Qianxin Lin b , Aixin Hou a , Irving A. Mendelssohn b,⇑ a Department of Environmental Sciences, Louisiana State University, Baton Rouge, LA 70803, USA b Department of Oceanography and Coastal Sciences, Louisiana State University, Baton Rouge, LA 70803, USA article info Keywords: Deepwater Horizon Macondo oil Oil spill Phragmites australis Growth response abstract We investigated impacts of Macondo MC252 oil from the Deepwater Horizon (DWH) spill on the common reed Phragmites australis (Cav.) Trin. ex Steud., a dominant species of the Mississippi River Delta. In green- house experiments, we simulated the most common DWH oiling scenarios by applying weathered and emulsified Macondo oil to aboveground shoots at varying degrees of coverage (0–100%) or directly to marsh soil at different dosages (0–16 L m 2 ). P. australis exhibited strong resistance to negative impacts when oil was applied to shoots alone, while reductions in above- and belowground plant growth were apparent when oil was applied to the soil or with repeated shoot-oiling. Although soil-oiling compro- mised plant function, mortality of P. australis did not occur. Our results demonstrate that P. australis has a high tolerance to weathered and emulsified Macondo oil, and that mode of exposure (aboveground versus belowground) was a primary determinant of impact severity. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction On April 20, 2010 the Deepwater Horizon (DWH) oil drilling plat- form exploded and released over the next 3 months a government- estimated 4.9 million barrels of oil (Macondo MC252), becoming the largest marine oil spill in United States history (National Commission on the BP Deepwater Horizon Oil Spill, 2011; Camilli et al., 2011). The sheer volume of oil caused widespread impacts including the oiling of approximately 796 km of Gulf Coast marshes; this included approximately 304 km of Louisiana marshes classified as moderately to heavily oiled (Michel et al., 2013, Supplementary materials). Although a majority of the im- pacted shoreline was salt marsh dominated by Spartina alterniflora, Juncus roemerianus, and Avicennia germinans, oiling of the Birdfoot Delta at the mouth of the Mississippi River impacted a sizable area of fresh and oligohaline marsh dominated by the common reed, Phragmites australis (P. australis (Cav.) Trin. ex Steud.). P. australis is a valuable species in south Louisiana because of its exceptional ability to accumulate and stabilize sediments as well as being adaptable to a wide variety of environmental conditions (Alizai and McManus, 1980; Hartog et al., 1989; Gorai et al., 2011). Also, the P. australis australis marshes in the Birdfoot Delta of the Missis- sippi River are unique in the United States in forming expansive deltaic wetlands similar to the P. australis-dominated deltas of the Volga and Danube Rivers in Europe. Although a relatively large body of literature has described the impact and recovery of a number of wetland species to oiling (e.g., Pezeshki et al., 2000; DeLaune et al., 2003; Mendelssohn et al., 2012), P. australis australis has received much less attention. Spe- cies such as S. alterniflora, Spartina patens, and J. roemerianus have been negatively impacted by oil exposure to soil and to above- ground tissues; these impacts range from reduced photosynthesis, stem density, and biomass to complete mortality (Lin and Mendelssohn, 1996; Pezeshki et al., 2000; Dowty et al., 2001; Anderson and Hess, 2012). However, not all wetland species are negatively impacted by oiling. For example, when Sagittaria lanci- folia, a dominant freshwater species in the southeastern United States, was exposed to soil oiling, regrowth was stimulated up to an oil dosage of 24 L m 2 (Lin and Mendelssohn, 1996). Limited re- search on P. australis after soil oiling has shown a reduction in pho- tosynthetic rates, above- and belowground biomass, and oxygenation of phyllospheres and rhizospheres causing failure of new buds to emerge (Dowty et al., 2001; Armstrong et al., 2009; Nie et al., 2010; Zhu et al., 2013). However, a more comprehensive understanding of the response of P. australis to oiling requires further examination given its demonstrated resilience to various other pollutants as evidenced by its use in wastewater treatment wetlands (Brix, 1987), for sludge stabilization (Cole, 1998; Bianchi et al., 2011; Borin et al., 2011), and for phytoremediation (Toyama et al., 2010, 2011). Impacts from the DWH oil spill have been documented for salt marsh species such as S. alterniflora and J. roemerianus, showing negative impacts at varying degrees of oiling applied to both the 0025-326X/$ - see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.marpolbul.2013.12.046 ⇑ Corresponding author. Tel.: +1 2255786425. E-mail address: [email protected] (I.A. Mendelssohn). Marine Pollution Bulletin 79 (2014) 69–76 Contents lists available at ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul
Transcript of Impacts of Macondo oil from Deepwater Horizon spill on the growth response of the common reed...

Marine Pollution Bulletin 79 (2014) 69–76
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Impacts of Macondo oil from Deepwater Horizon spill on the growthresponse of the common reed Phragmites australis: A mesocosm study
0025-326X/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.marpolbul.2013.12.046
⇑ Corresponding author. Tel.: +1 2255786425.E-mail address: [email protected] (I.A. Mendelssohn).
Chad R. Judy a, Sean A. Graham b, Qianxin Lin b, Aixin Hou a, Irving A. Mendelssohn b,⇑a Department of Environmental Sciences, Louisiana State University, Baton Rouge, LA 70803, USAb Department of Oceanography and Coastal Sciences, Louisiana State University, Baton Rouge, LA 70803, USA
a r t i c l e i n f o
Keywords:Deepwater HorizonMacondo oilOil spillPhragmites australisGrowth response
a b s t r a c t
We investigated impacts of Macondo MC252 oil from the Deepwater Horizon (DWH) spill on the commonreed Phragmites australis (Cav.) Trin. ex Steud., a dominant species of the Mississippi River Delta. In green-house experiments, we simulated the most common DWH oiling scenarios by applying weathered andemulsified Macondo oil to aboveground shoots at varying degrees of coverage (0–100%) or directly tomarsh soil at different dosages (0–16 L m�2). P. australis exhibited strong resistance to negative impactswhen oil was applied to shoots alone, while reductions in above- and belowground plant growth wereapparent when oil was applied to the soil or with repeated shoot-oiling. Although soil-oiling compro-mised plant function, mortality of P. australis did not occur. Our results demonstrate that P. australishas a high tolerance to weathered and emulsified Macondo oil, and that mode of exposure (abovegroundversus belowground) was a primary determinant of impact severity.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
On April 20, 2010 the Deepwater Horizon (DWH) oil drilling plat-form exploded and released over the next 3 months a government-estimated 4.9 million barrels of oil (Macondo MC252), becomingthe largest marine oil spill in United States history (NationalCommission on the BP Deepwater Horizon Oil Spill, 2011; Camilliet al., 2011). The sheer volume of oil caused widespread impactsincluding the oiling of approximately 796 km of Gulf Coastmarshes; this included approximately 304 km of Louisianamarshes classified as moderately to heavily oiled (Michel et al.,2013, Supplementary materials). Although a majority of the im-pacted shoreline was salt marsh dominated by Spartina alterniflora,Juncus roemerianus, and Avicennia germinans, oiling of the BirdfootDelta at the mouth of the Mississippi River impacted a sizable areaof fresh and oligohaline marsh dominated by the common reed,Phragmites australis (P. australis (Cav.) Trin. ex Steud.). P. australisis a valuable species in south Louisiana because of its exceptionalability to accumulate and stabilize sediments as well as beingadaptable to a wide variety of environmental conditions (Alizaiand McManus, 1980; Hartog et al., 1989; Gorai et al., 2011). Also,the P. australis australis marshes in the Birdfoot Delta of the Missis-sippi River are unique in the United States in forming expansivedeltaic wetlands similar to the P. australis-dominated deltas ofthe Volga and Danube Rivers in Europe.
Although a relatively large body of literature has described theimpact and recovery of a number of wetland species to oiling (e.g.,Pezeshki et al., 2000; DeLaune et al., 2003; Mendelssohn et al.,2012), P. australis australis has received much less attention. Spe-cies such as S. alterniflora, Spartina patens, and J. roemerianus havebeen negatively impacted by oil exposure to soil and to above-ground tissues; these impacts range from reduced photosynthesis,stem density, and biomass to complete mortality (Lin andMendelssohn, 1996; Pezeshki et al., 2000; Dowty et al., 2001;Anderson and Hess, 2012). However, not all wetland species arenegatively impacted by oiling. For example, when Sagittaria lanci-folia, a dominant freshwater species in the southeastern UnitedStates, was exposed to soil oiling, regrowth was stimulated up toan oil dosage of 24 L m�2 (Lin and Mendelssohn, 1996). Limited re-search on P. australis after soil oiling has shown a reduction in pho-tosynthetic rates, above- and belowground biomass, andoxygenation of phyllospheres and rhizospheres causing failure ofnew buds to emerge (Dowty et al., 2001; Armstrong et al., 2009;Nie et al., 2010; Zhu et al., 2013). However, a more comprehensiveunderstanding of the response of P. australis to oiling requiresfurther examination given its demonstrated resilience to variousother pollutants as evidenced by its use in wastewater treatmentwetlands (Brix, 1987), for sludge stabilization (Cole, 1998; Bianchiet al., 2011; Borin et al., 2011), and for phytoremediation (Toyamaet al., 2010, 2011).
Impacts from the DWH oil spill have been documented for saltmarsh species such as S. alterniflora and J. roemerianus, showingnegative impacts at varying degrees of oiling applied to both the

70 C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76
aboveground biomass and the soil profile in field and greenhousestudies (Lin and Mendelssohn, 2012; Wu et al., 2012; Biber et al.,2014). Impacts on these species range from minor, when oil is ap-plied to the aboveground biomass at low doses, to severe with theaddition of oil to the soil. In contrast to the documentation of im-pacts on salt marsh species, we are unaware of published researchconcerning the response of P. australis to the DWH oil spill. Due tothe relatively extensive weathering and emulsification of theMacondo oil before it made landfall, toxic components of the oilwere reduced, thereby making for a unique situation comparedto most spills of un-weathered oil (Mendelssohn et al., 2012).The effects of such weathered oil on P. australis require investiga-tion so that information on the sensitivity of this species to oilingcan be applied to cleanup and restoration efforts during futureoil spills in the Gulf of Mexico and elsewhere. In addition, muchof the oil contact occurred aboveground, but with varying degreesof plant coverage, yet the effects of differential aboveground oilinghave received little attention, especially for this species.
The objectives of this study were to (1) quantify the impacts ofweathered and emulsified Macondo oil on P. australis across a widerange of possible above- and belowground oiling scenarios, (2)determine the effects of different dosages of oil on plant growthto soil oiling, and (3) use this information to clarify how oil expo-sure (aboveground versus belowground) controls plant responses.We predict that P. australis growth response will be little affectedfrom oiling of aboveground plant tissues, while oil exposurebelowground will have growth-limiting impacts that increase withincreasing dosage.
2. Materials and methods
2.1. Experiment 1 – effects of common oiling scenarios on P. australisgrowth response
A greenhouse mesocosm study was conducted to determine theimpacts of weathered and emulsified Macondo oil from the DWHspill on common reed, P. australis. Thirty P. australis sods (28 cmdiameter � 30 cm deep) containing intact vegetation and soil werecollected in October 2011 from an unoiled marsh site located inMadisonville, Louisiana (30�23.2050N, 90�09.5510W). After collec-tion, the sods were immediately transported to a greenhouse(21–40 �C) at Louisiana State University (LSU) where they accli-mated for two months before the oil treatment began. During theacclimation period, non-treated soil samples from supplementalsods were collected using a 7 cm � 41 cm coring tube, dried to aconstant weight at 60 �C, and analyzed by the Coastal WetlandSoils Characterization Lab, School of Plant, Environmental and SoilSciences, LSU Agricultural Center. Soil textures was classified asloam with 27.9 ± 3.6% sand, 45.3 ± 3.3% silt, 26.8 ± 2.9% clay, and22.4 ± 2.8% organic matter (mean ± SE). Soil pH and salinity were6.4 ± 1.1 and 1.6 ± 0.5 psu, respectively.
2.1.1. Deepwater Horizon oil, weathering process and hydrocarbonanalysis
We used Macondo (MC252) source oil, the oil released duringthe DWH event, for this research. The Macondo MC252 oil has anAPI gravity of approximately 37, and contains a relatively largeproportion of light hydrocarbon compounds, as typical of southLouisiana crude (SLC) (Platts Oilgram, 2010). To simulate theweathering process the oil would have undergone between itsentry into Gulf of Mexico waters and contact with the plantmaterial, the oil was artificially weathered 40% by weight usingwind (greenhouse fan) and sunlight. After weathering, the oilwas then emulsified to approximately a 50% water contentby pouring the oil over 15 cm of 3% salt water (using Instant
Ocean™) and mixing with submersible pumps for a week (Linand Mendelssohn, 2012).
The weathered and emulsified Macondo oil was analyzed foralkanes (nC10–nC35) and polycyclic aromatic hydrocarbons(PAHs) by gas chromatography (GC)/mass spectrometry (MS) withan Agilent 6890A GC directly interfaced to an Agilent 5973 MS(Response & Chemical Assessment Team Laboratory, Departmentof Environmental Sciences, Louisiana State University). The GCwas configured with a 5% diphenyl/95% dimethyl polysiloxanehigh-resolution capillary column (30 m, 0.25 mm ID, 0.25 micronfilm). The injection temperature was set at 280 �C and only high-temperature, low thermal-bleed septa were used in the GC inlet.The GC was operated in the temperature program mode with aninitial column temperature of 60 �C for 3 min, and then increasingto 280 �C at a rate of 5 �C/min and held for 3 min. The oven wasthen heated from 280 �C to 300 �C at a rate of 1.5 �C/min and heldat 300 �C for two minutes. The interface to the MS was maintainedat 300 �C. The MS was operated in the Selective Ion Monitoring(SIM) to maximize the detection of the target analytes. The concen-tration of specific target oil analytes was determined by a 5-pointcalibration and internal standard method. Internal standardsincluded naphthalene-d8, acenaphthalene-d10, chrysene-d12,and perylene-d12. Recovery of all analytes was estimated usingtwo hydrocarbon surrogate standards: 5 alpha androstane(alkanes) and phenanthrene-d10 (PAHs). Total alkane and totalaromatic concentrations in the weathered and emulsified oil were38,339 lg g�1 and 5,917 lg g�1, respectively.
2.1.2. Experimental design and proceduresThe most common field oiling scenarios were applied in a
greenhouse to simulate and investigate oiling effects on P. australis(see Lin and Mendelssohn, 2012). The experimental design was arandomized block with five replicate blocks containing six sodseach. Within each block, a sod was randomly assigned one of sixoil treatment-levels: (1) oil coverage of the lower 30% of theshoot-height of P. australis (30-oil), (2) oil coverage of the lower70% of the shoot-height (70-oil), (3) repeated oil coverage of thelower 70% of the shoot-height (70-rep-oil), (4) 100% oil coverageof shoots (100-oil), (5) soil oiling with 8 L m�2 of oil added to thesoil surface and allowed to penetrate the sediment (Oil-Soil), and(6) no oil treatment (Control). For the soil oiling treatment, oilwas applied over standing water in each sod for even dispersal.The water was then drained through the soil column to facilitateoil contact and penetration into the sediment. The drained waterfrom each sod was collected in a receptacle and added back tothe same sod at the end of the day until oil was no longer clearlyvisible on the water surface, implying all oil had successfully ad-sorbed into the soil profile. Oil was applied to shoots by turningthe sod on its side and applying oil with a paint brush, making sureno oil contacted the soil. For the 70% repeated oil coverage treat-ment, shoot-oiling was repeated every four days for 10 weeks toapproximate an oil release that cannot be immediately stopped,similar to the DWH event.
2.1.3. Plant metricsData collection occurred four times over the course of the
24-week experiment, with each collection-event taking place overa five-day period. Plant stem density was determined by directlycounting the number of individual stems in each sod. Cumulativestem length was determined separately for green (indicating live)and brown (indicating oiled or dead) plant material by measuringand summing the lengths of each stem from the sod surface to thetip of the stem’s newest unfolded leaf. Relative growth rates weredetermined by tagging two stems from each sod, measuring theirheights, and then remeasuring their heights a second time after

C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76 71
five days. Relative growth rate (RGR) was then calculated from thetwo measurements using the following equation:
RGR ¼ ðln W2� ln W1Þ=ðt2� t1Þ
where W2 is the second height measurement, W1 is the firstmeasurement (both in mm), t2 minus t1 is the duration of growth(in days), and ln is the natural logarithm. Relative growth ratewas expressed as mm of growth per mm of stem length per day(mm mm�1 d�1).
Final aboveground biomass (live, dead, and total) was collectedat the conclusion of the experiment, 24 weeks after oiling, by har-vesting all aboveground plant material from the sods approxi-mately 2 cm above the soil surface. Collected plant material wasseparated into living and dead tissue. If there were dead leaveson living stems, they were stripped and added to the dead tissueportion. Plant material was then dried to a constant weight at60 �C. A final measurement of plant stem density was also col-lected at the same time as the aboveground biomass harvest.
2.1.4. Soil oil concentration and shear strengthTotal petroleum hydrocarbon (TPH) concentration was mea-
sured by collecting approximately 2 cm of surface soil from eachsod, extracting the oil with dichloromethane (DCM), and analyzinggravimetrically (Lin and Mendelssohn, 1996). Briefly, approxi-mately 5 g of soil were placed into a glass vial, the oil extractedfrom the soil using DCM, and the DCM extract transferred to apre-weighed foil dish. Once in the dish, the DCM evaporated leav-ing only the oil, which was weighed to the nearest 0.0001 g. TheTPH concentration was expressed as mg g�1 dry soil.
Soil shear strength was measured at the termination of theexperiment in each of the sods at four depth-intervals (0–5,5–10, 10–15 and 15–20 cm) with a hand vane (3.4 cm wide � 5 cmlong) and torque gauge (Geotechnics, Auckland, New Zealand).
2.2. Experiment 2 – effects of oil applied to the soil on P. australisgrowth response
2.2.1. Experimental design and proceduresAt the conclusion of Experiment 1, a second experiment was
initiated to further investigate the effects of oil added to the soil.The aboveground biomass from the 25 sods that received onlyshoot-oiling and the no-oil controls in Experiment 1 were har-vested and allowed to regrow for two months, at which pointDWH source oil, weathered and emulsified as in Experiment 1 (to-tal alkanes = 21,236 lg g�1; total aromatics = 3169 lg g�1), wasapplied to the marsh sods at rates of 0 (control), 4, 8, 12, and16 L m�2. The experimental design was a randomized block thatcontained five treatment-levels replicated in five blocks (i.e., 25sods in total).
2.2.2. Plant and soil metricsData collection occurred three times over 9-week period, with
each collection taking place over a 5-day period. All variables mea-sured during the first experiment were also measured during thesecond experiment (i.e., stem density, cumulative stem length,RGR, final aboveground biomass and stem density at week 9, andTPH). In addition, final belowground biomass was collected at theconclusion of the experiment with a thin-walled aluminum coretube (7 � 41 cm) and belowground material washed of soil overa 2-mm mesh screen. The remaining material was then separatedinto categories of live roots, live rhizomes, dead roots, and deadrhizomes and dried to a constant weight at 60 �C and weighed.
Soil redox potential (Eh) was measured in the second experi-ment at 1 cm and 15 cm into the soil column with an AccumentAP71 pH/mV/�C meter (Fisher Scientific, Waltham, MA) connectedto a Corning hi-stab calomel reference electrode and bright
platinum electrodes. Measurements were taken from each sodwith six platinum electrodes, three at each depth. The electrodeswere left in place for 24 h before data were collected to assurestable readings.
2.3. Statistical analyses
All statistical analyses were conducted using SAS (version 9.2,SAS Institute, Cary, NC). We used univariate mixed-model ANOVAs(PROC MIXED) to determine the effects of oil treatment, time (as arepeated measure when applicable), and their interaction on thefollowing dependent variables for Experiments 1 and 2 individu-ally: stem density, total stem length, RGR, soil redox, abovegroundbiomass, belowground biomass, and soil TPH. When ANOVAs weresignificant at p < 0.05, treatment means in PROC MIXED weretested using the least-square (LS) means procedure with aTukey–Kramer post-host adjustment to maintain an experiment-wise error rate of 5%. When necessary, these data were logarithmi-cally or square transformed prior to analysis to improve ANOVAassumptions of normality and homogeneity of variance. All mea-sures of significance were identified at p < 0.05 unless otherwisestated.
We were unable to analyze shoot branching data collectedduring Experiment 1 using ANOVA due to the number of zeros con-tained within the data set. Instead, we used logistic regression(PROC GLIMMIX) to determine the effects of oil treatment, time,and their interaction on the probability of shoot branching as a bin-ary distribution (i.e., branching present versus branching absent).However, the treatment by time interaction was not significantand was removed from the model prior to analysis because thetype III test of fixed effects are fully adjusted for this interaction,if included. As such, the final analysis contained fixed effects foroil treatment and time, individually.
3. Results
3.1. Experiment 1 – effects of common oiling scenarios on P. australisgrowth
As expected, soil TPH was significantly higher (p < 0.0001) whenoil was added to the soil (27 ± 2.7 mg g�1) compared to those treat-ment-levels that received oil coating of aboveground shoots/leavesalone as well as the controls (control = 1.1 ± 0.1 mg g�1; 30-oil =1.1 ± 0.1 mg g�1; 70-oil = 1.3 ± 0.2 mg g�1; 70-rep-oil = 1.3 ± 0.2 mg g�1;100-oil = 1.7 ± 0.4 mg g�1).
Oil exposure affected total (green + brown tissue) cumulativestem length, but this effect varied significantly over time(p < 0.0001; Fig. 1A). During the first six weeks following exposure,cumulative stem length was similar for all treatment-levels, butthereafter diverged to where at the end of the experiment 100%shoot-oiling resulted in significantly greater total cumulative stemlength than soil-oiling (p < 0.005; Fig. 1A). Trends were similar forboth live (p < 0.0001) and brown (p < 0.0001) tissue cumulativestem length when analyzed separately (data not shown). Side-branching, the production of new shoots from the nodes of oiledstems, was dramatic in the 100-oil treatment-level, and theprobability of shoot branching significantly decreased with lessshoot-oiling (Fig. 2). When side-branch lengths were subtractedfrom total cumulative stem length, the impact of oiling over timewas more apparent in the oil-soil and 70-rep-oil treatment-levels(Treatment*Week interaction: p < 0.0001; Fig. 1B).
Oil treatment also had significant effects on end-of-the-experi-ment live (p < 0.05) and dead (p < 0.06) aboveground biomass,respectively (Fig. 3). Live aboveground biomass in sods receivingthe oil-soil treatment-level was significantly lower than that for

Fig. 1. Effects of oil treatment-levels on cumulative stem length, with (A) and without (B) side-branches, from 0 to 23 weeks after oiling. The values are means ± standarderrors (n = 5).
72 C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76
the 70-oil treatment-level (p < 0.02), but none of the treatmentswere significantly different from the control (Fig. 3). Althoughthe oil-soil treatment-level had a significant effect on dead above-ground biomass at p < 0.06, absolute differences were small(Fig. 3). Total aboveground biomass showed similar significanttrends as the live (Fig. 3).
Stem density and RGR were unaffected by oiling. The mean den-sity of live, dead, and total (live + dead) stems were 17 ± 1stems sod�1, 1 ± 0.25 stems sod�1, and 18 ± 1 stems sod�1, respec-tively. Relative growth rate averaged 0.009 ± 0.001 mm�1 day�1.
3.2. Experiment 2 – effects of soil oiling on P. australis growth
Soil TPH levels increased significantly with each increasing oildosage to the soil (p < 0.0001) (Fig. 4). Correspondingly, soil redoxwas significantly lower (p < 0.001) than the control for all soil oil-ing treatments except 12 L m�2 (4 L m�2 = �111.9 ± 12.2 mV,
8 L m�2 =�125.1 ± 10.5 mV, 12 L m�2 =�110.2 ± 14.5 mV, 16 L m�2 =�145.7 ± 13.2 mV, control = �68.5 ± 15.3 mV).
Oil application to the soil significantly reduced the change in to-tal (green + brown tissue) cumulative stem length over an eight-week period (significant treatment effect, p < 0.0002; Fig. 5). Thecontrol treatment-level had a significantly greater increase in totalcumulative stem length compared to all other oiling treatment-levels (p < 0.0002). Also, the 4 L m�2 treatment-level had a signifi-cantly greater increase in cumulative stem length than the16 L m�2 treatment-level. Similarly, oil-treatment significantlyreduced live aboveground biomass (p < 0.0002; Fig. 6). The controlhad the highest live aboveground biomass and the 16 L m�2 oildosage the lowest with the other oil treatment-levels intermediate(Fig. 6). Total aboveground biomass (live + dead) showed similartrends (p < 0.0003) as the live, while the dead aboveground bio-mass did not significantly vary (p > 0.20) among treatment-levels(Fig. 6).
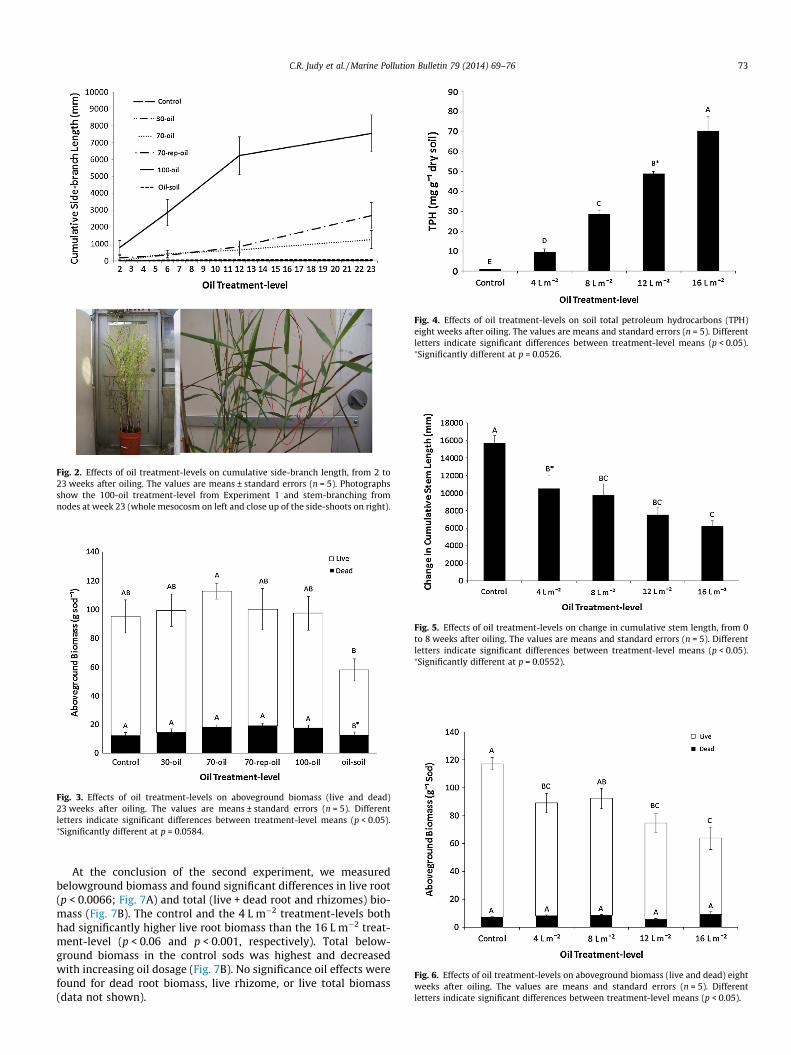
Fig. 2. Effects of oil treatment-levels on cumulative side-branch length, from 2 to23 weeks after oiling. The values are means ± standard errors (n = 5). Photographsshow the 100-oil treatment-level from Experiment 1 and stem-branching fromnodes at week 23 (whole mesocosm on left and close up of the side-shoots on right).
Fig. 3. Effects of oil treatment-levels on aboveground biomass (live and dead)23 weeks after oiling. The values are means ± standard errors (n = 5). Differentletters indicate significant differences between treatment-level means (p < 0.05).⁄Significantly different at p = 0.0584.
Fig. 4. Effects of oil treatment-levels on soil total petroleum hydrocarbons (TPH)eight weeks after oiling. The values are means and standard errors (n = 5). Differentletters indicate significant differences between treatment-level means (p < 0.05).⁄Significantly different at p = 0.0526.
Fig. 5. Effects of oil treatment-levels on change in cumulative stem length, from 0to 8 weeks after oiling. The values are means and standard errors (n = 5). Differentletters indicate significant differences between treatment-level means (p < 0.05).⁄Significantly different at p = 0.0552).
Fig. 6. Effects of oil treatment-levels on aboveground biomass (live and dead) eightweeks after oiling. The values are means and standard errors (n = 5). Differentletters indicate significant differences between treatment-level means (p < 0.05).
C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76 73
At the conclusion of the second experiment, we measuredbelowground biomass and found significant differences in live root(p < 0.0066; Fig. 7A) and total (live + dead root and rhizomes) bio-mass (Fig. 7B). The control and the 4 L m�2 treatment-levels bothhad significantly higher live root biomass than the 16 L m�2 treat-ment-level (p < 0.06 and p < 0.001, respectively). Total below-ground biomass in the control sods was highest and decreasedwith increasing oil dosage (Fig. 7B). No significance oil effects werefound for dead root biomass, live rhizome, or live total biomass(data not shown).

Fig. 7. Effects of oil treatment-levels on live root biomass (A) and total (live anddead, roots and rhizomes) biomass (B) eight weeks after oiling. The values aremeans and standard errors (n = 5). Different letters indicate significant differencesbetween treatment-level means (p < 0.05). ⁄Significantly different at p = 0.0599.
74 C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76
Just as in Experiment 1, RGR once again was unaffected by theoiling treatment or by its interaction with time, and averaged0.004 ± 0.001 mm mm�1 day�1 regardless of treatment. Also, theoiling treatment did not significantly affect live and total (live +dead) stem density (25 ± 2 stems sod�1 and 26 ± 1 stems sod�1,respectively). Oiling did not affect soil shear strength at any depth(overall mean and standard error = 31 ± 3 kPa).
4. Discussion
We investigated what had previously been identified as themost common oiling scenarios to coastal wetlands during theDeepwater Horizon oil release (Lin and Mendelssohn, 2012). Our re-search demonstrated the tolerance of P. australis to weathered andemulsified Macondo oil when applied to aboveground tissues andto the soil at lower doses. It became clear during the 23-weekstudy period of Experiment 1 that most of the growth in responseto the 100-oil treatment-level, and to a lesser degree the 70-rep-oiland 70-oil treatment-levels, occurred from the production of newshoots from the nodes of oiled stems. This production of side-shoots has not been noted in previous studies investigating theeffects of oiling on P. australis but has been known to occur inresponse to insect damage. For example, van der Toorn and Mook(1982) reported that stem borers of the genus Archanara can causeheavy damage to P. australis, but most of the shoots in their studysurvived because of side-shoot production. Tscharntke (1990)noted a similar response, finding that the stem boring caterpil-lar, Archanera geminipuncta, reduced shoot length greatly andinduced the production of narrow side-shoots. Tscharntke (1990)
concluded that the abundance of insect-damaged shoots was cor-related with the number of side-shoots. The present study, in com-bination with the aforementioned entomological work, suggeststhat the production of side-shoots by P. australis is a generalresponse to environmental stressors on aboveground tissues bythis species. Furthermore, the absence of an immediate shootresponse following soil-oiling was likely due to the fact that therewas no obvious stress to the shoots of P. australis with this treat-ment level. One might hypothesize that if there was an above-ground oil exposure in conjunction with the belowground oiling,the production of new side-shoots might have reduced the nega-tive impacts of the belowground oiling on total plant biomass.
Lin and Mendelssohn (2012) reported a similar response toshoot-oiling for S. alterniflora as was identified here for P. australis.After seven months under the same oil treatment-levels and withMacondo oil prepared in a similar fashion as the present study, S.alterniflora showed no difference in canopy height compared tothe control and only showed a negative impact to live stem densityin the 70-rep-oil treatment-level. In that same study, J. roemerianushad recovery of canopy height at the 30-oil and 70-oil treatment-levels while live stem density recovered from the 30-oil treatmentlevel. Also, when South Louisiana ‘‘sweet’’ crude oil (SLC), weath-ered to 87% and 67% of its original weight, was applied to thebottom 30 cm of S. alterniflora stems, rapid death occurred, butnew stems began growing within seven days of oiling (Pezeshkiet al., 1995). These results suggest that oiling, especially withweathered oil, to aboveground shoots of a number of marsh spe-cies does not cause lasting damage to plant vigor. Just as P. australisproduces side-branches from oiled shoots and quickly recoversfrom aboveground tissue oiling, S. alterniflora produces newgrowth from its underground rhizomes.
Similar to cumulative stem length, total and live abovegroundbiomasses were also impacted to a greater extent when oil wasapplied to the soil compared to the other treatment-levels. Thisresult was re-affirmed in Experiment 2, which demonstrated thatincreases in oil dosage caused decreases in cumulative stem length,aboveground biomass, and total and live-root belowground bio-mass. These findings are supported by others who have investi-gated the effects of oil on P. australis. Zhu et al. (2013) showedthat increasing TPH levels, associated with seepage around oilwells at the Chengdong oilfield in Eastern China, were directly cor-related with reduced aboveground biomass of P. australis. Dowtyet al. (2001) found a decrease in both above- and belowgroundbiomass of P. australis collected from the Manchac WildlifeManagement Area in south Louisiana when soil was exposed to5 L m�2 and 10 L m�2 of un-weathered SLC oil in a greenhousemesocosm study. Nie et al. (2010) also found a decrease in above-and belowground biomass of P. australis grown from seed in thegreenhouse when un-weathered crude oil from the Shengli oilfieldin China was mixed with P. australis soil at concentrations of6000 mg kg�1 and 12,000 mg kg�1. However, the latter two studiesused un-weathered oil, which would likely be much more toxicthan the weathered Macondo oil used in our research. Similarreductions with respect to shoot length were observed when P.australis shoots alone were exposed to liquid paraffin andun-weathered diesel oil (Armstrong et al., 2009). Collectively, theseresults demonstrate that oil exposure can negatively impactP. australis growth, but that the intensity of impact and this species’ability to rapidly regrow is dependent on the extent of oil weath-ering, the volume/concentration of the oil, and the mode ofexposure (aboveground versus belowground).
Research completed shortly after the DWH oil spill, usingsimilar weathered and emulsified Macondo oil as in the presentexperiment, found that aboveground biomass and live stem den-sity of both S. alterniflora and J. roemerianus, two dominant saltmarsh plants, were significantly impacted by moderate to heavy

C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76 75
soil oiling (Lin and Mendelssohn, 2012). This research was the firstto show that weathered and emulsified DWH oil was still toxic en-ough to impact plant growth, and it agrees with previous workdone on S. alterniflora after an oil spill in a bayou adjacent toGalveston Bay, Texas in January 1984 (Alexander and Webb,1987). Although the oil from the Texas study had much less timeto weather before contacting the salt marsh vegetation than theMacondo oil, it still showed that 5–51 mg/g of oil in the sedimentof a natural salt marsh caused significantly reduced growth ofS. alterniflora after 18 months. A study of oil effects on J. roemerianusthat used an unweathered mix of Empire and Saudi Arabian crudeoils found that a single dose of 1.5 L m�2 or 6–10 successivemonthly doses of 0.6 L m�2 completely killed the plants andcontinued to suppress growth for up to two years, although onlyan initial impact was seen at single low doses ranging from 0.25to 0.6 L m�2 (De la Cruz et al., 1981).
Field plots examined by Lin and Mendelssohn (2012), postDWH, contained TPH concentrations of approximately 500 mg g�1
in heavily oiled Louisiana salt marshes and approximately80 mg g�1 in moderately oiled marshes. They estimated that TPHconcentrations in heavily oiled marshes, where marsh plants wereseverely impacted, were equivalent to approximately 20 L oil m�2
as used in their mesocosm study. Although this dosage was 20%higher than the present study’s highest oil treatment-level(16 L m�2), it is not unreasonable to predict that similar impactscould have occurred in heavily oiled P. australis dominated sitesin the Mississippi River Birdfoot delta if the oil penetrated the soil.However, field data on P. australis response to the DWH oiling arepresently lacking.
When oil penetrates soil, the increase in labile carbon can pro-mote microbial activity and affect soil biogeochemistry (Ellis andAdams, 1961). We found that with greater oil dosages to the soiland resultant higher soil TPH concentrations, the soils becamemore biochemically reduced. Oil additions to the soil can stimulatemicrobial activity and soil respiration, which in turn reduces oxy-gen and other electron acceptors and thus lowers soil Eh (Ellis andAdams, 1961; Li et al., 1990; Llangovan and Vivekanandan, 1992;Nie et al., 2010). However, lower soil redox conditions associatedwith oil additions to the soil can be an initial impact that doesnot persist (Nyman, 1999; Nyman and McGinnis, 1999). Somestudies have found no effect on soil Eh by additions of oil to the soil(Lin and Mendelssohn, 1996; DeLaune et al., 1979), possibly due tothe transient nature of oil impacts on soil Eh (Nyman, 1999;Nyman and McGinnis, 1999).
5. Conclusion
Oiling intensity of P. australis following the DWH spill was quitevariable in the Birdfoot Delta of the Mississippi River, as it wasalong the entire coast of Louisiana. Consequently, this researchevaluated a suite of oiling scenarios and intensities. Our resultsshow that applying weathered and emulsified Macondo oil to theaboveground tissue of P. australis had no major impact on overallplant growth. In fact, shoot-oiling increased plant growth by induc-ing a vegetative stress response, whereby P. australis producedside-branches to compensate for the loss of photosynthetic tissuecaused by the mortality of oiled leaves. However, if the weatheredand emulsified oil penetrates the soil, the growth of P. australis canbe reduced, depending on the growth parameter and oil dosage.Based on comparable research (Lin and Mendelssohn, 2012),P. australis shows similar resilience to the Macondo oil asS. alterniflora, both of which are more resilient than J. roemerianus.These results support our original prediction that growth responsewill be little affected from oiling of aboveground plant tissues,while oil exposure belowground will have growth-limiting impacts
that increase with increasing dosage. Of course, the outcomes ofthis prediction, whether in the field or greenhouse, will varydepending on the degree to which the oil has weathered, thevolume and concentration of oil, the frequency of exposure, andthe influence of other environmental factors, e.g., salinity andflooding, that may modulate the effect of oiling. Future researchencompassing an even broader range of oiling conditions, includ-ing an endpoint of complete plant mortality, and other growthmodifying environmental variables would go a long way inquantifying the maximum oiling that this species could survive, apotentially important consideration in determining the intensityof spill response.
Acknowledgments
The Coastal Science Assistantship Program, sponsored by theLouisiana Department of Natural Resources and Louisiana SeaGrant, and the BP/Gulf of Mexico Research Initiative funded this re-search. Nick Keeney, Kris Ackoury, Luke Judy, Whitney Kiehn, andJason Pietroski provided field and laboratory assistance. We wouldalso like to thank two anonymous reviewers for improving theclarity of the manuscript.
References
Alexander, S.K., Webb, J.W., 1987. Relationship of Spartina alterniflora growth tosediment oil content following an oil spill. In: Proceedings of the 1987 Oil SpillConference. American Petroleum Institute, Washington, DC, pp. 445–449.
Alizai, S.A.K., McManus, J., 1980. The significance of reed beds on siltation in the Tayestuary. P. Roy. Soc. Lond. B. Bio. 78, s1–s13.
Anderson, C.J., Hess, T.A., 2012. The effects of oil exposure and weathering on black-needle rush (Juncus roemerianus) marshes along the Gulf of Mexico. Mar. Pollut.Bull. 64, 2749–2755.
Armstrong, J., Keep, R., Armstrong, W., 2009. Effects of oil on internal gas transport,radial oxygen loss, gas films and bud growth in Phragmites australis. Ann. Bot.London 103, 333–340.
Bianchi, V., Peruzzi, E., Masciandaro, G., Ceccanti, B., Mora Ravelo, S., Iannelli, R.,2011. Efficiency assessment of a reed bet pilot plant (Phragmites australis) forsludge stabilization in Tuscany (Italy). Ecol. Eng. 37, 779–785.
Biber, Patrick D., Wu, W., Peterson, M.S., Liu, Z., Pham, L., 2014. Oil contamination inMississippi saltmarsh habitats and the impacts to Spartina alterniflora photosynthesis.In: Alford, J.B., Peterson, M.S., Green, C.C. (Eds.), Impacts of Oil Spill Disasterson Marine Habitats and Fisheries in North America, CRC Press, Boca Raton,FL.
Borin, M., Milani, M., Salvato, M., Toscano, A., 2011. Evaluation of Phragmitesaustralis (Cav.) Trin. Evapotranspiration in northern and southern Italy. Ecol.Eng. 37, 721–728.
Brix, H., 1987. Treatment of wastewater in the rhizosphere of wetland plants- theroot-zone method. Water Sci. Technol. 19, 107–108.
Camilli, R., Di lorio, D., Bowen, A., Reddy, C.M., Techet, A.H., Yoerger, D.R., Whitcomb,L.L., Seewald, J.S., Sylva, S.P., Fenwick, J., 2011. Acoustic Measurement ofthe Deepwater Horizon Macondo well flow rate. P. Natl. Acad. Sci. USA 109,20235–20239.
Cole, S., 1998. The emergence of treatment wetlands. Environ. Sci. Technol. 32, 218–223.
De la Cruz, A.A., Hackney, C.T., Rajanna, B., 1981. Some effects of crude oil on aJuncus tidal marsh. J. Elisha Mitch. Soc. Sci. 97, 14–28.
DeLaune, R.D., Patrick Jr., W.H., Buresh, R.J., 1979. Effect of crude oil on a LouisianaSpartina alterniflora salt marsh. Environ. Pollut. 20, 21–30.
DeLaune, R.D., Pezeshki, S.R., Jugsujinda, A., Lindau, C.W., 2003. Sensitivity of USGulf of Mexico coastal marsh vegetation to crude oil: comparison of greenhouseand field responses. Aquat. Ecol. 37, 351–360.
Dowty, R.A., Shaffer, G.P., Hester, M.W., Childers, G.W., Campo, F.M., Greene, M.C.,2001. Phytoremediation of small-scale oil spills in fresh marsh environments: amesocosm simulation. Mar. Environ. Res. 52, 195–211.
Ellis, R., Adams Jr., R.J., 1961. Contamination of soils by petroleum hydrocarbons.Adv. Agron. 13, 197–213.
Gorai, M., Ennajeh, M., Khemira, H., Neffati, M., 2011. Influence of NaCl-salinity ongrowth, photosynthesis, water relations and solute accumulation in Phragmitesaustralis. Acta Physiol. Plant. 33, 963–971.
Hartog, D., Kvet, J., Sukopp, H., 1989. Reed a common species in decline. Aquat. Bot.35, 1–4.
Li, Y., Morris, J.T., Yoch, D.C., 1990. Chronic low level hydrocarbon amendmentsstimulate plant growth and microbial activity in salt-marsh microcosms. J. Appl.Ecol. 27, 159–171.
Lin, Q., Mendelssohn, I.A., 1996. A comparative investigation of the effects ofLouisiana crude oil on the vegetation of fresh, brackish, and salt marsh. Mar.Poll. Bull. 32, 202–209.

76 C.R. Judy et al. / Marine Pollution Bulletin 79 (2014) 69–76
Lin, Q., Mendelssohn, I.A., 2012. Impacts and recovery of the Deepwater Horizon oilspill on vegetation structure and function of coastal salt marshes in thenorthern Gulf of Mexico. Environ. Sci. Technol. 46, 3737–3743.
Llangovan, K., Vivekanandan, M., 1992. Effect of oil pollution on soil respiration andgrowth of Vigna mungo (L.) Hepper. Sci. Total Environ. 116, 187–194.
Mendelssohn, I.A., Andersen, G.L., Baltz, D.M., Caffey, R.H., Carman, K.R., Fleeger,J.W., 2012. Oil impacts on coastal wetlands: implications for the MississippiRiver delta ecosystem after the Deepwater Horizon Oil Spill. BioScience 62,562–574.
Michel, J., Owens, E.H., Zengel, S., Graham, A., Nixon, Z., Allard, T., Holton, W.,Reimer, P.D., Lamarche, A., White, M., Rutherford, N., Childs, C., Mauseth, G.,Challenger, G., Taylor, T., 2013. Extent and degree of shoreline oiling: DeepwaterHorizon oil spill, Gulf of Mexico, USA. PLOSone, 8, 1–9 and SupplementalInformation.
National Commission on the BP Deepwater Horizon Oil Spill and Offshore Drilling.Deep Water: The Gulf Oil Disaster and the Future of Offshore Drilling, Report tothe President, January 2011. Government Printing office, pp. 392. <http://www.oilspillcommission.gov/sites/default/files/documents/DEEPWATER_Reporttothepresident_FINAL.pdf>.
Nie, M., Yang, Q., Jiang, L.F., Fang, C.M., Chen, J.K., Li, B., 2010. Do plants modulatebiomass allocation in response to petroleum pollution? Biol. Lett. 6, 811–814.
Nyman, J.A., 1999. Effects of crude oil and chemical additives on metabolicactivity of mixed microbial populations in fresh marsh soils. Microbial Ecol. 37,152–162.
Nyman, J.A., McGinnis, T.E., 1999. Effects of crude oil and spill-response-options onmicrobial functions and oil disappearance in salt marsh soils: Year 2.Department of Biology, University of Southwestern Louisiana. LouisianaApplied and Educational Oil Spill Research and Development Program,OSRADP Technical Report Series, 99-007.
Pezeshki, R.S., DeLaune, R.D., Nyman, J.A., Lessard, R.R., Canevari, G.P., 1995.Removing oil and saving oiled marsh grass using a shoreline cleaner. In:Proceedings of the 1995 Oil Spill Conference. American Petroleum Institute,Washington, DC, pp. 529–533.
Pezeshki, S.R., Hester, Lin, Q., Nyman, J.A., 2000. The effects of oil spill and clean-upon dominant US Gulf coast marsh macrophytes: a review. Environ. Pollut. 108,129–139.
Platts Oilgram News/OPR Extra. June 15, 2010. <http://www.platts.com/IM.Platts.Content.InsightAnalysis/NewsFeature/2010/oilspill/20100615.Pdf>.
Toyama, T., Furukawa, T., Maeda, N., Inoue, D., Sei, K., Mori, K., Ike, M., 2010.Accelerated biodegradation of pyrene and benzo[a]pyrene in the Phragmitesaustralis rhizosphere by bacteria–root exudate interactions. Water Res. 45,1629–1638.
Toyama, T., Murashita, M., Kobayashi, K., Kikuchi, S., Sei, K., Tanaka, Y., Ike, M., Mori,K., 2011. Acceleration of nonylphenol and 4-tert-octylphenol degradation insediment by Phragmites australis and associated rhizosphere bacteria. Environ.Sci. Technol. 45, 6524–6530.
Tscharntke, T., 1990. Fluctuations in abundance of a stem-boring moth damagingshoots of Phragmites australis: causes and effects of overexploitation of food in alate-successional grass monoculture. J. Appl. Ecol. 27, 679–692.
Van der Toorn, J., Mook, J.H., 1982. The influence of environmental factors andmanagement on stands of Phragmites australis. I. Effects of burning, frost andinsect damage on shoot density and shoot size. J. Appl. Ecol. 19, 477–499.
Wu, W., Biber, P.D., Peterson, M., Gong, C., 2012. Modeling photosynthesis ofSpartina alterniflora (smooth cordgrass) impacted by the Deepwater Horizon oilspill using Bayesian inference. Environ. Res. Lett. 7, 045302.
Zhu, L., Zhao, X., Lai, L., Wang, J., Jiang, L., Ding, J., Liu, N., Yu, Y., Li, J., Xiao, N., Zheng,Y., Rimmington, G.M., 2013. Soil TPH concentration estimation using vegetationindices in an oil polluted area of eastern China. PLOS ONE 8, e54028.




















